
Construction and Biological Testing of Artificial Implants for ...
121
Construction and Biological Testing of Artificial Implants for Peripheral Nerve Repair Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der RWTH Aachen University zur Erlangung des akademischen Grades eines Doktors der Naturwissenschaften genehmigte Dissertation vorgelegt von Diplom-Biologe Andreas Kriebel aus Leverkusen Berichter: Prof. Dr. Jörg Mey Prof. Dr. Hermann Wagner Tag der mündlichen Prüfung: 26.11.2015 Diese Dissertation ist auf den Internetseiten der Universitätsbibliothek online verfügbar
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Construction and Biological Testing of Artificial Implants for ...

Construction and Biological Testing of Artificial
Implants for Peripheral Nerve Repair
Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der
RWTH Aachen University zur Erlangung des akademischen Grades eines
Doktors der Naturwissenschaften genehmigte Dissertation
vorgelegt von
Diplom-Biologe
Andreas Kriebel
aus Leverkusen
Berichter: Prof. Dr. Jörg Mey
Prof. Dr. Hermann Wagner
Tag der mündlichen Prüfung: 26.11.2015
Diese Dissertation ist auf den Internetseiten der Universitätsbibliothek online verfügbar


Table of Contents
Summary ......................................................................................................... 1
Zusammenfassung ......................................................................................... 3
General Introduction ..................................................................................... 5
The peripheral nervous system ................................................................................................. 5
Injuries to the peripheral nervous system................................................................................ 6
Peripheral nerve repair ............................................................................................................. 7
Aim and structure of the thesis ................................................................................................. 8
Chapter I
Theoretical background on peripheral nerve repair and design strategies for
artificial implants ............................................................................................. 9
1 Introduction ............................................................................................................................. 9
2 Physiological requirements of artificial nerve bridges ...................................................... 10
2.1 Peripheral nerve regeneration .......................................................................................... 10
2.2 Secreted signals in peripheral nerve regeneration ........................................................... 12
2.3 Surface-bound signals ...................................................................................................... 14
2.4 Desired properties of the scaffold .................................................................................... 15
3 Design strategies for artificial nerve bridges ...................................................................... 17
3.1 Hollow tubes and multichannel nerve conduits ............................................................... 17
3.2 Structured implants with polymeric fibers ....................................................................... 21
3.3 Structured implants with gels and scaffolds .................................................................... 24
3.4 Implantation of cells with artificial nerve bridges ........................................................... 25
4 Materials for artificial nerve conduits ................................................................................. 26
4.1 Natural materials .............................................................................................................. 26
4.2 Synthetic polymers .......................................................................................................... 30
4.3 Functionalization with ECM proteins and peptides ......................................................... 35
4.4 Controlled release of growth factors ................................................................................ 37
5 Conclusion.............................................................................................................................. 39
5.1 Standardized control of efficiency ................................................................................... 39
5.2 Structured three-dimensional implants ............................................................................ 40
5.3 Biomimetic functionalization of implants ....................................................................... 40
5.4 The thirty millimeter mark ............................................................................................... 40
Chapter II
Manufacture and characterization of three-dimensional fiber arrays and
assessment of guidance properties in vitro .................................................... 41
1 Introduction ........................................................................................................................... 41
2 Material and Methods .......................................................................................................... 42

2.1 Electrospinning of three-dimensional fiber arrays .......................................................... 42
2.2 Physical characterization of fiber arrays ......................................................................... 44
2.3 Inserting the 3D fiber arrays into a conduit ..................................................................... 45
2.4 Cell and organ culture ..................................................................................................... 45
2.5 Immunocytochemistry ..................................................................................................... 46
2.6 Fluorescence microscopy ................................................................................................ 46
2.7 Quantification of Schwann cell migration and axonal growth ........................................ 47
3 Results .................................................................................................................................... 47
3.1 Electrospinning of parallel fibers in a 3D configuration ................................................. 47
3.2 Characterization of electrospun 3D PCL-fiber arrays ..................................................... 48
3.3 Suspension and manipulation of 3D fiber arrays in biocompatible gels ......................... 48
3.4 Influence of PCL fibers on axonal growth ...................................................................... 51
3.5 Influence of PCL fibers on glial cell migration ............................................................... 53
3.6 Incorporation of gel/fiber composites in a tubular conduit ............................................. 53
4 Discussion .............................................................................................................................. 54
4.1 Three-dimensional configuration of guidance structures ................................................ 55
4.2 Biological functionalization of the growth substrate....................................................... 56
5 Conclusion ............................................................................................................................. 57
Chapter III
Assembly of artificial nerve implants and assessment of their suitability to
support nerve regeneration in vivo ................................................................. 59
1 Introduction .......................................................................................................................... 59
2 Material and Methods .......................................................................................................... 60
2.1 Manufacture of artificial implants ................................................................................... 60
2.2 Animals and surgical procedure ...................................................................................... 61
2.3 Tissue preparation and immunohistochemistry ............................................................... 62
2.4 Image analysis ................................................................................................................. 63
2.5 Electromyography ........................................................................................................... 63
2.6 Behavioral experiments ................................................................................................... 64
2.7 Statistical analysis ........................................................................................................... 64
3 Results .................................................................................................................................... 65
3.1 Artificial nerve implants and general observations ......................................................... 65
3.2 Tissue regeneration: macroscopic evaluation of the implants ......................................... 66
3.3 Schwann cell infiltration and axonal regeneration in the implants ................................. 66
3.4 Size distribution of regenerated fibers ............................................................................. 70
3.5 Functional regeneration: electromyography .................................................................... 71
3.6 Functional regeneration: recovery of motor functions .................................................... 75
3.7 Muscular atrophy and regeneration ................................................................................. 77
4 Discussion .............................................................................................................................. 78

4.1 Regeneration in the artificial nerve implants ................................................................... 79
4.2 The importance of an internal guidance structure in artificial implants .......................... 80
4.3 Implementation of biological signals in synthetic cell-free nerve grafts ......................... 81
5 Conclusion.............................................................................................................................. 82
General Discussion ....................................................................................... 83
Comparison to other studies ................................................................................................... 83
Possible augmentation of implant design ............................................................................... 85
Conclusion .................................................................................................... 87
References ..................................................................................................... 89
Abbreviations ............................................................................................. 107
Figures and Tables ..................................................................................... 109
Prior Publications and Contributions ...................................................... 111
Acknowledgments ...................................................................................... 113
Curriculum Vitae ....................................................................................... 114


1
Summary
In this thesis I documented the development of an artificial nerve conduit and subsequent
biological testing within an animal model of peripheral nerve injury. The focus for the
development of the conduit lied in the utilization of electrospun microfibers to serve as a guidance
structure for Schwann cells and axons during regeneration. Through the incorporation of such a
guidance structure into the lumen of a conventional hollow nerve conduit, Schwann cell migration
and axonal elongation across artificial nerve conduits was supposed to be facilitated, thereby
expanding the applicability of artificial nerve conduits to the repair of larger nerve gaps.
To achieve this I initially established a procedure to produce three-dimensional arrays of aligned
microfibers. Through embedding of these arrays into a biocompatible hydrogel I created a
scaffold with an integrated guidance structure, which can be incorporated into a conventional
nerve conduit. This scaffold was then used to confirm in vitro, whether the guidance properties,
the fibers exerted under two-dimensional conditions, are preserved in a three dimensional
environment. Since the hydrogel (e.g. collagen) could potentially compete with the fibers for the
interaction with cells due to its fibrillary structure.
Subsequently the new implants were assembled through incorporation of the scaffold into
conventional conduits and the benefit of the integrated structure was then evaluated in comparison
to the autologous nerve graft and the conventional hollow conduit by using an animal model of
peripheral nerve injury. For this, a sciatic nerve lesion was performed in rats (Rattus norvegicus),
which was subsequently repaired by insertion of the respective im-/transplant. Assessment of
recovery by means of immunohistological investigation of the regenerated nerve,
electromyography, determination of muscle weight as well as different behavioral tests, revealed
an advantage of the internally structured conduit over the conventional hollow conduit,
particularly when the integrated fibers contained proteins derived from the extracellular matrix.
Thus, even though the autologous nerve graft still performed considerably better than the other
implants tested, the new structured design offers several possibilities for further optimization and
thus presents a promising approach to narrow the gap between autologous nerve grafts and their
artificial alternatives.


3
Zusammenfassung
In der vorliegenden Dissertation ist die Entwicklung eines künstlichen Nervenimplantats bis hin
zur Überprüfung der Wirksamkeit innerhalb eines Tierversuchs dokumentiert. Der Schwerpunkt
bei der Entwicklung des künstlichen Nervenimplantats lag in der Verwendung von Mikrofasern
als Leitstruktur für Schwannzellen und Axone während der Regeneration. Durch Integration
dieser Leitstruktur in das Lumen eines konventionellen Nervenconduits sollte die Wegfindung
besagter Zellen über das Conduit verbessert, und die Anwendung künstlicher Implantate zur
Überbrückung größerer Läsionen ermöglicht werden.
Um dies zu erreichen, wurde zunächst ein Verfahren zur Erzeugung parallel ausgerichteter
Mikrofasern in dreidimensionaler Anordnung entwickelt. Durch Einbetten der Fasern in ein
Hydrogel wurde eine Matrix mit integrierten Leitstrukturen erzeugt, die in ein konventionelles
Conduit integriert werden kann. Mit dieser Matrix wurde zunächst in einem in vitro Experiment
überprüft, ob die Fähigkeit der Fasern, die Wuchsrichtung von Zellen vorzugeben, auch in einer
dreidimensionalen Umgebung erhalten bleibt. Da diese aufgrund ihrer Struktur potentiell mit den
Fasern um die Interaktion mit den Zellen konkurrieren könnte.
Anschließend wurde die Wirksamkeit des neu entworfenen Implantats im Vergleich zum
autologen Transplantat und einem konventionellen Conduit, in einem Tierversuch überprüft.
Hierzu wurde eine Läsion des Nervus ischiadicus in der Ratte (Rattus norvegicus) gesetzt, durch
das entsprechende Im-/Transplantat überbrückt, und der regenerative Erfolg über einen Zeitraum
von zwölf Wochen mittels immunhistologischer Färbung der Nervenregenerate,
Elektromyographie, Bestimmung des Muskelgewichts der betroffenen Gliedmaßen und
verschiedener Verhaltensexperimente bewertet. Dabei zeigte sich eine Überlegenheit des von uns
entworfenen Implantats gegenüber dem konventionellen Implantat, insbesondere bei
Anreicherung der Fasern mit Proteinen der extrazellulären Matrix. Allerdings war die
Überlegenheit des autologen Transplantats gegenüber künstlichen Implantaten auch in unseren
Experimenten augenscheinlich. Nichtsdestotrotz scheint der hier vorgestellte Ansatz, bezüglich
der Struktur künstlicher Implantate, angesichts zahlreicher Möglichkeiten zur weiteren
Optimierung, geeignet den Abstand zwischen autologem Transplantat und künstlichen
Implantaten zu verringern.


The peripheral nervous system
5
General Introduction
In general, peripheral nerve injuries (PNI) are not very frequent. Different studies and statistical
surveys show an incidence of approximately two percent for injuries to the extremities (Robinson,
2004; Taylor et al., 2008) accompanied by a prevalence of around three percent (Noble et al.,
1998). Nevertheless, their socio-economic implications should not be underestimated.
First of all, there is thought to be a relatively high number of cases were the diagnosis of the
actual PNI is delayed, because shortly after an accident the treatment of the apparent injuries is
usually in the focus. Thus a considerable number of cases might fail to enter the statistics
correctly. Furthermore data from the Swiss accident analysis and statistics from 1997 (UVG)
show that the duration of working inability caused by injuries to nerves and the spinal cord is
nearly twice as long as compared to injuries in general (26,8 days vs 14,7 days). Additionally PNI
are mostly affecting the upper extremities. Thus the functional impairment in particular of the
hands can notably reduce the patient’s professional capacity for a long time and over a wide range
of working fields. This is displayed by the high follow-up costs of such injuries. For injuries to
the median and ulnar nerve, lost production makes up 87 percent of the total costs and 67 percent
for injuries to the hand and forearm in general (Rosberg et al., 2005). On top of that, the patients
suffering from PNI are relatively young with most patients between 20 and 40 years of age (Noble
et al., 1998; Eser et al., 2009; Ciaramitaro et al., 2010). For this group of people, the functional
impairment and the accompanying loss of personal independence, together with the high risk of
enduring neuropathic pain, strongly diminishes the perceived quality of life.
To explain why the treatment of these injuries presents such a challenge to the physicians in
charge, I will briefly described the causes, consequences and possible therapies of PNI in the
following.
The peripheral nervous system
The peripheral nervous system (PNS) comprises all nerves and ganglia not located within the
brain or the spinal cord, i.e. the cranial nerves (except of the optic nerve) and the spinal nerves.
The main function of the PNS is the relay of sensory information from the periphery to the central
nervous system (CNS) and the propagation of information from the CNS to the musculature and
other organs in the periphery. It can be divided into the somatic nervous system enabling
voluntary control of skeletal musculature and the autonomic nervous system mainly involuntarily
influencing internal organs and related body functions. Thus a peripheral nerve (PN) can,
depending on the innervated target organ, consist of a composition of afferent sensory axons and
efferent motor or effector axons, which according to the individual fiber type, are more or less
enwrapped by the cell membrane of Schwann cells, the myelinating glia cell of the PNS. The

General Introduction
6
single axons are surrounded by the endoneurium, a connective tissue forming an endoneurial tube
along the complete length of the axon. Multiple axons are ensheathed by a lamellar arrangement
of perineurial cells (the perineurium) forming a fascicle. Several of these fascicles, together with
blood vessels, are embedded in connective tissue, called epineurium, to form a complete PN.
Injuries to the peripheral nervous system
The PNS is the essential relay station for the interaction of vertebrates with their environment and
the regulation of their homeostasis. However, unlike the central parts of the nervous system, i.e.
brain and spinal cord, which are surrounded by the cranium and the vertebrate column, the PNS is
not protected by bone tissue. Thus nerves of the PNS are particularly susceptible to traumatic
injuries, especially if located in the extremities. The main cause for these injuries are traffic
accidents, followed by gunshot and cutting wounds, and are primarily affecting the upper
extremities as confirmed by an international survey in 49 countries (Scholz et al., 2009). The
consequence of a PNI is a loss of function of the innervated organ or muscle. The extent and
duration of this loss of function is essentially dependent on the position and severity of the trauma
and also on the applicable therapy.
The mildest form of PNI is a neuropraxia according to Seddon (Seddon, 1943) or a grade one
injury according to Sunderland (Sunderland, 1951). It can be caused for example by a nerve
compression that results in a damage of the myelin sheath and thus impairs signal conduction.
Since the nerve fibers as such stay intact, there usually is a rapid recovery of function after nerve
conduction is restored.
If the axon itself is damaged, but the surrounding connective tissue stays intact, one talks about
axonotmesis (Seddon, 1943) or Sunderland grade two. In this case degenerative processes will be
initiated, which for example lead to a break down and degradation of myelin and axonal parts
distal to the injury. This is followed by axonal regeneration from the proximal stump towards the
original site of innervation. The speed of functional recovery after axonotmesis is obviously
limited by the growth rate of the regenerating axons and thus does strongly depend on the distance
between site of injury and the target organ. However, as the still intact endoneurial tube serves as
a guidance structure for the regrowing axons, the prospects for functional recovery are relatively
high.
The most severe case of a PNI is called neurotmesis according to Seddon, which is further divided
by Sunderland in grades three to five and ranges from transection of the axon and its endoneurial
tube up to a complete transection of the nerve. In the milder cases of neurotmesis, where damage
to connective tissue structures is limited, regeneration is possible without surgical intervention,
but there is increasing probability of aberrant sprouting and neuroma formation. In the case of a

Peripheral nerve repair
7
complete nerve transection, however, surgical intervention is obligatory to preserve a chance for
functional recovery.
Peripheral nerve repair
A complete nerve transection results in a gap between the two nerve stumps. They have to be
reconnected by a surgical intervention to enable regeneration. If the resultant gap is small (less
than a few mm), e.g. after a clean cut, it can be sufficient to bring both stumps together and secure
them by end to end neurorrhaphy. However, frequently the amount of tissue damage or delayed
repair, which generates the necessity for trimming the nerve stumps to get a plane surface, leads
to nerve gaps considerable bigger than that. In this case continuity must not be restored by a
neurorrhaphy, since this would apply strain on the nerve, having detrimental effects for
regeneration. Thus, if relocation of the nerve to avoid strain is not possible, the insertion of a
nerve bridge is necessary (005/010 S3-Leitlinie, 2013). For this purpose, several biological
materials, e.g. muscle or vein grafts, are conceivable and their applicability is experimentally
investigated (Meek et al., 2004; Nijhuis et al., 2013). Despite this, the standard procedure is the
insertion of an autologous nerve graft, which usually is a piece of sensory nerve, most frequently
the N. suralis, N. saphenus, N. cutaneus antebrachii medialis or lateralis (005/010 S3-Leitlinie,
2013). This has, by far, the best prediction for a recovery of function, but comes along with major
drawbacks. These are, a limited availability of donor material, which more or less applies to most
biological materials, and a loss of function at the donor site, which is a particular problem of
autologous nerve grafts. To overcome these constrictions, there has been continuous research in
the past decades to test and evaluate different artificial materials in their ability to substitute the
established autologous nerve graft. It has led to the availability of clinically approved artificial
nerve conduits of different biodegradable, non-toxic materials that hardly exhibit complications,
e.g. encapsulation or increased inflammatory reactions (Meek and Coert, 2008a). However, while
these nerve conduits show satisfactory results when used to bridge small nerve gaps, the predicted
functional recovery dramatically decreases with increasing gap size, compared to the autologous
nerve graft. Therefore 30 mm are usually considered as the maximum gap size to be bridged via
tubulization with artificial nerve conduits (Meek and Coert, 2012, 2014). Generally this is
attributed to a lack of guidance cues and trophic support. An autologous nerve graft offers
thousands of endoneurial tubes that regrowing axons can follow to reach the distal stump, while
being supported by growth factors released from Schwann cells inside the graft. Compared to that
directional support in the conventional artificial nerve conduit is very poor, as they consist of
hollow tubes with a saline filling. For this reason, a main focus of current research regarding
peripheral nerve repair is the implementation of structural guidance cues into artificial nerve
conduits.

General Introduction
8
Aim and structure of the thesis
The goal of the project was to develop and test a cell-free artificial nerve conduit for peripheral
nerve repair that made use of electrospun, sub-micron scale fibers to improve axonal guidance. In
chapter I of the thesis I reviewed the theoretical background regarding biological processes
involved in peripheral nerve injury and regeneration as well as the current state of research in
peripheral nerve repair. Particular emphasis was put on design strategies for the improvement of
axonal guidance, the choice of materials (biological and non-biological) for artificial nerve
conduits and the incorporation of biological signaling molecules.
To achieve the experimental goal of an artificial nerve conduit the following milestones were to
be reached:
1. Designing an electrospinning device that allowed producing three-dimensional (3D)
arrays of parallel fibers for axonal guidance. This included the characterization of fiber
properties associated with their applicability for peripheral nerve repair, e.g. fiber
diameter, density and degree of alignment.
2. Manufacturing a 3D scaffold which incorporated the guidance fibers. This included,
finding a biocompatible hydrogel with suitable physical properties and establishing a
procedure for the embedding of the fiber arrays in the gel.
3. Verification of the biological properties, specifically with respect to axonal guidance and
cell migration, of the 3D fiber/gel construct. Guidance properties of the fibers used had
been shown on 2D substrates in previous studies, but needed to be confirmed for the 3D
scaffolds with appropriate cell types in vitro.
4. Integration of the 3D scaffold into an implantable tube to be tested in vivo. This device
was intended to allow for long-term storage and should be quickly available if needed.
5. Biological testing of the conduit: the efficacy to support peripheral nerve regeneration
was to be tested in vivo using the sciatic nerve lesion model in rats. Regenerative success
was to be evaluated:
a. On functional level with behavioral experiments, i.e. toe spread analysis and grip
strength test
b. On physiological level with electromygraphical recordings from the foot
musculature after stimulation of the nerve
c. By preparation of immunohistological stainings of the regenerated nerve tissue
and determination of muscle weights in the affected limb.
The experiments associated with the above mentioned milestones one to three are presented in
chapter II of this thesis, while the respective experiments for milestones four and five are
presented in chapter III.

1 Introduction
9
Chapter I
Theoretical background on peripheral nerve repair and design
strategies for artificial implants
1 Introduction
Peripheral nerves transmit motor, sensory and autonomic information between the central nervous
system (CNS) and the rest of the body. If a peripheral nerve is severed, e.g., by an injury to the
face or limbs, these functions are lost. However, in contrast to the mostly non-regenerative
processes after spinal cord lesions, axons in the peripheral nervous system (PNS) are able to
regenerate, and the long-term functional outcome of peripheral nerve injury depends on the
severity of the trauma (Sunderland, 1951). The cell somata of PNS neurons are localized in the
ventral horn of the spinal cord (motor neurons), the dorsal root ganglia (DRG; sensory neurons),
or in the sympathetic chain ganglia close to the spinal column (neurons of the autonomic nervous
system). Damage to mammalian peripheral nerve fibers activates a growth program in the
axotomized neuronal cell bodies such that their axons regenerate, provided they find a growth-
permissive substrate. The ideal growth-promoting substrate is provided by the distal segment of a
lesioned peripheral nerve.
For this reason, peripheral nerves can be surgically repaired after simple transection injuries by
reconnecting the individual proximal and distal nerve fascicles. However, when the gap between
disrupted nerve stumps is too large, the surgeon is required to transplant a segment of an
autologous nerve that is taken from elsewhere in the patient (Pabari et al., 2010). Since sensory
nerves such as the sural or peroneal nerve are often used for such procedures, the inevitable
consequence is the loss of sensory function. Although sensory nerves are routinely transplanted
even for the repair of motor fibers, experimental data indicate that sensory grafts are less suitable
than motor nerves (Brenner et al., 2006). The limitation of available donor material for
autografting and the additional risk of donor site morbidity prompted the need for alternatives to
the autologous nerve transplantation. Although several alternative tissues have been investigated
for their potential to support PNS repair, such as muscles or veins (Meek et al., 2004), a major
research focus has been the development of a bioengineering approach to design artificial nerve
conduits. Recent advances include internal topographical features that assist in the guidance of
axonal regeneration. Among these developments, the use of polymeric nanofibers has provided
one of the most promising templates for the purpose of peripheral nerve repair (Deumens et al.,
2010; Gu et al., 2011).

Chapter I
10
2 Physiological requirements of artificial nerve bridges
2.1 Peripheral nerve regeneration
To identify the requirements that artificial nerve constructs must meet in order to substitute
autologous transplants, a good understanding of peripheral nerve regeneration is important.
Peripheral nerve injury invariably causes degeneration of the distal nerve stump. Since axons are
disconnected from their cell somata they are eventually degraded. Cytoskeleton and cell
membranes are broken up into their molecular constituents. The PNS glia, the Schwann cells,
shed their myelin. This anterograde degeneration of the distal nerve segments is referred to as
Wallerian degeneration. It coincides with the infiltration of hematogenous macrophages, which
clear myelin fragments and neuronal debris. When Schwann cells lose contact with living axons
they dedifferentiate and proliferate within endoneurial tubes of the nerve. In the process, the
aligned Schwann cells form the so-called bands of Büngner, which provide an excellent growth
substrate for axonal regeneration. At the same time, retrograde signals activate a physiological
program of regeneration in the neurons. Thus, growth cones are formed by severed axons in the
proximal nerve stump (Figure 1a, b).
The success of this regenerative response depends on the extent of the injury, especially on the
maintenance of connections between the proximal fiber fascicles and the endoneurium of the
severed, distal segments. This kind of injury (called axonotmesis) can be treated conservatively
because many injured neurons survive, and regenerating axons elongate in contact with the bands
of Büngner (Sulaiman et al., 2005). Injured peripheral nerves, especially the basement
membranes, provide an excellent growth substrate for axonal regeneration, where neurite
extension reaches velocities of several millimeters per day. Subsequently, the Schwann cells re-
myelinate regenerated axons (Boyd and Gordon, 2003). If axons reach their peripheral targets
they may form new synapses or end organs, and physiological function can be restored (Figure
1c, d). When the continuity of the endoneurial sheaths is disrupted, if fiber fascicles still remain
connected via the perineurium or epineurium (neurotmesis) recovery is also possible without
surgical intervention. After complete transaction, the stumps of the elastic nerves retract, and fiber
fascicles must be re-aligned and sutured (end-to-end neurorrhaphy) (Sunderland, 1951). Larger
nerves are supported by endogenous and exogenous blood vessels, and these, too, are surgically
restored. The problems described above may represent a microsurgical challenge, but do not call
for a tissue engineering approach. This is the case, however, when the injury causes larger gaps in
the nerve, where end-to-end suturing would lead to excessive tension, impair microvascular blood
flow, and result in scarring (Pabari et al., 2010). Any larger lesions are repaired with autologous
nerve grafts (Deumens et al., 2010) because no better substrate for nerve regeneration is known
than the injured peripheral nerve itself.

2 Physiological requirements of artificial nerve bridges
11
Consequently, natural peripheral nerves provide the ideal template for biomimetic designs of
artificial nerve conduits. Ideally, such constructs would also include molecular signals that guide
and support regeneration in the natural environment. Since research on molecular processes
associated with peripheral nerve regeneration has accumulated an immense corpus of data over
the last half century, I can only mention those pertinent signal transduction pathways that have
been considered in the functionalization of nerve constructs.
Figure 1 Peripheral nerve degeneration and regeneration.
(a) Neurons of the PNS are located in the ventral horns of the spinal cord (motor neurons), the dorsal
root ganglia (sensory neurons), or the sympathetic chain ganglia (autonomic nervous system). Axons
are myelinated by Schwann cells. (b) After nerve injury myelin sheaths and axons degenerate distal
to the lesion site. Schwann cells proliferate, and macrophages remove the debris of degenerating
fibers. At the lesion site, neurons form axonal growth cones. (c) Axons are able to regenerate along
longitudinal bands of glia and ECM. Subsequently, Schwann cells remyelinate the new axons. (d) After
complete transaction proximal and distal nerve stumps retract. Frequently, when axon sprouts fail to
cross the site of injury, a neuroma forms. The distal part of the nerve and muscle fibers that are no
longer innervated become atrophic

Chapter I
12
2.2 Secreted signals in peripheral nerve regeneration
Axonal injury causes depolarization and action potentials, resulting in an influx of Ca2+
ions and
the activation of an intracellular signal transduction machinery, which regulates formation of the
growth cone (Makwana and Raivich, 2005). Once axonal growth is initiated, cell survival and
continued axonal regeneration depend on the supply of trophic physiological growth factors. Most
of these are synthesized by Schwann cells and, to a lesser extent, by macrophages and possibly
fibroblasts. Provided that the different subpopulations of peripheral neurons are maintained,
interactions with the extracellular matrix (ECM) and with other cells guide elongation of the
nerve fibers. Attempts have been made to incorporate molecular signals that mediate these
processes in tissue engineered nerve grafts (Gu et al., 2011).
2.2.1 Regeneration signals that activate axonal growth
Following the immediate consequences of injury-induced depolarization, the effect of growth
factors sets in. Perhaps the single most important signaling molecules in this category are ciliary
neurotrophic factor (CNTF) and nerve growth factor (NGF). The neurocytokine CNTF is strongly
expressed by myelinating Schwann cells. Although expression of this factor declines after the
injury, it seems to be released from the damaged cells and triggers neuronal regeneration
(Sendtner et al., 1992). Another important signal is NGF, a member of the neurotrophin family,
the expression of which is strongly reduced immediately after the lesion (Heumann et al., 1987),
and the absence of which may be important for the initiation of the regenerative response
(Shadiack et al., 2001). Similar observations were made for insulin-like growth factor-I (IGF-I)
(Kanje et al., 1991). These and related factors at later stages support survival and regeneration of
injured neurons.
2.2.2 Survival factors for the damaged nerve cells
Even in the PNS, with its high regenerative potential, a large percentage of neuronal cell death
can occur after injury, especially if the lesion occurs close to the cell soma (Deumens et al.,
2010). A number of growth factors were found to be upregulated in regenerating peripheral
nerves and, as shown by blocking their activity in vivo, to be physiologically neuroprotective.
Thus, NGF, secreted by Schwann cells, is a neurotrophic factor for sensory and autonomic
neurons. Leukemia inhibitory factor (LIF), CNTF and glial-derived neurotrophic factor (GDNF)
support motoneurons. Related molecules, especially the neurotrophins brain-derived neurotrophic
factor (BDNF), neurotrophin-3 (NT-3), NT-4/5, the neuropoietic cytokines, e.g. interleukin-6 (IL-
6), and fibroblast growth factors (FGF-1, FGF-2) have also been found to enhance survival of
PNS neurons (Heumann et al., 1987; Sendtner et al., 1990; Huang and Reichardt, 2001). Most of
these molecules have been employed in attempts to support regeneration of peripheral nerve (PN)
injury. A complementary strategy to the supply of survival factors would be to interfere with

2 Physiological requirements of artificial nerve bridges
13
apoptotic pathways that are also initiated by nerve injury. To my knowledge this has not been
tested so far in the context of repair strategies with artificial nerve constructs.
2.2.3 Promoters of axonal regeneration
Many investigations have been conducted in search of regeneration-promoting molecules in vitro
and in vivo. Important candidates that were found to be involved in the physiological context are
IGF-I and IGF-II (Glazner et al., 1993; Emel et al., 2011), both of which are also beneficial when
given exogenously. BDNF and GDNF are important growth promoting
factors for motor neurons, especially in the case of chronic deafferentiation (Sendtner et al., 1990,
1992; Boyd and Gordon, 2003). Although a sufficient supply of these factors in vivo renders
additional pharmacological application useless to the injured nerve (Gordon, 2010), they may be
important tools for artificial nerve grafts. NGF is a powerful neuritogenic factor for sympathetic
neurons (Hoyle et al., 1993). It must be noted that neurotrophins can also have neurotoxic effects
when they act via the common low affinity NGF (p75) receptor instead of the specific receptor
tyrosine kinases (trkA, trkB, trkC) (Boyd and Gordon, 2003). Receptors for several GDNF family
members (c-ret and co-receptors GFRa2, GFRa3) are expressed in small unmyelinated sensory
neurons in the DRG and in sympathetic ganglia. Their signals enhance regeneration of
sympathetic and nociceptive neurons (Trupp et al., 1995; Keast et al., 2010).
2.2.4 Modulators of the Schwann cell response
Although the effect of neurons on Schwann cells and other non-neuronal cells in the nerve are
mediated to a large extent by cell surface contacts, soluble factors play a major part. The most
important inducer of Schwann cell differentiation and myelination is neuregulin-1 of the glial
growth factor (GGF) family. GGFs are expressed by neurons and have various glia and muscle
cells as recipients. In the PNS, neuregulin-1 can drive the entire pathway of Schwann cell
differentiation including axonal myelination (Nave and Salzer, 2006). Neuregulin receptors on the
Schwann cell are ErbB2/ErbB3 heterodimers, where the ErbB3 subunit binds the ligand while
ErbB2 has a kinase activity, which initiates the intracellular signaling cascade (Citri et al., 2003).
Peripheral nerve injury induces expression of neuregulins and their receptors (Carroll et al.,
1997). Other soluble signals include nuclear receptor ligands such as thyroid hormone, estrogen,
and retinoic acid. Triiodothyronine, estrogen, and progesterone enhance axonal regeneration after
peripheral nerve injury, though it is questionable whether these hormones are endogenous signals
in the process (Panaite and Barakat-Walter, 2010). Retinoic acid signaling, which is activated by
sciatic nerve crush (Zhelyaznik et al., 2003; Zhelyaznik and Mey, 2006) promotes peripheral
nerve regeneration (Taha et al., 2004) and seems to be a crucial downstream effect of the action of
neurotrophins (Corcoran and Maden, 1999), especially for NGF- and NT-3-dependent sensory
neurons (Corcoran et al., 2000). However, retinoid receptor expression is most prominent in
Schwann cells and macrophages. The transcription factor Krox20, which is an important regulator

Chapter I
14
of peripheral myelination, is activated by neuregulin and retinoic acid via a retinoid X receptor
(Latasa et al., 2010).
2.2.5 Signals for vascularization
Members of the large, heparin-binding fibroblast growth factor family, especially FGF-1 and
FGF-2 are strong promoters of angiogenesis. Another important paracrine factor in angiogenesis
is vascular endothelial growth factor (VEGF), which acts downstream of the FGFs, but also has
parallel, independent effect (Pola et al., 2004; Murakami and Simons, 2008). In addition to these
primary regulators, synergistic interaction with other factors, including granulocyte colony
stimulating factor and platelet-derived growth factor, influence blood vessel formation in
regenerating peripheral nerves (Pan et al., 2009).
2.3 Surface-bound signals
If artificial nerve implants are to mimic the physiological stimuli that initiate and maintain
regeneration in vivo, the most promising molecular candidates would be extracted from this list of
secreted molecules for intercellular communication. In addition, since axonal growth cones
advance only in contact with surfaces, regeneration and the infiltration of growth-promoting cells
depend on the presence of permissive substrates, which interact via surface-bound signals with
cell membrane receptors. Such substrates are (1) the ECM and (2) the plasmalemma of other
cells.
2.3.1 Extracellular matrix
The predominant growth-permissive surface that renders peripheral nerves such an ideal growth
substrate is the basal lamina laid down by the Schwann cells. It consists largely of laminin and
collagen IV (Koopmans et al., 2009). Laminin and collagen are therefore the most frequently used
ECM proteins in cell culture and three-dimensional constructs (see below). Nerve fibers are
grouped in distinct fiber fascicles, which are enclosed by a perineurial matrix (endo-, peri- and
epineurium). This matrix consists of fibrillar collagens type I, III, and V, and of fibronectin,
which serves as a connecter between cell surface receptors and collagen. The cellular receptors
for ECM molecules are heterodimeric transmembrane proteins called integrins (Geiger et al.,
2001). Axonal growth cones express integrins that are activated by specific epitopes of these
ECM molecules.
Functional integrin receptors consist of one a- and one b-subunit. On one hand, they interact with
the actin cytoskeleton and, on the other hand, they can activate numerous intracellular signaling
pathways. Among the two dozen integrins known today the β1-, αv- and α4-containing integrins
are particularly important as receptors of peripheral nerve ECM. They have multiple functions,
including the regulation of cell adhesion, cell motility, and differentiation (Hynes, 1992; Geiger et
al., 2001). A synergism of neuregulin-1, expressed by axons, and the laminins in the basal lamina

2 Physiological requirements of artificial nerve bridges
15
surrounding the Schwann cells is crucial for regulation of Schwann cell differentiation and
myelination (Nave and Salzer, 2006).
2.3.2 Surface-mediated cell–cell interactions
Several classes of molecules mediate communication between axons, Schwann cells, and other
cells in the PNS. Since the late 1970s the IgG-domain containing cell adhesion molecules (CAM)
are known, especially for their role in axonal growth. Important representatives are L1/Ng-CAM,
N-CAM and transient axonal glycoprotein-1, which appear to support axon fasciculation and
axonal growth on the surface of non-neuronal cells, including Schwann cells and fibroblasts
(Bixby et al., 1988; Martini and Schachner, 1988; Martini, 1994; Soares et al., 2005). Related
myelin-associated glycoprotein and protein zero are present in myelin membranes and are
important during myelination.
Cadherins are Ca2+-binding adhesion molecules that mediate homophilic interaction between
cells. Cadherins are particularly important for selective fasciculation of regenerating axons. For
instance, E-cadherin mediates attachment of unmyelinated sensory fibers and stabilizes glial
network (Hasegawa et al., 1996), and N-cadherin mediates axon–Schwann cell interactions during
regeneration (Bixby et al., 1988; Thornton et al., 2005).
A lot of research has been done on transcriptional changes and intracellular pathways that
correlate with axonal regeneration. The intricacies of lesion-induced intracellular signaling can
only concern us here in so far as these pathways have been considered as targets for
pharmacological intervention. Suffice it to say that Ras/ERK and PI3K cascades are primarily
involved, and that Rho–type GTPases (RhoA, Rac, Cdc42) and a panel of phosphorylation-
dependent transcription factors (most importantly STAT-3, ERK), determine neuronal survival,
the regenerative response of the growth cones, and also are involved in the glial responses
(Makwana and Raivich, 2005). A considerable overlap and cross-talk between the intracellular
signaling pathways of all soluble and surface-bound signals has been observed and is far from
understood.
2.4 Desired properties of the scaffold
From the description above I can deduce some functions that artificial implants must fulfill if they
are to promote nerve regeneration in vivo. A general requirement of biocompatibility, which
applies to materials implanted anywhere in the body, demands that the implanted construct must
not induce inflammatory reactions or tumor formation and is not rejected or encapsulated by scar
tissue.
2.4.1 Mechanical Properties
In addition to biochemical signals, mechanical properties are very important because irritation of
the tissue, e.g., due to stiffness of the material, may cause inflammation or fibrosis. The basic
design of virtually all artificial nerve guides is a flexible tube, either hollow or filled with interior

Chapter I
16
structures for nerve guidance. The choice of the material for the tube, the dimensions of its wall
and lumen, as well as the filling determine its biomechanical properties. Implanted devices should
have a similar degree of flexibility as the natural peripheral nerve, while at the same time they
must be stable enough not to collapse. The Young’s modulus of various mammalian nerves has
been determined, so present day scaffolds for implantation are mechanically compatible with the
surrounding tissue (Deumens et al., 2010; Gu et al., 2011).
2.4.2 Biodegradability
It is desirable that the bridge between nerve stumps remains in the body until axonal regeneration
is completed. Studies with earlier implants, which were made of nondegradable materials such as
silicone, showed some success, but also revealed disadvantages. The long-term presence of the
material can elicit an inflammatory foreign body reaction. In several human patients,
complications with silicon tube implants required follow-up surgery to remove the implants
(Dahlin and Lundborg, 2001). Thus, recent research studies and most clinical applications
concentrate on nerve scaffolds that are gradually degraded without the release of toxic products
(Meek and Coert, 2008b; Meek and Jansen, 2009).
2.4.3 Permeability for growth factors and gas exchange
Since regeneration and physiological function of the restored nerve need the exchange of gases,
water, and biological signals such as hormones or neurotrophins the artificial implant must be
sufficiently permeable. On the other hand, osmotic influx and swelling might reduce its lumen,
thereby exerting pressure against
the regenerating fibers (de Ruiter et al., 2009). This must be avoided.
2.4.4 Guidance of axon growth and Schwann cell migration
The central function of the implant, remains, of course, the guidance of regenerating axons from
the proximal to the distal nerve segment (Dalton and Mey, 2009). Consequently, the growth cones
have to recognize physical guidance structures, which promote axonal elongation by activating
specific receptors on the cell membrane. While biochemical signaling between cells and the
scaffold surface is crucial for any interaction to take place, the physical structure of the implant
must not only allow infiltration of cells and growth cones but should guide regeneration in the
longitudinal direction toward the distal end of the implant. Polymeric nanofibers oriented in
parallel can provide ideal scaffolds in this regard. Finally, axons have to enter into the distal
fascicles of the existing nerve, which have undergone Wallerian degeneration but still lead to the
peripheral targets that are to be innervated again. Thus, an additional requirement is that the
growing axons are not trapped inside the implant.
As mentioned above, Schwann cells are indispensable not only for the process of axonal
regeneration but also for the subsequent myelination and maintenance of physiological function.

3 Design strategies for artificial nerve bridges
17
Similar molecular interactions as with neurons should therefore allow migration of Schwann cells
from the host into the nerve bridge.
3 Design strategies for artificial nerve bridges
With respect to the properties of the implant that will determine its success, two aspects can be
distinguished: first, the physical structure of the scaffold (see Chap.I 3), and second, the
biochemical functions of its materials (see Chap.I 4). The basic structure of all artificial nerve
bridges is a tubular graft to connect the proximal with the distal peripheral nerve stump (Figure
2).
3.1 Hollow tubes and multichannel nerve conduits
In the simplest case, a hollow cylindrical tube with a single lumen is used. The original purpose of
this design was not to replace an entire nerve segment, but only to a bridge a short gap between
the proximal and distal nerve stumps, then direct ligature of the respective fiber fascicles might
cause a strain that would interfere with the natural process of regeneration. Different techniques
have been employed for fabrication of the tube, such as melt extrusion (Chiono et al., 2009),
particle leaching (Bian et al., 2009), injection molding, and electrospinning: a variety of materials
were also tested (Meek and Coert, 2008b; Gu et al., 2011).
Since a hollow tube is the simplest device conceivable as a nerve bridge, this type of implant was
soon also tried as a long-distance connector when pieces of nerve had to be replaced. Of all
designs, the empty tube has been tested by far most often in vivo (Figure 2a and 3a, b). The tube
may consist of natural (Chap.I 4.1) or synthetic materials (Chap.I 4.2) or composites. Although
case studies with a few patients have been undertaken using more sophisticated designs (Inada et
al., 2007; Fan et al., 2008), hollow tubes constitute the only nerve guide design that has been used
in larger clinical trials so far (Meek and Coert, 2008b; Gu et al., 2011). The many cell culture and
animal experiments that led to the clinical studies will not be reviewed here.
3.1.1 Empty nerve conduits
The non-degradable tubes that were first implanted in patients, consisted of silicon (Lundborg et
al., 1997b, 2004) or polytetrafluoroethylene (Stanec and Stanec, 1998; Pitta et al., 2001). In one
large study, 26 patients had suffered injuries to the median or the ulnar or to both nerves and
received tubular silicone implants as nerve bridges. Based on sensory and motor function and
pain, in 19 cases the outcome of the operation was rated very good or good; however, the implant

Chapter I
18
had to be removed in seven cases because of discomfort caused by the silicone tube (Braga-Silva,
1999). There are divided opinions about whether non-degradable implants are advisable because
they may cause a foreign body reaction, and whether the need to remove them in a subsequent
operation after regeneration is risky (Lundborg et al., 1997b). However, even Lundborg and
colleagues who have conducted successful clinical trials with silicon tubes state a preference for
Figure 2 Design strategies for artificial nerve bridges.
(a) Empty tube used for bridging gaps between proximal and distal nerve stumps; (b) tubular
implant with multiple intraluminal channels (e.g., de Ruiter et al., 2008; Yao et al., 2010); (c) aligned
microfibers as longitudinal guidance structures (e.g., Kim et al., 2008; Ribeiro-Resende et al., 2009);
(d) hydrogel filling of the conduit (e.g., Inada et al., 2004; Labrador et al., 1998); (e) sponge filled tube
(e.g., Inada et al., 2004; Toba et al., 2001); (f) longitudinally oriented pores within the scaffold (e.g.,
Ceballos et al., 1999; Möllers et al., 2009); (g) modification of the inner side of the tube with bioactive
coating (e.g., Chew et al., 2007; Shen et al., 2010)

3 Design strategies for artificial nerve bridges
19
biodegradable materials, provided their degradation does not cause further complications (Dahlin
and Lundborg, 2001). Thus, to date, biodegradable materials are the preferred solution. Three
types of such tubes have been approved for implantation in humans and are commercially
available. They have been tested in several hundreds of patients.
NeuraGen, consisting of type I collagen, is produced by Integra Neuroscience (www.integra-
ls.com). In the largest study so far, NeuraGen implants were used mostly for bridging sensory
nerves of the arm. Twenty-six patients were evaluated quantitatively to assess functional recovery
of nerve transmission. In 45 % of the patients an improvement of sensory functions was seen
(Wangensteen and Kalliainen, 2010). In a number of small clinical trials and case studies, the
collagen implant has demonstrated its usefulness (Ashley et al., 2006; Lohmeyer et al., 2007;
Farole and Jamal, 2008; Taras and Jacoby, 2008), though occasional failures were also reported
(Moore et al., 2009). In this latter report, another nerve conduit, Neurotube, was also tested,
though without better outcome.
Neurotube is a poly(glycolic acid) (PGA) implant, produced by Synovis Life Technologies
(www.synovismicro.com). It has been tested in several clinical studies. In one of these, a group of
46 patients received Neurotube implants for lesions of nerves innervating the hand. The results
showed good to excellent outcome in 74 % of the cases, which was not statistically different from
a control group who received autologous nerve transplantations (Weber et al., 2000). Other
successful studies with the PGA tubes have been published, with repair of median, ulnar, and
facial nerves being reported (Navissano et al., 2005; Donoghoe et al., 2007; Agnew and
Dumanian, 2010).
The third commercially available nerve scaffold is Neurolac, produced by Polyganics
(www.polyganics.com). It consists of a poly(lactic acid)/poly(ε-caprolactone) blend (PLA/PCL).
So far, two clinical studies and case reports have been published, with a total of 36 patients.
Although in the first report from 2003 no nerve regeneration was found, the second study reported
functional regeneration similar to results with autologous nerve transplants (Bertleff et al., 2005).
Again, failures to achieve recovery of sensory functions were reported, e.g., with digital nerve
reconstruction in the foot (Meek et al., 2006).
3.1.2 Multichannel devices
Without changing the basic design, tubular implants have been constructed to incorporate several
intraluminal channels that can accommodate different fiber fascicles within a peripheral nerve
(Figure 2b and 3c, d). Multichannel implants with up to seven compartments were produced using
a similar injection-molding technique as for the implant with a single lumen (de Ruiter et al.,
2008; Yao et al., 2010). On the level of fiber fascicles, the multichannel devices appeared to
reduce mixing of regenerating axons. Necessarily, these constructs reduced the available space
within the implant; and when numbers of regenerated fibers and behavioral improvement were

Chapter I
20
evaluated, the multichannel constructs were no better than hollow tubes (Yao et al., 2010). An
alternative construction design achieved the similar effect of longitudinal divisions of the
otherwise empty tube by integrating one or a few films into the construct. When tested in rat tibial
nerves, tubes with one film, i.e., just two compartments, were better than implants with none or
three films (Clements et al., 2009).
In animal experiments, many studies were performed with hollow tubular implants to assess the
biocompatibility of the material. Most often, grafts were tested as bridges of the sciatic nerve, and
by far most experiments were done with rats where nerve lesions rarely exceeded 20 mm. When
short nerve gaps are bridged with tubular implants, a fibrin matrix will fill this distance and allow
infiltration of Schwann cells from the nerve stumps. These cells can then form orientated bands of
Büngner, which may guide regenerating axons. Experiments with rabbits, cats, dogs, and
monkeys indicate that internal guidance structures are needed for larger cell-free implants (Brown
et al., 1996; Matsumoto et al., 2000; Ding et al., 2010). An extensive list of animal studies with
further references is given in a recent review (Shin et al., 2012). The construction of three-
dimensional guidance structures remains a technical challenge, where polymeric nano- and
microfibers take center stage.
Figure 3 Examples of artificial nerve guides that were successfully tested in animal
experiments.
(a) SEM images of a nerve tube consisting of electrospun PLGA/PCL microfibers; scale bar 500 µm
(Panseri et al., 2008). (b–d) Collagen conduits with one or multiple interior channels were fabricated
from molds with insertion of stainless wires; scale bar 1 mm (Yao et al., 2010). (e) Composite nerve
conduit of several hundred PCL filaments each having six leaflets in cross-section (insert); scale bar
150 µm (Whitlock et al., 2009). (f) Poly(glycolic acid) tubes filled with collagen sponge were
successfully used to bridge 80 mm gaps of canine peroneal nerves (Nakamura et al., 2004). (g)
Collagen scaffold produced with directional freezing; longitudinal sections demonstrate oriented
channels; extensive fenestration between adjacent channels can also be seen; scale bars 50 µm
(Möllers et al., 2009).

3 Design strategies for artificial nerve bridges
21
3.2 Structured implants with polymeric fibers
One successful strategy to provide such guidance structures in the longitudinal direction of the
implant is the use of polymer fibers (Figure 2c and 3e). These can be produced by a number of
different techniques such as drawing, template synthesis, phase separation, self-assembly, and
electrospinning. Historically, large diameter fibers were tested first. Lundberg and coworkers
filled silicone tubes with eight longitudinally oriented polyamide filaments of 250 µm diameter as
implants. Across a 15 mm gap in the rat sciatic nerve, this device was a substantial improvement
compared with hollow implants (Lundborg et al., 1997a). To mimic the natural situation, many
more and thinner fibers would provide topological cues and induce the formation of many
longitudinal strands of Schwann cells. Cell culture experiments demonstrated that small caliber
fibers (5–30 µm) had a stronger effect on the orientation of growing neurites than thicker fibers
(500 µm) (Smeal et al., 2005). In the context of peripheral nerve regeneration, the technique that
has been used most frequently for the manufacturing of small diameter fibers is electrospinning.
3.2.1 Electrospinning of nano-/microfibers
As a process to produce continuous fibers with very small diameters, electrospinning has been
known for more than a century. During the last two decades, the method has become increasingly
popular in the field of bioengineering because the method is easy to use, very flexible with respect
to fiber configuration in situ, and a great range of different materials can be used (Teo and
Ramakrishna, 2006). Biodegradable polymers that have been spun to nanofibers, tested in vitro,
and implanted in the nervous system will be discussed below. The typical electrospinning set-up
in a research laboratory consists of a high-voltage power supply (up to 30 kV), the spinneret (a
syringe with a flat tip needle), and collector electrodes (Figure 4). To induce formation of fibers, a
high voltage is applied via the spinneret to the polymer solution. This causes electrostatic
repulsion within the charged solution, which is stronger than its surface tension, such that a jet
erupts from the spinneret. At some distance, the jet enters a stage of bending instability, is
stretched under electrostatic forces toward the grounded target electrode, and the solvent
evaporates. With a low flow rate, polymer solution is pumped through the spinneret to ensure
continuous formation of fibers, which accumulate on the target. On a single plate electrode, used
as a target, mats of randomly coiled fibers accumulate. Several electrospinning devices were
tested in order to assemble nanofibers that are aligned in a parallel orientation. Rotating drums in
different configurations were devised by several groups (Matthews et al., 2002; Chew et al., 2005;
Kim et al., 2008). Alternatively, two parallel bars can be used as target electrodes, such that
parallel fibers accumulate between them (Schnell et al., 2007; Klinkhammer et al., 2009). Using
such devices, parallel fibers of diameter from less than 100 nm to more than 5 µm can be
produced from various synthetic polymers. A good review with drawings of various devices has
been written by Teo and Ramakrishna (Teo and Ramakrishna, 2006).

Chapter I
22
Electrospun micro- and nanofibers have high surface-to-volume ratios and provide growth
substrates for many neural cell types (Gerardo-nava et al., 2009; Bockelmann et al., 2011). Films
of electrospun fibers have been used to produce tubular implants and to subdivide nerve conduits
into longitudinal compartments (Clements et al., 2009). The specific advantage of nanofibers is,
of course, their property as individual guidance structures (Lietz et al., 2006; Kim et al., 2008). In
one study, PGA microfibers were incorporated in a chitosan tube as implants to bridge 30 mm
gaps in the sciatic nerve of beagles. The dogs, which were investigated after 6 months, recovered
function of the operated nerves. Skeletal muscles were re-innervated, and the artificial constructs
were completely degraded within the half year time-frame of the study (Wang et al., 2005). A
different research team stacked films of parallel fibers consisting of poly(acrylonitrile-co-methyl
acrylate) (PAN-MA) inside a polysulfone tube. These constructs were implanted in rats to bridge
sciatic nerve gaps of 17 mm. Sixteen weeks after surgery, target muscles were found to be
innervated again, and regeneration of sensory and motor nerve fibers could be demonstrated (Kim
et al., 2008). This represented a significant progress because the scientists incorporated oriented
nanofibers in a three-dimensional design. The largest distance of nerve repair has been reported
by Matsumoto, Toba and coworkers, who were able to bridge an 80 mm gap of the peroneal nerve
in dogs (Matsumoto et al., 2000). Their conduits consisted of tubes of PGA/collagen blend, filled
with laminin-coated collagen fibers. Regeneration was assessed histologically,
electrophysiologically, and with behavioral testing. Very sophisticated guidance structures have
Figure 4 Electrospinning of aligned fibers.
The electrospinning set-up used by the authors consists of a high voltage power supply (e.g., 20 kV),
the spinneret (a syringe with a flat tip needle) connected to a pump (flow rate e.g., 1 mL/h) and
parallel bars as target electrodes to collect parallel fibers (Schnell et al., 2007; Klinkhammer et al.,
2009). Another frequently used device to assemble aligned fibers is the rotating drum (Matthews et
al., 2002; Chew et al., 2005; Kim et al., 2008). Scanning electron microscopy images show
PCL/collagen fibers collected between parallel bars (Gerardo-nava et al., 2009)

3 Design strategies for artificial nerve bridges
23
been developed by Schlosshauer’s group, who also tested functionalization with NGF,
transforming growth factor-b1 (TGFb) and laminin in their experiments. Oriented PCL filaments
were made by melt extrusion using nozzles with six-leaf cross-sections. This resulted in yarns of
several hundred filaments, each with six longitudinal grooves (Figure 3e), which caused the
alignment of Schwann cells, “artificial bands of Büngner” (Ribeiro-Resende et al., 2009). As also
observed by others (Schnell et al., 2007), the topographic effect of the substrate on glia cells
indirectly affected the orientation of growing axons that need not have any contact with the fibers
themselves.
3.2.2 Self-assembling peptide scaffolds
The manufacture of self-assembling nanofibrous scaffolds is based on repetitive peptides that
consist of alternating hydrophilic and hydrophobic amino acids and spontaneously form stable b-
sheet structures. The self-assembling process takes place when the aqueous peptide solution is
introduced to a physiological salt-containing solution (i.e., saline, tissue culture media,
physiological solutions, or body fluids such as cerebrospinal fluid) and is mediated by non-
covalent bonds, such as van der Waals forces, hydrogen bonds, and electrostatic forces. As a
result of hydrophobic interactions, different b-sheets pack together and form double layered β-
sheet nanofibers with diameters of about 10 nm, much smaller than typical electrospun nanofibers
(Gelain et al., 2007; Cao et al., 2009). Scaffolds based on self-assembled peptides provide several
advantages over other biomaterials for their potential use as bridging materials for the nervous
system:
1. Their three-dimensional environment has dimensions that are similar to the
native ECM in peripheral nerves.
2. The peptides can be degraded into natural L-amino acids that are nontoxic and
may be recycled by surrounding cells.
3. They elicit only a minor, if any, inflammatory or immune response.
4. Modifications at the single amino acid level are possible, and various functional
motifs can be added to promote neurite outgrowth.
5. The self-assembly process takes place under physiological conditions without
the need of temperature changes (Gelain et al., 2007, 2010).
Several cell culture studies demonstrated that scaffolds made via self-assembling peptides provide
permissive substrates for cell attachment and growth as well as support for extensive neurite
outgrowth and synapse formation (Holmes et al., 2000; Silva et al., 2004; Ellis-Behnke et al.,
2006), including the migration of Schwann cells and axonal regeneration from DRG neurites
(DRG) (McGrath et al., 2010). Fewer reports have been published where such scaffolds were used
in the nervous system in vivo. In one study of CNS repair, a knife wound in the visual system of

Chapter I
24
2-day-old hamsters was successfully treated by injection of peptide solution into the lesion site.
Here self-assembling nanofiber scaffolds provided a permissive substrate for axonal regrowth,
and visual function could be restored (Ellis-Behnke et al., 2006). Peptide scaffolds implanted after
spinal cord dorsal column transection injuries of rats integrated very well with the host tissue and
supported the infiltration by blood vessels and axons (Guo et al., 2007). To my knowledge, only
one in vivo study focused on the potential of self-assembly nanofibers to repair a peripheral nerve
(McGrath et al., 2010). In this report, a 10 mm gap of the rat sciatic nerve was successfully
bridged with a tubular conduit filled with a self-assembling peptide nanofiber scaffold. The
distance of axonal regrowth 3 weeks after implantation was significantly enhanced by these
conduits when compared to control implants filled with alginate/fibronectin hydrogels, and
recovery of gastrocnemius muscle function was also better. However, grafted Schwann cells
improved the efficacy of the implants, and autologous nerve grafts still yielded the best outcome
(McGrath et al., 2010). Although these investigations demonstrated that self-assembling peptide
scaffolds can serve as permissive substrate in regenerative medicine, the nanofibers of these
constructs lack orientation. This is a desired property for long distance guidance within an
artificial nerve bridge, which is more easily achieved with the electrospinning method.
3.3 Structured implants with gels and scaffolds
A completely different approach for the internal structure of implants consists in gel scaffolds
with a narrow mesh of longitudinal pores or channels. Several laboratories used collagen
hydrogels to fill the lumen of different nerve guides, which themselves were made from a range
of biocompatible materials (Figure 2d). Based on histology and measurements of functional
recovery, such hydrogel fillings of collagen or laminin proved superior in comparison with saline-
filled implants. It is interesting to note that lower concentrations of collagen- or laminin-
containing gels (e.g., 1.28 mg/ml collagen, 4 mg/ml laminin) supported better functional recovery
than high gel concentrations (e.g., 1.92 mg/mL collagen and 12 mg/ml laminin) (Labrador et al.,
1998; Verdú et al., 2002).
Scientists from the University of Kyoto constructed a sophisticated nerve conduit that consists of
a polyglycolic acid tube, filled with a sponge of porcine collagen soaked with laminin (Figure 2e
and 3f) (Toba et al., 2001; Inada et al., 2004). Tubes of 3–4 mm inner diameter were implanted as
bridges into the peroneal nerves of beagle dogs. On the basis of histological and
electrophysiological evaluation, these implants were even superior to nerve autografts, although
some cases of neuromas occurred (Inada et al., 2004). Consequently, similar tubes were implanted
for repair in human patients. In two women, the frontal branch of the facial nerve was restored
with the construct. Five months after surgery both were able to lift their eyebrows symmetrically.
Electrophysiological tests showed recovery of compound muscle action potentials with normal
latency (Inada et al., 2007). This success was possible with the repair of small caliber nerves,

3 Design strategies for artificial nerve bridges
25
which also showed the best prognosis using hollow tubular implants. Nevertheless, the outcome
and the previous animal experiments, where bridges of up to 80 mm were implanted in dogs
(Toba et al., 2001; Siemionow et al., 2010), demonstrated that the inclusion of a collagen matrix
was a substantial advancement.
While these results are already very impressive, additional improvement is expected with the
design of gels with a fibrillar network that is oriented in the longitudinal direction of the nerve
guide (Figure 2f). One idea to achieve this is the application of strong magnetic fields that can
orient collagen or fibrin matrices. Such scaffolds promoted neurite elongation from chick DRG in
the desired direction (Dubey et al., 1999). In mice sciatic nerves, magnetically aligned collagen
gels supported regeneration and remyelination of axons over a 6 mm gaps better than control
tubes (Ceballos et al., 1999).
Another, very promising technique is controlled freeze drying of collagen suspensions (Figure
3g). When solvent is caused to freeze from one side of the solutions, collagen is displaced to the
side, thus resulting in walls of orientated guidance channels. The diameter of these longitudinal
pores can be controlled in the range of 20–120 µm. In vitro studies have already demonstrated
that such scaffolds can be easily infiltrated by Schwann cells, fibroblasts and regenerating axons,
which follow the desired orientation of the scaffold (Bozkurt et al., 2007, 2009; Möllers et al.,
2009). Three-dimensional constructs produced with this method are commercially available
(Matricel www.matricel.net) and have been tested successfully in animal experiments (Bozkurt et
al., 2012).
3.4 Implantation of cells with artificial nerve bridges
The closest thing to real nerve transplants are artificial constructs that are preseeded with growth
promoting cells, and many researcher believe that gaps longer than 30 mm will not be
successfully repaired unless supporting cells are included in the scaffold (Hood et al., 2009;
Deumens et al., 2010). These cells may derive from the host organism itself or from cultures.
They are intended to release physiological signals, many of which may be unknown and thus
cannot be mimicked with chemical modification of the implant material. Best suited for this
purpose are Schwann cells because they naturally populate the PNS, where they serve a number
of important functions during regeneration. As discussed above, Schwann cells secrete the basal
lamina that serves as axon guidance substrate, synthesize a host of necessary growth factors, and
even participate in the clearance of myelin debris. Later, they are required for myelination of the
regenerated fibers (Boyd and Gordon, 2003). Unfortunately, isolation of Schwann cells from a
peripheral nerve and expansion in culture are time-consuming and also carry the risk that culture
conditions alter the cellular phenotype.
Other cell sources have therefore been explored as alternatives. They include olfactory
ensheathing cells, embryonic stem cells, mesenchymal stem cells (Gu et al., 2011; Radtke et al.,

Chapter I
26
2011), and even cell lines such as the immortalized SCTM41 Schwann cells (Lago et al., 2009).
Genetic modification has been considered for implanting cells as a constant source of growth
factors. Most often, fibroblasts have been engineered with this intention, e.g., to produce
neurotrophins, CNTF and FGF-2 (Sørensen et al., 1998; Schmidt and Leach, 2003).
The majority of studies on PN regeneration, however, used Schwann cell primary cultures
(Guénard et al., 1992; Kim et al., 1994; Brown et al., 1996; Sørensen et al., 1998; May et al.,
2004; Radtke et al., 2005; Lohmeyer et al., 2007; Sinis et al., 2007). Unfortunately, the
implantation of any type of living cells is not without complications. First, the purity of the
primary culture is essential because the cells must not be contaminated, e.g., by fibroblasts in a
Schwann cell culture. Second, immune rejection of the cells is an issue. Heterologous
transplantation poses less of a problem between inbred strains of mice and rats, but it would
require immune suppression in humans. Ideally therefore, the cells would have to be taken from
the same patient who requires the PN implant. For a potential therapy of CNS lesions, an
interesting strategy is the preparation of olfactory ensheathing cells from the human olfactory
epithelium. In many physiological respects these glial cells are similar to Schwann cells (Kawaja
et al., 2009), and the cells could be prepared for autologous implantation. Olfactory ensheathing
cells are therefore being explored as a possibility to produce better implants also for peripheral
nerves (Radtke et al., 2011). Nonetheless, because of the fundamental difficulties of immune
rejection when foreign tissues are implanted, I consider the development of cell free artificial
nerve bridges as an important goal. Ideally, scaffolds of different lengths and diameters could be
produced in advance and stored to be implanted on demand.
4 Materials for artificial nerve conduits
Biocompatible materials are needed for the two essential components of the artificial nerve
bridge, i.e., the surrounding tube and its interior structure, the latter being either linear filaments
or a scaffold with longitudinal pores. The selected material will determine not only the
biochemical interaction between cells and graft but also the physical properties of the implant.
4.1 Natural materials
Many natural materials are hydrophilic and show excellent biocompatibility. Proteins and
carbohydrates of high molecular weight are extracted rather than synthesized chemically. On the
down side, when the material is derived from mammalian tissue there is a certain risk of disease
transmission and/or allergic reactions. Nevertheless, many biologically derived materials have just
the desired properties for applications in nerve repair.

4 Materials for artificial nerve conduits
27
4.1.1 Extracellular matrix proteins
The most straight forward approach is to use ECM material that gives structure to the peripheral
nerves in situ. Epineurium, perineurium and endoneurium consist largely of one family of ECM
proteins: collagen.
Collagen
Of all biologically derived material used in bioengineering, collagen is the most popular, not only
for PN conduits but for tissue reconstruction in many other applications. Collagens, which
account for more than a quarter of total body protein, are extracellular proteins with a
characteristic triple helix structure of three either identical or heteromeric a-chains (Koopmans et
al., 2009). Of at least 29 different forms, collagen I, III, and V form fibrils in the ECM of the
PNS. The non-fibrillar collagen IV is present in the basement membranes secreted by Schwann
cells. In vitro, collagen IV has superior qualities as a substrate for axonal growth, but the fibrillar
collagen I is more often used in scaffold production. As reviewed above, collagen has been used
to form tubular implants, e.g., NeuraGen (Ashley et al., 2006; Farole and Jamal, 2008; Taras and
Jacoby, 2008; Wangensteen and Kalliainen, 2010). It is also one of the materials frequently used
for interior guidance structures (Figure 3b–d, f, g): During matrix formation, collagen hydrogels
can be oriented with strong magnetic fields (Ceballos et al., 1999), and collagen sponges
combined with laminin (Toba et al., 2001; Inada et al., 2004) or chondroitin-6-sulfate
(Chamberlain et al., 1998) were very successful as nerve guides in animal experiments. Using the
directional freezing technique, scaffolds with longitudinally oriented pores were created and
tested for implantation in PNS and CNS (Bozkurt et al., 2007, 2009; Möllers et al., 2009). It is
also possible to produce collagen nanofibers by electrospinning (Matthews et al., 2002). Even
blends with other polymers (e.g., 25 % collagen, 75 % PCL) were significantly better for axon
guidance and Schwann cell migration than control fibers without collagen (Schnell et al., 2007).
Fibronectin
Axons and glia cells interact with the ECM via their integrin receptors. Although integrins can
directly bind to some epitopes on collagen, an important intermediate molecule between collagen
and the cells is the 440 kDa glycoprotein fibronectin, which is also part of the basement
membrane. It is primarily produced by fibroblasts. In addition, there is a soluble dimeric form of
fibronectin in the plasma, which together with fibrin has an important function in wound healing
(Pankov and Yamada, 2002). The biological activity of fibronectin has been shown to be strongly
influenced by the conformation of underlying surfaces (Dalton and Mey, 2009). Fibronectin
conduits have performed well in PN repair. For instance, tubular implants filled with a solution of
fibronectin, laminin, and fibronectin/laminin mixture permitted regeneration across an 18 mm gap
in rat sciatic nerves, and all of these ECM molecules promoted migration of Schwann cells into
the conduit (Bailey et al., 1993).

Chapter I
28
Laminin
This is the most important component of the basal lamina. Different laminins exist, all of which
are large glycoproteins (500–1,000 kDa) that consist of three polypeptide chains (α, β, and γ).
Applied in gels or solution together with collagen and other proteins, laminin enhanced nerve
regeneration in vivo and in vitro. In addition to the whole protein, integrin-activating peptide
sequences from laminin were identified that also mediate cell adhesion and growth. Well-
characterized sequences are those containing Arg-Gly-Asp (RGD, in one letter code) in laminin a-
chains (Hersel et al., 2003); YIGSR, located on the β1-chain; and IKVAV, which is present near
the C-terminal end of the α1-chain, close to a heparin-binding domain (Dalton and Mey, 2009).
When synthetic polymer fibers were functionalized with GRGDS, adhesion and speed of axonal
growth cones and of migrating cells were significantly enhanced (Bockelmann et al., 2011).
Laminins have already been used for a long time as substrates in cell culture experiments. The
commercially available Matrigel is a non-defined ECM solution derived from a mouse sarcoma
cell line. Its main component is laminin, but Matrigel also contains collagen IV, heparan sulfate
proteoglycans, and several growth factors. Used as a thermoreversible gel it is liquid at low
temperatures and solidifies at body temperature. As a filling in fiber containing implants, Matrigel
supported regeneration in the mouse sciatic nerve. Electromagnetic alignment of the material
further improved the outcome (Verdú et al., 2002; Lohmeyer et al., 2007).
4.1.2 Other natural proteins
Fibroin (Silk)
Silk is not immunogenic and has high tensile strength, toughness, and elasticity (Deumens et al.,
2010). Its core structural protein is fibroin, extracted from the cocoons of Bombyx mori
caterpillars (silkworm). The protein exhibited good biocompatibility with Schwann cells and
DRG (Yang et al., 2007). Fibroin was used for electrospinning of nanofibers (Madduri et al.,
2010b). A tubular fibroin construct, filled with about 20 longitudinally aligned fibroin fibers was
implanted as a 10 mm nerve bridge in rats and evaluated after 6 months. The detailed anatomical
evaluation demonstrated excellent results, though autologous nerve grafts were still superior
(Yang et al., 2007).
4.1.3 Polysaccharides
One risk with the implantation of foreign ECM proteins is development of scar tissue within the
regenerating area. This acts as a physical barrier to axonal elongation between the nerve stumps.
It appears that this problem is less likely with biocompatible polysaccharides.
Chitin and chitosan
The exoskeleton of arthropods consists largely of chitin, which is also present in the cell walls of
fungi. Second only to cellulose, chitin is the most abundant polysaccharide found in nature. The

4 Materials for artificial nerve conduits
29
monomeric building blocks of chitin are N-acetyl-D-glucosamine. Chitosan, a linear copolymer of
N-glucosamine and N-acetyl-D-glucosamine, is prepared via partial deacetylation of chitin. Due
to similarities with glucosaminoglycans side chains of ECM glycoproteins (e.g., chondroitin
sulfate, heparin sulfate, keratan sulfate), chitosan can interact with fibronectin, laminin, and
collagen. It has been used extensively in biomedical applications including nerve conduits (Wang
et al., 2005; Gu et al., 2011). In one construct, a double-layered tube was devised comprising an
outer chitosan film for mechanical strength and an inner layer of electrospun chitosan fibers
(Figure 2g). In addition, extended YIGSR peptides were covalently bound to the chitosan fibers to
enhance interaction with cells. As 10 mm peripheral nerve guides in rats, these constructs were
successful as autografts (Itoh et al., 2005; Wang et al., 2008). When chitosan/PLGA implants
were complemented with bone marrow-derived stem cells, even 50 mm sciatic gaps in dogs could
be repaired almost as well as with autologous nerve transplants (Ding et al., 2010). Chitosan
scaffolds were also prepared with longitudinal pores, which improved regeneration and functional
recovery (Huang et al., 2010). In other experiments, chitin/chitosan implants caused infiltration of
many inflammatory macrophages, and modification of the material was therefore considered
(Huang et al., 2010).
Alginate and agarose
Alginate, extracted from brown seaweed, consists of repeat units of mannuronic acid and
glucuronic acid. Having free carboxyl groups it can be cross-linked in the presence of calcium.
Most often, alginate is used as a gel inside implanted nerve tubes (Suzuki et al., 1999; Ohta et al.,
2004); however, alginate gel has been used as glue to connect nerve stumps 7 mm apart (Suzuki et
al., 2000), and hollow tubes were made of freeze-dried alginate/chitosan blend. For the delivery of
a growth factor, nerve tubes were filled with alginate and implanted in a rat sciatic nerve model.
This study showed that regeneration was better in empty conduits, suggesting that the presence of
alginate actually impeded regeneration (Mohanna et al., 2003). Agarose is also extracted from
seaweed and has been used extensively for cell cultures (Dalton and Mey, 2009). In the context of
artificial nerve implants, an interesting application has been the use of agarose as a material for
creating concentration gradients of growth factors (Mohanna et al., 2003, 2005).
Hyaluronic acid
Another important glucosaminoglycan of the ECM in connective tissues, especially cartilage and
skin, is hyaluronic acid. It consists of repeating disaccharide units of D-glucoronic acid and D-N-
acetyl-glucosamine. For biomedical applications, hyaluronic acid gels proved to be useful because
the molecule is involved in many aspects of wound repair, including angiogenesis, reduction of
scar formation, and moderation of the inflammatory reaction. A positive effect on PN
regeneration was achieved with local application (Ozgenel, 2003). Hydrogels consisting of

Chapter I
30
hyaluronic acid and ECM proteins were used as carrier matrix in cell-based repair strategies for
the PNS (Suri and Schmidt, 2010).
4.1.4 Self-assembling peptide scaffolds
As a new strategy in the design of scaffolds for nerve repair, self-assembling peptide nanofibers
have already been mentioned above. They consist of natural amino acids and form a three-
dimensional network of nanofibers through spontaneous self-assembly. The ability of EAK16-II
(AEAEAKAKAEAEAKAK; one-letter amino acid code for Glu, Ala, Lys), a 16 amino acid
peptide derived from the yeast protein zuotin, to self-assemble into a nanofibrous scaffold was
observed serendipitously when the peptide was tested for its cytotoxicity in cell culture (Zhang et
al., 1993). This finding has inspired the development of a number of additional self-assembling
peptides including RADA16-I (AcN-RADARADARADARADA-CNH2) and RADA16-II (AcN-
RARADADARARADADA-CNH2), in which Arg (R) and Asp (D) residues substitute Lys and
Glu (Holmes et al., 2000). Studies, which demonstrated the compatibility of self-assembling
peptides with neural tissue were cited above. In a few experiments, these were tested for nerve
repair (Rafiuddin Ahmed and Jayakumar, 2003; Silva et al., 2004; Ellis-Behnke et al., 2006;
McGrath et al., 2010). The RADA16 self-assembling peptide solution has been developed into a
commercial product intended for wound repair and three-dimensional cell cultures (3DM,
www.PuraMatrix.com).
4.2 Synthetic polymers
In comparison with biologically derived materials, synthetic polymers have several advantages
and disadvantages. Since they do not need to be extracted from animal tissue they pose a lower
risk of contamination and immunological reaction. Their physical and chemical properties can be
better controlled and manipulated. On the other hand, synthetic materials usually do not activate
specific cellular receptors and therefore do not by themselves elicit the desired cellular responses
such as axonal growth or cell migration. Some materials may even be rejected or encapsulated in
scar tissue.
4.2.1 Non-degradable materials
The first generation of tubular implants for PN repair were made of silicone rubber, which
showed some success (Lundborg et al., 1982, 1997b, 2004; Braga-Silva, 1999). Another non-
degradable material is poly(acrylonitrile-co-methylacrylate) (PAN-MA). Recently, a peripheral
nerve conduit was designed by stacking oriented, electrospun PAN-MA fibers in the two halves
of a longitudinally split polysulfone tube, which were resealed and implanted as a sciatic nerve
bridge in rats. In the absence of additional cells or growth factors this construct supported
excellent regeneration, which was monitored with histological, physiological, and behavioral
methods (Kim et al., 2008). This and other studies (Clements et al., 2009) demonstrated the
advantage of three-dimensional arrays of nanofibers as guidance structures in vivo (Figure 5).

4 Materials for artificial nerve conduits
31
Other non-degradable materials used for nerve implants are extended poly(tetrafluoroethylene)
(ePTFE, Gore-Tex) and poly(urethane) (Stanec and Stanec, 1998). Since non-degradable
materials may cause a chronic foreign body reaction and therefore need to be removed in a second
operation, they are no longer a preferred strategy for artificial nerve implants (Gu et al., 2011).
4.2.2 Aliphatic polyesters
Perhaps the most important category of biodegradable synthetic polymers with favorable
characteristics for nerve repair is that of aliphatic polyesters. The most important examples are
poly(a-hydroxy acids), i.e., poly(lactic acid) (PLA, with stereoisomers PLLA, PDLA),
poly(glycolic acid) (PGA), poly(lactic-co-glycolic acid) (PLGA), poly(ε-caprolactone) (PCL), and
poly(hydroxybutyric acid) (PHB). The compounds are completely biodegradable via hydrolytic
digestion, and many cell culture experiments demonstrated excellent biocompatibility with nerve
tissue of PLA (Yang et al., 2005; Liu et al., 2011), PGA (Ichihara et al., 2009), PLGA (Bini et al.,
2004; Ding et al., 2010; Yucel et al., 2010), PCL (Schnell et al., 2007; Klinkhammer et al., 2009;
Wang et al., 2009), and PHB (Mohanna et al., 2003, 2005; Aberg et al., 2009). However, they are
not soluble in water, do not directly activate biological signals, so possibilities for conjugation
with functional peptides are limited. For nerve bioengineering purposes, poly(a-hydroxy acids)
are dissolved in non-aqueous solvents and spun to fibers that can be used as material for the nerve
tube or as oriented guidance structures for growing axons and cells. Fibers can also be spun
directly from the polymer melts. Often blends of different polymers or blends with biologically
derived materials have been used successfully. A few important examples are given below, but, as
many other studies could be cited, this selection is somewhat arbitrary.
Poly(L-lactic acid)
Cai and coworkers produced porous tubular PLA conduits, which had an internal structure of
aligned PLA filaments. Ten weeks after implantation in a rat sciatic nerve lesion model, this
implant supported axon regeneration and remyelination much better than silicone conduits in
comparison (Cai et al., 2005). Tubes consisting of collagen-coated PGA and braided PLA/PGA
were compared in dogs for 40 mm repair of the peroneal nerve. Regeneration, including recovery
of muscular function after 12 months, was best in PLA/PGA-collagen tubes, where it reached
80 % of the positive control (Ichihara et al., 2009).
Poly(lactic-co-glycolic acid)
With electrospinning, PLGA fibers were collected on a rotating mandrel, and tubular nerve
conduits from this material were tested as 10 mm bridges in the rat PNS. One month after
implantation, no inflammatory response was recorded but successful regeneration was observed
only in 5 of 11 rats (Bini et al., 2004). Substantial improvements were possible with combinations
of PLGA and chitosan, coated with CNTF. Histological evaluations showed that such an implant

Chapter I
32
was able to induce regeneration across a 25 mm tibial nerve gap. In a canine model, the conduit
was as good as the autologous nerve transplant (Shen et al., 2010).
Poly(ε-caprolactone)
Electrospun PCL fibers alone are good substrates for axonal regeneration and the migration of

4 Materials for artificial nerve conduits
33
various glial cells (Gerardo-nava et al., 2009). These properties are significantly improved via
blending with collagen (Schnell et al., 2007) or covalent binding with ECM-derived peptides
(Bockelmann et al., 2011, see below). Oliveira and coworkers seeded PCL conduits with
mesenchymal stem cells and tested them for repair of median nerves in mice. Motor function after
8 weeks was assessed with a grasping test in addition to histological examination. The study
demonstrated good regeneration and a positive effect of the implanted cells (Oliveira et al., 2010).
Poly(hydroxybutyric acid)
In experiments with a PHB conduit, excellent results were achieved when the tube was filled with
a GGF-containing gel (Mohanna et al., 2003, 2005). In a spinal cord implantation experiment,
PHB constructs showed good integration and axonal regeneration within the graft (Novikova et
al., 2008). These descriptions make clear that the best results were always obtained when polymer
fibers were functionalized or combined with ECM proteins (Chap.I 4.3).
4.2.3 Polyphosphoesters
In addition to various aspects of biocompatibility, which they share with poly(a-hydroxy
carboxylic acids), fibers of polyphosphoesters (PPE) offer the potential for coupling of bioactive
molecules. PPE were therefore developed as matrices for PN implants (Wang et al., 2001). Films
of electrospun fibers of poly(caprolactone-co-ethyl ethylene phosphate) (PCLEEP) served also as
a delivery device for NGF (Chew et al., 2005) or GDNF (Chew et al., 2007). While it was
possible to spin parallel aligned fibers of the PCL/PPE copolymer on a rotating drum,
incorporation of the growth factor in the spinning solution made this difficult. When rolled films
Figure 5 Evaluation of artificial nerve implants in animal experiments.
(a–c) Electron microscopy of transverse sections: (a) non-lesioned sciatic nerve of the rat; (b)
regeneration in an autologous nerve transplant; (c) regeneration in an implant with gradients of NGF
and laminin after 4 months; distance 10 mm from the transection site; Ax axons, M myelin sheaths, Sc
Schwann cell, scale bar 2 mm (Dodla and Bellamkonda, 2008). (d, e) Immuno-histochemical staining
of longitudinal sections of regenerated axons in an implant with electrospun microfibers in rat sciatic
nerve after 4 months: above neurofilament, axons; below S100, Schwann cells (Kim et al., 2008). (f)
Retrograde tracing with FluoroGold from the distal end of a chitosan/PGA implant showing axonal
regeneration of sensory neurons from the dorsal root ganglion in dogs at 6 months after implantation
(Wang et al., 2005). (g) Electrophysiological recordings to assess functional regeneration cross a gap
of 80 mm in dogs after 12 months. Collagen tubes with laminin-coated fibers were implanted into the
peroneal nerve. Traces are shown for compound muscle action potential (CMAP) of the anterior
tibialis muscle after stimulation of the sciatic nerve; motor evoked potentials (MEP) in the tibialis
muscle after electrical stimulation of the motor cortex; somatosensory evoked potentials (SEP)
recorded from the cerebral cortex after muscle stimulation; and spinal cord evoked potentials (SCEP)
with stimulation of the nerve stump distal to the graft (Matsumoto et al., 2000). (h) Functional
demonstration of regeneration (through the implants shown in d and e) in rats. In the grid walking
test the number of mistakes are counted while the rat walks across a grid. Implants with fibers
orientated in parallel (asterisk) achieved better results than implants without or with non-orientated
fibers, yet autologous nerve transplants were still better (Kim et al., 2008). (i) The largest nerve gaps
repaired with artificial implants, which showed recovery of motor functions, were found with dogs.
Note movement of the left hind limb (Toba et al., 2001)

Chapter I
34
of fibers were tested as a 15 mm nerve bridge in rats, nerve regeneration was observed in all
animals after 3 months, even without the growth factor. Electrophysiological examination showed
a beneficial effect of the encapsulated growth factor (Chew et al., 2007).
4.2.4 Poly(ethylene glycol)
This polymer, which is non-adhesive to cells, is used to produce gels and matrices or serves as a
linker for proteins. From poly(ethylene glycol), the star-shaped NCOpoly(ethylene glycol)-stat-
poly(propylene glycol) (sPEG) with a backbone of 80 % ethylene oxide and 20 % propylene
oxide has been synthesized. A range of biochemical functionalities such as isocyanate, acrylate,
and vinyl sulfone can be introduced to provide the basis for covalent modification. Using sPEG
with reactive isocyanate end groups, blends of PCL and PCLdiol were functionalized with ECM
derived peptides, which improved their properties as guidance substrates for axons and Schwann
cells from DRG (Klinkhammer et al., 2010; Bockelmann et al., 2011). Recently, an easy method
for peptide functionalization of polyester fibers via sPEG has been published, which is promising
for in vivo applications (Grafahrend et al., 2011).
4.2.5 Poly(2-hydroxyethyl methacrylate) and related hydrogels
Synthetic hydrogels, a prime example being poly(2-hydroxyethyl methacrylate) (pHEMA), have a
very high water content after polymerization. Gels consisting of poly(HEMA-co-methyl
methacrylate) (pHEMA-MMA) implanted in the spinal cord of adult rats integrated well. Axonal
regeneration into the implant was found by 8 weeks (Tsai et al., 2004). Hydrogels can be used to
make scaffolds with structured channels or to encapsulate functional molecules when growth
factors are added to pHEMA sheets during polymerization in aqueous solution. A nerve tube
consisting of porous pHEMA-MMA was tested in 10 mm rat sciatic nerve gaps. After 8 weeks,
the outcome was similar to autografts, and after 16 weeks 60 % of the artificial tubes were still
comparable to this positive control (Belkas et al., 2005b). In an experimental medical application,
pHEMA-MMA tubes filled with collagen resulting in a gel-like outer and porous inner structure.
As implants in the adult rat sciatic nerve, such conduits were very successful over a period of 8
weeks. In long-term survival studies, however, some of these implants tended to collapse and also
evoked inflammatory reactions as suggested by the presence of macrophages and giant cells in the
implants after 16 weeks (Belkas et al., 2005a). Copolymerization of HEMA with 2-aminoethyl
methacrylate provided primary amine groups for covalent binding of laminin-derived
oligopeptides. This approach was combined with physical modification of the gel, which
contained multiple longitudinal channels (Yu and Shoichet, 2005).
4.2.6 Oxidized polypyrrole
The electrically conducting polymer poly(pyrrole) (PPy) is interesting for neural applications
because electrical stimulation can influence axonal growth. In one study, electrospun PLGA fibers
were coated with PPy, and electrical stimulation of the conducting nanofibers enhanced neurite

4 Materials for artificial nerve conduits
35
outgrowth (Lee et al., 2009). This work and related pioneering studies (Gomez and Schmidt,
2007) were done with a PC12 cell line, however, this has only limited predictive value for
neurons in vivo. While the material properties of PPy, which was discovered in the 1960s, have
been thoroughly characterized, there is still limited experience with PPy as implant material in
vivo (Schmidt et al., 1997; Wang et al., 2004; George et al., 2009). In a tube of combined PPy and
silicone implanted to bridge a 10 mm gap in the rat, regeneration of nerve tissue was only
marginally better than that in the plain silicone tube (Schmidt et al., 1997; Wang et al., 2004;
George et al., 2009). Future studies will show whether the electrical conductance of PPy and other
conducting polymers (polyaniline, polythiophene) is a real advantage in PN implants in vivo.
4.3 Functionalization with ECM proteins and peptides
As noted above (Chap.I 3.3), tubular implants and scaffolds with longitudinal pores have been
made of collagen with very promising results. Similar success was obtained with artificial
polymers that were blended or coated with collagen and laminin (Inada et al., 2004, 2007;
Ichihara et al., 2009). On the other hand, synthetic polymers proved very useful in the
construction of nerve guides, not least because of their superior physical properties. For instance,
many hydrophobic polymers can be spun much more easily than ECM proteins (Klinkhammer et
al., 2009). Unfortunately, they lack the specific biochemical functions involved in nerve
regeneration (Chap.I 2.2 and 2.3). The purpose of functionalization is therefore to endow
synthetic polymers with molecular moieties that activate endogenous cells (neurons, Schwann
cells, endothelial cells) in a way that promotes axonal growth. Corresponding to the distinction
between secreted and surface-bound signals, two fundamental approaches have been employed:
binding of ECM molecules and controlled release of soluble growth factors.
4.3.1 Blends of synthetic polymers with proteins
The use of ECM molecules from the PNS is suggested because axons regenerate naturally along
the basal lamina. Thus, ECM proteins or peptides are coupled to synthetic polymers either by
blending, adsorption, or covalent binding (Koh et al., 2008) (Figure 6). This should promote cells
to migrate into the artificial implant and close the gap between the nerve stumps. As discussed
above, the main components of the basement membrane are the trimeric laminins plus collagen
IV. Fibrous collagens (I, III, V) are major components of the ECM (Koopmans et al., 2009).
Thus, a logical strategy was to blend synthetic polymers (e.g., of PLA with laminin (Koh et al.,
2008) or PCL with collagen (Zhang et al., 2005; Schnell et al., 2007)) and spin this material to
fibers, which could then be used for the fabrication of tubular implants or guidance structures.
PLA/laminin blends and PCL/collagen blends significantly improved the surface characteristics of
electrospun nanofibers in comparison with the pure synthetic polymers. Some groups were able to
spin fibers of pure ECM proteins (Chew et al., 2005; Heydarkhan-Hagvall et al., 2008); however,

Chapter I
36
some found it difficult to produce aligned fiber substrates of good quality with this method
(Klinkhammer et al., 2009).
4.3.2 Covalent functionalization of polymer fibers
Instead of whole ECM proteins, which are extracted from biological sources, specific peptide
sequences from laminin, fibronectin, or collagen can be synthesized (Wang et al., 2008; Dalton
and Mey, 2009). Their design was made possible by the knowledge of specific amino acid
Figure 6 Functionalization of polymer fibers for nerve regeneration.
(a) Functionalization with adsorption or covalent binding of ECM molecules to electrospun polymer
fibers (Koh et al., 2008). (b) Functionalization by blending of ECM molecules with synthetic polymers
before electrospinning (Schnell et al., 2007). (c) Functionalization by chemical bulk modification of
the electrospinning solution (Klinkhammer et al., 2010)

4 Materials for artificial nerve conduits
37
sequences in the ECM that bind to integrin receptors of Schwann cells and neurons. These
peptides are adhesive because of the intracellular connection of integrins to the cytoskeleton. In
addition, they activate a number of signal transduction cascades within the cells (Geiger et al.,
2001). So far, peptides with the amino acid sequences RGD, YIGSR, IKVAV, and longer
versions of those have been successfully applied. People from my laboratory showed, for
instance, that covalent binding of GRGDS to electrospun nanofibers of PCL improved their
ability to guide axonal growth (Bockelmann et al., 2011). Several functionalization strategies with
ECM molecules are also being tested in vivo: The interior surface of a chitosan tube was modified
with various laminin peptides. As an artificial bridge of the rat sciatic nerve, the tube with one of
these sequences promoted regeneration to a similar degree as a transplanted nerve, although not as
well as autologous implants (Wang et al., 2008). In addition to ECM proteins, the cell adhesion
molecules (CAMs, important examples for nerve regeneration are L1, N-CAM) constitute another
group of surface-bound signals that can induce axonal growth. These transmembrane proteins,
which are exposed on cell membranes and are not part of the ECM, are also being considered for
functionalization (Webb et al., 2001).
In conclusion, the chemical functionalization of synthetic materials with ECM molecules was an
important step in the development of artificial implants with biological activities. Apart from
these signals, which are always fixed to surfaces, peripheral nerve regeneration in vivo depends
on a number of signals that are secreted from Schwann cells or the neurons themselves (Chap.I
2.2).
4.4 Controlled release of growth factors
To apply soluble factors in vivo, the first ideas were to fill implanted devices with a solution of
the growth factor or with growth-factor-secreting cells. In a more sophisticated manner, carriers
are incorporated that gradually release the active molecules over longer periods. Several problems
have yet to be solved (de Ruiter et al., 2009). For instance, the biological activity should be
present for several weeks to months, especially in the case of longer nerve lesions. Also,
sterilization should be possible without compromising the function of the signals. As these issues
have been studied in several experiments, examples for the major paths of delivery are given
below. The reader can find additional references in (Siemionow et al., 2010).
4.4.1 Immobilization in the tubular wall
A common feature of artificial nerve conduits is an outer wall in the form of a tube. Different
techniques have been developed to immobilize growth factors such that they are released over
time from the scaffold wall to the inside. In one study, tubes were fabricated by dip-molding from
a solution of ethylene-vinyl acetate, bovine serum albumin, and FGF-2. In vitro assays revealed a
burst phase of FGF release for the first 3 days (50 % of total protein), which then declined to a
rate of 0.1 – 0.5 % per day. In vivo, after 4 weeks these tubes were clearly superior in bridging a

Chapter I
38
15 mm sciatic nerve gap compared to tubes without the growth factor (Aebischer et al., 1989).
Madduri and coworkers incorporated NGF and GDNF in conduits consisting of collagen
(Madduri et al., 2010a) or fibroin (silk) fibers (Madduri et al., 2010b). During biological
degradation of the scaffold, the neurotrophic factors were gradually released.
4.4.2 Growth factors released from gels
Advanced conduit designs involve internal scaffolds for stabilization and guidance of the
regenerating tissue. Hence, an obvious approach is to incorporate the growth factor into this
polymer by mixing before gelling. This was done with NGF and NT-3 in pHEMA (Moore et al.,
2006). The growth factor can be equally distributed over the conduit or in the form a gradient to
attract continued regeneration in the distal direction of the implant. In some cases, it could
become a problem that axons are trapped within the chemoattractive environment of the conduit
(“candy store phenomenon”). In a successful example of the gradient strategy, 40 mm gaps in the
peroneal nerve of rabbits were bridged within little more than 6 weeks. This was achieved using
PHB conduits filled with GGF (pro-neuregulin-1), which was suspended in alginate (Mohanna et
al., 2005). Neither empty nor alginate-filled tubes without GGF were able to support growth of
Schwann cells or axons across the complete gap, even after 9 weeks (Mohanna et al., 2003).
Integration of polysaccharides into the polymer can enhance growth factor retention due to
hydrophobic, electrostatic, or ionic interaction. This is a specific activation effect with heparan
sulfate proteoglycans and growth factors FGF-2, VEGF, and GDNF (Barnett et al., 2002; Forsten-
Williams et al., 2008). This synergistic interaction was exploited by using alginate/heparin gels as
a matrix for FGF-2, which increased vascularization and faster axonal growth through the
artificial implants (Ohta et al., 2004). Combining growth factors with ECM proteins was also
promising. Dodla and colleagues devised an agarose gel to apply gradients of NGF and laminin.
Both gradients had a positive effect on nerve regeneration (Dodla and Bellamkonda, 2008).
4.4.3 Microspheres as delivery vectors
Instead of inducing growth factors into the conduit wall or the inner matrix, they can be
encapsulated in microspheres before incorporation. With this approach, the release kinetics can be
tested and tuned beforehand in vitro. Microspheres of PLGA and PLA with GDNF were
embedded into the inner half of a PCL-tube fabricated by dip-coating. The microspheres were
formed by adding an oil-in-oil emulsion of PLA and PLGA (the latter containing GDNF) drop-
wise to an aqueous solution, followed by centrifugation and lyophilization. The GDNF release
kinetics observed in vitro showed a strong release of 4.6 ng GDNF per mg microsphere during the
first day. The subsequent release approached zero-order kinetics and reached a cumulative release
of 6.4 ng/mg microsphere at day 64. The bridging of a 15 mm rat sciatic nerve gap with this nerve
conduit was much better than without the GDNF, similar to results obtained with nerve autografts
(Kokai et al., 2011). In another study, nerve conduits containing PPE microspheres with NGF

5 Conclusion
39
were successfully used to bridge a 10 mm rat sciatic nerve gap. Three months after implantation,
rats with NGF-microsphere conduits showed a higher percentage of positive reflex response than
controls after pinching distal nerve trunks (Xu et al., 2003).
4.4.4 Release from electrospun fibers
Aligned electrospun fibers, which are perhaps the most promising topological cues for axon
guidance, were also used to deliver growth factors. For example, PCLEEP fibers containing
GDNF have been obtained by mixing GDNF into the polymer solution followed by
electrospinning. The fibers were spun into a PCLEEP film, which was later rolled to form a tube
on the inner surface of a nerve guide (Figure 2g). Release kinetics were characterized by an initial
burst of about 30 % of protein followed by relatively stable release until leveling off after almost
2 months. In vivo investigations with 15 mm rat PN lesions revealed a significantly higher
number of myelinated axons inside the GDNF-implant compared to the controls (Chew et al.,
2007). Recently, a conduit was designed that consisted of a PLA-co-PCL shell made of
electrospun nanofibers filled with an NGF-containing core of bovine serum albumin. Again, when
tested as a sciatic nerve bridge of 10 mm in rats, results after 12 weeks were similar to those
obtained with autografts (Liu et al., 2011).
In summary many different and equally promising strategies of functionalization have been
followed during the last decade (Figure 5 and Figure 6). Unfortunately, since there are hardly any
studies that compare different strategies of functionalization, at this moment it is difficult to
decide which molecules should be preferred in the future and which delivery strategy is most
suitable.
5 Conclusion
In contrast to long-distance connections within the CNS such as the optic nerve or the white
matter tracts of the spinal cord, lesioned peripheral nerves contain the inherent cellular and
molecular components that are required to support successful axonal regeneration. The
development of axon growth-promoting devices that incorporate a number of these components
has become an expanding area of research. Although a number of biocompatible materials have
been developed, which offer great potential for therapies of peripheral nerve injury, several
technical or scientific challenges need to be addressed.
5.1 Standardized control of efficiency
Many experiments carried out using different biomaterials are not comparable with each other
because different cell cultures, animal models, or criteria of success were used. In general, I have
observed that a considerable number of publications reporting innovative materials failed to use
appropriate biological tests. Papers falling into this category have not been discussed in the
present review. Even in many animal studies, the inflammatory reactions, effects on gene

Chapter I
40
expression, and long-term results are rarely investigated. Clearly, a systematic approach with
internationally agreed standards would be desirable for such investigations to allow a better
degree of comparison.
5.2 Structured three-dimensional implants
So far, most studies of PN regeneration in vivo have only implanted hollow tubes, where axons
start to grow on the inner surface of the implant. Thus a major technical challenge is the
construction of three-dimensional nerve bridges with an internal architecture that is capable of
guiding axonal growth and migration of the endogenous Schwann cells. In my opinion, the most
promising approaches are the three-dimensional integration of microfibers in parallel orientation
and the production of orientated channels in gels or matrices of ECM proteins.
5.3 Biomimetic functionalization of implants
With the use of proteins and peptides derived from ECM, artificial implants can already mimic
the signals that regenerating axons encounter in the environment of the lesioned PNS. However,
the concept of targeted use of factors that specifically activate Schwann cells, sensory neurons, or
motor neurons is still in its infancy. In addition to the selection of the appropriate combination of
molecular signals, the ideal temporal and spatial presentation of such molecules for the efficient
support of axon regeneration and tissue repair remains unknown. In this context, it is possible that
neurotrophic signals might be required for several months and with increasing gradients towards
the distal end of the implant.
5.4 The thirty millimeter mark
Studies have demonstrated that peripheral nerve regeneration through artificial implants can cover
distances of up to 30 mm but rarely beyond. Successful motor fiber regeneration appears to be
more difficult than that of sensory axons, and thin fascicles regenerate more easily than large
caliber nerves. Since larger gaps must occasionally be bridged in cases of human nerve injury, the
usefulness of future constructs will be judged by their ability to support motor and sensory axon
regeneration over long distances. For this, it is likely that experiments with animals larger than the
rat will be required. I am convinced that results from basic research on mechanisms of axonal
growth in combination with engineering methods (such as electrospinning, freeze-drying, and
chemical functionalization) will result in further progress, with the development of artificial nerve
implants that approach or even exceed the efficiency of the autograft. Some very substantial
advances have already been made in the last few years. Thus, there is great hope and expectation
that such strategies will provide the neurosurgeon with a range of cell-free scaffolds that can be
used on demand.

1 Introduction
41
Chapter II
Manufacture and characterization of three-dimensional fiber arrays
and assessment of guidance properties in vitro
1 Introduction
Peripheral nerve (PN) lesions can be surgically repaired by suturing the proximal and distal nerve
stumps together. In these cases, axons regenerating from injured motor, sensory, and autonomic
neurons are able to grow within the scaffolds of Schwann cells and extracellular matrix (ECM) of
the injured nerve (Dalton and Mey, 2009). When PN injuries result in larger gaps between
transected nerve ends, these have to be bridged, for instance, by transplanting a sensory nerve
from the same patient. Unfortunately, this approach is functionally successful only in about half
of the patients and has other disadvantages, such as sensory loss at the donor site and a limited
availability of transplant material (Deumens et al., 2010). Currently, much research is therefore
devoted to developing artificial nerve conduits that can replace autologous transplants (Deumens
et al., 2010; Siemionow et al., 2010; Mey et al., 2012).
For clinical use in humans, only simple tubular conduits have so far been approved in the United
States and Europe (Siemionow et al., 2010). The majority of animal experiments are also based on
the design of the hollow tube. Because axons grow on surfaces and must be guided toward their
peripheral target, better results are expected if internal structures could support axonal growth and
induce colonization with host-derived Schwann cells, which are required for trophic support and
the subsequent remyelination of regenerated nerve fibers (Hoffman-Kim et al., 2010). Recent
advances in the design of artificial nerve implants therefore focus on topographical features that
guide axonal regeneration and cell migration. Basic design strategies for nerve bridges with
internal guidance structures are based on (1) large interior channels to separate individual fiber
fascicles (Clements et al., 2009; Yao et al., 2010), (2) scaffolds with many longitudinally
orientated pores (Ceballos et al., 1999; Möllers et al., 2009), or (3) parallel aligned polymer fibers
produced by melt extrusion (Matsumoto et al., 2000; Ribeiro-Resende et al., 2009) or
electrospinning (Teo and Ramakrishna, 2006; Kim et al., 2008) (Figure 7).
The approach presented here is based on electrospinning of the biodegradable polyester poly(ε-
caprolactone) (PCL) using two separated target electrodes, such that parallel fibers accumulate
between them (Schnell et al., 2007; Klinkhammer et al., 2009). The resulting microfibers have
been shown to be excellent growth substrates for glial cells and axons of sensory neurons
(Gerardo-nava et al., 2009; Bockelmann et al., 2011). Given successful in vitro tests of several
two-dimensional growth substrates a major challenge in scaffold design consists in the
construction of three-dimensional (3D) structures that can be implanted in vivo. In this study, I
describe a technique for 3D spinning of parallel microfibers, their embedding in collagen gels,

Chapter II
42
and incorporation in biodegradable PCL tubes. The biocompatibility of the resulting 3D scaffold
was investigated using dorsal root ganglia (DRG) explants.
2 Material and Methods
2.1 Electrospinning of three-dimensional fiber arrays
Solvents and PCL used for electrospinning were purchased from Sigma-Aldrich Chemie GmbH
(Munich, Germany). Electrospinning was performed with 8 wt% and 12 wt% PCL (Mn = 70,000 –
90,000 g/mol) dissolved in chloroform/methanol (3:1). Unless otherwise indicated, data are
reported for fibers made from 12 % PCL. The spinneret consisted of a syringe with a 20G flat
tipped needle connected to a metal disc with 90 mm diameter. After various target electrodes had
been tested, the standard target configuration consisted of two opposing aluminum bars with a
distance of 18 mm between them. These were mounted on a tapered base, which was connected
with its narrow end to a grounded metal disc (Figure 8a). Of the different target devices, all made
Figure 7 Conduit design for artificial nerve guides. (a) Empty tube used for bridging gaps between proximal and distal nerve stumps,used in most experiments so far; (b) tubular implant with multiple intraluminal channels (e.g. de Ruiter et al., 2008; Yao et al., 2010); (c) tube filled with an internal matrix structured by longitudinally oriented pores or channels (e.g. Bozkurt et al., 2012); (d) aligned polymer fibers as longitudinal guidance structures (e.g. Kim et al., 2008; Ribeiro-Resende et al., 2009); (e) present construct consisting of 3D aligned PCL fibers embedded in layers of collagen gel.

2 Material and Methods
43
of aluminum (Figure 8b–e), a stair case-like target was used for spinning of 3D fiber arrays
(different sizes were successfully tested, e.g., 12 steps, 20 mm breadth, 1 mm step size; five steps,
13 mm breadth, 1 mm step size; (Figure 8d,e and Figure 9a). Standard spinning conditions were
17 kV, with 20 cm distance to the collector without mechanical ejection of the polymer solution
by a syringe pump resulting in a feed rate of about 0.5 ml/h. However, depending on ambient
conditions the distance between spinneret and target was adjusted between 17 and 20 cm to give
Figure 8 Electrospinning setup for the collection of 3D fiber arrays. (a) Schematic diagram of the spinning device consisting of a syringe, 20 gauge needle connected with a metal plate (spinneret), high volt- age power supply, and grounded collection target, which is mounted on a conducting v-shaped base likewise connected to a metal plate. (b–e) Photos of different target designs to illustrate accumulation of parallel fibers. Two parallel bars (b) resulted in a single layer of paral- lel fibers accumulating on the upper edge. Substituting edged bars with a curvature allowed (c) accumulation on a slightly wider depth on the target. The best target for 3D-spinning is characterized by a stepped design (d) resulting in a sequential accumulation of fibers from bottom to the top edge (e, arrows).

Chapter II
44
Figure 9 Three-dimensional arrays of parallel fibers. (a) Electrospun fibers suspended in a step target are shown from the side. (b) Projection of a 400-mm confocal scan from above demonstrates the parallel alignment of the fibers. (c) SEM image of fibers with 200-fold and 5000-fold magnification. High magnification photos were used to determine the fiber diameters (fiber seen as a vertical line in the inset; 808 ± 175 nm, mean ± SD)
best results. For spinning of one batch of fibers I applied the voltage for approximately 30 s to the
spinneret. To handle the three-dimensional array of fibers after spinning, the bars were attached to
a U-shaped frame before disassembling it from the base. To visualize electrospun fibers with
fluorescence microscopy, the lipophilic dye DiI [Molecular Probes, D3911, 75 ml DiI/DMSO
(1/1000) in 20 ml] was added to the PCL solution.
2.2 Physical characterization of fiber arrays
Electrospun fibers suspended on the targets were examined with a Leica TCS SP2 confocal
microscope (Leica Microsystems GmbH, Wetzlar, Germany). For documentation of alignment
and density of the fiber arrays image stacks were produced from the upper most fiber plane
reaching 2 mm down into the lowest fiber plane. The image stacks were processed for further
analysis using Image J software. For evaluation of fiber density, I divided a stack into five bins of
400 µm, made a projection of all images in one bin and counted the number of fibers crossing the
midline. For assessing fiber alignment, the mean direction of all fibers in a stack was determined
and the deviation angles of the individual fibers were plotted. The standard deviation (SD) of
deviation angles was calculated using MATLAB. Fiber diameter was determined by sputter
coating the samples with 30 nm gold and
performing scanning electron microscopy
using a FEI/Philips XL30 FEG ESEM
with a 10 kV electron beam. Using high
magnification (5000×) images of single
fibers I averaged the thickness of each
fiber from five positions within an image.
Mean fiber thickness and standard error
(SEM) were calculated from 23 fibers of
five different samples.

2 Material and Methods
45
2.3 Inserting the 3D fiber arrays into a conduit
For biological testing, the electrospun fibers were inserted in gels that consisted of type I
collagen. For the matrix, I prepared a 2.4 mg/ml solution of rat tail collagen I (Life Technologies,
A1048301) according to the manufacturer’s protocol. In brief, one part 10× basal medium Eagle
was added to eight parts rat tail collagen I solution. I neutralized the pH with 1 M NaOH and then
filled up to 10 parts with distilled water (DRG cultures) or with cell suspension to obtain
106 cells/ml. To prevent premature gelation, the collagen solution was kept on ice during the
whole procedure. To produce composites of collagen and PCL fibers, a drop of 50 ml collagen
solution was applied onto a poly-L-lysine coated cover slip. Using a U-shaped frame to hold the
target electrodes the fiber array was deposited into the collagen solution and detached from the
frame using hot forceps. To induce gelation, the composites were placed in an incubator for 30 –
40 min at 37°C and 95 % humidity. This step could be repeated several times to incorporate
additional layers of fibers.
For manufacturing implantable tubes to be filled with the composite of gel and fibers, porous,
biodegradable PCL tubes were made by dip coating. Crystalline NaCl was ground to fine powder
in a mortar and was mixed with an equal amount (wt) of a 12 % PCL-solution in
chloroform/methanol (3:1). Steel wires of 2 mm diameter were repeatedly dipped into the
NaCl/PCL suspension to produce conduits of 1 – 2 cm length and 500 µm wall thickness (for a
final construct of 1.5 cm). The surface was leveled after 1 min drying at RT. After another 5 h of
drying, the conduits were removed from the wires and transferred to a Petri dish with stirred
deionized water for 24 h to dissolve the NaCl(Chang, 2009). Water was changed twice. Conduits
were cut lengthwise in half and filled with gel/fiber stacks as described above for glass cover
slips. Subsequently, half-tubes were resealed using the PCL solution and equilibrated with cell
culture medium. All steps were performed under sterile conditions. The thickness of the tube wall
was determined with scanning electron microscopy.
2.4 Cell and organ culture
All cell culture reagents were purchased from Life Technologies GmbH (Darmstadt, Germany).
Before the experiments, astrocytoma cells (U373) were grown in Dulbecco’s modified Eagle
medium (DMEM) with Glutamax and enriched with 10 % fetal calf serum (FCS) and 0.2 %
penicillin/streptomycin at 37°C, 5 % CO2. The culture medium was changed every second day
and cultures were split twice a week. For the experiments, cells were washed with phosphate
buffered saline (PBS) and detached from the surface of the culture flask using 0.25 %
trypsin/ethylenediaminetetraacetic acid (EDTA). After trypsination was stopped by adding 5 ml
serum containing medium, cells were centrifuged and resuspended in culture medium. Cell
numbers were determined using a Thoma chamber.

Chapter II
46
For organ cultures, I dissected DRG from 10-day-old chicken embryos. The embryos were
decapitated and the skin removed. Accessing from the dorsal side, sharp forceps were inserted
close to the lumbar vertebral column. By opening the forceps the hind limb could then be
separated from the torso. The DRG, remaining on the inside of the thigh, were removed and
collected in cold, sterile PBS, where they were kept until being transferred to the collagen matrix
(< 30 min) and before adding of the PCL-fibers. After gel formation, the samples were placed in a
24-well plate with 1 ml culture medium and incubated at 37°C and 5 % CO2 for six (single cells)
or seven days (DRG). The medium was changed once after three days.
The biological compatibility of the complete conduits was also tested with DRG cultures. After
coating the inner side of the longitudinally sectioned half-conduits with poly-L-lysine, several
layers of collagen and PCL-fibers were added together with a DRG explant. To allow microscopic
evaluation, I sealed the surface with a poly-L-lysine coated cover slip and transferred the
assembly upside down to a culture well, where culture media could only enter through the wall or
the endings of the conduit. After 30 min gelation at 37°C I added 1 ml of culture medium to the
well. The culture was maintained for seven days with medium change once after three days in
vitro. Axonal growth and cell migration was investigated in whole mounts.
2.5 Immunocytochemistry
Samples were fixed 1 h with cold 4 % paraformaldehyde/phosphate buffer, rinsed with PBS and
blocked with 1 % bovine serum albumin, 4 % normal goat serum and 0.1 % TritonX-100 in PBS
for 1 h. Incubation with the primary antibody [either 1/500 rabbit-anti-S100 (Sigma-Aldrich,
Munich, Germany, S2644) or 1/500 rabbit-anti-NF200 (Sigma-Aldrich, N4142)] was performed
over night at 4°C in blocking solution. Incubation with secondary antibody, Alexa Fluor 488 goat-
anti-rabbit (Life Technologies GmbH, Darmstadt, Germany, A11008), 1/500 in blocking solution,
was done for 1 h at room temperature followed by nuclear staining with 4’,6-diamidino-2-
phenylindole (DAPI, 1:5000) for 10 min. Negative controls were performed without primary
antibodies. Samples were placed upside down on object slides using Fluoprep (Biomerieux) to
stabilize fluorochromes.
2.6 Fluorescence microscopy
Images of the U373 astrocytoma cell cultures were captured using a Leica TCS SP2 confocal
microscope and were further processed using Image J software. Cell migration and axonal
outgrowth from DRG were analyzed with a Leica DM5000B epifluorescence microscope using
filter cubes L5, N2.1 and A4 for Alexa Fluor 488, DiI and DAPI nuclear staining. Photos were
recorded with a Leica DFC350 FX digital camera. Composite pictures of entire DRG and all
outgrowing cells and axons were assembled by means of a Leica DMI6000 microscope with
LASAF software. Individual cells and their interaction with PCL fibers were imaged using Leica
TCS-SP5 confocal microscopy and further processed using Image J.

3 Results
47
2.7 Quantification of Schwann cell migration and axonal growth
On composite photographs of DRG four tangents were drawn to the edges of the DRG in parallel
and orthogonal to the fiber direction or, in case of the controls, horizontal and vertical with
respect to the photographic frame. For cell migration, I measured the distance between the farthest
cell body and the corresponding tangent. For axonal outgrowth, I counted the number of visible
axonal endings and measured the distance between the 10 % longest axons and the corresponding
tangent. In the statistical analysis the individual DRG were considered as independent samples.
Conditions were compared using Student’s t-test, calculated with MATLAB.
3 Results
3.1 Electrospinning of parallel fibers in a 3D configuration
The configuration of the fiber array produced with electrospinning depends on the geometry of
the target which is used. Although a simple plate target results in a mat of non-oriented fibers,
parallel fiber alignment can be obtained with targets consisting of a rotating drum or two parallel
bars. Using the latter design, I collected PCL fibers that were suspended in the air between the
two target bars (Figure 8a). Fibers were aligned in parallel but formed one layer as they attached
to the upper edge of the bars (Figure 8b). To achieve a 3D array of the accumulating fibers,
various target configurations were tested by modifying the bars. For instance, replacing the sharp
edges with a rounder curvature led to the accumulation of fibers on a slightly wider area of the
target, yet the distribution was not easy to control and was frequently different between the left
and right bar (Figure 8c). Taking advantage of the fact that the fibers tend to accumulate on edges,
where electric field lines converge, I designed step-like targets with multiple edges facing each
other (Figure 8d). On this kind of target, the accumulation of fibers starts at the lowest edges.
With increasing density as fibers accumulate on these edges, electrostatic repulsion between them
forced further fiber accumulation towards the next higher steps. In this way, a consecutive
accumulation of fibers from bottom to top was achieved (Figure 8e) resulting in a 3D
configuration of PCL fibers. A view from the side of a 3D fiber array spun onto the final version
of the target is shown in Figure 9a. Microscopic evaluation of fluorescently labeled fibers (Figure
9b) and SEM visualization (Figure 9c) demonstrated the degree of their alignment.

Chapter II
48
Figure 10 Quantitative characterization of the 3D fiber array. (a) Fiber density was determined from sequential confocal scans of 400 mm thickness through the fiber arrays suspended in the target frame (mean ± SEM, n = 3 targets). The fiber density decreases from top to bottom with the bottom layer still containing more than 18 fibers/mm2. (b) The histogram depicts variability of fiber orientation as individual deviation from the mean orientation (n = 559 fibers from six samples. Angular standard deviation is 2.46°).
3.2 Characterization of electrospun 3D PCL-fiber arrays
To quantitatively characterize the fiber arrays, the fiber density at different depths inside the array
was binned (bin 1 to 5; Figure 10a). The fiber density decreased from the top to the bottom of the
3D array, with the highest and lowest fiber density, 86.7 ± 5.1 fibers/mm2 (area of cross section, n
= 5 fiber stacks, mean ± SEM) and 18.3 ± 6.4 fibers/mm2
in the first and last bin, respectively
(ANOVA p < 0.005; F = 8.00; df = 4). The mean fiber density was 48.8 ± 4.5 fibers/mm2.
To assess fiber alignment, I determined the direction of the individual fibers, calculated the mean
orientation of all fibers and for every fiber its deviation angle from the mean orientation (Figure
10b). The alignment of electrospun fibers was high, as fiber angles showed a narrow Gaussian
distribution with around 70 % of the fibers deviating less than 2.5° from the mean fiber
orientation (n = 559 fibers; SD = 2.46°). Only about 1 % of all measured fibers deviated more
than 10° from the mean. The maximum deviation was 15°.
Fiber diameters were determined on the basis of scanning electron microscopy photos (Figure 9c).
As described previously (Jha et al., 2011), fiber diameter was dependent on the PCL
concentration of the spinning solution used. A concentration of 8 % PCL resulted in a fiber
diameter of 660 ± 190 nm (mean ± SEM). By raising the concentration to 12 %, the fiber diameter
was increased to 808 ± 175 nm (p < 0.05; compared by student’s t-test).
3.3 Suspension and manipulation of 3D fiber
arrays in biocompatible gels
The 3D arrays of PCL fibers were embedded into a
collagen matrix by inserting the fibers into the gel
(Figure 11a). To handle the 3D fiber stacks for this
purpose, electrodes were fixed to a metal frame
(Figure 11b) and detached from the spinning setup
with the suspended fibers between them. The
embedding caused some compression of the array
thereby increasing the fiber density.

3 Results
49
To achieve larger 3D stacks that were not limited by the size of the electrospinning targets, this
procedure could be repeated several times by stacking alternate layers of fiber arrays (Figure 11c–
e). The thickness of the gel layers between 3D fiber arrays was determined by the viscosity of the
gel solution. To reduce displacement of the fibers by the collagen solution it was important to
introduce them before gelation of the collagen took place. Gelation was initiated by heating the
collagen to 37°C, but to some extent changes in viscosity of the collagen already occurred at room
Figure 11 Embedding of 3D fiber stacks in collagen. (a) Schematic drawing of the embedding process. The collagen solution is applied onto a poly(lysine)-coated coverslip. Subsequently, fibers are dipped into the collagen solution and detached from the frame using hot forceps. Gelling was induced by incubation of the composite at 37 C for 30 min. To add fiber layers, the process was repeated several times. (b) For handling of the fiber stacks a U-shaped frame was used to keep the two parts of the target in place when detached from the base. (c–e) Arrangement of fiber stacks in the collagen gel shown in cross section after one (c), two (d), and 10 (e) rounds of deposition. (f) One single 3D array of PCL fibers was maintained in a gelatin gel which was cast over the fibers suspended between the target electrodes.

Chapter II
50
temperature. Thus with collagen gels I obtained a matrix with several 3D arrays of fibers
separated by regions devoid of fibers (Figure 11d and Figure 7e). A standard array now consists
of 10 layers of fibers and gels with a distance of 70 – 120 µm between the layers (Figure 11e)].
An alternative method to contain the 3D fiber array within a hydrogel consisted in casting the gel
solution over the fibers (Figure 11f)].
The effect of fiber-containing collagen matrices on cell shape was first tested using the glial cell
line U373. These cells grew well within the scaffolds and produced cellular processes. When the
cells were cultivated inside the 3D collagen matrix without PCL fibers, I observed a more diverse
morphology than in 2D cultures. In the 3D gels, many cells had complex cell bodies with several
processes in various directions (Figure 12a). When PCL fibers were added in 2D or 3D cultures
cell bodies tended to attach to the polymer fibers (Figure 12b). In 3D collagen matrices, all cells
Figure 12 Gel-embedded PCL fibers affect cell morphology. To visual- ize the effects on cell shapes U373 cells were cultivated for 6 days in a 3D collagen matrix with and without PCL fibers. (a,b) Confocal microscopy images show immunostaining against S100 (green) and nuclear staining (DAPI, blue). PCL fibers were stained red using DiI. Panels (a) and (c) show cell morphologies in the absence of PCL fibers, panels (b) and (d) with PCL fibers in the gel. Different magnifi- cation are shown in (a) and (b) with all scale bars measuring 50 mm. (c,d) Reconstructions of cells from both conditions to display the dif- ferent cellular morphology in space. The red arrow in (d) points at the PCL fiber.
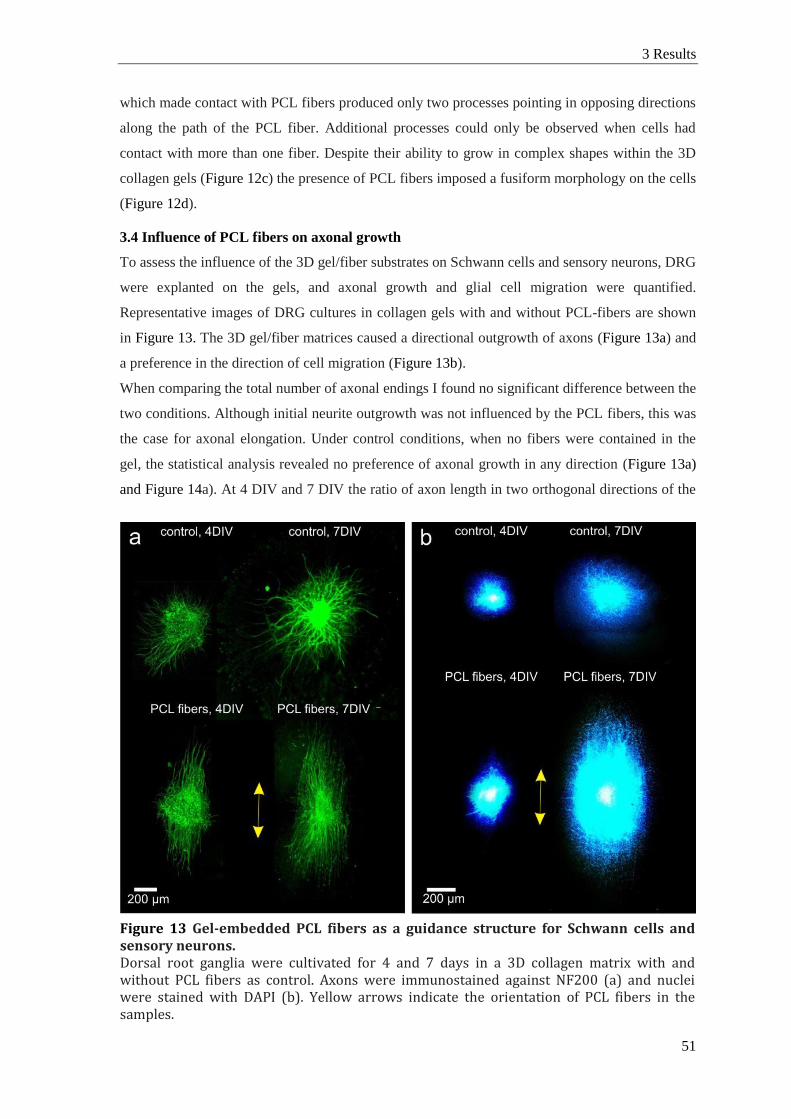
3 Results
51
which made contact with PCL fibers produced only two processes pointing in opposing directions
along the path of the PCL fiber. Additional processes could only be observed when cells had
contact with more than one fiber. Despite their ability to grow in complex shapes within the 3D
collagen gels (Figure 12c) the presence of PCL fibers imposed a fusiform morphology on the cells
(Figure 12d).
3.4 Influence of PCL fibers on axonal growth
To assess the influence of the 3D gel/fiber substrates on Schwann cells and sensory neurons, DRG
were explanted on the gels, and axonal growth and glial cell migration were quantified.
Representative images of DRG cultures in collagen gels with and without PCL-fibers are shown
in Figure 13. The 3D gel/fiber matrices caused a directional outgrowth of axons (Figure 13a) and
a preference in the direction of cell migration (Figure 13b).
When comparing the total number of axonal endings I found no significant difference between the
two conditions. Although initial neurite outgrowth was not influenced by the PCL fibers, this was
the case for axonal elongation. Under control conditions, when no fibers were contained in the
gel, the statistical analysis revealed no preference of axonal growth in any direction (Figure 13a)
and Figure 14a). At 4 DIV and 7 DIV the ratio of axon length in two orthogonal directions of the
Figure 13 Gel-embedded PCL fibers as a guidance structure for Schwann cells and sensory neurons. Dorsal root ganglia were cultivated for 4 and 7 days in a 3D collagen matrix with and without PCL fibers as control. Axons were immunostained against NF200 (a) and nuclei were stained with DAPI (b). Yellow arrows indicate the orientation of PCL fibers in the samples.

Chapter II
52
image was close to one (Figure 14b). In contrast, collagen gels that contained aligned PCL-fibers
had the effect that axons in direction of the PCL fibers were significantly longer than in the
absence of fibers, and axons in the direction orthogonal to the PCL fibers were significantly
shorter (Figure 14a), t-test p < 0.01 at 4 DIV]. For instance, after 4 DIV mean axon lengths in
parallel with PCL fibers was 1148 ± 81 µm, whereas in the orthogonal direction they only reached
378 ± 36 µm.
As axonal growth along the electrospun fibers continued in the cultures, the difference between
the two directions was even larger at 7 DIV. Axons also continued to grow in collagen gels
without PCL fibers. For this reason the difference in axon length between gels without PCL fibers
and the preferred direction in the collagen/PCL fiber matrix was no longer significant. However,
the orientation index demonstrated quantitatively that the fiber containing matrices continued to
Figure 14 Quantification of the guidance properties of gel-embedded PCL fibres. (a) Axon length in different directions is plotted as a result of the different 3D substrates. Data show mean of the 10 % longest axons of every explant. Substrates were collagen gels without or with PCL fibers. For the controls axonal growth was analyzed in arbitrarily defined horizontal and vertical (h and v) directions; for fiber containing gels axon lengths was assessed in parallel and orthogonal to the fiber direction (p and o). (b) An orientation index for axon length was determined as the ratio between the two directions measured for every condition (h/v or p/o). (c) The distance of cell migration from the DRG was quantified in a similar manner, and (d) an orientation index was calculated accordingly. Asterisks indicate significant differences between the PCL fiber sample and control; pound symbols indicate differences between the two directions within PCL fiber samples. Bars show mean ± SEM (n ≥ 6 DRG explants; Student’s t-test; p* < 0.05; p*** < 0.001; p### < 0.001).

3 Results
53
exert a strong guidance effect on the growing axons, whereas outgrowth was random in isotropic
collagen gels (Figure 14b). Comparing the results at both time points indicated that under all
conditions axonal elongation slowed down during the time course of cultivation.
3.5 Influence of PCL fibers on glial cell migration
The data for cell migration demonstrated a similar effect of the aligned PCL fibers (Figure 13b
and Figure 14c,d). Under the control condition of 3D gels without fibers there was no significant
difference in the distance of cells that migrated from the DRG in vertical or horizontal direction
(e.g., 4 DIV: 834 ± 52 µm vs. 780 ± 60 µm). When cultivated in the presence of aligned PCL-
fibers, cell migration parallel to the fiber orientation reached significantly longer distances (e.g.,
at 4 DIV: 1509 ± 55 µm) in comparison with cell migration perpendicular to the PCL fibers
(4 DIV, 7 DIV: both p < 0.001) and also in comparison with cell migration in 3D gels without
fibers (4 DIV: p < 0.05; 7 DIV: p < 0.001). As for axonal growth the orientation index was highly
significant at 4 DIV and 7 DIV (Figure 14d).
3.6 Incorporation of gel/fiber composites in a tubular conduit
To use the gel/fiber composite scaffold as a substrate for PN regeneration I inserted it into a PCL-
conduit, which was manufactured by dip-coating a steel rod with PCL solution. The addition of
NaCl to the PCL-solution and subsequent leaching resulted in the formation of a tubular conduit
wall with a porous surface (Figure 15). It also increased the flexibility of the conduit and hence
the suitability for in vivo applications. Surfaces and cross sections of tubes were examined with
scanning electron microscopy (Figure 15a,b), and parameters of the dipping technique
(concentration of the solutions, repetitions of coating) were adjusted according to the results.
PCL tubes were longitudinally cut in half, gel/fiber scaffolds were inserted into the half tubes, and
artificial nerve bridges were then produced by sealing two half tubes with PCL-solution (Figure
15c). To investigate the biocompatibility of the constructs, DRG explants were cultivated inside
the half conduits, which were sealed with a cover slip, so that exchange of metabolic matter could
only occur through the gel and PCL wall or via the endings of the conduit. The cultivated DRG
showed no visible change in appearance compared with those cultivated in open 3D gels. After
7 DIV, axon growth and cell migration had occurred within the 3D matrix (Figure 15d) and on the
surface of the cover slips. Anti-NF200 immunohistochemistry revealed widespread axonal growth
and similar axon lengths to those observed in 3D cultures that were not enclosed by tubular
conduits. Nuclear DAPI staining of cells that had migrated out of the DRG indicated cell
migration was also not impaired.

Chapter II
54
4 Discussion
To develop artificial scaffolds for PN regeneration I have described a method for the 3D assembly
of electrospun PCL fibers in parallel alignment. Consecutive embedding of these fibers in layers
of collagen gels and incorporation in a tubular conduit resulted in a 3D scaffold for axonal
guidance, whose biological use was tested with DRG explants. Schwann cells and sensory axons
adhered to the PCL fiber substrate and were guided by the orientated structure inside the gel/fiber
scaffold. Within the 3D collagen gels contact with the microfibers caused the cells to assume a
spindle-shaped morphology following the fiber direction.
Previous developments of fiber substrates revealed two major challenges with respect to the goal
of an artificial nerve implant, (1) the incorporation of microfibers in a 3D configuration, and (2)
the functionalization of the growth substrate to attract axonal growth over long distances
Figure 15 Integration of the composit scaffold into a biocompatible conduit. Conduits were produced from a mixture of a 12 % PCL solution and NaCl by dipmolding and subsequent leaching of the NaCl fraction. (a,b) SEM images depict the porous structure of the conduits wall (cross-section in a) and surface (b). (c) Scheme of the culture model used to verify biocompatibility of the conduit. In the inset a photograph of the collagen-fiber scaffold integrated into the tubular PCL conduit is shown. (d) Axonal growth and cell migration occurred within the structured 3D conduit. Axons are marked with NF200 immunostaining, migrated cells with nuclear DAPI stain.

4 Discussion
55
(Bockelmann et al., 2011). The present study contributes to solving the first of these problems and
provides a 3D platform to address the second.
4.1 Three-dimensional configuration of guidance structures
Artificial nerve bridges are designed as tubular grafts that connect the proximal with the distal PN
stump (Siemionow et al., 2010; Mey et al., 2012). When hollow cylinders with a single lumen are
used, regenerating axons and migrating cells accumulate on the inside of the tubular walls.
Constructs with several lumens were fabricated to increase that surface area and to separate
different fiber bundles within the conduit (de Ruiter et al., 2008; Clements et al., 2009; Yao et al.,
2010). However, increasing the number of compartments with these methods, for example by
using seven channels or by incorporating three films, did not improve regeneration (Clements et
al., 2009; Yao et al., 2010). Although implants have also been developed that were filled with
non-orientated “isotropic” matrices (Labrador et al., 1998; Toba et al., 2001; Inada et al., 2004;
McGrath et al., 2010), most studies indicate that orientated 3D structures with a large surface area
are required to provide an adequate substrate for long distance axonal guidance (Hoffman-Kim et
al., 2010; Mey et al., 2012). I can distinguish two basic designs that provide such internal
guidance structures within tubular implants (Figure 7). The tubes contain either longitudinal fibers
or a porous matrix with longitudinal orientation of the pores. Conduits that consist of polymer
fibers can be produced by a number of different techniques such as drawing from a melt, template
synthesis, phase separation, self-assembly and electrospinning (Teo and Ramakrishna, 2006;
Schnell et al., 2007; Kim et al., 2008). A major challenge consists in filling the conduit with the
orientated fibers.
One strategy consists of stacking 2D films of parallel fibers above one another. In one previous
study, this was done with 10 – 12 films of electrospun poly(acrylonitrile-comethylacrylate) fibers
inside longitudinally split polysulfone tubes, which were then sealed and implanted to bridge
17 mm sciatic nerve gaps in rats (Kim et al., 2008). More recently, an improved method for
manufacturing 3D fiber scaffolds was published (Yang et al., 2011). Highly aligned parallel fibers
were electrospun onto individual non-conductive frames using a conductive wire as target. The
holding frames with suspended microfibers were then placed across the surface of a hydrogel,
covered with a spacer frame and another layer of hydrogel was added. This process was repeated
several times to create a 3D microfiber scaffold (Yang et al., 2011). The procedure explained in
the present study is based on the 3D accumulation of fibers already during the spinning process.
This could be used also for successive stacking of fiber and gel layers (Figure 11d,e) or for
casting a gel over the 3D fiber array (Figure 11f).
While I modified the physical design of the target electrode to achieve a 3D fiber configuration, a
similar technique of “air gap electrospinning” also used two horizontally separated targets, each
connected to a common voltage with respect to the spinneret (Jha et al., 2011). To manufacture an

Chapter II
56
implantable scaffold, an exterior coating of poly(glycolic acid)/poly(lactic acid) copolymer was in
this case deposited onto the outside of the cylindrical fiber configuration. For this, a circular steel
plate was placed behind the fibers, and the spinneret was used for subsequent electrospraying of
the copolymer (Jha et al., 2011). A different technique was developed by Schlosshauer’s group,
who produced oriented PCL filaments by hot melt extrusion through nozzles with six-leaf cross
sections. After extrusion, the filaments were drawn in a separate process. This resulted in yarns of
several hundred filaments, each with six longitudinal grooves, which could be inserted in a
tubular conduit (Ribeiro-Resende et al., 2009). These structured filaments have been
functionalized in various ways including siRNA (Hoffmann et al., 2011) and were successfully
used to fill epineurial tubes in vivo (Hajosch et al., 2010). Using the method of injection molding
chitosan conduits were produced that contained several hundreds of longitudinally aligned PLGA
fibers (Wang et al., 2005; Ding et al., 2010).
The second approach to achieve internally structured implants aims at scaffolds with longitudinal
pores or channels. One method to achieve this is the application of strong magnetic fields which
orient collagen or fibrin matrices (Ceballos et al., 1999; Dubey et al., 1999; Verdú et al., 2002).
Another promising technique consists in the controlled freeze drying of collagen suspensions.
When the solvent is caused to freeze from one side of the solution, collagen is displaced, thus
resulting in collagen walls of longitudinally orientated guidance channels. The diameter of these
pores can be controlled to be in the range of 20 – 120 µm (Bozkurt et al., 2007, 2009; Möllers et
al., 2009). Structured collagen conduits, which are commercially available (Perimaix, Matricel
GmbH), were implanted as sciatic nerve grafts in rats, and axonal regeneration followed the
longitudinal pores of the artificial nerve guide. In these experiments better outcomes were
achieved by co-implanting Schwann cells (Bozkurt et al., 2012), though fiber containing implants
appear to achieve similar or better results without the need to implant live cells (Wang et al.,
2005; Hu et al., 2013).
4.2 Biological functionalization of the growth substrate
Although pure polymer fibers, such as PCL or poly(lactic acid), already proved useful as guidance
structures, several studies showed that functionalization either by covalent binding of ECM
peptides (Bockelmann et al., 2011) or blending with ECM proteins (Schnell et al., 2007)
improved the ability of such fibers to guide cell migration and axonal growth. In the present
experiments the surrounding gel already consisted of collagen with signal sequences that can
activate cellular integrin receptors. With the use of hydrogels that contain signaling molecules one
might therefore use artificial polymer fibers without the further need of functionalization. I found
that axonal growth along non-functionalized PCL fibers in a 3D collagen environment was
initially faster than in pure collagen, however, with time, that is after 1 week, this difference
disappeared. Possibly, activation of axonal integrin receptors by the collagen supported axonal

5 Conclusion
57
elongation whereas no such signaling was initiated by the PCL. This outcome suggests that
additional modification of the fibers with signaling molecules may be beneficial.
Coating of artificial polymer fibers with laminin or fibronectin enabled axons to grow faster than
the migration of Schwann cells (Hoffman-Kim et al., 2010). An easy technique to achieve such
functionalization consists of blending a synthetic polymer with fibronectin, collagen or laminin
when preparing the electrospinning solution. Cell culture investigations demonstrated this
approach to be even superior, with respect to certain aspects, to more sophisticated chemical
modification (Koh et al., 2008). Recently, an innovative method for the synthesis of covalently
functionalized poly(D,L-lactide-coglycolide) fibers has been published. The polyester was made
hydrophilic by adding star-shaped poly(ethylene oxide) followed by covalent binding of cell-
adhesion mediating peptides. The resulting fiber matrix was bioactive and promoted cell adhesion
via recognition of the immobilized binding motifs (Grafahrend et al., 2011). For functionalization
through blending or covalent modification in the electrospinning solution the present 3D spinning
setup with staircase-like targets can be used without further modifications.
When, as here, electrospun fibers are embedded in a biocompatible hydrogel, the preferential
adherence of axons to the electrospun fibers is an issue. In previous 2D studies aligned fibers
were placed on non-treated tissue culture plastic (Kim et al., 2008) or on cell repellent substrates
(Schnell et al., 2007; Klinkhammer et al., 2009), which left the cells little choice but to attach to
the fiber substrate. In 3D composites that consist of guidance fibers suspended in a supportive
matrix a suitable balance of cell surface attachment between the different material components
must be struck. With the imbedding of pure PCL fibers in a collagen gel the guidance effect or the
morphological alignment was maintained. It remains to be seen whether this remains when
additional growth promoting factors, for example neurotrophins, are introduced into the matrix
(Dalton and Mey, 2009). Different combinations of gels and modification of artificial polymers
can now be tested.
5 Conclusion
I report a technique for the 3D electrospinning of parallel microfibers that were suspended in
collagen gels and incorporated in a tubular nerve implant. In contrast to conventional hollow
nerve conduits, this nerve guide with aligned arrays of anisotropic fibers directed axonal
regeneration and Schwann cell migration in the longitudinal axis of the scaffold. The comparative
advantage of this design in comparison with alternative designs will now be tested in vivo.


1 Introduction
59
Chapter III
Assembly of artificial nerve implants and assessment of their
suitability to support nerve regeneration in vivo
1 Introduction
Traumatic injuries are frequently accompanied by damage to a peripheral nerve (PN). A complete
nerve transection considerably prolongs the dysfunction of an affected body part far beyond the
damage to bones or muscular tissue and hence impairs the patient’s autonomy, even when most of
the injured tissue has already been restored. As a result, it can aggravate the patient’s emotional
distress and produce relevant social and economic costs (Rosberg et al., 2005). Despite these
serious consequences, options for clinical treatment of PN injuries are limited (Lundborg, 2000;
Deumens et al., 2010). Since an end-to-end suture is only possible when the two stumps can be
brought together in a tensionless manner, the first choice for treating a PN gap is the insertion of
an autologous nerve graft (Meek and Coert, 2014). However, this approach implicates donor site
morbidity and may cause other complications associated with the necessity of a second operation.
For short nerve gaps it is possible to bridge the two stumps with saline-filled but otherwise empty
tubes of biodegradable materials. These are the only artificial nerve bridges that are clinically
approved at the moment (Meek and Coert, 2014). Their use circumvents the drawbacks of
autologous nerve transplants, however at the cost of a generally reduced prospect of regeneration,
and the disadvantage increases with the size of the gap (Isaacs, 2010; Konofaos and Ver Halen,
2013). Intensive research is therefore being carried out to develop new scaffold designs and
materials that allow functional recovery after PN injury (Mey et al., 2012; Jager et al., 2014).
The reduced efficacy of conventional hollow nerve conduits is attributed to the lack of supportive
cues, which are present in the distal nerve stump or in an autologous nerve graft (Meek and Coert,
2014). These include endogenous trophic factors acting on regenerating axons, including
neurotrophins, glial cell-derived neurotrophic factor (GDNF) or the fibroblast growth factors
(FGFs; Dalton and Mey, 2009). They can be incorporated with microspheres that gradually
release growth factors (Kokai et al. 2010). Alternatively, the delivery can be achieved by
introducing cells with the implant. This approach was recently chosen by Allodi et al. (2014), who
used a collagen matrix seeded with transfected Schwann cells overexpressing FGF-2 inside a
silicone tube to repair a 6 mm sciatic nerve gap in rats. Other cell types that support PN
regeneration are several types of stem cells (di Summa et al., 2010; Hu et al., 2013) and olfactory
ensheathing cells (Dombrowski et al., 2006; You et al., 2011). In addition, structural cues in the
extracellular matrix, most importantly the Schwann cell-derived basal lamina, play an important
role in axonal guidance (Johnson et al., 2011; Brooks et al., 2012). To provide a physical substrate
a number of researchers used biological materials, e.g. silk fibroin and collagen, or synthetic

Chapter III
60
polyesters, e.g. poly(ε-caprolactone) (PCL) and poly-glycolic acid (PGA). Bozkurt et al. (2007)
developed a collagen matrix with longitudinal pores to facilitate nerve regeneration, and used it to
reconstruct a biopsy induced sural nerve gap (Bozkurt et al., 2014). Particular attention has been
paid to the application of longitudinal filaments either as a part of the conduit’s wall or in its
lumen (Kim et al., 2008; Madduri et al., 2010b; Jha et al., 2011). In a clinical case study, Fan et
al. (2008) used a chitosan tube with internal filaments of PGA for repair of a 35 mm median nerve
defect in a patient. Despite these promising studies, the available artificial conduits are not
considered satisfactory, especially in comparison with autologous nerve grafts. Implants that
contain live cells are more successful, however, here the issues of immune suppression or of
finding an appropriate source for the cellular implants arise (e.g. Mey et al., 2012). For this
reason, I am aiming at a cell-free, artificial implant design that mimics structural features of the
natural ECM. In preceding experiments, I showed that arrays of electrospun PCL fibers aligned in
parallel determined the direction of Schwann cell migration and axonal growth. This effect could
be enhanced when fibers included biological signals, for example by blending PCL with collagen
(Schnell et al., 2007) or by covalent binding of integrin-activating peptides (Bockelmann et al.,
2011). This was shown first on two-dimensional substrates in vitro and recently transferred to
three-dimensional (3D) substrates with fibers embedded in collagen hydrogels (see chapter II or
Kriebel et al., 2014). The design developed in this study showed the potential of such fibers to
partly mimic basal lamina functions. Based on these results I designed an artificial conduit for PN
regeneration with an internal structure of sub-micron scale PCL or c/PCL fibers embedded in a
gelatin matrix.
In the present study, the suitability of these implants was tested by repairing a 15 mm sciatic
nerve gap in rats immediately after transection. Functional recovery was assessed via weekly
behavioral experiments followed by electromyography and histological evaluation of nerves,
implants and peripheral muscles 12W after surgery.
2 Material and Methods
2.1 Manufacture of artificial implants
To manufacture structured implants of sub-micron scale PCL-fibers in a 3D gelatin matrix
(implant design referred to as “G1”), 12 wt% PCL in chloroform/methanol (3:1) were electrospun
onto a stepped target. In this way, a 3D configuration of fibers (Figure 16a and b). For more
detailed information on the setup, spinning process and fibers see chapter II or Kriebel et al.,
2014. Subsequently, sterile 5% gelatin, dissolved in PBS, was applied onto an object slide to form
a 1.5 mm thick film into which the fiber array was submerged (Figure 16c). Using hot forceps the
fibers were detached from the target, and the gelatin was kept on ice for 15 minutes. Two
razorblades with a fixed distance of 1.5 mm were used to punch out stripes of the gelled films

2 Material and Methods
61
with longitudinal fibers. The stripes were frozen and inserted into hollow collagen conduits to
form the artificial implant. Empty spaces between conduit wall and internal scaffold were
cautiously filled up with gelatin solution followed by immediate cooling. For the second design
(G2) blended c/PCL fibers were spun from a solution of 12 wt% collagen/PCL (1:3), dissolved in
1,1,1,3,3,3-hexafluoro-2-propanol. The rest of the procedure was carried out as for G1. The empty
collagen conduits were provided by a commercial manufacturer and, for comparison, were used
also in a third group of non-structured implants (ET).
2.2 Animals and surgical procedure
For the experiments 10-week old, female Lewis rats of ca. 200 g body were used weight. Animal
housing and treatment was carried out in accordance with SfN guidelines for animal
experimentation and with permission of the Landesamt für Natur, Umwelt und Verbraucherschutz
Nordrhein-Westfalen. There were five experimental groups with 15 rats per group. Of these, three
were sacrificed after 2W for histological assessment of the biocompatibility of the implants, seven
after 12W (electrophysiology, behavioral assays, histology) and five after 14 weeks (behavioral
assays, neuroanatomical tracing, tracing data not included in this study). For surgery, anesthesia
was induced and maintained via inhalation of an isoflurane/air mixture of 5 % and 3 %,
respectively, using the Univentor 400 (Univentor High Precision Instruments). The depth of the
anesthesia was checked by testing of pedal withdrawal and corneal reflexes. The surgical area was
shaved and disinfected. Animals were placed in a prone position onto a thermostatically regulated
heating pad to maintain a constant body temperature. Subsequently, the right sciatic nerve, from
the sciatic notch to the point of trifurcation into peroneal, sural and tibial nerve, was exposed via
Figure 16 Brief documentation of the fiber production and embedding in gelatin. (a) Stair case-like target electrodes for electrospinning were attached to a U-shaped frame to allow handling of the fiber array. (b) Electrospun fibers accumulated at the target electrodes during the spinning process. (c) Gelatin (ca. 40°C) was applied onto a glass cover slip and fibers were submerged into the gelatin and detached from the target electrodes. Subsequently, the gelatin fiber composite was chilled to 4°C. From this, composite stripes were produced and inserted into collagen tubes to form implants G1 and G2.

Chapter III
62
incision of the skin and blunt dissection of the thigh musculature. This was followed by the
excision of a 15 mm piece of the sciatic nerve. According to experimental group, animals
immediately received one of the following treatments. For an autologous nerve transplantation
(AT) the excised piece of nerve was re-implanted with epineural sutures using 10-0 monofilament
nylon (Ethilon, Ethicon Inc., Somerville, USA). For groups G1, G2 and ET the respective
artificial implants were trimmed to a length of 17 mm and equilibrated for three minutes in cold
saline. Then the nerve stumps were inserted 1 mm into the ends of the implant and secured with a
single 10-0 mattress suture. Any residual air bubbles inside the conduits were removed before
insertion of the proximal stump. In the negative control group (lesion only, LO), the proximal
stump was embedded in the neighboring thigh muscle and secured with a 10-0 stitch to prevent
regeneration of axons into the distal stump. Finally, the thigh muscles and the skin were closed
with 7-0 and a 4-0 suture respectively. For pain relief animals received a subcutaneous injection
of 0.05 mg/kg body weight buprenorphine hydrochloride (Temgesic).
2.3 Tissue preparation and immunohistochemistry
At 2W after surgery and subsequent to electromyography at 12W after surgery, anesthetized rats
were transcardially perfused using 4 % buffered paraformaldehyde (PFA). Immediately after
perfusion, the sciatic nerve and muscles of the lower hind limb from both sides were dissected
post-fixed 24 h with 4 % buffered PFA and stored in PBS containing 0.05 % sodium azide.
Muscular atrophy: Since the susceptibility to atrophy depends on the predominant muscle fiber
type (Yamakuchi et al., 2000; Desaphy et al., 2001) four different muscles were weighed, which
vary in fiber type composition or size. The small Musculus soleus and M. extensor digitorum
longus have comparable size, but vary in fiber type composition with approximately 95 % slow
twitch/5 % fast twitch in the first and mostly fast twitch fibers in the second muscle (Soukup et
al., 2002). The M. gastrocnemius and its antagonist the M. tibialis anterior were more
heterogeneous and much larger. For muscle weight analysis, muscles were briefly dabbed dry
with tissue paper and weighed. Muscle weight on the operated side was normalized to the
contralateral muscle of the individual animal and subsequently averaged per group.
Nerve regeneration: For sectioning, nerve samples were cryoprotected with 30 % buffered
sucrose, embedded in Tissue-Tek and snap frozen in isopentane (-80 °C). Using a cryostat, cross
sections (16 µm) were made from the nerve proximal and distal to the implant (including tibial,
sural and peroneal branches) as well as from the medial part of the implant, and longitudinal
sections (16 µm) from the proximal and distal coaptation sites. For immunohistochemistry, the
sections were blocked for 1 h with 0.1 M PBS containing 5 % normal goat serum (NGS) and 0.5
% Triton X100, incubated overnight at room temperature in a mixture of primary antibodies,
diluted in 1 % NGS and 0.5 % TritonX100. A mouse monoclonal antibody was used against β-
tubulin III (T8660, Sigma, 1:1000), rabbit polyclonal antisera against S100 (S2644, Sigma,

2 Material and Methods
63
1:1000) and choline acetyltransferase (ChAT; AB143, Millipore, 1:200). Subsequently, sections
were washed and incubated for 2 h with Alexa Fluor-conjugated secondary antibodies goat anti
rabbit IgG (Alexa Fluor 488, A11008, Invitrogen, 1:1000) and goat anti mouse IgG (Alexa Fluor
546, A11003, Invitrogen, 1:1000) and 4',6-diamidino-2-phenylindole (DAPI, Sigma-Aldrich; 1
µg/ml) in 1 % NGS and 0.5 % TritonX100. Histological sections were visualized with a Leica
TCS SP2 confocal microscope (Wetzlar, Germany).
2.4 Image analysis
Cross sectional areas of the regenerated tissue in the medial segment of the implant/autologous
nerve graft were determined using ImageJ 1.46r (NIH, USA). Briefly, the regenerated tissue was
encircled using the polygon selection tool followed by an area measurement of the region of
interest. One image per animal and all animals (N ≥ 5) per group were analyzed.
Axon numbers were counted and the cross sectional area of individual axons determined with
custom-made MATLAB software (TheMathWorks, Natick, MA). Images of cross sections of the
medial segment, stained against βIII-tubulin, were converted into binary images using individual
thresholds set by an observer blind to the treatment conditions of the samples. Thresholds were
chosen so that during conversion the original data of axon size were conserved as far as possible
while avoiding confluence of images of individual axons. From the converted binary images, the
software automatically determined number and size of individual axons. The obtained data were
checked with manual counting (in 2-3 samples per experimental group). To exclude profiles of
non-plausible sizes, the software was set to detect only objects within a range of 0.2 -78 µm2
(≈0.5 - ≈10 µm diameter).
For analyzing the percentage of motoraxons in the total axonal population, images of medial cross
sections from three individual animals per group, co-stained against βIII-tubulin and ChAT, were
used. The percentage of ChAT-positive axon profiles that were also positive for βIII-tubulin was
determined in randomly chosen regions of interest.
2.5 Electromyography
The experiments were conducted 12W after initial surgery. Anesthesia was performed as
described above, and body temperature was controlled using a heating pad. After exposing the
sciatic nerves on both sides, a bipolar hook electrode was used to stimulate the sciatic nerve were
proximal to the lesion, close to the sciatic notch. Compound muscle action potentials (CMAPs)
were recorded from the plantar Musculus flexor digitorum brevis and the M. interosseus pedis
dorsalis between toes two and three using a monopolar needle electrode. The reference electrode
was positioned at the contralateral site between the separated thigh musculature. Stimuli were
rectangular current pulses of 100 µs duration and 30 % supramaximal intensity, determined by
stepwise increase of the injected current until CMAP amplitude reached a plateau. Absolute

Chapter III
64
intensity was on average 1.76 ± 1.78 mA (mean ± SD). For a single measurement, ten repetitions
with a frequency of 1 Hz were averaged.
Signal detection and analysis was performed using custom made MATLAB software. For
detection of the evoked potential, a threshold slope and, in case of small signals, an amplitude
threshold was used. The software automatically analyzed the time to first peak, in the following
referred to as “latency”, and the potential difference between highest and lowest peak, referred to
as “amplitude”. To correct for differences in amplitude caused by a different impedance of the
recording electrodes, CMAPs on the operated side were normalized to the corresponding
contralateral side of the same animal, which was recorded immediately afterwards. For the final
evaluation, means and SD (N ≥ 5 animals) were calculated for every experimental group.
2.6 Behavioral experiments
To assess the recovery of motor functions, toe spread and grip strength baseline values were
determined on three consecutive days prior to surgery, followed by weekly testing starting 1W
after surgery.
For toe spread measurements, animals were placed on a transparent surface with a camera system
underneath. The animals were lifted up by the experimenter and a sequence of images with 1 Hz
frequency was produced to gather at least six images, in which the animal was not moving and
had both feet in one plane. Custom made MATLAB software was used to present the images in a
randomized fashion to a single blinded observer, who had to mark the position of the 1st and 5th
toe of both feet. Subsequently, the software determined the mean ratio of the distances between
the toes of the operated and non-operated hind limb.
Grip strength was determined using the “Bioseb grip strength test” device. For this, while the
animal was held by the experimenter, its hind paw was placed on a metal grid fixed to the test
device. Then the animal was pulled away from the grid, while the maximum force which the
animal could exert on the bars was measured. Three trials were averaged per animal and time
point. For the final analysis of toe spread and grip strength measurements, means and SD of all
animals (N ≥ 11) in the respective groups at a given time point were calculated.
2.7 Statistical analysis
Individual animals were considered as independent samples, and means, SD and SEM were
determined for the experimental groups and time points. Data processing, except for
electromyography, and statistical analysis were performed using IBM SPSS Statistics Version 21
(IBM Corp., Armonk, NY, USA). Intergroup differences were tested with ANOVA. After
Levene’s test, post-hoc comparisons were performed according to Dunnet T3 for unequal
variances (i.e. analysis of grip strength, toe spread, muscle weight and median axon size) and
Scheffé for equal variances (i.e. analysis of nerve cross section area and axon count). Statistical

3 Results
65
analysis of electromyography data was performed with the MATLAB software. Intergroup
differences were tested with ANOVA using post-hoc Bonferroni-corrected t-tests.
3 Results
3.1 Artificial nerve implants and general observations
Based on a recently published technique (Kriebel et al., 2014) artificial nerve guides were
produced that consisted of a biodegradable tube with a three-dimensional array of longitudinally
oriented PCL and c/PCL fibers embedded in a gelatin matrix (Figure 16).
Following surgery the rats recovered well and did not display signs of pain, self-mutilation or
secondary complications, such as inflammation, throughout the study. Three animals of each
experimental group were sacrificed at
2W after implantation. In all cases
the nerve guides or autologous
transplants appeared well integrated
within the host tissue and showed no
macroscopic signs of neuroma
formation, or excessive encapsulation
of the conduit by fibrotic scar tissue.
At this point in time (2W), the
collagen tubes were still intact, but light-microscopical investigation revealed no vestiges of the
electrospun fibers. At 12W after surgery the tubes had been completely resorbed in all animals,
whereas the non-degradable suture material was still visible (Figure 17).
Figure 17 Tissue regeneration following sciatic nerve transection (15 mm gap size) and subsequent repair using different nerve bridges. The photographs show (a) a non-lesioned sciatic nerve (contralateral control, CL),b) regenerated tissue 2 weeks after the insertion of an artificial implant (as an example for all artificial conduits, which looked similar) and (c) regenerated tissue 12 weeks after the insertion of an autologous nerve graft (AT) and different artificial implants (ET, G1 and G2; see text).

Chapter III
66
3.2 Tissue regeneration: macroscopic evaluation of the implants
Gross examination of the artificial nerve conduits (groups G1, G2 or ET) showed that the conduit
was surrounded by a thin layer of connective tissue. At the proximal suture sites, the nerve stumps
were slightly swollen, visibly sealing the ends of the tubes. Inside the conduit a tissue cable with
clearly visible blood vessels had formed spanning the entire length of the lesion (Figure 17b). At
the later time point (12W), when the collagen tube had been degraded, the swelling at the suture
site was reduced at the proximal and had completely disappeared at the distal interface. At both
proximal and distal coaptation points, a network of blood vessels emerged. The regenerated tissue
between the proximal and distal nerve stumps had increased in diameter compared to the earlier
time point in all experimental groups (except the non-implanted group, which lacked a
regenerated tissue connection between the stumps; not shown). After 12W there was no apparent
difference between implants G1, G2 and ET, which were still thinner than the autologous nerve
grafts. Macroscopically, the autografts resembled the non-operated sciatic nerves (Figure 17c).
3.3 Schwann cell infiltration and axonal regeneration in the implants
To assess the regeneration inside the implants, longitudinal sections of the coaptation sites
between nerve stumps and the implants, and cross sections of the proximal, the distal nerve
stumps and of the mid-section of the implanted conduits were produced.
An analysis of the cross sectional area of the different implants after 12W confirmed the
macroscopic assessment of substantial growth in all artificial nerve bridges (Figure 18). The
regenerated tissue was similar in size in all experimental scaffolds (ET, G1, and G2) and still
significantly smaller than the resected and
re-implanted nerves (AT), which had the
same diameters as the non-operated control
nerves.
The inspection of the proximal connections
between nerves and implants 2W after
surgery indicated a better ingrowth of
axons (III-tubulin labeling) and Schwann
cells (S100 staining) into the internally
structured implants (G1, G2) than into the
empty tubes (ET; Figure 19). All groups
had intense staining of axons and S100+
cells proximal to the lesion. Substantial
infiltration of Schwann cells and axonal
elongation was visible in G1, G2 and AT
grafts at the proximal interface, and some
Figure 18 Cross sectional area of the regenerated tissue in the medial segment 12W after nerve transection and repair. The cross sectional areas of the autologous nerve grafts (AT) were unchanged 12W after nerve repair, resembling the non-operated situation (CL), tissue that regenerated through one of the artificial implants had significantly smaller cross sections. There were no differences between ET, G1 and G2. Symbols indicate significant differences against groups CL (*) and AT (#) (p** < 0.01; p*** < 0.001; N ≥ 6).

3 Results
67
axonal and S100+
profiles could be found in cross sections of the medial segment. In the ET
group, in contrast, the staining intensity declined abruptly at the proximal interface. At 2W there
were few axons and S100+ cells penetrating the ET conduit, but none were detected in the
midsection of the implanted segments. Distal to the graft no axons were found in any of the
groups at this early time point (2W; Figure 19).
The same assessment was performed 12W after surgery (Figure 20). In the physiological
situation, represented by the contralateral, non-lesioned sciatic nerve (CL), axons were evenly
distributed in the nerve and grouped into a few fascicles. The autograft after 12W showed a
higher degree of fasciculation but generally resembled the non-lesioned nerve. In the artificial
Figure 19 Schwann cell infiltration and axonal regeneration at the proximal interface between nerve and implant 2W post surgery. Longitudinal sections from groups CL (top), ET (middle) and G2 (bottom) stained against βIII-tubulin (a, c, e) and S100 (b, d, f) to visualize axons and Schwann cells, respectively. Yellow arrowheads point to the collagen tube, which is still present 2W post surgery. The weaker staining intensity beyond the interface of nerve and implant in group ET compared to G2 indicates a slower infiltration of Schwann cells and axons into the implant.
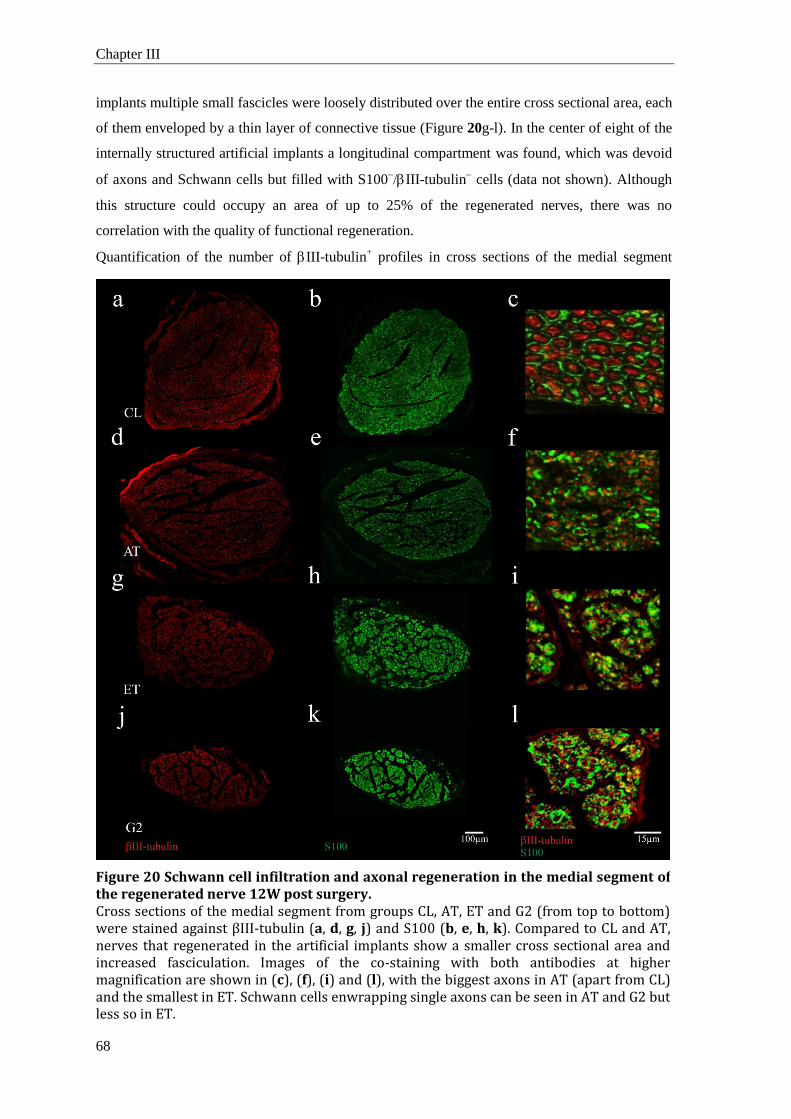
Chapter III
68
implants multiple small fascicles were loosely distributed over the entire cross sectional area, each
of them enveloped by a thin layer of connective tissue (Figure 20g-l). In the center of eight of the
internally structured artificial implants a longitudinal compartment was found, which was devoid
of axons and Schwann cells but filled with S100III-tubulin
cells (data not shown). Although
this structure could occupy an area of up to 25% of the regenerated nerves, there was no
correlation with the quality of functional regeneration.
Quantification of the number of III-tubulin+ profiles in cross sections of the medial segment
Figure 20 Schwann cell infiltration and axonal regeneration in the medial segment of the regenerated nerve 12W post surgery. Cross sections of the medial segment from groups CL, AT, ET and G2 (from top to bottom) were stained against βIII-tubulin (a, d, g, j) and S100 (b, e, h, k). Compared to CL and AT, nerves that regenerated in the artificial implants show a smaller cross sectional area and increased fasciculation. Images of the co-staining with both antibodies at higher magnification are shown in (c), (f), (i) and (l), with the biggest axons in AT (apart from CL) and the smallest in ET. Schwann cells enwrapping single axons can be seen in AT and G2 but less so in ET.

3 Results
69
revealed no differences between the non-injured nerves and the autologous implants, whereas the
artificial implants contained significantly fewer axons (Figure 21a). In all three groups (ET, G1,
G2) about 3.400 axonal profiles per cross section were counted, about half of the number
contained in the control nerves. Interestingly, the number of III-tubulin+ profiles in the medial
segments of the AT group was higher than in control nerves (CL) and also higher than proximal
to the lesion, even though this difference was not statistically significant. In all nerve repair
groups with artificial implants an increase in the number of axonal profiles was observed from the
midsection of the implant to the distal segment (significant for G1 and G2; p < 0.05 and p <
0.001, respectively), suggesting that sprouting of regenerating axons at the distal coaptation point
had taken place.
Apart from the number of all regenerating axons, the efficacy of the different implants to support
regeneration by motor neurons was investigated. Using immunostaining against ChAT (Figure
22a-b) the percentage of motor axons among the total population of axons (III-tubulin+, i.e.
motor, sensory and autonomic axons) in the midsection of control and regenerated nerves were
Figure 21 Quantitative analysis of the regenerated nerve tissue. Panel (a) shows the number of individual βIII-tubulin+ profiles (regenerated axons) in cross sections of the medial segment (significant difference against CL and AT are indicated as * and #, respectively; p<0,001; N≥6), (b) size distributions of βIII-tubulin+ profiles, step size 1µm2, and (c) their cross sectional areas in µm2 in the different groups. The solid and dashed lines in c indicate median and mean, respectively; the boxes 25 - 75 %, whiskers 10 – 90 % and dots 1 % and 99 %.

Chapter III
70
determinded. Depending on the experimental group this percentage ranged from 32.1 ± 1.2 %
(G1) to 37,1 ± 5.4% (CL) with no significant difference between any of the groups or the non-
operated control nerve (Figure 22c).
3.4 Size distribution of regenerated fibers
High magnification images revealed considerable differences in the thickness of axons and
association of Schwann cells with the fibers in the different groups. Cross sections of non-
lesioned sciatic nerves showed many large and roughly circular or elliptical axonal profiles that
were regularly encircled by S100+ Schwann cells (Figure 20c). Non-lesioned nerves exhibited
regions of predominantly bigger or smaller axons with a gradual transition between them. This
was different in the tissue 12W after nerve repair (Figure 20f, i, l) Axons were generally smaller
and the fewer large diameter axons were randomly intermingled with the mass of small axons.
The diameters of III-tubulin+ profiles were for CL: 6.0 ± 3.2 µm (mean +/- SD), AT: 3.4 ± 2.0
µm, G1: 3.9 ± 1.8 µm, G2: 5.1 ± 2.1 µm, and ET: 3.1 ± 1.5 µm. The size distribution of III-
Figure 22 Similar percentage of regenerating motoraxons in all implants. Immunostaining against βIII-tubulin (red) and ChAT (green) was used to identify motoraxons in cross sections from repaired (mid of lesion) and non-operated sciatic nerve samples (exemplary image from CL). Only motoraxons are characterized by co-localization of both antibodies (examples: white arrowheads). Quantification of the ratio of motoraxons to the total axonal population revealed no difference between the different experimental conditions and the non-operated contralateral nerve.

3 Results
71
tubulin+ profiles (Figure 21b) confirmed that small axon diameters were relatively more abundant
in the regenerated nerves. When comparing these groups, I found the number of large caliber
axons (larger than 10 µm2 cross section) to decrease from AT (548 ± 94; mean ± SEM) to G2
(389 ± 44) and G1 (307 ± 74) to ET (209 ± 29). For the non-lesioned nerve the value was 1378 ±
130. The median axon diameter was largest in the non-injured nerves, and this was significantly
different only from the ET group (p < 0.05), which had the smallest fibers (Figure 21c). The only
other experimental group with a significantly larger median of axon diameters compared to ET
was the G2 construct. It was not the case for AT because of a large number of regenerated small
fibers.
As with the axons, the appearance of S100+ positive profiles was less regular in the repaired
situation compared to control nerves. However, most S100+ positive profiles in samples from
group AT displayed a clear circular shape suggesting a myelinating phenotype, and this was
confirmed with staining against myelin basic protein (data not shown). This pattern was also
observed in groups G1 and G2, however there were also Schwann cells that did not show a clear
interaction with one axon, thus likely to have a non-myelinating phenotype. This observation was
even more pronounced in group ET (Figure 20i). In summary, judging the maturity of regenerated
axons by diameter and state of myelination, I saw the highest similarity with the non-lesioned
nerve in the AT group, a lesser degree in the structured implants G1, G2, and very little of this
organization in the ET control group.
3.5 Functional regeneration: electromyography
At the endpoint of this study, 12W after sciatic nerve transection and repair, functional
regeneration was assessed by recording of CMAPs from the plantar Musculus flexor digitorum
brevis (Mfdb) and M. interosseus pedis dorsalis (Mipd), both innervated by the tibial branch of
the sciatic nerve. CMAPs were induced with electrical stimulation of the sciatic nerve proximal to
the lesion site (Figure 23a, traces of typical recordings). The successful recording of induced
electrical muscle activity in all rats that received autologous transplants (AT) or scaffolds with
PCL-fibers (G1) and in five of the six animals that received c/PCL-fiber constructs (G2)
demonstrated that the new artificial implants supported nerve regeneration including re-
innervation of target tissue. In contrast, I was only able to record CMAPs in half of the animals in
the group that received an empty collagen tube (ET). As anticipated, no CMAPs could be
recorded from the negative control group (lesion only, LO; Figure 23b).
To evaluate the functional quality of regeneration in more detail, I analyzed amplitude and latency
of the recorded potentials (Figure 23c, d). The CMAP amplitude is dependent on the number of
functional motor units and the temporal coherence of nerve excitation in the vicinity of the
electrode, hence gives an indication of the number and quality of regenerated motor axons. For
group AT I found the strongest recovery with 48 % of the CMAP amplitude recorded, compared

Chapter III
72
to the non-lesioned contralateral side (Mipd). The second highest CMAP amplitudes were found
in G2 (22 %) followed by G1 (16 %) and the lowest in ET (10 %, Table 1). The differences
between the artificial constructs were not significant.
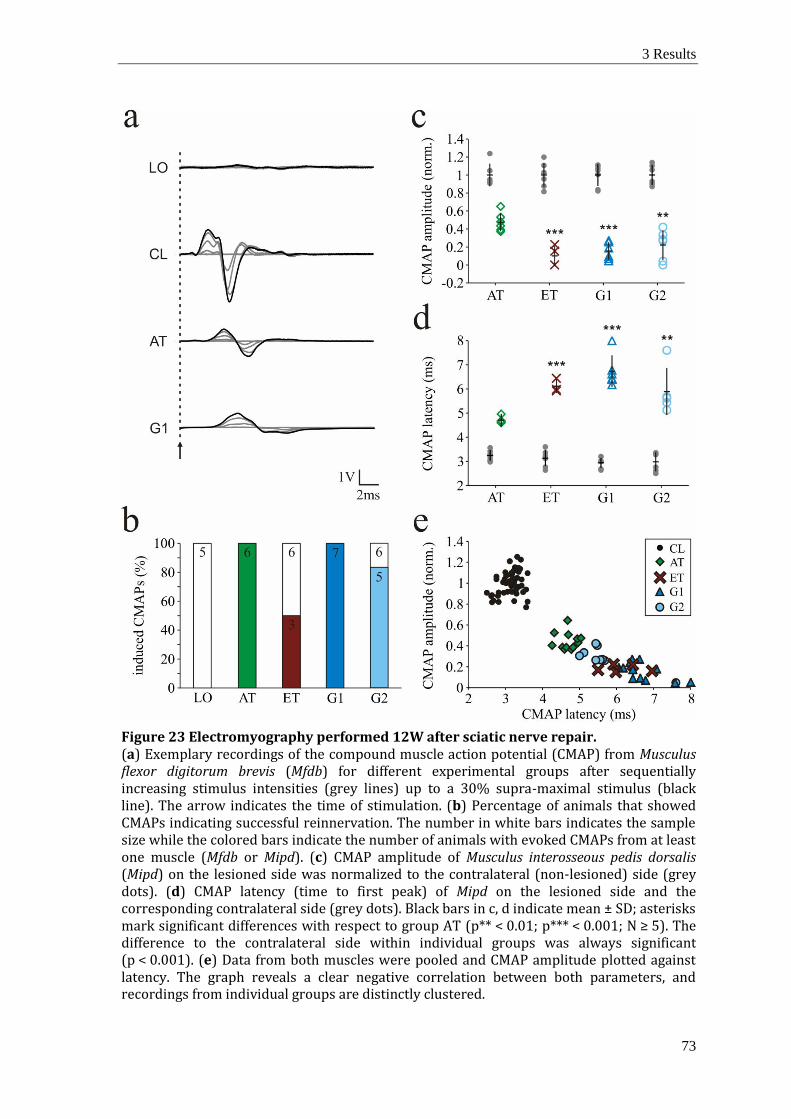
3 Results
73
Figure 23 Electromyography performed 12W after sciatic nerve repair. (a) Exemplary recordings of the compound muscle action potential (CMAP) from Musculus flexor digitorum brevis (Mfdb) for different experimental groups after sequentially increasing stimulus intensities (grey lines) up to a 30% supra-maximal stimulus (black line). The arrow indicates the time of stimulation. (b) Percentage of animals that showed CMAPs indicating successful reinnervation. The number in white bars indicates the sample size while the colored bars indicate the number of animals with evoked CMAPs from at least one muscle (Mfdb or Mipd). (c) CMAP amplitude of Musculus interosseous pedis dorsalis (Mipd) on the lesioned side was normalized to the contralateral (non-lesioned) side (grey dots). (d) CMAP latency (time to first peak) of Mipd on the lesioned side and the corresponding contralateral side (grey dots). Black bars in c, d indicate mean ± SD; asterisks mark significant differences with respect to group AT (p** < 0.01; p*** < 0.001; N ≥ 5). The difference to the contralateral side within individual groups was always significant (p < 0.001). (e) Data from both muscles were pooled and CMAP amplitude plotted against latency. The graph reveals a clear negative correlation between both parameters, and recordings from individual groups are distinctly clustered.

Chapter III
74
Table 1 Compound muscle action potentials after PN repair with artificial implants Data are means ± standard deviation. The number of rats of whom CMAPs could be recorded were for AT: n = 6, ET: n = 3 G1: n = 7, G2: n = 5 (animals without CMAP recording in groups ET and G2 are excluded from latency data). CMAP amplitude of the non-lesioned nerve was set to 100 % individually.
The maturity of regenerated axons can be assessed by measuring CMAP latency, which are
considered to be influenced by axon diameters and the degree of myelination. The same relative
ranking between the experimental groups that I found with response amplitude was also observed
when comparing latencies between electrical stimulation and CMAP (Table 1). Shortest latencies
were determined in group AT. While differences between the three artificial implants were not
significant, the shorter mean latencies of construct G2 compared with G1 and ET again indicated
an advantage of the c/PCL-fiber containing matrix.
Latency and amplitude of the CMAPs were negatively correlated. The CMAPs of non-lesioned
nerves had relatively high amplitudes (many motor units were activated simultaneously)
combined with short latencies (high conduction velocities of the stimulated fibers), whereas a
combination of low amplitudes and long latencies were seen when regeneration was poor.
When plotting CMAP amplitude vs latency the graph describes a hyperbola that asymptotically
approaches the abscissa (Figure 23e). Data of the non-injured controls and the two implant groups
AT and G2, form clusters along this function, illustrating the rank order between these groups that
are characterized by successful recovery. Data points for groups ET and G1 tend to form a single
cluster at the bottom end of the function indicating an inferior recovery according to
electromyography, compared to the other treatments.
CMAP amplitude
[% of control] CMAP latency
[ms]
M. flex. dig. brev.
M. inteross. ped. dors.
M. flex. dig. brev. M. inteross. ped.
dors.
non lesioned side (CL)
100 100 3.21 ± 0.22 3.07 ± 0.31
lesion only (LO) 0 0 n.a. n.a.
autograft (AT) 41.4 ± 5.5 47.7 ± 10.3 4.67 ± 0.24 4.71 ± 0.26
empty tube (ET) 5.7 ± 8.9 10.1 ± 11.4 6.23 ± 1.03 6.11 ± 0.28
PCL-implant (G1) 17.1 ± 7.9 15.6 ± 6.9 6.65 ± 0.51 6.74 ± 0.65
C/PCL-implant (G2) 21.5 ± 15.8 22.1 ± 16.4 5.81 ± 1.01 5.89 ± 0.98

3 Results
75
3.6 Functional regeneration: recovery of motor functions
The crucial outcome with respect to PN regeneration is the recovery of appropriate motor
functions. To investigate the implications of nerve transection and the progress of regeneration
after nerve repair using different implants, behavioral experiments were performed weekly
starting 1W after surgery. At that time point, all animals exhibited paralysis and avoided the use
of the lower hind limb on the operated side. Behavioral observations of the rats confirmed the
completeness of the nerve transection and the gradual regeneration. Two quantitative measures,
toe spread on response to lifting the animal and grip strength, revealed when the animals regained
motor functions of their hind paws.
The toe spread score measures the ability to extend and abduct the digits. In the days following
surgery the affected paw showed a flat or passive posture with extended toes and reduced spread
between digits 1 and 5 compared to baseline (Figure 24 a, b). In the following weeks, toe spread
decreased further, reaching a minimum of approximately 30 % of baseline 3W after surgery. For
the animals with sciatic nerve lesion but no implant, the toe spread value remained at this value
without improvement until the end of the experiment at 12W. In the animals with nerve implants,
toe spread began to increase and by 12W had recovered to a large extent (up to 75% compared to
the contralateral paw). Compared to the non-repair group this recovery was significant for groups
AT, G1 and G2 but not for the ET group (Figure 18b).
The grip strength test measures the muscular force exerted by the digits when flexing or grasping
a metal grid. Consistent with the paralysis in the operated lower hind limb, all animals failed to
exert any additional force beyond mere friction as the paw moved across the metal bar surface
during the first weeks after surgery (i.e. below 15 N, dashed line in Figure 24c). Starting at 6W
after surgery, the first animal (in group AT) exhibited a change from an extended to a flexed
posture of the affected toes. Mean grip strength of group AT started to recover 7W after surgery
and, from then onwards, showed a steady and linear increase. The groups with artificial nerve
grafts also showed regeneration starting to recover in approximately one week intervals, with
group G2 being first, followed by G1 and ET last. In these cases the observed functional recovery
was more modest, starting later and developing with a lower slope (Figure 24c). At the endpoint
(12W after surgery) mean grip strength for all groups with the exception of ET was significantly
larger than that of the group without implant (LO, Figure 24d). This outcome was reflected in the
higher number of ET individuals compared to the other groups, that did not exhibit grip strength
above 15 N, considered as no recovery, during the time of the study. While this threshold was
reached by 100 % (11/11) of the individuals in groups AT and G1, and by 75 % (9/12) in group
G2, only 25 % (4/12) in ET exhibited a detectable increase in grip strength.

Chapter III
76
Figure 24 Behavioral analysis of nerve regeneration. Following surgery the recovery of the toe spread reflex and grip strength was investigated once per week. (a) The toe spread reflex was investigated by measuring the distance between toes 1 and 5 on the operated and non-operated legs. (b) The toe spread ratio of the operated to the non-operated side is plotted for the different conditions at the 3W (white bars) and 12W (colored bars) time point. It drops immediately after the operation, declines further to a minimum at 3W for all groups and recovers in groups AT, G1 and G2. (c) Grip strength development over time illustrates the degree of functional regeneration. A threshold of resistance without muscular activity (dashed line) was never surpassed by any rat of group LO. Plot (d) shows the grip strength before surgery (baseline) as well as 1W (white bars) and 12W (colored bars) post-surgery, respectively. Error bars indicate SEM, asterisks significant differences to the mean of group LO 12W post surgery (p** < 0.01; p*** < 0.001; N ≥ 11).

3 Results
77
3.7 Muscular atrophy and regeneration
A secondary effect of the denervation after nerve transection is muscular atrophy due to the lack
of activity. To the extent that this is caused by the degradation of proteins in muscle fibers which
survive, reinnervation can initiate new growth of muscle mass. To include different muscle fiber
types in the analysis the weight of four muscles of the lower hind limb were measured, i.e. M.
soleus, M. extensor digitorum longus, M. gastrocnemius and its antagonist the M. tibialis anterior.
(Table 2 and Figure 25). Denervated muscles lost approximately 50 % of their weight in the first
2W after nerve transection. This was similar in all experimental groups and independent of the
type of implant the animals had received. With sustained denervation, as in the non-repaired
situation (LO), muscle weight decreased further, reaching 26 % (average of all four muscles) after
12W. In animals that received one of the artificial implants this continuing decline in muscle
weight was prevented. After termination of the experiment their muscles were only slightly
smaller than 2W after surgery, and this difference was not significant. Given that reuse of re-
innervated muscles began later than 6W even in the AT group (Figure 24c), these results indicate
a recovery of muscle weight after reaching a minimum sometime between 2W and 12W. Only
animals treated with an autologous nerve graft (AT) showed considerable recovery of muscle
weights beyond the values obtained 2W after surgery. The average muscle weight in group AT
reached 71 % of the non- lesioned side.
Figure 25 Determination of muscular atrophy. Following nerve transection the denervated muscles start to atrophy. (a) Tibialis anterior muscles from the operated leg 12W post surgery of experimental groups (LO, AT, G1) exhibit different degrees of atrophy. For comparison, the corresponding contralateral muscle (CL) is shown. (b) Atrophy was quantified by determination of the weight of four muscles of the lower hind limb (see text) at 2W (white bars) and 12W (colored bars) post surgery. Muscle weight of the operated leg was normalized individually to the contralateral side. For the graph the relative weight of all four muscles of every animal was averaged. Error bars indicate SEM, asterisks depict significant difference to LO at the respective time point (p*** < 0.001; N2W ≥ 2; N12W ≥ 6).

Chapter III
78
4 Discussion
The quality of autologous nerve grafts to support functional recovery after PN injury remains
unmatched. Yet, their disadvantages, including the need for additional operations and the limited
availability of donor material create a demand for alternatives based on the use of acellular,
allogenic implants or artificial implants. In the present study, I tested the efficacy of an internally
structured scaffold to promote PN regeneration over a 15 mm sciatic nerve gap in rats. These
artificial implants contained aligned sub-micron scale PCL-fibers as longitudinal guidance cues
embedded in a gelatin matrix. It was derived from the design I documented in chapter I (see also
Kriebel et al., 2014), but with modifications regarding the gelatin hydrogel for embedding the
fibers (instead of collagen), and the choice of the surrounding tube. In its molecular composition
gelatin is closely related to collagen. I found that the gelling characteristics of gelatin facilitated
implant assembly and storage of the prepared scaffolds, while offering the possibility to sterilize it
before use. In addition, gelatin is much cheaper than collagen. For the hollow nerve conduit I used
a collagen tube instead of the previously used hand-made PCL tubes. Based on the recovery of
motor functions and on electrophysiological assessment of the regenerated nerves the structured
implants represent a significant improvement on the conventional hollow conduits and offer
possibilities for further optimization. A modification of the fibers by blending the PCL with
collagen before electrospinning produced an additional positive effect on regeneration.
Table 2 Muscle weight normalized to the corresponding muscle on the contralateral side 2 and 12 weeks after respective treatment. Data are % of individual contralateral muscle (CL) given as means ± standard deviation.
M. gastrocnemius
M. tibialis anterior
M. soleus M. extensor
digitorum longus
2
weeks 12
weeks 2
weeks 12
weeks 2
weeks 12
weeks 2
weeks 12
weeks
non lesioned side (CL)
1,38g ± 0,18g
1,16g ± 0,09g
0,51g ± 0,10g
0,40g ± 0,04g
0,13g ± 0,01g
0,12g ± 0,01g
0,12g ± 0,02g
0,09g ± 0,01g
lesion only (LO)
47,76 ± 5,20
17,30 ± 1,92
47,12 ± 3,59
24,24 ± 1,90
46,73 ± 4,08
34,69 ± 6,97
48,78 ± 6,22
29,13 ± 1,51
autograft (AT) 43,69 ±
1,43 55,95 ±
5,69 48,07 ±
0,06 72,94 ±
3,57 50,82 ±
1,39 69,50 ± 14,56
46,14 ± 0,16
86,06 ± 6,70
empty tube (ET)
44,80 ± 1,06
31,64 ± 12,06
50,46 ± 4,80
46,97 ± 19,79
51,11 ± 0,47
42,40 ± 19,92
51,63 ± 0,89
51,36 ± 18,90
PCL-implant (G1)
47,50 ± 2,60
40,65 ± 8,37
54,66 ± 4,37
49,33 ± 10,12
53,95 ± 7,35
45,48 ± 6,69
58,73 ± 0,76
60,62 ± 8,98
C/PCL-implant (G2)
48,45 ± 0,68
37,74 ± 9,91
53,76 ± 5,49
50,45 ± 16,63
51,30 ± 3,91
43,18 ± 11,60
67,30 ± 3,74
57,81 ± 18,53

4 Discussion
79
4.1 Regeneration in the artificial nerve implants
Following a PN crush lesion, the distal sections of the transected axons degenerate, and activated
Schwann cells and macrophages phagocytose cellular debris (Deumens et al., 2010). Structural
reorganization follows, characterized by a longitudinal alignment of proliferating Schwann cells
and basement membranes. These form bands of Büngner, which guide regenerating axons from
the proximal stump to targets in the periphery (Christie and Zochodne, 2013). Similar processes
promote regeneration in an autologous nerve transplant. Although the ECM and secreted factors
of an injured PN create a permissive environment for axonal growth, regeneration is initially
slow, because proximal axon endings degenerate back to the last node of Ranvier before the
metabolic changes take place that induce the formation of growth cones (Deumens et al., 2010).
Even in case of a direct end-to-end suture after sciatic nerve transection in mice it was reported
that only 17% of the axons had crossed the repair site after five days (Witzel et al., 2005).
When a large nerve gap is bridged by fluid-filled tubular implants, bands of Büngner are not
available, which complicates regeneration: Within the first week after surgery a reconnection of
the proximal and distal nerve stumps is initiated by the formation of a cord of longitudinal fibrin
(Weis et al., 1994; McDonald and Zochodne, 2003). How this first tissue bridge is produced in
detail remains to be elucidated. Several publications indicate that perineurial glia cells and
fibroblasts are involved in the process (Scaravilli, 1984; Weis et al., 1994), and my observation of
central strands filled with S100-/IIItubulin
-/MBP
- cells in regenerated nerves within artificial
conduits at 12W may reflect it.
Subsequently, Schwann cells infiltrate the lumen of the conduit. Migrating along the bridge of
connective tissue, they proliferate and deposit other ECM material including a laminin-containing
basement membrane. On this substrate regenerating axons from the proximal stump follow up
closely (Zochodne, 2012). In my experiments, axons were detectable at 2W in the medial segment
of the internally structured implants, but they were few compared to the endpoint observation at
12W. This may be expected, given that previous studies (Witzel et al., 2005) demonstrated that
axonal regeneration does not occur simultaneously but successively, with a few pioneering axons,
which are followed by consecutive waves of regenerating fibers. Based on the histological
evidence of an internal connective tissue fascicle after 12W that was surrounded by Schwann cells
and axons, I surmise that the formation of a connecting fibrin cable occurred also in the gel- and
fiber-containing implants. Although the physiological and functional data demonstrated better
regeneration in the structured implants compared to the merely fluid-filled tubes, the histological
evaluation was not suitable to demonstrate that axons or Schwann cells actually moved in contact
with these substrates. It is also possible, that the positive effect of the internal structure was
primarily based on aiding the formation and stabilization of the fibrin cable, the very first step in
regeneration, as suggested by Koh et al. (2010). Axon counting revealed that not all axons entered
into the artificial implants but that axonal branching must have occurred when axons crossed the

Chapter III
80
distal coaptation site and regrew into the denervated distal stump. Branching of fibers was also
observed within the autologous transplants at the proximal coaptation site, as reported before
(Jenq and Coggeshall, 1985; Miyamoto et al., 1985; Witzel et al., 2005). This observation
suggests that branching of axons is induced through contact or trophic factors released from a
nerve undergoing Wallerian degeneration, which could be either the distal stump or an autologous
nerve graft.
Since the time needed for reinnervation of a target tissue depends on the type, -size and location
of injury, it is difficult to compare the speed of recovery in the present implants with data from
the literature (Cao et al., 2011; Yu et al., 2011; van Neerven et al., 2012; Kim et al., 2014).
Nevertheless, my analyses of grip strength recovery and muscular atrophy are in accordance with
previous studies. After making a 10 mm sciatic nerve lesion in rats at mid-thigh level and
subsequent repair with an autologous nerve graft, a duration of 6-7 weeks until reinnervation of
the gastrocnemius muscle has been reported (Korte et al., 2011; Kim et al., 2014). Other
researchers (McCallister et al., 2001) autografted a 10 mm gap in the rat sciatic nerve and
observed a remaining muscle weight of the M. tibialis anterior of 62 % at 12w after injury (73%
in the present study) while the controls without repair had 29 % (24 % in the present study). The
better results of the present autograft is likely to be due to the fact that McCallister and colleagues
reversed the orientation of their short nerve graft before re-implantation, whereas larger lesions of
the present study and also those of Bozkurt et al. (2012) made the reversal of the graft impractical,
and therefore was not performed. Given the general agreement with published data concerning
autologous transplants the data about the structured artificial implants presented here may be put
into perspective. Thus, the novel PCL- and c/PCL-fiber containing gels represent a significant
improvement on standard implants without internal guidance structure, although they do not reach
the quality of autologous nerve grafts or allografts (Brooks et al., 2012).
4.2 The importance of an internal guidance structure in artificial implants
The only FDA and CE approved alternatives to autologous transplants are simple tubes
applicable for short distance repair (less than 2-3 cm). As corroborated here, they are inferior to
autologous transplants, and the integration of molecular and structural cues for reaching a
comparable functional outcome remains a challenge (Lundborg, 2000; Isaacs, 2010; Konofaos
and Ver Halen, 2013). Since fluid-filled tubes provide no substrate for axonal growth in their
lumen, it is only after some time, when a continuous tissue bridge has formed and infiltration of
perineurial and Schwann cells has occurred, that axons can start to grow into the implant (Lewis
and Kucenas, 2014). On the other hand, the potential for successful regeneration diminishes with
the time after injury (Xu et al., 2014), implying that the presence of a growth substrate may be an
important factor for functional recovery within a critical window of opportunity. I therefore
attribute the better functional outcome in my experimental groups to the incorporation of

4 Discussion
81
electrospun fibers that serve as longitudinal guidance cues (as shown in vitro: Schnell et al., 2007;
Kriebel et al., 2014) and not to the gelatin substrate surrounding them. This statement does not
diminish the importance of the choice of the hydrogel and its physical properties. Generally, cells
need surfaces to grow on and migrate, and in situations when the hydrogel forms a distinct
boundary with the surrounding medium, as observed with alginate and fibrin gels, cells tend to
grow along this interface rather than penetrating the gel (Mohanna et al., 2003; Gerardo-Nava et
al., 2014). This prevents regeneration within the intended structure. To avoid such effects I
decided to use non-fixed gelatin as a support for the fiber arrays. Kept cold, it allowed handling
and incorporation of the fibers into the tube, while at body temperature it became liquid again and
thus did not pose an obstacle for cells involved in regeneration.
4.3 Implementation of biological signals in synthetic cell-free nerve grafts
Under the condition of nerve crush with intact perineurial sheaths (axonotmesis) multiple
molecular signals on the ECM and molecules released from the Schwann cells induce axonal
regeneration. Hydrogels and electrospun fibers can provide a platform to present selected signals
in artificial implants (Dalton and Mey, 2009). Soluble growth factors, e.g. nerve growth factor
(Xu et al., 2003) and glial growth factor (Mohanna et al., 2003) were successfully used in this
context. Another option consists in functionalization of implanted surfaces with activators of
integrins and cell adhesion molecules (Klinkhammer et al., 2010; Grafahrend et al., 2011).
Already the implementation of one type of integrin-activating peptide (RGDS) on electrospun
fibers increased their ability to induce Schwann cell migration and axonal growth (Bockelmann et
al., 2011).
Considering that the implantation of living cells has serious practical disadvantages, such as
additional operations for harvesting endogenous cells or immune suppression, my strategy
focuses on the development of cell-free artificial nerve bridges. Two recently developed acellular
products that contain internal guidance structures are the Avance® and Neuromaix® nerve
implants. Both incorporate some biological activity because they are derived from natural
materials. The Avance (AxoGen Inc., Alachua, FL) nerve graft is marketed as the “only off-the-
shelf commercially available processed nerve allograft intended for the surgical repair of PN
discontinuities”. Recovered from human PN, which are cleaned from cellular debris, the grafts
preserve the ECM structure of the PN and have been reported to show similar axon regenerating-
promoting properties as autologous nerve grafts (Brooks et al., 2012). Neuromaix (Matricel
GmbH; Herzogenrath, Germany) consists of an orientated scaffold with longitudinal pores that is
produced from porcine collagen type I using a patented freezing protocol. Animal experiments
with this scaffold were also successful (Bozkurt et al., 2012), and a clinical phase 1 trial is
currently under way (Bozkurt et al., 2014).

Chapter III
82
Several disadvantages with these products make the further development of synthetic nerve grafts
with specific biological signals desirable: The AxoGen scaffolds still require harvesting of human
tissue and result in limited control over the material properties. The Matricel scaffold has mostly
the signaling properties of collagen type I via α2β1 and integrins (Bozkurt et al., 2007; Knight et
al., 1999). While the constructs presented here are not likely to be superior to these products, my
approach opens the possibility to use completely synthetic hydrogels and fibers and to endow
them with signaling peptides designed to activate integrin receptors. Further improvements of the
present scaffold may be achieved by additional functionalization of the electrospun fibers
(Bockelmann et al., 2011; Grafahrend et al., 2011) and/or the application of growth factors that
are gradually released (Lee, 2003; Solorio and Zwolinski, 2010)
5 Conclusion
Based on the recovery of motor functions and on physiological assessment of the regenerated
nerves the novel structured implants represent an improvement on conventional hollow conduits
and offer possibilities for further optimization. By elaboration of structural properties (e.g. fiber
density) or by providing more information on the interaction of the involved cell types with
biological signals it should be possible to further narrow the functional gap between artificial
implants and autologous nerve grafts.

Comparison to other studies
83
General Discussion
Peripheral nerve lesions not only may have drastic consequences for the affected individual but, at
the level of the population, also considerable socio-economic impact. This is particularly due to
the high follow up costs generated by these injuries. Reasons for that are, for example, the long
term physical therapy the patient has to undergo to support functional recovery. Furthermore,
until this is achieved to a satisfactory degree, if at all, the patient will not be at his full
professional capacity. To facilitate best possible recovery for the patient and reduce the total costs
for the society, it is necessary to have treatments available with a prospect for recovery at least on
the level of the autologous nerve graft. However, as stated before, a direct end-to-end
neurorrhaphy is not always possible, nor is an autologous nerve graft always available. Hence, at
the moment there are cases were the physician in charge has to make use of alternatives that
might be less promising than the aforementioned solutions, e.g. an artificial implant. Therefore,
there is great interest in improving the performance of artificial implants especially for the
treatment of bigger nerve gaps.
To achieve this, different strategies can be applied, as elaborated in Chapter I of this thesis. The
approach presented here aimed for the integration of sub-micron scale fibers as a guidance
structure into a hollow conduit to obtain a cell-free artificial nerve implant with improved
performance, compared to the hitherto available conventional implants. For this, I designed and
established an electrospinning device for the production of 3D arrays of aligned fibers. These
arrays were embedded in a hydrogel matrix to form a scaffold with an internal guidance structure.
After showing via in vitro experiments that the fibers keep their guidance properties inside the
hydrogel, fiber/gel constructs were incorporated into collagen tubes to obtain implantable devices.
Subsequently these were used to bridge a 15 mm rat sciatic nerve gap in vivo. It showed that, even
though the autologous nerve graft still performed notably better, the incorporation of the fiber/gel
constructs proved to be beneficial for the outcome of regeneration compared to hollow collagen
tubes. Thus I consider this approach a promising basis for further improvements. However, there
have also been other approaches in the past years, of which a representative selection will be
presented briefly in the following.
Comparison to other studies
The suitability of electrospun fibers as guidance structures for peripheral nerve regeneration has
also been demonstrated in other studies. In two consecutive in vivo studies with rats by Ravi V.
Bellamkonda‘s group (Kim et al., 2008; Clements et al., 2009), films of longitudinally aligned
fibers from PAN-MA were incorporated into polysulfone nerve conduits to successfully bridge a
tibial nerve gap of 17 and 14 mm, respectively. In the first study they also performed nerve repair
with an empty (saline-filled) polysulfone conduit and an autologous nerve graft, which, to a

General Discussion
84
certain extent, allows a comparison between their approach and mine. In their study, as in mine,
the best regeneration was found in the group of animals that was treated with an autologous nerve
graft. This was relatively closely followed by the aligned fiber-film group, and the differences
between autograft group and aligned fiber-film group seem smaller than in the present study. This
observation can be explained by several reasons. First of all, the more distal location of the nerve
defect (tibial versus sciatic nerve) generally has a better prediction for functional recovery. In
addition, the outcome of regeneration was assessed only 16 weeks after implantation, which is
four weeks later than in the present study. Surprisingly, in contrast to my study, they did not find
any signs of regeneration in the empty conduits even though the nerve gap to be bridged was only
2 mm longer in their study (17 versus 15 mm). This creates the impression that differences
between autograft group and aligned fiber-film group are smaller than observed in the present
study, allthough this might not be true. However, the data in general suggest a slightly better
performance of their approach compared to mine.
Another example for the successful application of electrospun fibers to perform a PN repair in rats
is the work by Jha et al. (2011). The unique feature of this work is the very convenient procedure
to obtain the internally structured implants. By electrospinning onto two rounded target electrodes
a bundle of aligned fibers was created. In a second step an exterior coating was added to the
construct by electrospraying PGA:PLA copolymer onto the outside of the bundle and thus
forming the final construct. Unfortunately they only used a nerve gap size of 10 mm for biological
testing and stopped the experiment seven weeks after implantation. At this time point the animals
showed a withdrawal reflex in response to foot pinch, indicating sensory reinnervation. This
coincides with the regain of grip strength in the present study, but a more detailed comparison is
hindered by the differences in experimental conditions.
Besides the application of fibers, specifically processed hydrogels have also been used to
implement directional cues into nerve conduits. One way to achieve this was demonstrated by
Yao et al. (2010). They produced collagen nerve conduits with multiple internal channels with
diameters of approximately 500 µm to limit axonal dispersion inside the conduit and thus reduce
inappropriate target reinnervation. While this aspect of the study seemed to be a success, as they
found that even fewer neurons showed double projections to the tibial and the peroneal branch in
the 4-channel group as in the autograft group, the actual regenerative outcome did not differ
between their implants and a conventional hollow collagen conduit when used to repair a 10 mm
sciatic nerve gap in rats.
An alternative approach was used by Bozkurt et al. (2012) who employed a conduit material
called Perimaix. This material, developed by Matricel GmbH (Herzogenrath, Germany), is
derived from porcine collagen that was further processed through cross-linking and directional
freezing to create longitudinal pores for cellular and axonal guidance (Bozkurt et al., 2009).
Bozkurt and coworker used the Perimaix conduit to bridge a 2 cm sciatic nerve gap in rats. This is

Possible augmentation of implant design
85
expected to be a challenging gap size for the application of artificial implants and probably the
largest possible distance to repair in this animal model. Nevertheless they were able to show that
six weeks after implantation axons had crossed the complete implant and depicted the same
diameters as in the autograft group. However, axon density and myelin thickness was reduced to
approximately 50 and 66 %, respectively, of that determined for the autograft group
demonstrating that the nerve transplant was still superior to the artificial scaffold. Functional
recovery was not assessed in this study. A notable difference to most other studies, which might
have had a positive effect on regeneration is the absence of a conventional tubing material.
Instead they opened the nerves epineurium by a longitudinal incision at the site of nerve defect
and later on used it to ensheath the inserted implant, embedding it in a much more natural
environment. Again, the differences in the experimental conditions in addition to the lack of a
comparable negative control (empty tube) do not permit a detailed comparison between this and
the study presented here. However the quality of axonal regeneration across such a defect size is
impressive and led to consecutive clinical testing of the material in humans (Bozkurt et al., 2014).
To conclude, one first of all one has to recognize that comparison between different studies is
difficult in general. Even though the selection presented here is restricted to only one of several
animal species (e.g. rabbit, dog, sheep) used for PN repair studies, there is still a considerable
variability of experimental conditions impeding detailed comparison. These are, for example, the
choice of nerve, defect size, time of recovery and methods for its assessment. Thus, even though
there is a lot of research going on in the field, it is difficult to rank the success of my own
approach in relation to alternative ones. A demand for standardization of peripheral nerve repair
studies with rodents has already been voiced by others (Bozkurt et al., 2012). This would greatly
help to identify the most promising strategies, discard others and accelerate progress in this field
of research.. Apart from that, the selection of studies presented here suggests that my implant
design is not generally superior but competitive to other approaches with internal guidance
structures. However, it seems that the incorporation of physical/topographical guidance cues only
is not sufficient to achieve results on the level of autologous nerve grafts. Further augmentation of
functionality seems to be necessary to obtain a true alternative to the current “gold standard“, the
autologous nerve graft.
Possible augmentation of implant design
The arrangement of guidance cues can influence an implant’s performance in terms of
regeneration as shown, for example, by Clements et al. (2009). Thus a further improvement of my
implant’s performance by adjustment of certain parameters is conceivable, e.g. by increasing the
amount of fibers and their dispersion inside the implant. However, in the past decade there has
been a variety of different design approaches for artificial nerve conduits with or without

General Discussion
86
incorporated physical/topographical guidance cues, and, to my knowledge, none of them was able
to compete with the autologous nerve graft with respect to regeneration and functional recovery in
gap sizes bigger than 1 cm. Apparently the incorporation of physical/topographical guidance cues
has beneficial effect, but further augmentation, e.g. through implementation of biological
signaling, seems necessary to obtain results equivalent to what can be achieved with autologous
nerve grafts.
There are several possibilities to equip an implant with biological signals that support axonal
regeneration. An obvious way to achieve this is preseeding the conduit with appropriate cells
before implantation. The positive effect of preseeded cells was shown in numerous studies with a
variety of cell types (di Summa et al., 2010; Ding et al., 2010; Bozkurt et al., 2012; Allodi et al.,
2014; Beigi et al., 2014). As an example, by preseeding the Perimaix conduits with Schwann cells
for repairing a 2 cm sciatic nerve lesion in rats Bozkurt et al. (2012) achieved regeneration that
was almost as effective as when an autologous nerve graft was used. However, for the application
in humans, which is the final therapeutic goal of all this research, preseeded conduits involve
certain complications, as discussed earlier (Chapter I 3.4). Nevertheless, it is conceivable to
mimic certain features those cells exhibit to support regeneration.
One of those features is the secretion of ECM molecules. The ECM is equipped with a variety of
molecular components that can be recognized by axonal receptors, e.g. integrins. Hence, a way to
improve the performance of artificial implants is the incorporation of selected ECM components.
In the present study, I used a rather simple approach to achieve an enhancement of the fibers
guidance properties, namely by blending the PCL solution with collagen, a major ECM
constituent. This was shown to have positive effects on Schwann cell migration and axonal
regeneration in vitro (Schnell et al., 2007; Mey et al., 2012) and also led to slightly better
performance in my in vitro experiments compared to pure PCL. More sophisticated methods
involve chemical modification of materials by covalent binding of signaling peptides, e.g. an
RGD sequence (arginine-glycine-aspartic acid), which cells can bind to via certain integrins (Koh
et al., 2008; Bockelmann et al., 2011; Grafahrend et al., 2011). However, an advantage of
covalent binding of signaling peptides to the guidance structure over mere blending of petides to
the electrospinning solution could not be shown yet.
Another beneficial feature cells exhibit to support regeneration is the release of growth factors.
There is a variety of different growth factors known to support the regeneration of axons after
PNI, e.g. NGF, GDNF, NT-3, CNTF etc. (Kokai et al., 2010, 2011; Allodi et al., 2011).
Depending on the factor, the positive effect on regeneration is mediated through different
mechanisms, for example the promotion of axonal outgrowth or neuronal survival (de Ruiter et
al., 2009). Thus, by the combination of different growth factors, synergistic effects are to be
expected that could lead to better support than the application of only a single type of growth
factor (Madduri et al., 2010a). However more comprehensive analysis of the dose-dependency of

Possible augmentation of implant design
87
different factors and their possible interactions are desirable to choose the best combination of
growth factors for an artificial nerve conduit. Possibilities for the incorporation of growth factors
into artificial conduits have already been presented in a number of studies. Usually systems for
the sustained delivery of factors over a longer period of time are preferred. This can be achieved
by encapsulating the factors into slowly degradable materials, e.g. PCL or cross-linked gelatin
(Xu et al., 2003; Moore et al., 2006; Chang, 2009; Valmikinathan et al., 2009; Solorio and
Zwolinski, 2010; Kokai et al., 2011; Liu et al., 2011).
Conclusion
As shown by the experiments presented in this thesis, I successfully designed and manufactured a
cell-free artificial nerve conduit for the support of peripheral nerve regeneration. The new design
with the incorporated fiber/gel scaffold represents an improvement over a hollow collagen
conduit. As the performance of my implant design with respect to the support of regeneration and
functional recovery is generally on a comparable level with other designs presented so far, further
effort in advancing the design for the use in humans is justified. To reach this goal I suggest the
following steps to be taken.
First of all, the physical structure of the internal scaffold should be optimized. The distribution of
fibers inside the implants, used in the present study, was not homogenous. The fibers were mainly
localized in the center of the scaffold in an area of approximately 1 mm width and 0.5 mm heights
(when looking at a cross section). Here, one should strive for an adjustment of the procedure for
fiber embedding and incorporation into the tube to obtain a more homogenous distribution of
fibers throughout the whole cross section of the conduit. In my opinion a moderate increase in
fiber density would also be favorable. This might prove beneficial for regeneration, while
detrimental effects are not to be expected. At least I conclude this from comparisons with other
studies were the process for manufacturing of the implants suggests a final fiber density that was
higher than in the case presented here, e.g. (Jha et al., 2011).
Secondly, biological signaling should be implemented into the conduit. To obtain the best
possible results, I suggest to use a combination of functionalizing the fibers through modification
of their surface with ECM molecules, as suggested by (Grafahrend et al., 2011), and the
incorporation of a release system for growth factors. For this purpose, the use of microspheres
encapsulating the growth factors seems reasonable, since they are already widely used in this
context and can be incorporated into the conduit wall (Kokai et al., 2010). Additionally I suggest
the use of cocktail of different growth factor to make use of synergistic effects. However ideal
combinations and concentrations remain to be determined.
Finally, before testing the optimized implants in humans, a second animal study in vivo, using the
optimized implants, would be desirable. In this case, I suggest to choose a more challenging gap

Conclusion
88
size to be repaired. This way one should obtain more distinct differences between experimental
groups, which eases the evaluation of the implants performance. However this requires a bigger
animal model for example, e.g. Beagle dogs (Wang et al., 2005; Ding et al., 2010; Huang et al.,
2013). Also, in contrast to the present study, the assessment of functional recovery should be
expanded to sensory function to give a more detailed insight into different aspects of regeneration.
A successful completion of the suggested measures would, in my opinion, justify the start of
clinical trials in humans.

89
References
005/010 S3-Leitlinie: Versorgung peripherer Nervenverletzungen (2013)
www.awmf.org/leitlinien/detail/ll/005-010.html
Aberg M, Ljungberg C, Edin E, Millqvist H, Nordh E, Theorin A, Terenghi G, Wiberg M (2009)
Clinical evaluation of a resorbable wrap-around implant as an alternative to nerve repair: a
prospective, assessor-blinded, randomised clinical study of sensory, motor and functional
recovery after peripheral nerve repair. J Plast Reconstr Aesthet Surg 62:1503–1509.
Aebischer P, Salessiotis a N, Winn SR (1989) Basic fibroblast growth factor released from
synthetic guidance channels facilitates peripheral nerve regeneration across long nerve gaps.
J Neurosci Res 23:282–289.
Agnew SP, Dumanian G a (2010) Technical use of synthetic conduits for nerve repair. J Hand
Surg Am 35:838–841.
Allodi I, Guzmán-Lenis M-S, Hernàndez J, Navarro X, Udina E (2011) In vitro comparison of
motor and sensory neuron outgrowth in a 3D collagen matrix. J Neurosci Methods 198:53–
61.
Allodi I, Mecollari V, González-Pérez F, Eggers R, Hoyng S, Verhaagen J, Navarro X, Udina E
(2014) Schwann cells transduced with a lentiviral vector encoding Fgf-2 promote motor
neuron regeneration following sciatic nerve injury. Glia 62:1736–1746.
Ashley WW, Weatherly T, Park TS (2006) Collagen nerve guides for surgical repair of brachial
plexus birth injury. J Neurosurg 105:452–456.
Bailey SB, Eichler ME, Villadiego A, Rich KM (1993) The influence of fibronectin and laminin
during Schwann cell migration and peripheral nerve regeneration through silicon chambers.
J Neurocytol 22:176–184.
Barnett MW, Fisher CE, Perona-Wright G, Davies JA (2002) Signalling by glial cell line-derived
neurotrophic factor (GDNF) requires heparan sulphate glycosaminoglycan. J Cell Sci
115:4495–4503.
Beigi M-H, Ghasemi-Mobarakeh L, Prabhakaran MP, Karbalaie K, Azadeh H, Ramakrishna S,
Baharvand H, Nasr-Esfahani M-H (2014) In vivo integration of poly(ε-caprolactone)/gelatin
nanofibrous nerve guide seeded with teeth derived stem cells for peripheral nerve
regeneration. J Biomed Mater Res A 102:4554–4567.
Belkas JS, Munro CA, Shoichet MS, Johnston M, Midha R (2005a) Long-term in vivo
biomechanical properties and biocompatibility of poly(2-hydroxyethyl methacrylate-co-
methyl methacrylate) nerve conduits. Biomaterials 26:1741–1749.
Belkas JS, Munro CA, Shoichet MS, Midha R (2005b) Peripheral nerve regeneration through a
synthetic hydrogel nerve tube. Restor Neurol Neurosci 23:19–29.
Bertleff MJOE, Meek MF, Nicolai J-PA (2005) A prospective clinical evaluation of
biodegradable neurolac nerve guides for sensory nerve repair in the hand. J Hand Surg Am
30:513–518.

References
90
Bian Y-Z, Wang Y, Aibaidoula G, Chen G-Q, Wu Q (2009) Evaluation of poly(3-
hydroxybutyrate-co-3-hydroxyhexanoate) conduits for peripheral nerve regeneration.
Biomaterials 30:217–225.
Bini T, Gao S, Tan T, Wang S, Lim A, Hai L, Ramakrishna S (2004) Electrospun poly(L-lactic-
co-glycolide) biodegradable polymer nanofibre tubes for peripheral nerve regeneration.
Nanotechnology 15:1459.
Bixby JL, Lilien J, Reichardt LF (1988) Identification of the major proteins that promote neuronal
process outgrowth on Schwann cells in vitro. J Cell Biol 107:353–361.
Bockelmann J, Klinkhammer K, von Holst A, Seiler N, Faissner A, Brook GA, Klee D, Mey J
(2011) Functionalization of electrospun poly(ε-caprolactone) fibers with the extracellular
matrix-derived peptide GRGDS improves guidance of schwann cell migration and axonal
growth. Tissue Eng Part A 17:475–486.
Boyd JG, Gordon T (2003) Neurotrophic factors and their receptors in axonal regeneration and
functional recovery after peripheral nerve injury. Mol Neurobiol 27:277–324.
Bozkurt A, Brook G a, Moellers S, Lassner F, Sellhaus B, Weis J, Woeltje M, Tank J, Beckmann
C, Fuchs P, Damink LO, Schügner F, Heschel I, Pallua N (2007) In vitro assessment of
axonal growth using dorsal root ganglia explants in a novel three-dimensional collagen
matrix. Tissue Eng 13:2971–2979.
Bozkurt A, Deumens R, Beckmann C, Olde Damink L, Schügner F, Heschel I, Sellhaus B, Weis
J, Jahnen-Dechent W, Brook G a, Pallua N (2009) In vitro cell alignment obtained with a
Schwann cell enriched microstructured nerve guide with longitudinal guidance channels.
Biomaterials 30:169–179.
Bozkurt A, Lassner F, O’Dey D, Deumens R, Böcker A, Schwendt T, Janzen C, Suschek C V,
Tolba R, Kobayashi E, Sellhaus B, Tholl S, Eummelen L, Schügner F, Damink LO, Weis J,
Brook GA, Pallua N (2012) The role of microstructured and interconnected pore channels in
a collagen-based nerve guide on axonal regeneration in peripheral nerves. Biomaterials
33:1363–1375.
Bozkurt A, van Neerven SGA, Claeys KG, O’Dey DM, Sudhoff A, Brook G a, Sellhaus B,
Schulz JB, Weis J, Pallua N (2014) The proximal medial sural nerve biopsy model: a
standardised and reproducible baseline clinical model for the translational evaluation of
bioengineered nerve guides. Biomed Res Int 2014:11.
Braga-Silva J (1999) The use of silicone tubing in the late repair of the median and ulnar nerves in
the forearm. J Hand Surg Br 24:703–706.
Brenner MJ, Hess JR, Myckatyn TM, Hayashi A, Hunter DA, Mackinnon SE (2006) Repair of
motor nerve gaps with sensory nerve inhibits regeneration in rats. Laryngoscope 116:1685–
1692.
Brooks DN, Weber R V, Chao JD, Rinker BD, Zoldos J, Robichaux MR, Ruggeri SB, Anderson
KA, Bonatz EE, Wisotsky SM, Cho MS, Wilson C, Cooper EO, Ingari J V, Safa B, Parrett
BM, Buncke GM (2012) Processed nerve allografts for peripheral nerve reconstruction: a
multicenter study of utilization and outcomes in sensory, mixed, and motor nerve
reconstructions. Microsurgery 32:1–14.

91
Brown RE, Erdmann D, Lyons SF, Suchy H (1996) The use of cultured Schwann cells in nerve
repair in a rabbit hind-limb model. J Reconstr Microsurg 12:149–152.
Cai J, Peng X, Nelson KD, Eberhart R, Smith GM (2005) Permeable guidance channels
containing microfilament scaffolds enhance axon growth and maturation. J Biomed Mater
Res A 75:374–386.
Cao H, Liu T, Chew SY (2009) The application of nanofibrous scaffolds in neural tissue
engineering. Adv Drug Deliv Rev 61:1055–1064.
Cao J, Sun C, Zhao H, Xiao Z, Chen B, Gao J, Zheng T, Wu W, Wu S, Wang J, Dai J (2011) The
use of laminin modified linear ordered collagen scaffolds loaded with laminin-binding
ciliary neurotrophic factor for sciatic nerve regeneration in rats. Biomaterials 32:1–10.
Carroll SL, Miller ML, Frohnert PW, Kim SS, Corbett JA (1997) Expression of neuregulins and
their putative receptors, ErbB2 and ErbB3, is induced during Wallerian degeneration. J
Neurosci 17:1642–1659.
Ceballos D, Navarro X, Dubey N, Wendelschafer-Crabb G, Kennedy WR, Tranquillo RT (1999)
Magnetically aligned collagen gel filling a collagen nerve guide improves peripheral nerve
regeneration. Exp Neurol 158:290–300.
Chamberlain LJ, Yannas I V, Hsu HP, Strichartz G, Spector M (1998) Collagen-GAG substrate
enhances the quality of nerve regeneration through collagen tubes up to level of autograft.
Exp Neurol 154:315–329.
Chang CJ (2009) Effects of nerve growth factor from genipin-crosslinked gelatin in
polycaprolactone conduit on peripheral nerve regeneration - In vitro and in vivo. J Biomed
Mater Res - Part A.
Chew SY, Mi R, Hoke A, Leong KW (2007) Aligned Protein-Polymer Composite Fibers Enhance
Nerve Regeneration: A Potential Tissue-Engineering Platform. Adv Funct Mater 17:1288–
1296.
Chew SY, Wen J, Yim EKF, Leong KW (2005) Sustained release of proteins from electrospun
biodegradable fibers. Biomacromolecules 6:2017–2024.
Chiono V, Vozzi G, Vozzi F, Salvadori C, Dini F, Carlucci F, Arispici M, Burchielli S, Di Scipio
F, Geuna S, Fornaro M, Tos P, Nicolino S, Audisio C, Perroteau I, Chiaravalloti A,
Domenici C, Giusti P, Ciardelli G (2009) Melt-extruded guides for peripheral nerve
regeneration. Part I: poly(epsilon-caprolactone). Biomed Microdevices 11:1037–1050.
Christie KJ, Zochodne D (2013) Peripheral axon regrowth: new molecular approaches.
Neuroscience 240:310–324.
Ciaramitaro P, Mondelli M, Logullo F, Grimaldi S, Battiston B, Sard A, Scarinzi C, Migliaretti G,
Faccani G, Cocito D (2010) Traumatic peripheral nerve injuries: epidemiological findings,
neuropathic pain and quality of life in 158 patients. J Peripher Nerv Syst 15:120–127.
Citri A, Skaria KB, Yarden Y (2003) The deaf and the dumb: the biology of ErbB-2 and ErbB-3.
Exp Cell Res 284:54–65.

References
92
Clements IP, Kim Y, English AW, Lu X, Chung A, Bellamkonda R V (2009) Thin-film enhanced
nerve guidance channels for peripheral nerve repair. Biomaterials 30:3834–3846.
Corcoran J, Maden M (1999) Nerve growth factor acts via retinoic acid synthesis to stimulate
neurite outgrowth. Nat Neurosci 2:307–308.
Corcoran J, Shroot B, Pizzey J, Maden M (2000) The role of retinoic acid receptors in neurite
outgrowth from different populations of embryonic mouse dorsal root ganglia. J Cell Sci
113 ( Pt 1:2567–2574.
Dahlin L, Lundborg G (2001) The use of silicone tubing in the late repair of the median and ulnar
nerves in the forearm. J Hand Surg Br 26:393–394.
Dalton PD, Mey J (2009) Neural interactions with materials. Front Biosci 14:769–795.
De Ruiter GC, Spinner RJ, Malessy MJA, Moore MJ, Sorenson EJ, Currier BL, Yaszemski MJ,
Windebank AJ (2008) Accuracy of motor axon regeneration across autograft, single-lumen,
and multichannel poly(lactic-co-glycolic acid) nerve tubes. Neurosurgery 63:144–153;
discussion 153–155.
De Ruiter GCW, Malessy MJA, Yaszemski MJ, Windebank AJ, Spinner RJ (2009) Designing
ideal conduits for peripheral nerve repair. Neurosurg Focus 26:E5.
Desaphy JF, Pierno S, Léoty C, George AL, De Luca A, Camerino DC (2001) Skeletal muscle
disuse induces fibre type-dependent enhancement of Na(+) channel expression. Brain
124:1100–1113.
Deumens R, Bozkurt A, Meek MF, Marcus MAE, Joosten EAJ, Weis J, Brook GA (2010)
Repairing injured peripheral nerves: Bridging the gap. Prog Neurobiol 92:245–276.
Di Summa PG, Kingham PJ, Raffoul W, Wiberg M, Terenghi G, Kalbermatten DF (2010)
Adipose-derived stem cells enhance peripheral nerve regeneration. J Plast Reconstr Aesthet
Surg 63:1544–1552.
Ding F, Wu J, Yang Y, Hu W, Zhu Q, Tang X, Liu J, Gu X (2010) Use of tissue-engineered nerve
grafts consisting of a chitosan/poly(lactic-co-glycolic acid)-based scaffold included with
bone marrow mesenchymal cells for bridging 50-mm dog sciatic nerve gaps. Tissue
Engeneering Part A 16:3779–3790.
Dodla MC, Bellamkonda R V. (2008) Differences between the effect of anisotropic and isotropic
laminin and nerve growth factor presenting scaffolds on nerve regeneration across long
peripheral nerve gaps. Biomaterials 29:33–46.
Dombrowski MA, Sasaki M, Lankford KL, Kocsis JD, Radtke C (2006) Myelination and nodal
formation of regenerated peripheral nerve fibers following transplantation of acutely
prepared olfactory ensheathing cells. Brain Res 1125:1–8.
Donoghoe N, Rosson GD, Dellon AL (2007) Reconstruction of the human median nerve in the
forearm with the NeurotubeTM
. Microsurgery 27:595–600.
Dubey N, Letourneau PC, Tranquillo RT (1999) Guided neurite elongation and schwann cell
invasion into magnetically aligned collagen in simulated peripheral nerve regeneration. Exp
Neurol 158:338–350.

93
Ellis-Behnke RG, Liang Y-X, You S-W, Tay DKC, Zhang S, So K-F, Schneider GE (2006) Nano
neuro knitting: peptide nanofiber scaffold for brain repair and axon regeneration with
functional return of vision. Proc Natl Acad Sci U S A 103:5054–5059.
Emel E, Ergün SS, Kotan D, Gürsoy EB, Parman Y, Zengin A, Nurten A (2011) Effects of
insulin-like growth factor-I and platelet-rich plasma on sciatic nerve crush injury in a rat
model. J Neurosurg 114:522–528.
Eser F, Aktekin LA, Bodur H, Atan C (2009) Etiological factors of traumatic peripheral nerve
injuries. Neurol India 57:434–437.
Fan W, Gu J, Hu W, Deng A, Ma Y, Liu J, Ding F, Gu X (2008) Repairing a 35-mm-long median
nerve defect with a chitosan/PGA artificial nerve graft in the human: A case study.
Microsurgery 28:238–242.
Farole A, Jamal BT (2008) A bioabsorbable collagen nerve cuff (NeuraGen) for repair of lingual
and inferior alveolar nerve injuries: a case series. J Oral Maxillofac Surg 66:2058–2062.
Forsten-Williams K, Chu CL, Fannon M, Buczek-Thomas JA, Nugent MA (2008) Control of
growth factor networks by heparan sulfate proteoglycans. Ann Biomed Eng 36:2134–2148.
Geiger B, Bershadsky A, Pankov R, Yamada KM (2001) Transmembrane crosstalk between the
extracellular matrix--cytoskeleton crosstalk. Nat Rev Mol Cell Biol 2:793–805.
Gelain F, Horii A, Zhang S (2007) Designer self-assembling peptide scaffolds for 3-d tissue cell
cultures and regenerative medicine. Macromol Biosci 7:544–551.
Gelain F, Unsworth LD, Zhang S (2010) Slow and sustained release of active cytokines from self-
assembling peptide scaffolds. J Control Release 145:231–239.
George PM, Saigal R, Lawlor MW, Moore MJ, LaVan DA, Marini RP, Selig M, Makhni M,
Burdick JA, Langer R, Kohane DS (2009) Three-dimensional conductive constructs for
nerve regeneration. J Biomed Mater Res A 91:519–527.
Gerardo-nava J, Führmann T, Klinkhammer K, Seiler N, Mey J, Klee D, Möller M, Dalton PD,
Brook GA (2009) Human neural cell interactions with orientated electrospun nanofibers in
vitro. Nanomedicine 4:11–30.
Gerardo-Nava J, Hodde D, Katona I, Bozkurt A, Grehl T, Steinbusch HWM, Weis J, Brook GA
(2014) Spinal cord organotypic slice cultures for the study of regenerating motor axon
interactions with 3D scaffolds. Biomaterials 35:4288–4296.
Glazner GW, Lupien S, Miller JA, Ishii DN (1993) Insulin-like growth factor II increases the rate
of sciatic nerve regeneration in rats. Neuroscience 54:791–797.
Gomez N, Schmidt CE (2007) Nerve growth factor-immobilized polypyrrole: bioactive
electrically conducting polymer for enhanced neurite extension. J Biomed Mater Res A
81:135–149.
Gordon T (2010) The physiology of neural injury and regeneration: The role of neurotrophic
factors. J Commun Disord 43:265–273.

References
94
Grafahrend D, Heffels K-H, Beer M V, Gasteier P, Möller M, Boehm G, Dalton PD, Groll J
(2011) Degradable polyester scaffolds with controlled surface chemistry combining minimal
protein adsorption with specific bioactivation. Nat Mater 10:67–73.
Gu X, Ding F, Yang Y, Liu J (2011) Construction of tissue engineered nerve grafts and their
application in peripheral nerve regeneration. Prog Neurobiol 93:204–230.
Guénard V, Kleitman N, Morrissey TK, Bunge RP, Aebischer P (1992) Syngeneic Schwann cells
derived from adult nerves seeded in semipermeable guidance channels enhance peripheral
nerve regeneration. J Neurosci 12:3310–3320.
Guo J, Su H, Zeng Y, Liang Y-X, Wong WM, Ellis-Behnke RG, So K-F, Wu W (2007)
Reknitting the injured spinal cord by self-assembling peptide nanofiber scaffold.
Nanomedicine 3:311–321.
Hajosch R, Grupp L, Nichterwitz S, Schlosshauer B (2010) A novel microsurgical nerve
implantation technique preserving outer nerve layers. J Neurosci Methods 189:205–209.
Hasegawa M, Seto A, Uchiyama N, Kida S, Yamashima T, Yamashita J (1996) Localization of E-
cadherin in peripheral glia after nerve injury and repair. J Neuropathol Exp Neurol 55:424–
434.
Hersel U, Dahmen C, Kessler H (2003) RGD modified polymers: biomaterials for stimulated cell
adhesion and beyond. Biomaterials 24:4385–4415.
Heumann R, Korsching S, Bandtlow C, Thoenen H (1987) Changes of nerve growth factor
synthesis in nonneuronal cells in response to sciatic nerve transection. J Cell Biol 104:1623–
1631.
Heydarkhan-Hagvall S, Schenke-Layland K, Dhanasopon AP, Rofail F, Smith H, Wu BM,
Shemin R, Beygui RE, MacLellan WR (2008) Three-dimensional electrospun ECM-based
hybrid scaffolds for cardiovascular tissue engineering. Biomaterials 29:2907–2914.
Hoffman-Kim D, Mitchel JA, Bellamkonda R V (2010) Topography, cell response, and nerve
regeneration. Annu Rev Biomed Eng 12:203–231.
Hoffmann N, Mittnacht U, Hartmann H, Baumer Y, Kjems J, Oberhoffner S, Schlosshauer B
(2011) Neuronal and glial responses to siRNA-coated nerve guide implants in vitro.
Neurosci Lett 494:14–18.
Holmes TC, de Lacalle S, Su X, Liu G, Rich A, Zhang S (2000) Extensive neurite outgrowth and
active synapse formation on self-assembling peptide scaffolds. Proc Natl Acad Sci U S A
97:6728–6733.
Hood B, Levene HB, Levi AD (2009) Transplantation of autologous Schwann cells for the repair
of segmental peripheral nerve defects. Neurosurg Focus 26:E4.
Hoyle GW, Mercer EH, Palmiter RD, Brinster RL (1993) Expression of NGF in sympathetic
neurons leads to excessive axon outgrowth from ganglia but decreased terminal innervation
within tissues. Neuron 10:1019–1034.

95
Hu N, Wu H, Xue C, Gong Y, Wu J, Xiao Z, Yang Y, Ding F, Gu X (2013) Long-term outcome
of the repair of 50 mm long median nerve defects in rhesus monkeys with marrow
mesenchymal stem cells-containing, chitosan-based tissue engineered nerve grafts.
Biomaterials 34:100–111.
Huang EJ, Reichardt LF (2001) Neurotrophins: roles in neuronal development and function. Annu
Rev Neurosci 24:677–736.
Huang J, Lu L, Hu X, Ye Z, Peng Y, Yan X, Geng D, Luo Z (2010) Electrical stimulation
accelerates motor functional recovery in the rat model of 15-mm sciatic nerve gap bridged
by scaffolds with longitudinally oriented microchannels. Neurorehabil Neural Repair
24:736–745.
Huang J, Xiang J, Yan Q, Li S, Song L, Cai X (2013) Dog tibial nerve regeneration across a 30-
mm defect bridged by a PRGD/PDLLA/beta-TCP/NGF sustained-release conduit. J
Reconstr Microsurg 29:77–87.
Hynes RO (1992) Integrins: versatility, modulation, and signaling in cell adhesion. Cell 69:11–25.
Ichihara S, Inada Y, Nakada A, Endo K, Azuma T, Nakai R, Tsutsumi S, Kurosawa H, Nakamura
T (2009) Development of new nerve guide tube for repair of long nerve defects. Tissue Eng
Part C Methods 15:387–402.
Inada Y, Hosoi H, Yamashita A, Morimoto S, Tatsumi H, Notazawa S, Kanemaru S, Nakamura T
(2007) Regeneration of peripheral motor nerve gaps with a polyglycolic acid-collagen tube:
technical case report. Neurosurgery 61:E1105–E1107; discussion E1107.
Inada Y, Morimoto S, Takakura Y, Nakamura T (2004) Regeneration of peripheral nerve gaps
with a polyglycolic acid-collagen tube. Neurosurgery 55:640–646; discussion 646–648.
Isaacs J (2010) Treatment of acute peripheral nerve injuries: current concepts. J Hand Surg Am
35:491–497.
Itoh S, Matsuda A, Kobayashi H, Ichinose S, Shinomiya K, Tanaka J (2005) Effects of a laminin
peptide (YIGSR) immobilized on crab-tendon chitosan tubes on nerve regeneration. J
Biomed Mater Res B Appl Biomater 73:375–382.
Jager SB, Ronchi G, Vaegter CB, Geuna S (2014) TheMouse Median Nerve Experimental Model
in Regenerative Research. Biomed Res Int 2014:13–17.
Jenq C-B, Coggeshall RE (1985) Numbers of regenerating axons in parent and tributary
peripheral nerves in the rat. Brain Res 326:27–40.
Jha BS, Colello RJ, Bowman JR, Sell S a, Lee KD, Bigbee JW, Bowlin GL, Chow WN, Mathern
BE, Simpson DG (2011) Two pole air gap electrospinning: Fabrication of highly aligned,
three-dimensional scaffolds for nerve reconstruction. Acta Biomater 7:203–215.
Johnson PJ, Newton P, Hunter DA, Mackinnon SE (2011) Nerve endoneurial microstructure
facilitates uniform distribution of regenerative fibers: a post hoc comparison of midgraft
nerve fiber densities. J Reconstr Microsurg 27:83–90.
Kanje M, Skottner A, Lundborg G, Sjöberg J (1991) Does insulin-like growth factor I (IGF-1)
trigger the cell body reaction in the rat sciatic nerve? Brain Res 563:285–287.

References
96
Kawaja MD, Boyd JG, Smithson LJ, Jahed A, Doucette R (2009) Technical strategies to isolate
olfactory ensheathing cells for intraspinal implantation. J Neurotrauma 26:155–177.
Keast JR, Forrest SL, Osborne PB (2010) Sciatic nerve injury in adult rats causes distinct changes
in the central projections of sensory neurons expressing different glial cell line-derived
neurotrophic factor family receptors. J Comp Neurol 518:3024–3045.
Kim DH, Connolly SE, Kline DG, Voorhies RM, Smith A, Powell M, Yoes T, Daniloff JK (1994)
Labeled Schwann cell transplants versus sural nerve grafts in nerve repair. J Neurosurg
80:254–260.
Kim D-Y, Choi Y-S, Kim S-E, Lee J-H, Kim S-M, Kim Y-J, Rhie J-W, Jun Y-J (2014) In vivo
effects of adipose-derived stem cells in inducing neuronal regeneration in sprague-dawley
rats undergoing nerve defect bridged with polycaprolactone nanotubes. J Korean Med Sci
29:183–192.
Kim Y-T, Haftel VK, Kumar S, Bellamkonda R V (2008) The role of aligned polymer fiber-based
constructs in the bridging of long peripheral nerve gaps. Biomaterials 29:3117–3127.
Klinkhammer K, Bockelmann J, Simitzis C, Brook G a, Grafahrend D, Groll J, Möller M, Mey J,
Klee D (2010) Functionalization of electrospun fibers of poly(epsilon-caprolactone) with
star shaped NCO-poly(ethylene glycol)-stat-poly(propylene glycol) for neuronal cell
guidance. J Mater Sci Mater Med 21:2637–2651.
Klinkhammer K, Seiler N, Grafahrend D, Gerardo-Nava J, Mey J, Brook G a, Möller M, Dalton
PD, Klee D (2009) Deposition of electrospun fibers on reactive substrates for in vitro
investigations. Tissue Eng Part C Methods 15:77–85.
Koh HS, Yong T, Chan CK, Ramakrishna S (2008) Enhancement of neurite outgrowth using
nano-structured scaffolds coupled with laminin. Biomaterials 29:3574–3582.
Koh HS, Yong T, Teo WE, Chan CK, Puhaindran ME, Tan TC, Lim A, Lim BH, Ramakrishna S
(2010) In vivo study of novel nanofibrous intra-luminal guidance channels to promote nerve
regeneration. J Neural Eng 7:14.
Kokai LE, Bourbeau D, Weber D, McAtee J, Marra KG (2011) Sustained Growth Factor Delivery
Promotes Axonal Regeneration in Long Gap Peripheral Nerve Repair. Tissue Eng Part A
17:1263–1275.
Kokai LE, Ghaznavi AM, Marra KG (2010) Incorporation of double-walled microspheres into
polymer nerve guides for the sustained delivery of glial cell line-derived neurotrophic factor.
Biomaterials 31:2313–2322.
Konofaos P, Ver Halen JP (2013) Nerve Repair by Means of Tubulization: Past, Present, Future. J
Reconstr Microsurg 29:149–163.
Koopmans G, Hasse B, Sinis N (2009) The role of collagen in peripheral nerve repair., 1st ed.
Elsevier Inc.
Korte N, Schenk HC, Grothe C, Tipold A, Haastert-Talini K (2011) Evaluation of periodic
electrodiagnostic measurements to monitor motor recovery after different peripheral nerve
lesions in the rat. Muscle Nerve 44:63–73.

97
Kriebel A, Rumman M, Scheld M, Hodde D, Brook G, Mey J (2014) Three-dimensional
configuration of orientated fibers as guidance structures for cell migration and axonal
growth. J Biomed Mater Res B Appl Biomater 102:356–365.
Labrador RO, Butí M, Navarro X (1998) Influence of collagen and laminin gels concentration on
nerve regeneration after resection and tube repair. Exp Neurol 149:243–252.
Lago N, Casas C, Muir EM, Rogers J, Navarro X (2009) Effects of Schwann cell transplants in an
experimental nerve amputee model. Restor Neurol Neurosci 27:67–78.
Latasa M-J, Ituero M, Moran-Gonzalez A, Aranda A, Cosgaya JM (2010) Retinoic acid regulates
myelin formation in the peripheral nervous system. Glia 58:1451–1464.
Lee AC (2003) Controlled release of nerve growth factor enhances sciatic nerve regeneration. Exp
Neurol 184:295–303.
Lee JY, Bashur CA, Goldstein AS, Schmidt CE (2009) Polypyrrole-coated electrospun PLGA
nanofibers for neural tissue applications. Biomaterials 30:4325–4335.
Lewis GM, Kucenas S (2014) Perineurial glia are essential for motor axon regrowth following
nerve injury. J Neurosci 34:12762–12777.
Lietz M, Dreesmann L, Hoss M, Oberhoffner S, Schlosshauer B (2006) Neuro tissue engineering
of glial nerve guides and the impact of different cell types. Biomaterials 27:1425–1436.
Liu J-J, Wang C-Y, Wang J-G, Ruan H-J, Fan C-Y (2011) Peripheral nerve regeneration using
composite poly(lactic acid-caprolactone)/nerve growth factor conduits prepared by coaxial
electrospinning. J Biomed Mater Res A 96:13–20.
Lohmeyer JA, Shen Z-L, Walter GF, Berger A (2007) Bridging extended nerve defects with an
artifcial nerve graft containing Schwann cells pre-seeded on polyglactin filaments. Int J Artif
Organs 30:64–74.
Lundborg G (2000) A 25-year perspective of peripheral nerve surgery: evolving neuroscientific
concepts and clinical significance. J Hand Surg Am 25:391–414.
Lundborg G, Dahlin L, Dohi D, Kanje M, Terada N (1997a) A new type of “bioartificial” nerve
graft for bridging extended defects in nerves. J Hand Surg Br 22:299–303.
Lundborg G, Dahlin LB, Danielsen N, Gelberman RH, Longo FM, Powell HC, Varon S (1982)
Nerve regeneration in silicone chambers: influence of gap length and of distal stump
components. Exp Neurol 76:361–375.
Lundborg G, Rosén B, Dahlin L, Danielsen N, Holmberg J (1997b) Tubular versus conventional
repair of median and ulnar nerves in the human forearm: early results from a prospective,
randomized, clinical study. J Hand Surg Am 22:99–106.
Lundborg G, Rosén B, Dahlin L, Holmberg J, Rosén I (2004) Tubular repair of the median or
ulnar nerve in the human forearm: a 5-year follow-up. J Hand Surg Br 29:100–107.
Madduri S, Feldman K, Tervoort T, Papaloïzos M, Gander B (2010a) Collagen nerve conduits
releasing the neurotrophic factors GDNF and NGF. J Control Release 143:168–174.

References
98
Madduri S, Papaloïzos M, Gander B (2010b) Trophically and topographically functionalized silk
fibroin nerve conduits for guided peripheral nerve regeneration. Biomaterials 31:2323–2334.
Makwana M, Raivich G (2005) Molecular mechanisms in successful peripheral regeneration.
FEBS J 272:2628–2638.
Martini R (1994) Expression and functional roles of neural cell surface molecules and
extracellular matrix components during development and regeneration of peripheral nerves.
J Neurocytol 23:1–28.
Martini R, Schachner M (1988) Immunoelectron microscopic localization of neural cell adhesion
molecules (L1, N-CAM, and myelin-associated glycoprotein) in regenerating adult mouse
sciatic nerve. J Cell Biol 106:1735–1746.
Matsumoto K, Ohnishi K, Kiyotani T, Sekine T, Ueda H, Nakamura T, Endo K, Shimizu Y
(2000) Peripheral nerve regeneration across an 80-mm gap bridged by a polyglycolic acid
(PGA)-collagen tube filled with laminin-coated collagen fibers: a histological and
electrophysiological evaluation of regenerated nerves. Brain Res 868:315–328.
Matthews JA, Wnek GE, Simpson DG, Bowlin GL (2002) Electrospinning of collagen
nanofibers. Biomacromolecules 3:232–238.
May F, Weidner N, Matiasek K, Caspers C, Mrva T, Vroemen M, Henke J, Lehmer A,
Schwaibold H, Erhardt W, Gänsbacher B, Hartung R (2004) Schwann cell seeded guidance
tubes restore erectile function after ablation of cavernous nerves in rats. J Urol 172:374–377.
McCallister W V, Cober SR, Norman A, Trumble TE (2001) Using intact nerve to bridge
peripheral nerve defects: an alternative to the use of nerve grafts. J Hand Surg Am 26:315–
325.
McDonald DS, Zochodne DW (2003) An injectable nerve regeneration chamber for studies of
unstable soluble growth factors. J Neurosci Methods 122:171–178.
McGrath AM, Novikova LN, Novikov LN, Wiberg M (2010) BDTM
PuraMatrixTM
peptide
hydrogel seeded with Schwann cells for peripheral nerve regeneration. Brain Res Bull
83:207–213.
Meek MF, Coert JH (2008a) US Food and Drug Administration/Conformit Europe-Approved
Absorbable Nerve Conduits for Clinical Repair of Peripheral and Cranial nerves. Ann Plast
Surg.
Meek MF, Coert JH (2008b) US Food and Drug Administration /Conformit Europe- approved
absorbable nerve conduits for clinical repair of peripheral and cranial nerves. Ann Plast Surg
60:466–472.
Meek MF, Coert JH (2012) Nerve Conduits for Nerve Repair or Reconstruction. J Am Acad
Orthop Surg 20:63–68.
Meek MF, Coert JH (2014) Peripheral Nerve Reconstruction after Injury : A Review of Clinical
and Experimental Therapies. Biomed Res Int 2014:13.

99
Meek MF, Jansen K (2009) Two years after in vivo implantation of poly(DL-lactide-epsilon-
caprolactone) nerve guides: has the material finally resorbed? J Biomed Mater Res A
89:734–738.
Meek MF, Nicolai J-PA, Robinson PH (2006) Secondary digital nerve repair in the foot with
resorbable p(DLLA-epsilon-CL) nerve conduits. J Reconstr Microsurg 22:149–151.
Meek MF, Varejão ASP, Geuna S (2004) Use of skeletal muscle tissue in peripheral nerve repair:
review of the literature. Tissue Eng 10:1027–1036.
Mey J, Brook G, Hodde D, Kriebel A (2012) Electrospun Fibers as Substrates for Peripheral
Nerve Regeneration. Adv Polym Sci 246:131–170.
Miyamoto Y, Sugita T, Higaki T, Ikuta Y, Tsuge K (1985) The duration of denervation and
regeneration in nerve grafting Quantitative histological assessment in the rat. Int Orthop
9:271–276.
Mohanna P-N, Terenghi G, Wiberg M (2005) Composite PHB-GGF conduit for long nerve gap
repair: a long-term evaluation. Scand J Plast Reconstr Surg Hand Surg 39:129–137.
Mohanna PN, Young RC, Wiberg M, Terenghi G (2003) A composite poly-hydroxybutyrate-glial
growth factor conduit for long nerve gap repairs. J Anat 203:553–565.
Möllers S, Heschel I, Damink LHHO, Schügner F, Deumens R, Müller B, Bozkurt A, Nava JG,
Noth J, Brook GA (2009) Cytocompatibility of a novel, longitudinally microstructured
collagen scaffold intended for nerve tissue repair. Tissue Eng Part A 15:461–472.
Moore AM, Kasukurthi R, Magill CK, Farhadi HF, Borschel GH, Mackinnon SE (2009)
Limitations of conduits in peripheral nerve repairs. Hand (N Y) 4:180–186.
Moore K, MacSween M, Shoichet M (2006) Immobilized concentration gradients of neurotrophic
factors guide neurite outgrowth of primary neurons in macroporous scaffolds. Tissue Eng
12:267–278.
Murakami M, Simons M (2008) Fibroblast growth factor regulation of neovascularization. Curr
Opin Hematol 15:215–220.
Nakamura T, Inada Y, Fukuda S, Yoshitani M, Nakada A, Itoi S, Kanemaru S, Endo K, Shimizu
Y (2004) Experimental study on the regeneration of peripheral nerve gaps through a
polyglycolic acid-collagen (PGA-collagen) tube. Brain Res 1027:18–29.
Nave K-A, Salzer JL (2006) Axonal regulation of myelination by neuregulin 1. Curr Opin
Neurobiol 16:492–500.
Navissano M, Malan F, Carnino R, Battiston B (2005) Neurotube for facial nerve repair.
Microsurgery 25:268–271.
Nijhuis THJ, Bodar CWJ, van Neck JW, Walbeehm ET, Siemionow M, Madajka M, Cwykiel J,
Blok JH, Hovius SER (2013) Natural conduits for bridging a 15-mm nerve defect:
comparison of the vein supported by muscle and bone marrow stromal cells with a nerve
autograft. J Plast Reconstr Aesthet Surg 66:251–259.

References
100
Noble J, Munro CA, Prasad VS, Midha R (1998) Analysis of upper and lower extremity
peripheral nerve injuries in a population of patients with multiple injuries. J Trauma 45:116–
122.
Novikova LN, Pettersson J, Brohlin M, Wiberg M, Novikov LN (2008) Biodegradable poly-beta-
hydroxybutyrate scaffold seeded with Schwann cells to promote spinal cord repair.
Biomaterials 29:1198–1206.
Ohta M, Suzuki Y, Chou H, Ishikawa N, Suzuki S, Tanihara M, Suzuki Y, Mizushima Y, Dezawa
M, Ide C (2004) Novel heparin/alginate gel combined with basic fibroblast growth factor
promotes nerve regeneration in rat sciatic nerve. J Biomed Mater Res A 71:661–668.
Oliveira JT, Almeida FM, Biancalana A, Baptista AF, Tomaz MA, Melo PA, Martinez AMB
(2010) Mesenchymal stem cells in a polycaprolactone conduit enhance median-nerve
regeneration, prevent decrease of creatine phosphokinase levels in muscle, and improve
functional recovery in mice. Neuroscience 170:1295–1303.
Ozgenel GY (2003) Effects of hyaluronic acid on peripheral nerve scarring and regeneration in
rats. Microsurgery 23:575–581.
Pabari A, Yang SY, Seifalian AM, Mosahebi A (2010) Modern surgical management of
peripheral nerve gap. J Plast Reconstr Aesthet Surg 63:1941–1948.
Pan H-C, Wu H-T, Cheng F-C, Chen C-H, Sheu M-L, Chen C-J (2009) Potentiation of
angiogenesis and regeneration by G-CSF after sciatic nerve crush injury. Biochem Biophys
Res Commun 382:177–182.
Panaite P-A, Barakat-Walter I (2010) Thyroid hormone enhances transected axonal regeneration
and muscle reinnervation following rat sciatic nerve injury. J Neurosci Res 88:1751–1763.
Pankov R, Yamada KM (2002) Fibronectin at a glance. J Cell Sci 115:3861–3863.
Panseri S, Cunha C, Lowery J, Del Carro U, Taraballi F, Amadio S, Vescovi A, Gelain F (2008)
Electrospun micro- and nanofiber tubes for functional nervous regeneration in sciatic nerve
transections. BMC Biotechnol 8:39.
Pitta MC, Wolford LM, Mehra P, Hopkin J (2001) Use of Gore-Tex tubing as a conduit for
inferior alveolar and lingual nerve repair: experience with 6 cases. J Oral Maxillofac Surg
59:493–496; discussion 497.
Pola R, Aprahamian TR, Bosch-Marcé M, Curry C, Gaetani E, Flex A, Smith RC, Isner JM,
Losordo DW (2004) Age-dependent VEGF expression and intraneural neovascularization
during regeneration of peripheral nerves. Neurobiol Aging 25:1361–1368.
Radtke C, Akiyama Y, Lankford KL, Vogt PM, Krause DS, Kocsis JD (2005) Integration of
engrafted Schwann cells into injured peripheral nerve: axonal association and nodal
formation on regenerated axons. Neurosci Lett 387:85–89.
Radtke C, Wewetzer K, Reimers K, Vogt PM (2011) Transplantation of olfactory ensheathing
cells as adjunct cell therapy for peripheral nerve injury. Cell Transplant 20:145–152.
Rafiuddin Ahmed M, Jayakumar R (2003) Peripheral nerve regeneration in RGD peptide
incorporated collagen tubes. Brain Res 993:208–216.

101
Ribeiro-Resende VT, Koenig B, Nichterwitz S, Oberhoffner S, Schlosshauer B (2009) Strategies
for inducing the formation of bands of Büngner in peripheral nerve regeneration.
Biomaterials 30:5251–5259.
Robinson LR (2004) traumatic injury to peripheral nerves. Suppl Clin Neurophysiol 57:173–186.
Rosberg H-E, Carlsson KS, Dahlin LB (2005) Prospective study of patients with injuries to the
hand and forearm: costs, function, and general health. Scand J Plast Reconstr Surg Hand
Surg 39:360–369.
Scaravilli F (1984) Regeneration of the perineurium across a surgically induced gap in a nerve
encased in a plastic tube. J Anat 139:411–424.
Schmidt CE, Leach JB (2003) Neural tissue engineering: strategies for repair and regeneration.
Annu Rev Biomed Eng 5:293–347.
Schmidt CE, Shastri VR, Vacanti JP, Langer R (1997) Stimulation of neurite outgrowth using an
electrically conducting polymer. Proc Natl Acad Sci U S A 94:8948–8953.
Schnell E, Klinkhammer K, Balzer S, Brook G, Klee D, Dalton P, Mey J (2007) Guidance of glial
cell migration and axonal growth on electrospun nanofibers of poly-epsilon-caprolactone
and a collagen/poly-epsilon-caprolactone blend. Biomaterials 28:3012–3025.
Scholz T, Krichevsky A, Sumarto A, Jaffurs D, Wirth GA, Paydar K, Evans GRD (2009)
Peripheral nerve injuries: an international survey of current treatments and future
perspectives. J Reconstr Microsurg 25:339–344.
Seddon HJ (1943) Three types of nerve injury. Brain 66:237–288.
Sendtner M, Kreutzberg GW, Thoenen H (1990) Ciliary neurotrophic factor prevents the
degeneration of motor neurons after axotomy. Nature 345:440–441.
Sendtner M, Stöckli KA, Thoenen H (1992) Synthesis and localization of ciliary neurotrophic
factor in the sciatic nerve of the adult rat after lesion and during regeneration. J Cell Biol
118:139–148.
Shadiack AM, Sun Y, Zigmond RE (2001) Nerve growth factor antiserum induces axotomy-like
changes in neuropeptide expression in intact sympathetic and sensory neurons. J Neurosci
21:363–371.
Shen H, Shen Z-L, Zhang P-H, Chen N-L, Wang Y-C, Zhang Z-F, Jin Y-Q (2010) Ciliary
neurotrophic factor-coated polylactic-polyglycolic acid chitosan nerve conduit promotes
peripheral nerve regeneration in canine tibial nerve defect repair. J Biomed Mater Res B
Appl Biomater 95:161–170.
Shin JE, Cho Y, Beirowski B, Milbrandt J, Cavalli V, DiAntonio A (2012) Dual leucine zipper
kinase is required for retrograde injury signaling and axonal regeneration. Neuron 74:1015–
1022.
Siemionow M, Bozkurt M, Zor F (2010) Regeneration and repair of peripheral nerves with
different biomaterials: Review. Microsurgery 30:574–588.

References
102
Silva GA, Czeisler C, Niece KL, Beniash E, Harrington DA, Kessler JA, Stupp SI (2004)
Selective differentiation of neural progenitor cells by high-epitope density nanofibers.
Science 303:1352–1355.
Sinis N, Schaller H-E, Becker ST, Schlosshauer B, Doser M, Roesner H, Oberhoffner S, Müller
H-W, Haerle M (2007) Long nerve gaps limit the regenerative potential of bioartificial nerve
conduits filled with Schwann cells. Restor Neurol Neurosci 25:131–141.
Smeal RM, Rabbitt R, Biran R, Tresco PA (2005) Substrate curvature influences the direction of
nerve outgrowth. Ann Biomed Eng 33:376–382.
Soares S, Traka M, von Boxberg Y, Bouquet C, Karagogeos D, Nothias F (2005) Neuronal and
glial expression of the adhesion molecule TAG-1 is regulated after peripheral nerve lesion or
central neurodegeneration of adult nervous system. Eur J Neurosci 21:1169–1180.
Solorio L, Zwolinski C (2010) Gelatin microspheres crosslinked with genipin for local delivery of
growth factors. J Tissue Eng Regen Med 4:514–523.
Sørensen J, Haase G, Krarup C, Gilgenkrantz H, Kahn A, Schmalbruch H (1998) Gene transfer to
Schwann cells after peripheral nerve injury: a delivery system for therapeutic agents. Ann
Neurol 43:205–211.
Soukup T, Zacharová G, Smerdu V (2002) Fibre type composition of soleus and extensor
digitorum longus muscles in normal female inbred Lewis rats. Acta Histochem 104:399–
405.
Stanec S, Stanec Z (1998) Ulnar nerve reconstruction with an expanded polytetrafluoroethylene
conduit. Br J Plast Surg 51:637–639.
Sulaiman O, Boyd J, Gordon T (2005) Axonal regeneration in the peripheral nervous system of
mammals. In: Neuroglia (Kettenmann H, Ransom BH, eds), pp 454–466. Oxford: Oxford
University Press.
Sunderland S (1951) A classification of peripheral nerve injuries producing loss of function. Brain
74:491–516.
Suri S, Schmidt CE (2010) Cell-laden hydrogel constructs of hyaluronic acid, collagen, and
laminin for neural tissue engineering. Tissue Eng Part A 16:1703–1716.
Suzuki K, Suzuki Y, Ohnishi K, Endo K, Tanihara M, Nishimura Y (1999) Regeneration of
transected spinal cord in young adult rats using freeze-dried alginate gel. Neuroreport
10:2891–2894.
Suzuki K, Suzuki Y, Tanihara M, Ohnishi K, Hashimoto T, Endo K, Nishimura Y (2000)
Reconstruction of rat peripheral nerve gap without sutures using freeze-dried alginate gel. J
Biomed Mater Res 49:528–533.
Taha MO, Rosseto M, Fraga MM, Mueller SF, Fagundes DJ, Novo NF, Caricati-Neto A (2004)
Effect of retinoic acid on tibial nerve regeneration after anastomosis in rats: histological and
functional analyses. Transplant Proc 36:404–408.
Taras JS, Jacoby SM (2008) Repair of lacerated peripheral nerves with nerve conduits. Tech Hand
Up Extrem Surg 12:100–106.

103
Taylor CA, Braza D, Rice JB, Dillingham T (2008) The incidence of peripheral nerve injury in
extremity trauma. Am J Phys Med Rehabil 87:381–385.
Teo WE, Ramakrishna S (2006) A review on electrospinning design and nanofibre assemblies.
Nanotechnology 17:R89–R106.
Thornton MR, Mantovani C, Birchall MA, Terenghi G (2005) Quantification of N-CAM and N-
cadherin expression in axotomized and crushed rat sciatic nerve. J Anat 206:69–78.
Toba T, Nakamura T, Shimizu Y, Matsumoto K, Ohnishi K, Fukuda S, Yoshitani M, Ueda H,
Hori Y, Endo K (2001) Regeneration of canine peroneal nerve with the use of a polyglycolic
acid-collagen tube filled with laminin-soaked collagen sponge: a comparative study of
collagen sponge and collagen fibers as filling materials for nerve conduits. J Biomed Mater
Res 58:622–630.
Trupp M, Rydén M, Jörnvall H, Funakoshi H, Timmusk T, Arenas E, Ibáñez CF (1995)
Peripheral expression and biological activities of GDNF, a new neurotrophic factor for avian
and mammalian peripheral neurons. J Cell Biol 130:137–148.
Tsai EC, Dalton PD, Shoichet MS, Tator CH (2004) Synthetic hydrogel guidance channels
facilitate regeneration of adult rat brainstem motor axons after complete spinal cord
transection. J Neurotrauma 21:789–804.
UVG www.unfallstatistik.ch/d/neuza/med_stat/diag_VerlNerven_d.htm.
Valmikinathan CM, Defroda S, Yu X (2009) Polycaprolactone and bovine serum albumin based
nanofibers for controlled release of nerve growth factor. Biomacromolecules 10:1084–1089.
Van Neerven SGA, Bozkurt A, O’Dey DM, Scheffel J, Boecker AH, Stromps J-P, Dunda S,
Brook GA, Pallua N (2012) Retrograde tracing and toe spreading after experimental
autologous nerve transplantation and crush injury of the sciatic nerve: a descriptive
methodological study. J Brachial Plex Peripher Nerve Inj 7:5.
Verdú E, Labrador RO, Rodríguez FJ, Ceballos D, Forés J, Navarro X (2002) Alignment of
collagen and laminin-containing gels improve nerve regeneration within silicone tubes.
Restor Neurol Neurosci 20:169–179.
Wang S, Wan AC, Xu X, Gao S, Mao HQ, Leong KW, Yu H (2001) A new nerve guide conduit
material composed of a biodegradable poly(phosphoester). Biomaterials 22:1157–1169.
Wang S, Yaszemski MJ, Knight AM, Gruetzmacher JA, Windebank AJ, Lu L (2009) Photo-
crosslinked poly(epsilon-caprolactone fumarate) networks for guided peripheral nerve
regeneration: material properties and preliminary biological evaluations. Acta Biomater
5:1531–1542.
Wang W, Itoh S, Matsuda A, Aizawa T, Demura M, Ichinose S, Shinomiya K, Tanaka J (2008)
Enhanced nerve regeneration through a bilayered chitosan tube: the effect of introduction of
glycine spacer into the CYIGSR sequence. J Biomed Mater Res A 85:919–928.
Wang X, Gu X, Yuan C, Chen S, Zhang P, Zhang T, Yao J, Chen F, Chen G (2004) Evaluation of
biocompatibility of polypyrrole in vitro and in vivo. J Biomed Mater Res A 68:411–422.

References
104
Wang X, Hu W, Cao Y, Yao J, Wu J, Gu X (2005) Dog sciatic nerve regeneration across a 30-
mm defect bridged by a chitosan/PGA artificial nerve graft. Brain a J Neurol 128:1897–
1910.
Wangensteen KJ, Kalliainen LK (2010) Collagen tube conduits in peripheral nerve repair: a
retrospective analysis. Hand (N Y) 5:273–277.
Webb K, Budko E, Neuberger TJ, Chen S, Schachner M, Tresco PA (2001) Substrate-bound
human recombinant L1 selectively promotes neuronal attachment and outgrowth in the
presence of astrocytes and fibroblasts. Biomaterials 22:1017–1028.
Weber R a, Breidenbach WC, Brown RE, Jabaley ME, Mass DP (2000) A randomized
prospective study of polyglycolic acid conduits for digital nerve reconstruction in humans.
Plast Reconstr Surg 106:1036–1045; discussion 1046–1048.
Weis J, May R, Schroder JM (1994) Fine structural and immunohistochemical identification of
perineurial cells connecting proximal and distal stumps of transected peripheral nerves at
early stages of regeneration in silicone tubes. Acta Neuropathol 88:159–165.
Whitlock EL, Tuffaha SH, Luciano JP, Yan Y, Hunter DA, Magill CK, Moore AM, Tong AY,
Mackinnon SE, Borschel GH (2009) Processed allografts and type I collagen conduits for
repair of peripheral nerve gaps. Muscle Nerve 39:787–799.
Witzel C, Rohde C, Brushart TM (2005) Pathway sampling by regenerating peripheral axons. J
Comp Neurol 485:183–190.
Xu C, Kou Y, Zhang P, Han N, Yin X, Deng J, Chen B, Jiang B (2014) Electrical stimulation
promotes regeneration of defective peripheral nerves after delayed repair intervals lasting
under one month. PLoS One 9:e105045.
Xu X, Yee W-C, Hwang PYK, Yu H, Wan ACA, Gao S, Boon K-L, Mao H-Q, Leong KW, Wang
S (2003) Peripheral nerve regeneration with sustained release of poly(phosphoester)
microencapsulated nerve growth factor within nerve guide conduits. Biomaterials 24:2405–
2412.
Yamakuchi M, Higuchi I, Masuda S, Ohira Y, Kubo T, Kato Y, Maruyama I, Kitajima I (2000)
Type I muscle atrophy caused by microgravity-induced decrease of myocyte enhancer factor
2C (MEF2C) protein expression. FEBS Lett 477:135–140.
Yang F, Murugan R, Wang S, Ramakrishna S (2005) Electrospinning of nano/micro scale poly(L-
lactic acid) aligned fibers and their potential in neural tissue engineering. Biomaterials
26:2603–2610.
Yang Y, Ding F, Wu J, Hu W, Liu W, Liu J, Gu X (2007) Development and evaluation of silk
fibroin-based nerve grafts used for peripheral nerve regeneration. Biomaterials 28:5526–
5535.
Yang Y, Wimpenny I, Ahearne M (2011) Portable nanofiber meshes dictate cell orientation
throughout three-dimensional hydrogels. Nanomedicine 7:131–136.
Yao L, de Ruiter GCW, Wang H, Knight AM, Spinner RJ, Yaszemski MJ, Windebank AJ, Pandit
A (2010) Controlling dispersion of axonal regeneration using a multichannel collagen nerve
conduit. Biomaterials 31:5789–5797.

105
You H, Wei L, Liu Y, Oudega M, Jiao S-S, Feng S-N, Chen Y, Chen J-M, Li B-C (2011)
Olfactory ensheathing cells enhance Schwann cell-mediated anatomical and functional
repair after sciatic nerve injury in adult rats. Exp Neurol 229:158–167.
Yu TT, Shoichet MS (2005) Guided cell adhesion and outgrowth in peptide-modified channels for
neural tissue engineering. Biomaterials 26:1507–1514.
Yu W, Zhao W, Zhu C, Zhang X, Ye D, Zhang W, Zhou Y, Jiang X, Zhang Z (2011) Sciatic
nerve regeneration in rats by a promising electrospun collagen/poly(ε-caprolactone) nerve
conduit with tailored degradation rate. BMC Neurosci 12:68.
Yucel D, Kose GT, Hasirci V (2010) Polyester based nerve guidance conduit design. Biomaterials
31:1596–1603.
Zhang S, Holmes T, Lockshin C, Rich A (1993) Spontaneous assembly of a self-complementary
oligopeptide to form a stable macroscopic membrane. Proc Natl Acad Sci U S A 90:3334–
3338.
Zhang YZ, Venugopal J, Huang Z-M, Lim CT, Ramakrishna S (2005) Characterization of the
surface biocompatibility of the electrospun PCL-collagen nanofibers using fibroblasts.
Biomacromolecules 6:2583–2589.
Zhelyaznik N, Mey J (2006) Regulation of retinoic acid receptors alpha, beta and retinoid X
receptor alpha after sciatic nerve injury. Neuroscience 141:1761–1774.
Zhelyaznik N, Schrage K, McCaffery P, Mey J (2003) Activation of retinoic acid signalling after
sciatic nerve injury: up-regulation of cellular retinoid binding proteins. Eur J Neurosci
18:1033–1040.
Zochodne DW (2012) The challenges and beauty of peripheral nerve regrowth. J Peripher Nerv
Syst 17:1–18.


107
Abbreviations
BDNF Brain-derived neurotrophic factor
CAM Cell adhesion molecule
CNS Central nervous system
CNTF Ciliary neurotrophic factor
DRG Dorsal root ganglion
ECM Extracellular matrix
ePTFE Extended poly(tetrafluoroethylene)
ErbB Erythroblastic leukemia viral oncogene (epidermal growth factor receptor
family)
FGF Fibroblast growth factor
GDNF Glial-derived neurotrophic factor
GFR Receptor of GDNF-family ligands
GGF Glial growth factor
IGF Insulin-like growth factor
IL Interleukin
LIF Leukemia inhibitory factor
NGF Nerve growth factor
NT Neurotrophin
PAN-MA Poly(acrylonitrile-co-methacrylate)
PCL Poly(ε-caprolactone)
PCLEEP Poly(caprolactone-co-ethyl ethylene phosphate)
PGA Poly(glycolic acid)
PHB Poly(hydroxybutyric acid)
pHEMA Poly(2-hydroxyethyl methacrylate)
pHEMA-MMA Poly(2-hydroxyethyl methacrylate-co-methyl methacrylate)
PLA Poly(lactic acid); the D- and L-isomers are often referred to as PDLA,
PLLA, respectively
PLGA Poly(lactic-co-glycolic acid)
PN Peripheral nerve
PNI Peripheral nerve injury
PNS Peripheral nervous system
PPE Polyphosphoester
PPy Poly(pyrrole)
sPEG Star-shaped poly(ethylene glycol)
STAT Signal transducer and activator of transcription
TGF Transforming growth factor
trk Tyrosine kinase/tropomyosin receptor kinase (neurotrophin receptor)
VEGF Vascular endothelial growth factor


109
Figures and Tables
Figure 1 Peripheral nerve degeneration and regeneration. ..................................................... 11
Figure 2 Design strategies for artificial nerve bridges. ............................................................. 18
Figure 3 Examples of artificial nerve guides that were successfully tested in animal
experiments. .................................................................................................................................. 20
Figure 4 Electrospinning of aligned nanofibers. ....................................................................... 22
Figure 5 Evaluation of artificial nerve implants in animal experiments. ................................ 33
Figure 6 Functionalization of polymer fibers for nerve regeneration. .................................... 36
Figure 7 Conduit design for artificial nerve guides. .................................................................. 42
Figure 8 Electrospinning setup for the collection of 3D fiber arrays....................................... 43
Figure 9 Three-dimensional arrays of parallel fibers. .............................................................. 44
Quantitative characterization of the 3D fiber array. .................................................................. 48
Figure 11 Embedding of 3D fiber stacks in collagen. ................................................................ 49
Figure 12 Gel-embedded PCL fibers affect cell morphology. ................................................... 50
Figure 13 Gel-embedded PCL fibers as a guidance structure for Schwann cells and sensory
neurons. ......................................................................................................................................... 51
Figure 14 Quantification of the guidance properties of gel-embedded PCL fibres. ............... 52
Figure 15 Integration of the composit scaffold into a biocompatible conduit. ....................... 54
Figure 16 Brief documentation of the fiber production and embedding in gelatin. .............. 61
Figure 17 Tissue regeneration following sciatic nerve transection (15 mm gap size) and
subsequent repair using different nerve bridges. ...................................................................... 65
Figure 18 Cross sectional area of the regenerated tissue in the medial segment 12W after
nerve transection and repair. ...................................................................................................... 66
Figure 19 Schwann cell infiltration and axonal regeneration at the proximal interface
between nerve and implant 2W post surgery. ........................................................................... 67
Figure 20 Schwann cell infiltration and axonal regeneration in the medial segment of the
regenerated nerve 12W post surgery. ........................................................................................ 68
Figure 21 Quantitative analysis of the regenerated nerve tissue. ........................................... 69
Figure 22 Similar percentage of regenerating motoraxons in all implants. ........................... 70
Figure 23 Electromyography performed 12W after sciatic nerve repair. .............................. 73
Figure 24 Behavioral analysis of nerve regeneration. .............................................................. 76
Figure 25 Determination of muscular atrophy. ........................................................................ 77
Table 1 Compound muscle action potentials after PN repair with artificial implants ........... 74
Table 2 Muscle weight normalized to the corresponding muscle on the contralateral side 2
and 12 weeks after respective treatment. .................................................................................. 78


111
Prior Publications and Contributions
The experiments reported in this thesis represent a joint effort of myself and several scientists,
who contributed to the investigation, supervised me or were supervised by me. I performed the
majority of all experimental work in the laboratory and I had a key role in designing, evaluating
and writing the studies that have already been submitted for publication. This is documented by
the respective lists of co-authors (see below).
Animal surgery was carried out by Gary Brook (with my assistance), programming and design of
the electrophysiology was the work of Thomas Kuenzel.
Electrospun Fibers as Substrates for Peripheral Nerve Regeneration
Jörg Mey, Gary Brook, Dorothee Hodde and Andreas Kriebel
Published in Advanced Polymer Science (2012) 246: 131–170
Three-dimensional configuration of orientated fibers as guidance structures for cell
migration and axonal growth
Andreas Kriebel, Muhammad Rumman, Miriam Scheld, Dorothee Hodde, Gary Brook and Jörg
Mey
Published in Journal of Biomedical Materials Research: Part B (2014) 102B: 356-365
Cell-free artificial nerve implants with electrospun fibers in a three-dimensional gelatin
matrix support nerve regeneration in vivo
Andreas Kriebel, Dorothee Hodde, Thomas Kuenzel, Jessica Engels, Gary Brook and Jörg Mey
Submitted for publication (March 2015)


113
Acknowledgments
Zuallererst möchte ich mich bei Jörg bedanken, dass er mir das Vertrauen geschenkt hat, dieses
Projekt zu bearbeiten. Ohne immer einer Meinung zu sein, sind wir dennoch gut miteinander
ausgekommen. Es hat Spaß gemacht mit Dir zu arbeiten.
Bedanken möchte ich mich auch bei Hermann, der mir uneigennützig Labor und Arbeitsplatz zur
Verfügung gestellt hat und auch sonst stets ein offenes Ohr für mich hatte.
Two persons that contributed essentially to this project are Gary and Doro. I dont know, whether I
could have spend all this time working with the animals without having Doro to talk to. I could
always rely on her and she helped whenever she could. The same accounts to Gary. I had a great
time assisting him with the operations. I have rarely seen a PI contributing so much to the
practical parts of a project. Besides that, he kept up my motivation, when things did not work out
as supposed to. That really helped a lot. You were a great team to work with, I hope I was able to
return something to you.
Thomas und Marcus danke ich für die vielen Ratschläge, die ich mir bei Ihnen geholt habe. Vor
allem Thomas tatkräftige Unterstützung hat nennenswert zum erfolgreichen Abschluss dieser
Arbeit beigetragen.
Darüber hinaus möchte ich auch den Bachelorstudenten danken, die durch ihre Abschlussarbeiten
Anteil an diesem Projekt hatten.
Natürlich darf auch der Rest des Instituts nicht vergessen werden. Ich habe mich in den nunmehr
fast sechs Jahren, die ich hier tätig war immer sehr wohl gefühlt. Besonders bei Michael muss ich
mich bedanken. Er hat zwar nie Zeit, aber nimmt sie sich doch immer, wenn man seine Hilfe
braucht, und die brauchte ich öfters. Roland muss ich auch danken, denn ohne seine Kompetenz
im Umgang mit Word wären vermutlich einige meiner PC-Komponenten aus dem Fenster
geflogen. Philipp, Lutz und Julius muss ich nicht danken…die wissen schon, dass sie für mich
mehr als bloß Kollegen sind und ich hoffe, dass es auch noch lange so bleibt.
Bleibt noch der Dank an meine Familie.
Katharina, Maya und Adrian…ihr seid der Antrieb und gleichzeitig die willkommenste
Ablenkung in meinem Leben. Meinem Vater danke ich dafür, dass er mir immer meine
Möglichkeiten aufgezeigt hat, ohne meine Entscheidungen treffen zu wollen.
Zu guter Letzt bedanke ich mich bei meiner Mutter. Sie hat mich zu dem Menschen gemacht hat,
der ich heute bin. Ich habe nun wohl fast alles erreicht, das sie sich für mich gewünscht hat. Ich
wünschte, sie hätte es erleben können.
Dieses Projekt wurde von der Deuschen Forschungsgemeinschaft finanziert (grant ME 1261/11-1)


115
Curriculum Vitae
Persönliche Daten
Name Andreas Kriebel
Geburtstag 11.12.1981
Geburtsort Leverkusen
Staatsangehörigkeit Deutsch
Werdegang
2001 Erlangung der allgemeinen Hochschulreife an der Landrat-
Lucas-Schule in Leverkusen-Opladen
2001-2002 Grundwehrdienst im Fallschirmjägerbataillon 263 in
Zweibrücken
2002-2003 Studium des Maschienenbaus, RWTH Aachen University
2003-2010 Studium der Biologie, RWTH Aachen University
Abschluss: Diplom-Biologe
2010-2014 Wissenschaftlicher Angestellter
Institut für Biologie II, RWTH Aachen University
2010-2015 Promotionsstudium, RWTH Aachen University




















