
Inflation is under control and does not exert any impact on business ...
Upload
independentCategory
view
2download
0
Journal of Leukocyte Biology 49:126-138 (1991)
© 1991 Wiley-Liss, Inc.
Splenic Macrophages From Tumor-Bearing MiceCo-Expressing MAC-I and MAC-2 Antigens Exert
Immunoregulatory Functions Via TwoDistinct Mechanisms
Gordon A. Watson, Yang-Xin Fu, and Diana M. LopezDepartment of Microbiology and Immunology, University of Miami School of Medicine (G.A.W., Y.-X.F., D.M.L.), and Sylvester
Comprehensive Cancer Center (D.M.L.), Miami, Florida
Tumor burden has been shown to induce a variety of phenotypic and functional changesin the cellular constituents of the host’s immune system. These changes have beenimplicated as mechanisms by which tumors avoid rejection. Studies of BALB/c micebearing a D1-DMBA-3 mammary adenocarcinoma showed alterations of the splenocytepopulations. There was a five-fold increase of macrophages (M4)that were phenotypicallyand functionally analyzed to establish their role in tumor-induced modifications of thehost’s immune response. Monoclonal antibody staining defined a Mac-i � populationwhich comprised up to 20% of the splenocytes in tumor-bearers (TB), but is negligible inspleens from normal mice. These Mac-i � M4 were found to mediate down-regulationof both polyclonal and antigen-specific T and B cell responses in vitro and in vivo.Although B cell responses were suppressed via prostaglandin E2 (PGE2) production bythe TB M4, T cell responses were relatively refractory to PGE2-mediated down-regulation.Instead, they were suppressed by a contact-dependent T cell-M4 interaction. Further-more, tumor-derived factors such as granulocyte-M4 colony-stimulating factor (GM-CSF)seem to play an Important role in the induction and expansion of the Mac-i � M4. Thesecells appear to mediate down-regulation of the host immune responses by at least twodistinct mechanisms: i) PGE2 production and 2) a cell contact-dependent, but non-major-histocompatibility-complex-specific, interaction.
Key words: prostaglandin E2, mixed leukocyte culture, down-regulation, GM-CSF
INTRODUCTION
Ineffective immune responses to a progressively grow-ing, yet immunogenic, tumor define the central paradox
in the study of tumor immunology. Among the mecha-nisms proposed to explain how tumors escape the con-
sequences of the host’s immunity are modifications ofimmunologic reactions [1-5] and altered regulation of
antitumor responses [6-10]. Phenotypic changes of themacrophages (M4) accompanied by functional alter-ations may be of major importance in defining theimbalance between effective host immune response andtumor progression.
The development of monoclonal antibodies raised
against M� surface antigens (anti-Mac-l and anti-Mac-2)by Springer and co-workers [ 1 1-1 3] has permitted thecharacterization of distinct M4 subsets. Mac-i, which
functions as the C3b receptor, is a glycoprotein of theLFA family of molecules. This molecule may play a rolein adhesion and target cell recognition, as well as being
an early differentiation antigen of M4 lineage. Mac-2 isa glycoprotein of 32 kDa molecular weight and its
expression appears to be a marker of M4 activationand/or differentiation.
In our model system, consisting of BALB/c mice
bearing an immunogenic, non-metastatic , murine mam-mary adenocarcinoma induced by dimethylbenzan-thracene (Dl-DMBA-3), we have previously reported areduced proliferative response to mitogens and tumor-
associated antigens. Additionally, there is decreasednatural killer cell and cytotoxic T lymphocyte (CTL)activities beginning 2 weeks post tumor implantation[14-16]. We also observe that this immune suppression
parallels a systemic increase of M4 lineage cells in theTB mice [17] . Macrophages from TB hosts are reportedto have different immunoregulatory activities compared
Received May 9, 1990, accepted June 8, 1990.
This research was supported by grant R37 CA 25583 from NCI, NIH.
Reprint requests: Diana M. Lopez, University of Miami School ofMedicine, Microbiology and Immunology (R-l38), P0 Box 016960,Miami, FL 33101.

Immunoregulation by Mac-i � Macrophages 127
to those from normal controls [6-8 , 18] . These functionaldifferences may, as with T and B cells, be reflected byphenotypic variations. In this study we present evidencethat TB murine splenic M� predominantly coexpressMac- 1 and Mac-2 antigens in contrast to only Mac- 1
antigen expression on most M� from normal mice.
Comparison of the immunoregulatory function of theseM4 show that Mac-l �2 M4 from normal mice act
positively as accessory cells in mitogen-induced andantigen-specific lymphocyte proliferation, whereas Mac-1 � M4 from TB mice significantly down-regulate Tand B cell proliferation. Secretion of POE2 by these TB
M4 appears to be the main soluble mediator of thissuppression. However, a MHC unrestricted contact-dependent mechanism seems to be responsible for thedown-regulation of T cell responses by Mac-l � M�.
The data supports a correlation of Mac antigen expressionand the immunoregulatory function of an M4 subset intumor-bearing hosts.
MATERIALS AND METHODSAnimals
The mice used in this study were 10-16 week age-
matched BALB/c (H-2”), C57B1I6 (H�2b), CBAJJ (H-2”),and DBAI2 (H�2d). All mice were bred and maintained inour facilities.
Tumor
The Dl-DMBA-3 transplantable mammary adeno-carcinoma was derived from a non-viral, non-carcino-
gen-induced hyperplastic outgrowth treated with 7 , 12-di-methylbenzanthracene [19] . This immunogenic tumorwas maintained by subcutaneous injection of 0.05 ml of
minced tumor suspension (approx. 1 X 106 viable cells).BALB/c mice were injected 28 days prior to use unless
otherwise noted. Littermate or age-matched mice were
used as normal controls.
Reagents
All standard reagents were obtained from GIBCO
(Grand Island, NY) unless otherwise noted. RPMI 1640supplemented with 20 mM HEPES, 5 X l0� mercap-toethanol (Sigma, St. Louis, MO), sodium pyruvate,L-glutamine, non-essential amino acids, MEM vitaminswas termed RPMI-S, and depending upon the assay, 5%or 10% heat-inactivated fetal calf serum (Hyclone, Lo-gan, Utah) was also added. Monoclonal anti-Mac-l and
Mac-2 antibodies were purified from supernatants ofhybridomas provided by Dr. T. Springer. MK-D6 (anti-I-A�) and 25-9-3S (anti-I-A1’) monoclonal antibodieswere obtained from Dr. P. Marrack. In some experimentsthe purified monoclonal antibodies were conjugated with
either biotin or fluorescein-isothiocyanate (FITC) by
standard protocols. Streptavidin-phycoerythrin (PE) wasused as the second step reagent for the biotinylatedantibodies. Dr. D. Mochizuki of Immunex Corp. (Seat-tle, WA) provided the rGM-CSF.
Histology
All histological sections were prepared by standardprocedures. For morphological evaluations splenocyte
suspensions were cytocentrifuged onto glass slides, airdried, and after methanol fixation stained with eitherMay-Grunwald-Giemsa or Diff-Quik Wright’s stain [20]and observed under oil immersion on a Nikon Optiphot
microscope. For nuclear-cytoplasmic ratio determina-tions, cells were cytocentrifuged, air dried, fixed at 4#{176}Cfor 1 mm in formalin/methanol, rinsed, and incubatedwith 0.2 mg/ml ribonuclease for 1 h at 37#{176}C.The cellswere then stained for 30 mm in 5% aqueous pyronine and
2% methyl green in acetic acid, rinsed, and read under oil
immersion.
Cell Separations
To obtain normal and TB purified M4, splenic single
cell suspensions were prepared and the cells fractionatedby centrifugation on Percoll (Pharmacia, Upsalla, Swe-den) continuous density gradients using a variation on the
method of Kurnick et al. [21]. Briefly, 20 ml of a I .070g/ml Percoll solution was centrifuged in a 25 ml poly-carbonate tube at 30,000g in a Beckman Type 30 rotor for15 mm at 4#{176}Cand decelerated with no brake. Up to1 x 108 mononuclear cells in HBSS were placed on thegradient and centrifuged at 400g for 20 mm at 4#{176}C.The
resulting bands were washed and analyzed. Further
enrichment for M� and M4 subsets was done by cellsorting on a FACS Ill (B-D, Mt. View, CA). The M4were separated by positive selection after staining with
fluorescein-conjugated anti-Mac- 1 and anti-Mac-2. Tand B cell subsets were obtained as previously described[15] with preparations routinely >95% pure and >90%viable.
Flow Cytometric Analyses
Cells for flow cytometric analyses and sorting weresuspended at 1 x 106 cells/ml in either phosphate buff-ered saline (PBS), or PBS with 1% bovine serum albumin(BSA) and 0.03 M sodium azide and stained at 4#{176}C.Cellphenotype was determined by fluorescent staining with
the appropriate positive and negative controls. Routinelythe following anti-mouse antibodies were utilized: anti-Mac-l , anti-Mac-2, anti-L3T4 (Dr. T. Malek), anti-Lyt
2.2 and anti-Thy 1 .2-FITC (B-D, Mt. View, CA), andanti-slg-FITC (Hybritech/Boehringer, Indianapolis, IN).Second step reagents, also added alone as controls, wereanti-rat-kappa-FITC and anti-rat-IgG-PE, both purchased
from HybritechlBoehringer (Indianapolis, IN). The

128 Watson et al.
FACS III, interfaced to an HP-86 micro-computer, wasused for cell analysis and sorting. Machine performancewas always checked against known standards (Micro-beads, B-D, Mt. View, CA) and unstained cells. The M�subsets were enriched by positive selection after staining
with either FITC or PE-conjugated anti-Mac-l and anti-Mac-2. Forward light-scatter and one- or two-colorfluorescence were analyzed according to standard proto-cols [22]. Though not reported, right angle scatter
patterns were occasionally done to further check cellpurity by size and/or density. These patterns wereconstant every time they were tested.
Ectoenzyme Assays
Lysates of 1 x l0� M4 were obtained by mixing thecells with 200 �.tl offreshly prepared 0.05% Triton X-lOO(Sigma, St. Louis, MO). The lysates were aliquoted forimmediate analysis or were stored at - 70#{176}Cfor assay ata later date. Protein concentration was determined using
a Bio-Rad kit with BGG as a standard. The proceduredescribed by Edelson and Cohn [23] using 0. 15 mM[3H]AMP was used to assay for 5’-nucleotidase activity.Specific activity was expressed as nM of AMP hydro-lyzed per minute per mg protein. Alkaline phosphodi-
esterase activity was assayed using 1 .5 mM p-nitrophenyl5 ‘-monophosphate as substrate [24]. Specific activitywas expressed as nM of product per minute per mg
protein. Leucine aminopeptidase activity was assayed
using 1 mM leucine p-nitroanilide [25].
Lymphocyte Transformation Assay
Mitogen lymphocyte transformation assays were per-formed as previously described [15]. The non-specificimmunoregulatory effects of TB Mac-l � cells were
assayed by co-culturing titrated numbers ofthese cells, orM4 from control animals, with purified normal cellsresponsive to a specific mitogen in RPMI-S medium. The
Mac- 1 � cells were exposed to 1 ,500 rads beforeculture, and in some experiments indomethacin at2 x l0� M was added to inhibit POE2 production. The
concentration of indomethacin used was determined tomaximally inhibit M4 POE2 secretion (80-90%, as
determined by RIA) while not affecting lymphocyteviability or proliferation. A lower concentration of in-
domethacin, i.e., l0� M, will inhibit approximately50% of PGE2 production, but this amount of inhibitionwas not sufficient for our studies. The possible effects ofother inhibitory factors were controlled for by the directaddition of PGE2 to cultures without indomethacin. This
protocol resulted in inhibition of lymphocyte prolifera-tion similar to that seen in cultures with indomethacin.
On day 2 of culture 0.5 1iCi [3H]TdR was added, the cellswere harvested 18 h later, and [3H]TdR incorporationwas measured in a liquid scintillation counter (Beckman
Instruments, Inc., Irvine, CA).
Mixed Lymphocyte Reaction (MLR)
In these assays, either BALB/c (H-2”) responder
splenic T-lymphocytes obtained by G-l0 and nylon wool
passage or lymph node cells were used. First, 2 x i05cells were co-cultured with 1 X l0� allogeneic C57BI/6
(H-21’) � CBA/J (H�2k) stimulator cells which had beenexposed to 1 ,500 rads. Normal splenic BALB/c Mac-1 �2 M4 or Mac- 1 � M4 from TB spleens in titrated
amounts of 0. 1-2.0 x l0� cells/well were used in MLRcultures to assess their effect on alloantigen-specificproliferation. Indomethacin at 2 x l0� M was added atday 0 in some experiments to inhibit POE2 production by
the added M4. On day 4 of culture 0.5 �xCi [3H]TdR wasadded, the cells were harvested 18 h later, and [3H]TdRincorporation was measured by liquid scintillation.
Prostaglandin Assay
Levels of POE2 in serum and tissue culture superna-tants were assayed using the protocol provided in the
PGE2 RIA kit from Seragen (Boston, MA). Prostaglan-din in the samples was extracted with ethyl acetate anddimethylformamide and the POE2 containing residue was
evaporated to dryness under a stream of liquid nitrogen.The samples were then subjected to cold competitionRIA. Extraction efficiency was determined by subjectinga known concentration of [‘25I]PGE2 standard to thesame procedure. POE2 levels were determined by extrap-olation of a linear graph of cpm vs. a known concentra-tion of a PGE2 standard.
Marbrook Chamber Assay
To directly assess the relative contributions of solublefactor(s) produced by Mac-l � M4 and/or cell contact-
dependent mechanisms, a Marbrook culture chambersystem from CoStar (Cambridge, MA) was used. The
two chambers were separated by a filter with a pore sizesufficient for the passage of < 120 kDa MW material.M� were obtained by Percoll density gradient centrifu-gation and irradiated with 1 ,500 rad prior to co-culture ina Con A proliferation assay with responder splenocytes at
2.5 x l0� per well. Indomethacin was added at a con-centration of 2 x l0� M. Results are expressed as themean ± SEM of 4 wells in each of 4 experiments.
RESULTSIdentification of Mac-I � Macrophagesin TB Spleens
An increase in the percentage of splenic M4 wasobserved with increasing tumor burden (approximately
3% in the spleens of normal mice to 17% in the spleensof TB). A distinct M� population was observed aftertumor implantation in splenic imprints and histopatho-
logical sections. These M4 resemble resident bone

NORMAL4
10
210
10
TUMOR BEARER
wUzU
w0:D-JLA.0
LU
00
3
2
10
10
10
LUUzLUUU)LU
0:�-JLA.
LU
00-A
.. .
. .;......�.�‘:
�:::
0 1 2 310 10 10 10
A LOG GREEN FLUORESCENCE
Fig. 1 . Flow cytometric analyses of two-color immunofluores-cence patterns of splenic M4 populations, enriched by Percolldensity gradient centrifugatlon, from normal (A) and TB (B).Cells were stained wIth anti-Mac-i-FITC and biotinilated anti-Mac-2 followed by streptavidin-PE. The percentages of cells of
0 1 2 3
10 10 10 10
B LOG GREEN FLUORESCENCE
each distinct phenotype are given below (cell phenotype, M4from normal mice, M4 from TB). Panel i : Mac-i 2, i6.7%, 9.4%.Panel No. 2: Mac-P2, 78.1%, 4.7%. Panel No. 3: Mac-i2�,4.0%, 6.i%. Panel No. 4: Mac-i � i.2%, 79.8%.
Immunoregulation by Mac-i � Macrophages 129
marrow M4 with a mean diameter > 15 p.m and a
relatively high nuclear/cytoplasmic ratio. Pyronine stain-ing demonstrated diffuse DNA staining in these cells
suggestive of active proliferation. Splenocytes, isolatedon continuous Percoll density gradients, formed five
bands. Most of the M4 from the TB were found in theband at density 1 .055 g/ml. The majority of the M4 fromnormal mice, however, banded at density 1 .050 g/ml.Expression of Mac- 1 and Mac-2 were of interest sincethey have been shown to be important markers of M4
differentiation and/or activation [1 1-13]. The Percollgradient-enriched splenic M4 were analyzed using dual
immunofluorescence staining for Mac- 1 and Mac-2 an-tigen expression. The flow cytometric histograms
(Fig. 1 ) show the presence in TB mice of a cellpopulation co-expressing Mac- 1 and Mac-2 antigens. Incontrast, in normal BALB/c mice most of the M4 wereMac- 1 �2 - . The TB Percoll purified M� populations
contained up to 80% Mac-l � cells; however, this
phenotype accounted for only background levels of 1-2%in the Percoll-purified normal mouse splenocyte popula-
tions. Furthermore, the Mac- 1 � cells averaged 17.4%of the total splenocytes in mice bearing 4 week tumors,
whereas Mac-i 2� cells accounted for <3% of the total
splenic population in both normal and TB mice.
Ectoenzyme Assays
A study of the ectoenzyme profiles of TB splenic M4
(Table 1) showed that they have lower levels of 5’-
nucleotidase, and higher levels of alkaline phosphodies-
terease than splenic M4 from normal mice. In contrast,the levels of non-specific esterase, myloperoxidase,
leucine aminopeptidase, and N-acetyl-B-D-glucosamin-idase activities were comparable to those from normalmouse spleen cells. It is known that resident peritoneal
and splenic M4 from normal mice express 5’-nucleoti-
3 4
ii I � )-:t��.k �. 2
TABLE I . Ectoenzyme Profiles in Splenlc Macrophages From Normal and Tumor-Bearing Micea
Enzyme
Macrophage sourceb
Normal splenocytes TB splenocytes
Nonspecific esterase >95% >95%5’-nucleotidase (nM/mm/mg protein) 46.8 ± 1.3 24.1 ± 3.5Leucine aminopeptidase [U(MM/min/l09 cells)l 1.2 ± 0.1 2.5 ± 1.3Alkaline phosphodiesterase (nM/mm/mg protein) 15.6 ± 1.3 22.4 ± 1.7N-acetyl-B-D-glucosaminidase (U/hr/lO �.tg DNA) 426.0 ± 21.0 469.0 ± 74.0Myeloperoxidase - -
aResults expressed as mean cpm X l0� ± S.E.M. of 4 experiments.
bNormal and TB splenic M#{231}6obtained by plastic adherence.

130 Watson et al.
dase and N-acetyl-B-D-glucosaminidase, but the levels
of these enzymes are diminished on inflammatory cells oron cells activated immunologically. Therefore, failure toexpress these ectoenzymes is thought to be a character-istic of activated M4. Expression of leucine aminopep-
tidase and alkaline phosphodiesterase is higher in acti-vated M4 than in resting M4. These results suggest thatthe tumor-induced splenic M4 are at least in a potentially
more metabolically active state than normal splenic M4.
The lack of staining with myeloperoxidase, which stainsPMN and myeloid progenitors, but not cells of themonocyte series, argues against the possibility that theseM4 belong to the myeloid series.
Down-Regulation of Lymphocyte Responses byMac-i � Macrophages
We have previously documented that splenic lympho-
cytes from tumor-bearing animals respond poorly tomitogenic stimulation [14, 15] . An altered accessory cell
function may be partly responsible for this downregula-tion. Normal splenocytes were depleted of M� by two
passages over Sephadex G- 10 and separated into B and T
cell-enriched populations by nylon wool fractionation.Splenic Mac-l �2 cells from normal mice were Percollgradient purified. These cells were used for normalcontrols since they represent the majority of the M4 fromanimals without tumors. Mac-l � cells from TB wereobtained from Percoll gradients and further enriched bysorting based on the expression of Mac-2 antigen on their
surface. As mentioned above, this population defined byflow cytometry comprises the majority of the M4 fromTB. The normal T cell proliferative responses to Con A,known to be accessory cell dependent, were assessed inthe presence of varying concentrations of these M4.Although B cell polyclonal stimulation by LPS is not
thought to be accessory dependent, Mac- 1 � 2 � M4 werealso co-cultured with B cells to determine the ability ofM4 from TB to modulate B cell proliferation. The results(Table 2) show that the T cell Con A response at a 10:1
responder:accessory cell ratio was optimal in the pres-ence of M4 from normal mice. M4 from TB, however,
did not provide efficient accessory cell function at anytested ratio. Less than 10% of the normal T cell prolif-
erative response to Con A could be elicited under those
TABLE 2. Effect of Macrophages From Tumor-Bearing Mice on Mitogen-InducedBlastogenic Transformation
ponder cells
Additions to culture
Res No. accessory cellsa Con Ab LPS
2 X l0� T cellsNone 0.8 ± 0.3 ND.
Normal M#{248}
1 X l0�
2 X l0�4 X I0�
127.8 ± 13.6
155.2 ± 9.586.9 ± 5.7
1.3 ± 0.6
ND.ND.
M� from TB1 X l0�2 X l0�4 X l0�
12.7 ± 4.2 (90.1Y�*4.2 ± 1.5 (973)*5.6 ± 0.9 (93.6)*
3.2 ± 1.2ND.ND.
2 X l0� Normal M�+
M#{248}from TBI X l0�2 X l0�4 X iO�
79.7 ± 3.9 (48.6)*51.4 ± 2.8 (66.9)*38.6 ± 3.4 (75.1)*
5.3 ± 0.7ND.ND.
2 X l0� B cellsNone 1.4 ± 0.6 78.2 ± 6.5
M#{248}from TB1 X l0�2 X l0�4 X l0�
ND.ND.ND.
21.2 ± 3.5 (72.9)8.5 ± 2.7 (89.1)7.4 ± 1.8 (90.5)
aMacrophages from normal and tumorbearing mice were purified by Percoll gradients as described in
“Materials and Methods.”bResults expressed as cpm X I O� minus control cpm from parallel cultures without mitogen ± S.E.M.
CNos in parentheses represent percent in the reduction of I3HITdR incorporation.
*Values are significantly (P< .01 by the Student t test) different from cultures containing Mac- I +2
accessory cells from normal mouse spleens.

0.1 0.5 1.0 2.0
Number of Macrophages x 10
Fig. 2. C57BI/6 T cells stimulated by allogeneic M4 fromnormal and TB BALB/c mice. G-iO and nylon wool-purifiedC57BI/6 (H�2b) responder T cells were cultured with normalsyngeneic (C57Bl/6) or allogeneic (BALB/c) normal and TBsplenic M4. Mac-i � M4 obtained from 4 week TB BALB/c micewere purified by Percoll gradients. All stimulator cells weretreated with 500 rads prior to culture. Values represent meancpm ± S.E.M. of 4 wells from 6 experiments.
Immunoregulation by Mac-i � Macrophages 131
conditions. In parallel crowding control cultures, irradi-
ated splenocytes from normal mice were added in appro-priate numbers. Such cultures gave the same results asthose to which no extra cells were added. Furthermore,
addition of low numbers of M4 from TB to cultures inwhich appropriate antigen presenting cells (Mac-l �2)
from normal mice were added resulted in a significantreduction ofthe Con A response. This indicated that therewas an actual down-regulatory function exerted by the
M4 from TB mice. In previous studies we had deter-mined that the Percoll-purified M4 from normal micehad no effect on B cell responses to LPS. However, at alltested ratios, co-culture with tumor M4 from TB resulted
in a 73-91% decrease in B cell polyclonal proliferation.Crowding controls with irradiated splenocytes did not
affect these results.Macrophages from normal and TB mice were used as
stimulators in MLR cultures with H-2�’ allogeneic re-
sponder T cells from C57 BL/6 mice to determine if therewas an alteration in their potential to induce immunopro-liferation across an MHC barrier. A significant decreaseof alloreactivity was observed when TB M4 were used atall ratios tested (Fig. 2). A similar phenomenon wasobserved when responders belonged to a phenotype ofH�2k, i.e., CBAIJ (data not shown).
Soluble Factor(s) Released byMac-i � Macrophages
To investigate the possible role of soluble factor(s) inthe decrease of lymphoproliferative responses by Mac-1� TB M4, cultures were set up with either Mac-
0
EU
l�2 M4 from normal mice or Mac-l � M4 from TBalone, or in combination with normal mouse T cells thathad been stimulated with Con A. The cultures wereincubated for 48 h and the supernatants were harvested.
Serial dilutions of these supernatants plus either Con A orLPS were added to mitogenic transformation assaysemploying normal whole spleen cells as responders(Table 3). The data show that TB M4 produce solublematerial(s) capable of suppressing the mitogenic re-
sponses of normal splenic T and B cells. Mac-i � TBM4 constitutively produce the putative factor(s), sincesimply culturing these cells for a 48 h period allowed for
sufficient accumulation of a suppressive product. Inaddition, supernatants from the above M� cultures wereadded to allogeneic MLR cultures to determine if theirwas a down-regulation of immune proliferation across an
MHC barrier by soluble factor(s). Responder cells from
C57B1/6 (H-21’) animals were cultured with irradiatednormal BALB/c splenocytes as stimulators. A decreasedproliferation of MLR cultures in response to histoincom-patible stimulators was observed in secondary cultures
treated with a 1 :2 dilution of supernatants from primarycultures of M4 from TB. This effect was shown by adecrease in [3H]TdR from an initial level of 53,000
± 3,000 cpm to 22,000 ± 3,500 cpm. Supernatants, at
the same dilution, from primary M4 cultures from
normal mice did not cause any reduction in [3H]TdRincorporation (56,000 ± 8,000 cpm). Similar results
were obtained using other strains of mice such as CBAIJ(H�2k) as responders. As seen in Table 3, a higher levelof suppressor activity was detected in the supernatantsfrom T cell-Con A : M4 from TB co-cultures than inthose from TB M4 alone in both mitogen and alloantigenexperiments. These results indicate that the productionand release of suppressive factors was greatly enhancedby the interaction between activated T cells and M4,which is more reflective of the in vivo situation in TB.
PGE2 Production by Macrophages
The production and release of PGE2 by M4 has oftenbeen associated with non-specific suppression of immuneresponses [27,28]. Therefore, culture supernatants from
M4 from normal mice and TB were tested for PGE2activity by radioimmunoassay (Table 4). Significantlyhigher amounts of PGE2 were detected in the TB M4cultures than in those from normal mice. The inclusion of2 x l0� M indomethacin, a cyclooxygenase inhibitor,
decreased PGE, synthesis and release by 70-80% in bothMac-i �2 normal and Mac-i � TB M4 cultures.Since it has been suggested that inhibition of PGE,
synthesis by indomethacin is also due to over-production
of toxic oxygen radicals, catalase was added to culturescontaining indomethacin. No change could be observedin the results after such treatment. To determine if an
abnormal level of POE2 was present in TB, in vivo serum
i32 Watson et al.
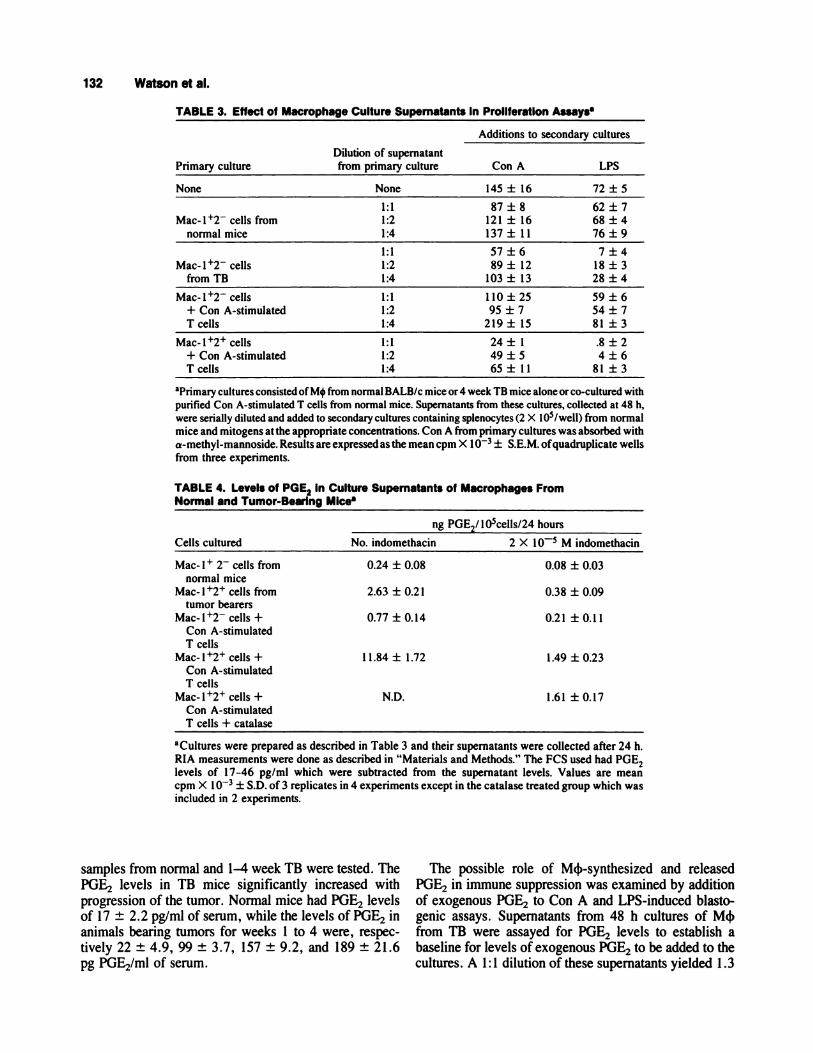
TABLE 3. Effect of Macr ophage Culture Supernatant s In Proliferation Assaysa
Dilution of supernatant
Additions to secondary cultures
Primary culture from primary culture Con A LPS
None None 145 ± 16 72 ± 5
1:1 87±8 62±7Mac-l�2 cells from 1:2 121 ± 16 68 ± 4
normal mice 1:4 137 ± I 1 76 ± 9
1:1 57±6 7±4
Mac-l�2 cells 1:2 89 ± 12 18 ± 3from TB 1:4 103 ± 13 28 ± 4
Mac- 1 �2 cells 1 : 1 1 1 0 ± 25 59 ± 6
+ Con A-stimulated 1:2 95 ± 7 54 ± 7T cells 1:4 219 ± 15 81 ± 3
Mac-l�2� cells 1:1 24 ± 1 .8 ± 2+ Con A-stimulated 1:2 49 ± 5 4 ± 6T cells 1:4 65 ± 1 1 81 ± 3
aprimary cultures consisted of M4 from normal BALB/c mice or 4 week TB mice alone or co-cultured with
purified Con A-stimulated I cells from normal mice. Supernatants from these cultures, collected at 48 h,were serially diluted and added to secondary cultures containing splenocytes (2 X l05/well) from normalmice and mitogens at the appropriate concentrations. Con A from primary cultures was absorbed with
a-methyl-mannoside. Results are expressed as the mean cpm X I O� ± S.E.M. ofquadruplicate wellsfrom three experiments.
TABLE 4. Levels of PGE2 in Culture Supernatants of Macrophages FromNormal and Tumor-Bearing MIce8
Cells cultured
ng PGE2/ lO5cells/2 4 hours
No. indomethacin 2 X iO� M indomethacin
Mac- I � 2 cells from 0.24 ± 0.08 0.08 ± 0.03normal mice
Mac- 1 � cells from 2.63 ± 0.2 1 0.38 ± 0.09tumor bearers
Mac-l�2 cells + 0.77 ± 0.14 0.21 ± 0.11Con A-stimulatedT cells
Mac- 1 � cells + I 1.84 ± 1.72 1 .49 ± 0.23Con A-stimulatedT cells
Mac-l�2� cells + ND. 1.61 ± 0.17Con A-stimulatedT cells + catalase
aCultures were prepared as described in Table 3 and their supernatants were collected after 24 h.
RIA measurements were done as described in “Materials and Methods.” The FCS used had POE2levels of 1 7-46 pg/ml which were subtracted from the supernatant levels. Values are mean
cpm X 1 � ± S.D. of 3 replicates in 4 experiments except in the catalase treated group which wasincluded in 2 experiments.
samples from normal and 1-4 week TB were tested. ThePOE2 levels in TB mice significantly increased withprogression of the tumor. Normal mice had PGE2 levelsof 17 ± 2.2 pg/mi of serum, while the levels of PGE2 inanimals bearing tumors for weeks 1 to 4 were, respec-tively 22 ± 4.9, 99 ± 3.7, 157 ± 9.2, and 189 ± 21.6pg PGE2/ml of serum.
The possible role of M�-synthesized and released
POE2 in immune suppression was examined by additionof exogenous PGE2 to Con A and LPS-induced blasto-genic assays. Supernatants from 48 h cultures of M�from TB were assayed for POE2 levels to establish abaseline for levels of exogenous PGE2 to be added to thecultures. A 1 : 1 dilution of these supernatants yielded 1.3

-0--- Con A
. Ips
10 20
-a- C5761/6
. CBA/J
30
20
10
0 10 20 30
Immunoregulation by Mac-i � Macrophages 133
ng/ml PGE2 and this calculation was the basis for the
culture conditions used. The results in Figure 3 (upperpanel) show that levels of POE2 similar to those producedin culture by M4 from TB were capable of decreasing[3H]TdR incorporation by Con A-stimulated normal Tcells and LPS-activated normal B cells. The B cells weremore susceptible to PGE�-mediated suppression (P < .05)
than T cells. The addition of titrated amounts of PGE2 toallogeneic MLR cultures was done to determine if thissoluble factor also contributed to decreased responses toalloantigen seen with the addition of TB M4 culture
supernatants. The data in Figure 3 (lower panel) indicate
that the alloantigen responses of C57B1/6 or CBAIJ Tcells stimulated by irradiated BALB/c splenocytes werediminished by POE2 only at relatively high concentra-tions (>5 ng). The data provided evidence that POE2 islikely to be partially responsible for the suppression ofimmune responses. To further define the role of TB
M4-produced PGE2, coculture experiments with irradi-
0
xE
U
0
x
EU
Effect of PGE 2 on Mitogen Response5
PGE 2(ng)
Effect of PGE2On MLR
PGE 2(ng)
Fig. 3. Con A and LPS blastogenesis assays (upper) wIthnormal BALB/c mouse spleen cells as responders and MLRassays (lower) with C57BI/6 and CBA/J cells as responders andBALB/c splenocyte stimulators as described In “Materials andMethods.” PGE2 was added at the beginning of the cultures. Allvalues are the mean cpm ± S.E.M. of 4 wells from 4 experi-ments.
ated M4 and mitogen-stimulated splenocytes were per-
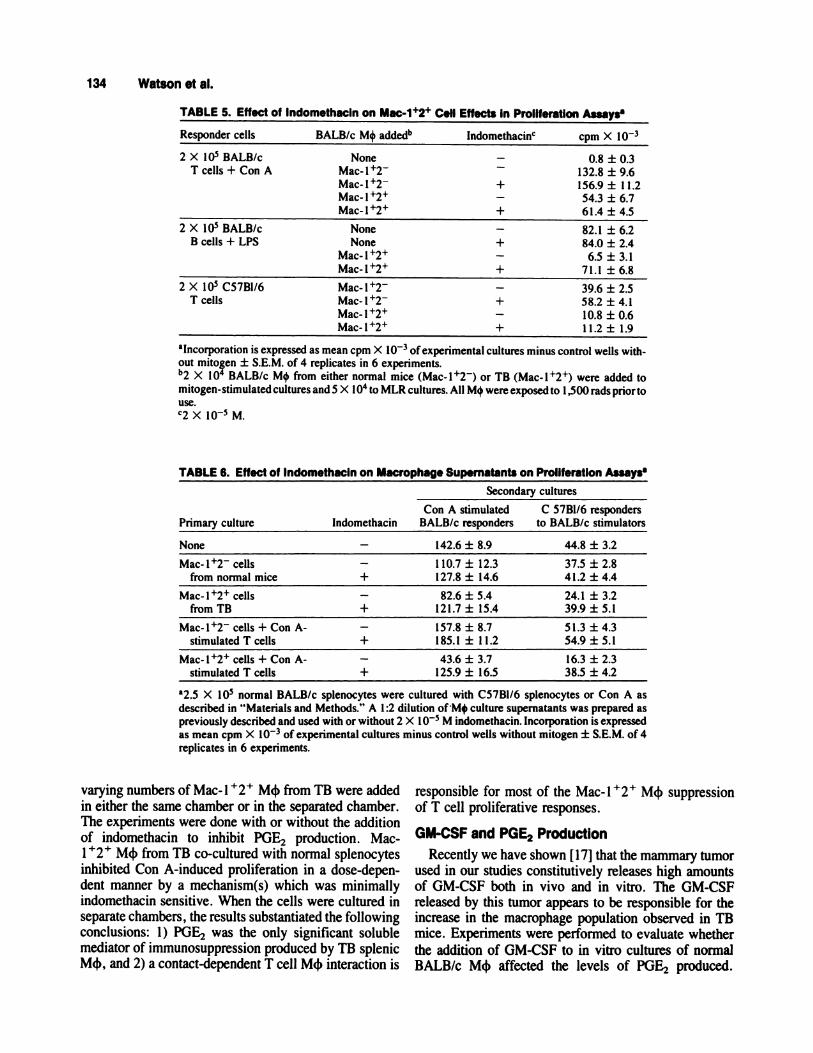
formed in the presence or absence of indomethacin.Table 5 shows that 2 x iO� M indomethacin re-stores >85% of B cell responses to LPS. This suggeststhat POE2 released by TB M� is the main inhibitor of thepolyclonal B cell response to mitogens in vitro. Poly-clonal T cell responses to Con A, however, were stillsignificantly inhibited by M4 from TB in the presence of
indomethacin. Furthermore, indomethacin did not re-verse the decrease in allogeneic-induced proliferation
assays using C57B 1/6 mice in the presence of M4 from
TB (Table 5). It should be noted that indomethacin is
capable of overcoming the slight reduction which existedin these assays by addition of Mac-i �2 M4. This
indicates that in high numbers even M4 from normalmice may exert an inhibitory activity on MLR. These
data indicate that suppression by the Mac- 1 � M� fromTB on the polyclonal T cell response is due to amechanism other than POE2 and this mechanism isdominant when POE2 production is reduced.
At least two possibilities exist to explain the T cellsuppression observed above. There is another solublefactor present, or M4-T cell contact may be necessary.To test these alternatives, supernatants from M4 primary
cultures from normal mice and TB were added to ConA-induced T cell proliferation assays in the presence orabsence of indomethacin (Table 6). Addition of super-
natants from Mac-i �2 normal primary cultures did notsignificantly alter Con A responses in the cell prolifera-
tive assays whether or not indomethacin was added.However, the addition of primary culture supernatants ofMac- 1 � 2 � cells from TB was very suppressive in the
proliferation assay and could be completely reversed bythe addition of indomethacin. Furthermore, similar re-suits were obtained with alloantigen-stimulated T cellsfrom C57B1/6 were utilized in the secondary cultures.
These results are in contrast with those observed inpreceding studies (Table 5) when Mac-i � cells were
directly added to the Con A-simulated T cells. In thoseexperiments, indomethacin could not reverse the sup-pressive effect on T cells by Mac-l � M4 while itreadily could reverse suppression caused by a soluble
factor(s) alone. A slight enhancement of the proliferativeactivity was also observed in indomethacin-treated cul-tures of Mac-l �2 cells from normal mice. The resultsin Table 6 suggest that POE2 may be the major soluble
suppressor product of polyclonal T cell proliferation byM4 from TB, while a second cell-to-cell contact-depen-dent suppression may be dominant in co-cultures.
Cell Contact-Dependent Suppression
In order to further assess the relative contribution insuppression of soluble factor(s) and/or cell contact-dependent mechanisms by Mac- 1 � cells, Marbrookchamber experiments were performed (Table 7). NormalBALB/c splenocytes were placed in one chamber and
i34 Watson et al.
TABLE 5. Effect of lndomethacln on Mac-i’�2’ Cell Effects in Proliferation Assays
Responder cells BALB/c M4 added�’ Indomethacin cpm X l0�
2 X l0� BALB/c
T cells + Con ANone
Mac-l12Mac-l�2Mac-l�2�Mac- I �
-
+
-
+
0.8 ± 0.3132.8 ± 9.6156.9 ± I 1.2
54.3 ± 6.76 1.4 ± 4.5
2 X l0� BALB/cB cells + LPS
NoneNone
Mac-1�2�
Mac-l�2�
-
+
-
+
82.1 ± 6.284.0 ± 2.4
6.5 ± 3.171.1 ± 6.8
2 X l0� C57B1/6T cells
Mac-l�2Mac-l�2
Mac-I�2�Mac-l�2�
-
+
-
+
39.6 ± 2.558.2 ± 4.1
10.8 ± 0.611.2 ± 1.9
alncomoration is expressed as mean cpm X l0� ofexperimental cultures minus control wells with-
out mitogen ± S.E.M. of 4 replicates in 6 experiments.b2 x l0� BALB/c M4 from either normal mice (Mac-l�2) or TB (Mac-l�2�) were added to
mitogen-stimulated cultures and 5 X iO� to MLR cultures. All M4 were exposed to 1,500 rads prior touse.C2 X l0� M.
TABLE 6. Effect of Indom ethacin on Macro phage Supernatants on Proliferation Assays
Primary culture Indomethacin
Secondary cultures
Con A stimulated C 57Bl/6 respondersBALB/c responders to BALB/c stimulators
None - 142.6 ± 8.9 44.8 ± 3.2
Mac-l�2 cellsfrom normal mice
-
+1 10.7 ± 12.3 37.5 ± 2.8127.8 ± 14.6 41.2 ± 4.4
Mac-l�2� cellsfrom TB
-
+82.6 ± 5.4 24.1 ± 3.2
121.7 ± 15.4 39.9 ± 5.1
Mac-l�2 cells + Con A-
stimulated T cells
-
+157.8 ± 8.7 51.3 ± 4.3185.1 ± 1 1.2 54.9 ± 5.1
Mac-l�2� cells + Con A-stimulated T cells
-
+43.6 ± 3.7 16.3 ± 2.3
125.9 ± 16.5 38.5 ± 4.2
a�5 x l0� normal BALB/c splenocytes were cultured with C57B1/6 splenocytes or Con A as
described in “Materials and Methods.” A 1 :2 dilution ofM+ culture supernatants was prepared aspreviously described and used with or without 2 X I 0� M indomethacin. Incorporation is expressed
as mean cpm X l0� ofexperimental cultures minus control wells without mitogen ± S.E.M. of 4replicates in 6 experiments.
varying numbers of Mac- 1 � 2 � M4 from TB were added
in either the same chamber or in the separated chamber.The experiments were done with or without the additionof indomethacin to inhibit POE2 production. Mac-1 � M4 from TB co-cultured with normal splenocytesinhibited Con A-induced proliferation in a dose-depen-dent manner by a mechanism(s) which was minimallyindomethacin sensitive. When the cells were cultured in
separate chambers , the results substantiated the followingconclusions: 1) POE2 was the only significant solublemediator of immunosuppression produced by TB splenicM4, and 2) a contact-dependent T cell M4 interaction is
responsible for most of the Mac-i � M4 suppression
of T cell proliferative responses.
GM-CSF and PGE2 Production
Recently we have shown [17] that the mammary tumor
used in our studies constitutively releases high amountsof OM-CSF both in vivo and in vitro. The OM-CSFreleased by this tumor appears to be responsible for theincrease in the macrophage population observed in TBmice. Experiments were performed to evaluate whether
the addition of OM-CSF to in vitro cultures of normalBALB/c M� affected the levels of POE2 produced.

Immunoregulation by Mac-i � Macrophages 135
TABLE 7. The Effect of Marbrook Chamber Separation onImmunoregulatlon of Con A Responses by Mac�i+2+ Cellst
ChamberCells added separation lndomethacin cpm X l0� % of control
None -
-
-
+
149.7 ± 9.1
163.4 ± 15.2
100”100B
1 X l0� Mac-l�2� - - 83.3 ± 10.4 55.6acells from TB -
+
+
+-
+
95.5 ± 10.6120.3 ± 15.9
144.7 ± 13.9
584b
80.3a885b
2 X 1O� Mac- 1� - - 69.2 ± 10.4 46.2a
cells from TB -
+
+
+-
+
81.9 ± 15.71 10.2 ± 20.9136.7 ± 21.2
501b
73.6k’836b
4 X iO� Mac-1�2� - - 41.4 ± 17.1 27.7a�
cells from TB -
+
+
+-
+
62.9 ± 10.2107.1 ± 20.4129.1 ± 12.8
385b
71.5’790b
t2.5 X l0� normal BALB/c splenocytes were used as responders in Con A (10 �g/ml) inducedproliferative assays using Marbrook chambers as described in “Materials and Methods.” Where
indicated 2 X 1O� M indomethacin in EtOH was added. Mac- 1 � M4 were obtained by density
centrifugation and further purified by positive cell sorting selection. Incorporation is expressed asmean cpm ofexperimental cultures minus control wells without mitogen ± S.E.M. of4 replicates in 4
experiments. Percent of control was calculated as follows: values for a (without indomethacin) or b(with indomethacin) divided by either A or B (stimulation with no chamber separation and no added
Mac-1�2� cells from TB) X 100%.
TABLE 8. Effect ofin Vitro rGM-CSF Treatment onBone Marrow, and Perltoneal Exudate Cells
PGE2 Production by Cultured Spleen,
POE2 levels (ng/ml)
Cell source Media alone Media + rGM-CSF (500 U/mI)
Spleen (5 X l05/ml) 0.8Bone marrow (5 X l05/ml) 1.5PEC (1 X 105/ml) 0.6
3.89.85.0
aSupernatants from untreated or rGM-CSF treated cultures were harvested at day 5 and assayed for
POE2 levels by RIA as described in “Materials and Methods.” Values are the mean of duplicate
cultures of a representative experiment.
Table 8 shows that rOM-CSF added in vitro to cultures ofspleen, bone marrow, and peritoneal cells for 5 daysincreased the culture supernatant levels of POE2 ascompared to untreated cultures. The data, therefore,suggests that tumor-derived OM-CSF may be a possible
cause of increased POE2 production by M4 from TB
mice.
DISCUSSION
Splenic lymphocytes from BALB/c TB mice have been
shown to have reduced mitogenic proliferative responses[15] and decreased MLR responses when they arestimulated with ailoantigens from H�2b or H�2k mice
[16]. It is possible that there are host cells induced by the
tumor presence which act to down-regulate the immuneresponse. A histologically unique population of spleno-
cytes of the macrophage lineage has been found to beassociated with mammary tumor progression. This studyanalyzes these cells and the role they may play in theregulation of the BALB/c host immune response to the
Dl-DMBA-3 mammary adenocarcinoma.
There is an influx, or expansion of a resident popula-
tion, of cells which we characterized as Mac- 1 �macrophages. This population was seen by 2 weeks aftertumor implantation and peaked at 4 weeks. Such cellscomprise less than 2% ofthe splenocytes in normal mice,but account for 15-25% ofTB splenic leukocytes. Garner

136 Watson et al.
and Elgert [1 8] extensively analyzed changes in mac-rophage-antigen expression in BALB/c mice with a
methylcholanthrene-induced transplantable non-metastaticfibrosarcoma. Using a series of monoclonal antibodiesspecific for M4 cell-surface antigens, they found a shiftin M4 subpopulations associated with changes in tumor
burden . In normal mice Mac- 1 was detected on 39% ofsplenic adherent cells, whereas 63% of equivalent TB
cells expressed this antigen. Additionally, 27% of thenormal splenic adherent cells were Mac-2 � , compared to40% in TB . In other studies by this group [28] it was
found that the Mac-2� M4 from TB were responsible fora factor-mediated suppression of MLR which was in
parallel to the production of POE2 . Garner et al. [29] andYurochko et al. [30] also have presented evidence that the
shift in M4 phenotypes during tumor growth is associ-ated with their immunosuppressive activity. In our stud-ies, after isolation by cell sorting based on forward andright angle light scatter and/or Percoll density gradient
centrifugation, we found that splenic M4 from bothnormal and TB BALB/c mice could be significantlyenriched (85-90%). This methodology allows for much
greater reproducibility of results by excluding adherentcells of non-macrophage/monocyte lineage such as den-dritic cells, and including M� that are weakly adherent,such as immature macrophage/monocytes. Two colorflow cytometric determinations demonstrated that, in ourmodel system, 80% of the purified tumor splenic M�co-expressed Mac- 1 and Mac-2 antigens. Further, the
Mac-i � 2 � M� of the mammary TB , morphologicallyand histochemically, closely resembles immature, ac-tively cycling, but relatively metabolically non-activated
cells, in spite of the presence of the Mac-2 antigen, amarker of differentiation and activation. Our experimentsshowed that the increase in the Mac-i � populationcorrelates with a profound down-regulation of immuneresponses. In interpreting whether the important featureof the cells, in terms of their function, is the shift inphenotype or an increase in the overall number of M4,we favor the first alternative. Thus, we believe that the
phenotypic change from a majority of Mac- 1 �2 M4 innormal mice to Mac- 1 � M4 in TB causes the func-tional alterations that lead to the down-regulatory prop-erties of these cells. This is based on the fact that we add
the same numbers of Mac-i �2 cells to the controlcultures and the experimental group containing Mac-
l�2� M� from TB.Cells of the macrophage/monocyte lineage have been
reported to function as effectors and/or mediators ofimmune suppression in different primary and transplant-able tumor systems including those of spontaneous, viral,or chemical origin [6-10,26,27,29,30]. Hyde et al. [26]found results similar to ours in a study of BALB/c
splenocyte response to PHA after injection of a herpes-
transformed embryonic tumor line, but characterized
their “suppressor” M4 only by phagocytic properties and
0-10 adherence. Bonaventure et ai. [7], using Fischerrats bearing an ascites mammary adenocarcinoma, pre-
sented data correlating an increase in the splenic mac-rophage:lymphocyte ratio with tumor burden and thepresence of adherent splenocytes with the ability toinhibit Con A responses by T lymphocytes. Immunosup-pression may also be mediated by a variety of solubleproducts secreted by M4 or tumor [26-28,3 1-34]. In
some studies, M4-derived POE, was reported to be theonly mechanism that suppressed the T cell function [27].In our model system, however, although POE2 is the
main factor involved in the downregulation of B cellfunctions, direct cell-cell contact appears to be the majormechanism of suppression of T cells in TB mice.
Mac- 1 �2 � M4 were found to be ineffective accessorycells in normal T cell mitogen proliferation or alloantigenresponses. Additionally, those cells completely inhibitedB cell responses to LPS, which is not accessory-celldependent. Upon co-culture with normal M4 as alterna-tive accessory cells, the TB M4 inhibited the T cell
responses, suggesting that they do play an active role inthe inhibitory process. In fact, our studies with theMarbrook chambers indicate that TB Mac- 1 � cellsmainly suppress T cell functions by a contact-dependentcell-to-cell interaction. The B cell responses, however,were shown to be mainly suppressed by POE2-mediateddown-regulation. Correlation of the kinetics of down-regulated host immune function and the appearance ofsplenic Mac- 1 � M� suggests that these TB splenicM� are one of the major effectors of immune down-
regulation during mammary tumorigenesis.We have recently found that the tumor utilized in this
study releases high amounts of OM-CSF, both in vivo
and in vitro, which appears to contribute to the expansionof cells of the M� lineage in TB mice [17]. Similarfindings have also been reported by Tsuchiya et al. [35].
These investigators have found that several tumors pro-duce OM-CSF that upon incubation with splenic mono-nuclear cells promotes the induction of Mac-i � IA�Thy 1 - L3T4 Lyt 2 1g phenotypes with suppressive
functions. In vitro splenocytes, bone marrow, and peri-toneal cells from normal mice can be stimulated to releasePOE2 by rOM-CSF. These data imply that the increase of
POE2 production observed in TB M� may be due to theGM-CSF constitutively produced by the tumor itself.Additionally, preliminary in vivo data show that an
increase of Mac-i � and Mac-2� cells in the spleen andbone marrow of normal mice can be induced by injectionof 400 ng of rGM-CSF twice daily for 2 weeks.Heidenreich et al. [36] have demonstrated that LPS andy-INF activation of M� to become highly cytotoxic couldbe reversed by 20 h of pretreatment with rGM-CSF. The
mechanism of this depressed M� function appears to be
due to the induction of increased levels of PGE2 which inturn suppress TNF-a release. In previous studies we havefound that M� from TB , as compared to those of normal

Immunoregulation by Mac-i � Macrophages 137
mice, had a decreased capacity to become cytotoxicagainst tumor cells [16]. Taken together, the data supportthe hypothesis that tumor derived factor(s) may directly
induce and/or expand a population of suppressor M4 inTB mice in order to circumvent the host’s efforts toeliminate the tumor. Phenotypic and functional changesof M� observed during tumor progression suggest that
the tumor induces the expansion of a population of hostM4 that effect immunosuppression by releasing solublefactor(s) and/or by a cell-to-cell contact-dependent mech-anism. Our results with the D1-DMBA-3 tumor system,therefore, suggest that the changes in the macrophage/
monocyte compartment during tumor growth may be animportant factor associated with non-specific host immu-nosuppression.
ACKNOWLEDGMENTS
The excellent technical assistance of Mantley Dorsey,Jr. is gratefully acknowledged. We would also like tothank Dr. Mayra Lopez-Cepero and Dr. Eduardo Soto-mayor for helpful review and discussions.
REFERENCES
I . Moller, 0. Effect on tumor growth in syngeneic recipients of
antibodies against tumor-specific antigens in methylcholanthreneinduced mouse sarcomas. Nature 204,846, 1964.
2. North, R.J. Development of the antitumor immune response.
Adv. Cancer Res. 45.1, 1985.3. Nelson, K.A. , George, E. , Swenson, C. , Forstrom, J.W. , and
Hellstrom, K.E. Immunotherapy of murine sarcomas with auto-anti-idiotypic monoclonal antibodies which bind to tumor specific
Tcells. J. Immunol. 137,2110, 1987.4. Hoon, D.B.S. , Bowker, R.J. , and Cochran, A.J. Suppressor cell
activity in melanoma draining lymph nodes. Cancer Res.47,1740, 1987.
5. Goodwin, J.S. , and Webb, DR. Regulation of the immuneresponse by prostaglandins . Clin . Immunol. Immunopathol.15,106, 1980.
6. Varesio, L. , Giovarelli, M. , Landolfo, S. , and Forni, 0. Sup-pression of proliferative response and lymphokine productionduring the progression of a spontaneous tumor. Cancer Res.
39,4983, 1979.
7. Bonaventure, J. , Nigram, V.N. , and Brailovsky, CA. Modula-tion of the immuno-suppressive effects of splenic macrophages inFischer rats bearing adenocarcinoma I 3762 . Cancer Immunol.Immunother. 20,167, 1985.
8. Allison, AC. Mechanism by which activated macrophages inhibitlymphocyte responses. Immunol. Rev. 40,3, 1985.
9. Nathan, C. , Brukner, L. , Kaplan, 0. , Unkeless, J. , and Cohn, Z.
Role of activated macrophages in antibody-dependent lysis oftumor cells. J. Exp. Med. 152,183, 1980.
10. Grabstein, K.H. , Urdal, DL. . Tushinski, R.J. , Mochizuki,
DY. , Prive, V.L. , Cantrell, MA. , Gillis, S. , and Conlon, P.L.Induction of macrophage tumoricidal activity by granulocyte-macrophage colony stimulating factor. Science 232,506, 1986.
11. Ho, M., and Springer, TA. Mac-2, a novel 32,000 Mr mousemacrophage subpopulation-specific antigen defined by mono-clonal antibodies. J. Immunol. 128,1221, 1982.
12. Ho, M. , and Springer. TA. Preparation and use of monoclonal
antimacrophage antibodies. In Methods in Enzymology, (Sabato,GD. , Langone, J.J. , and Vunakis, H.V. , Eds.) New York: Acad.
Press, p. 313, 1984.
13. Springer, TA. , Galfre, 0. , Secher, D.S. , and Milstein, C.Mac-I : A macrophage differentiation antigen identified by mono-clonal antibody. Eur. J. Immunol. 9,301, 1979.
14. Sigel, MM. , Lopez, D.M. , and Ortiz-Muniz, 0. In vitro immune
responses to viral and tumor antigens in murine breast cancer.Cancer Res 36,748, 1976.
15. Lopez, D.M. , and Paul, RD. Analysis of lymphoid cells respond-ing to virus- and tumor-associated antigens in spleens of syngeneic
mouse mammary tumor systems with low and high oncogenicpotentials. J.N.C.I., 69,1403, 1982.
16. Lopez, D.M. , Padmanabhan, R.R. , Rivera, L.M. , Paul, RD.,
Herbert, L. , Cray, C. , and Watson, GA. Cytotoxic effectormechanisms involved in the immunity against mammary tumors.In Breast Cancer: Scientific and Clinical Progress, (Rich, MA.,
Hager, J.C., and Lopez, D.M. , his.) Boston: Kluwer Acad.Publ., p. 185, 1988.
17. Fu, Y.-X. , Watson, GA. , Jimenez, ii. , Wang, Y. , and Lopez,D.M. Expansion of immunoregulatory macrophages by granulo-
cyte-macrophage colony-stimulating factor derived from a murinemammary tumor. Cancer Res. 50,227, 1990.
18. Garner, RE. , and Elgert, K.D. Changes in splenic macrophages’Mac antigen expression during tumor growth: A kinetic study ofaccessory cell function and antigen-defined phenotypes. J. Leu-kocyte Biol. 40,709, 1986.
19. Medina, D., and DeOme, KB. Response ofhyperplastic alveolar
nodule outgrowth line Dl to mammary tumor virus, nodule-inducing virus, and prolonged hormonal stimulation acting singly
and in combination. J.N.C.I. 42,303, 1969.20. Thompson, SW. , and Hunt, RD. Romanowsky-type staining
techniques: Wright’s stain, Giemsa’s stain, May-Grunwald Gi-emsa’s stain. In Selected Histochemical and HistopathologicalMethods, 2nd Ed. (Thompson, SW. , Ed.) Springfield: Thomas
Co. , p. 757, 1966.21 . Kurnick, J.T. , Ostberg, L. , and Stegagno, M. A rapid method for
the separation of functional lymphoid cell populations of humanand animal origin on PVP-Silica (Percoll) density gradients.
Scand. J. Immunol. 10,563, 1979.22. Thompson, J.T. , Gralow, JR. , Levy, R. , and Miller, R.A. The
optimal application of forward and ninety-degree light scatter in
flow cytometry for the gating of mononuclear cells. Cytometry
6,401, 1985.
23. Edelson, P.J. , and Cohn, Z.A. 5’-nucleotidase activity of mouseperitoneal macrophages. I. Synthesis and degradation in residentand inflammatory populations. J. Exp. Med. 144,1581 , 1976.
24. Edelson, P.J. , and Erbs, C. Plasma membrane localization andmetabolism of alkaline phosphodiesterase I in mouse peritonealmacrophages. J. Exp. Med. 147,77, 1978.
25. Wachsmuth, ED. Aminopeptidase as a marker for macrophage
differentiation. Exp. Cell. Res. 96,409, 1975.26. Hyde, AM. . Stagg. RB. , McEachron, R. and Nuner, R.L.
Effects of the removal of adherent and phagocytic cells on thespleen cell lymphoproliferative response of tumor-bearing mice.Cancer Immunol. Immunother. 20,97, 1985.
27. Parhar, R. . , and Lala, P.K. Prostaglandin E,-mediated inactiva-tion of various killer lineage cells by tumor-bearing host mac-
rophages. J. Leukocyte Biol. 44,474, 1988.28. Malick, A.P. , Elgert, K.D. , Garner, RE. , and Adkinson, N.F.,
Jr. , Prostaglandin E2 production by Mac-2� macrophages: Tu-mor-induced population shift. J. Leukocyte Biol. 42,673, 1987.
29. Garner, RE. , Malick, A.P. , Yorochko, AD. , and Elgert, K.D.Shifts in macrophage (M4) surface phenotypes during tumorgrowth: Association of Mac-V and Mac-3� M4 with immuno-
suppressive activity. Cell. Immunol. 108,255, 1987.

i38 Watson et al.
30. Yurochko, AD. , Pyle, RH. , and Elgert, K.D. Changes in
macrophage populations: Phenotypic differences between normaland tumor-bearing host macrophages. Immunobiology 178,416,1989.
31 . Krakuer, T. A macrophage-derived factor that inhibits the pro-
duction and action of interleukin-2. J. Leukocyte Biol. 38,429,1985.
32. Stallcup, K.C., Liu, Y.N., Dorf, ME., and Mescher, M.F.
Inhibition of lymphoid cell growth by a lipid-like component ofmacrophage hybridoma cells.J. Immunol. 136,2723, 1986.
33. Fujiwara, H., and Ellner, J.J. Spontaneous production of asuppressor factor by the human macrophage-like cell line U937.
I. Suppression of interleukin 1, interleukin 2, and mitogen-induced blastogenesis in mouse thymocytes. J. Immunol.
l36(l),l8l, 1986.
34. Young, MR. , Endicott, R.A. , Duffie, OP. , and Wepsic, H.T.Suppressor alveolar macrophages in mice bearing metastaticLewis lung carcinoma tumors. J. Leukocyte Biol. 42,682, 1987.
35. Tsuchiya, Y. Igarashi, M. , Suzuki, R. , and Kumagai, K.Production of colony-stimulating factor by tumor cells and the
factor-mediated induction of suppressor cells. J. Immunol.141,699, 1988.
36. Heidenreich, S., Gong, J.-H., Schmidt, A., Nain, M., andGemsa, D. Macrophage activation by granulocyte/macrophagecolony-stimulating factor priming for enhanced release of tumornecrosis factor--y and prostaglandin E,. J. Immunol. 143,1198,
1989.
Copyright © 2022 FDOKUMEN










![NK Cells Mediate Increase of Phagocytic Activity but Not of Proinflammatory Cytokine (Interleukin6 [IL6], Tumor Necrosis Factor Alpha, and IL12) Production Elicited in Splenic Macrophages](https://static.fdokumen.com/doc/165x107/632147240c12e1161503b7d3/nk-cells-mediate-increase-of-phagocytic-activity-but-not-of-proinflammatory-cytokine.jpg)




![7H-Dibenzo[ c, g]carbazole and 5,9-dimethyldibenzo[ c, g]carbazole exert multiple toxic events contributing to tumor promotion in rat liver epithelial ‘stem-like’ cells](https://static.fdokumen.com/doc/165x107/63163ec03ed465f0570bf28c/7h-dibenzo-c-gcarbazole-and-59-dimethyldibenzo-c-gcarbazole-exert-multiple.jpg)




