
Splenic marginal zone antigen-presenting cells are critical for the primary allo-immune response to...
8
ORIGINAL ARTICLE Splenic marginal zone antigen-presenting cells are critical for the primary allo-immune response to therapeutic factor VIII in hemophilia A A. NAVARRETE,* à S. DASGUPTA,* à S. DELIGNAT,* à G. CALIGIURI,* à O. D. CHRISTOPHE,§ J. BAYRY,* à A. NICOLETTI,* à S. V. KAVERI* à and S. LACROIX-DESMAZES* à *Centre de Recherche des Cordeliers, Universite ´ Pierre et Marie Curie-Paris6, UMRS 872, Paris; Universite ´ Paris Descartes, UMRS872, Paris; àINSERM, U872, Paris; and §INSERM U770, Universite ´ Paris-Sud, Faculte ´ de me ´decine Paris-Sud, IFR93, Le Kremlin-Bice ˆtre, France To cite this article: Navarrete A, Dasgupta S, Delignat S, Caligiuri G, Christophe OD, Bayry J, Nicoletti A, Kaveri SV, Lacroix-Desmazes S. Splenic marginal zone antigen-presenting cells are critical for the primary allo-immune response to therapeutic factor VIII in hemophilia A. J Thromb Haemost 2009; 7: 1816–23. Summary. Background: Alloimmune responses to intrave- nously administered protein therapeutics are the most common cause of failure of replacement therapy in patients with defective levels of endogenous proteins. Such a situation is encountered in some patients with hemophilia A, who develop inhibitory anti- factor (F)VIII alloantibodies after administration of FVIII to treat hemorrhages. Objectives: The nature of the secondary lymphoid organs involved in the initiation of immune responses to human therapeutic has not been studied. We therefore investigated this in the case of FVIII, a self-derived exogenous protein therapeutic. Methods: The distribution of intravenously administered FVIII was followed after FVIII-deficient mice were injected with radiolabeled FVIII and using immunohisto- chemistry. The role of the spleen and antigen-presenting cells (APC) in the onset of the anti-FVIII immune response was analyzed upon splenectomy or treatment of the mice with APC- depleting compounds. Results: FVIII preferentially accumu- lated in the spleen at the level of metallophilic macrophages in the marginal zone (MZ). Surgical removal of the spleen or selective in vivo depletion of macrophages and CD11c-positive CD8a-negative dendritic cells resulted in a drastic reduction in anti-FVIII immune responses. Conclusions: Using FVIII- deficient mice as a model for patients with hemophilia A, and human pro-coagulant FVIII as a model for immunogenic self- derived protein therapeutics, our results highlight the impor- tance of the spleen and MZ APCs in the initiation of immune responses to protein therapeutics. Identification of the receptors implicated in retention of protein therapeutics in the MZ may pave the way towards novel strategies aimed at reducing their immunogenicity. Keywords: allo-immunization, factor VIII, FVIII inhibitors, hemophilia A, protein therapeutics, spleen. Introduction Replacement therapy is the treatment of choice for patients with endogenous deficiencies of self-proteins. In several instances, patients develop antibodies that inhibit the admin- istered therapeutics [1]. Such a situation is encountered in patients with hemophilia A, a X-linked bleeding disorder consecutive to the lack of functional endogenous factor (F)VIII: in 10%–30% of the patients, administration of FVIII to treat hemorrhages results in induction of inhibitory anti- FVIII IgG [2]. The development of allo-immune responses to therapeutic FVIII suggests that exogenous FVIII is not solely attracted to bleeding sites and trapped by catabolic organs, but also reaches lymphoid organs where it is processed by antigen- presenting cells (APCs) and presented to immune effectors. The spleen plays a primary role in filtration of blood by removing particulate components such as immune complexes, bacteria or colloidal particles, and supports the development of immunity, especially towards blood-borne antigens [3]. The spleen contains three major types of professional APCs: macrophages, B lymphocytes and dendritic cells (DCs) [4]. Blood-borne antigens reach the spleen through the splenic artery which branches either towards the red pulp, and interacts with red pulp macrophages, or towards the marginal zone (MZ) of the spleen where B cells, metallophilic macrophages and MZ macrophages reside [3]. Initiation of the immune responses requires the antigens to reach the white pulp of the spleen [5]. Depending on their size, solubility and concentra- tion, blood-borne molecules diffuse directly to the white pulp from the MZ [6] or are delivered to the follicles by APCs [7]. Correspondence: Se´bastien Lacroix-Desmazes, INSERM UMR 872 Equipe 16, Centre de Recherche des Cordeliers, Paris F-75006, France. Tel.:+33 1 55 42 82 65; fax: +33 1 55 42 82 62. E-mail: [email protected] AMN, SuDa and SaDe contributed equally to the work. Received 4 June 2009, accepted 13 July 2009 Journal of Thrombosis and Haemostasis, 7: 1816–1823 DOI: 10.1111/j.1538-7836.2009.03571.x Ó 2009 International Society on Thrombosis and Haemostasis
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Splenic marginal zone antigen-presenting cells are critical for the primary allo-immune response to...

ORIGINAL ARTICLE
Splenic marginal zone antigen-presenting cells are critical forthe primary allo-immune response to therapeutic factor VIII inhemophilia A
A. NAVARRET E ,*�� S . DASGUPTA ,*�� S . DEL IGNAT ,*�� G. C AL I G I UR I ,*�� O. D . CHR ISTOPHE ,§
J . BAYR Y ,*�� A . N I COLETT I , *�� S . V . KA VER I *�� and S . LACROIX-DESMAZE S*��*Centre de Recherche des Cordeliers, Universite Pierre et Marie Curie-Paris6, UMRS 872, Paris; �Universite Paris Descartes, UMRS872, Paris;
�INSERM, U872, Paris; and §INSERM U770, Universite Paris-Sud, Faculte de medecine Paris-Sud, IFR93, Le Kremlin-Bicetre, France
To cite this article: Navarrete A, Dasgupta S, Delignat S, Caligiuri G, Christophe OD, Bayry J, Nicoletti A, Kaveri SV, Lacroix-Desmazes S. Splenic
marginal zone antigen-presenting cells are critical for the primary allo-immune response to therapeutic factor VIII in hemophilia A. J Thromb
Haemost 2009; 7: 1816–23.
Summary. Background: Alloimmune responses to intrave-
nously administered protein therapeutics are the most common
cause of failure of replacement therapy in patientswith defective
levels of endogenous proteins. Sucha situation is encountered in
some patients with hemophilia A, who develop inhibitory anti-
factor (F)VIII alloantibodies after administration of FVIII to
treat hemorrhages. Objectives: The nature of the secondary
lymphoid organs involved in the initiation of immune responses
to human therapeutic has not been studied. We therefore
investigated this in the case of FVIII, a self-derived exogenous
protein therapeutic.Methods:The distribution of intravenously
administered FVIII was followed after FVIII-deficient mice
were injected with radiolabeled FVIII and using immunohisto-
chemistry. The role of the spleen and antigen-presenting cells
(APC) in the onset of the anti-FVIII immune response was
analyzed upon splenectomy or treatment of themicewithAPC-
depleting compounds. Results: FVIII preferentially accumu-
lated in the spleen at the level of metallophilic macrophages in
the marginal zone (MZ). Surgical removal of the spleen or
selective in vivo depletion of macrophages and CD11c-positive
CD8a-negative dendritic cells resulted in a drastic reduction in
anti-FVIII immune responses. Conclusions: Using FVIII-
deficient mice as a model for patients with hemophilia A, and
human pro-coagulant FVIII as a model for immunogenic self-
derived protein therapeutics, our results highlight the impor-
tance of the spleen and MZ APCs in the initiation of immune
responses to protein therapeutics. Identification of the receptors
implicated in retention of protein therapeutics in the MZ may
pave the way towards novel strategies aimed at reducing their
immunogenicity.
Keywords: allo-immunization, factor VIII, FVIII inhibitors,
hemophilia A, protein therapeutics, spleen.
Introduction
Replacement therapy is the treatment of choice for patients
with endogenous deficiencies of self-proteins. In several
instances, patients develop antibodies that inhibit the admin-
istered therapeutics [1]. Such a situation is encountered in
patients with hemophilia A, a X-linked bleeding disorder
consecutive to the lack of functional endogenous factor
(F)VIII: in 10%–30% of the patients, administration of FVIII
to treat hemorrhages results in induction of inhibitory anti-
FVIII IgG [2]. The development of allo-immune responses to
therapeutic FVIII suggests that exogenous FVIII is not solely
attracted to bleeding sites and trapped by catabolic organs, but
also reaches lymphoid organs where it is processed by antigen-
presenting cells (APCs) and presented to immune effectors.
The spleen plays a primary role in filtration of blood by
removing particulate components such as immune complexes,
bacteria or colloidal particles, and supports the development of
immunity, especially towards blood-borne antigens [3]. The
spleen contains three major types of professional APCs:
macrophages, B lymphocytes and dendritic cells (DCs) [4].
Blood-borne antigens reach the spleen through the splenic
artery which branches either towards the red pulp, and interacts
with red pulp macrophages, or towards the marginal zone
(MZ) of the spleen where B cells, metallophilic macrophages
and MZ macrophages reside [3]. Initiation of the immune
responses requires the antigens to reach the white pulp of the
spleen [5]. Depending on their size, solubility and concentra-
tion, blood-borne molecules diffuse directly to the white pulp
from the MZ [6] or are delivered to the follicles by APCs [7].
Correspondence: Sebastien Lacroix-Desmazes, INSERM UMR 872
Equipe 16, Centre de Recherche des Cordeliers, Paris F-75006, France.
Tel.:+33 1 55 42 82 65; fax: +33 1 55 42 82 62.
E-mail: [email protected]
AMN, SuDa and SaDe contributed equally to the work.
Received 4 June 2009, accepted 13 July 2009
Journal of Thrombosis and Haemostasis, 7: 1816–1823 DOI: 10.1111/j.1538-7836.2009.03571.x
� 2009 International Society on Thrombosis and Haemostasis

The localization and identification of the type of APCs
implicated in immune responses have been studied in the case
of exogenous model antigens [8], often as a prerequisite to
therapeutic targeting of defined APCs in vaccination strate-
gies [9]. To our knowledge, the identification of the lymphoid
organs and APCs implicated in immune responses to blood-
borne therapeutic self-proteins, a prerequisite to the selective
prevention of deleterious iatrogenic allo-immune responses,
has not been addressed. In the present study, we hypothe-
sized that intravenously administered therapeutic FVIII
accumulates in the spleen. Using FVIII-deficient mice, a
mouse model of hemophilia A, we demonstrate the relevance
of the spleen and of splenic phagocytes for the onset of the
naıve anti-FVIII allo-immune response. We further seek to
identify the populations of endocyting cells implicated in the
capture of FVIII.
Materials and methods
Animals
Mice used in the experiments were 7- to 12-week-old 129/B6
(H-2Db background) exon 16 FVIII-deficientmales (a kind gift
from Professor H.H. Kazazian, Department of Genetics,
University of Pennsylvania School of Medicine, Philadelphia,
USA). Animals were handled in agreement with local ethical
authorities (Comite regional d�ethique p3/2005/002).
Coupling of iodine125 to FVIII
Coupling of FVIII to iodine125 (I125) was done by the IODO-
GEN� (Pierce Chemical Co, Rockford, IL, USA) method.
Briefly, recombinant human FVIII (Kogenate�; Bayer
Healthcare, Lille, France; or Helixate�; CSL-Behring, Paris,
France; Kogenate� and Helixate� are the same molecule) was
solubilized in water and dialyzed against 100 mM Borate
buffer for 4 h. Dialyzed FVIII was incubated at room
temperature for 6 min with iodine125 (Amersham GE Health-
care, Buckinghamshire, UK) at a ratio of 10 lCi lg)1 of
FVIII. Radio-labeled FVIII was eluted using a PD-10 column
(Amersham GE Healthcare). FVIII-containing fractions were
eluted and passed on a gamma-counter. The specific activity
of coupling was 1065 cpm fmol)1 of FVIII. Comparison of
the binding of unlabeled and radiolabeled FVIII to von
Willebrand factor (VWF) and to a monoclonal anti-C2 IgG
(Bo2C11, a kind gift from Professor JM Saint-Remy, Leuven,
Belgium) in radio-immuno assays, indicated that labeled
FVIII retained 65%–75% of its capacity to bind VWF.
Organ distribution of I125-FVIII
FVIII-deficient mice were injected intravenously with I125-
FVIII (1 lg per mouse in 200 lL PBS). Mice were sacrificed
at different time points after injection. Organs were collected,
weighed and incorporated radioactivity was measured with a
gamma-counter. Five mice per time point were utilized.
Kinetics of I125-FVIII clearance were computed using a
double-exponential model: Q = Q1e�k1t þQ2e
�k2t, where Q
represents the quantity of FVIII (cpm lg)1 tissue) at a given
time point, k1 and k2 are the rate constants corresponding to
fast and slow phases of FVIII elimination in min)1, and Q1
and Q2 are quantities of FVIII removed during the fast and
slow phases of clearance, respectively. In VWF-deficient mice,
FVIII is eliminated in < 10 min. The 35%–25% of iodinated
FVIII molecules that do not bind to VWF will thus be
eliminated very rapidly after intravenous injection, and,
hence, will not bias our experimental results on FVIII half-
life.
Administration of FVIII to splenectomized mice
Anesthetized mice were splenectomized or sham-operated. One
day after surgery, mice were injected intravenously or subcu-
taneously with FVIII. In the case of intravenous injections,
recombinant human FVIII was administered (0.2 lg per
mouse) once a week for 4 weeks. For subcutaneous injection,
FVIII (20 lg per mouse) in the presence of Freund�s adjuvantwas injected three times at 1-week intervals. Blood samples
were collected 1 week after the last injection. Serumwas kept at
)20 �C until use.
Titration of anti-FVIII IgG and of inhibitors was performed
by ELISA and chromogenic assay respectively, as described
[10].
Administration of FVIII to clodronate-containing liposomes-
treated mice
Clodronate was a gift of Roche Diagnostics GmbH, Mann-
heim, Germany. It was encapsulated in liposomes as described
earlier [11]. FVIII-deficient mice were administered intrave-
nously with clodronate- or phosphate-buffered saline (PBS)-
containing liposomes (200 lL per mouse) twice with a weekly
interval. FVIII (0.2 lg per mouse) was injected intravenously
48 h after the first and second administrations of liposomes,
and two additional times at weekly intervals. Under such
conditions, clodronate-containing liposomes eliminate phago-
cytes [11]. Repopulation of macrophages starts 9 days after
treatment in the red pulp and 15 days in the MZ [12]. Blood
samples were collected 1 week after the last injection of FVIII.
Serum was kept at )20 �C until use. In some cases, mice were
sacrificed 48 h after the first administration of liposomes.
Spleens were collected and analyzed by immunohistochemistry
and flow cytometry to assess the extent of cell depletion.
Analysis of splenocytes by multi-color flow cytometry
Mice were sacrificed and spleens collected. Spleens were
digested with a solution containing 0.42 lg mL)1 Liberase
and 0.1 lg mL)1 DNAse (Roche, Indianapolis, IN, USA).
Thirty minutes later, single cell suspensions were prepared.
Splenocytes were washed, counted and incubated with anti-
mouse CD11c-PE, anti-mouse CD8a-pacific blue, anti-mouse
Allo-immunization to therapeutic FVIII 1817
� 2009 International Society on Thrombosis and Haemostasis

B220 PE-TexRed (Pharmingen, BD bioscience, CA, USA)
and anti-mouse F4/80-APC (EBioscience, San Diego, CA,
USA) monoclonal rat antibodies, and analyzed using a
LSRII flow cytometer (BD bioscience). Dead cells were gated
using positive annexin V-FITC (Immunotech, Marseille,
France) staining and excluded from the analysis. More than
400 000 events were acquired in the annexin V-FITC negative
gate. Analysis used the FACSDIVA software v5.0.1 (BD
bioscience).
Analysis of splenocytes by immuno-histochemistry
Mice were sacrificed and spleens snap-frozen in liquid nitrogen.
Spleen sections (10 lm) were fixed in acetone at 4 �C for
10 min. Endogenous peroxidase and avidin-biotin activity were
eliminated (Dako Cytomation, Glostrup, Denmark). Metallo-
philic macrophages and macrophage-associated receptor with
collagenous structure (MARCO)-positive MZ macrophages
were identified with monoclonal rat anti-sialic acid-binding
lectin-1 (MOMA-1) and anti-MARCO antibodies (Abd Sero-
tec, Oxford, UK), followed by a rabbit anti-rat biotinylated
antibody (Dako). DCs were detected with a monoclonal
hamster anti-CD11c antibody (Pharmingen, BD Biosciences,
CA, USA), followed by a goat anti-hamster biotinylated
antibody. Red pulp macrophages were labeled with a mono-
clonal rat anti-F4/80 biotinylated antibody (eBiosciences) and
revealed with streptavidine-peroxydase from the ABC kit
(DAKO) and DAB substrate (Vector laboratories, CA, USA).
Analysis of FVIII distribution in the spleen by
immuno-fluorescence
Mice were injected with 3 lg of human recombinant FVIII.
Thirty minutes post-injection, the mice were sacrificed and
spleens collected and snap-frozen in liquid nitrogen. Serial
cryosections (10 lm)were air-dried and fixed in acetone. FVIII
was detected with a sheep polyclonal anti-human FVIII IgG
(Kordia, Leiden, the Netherlands) followed by a donkey anti-
sheep IgG conjugated to FITC (Abcam, Cambridge, UK) or to
Alexa Fluor 647 (Invitrogen, Orlando, FL, USA). Metallo-
philic macrophages, MZ macrophages and red pulp macro-
phages were stained using monoclonal rat anti-MOMA-1
FITC, rat anti-MARCO FITC (AbD Serotec, Oxford, UK)
and biotinylated anti-F4/80 (eBiosciences) antibodies followed
by Avidin-Alexa Fluor 555 (Invitrogen), respectively. DCs
were identified with a monoclonal hamster anti-CD11c,
followed by biotinylated goat anti-hamster polyclonal antibody
and Avidin-Alexa Fluor 555. Nuclei were counterstained with
0.1 lg mL)1 Hoechst 33342 and tissue sections were mounted
with ProLong� Gold antifade reagent (Invitrogen). Images
were acquired using an Axiovert� M200 microscope (Zeiss)
equipped with Apoptome� and four filters (Dapi, FITC,
Rhodamine, APC) connected to a monochromatique CCD
camera. Digital images were captured with AxioVision�
software and analyzed with Adobe Photoshop V.CS3 (Adobe,
Dublin, Ireland).
Results
FVIII accumulates in the spleen of FVIII-deficient mice after
intravenous administration
We investigated the distribution of intravenously administered
FVIII in the liver, spleen, blood, kidneys, lymph nodes and
lungs of FVIII-deficient mice after 15, 30, 60, 120, 240 and
480 min. FVIII declined from the circulation whereas it
accumulated in the spleen and liver (Fig. 1). FVIII density in
lungs, kidneys and lymph nodes was marginal. FVIII clearance
from blood, liver and spleen required a double exponential
model to fit the data (Table 1), with a fast phase and a slow
phase of FVIII elimination.
During the first 30 min after injection, the FVIII concentra-
tiondecreasedby7.6% ± 11.0%inthecaseofthespleenandby
11.0% ± 8.8% in the case of blood. It dropped by
48.2% ± 12.0% in the case of the liver (P < 0.01 as compared
with the spleen and blood using Mann–Whitney U-test).
Accordingly, the rate constant for the fast phase of FVIII
elimination tended to be significantly higher in the case of the
liver (0.078 ± 0.037 min)1) than the spleen (0.025 ± 0.025
min)1, P = 0.076). Together, the data indicate a longer
residence time and a higher density of FVIII in the spleen as
compared with other organs, including liver and lymph nodes.
The spleen is essential for mounting an allo-immune response
to therapeutic FVIII
In order to determine the importance of the spleen in a specific
immune response to therapeutic FVIII, FVIII-deficient mice
1600
Lymph nodesBloodLiverLungSpleenKidney
1200
800
400
010 100 1000
Time (min)
FV
III (
cpm
per
mg
tissu
e)
Fig. 1. Tissue distribution of factor (F)VIII after intravenous adminis-
tration. I125-FVIII (1 lg) was injected intravenously to FVIII-deficient
mice. At the indicated time points, mice were sacrificed. Lymph nodes (full
circles), blood (empty circles), liver (empty squares), lung (X), spleen (full
squares) and kidney (+) were collected, weighed and the incorporated
radioactivity was measured. Results depict the density of FVIII (cpm per
lg of tissue). Data represent mean values from five mice at each time point
and are representative of two independent experiments.
1818 A. Navarrete et al
� 2009 International Society on Thrombosis and Haemostasis

were splenectomized and injected with FVIII. After the fourth
injection of FVIII, the anti-FVIII IgG titer was 4.4-fold lower
in splenectomized mice (81.3 ± 46.8 lg mL)1 mAb6-equiva-
lent) as compared with sham-operated animals (359.9 ±
204.4 lg mL)1 mAb6-equivalent, P < 0.05, Fig. 2A). Fur-
ther, the inhibitory activity in the serum of sham-operated mice
(494 ± 304 BU mL)1) was 4.3-fold higher than in the serum
of splenectomized animals (115 ± 104 BU mL)1, P < 0.05)
(Fig. 2B). Interestingly, total IgG titers were similar in both
groups of mice (Fig. 2C), indicating that removal of the spleen
does not alter the levels of circulating IgG over a period of
4 weeks after surgery. Furthermore, sham-operated and sple-
nectomized mice developed similar titers of anti-FVIII IgG
after subcutaneous immunization with FVIII in Freund�sadjuvant (data not shown), indicating that removal of the
spleen does not hamper the capacity of the mice to mount a
specific immune response to an antigen that is targeted to the
lymph nodes. Together, these results demonstrate that the
spleen is the major secondary lymphoid organ for the
development of an anti-FVIII immune response.
Accumulation of therapeutic FVIII at the level of macrophages
in the spleen
In order to identify the cells co-localizing with FVIII in the
spleen, we injected intravenously near to therapeutic doses of
exogenous FVIII (3 lg) to naıve FVIII-deficient mice. Fig-
ure 3A documents the accumulation of FVIII in the splenic
MZ after intravenous injection of FVIII (3 lg). Immunohis-
tochemistry experiments demonstrated preferential FVIII co-
localization with metallophilic macrophages (MOMA-1-posi-
tive, Fig. 3B) and, to a lesser extent, MARCO-positive MZ
macrophages (Fig. 3C). Occasionally FVIII was co-localized
with CD11c-positive DCs in theMZ (Fig. 3E). FVIII was only
sporadically detected with red pulp F4/80-positive macrophag-
es (Fig. 3D). FVIII was not detected at the level of B
lymphocytes in naıve mice (data not shown).
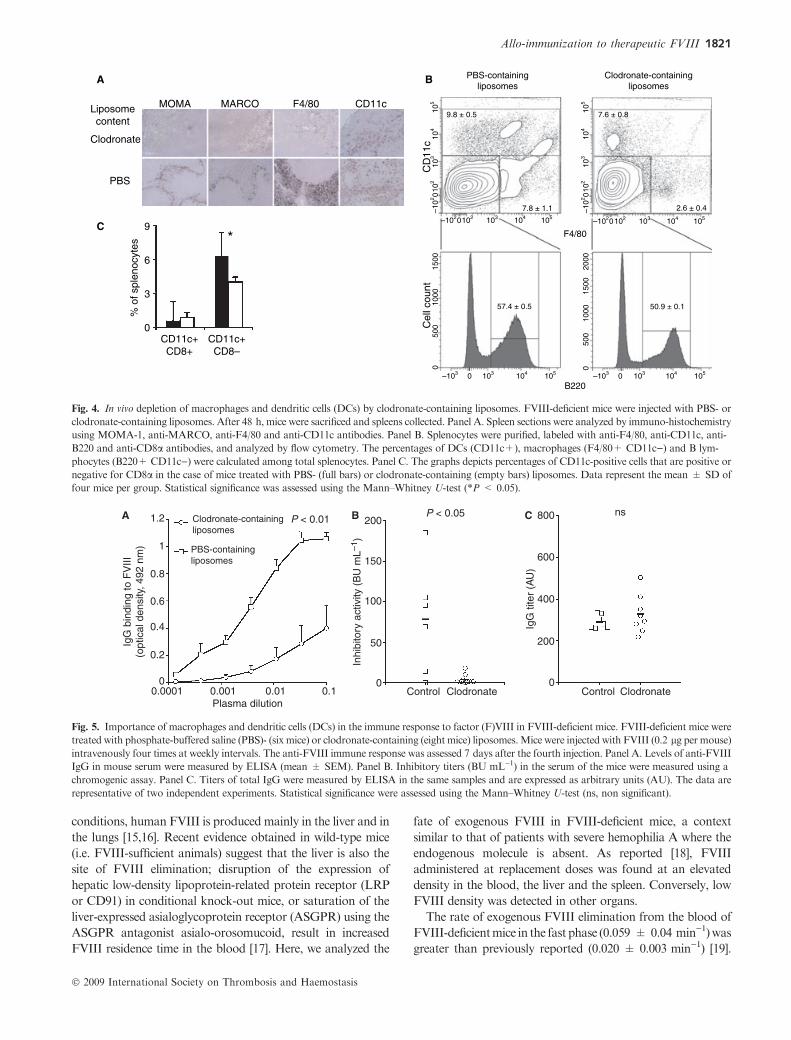
Depletion of macrophages hampers the development of the
anti-FVIII immune response
To determine the importance of macrophages in an anti-FVIII
immune response, mice were injected with clodronate-contain-
ing liposomes. Immuno-histochemistry confirmed depletion of
bothF4/80-positive redpulpmacrophagesandMZ(MOMA-1-
andMARCO-positive)macrophages from clodronate-contain-
ing liposome-treated mice (Fig. 4A). Numbers of red pulp F4/
80-positive macrophages and CD11c-positive DCs were
reduced by 3.1 (from 7.8% ± 1.1% to 2.6% ± 0.4% of the
total splenocyte population) and 1.3-fold (from 9.8% ± 0.5%
to 7.6% ± 0.8%) as compared with control mice, respectively
(P < 0.05, Fig. 4B). Changes in B lymphocytes numbers were
marginal (from 57.4% ± 0.5% to 50.9% ± 0.1%).
Table 1 Clearance of 125I-FVIII from the blood, liver and spleen of FVIII-
deficient mice
Organ
Q1 Q2 k1 k2
cpm lg)1 min)1
Blood 0.48 ± 0.25*� 0.55 ± 0.08* 0.059 ± 0.049 0.005 ± 0.001
Liver 2.87 ± 1.92* 0.36 ± 0.12* 0.078 ± 0.037� 0.006 ± 0.002
Spleen 2.16 ± 1.10� 0.81 ± 0.48 0.025 ± 0.025� 0.006 ± 0.005
The values (mean ± SD) of the kinetic rate constants k1 and k2, that
correspond to the fast and slow phases of FVIII clearance, and the
amount of FVIII removed during these two phases (Q1 and Q2,
respectively, expressed as cpm lg)1 of tissue) were determined by fit-
ting the clearance data depicted in Fig. 1 to the double exponential
decay curve: Q = Q1e�k1t þQ2e
�k2t, where t is the time in minutes.
Differences were statistically compared using the Mann–Whitney U-
test (*, blood vs. liver; �, blood vs. spleen; �, liver vs. spleen).
Underscored letters indicate tendency towards significance (P < 0.1),
otherwise significance (P < 0.05).
P < 0.05 P < 0.051000
800
600
400
Inhi
bito
ry a
ctiv
ity (
BU
mL–
1 )
200
0
SplnxSham
SplnxSham
1.6A B C
1.2
0.8
0.4
00.0001 0.001 0.01 0.1
Plasma dilution
IgG
bin
ding
to F
VIII
(opt
ical
den
sity
, 492
nm
)
ns800
600
400
Tota
l IgG
(A
U)
200
0SplnxSham
Fig. 2. Importance of the spleen in the immune response to factor (F)VIII in FVIII-deficient mice. FVIII-deficient mice were splenectomized (Splnx, eight
mice) or sham operated (five mice) and injected with FVIII (0.2 lg per mouse) intravenously four times at weekly intervals. Panel A. Levels of anti-FVIII
IgG in the serum of the mice weremeasured by (mean ± SEM). Panel B. Inhibitory titers (BUmL)1) in mouse serumwere measured using a chromogenic
assay. Panel C. Titers of total IgG were measured by ELISA in the same samples and are expressed as arbitrary units (AU). The data are representative of
two independent experiments. Statistical significance was assessed using the Mann–Whitney U-test (ns, non significant).
Allo-immunization to therapeutic FVIII 1819
� 2009 International Society on Thrombosis and Haemostasis

T lymphocytes numbers were not affected by clodronate-
containing liposome treatment (data not shown). Among
CD11c-positive DCs, CD8a-negative cells were significantly
reduced in numbers (from 6.3% ± 2.1% to 4.0% ± 0.4% of
the total splenocyte population, P < 0.05, Fig. 4C).
Clodronate-containing liposome-treated mice also demon-
strated significantly reduced titers of anti-FVIII IgG
(28.8 ± 20.0 vs. 573.7 ± 154.5 lg mL)1 mAb6-equivalent,
P < 0.01) and of inhibitory activity towards FVIII, as
compared with control mice (Fig. 5A and B). Clodronate-
containing liposomes treatment had no effect on the total levels
of circulating IgG over a period of 4 weeks after the initial
liposome treatment (Fig. 5C).
Discussion
Pharmacokinetics of FVIII have been studied in different
animal models and in human [13,14]. Under physiological
A
B
C
D
E
20 µm RP
RP
RP
RP
RP
RP
MZ
MZ
MZ
MZ
WP
WP
WP
WP
WP
Fig. 3. In situ localization of factor (F)VIII in the spleen of FVIII-deficient mice. FVIII-deficient mice were injected with FVIII or phosphate-buffered
saline (PBS), and sacrificed 30 min later. Panel A. FVIII (green) was detected on histological spleen sections in the case of FVIII- (left panel) and
PBS-treated mice (right panel). Panels B, C, D and E. Histological sections of spleens of FVIII-treated mice were labeled with anti-FVIII antibodies
(Green), and with MOMA-1, anti-MARCO, anti-F4/80 or anti-CD11c (panels B, C, D and E, respectively) antibodies (Red). Left panels show FVIII
staining alone, middle panels showMOMA-1, MARCO, F4/80 or CD11c staining alone and right panels showmerged images. Data are representative of
3–4 mice in each group. MZ, marginal zone, WP, white pulp, RP, red pulp, as identified using MOMA-1 and anti-F4/80 antibodies. Magnification 40·.
1820 A. Navarrete et al
� 2009 International Society on Thrombosis and Haemostasis
conditions, human FVIII is produced mainly in the liver and in
the lungs [15,16]. Recent evidence obtained in wild-type mice
(i.e. FVIII-sufficient animals) suggest that the liver is also the
site of FVIII elimination; disruption of the expression of
hepatic low-density lipoprotein-related protein receptor (LRP
or CD91) in conditional knock-out mice, or saturation of the
liver-expressed asialoglycoprotein receptor (ASGPR) using the
ASGPR antagonist asialo-orosomucoid, result in increased
FVIII residence time in the blood [17]. Here, we analyzed the
fate of exogenous FVIII in FVIII-deficient mice, a context
similar to that of patients with severe hemophilia A where the
endogenous molecule is absent. As reported [18], FVIII
administered at replacement doses was found at an elevated
density in the blood, the liver and the spleen. Conversely, low
FVIII density was detected in other organs.
The rate of exogenous FVIII elimination from the blood of
FVIII-deficientmice in the fast phase (0.059 ± 0.04 min)1)was
greater than previously reported (0.020 ± 0.003 min)1) [19].
Liposomecontent
Clodronate
PBS
Cel
l cou
ntC
D11
c10
510
410
310
2–1
0215
0010
0050
00
2000
1500
1000
500
0
0
105
104
103
102
–102
0
105
7.8 ± 1.1
F4/80
PBS-containingliposomes
Clodronate-containingliposomes
B220
9.8 ± 0.5 7.6 ± 0.8
2.6 ± 0.4
50.9 ± 0.157.4 ± 0.5
104103102–1020
105104103–103 0 105104103–103 0
105104103102–1020
MOMA
A
C
B
MARCO F4/80 CD11c
9*
6
3
0
% o
f spl
enoc
ytes
CD11c+CD8+
CD11c+CD8–
Fig. 4. In vivo depletion of macrophages and dendritic cells (DCs) by clodronate-containing liposomes. FVIII-deficient mice were injected with PBS- or
clodronate-containing liposomes. After 48 h, mice were sacrificed and spleens collected. Panel A. Spleen sections were analyzed by immuno-histochemistry
using MOMA-1, anti-MARCO, anti-F4/80 and anti-CD11c antibodies. Panel B. Splenocytes were purified, labeled with anti-F4/80, anti-CD11c, anti-
B220 and anti-CD8a antibodies, and analyzed by flow cytometry. The percentages of DCs (CD11c+), macrophages (F4/80+ CD11c)) and B lym-
phocytes (B220+ CD11c)) were calculated among total splenocytes. Panel C. The graphs depicts percentages of CD11c-positive cells that are positive or
negative for CD8a in the case of mice treated with PBS- (full bars) or clodronate-containing (empty bars) liposomes. Data represent the mean ± SD of
four mice per group. Statistical significance was assessed using the Mann–Whitney U-test (*P < 0.05).
1.2
1
0.8
0.6
0.4
0.2
00.0001 0.001 0.01 0.1
Plasma dilution
P < 0.05P < 0.01
ns800
600
400
IgG
tite
r (A
U)
200
0
200
150
100
Inhi
bito
ry a
ctiv
ity (
BU
mL–
1 )
50
0Control Clodronate Control Clodronate
Clodronate-containingliposomes
PBS-containingliposomes
IgG
bin
ding
to F
VIII
(opt
ical
den
sity
, 492
nm
)
A B C
Fig. 5. Importance of macrophages and dendritic cells (DCs) in the immune response to factor (F)VIII in FVIII-deficient mice. FVIII-deficient mice were
treated with phosphate-buffered saline (PBS)- (six mice) or clodronate-containing (eight mice) liposomes.Mice were injected with FVIII (0.2 lg per mouse)
intravenously four times at weekly intervals. The anti-FVIII immune response was assessed 7 days after the fourth injection. Panel A. Levels of anti-FVIII
IgG in mouse serum were measured by ELISA (mean ± SEM). Panel B. Inhibitory titers (BU mL)1) in the serum of the mice were measured using a
chromogenic assay. Panel C. Titers of total IgG were measured by ELISA in the same samples and are expressed as arbitrary units (AU). The data are
representative of two independent experiments. Statistical significance were assessed using the Mann–Whitney U-test (ns, non significant).
Allo-immunization to therapeutic FVIII 1821
� 2009 International Society on Thrombosis and Haemostasis

Similar rates were observed in wild-type mice under the same
experimental conditions (data not shown). Both the liver and
spleen demonstrated accumulation of exogenous FVIII early
after injection, and a biphasic elimination pattern of FVIII. The
two organs displayed different behaviors as far as FVIII
elimination is concerned.While the amount of FVIII in the liver
rapidly decreased during the first 30 min after administration, it
remained stable in the case of the spleen. The data may reflect
the different capacities of the two organs to catabolize FVIII, as
well as their differences in size. Thus, during the first 30 min
after intravenous administration, the catabolism of FVIII in the
spleen is balanced by its constant influx from the blood which
maintains a constant level of FVIII.
Blood-borne antigens have been shown to reach theMZ and
red pulp of the spleen and to be captured by resident APCs
[6,20]. However, such studies employedwild-type animals using
model antigens such as albumin. In contrast, here we have used
human pro-coagulant FVIII as a model for immunogenic self-
derived protein therapeutics administered to FVIII-deficient
mice, a suitable model for patients with the X-linked hemor-
rhagic disorder hemophilia A. In addition, the amount of
FVIII injected in our model was close to therapeutic doses, i.e.
‡ 30-fold lower than that used in the case of albumin.
During the first 30 min after intravenous administration,
accumulation of FVIII in the spleen may be of importance to
its immunogenicity. Indeed, the spleen screens the blood and
provides the immunological environment for initiation of
adaptive immune responses against blood-borne antigens [3].
Using FVIII-deficient mice, we show that the onset of the anti-
FVIII immune response occurs in the spleen. Removal of the
spleen resulted in a significant reduction in the amplitude of the
anti-FVIII humoral response. Our results are in line with
previous observations wherein disruption of splenic germinal
centers by intravenous injection of anti-CD154 antibodies was
followed by a reduction in anti-FVIII antibody titers and
abolition of T-cell responses to FVIII [21]. Interestingly,
development of a detectable anti-FVIII immune response to
therapeutic FVIII was observed in splenectomized animals,
indicating that alternative secondary organs, the lymph nodes
or possibly the bone marrow, may be involved in the immune
response to blood-borne protein therapeutics as well [22].
Whether such secondary lymphoid organs are also at play
when the spleen is present, remains to be validated.
Our data suggest a role for residentmacrophages andDCs in
trappingFVIII in themarginal zone:FVIII clustered at the level
of MZ macrophages, preferentially MOMA-1-positive cells
and, toa lesser extent,withMARCO-positivemacrophages and
CD11c-positive DCs in the marginal zone. Localization of
exogenously administered proteins at the level of MZ macro-
phages hasbeendocumented in the caseof the administrationof
elevated amounts of ovalbumin (OVA) [6]. In contrast, intra-
venously administered VWFwas recently found to accumulate
at the level of red pulp macrophages and to be absent from the
MZ [18].
MZ macrophages play a role in both T-dependent and
T-independent immune responses [12,23]. Here, depletion of
macrophages from the spleen of naıve FVIII-deficient mice
with clodronate-containing liposomes, prior to administration
of therapeutic doses of FVIII, resulted in close to complete
abrogation of the humoral anti-FVIII immune response.
Elimination of MZ macrophages by clodronate-containing
liposomes may have diverse side effects. Indeed, splenic MZ
macrophages have important alternative roles in the dynamic
equilibrium of the immune system. They secrete cytokines and
chemokines that determine the fate and intensity of immune
responses [24]. Further, they have been proposed to transport
and deliver antigens to immune effectors in the follicles [25].
Treatment of mice with clodronate-containing liposomes
also reduced the CD11c+CD8 a-negative DC population, as
described previously [26]. Targeting of CD8a-negative DCs
using ovalbumin-coupled 33D1-specific antibodies drives anti-
gen presentation through the MHC II pathway [8]. Our
observations of a drastically reduced immune response to
FVIII in clodronate-containing liposome-treated animals may
thus also result from the reduction in FVIII internalization by
CD11c+CD8a-negative DCs and presentation to T cells in
splenic germinal centers [27].
Several endocytic receptors specific for different FVIII
moieties have been characterized. Members of the low-density
lipoproteinreceptor(LDLR)familyrecognizeproteinstructures
in the heavy and light chains of FVIII [19,28,29]. ASGPRbinds
to galactose-ending glycans of the B domain of FVIII [17].
Recently,wehaveshownthatthemacrophagemannosereceptor
(MMR/CD206) interacts with mannose-ending glycans on the
A1 and C1 domains of the molecule [30]. The constitutive
expressionofMARCOandMOMA-1byMZmacrophageshas
been implicated in the capture of blood-borne antigens [31–33].
Whether the latter receptorsmaybe implicated in recognition of
FVIII by macrophages is being currently investigated.
Addendum
Designed research: S. Dasgupta, J. Bayry, A. Navarrete, S.V.
Kaveri, S. Lacroix-Desmazes; performed research: A. Navar-
rete, S.Dasgupta,S.Delignat,G.Caligiuri,O.D.Christophe,A.
Navarrete, S. Lacroix-Desmazes; analyzed data: A. Navarrete,
S. Dasgupta, S. Delignat, G. Caligiuri, O.D. Christophe, A.
Navarrete, S. Lacroix-Desmazes; wrote the paper: A. Navar-
rete, S. Delignat, J. Bayry, S.V. Kaveri, S. Lacroix-Desmazes.
Acknowledgements
This work was supported by INSERM, by CNRS, by UPMC-
Paris6, and by grants from Agence Nationale de la Recherche
(ANR-05-MRAR-030-1, ANR-07-JCJC-0100-01, ANR-07-
RIB-002-02) and Grifols (Barcelona, Spain). SuD and AMN
were recipients of fellowships from Fondation de la Recherche
Medicale and Region Ile-de-France (Paris, France).
We thank A. Rice (Mater Medical Research Institute and
University of Queensland, Australia) for giving us anti-CD11c
antibodies. Kogenate�, Helixate� and Advate� were kind gifts
from Bayer Healthcare (Lille, France), from CSL-Behring
1822 A. Navarrete et al
� 2009 International Society on Thrombosis and Haemostasis

(Paris, France) and from Baxter (Maurepas, France), respec-
tively. We thank C. Klein for advice with immunohistochem-
istry (�Cellular imaging and cytometry� facility, CRC,
INSERM U872, Paris).
Disclosure of Conflict of Interests
The authors state that they have no conflict of interest.
References
1 De Groot AS, Moise L. Prediction of immunogenicity for therapeutic
proteins: state of the art.Curr Opin DrugDiscov Devel 2007; 10: 332–40.
2 Ehrenforth S, Kreuz W, Scharrer I, Linde R, Funk M, Gungor T,
Krackhardt B, Kornhuber B. Incidence of development of factor VIII
and factor IX inhibitors in haemophiliacs. Lancet 1992; 339: 594–8.
3 Mebius RE, Kraal G. Structure and function of the spleen. Nat Rev
Immunol 2005; 5: 606–16.
4 Vremec D, Pooley J, Hochrein H, Wu L, Shortman K. CD4 and CD8
expression by dendritic cell subtypes in mouse thymus and spleen. J
Immunol 2000; 164: 2978–86.
5 Nolte MA, Hoen EN, van Stijn A, Kraal G, Mebius RE. Isolation of
the intact white pulp. Quantitative and qualitative analysis of the cel-
lular composition of the splenic compartments. Eur J Immunol 2000;
30: 626–34.
6 Nolte MA, Belien JA, Schadee-Eestermans I, Jansen W, Unger WW,
van Rooijen N, Kraal G, Mebius RE. A conduit system distributes
chemokines and small blood-borne molecules through the splenic
white pulp. J Exp Med 2003; 198: 505–12.
7 Cinamon G, Zachariah MA, Lam OM, Foss FW Jr, Cyster JG.
Follicular shuttling of marginal zone B cells facilitates antigen trans-
port. Nat Immunol 2008; 9: 54–62.
8 Dudziak D, Kamphorst AO, Heidkamp GF, Buchholz VR, Trump-
fheller C, Yamazaki S, CheongC, LiuK, LeeHW, ParkCG, Steinman
RM, Nussenzweig MC. Differential antigen processing by dendritic
cell subsets in vivo. Science 2007; 315: 107–11.
9 Breukels MA, Zandvoort A, van Den Dobbelsteen GP, van Den
Muijsenberg A, Lodewijk ME, Beurret M, Klok PA, Timens W,
Rijkers GT. Pneumococcal conjugate vaccines overcome splenic
dependency of antibody response to pneumococcal polysaccharides.
Infect Immun 2001; 69: 7583–7.
10 Delignat S, Dasgupta S, Andre S, Navarrete A, Kaveri S, Bayry J,
Andre M, Chtourou S, Tellier Z, Lacroix-Desmazes S. Comparison of
the immunogenicity of different therapeutic preparations of human
factor VIII in the murine model of hemophilia A.Haematologica 2007;
92: 1423–6.
11 van Rooijen N, Sanders A. Liposome mediated depletion of macro-
phages: mechanism of action, preparation of liposomes and applica-
tions. J Immunol Methods 1994; 174: 83–93.
12 Kraal G, Rodrigues H, Hoeben K, van Rooijen N. Lymphocyte
migration in the spleen: the effect of macrophage elimination. Immu-
nology 1989; 68: 227–32.
13 Morfini M, Longo G, Messori A, Lee M, White G, Mannucci P.
Pharmacokinetic properties of recombinant factor VIII compared with
a monoclonally purified concentrate (Hemofil M). The Recombinate
Study Group. Thromb Haemost 1992; 68: 433–5.
14 Mordenti J, Osaka G, Garcia K, Thomsen K, Licko V, Meng G.
Pharmacokinetics and interspecies scaling of recombinant human
factor VIII. Toxicol Appl Pharmacol 1996; 136: 75–8.
15 Jacquemin M, Neyrinck A, Hermanns MI, Lavend�homme R,
Rega F, Saint-Remy JM, PeerlinckK, vanRaemdonckD,Kirkpatrick
CJ. FVIII production by human lung microvascular endothelial cells.
Blood 2006; 108: 515–7.
16 Stel HV, van der Kwast TH, Veerman EC. Detection of factor VIII/
coagulant antigen in human liver tissue. Nature 1983; 303: 530–2.
17 Bovenschen N, Rijken DC, Havekes LM, Vlijmen BJ,Mertens K. The
B domain of coagulation factor VIII interacts with the asialoglyco-
protein receptor. J Thromb Haemost 2005; 3: 1257–65.
18 van Schooten CJ, Shahbazi S, Groot E, Oortwijn BD, van den Berg
HM, Denis CV, Lenting PJ. Macrophages contribute to the cellular
uptake of von Willebrand factor and factor VIII in vivo. Blood 2008;
112: 1704–12.
19 Saenko E, Yakhyaev A, Mikhailenko I, Strickland D, Sarafanov A.
Role of the low density lipoprotein-related protein receptor in media-
tion of factor VIII catabolism. J Biol Chem 1999; 274: 37685–92.
20 Burgdorf S, Lukacs-Kornek V, Kurts C. The mannose receptor
mediates uptake of soluble but not of cell-associated antigen for cross-
presentation. J Immunol 2006; 176: 6770–6.
21 Qian J, Burkly L, Smith E, Ferrant J, Hoyer L, Scott D, Haudenschild
C. Role of CD154 in the secondary immune response: the reduction of
pre-existing splenic germinal centers and anti-factor VIII inhibitor ti-
ter. Eur J Immunol 2000; 30: 2548–54.
22 FeuererM, Beckhove P, Garbi N,Mahnke Y, Limmer A, HommelM,
Hammerling GJ, Kyewski B, Hamann A, Umansky V, Schirrmacher
V. Bone marrow as a priming site for T-cell responses to blood-borne
antigen. Nat Med 2003; 9: 1151–7.
23 Kraal G, JanseM, Claassen E.Marginal metallophilic macrophages in
the mouse spleen: effects of neonatal injections of MOMA-1 antibody
on the humoral immune response. Immunol Lett 1988; 17: 139–44.
24 Eloranta ML, Alm GV. Splenic marginal metallophilic macrophages
and marginal zone macrophages are the major interferon-alpha/beta
producers in mice upon intravenous challenge with herpes simplex
virus. Scand J Immunol 1999; 49: 391–4.
25 TaylorPR,ZamzeS,StillionRJ,WongSY,GordonS,Martinez-Pomares
L. Development of a specific system for targeting protein to metallo-
philic macrophages. Proc Natl Acad Sci U S A 2004; 101: 1963–8.
26 Ciavarra RP, Taylor L, Greene AR, Yousefieh N, Horeth D, van
Rooijen N, Steel C, Gregory B, BirkenbachM, SekellickM. Impact of
macrophage and dendritic cell subset elimination on antiviral immu-
nity, viral clearance and production of type 1 interferon. Virol 2005;
342: 177–89.
27 Pooley JL, HeathWR, ShortmanK. Cutting edge: intravenous soluble
antigen is presented to CD4 T cells by CD8) dendritic cells, but cross-
presented to CD8 T cells by CD8+ dendritic cells. J Immunol 2001;
166: 5327–30.
28 Bovenschen N, Mertens K, Hu L, Havekes L, van Vlijmen B. LDL
receptor cooperates with LDL receptor-related protein in regulating
plasma levels of coagulation factorVIII in vivo.Blood 2005; 106: 906–12.
29 Lenting P, Neels J, van den Berg B, Clijsters P, Meijerman D,
Pannekoek H, van Mourik J, Mertens K, van Zonneveld A. The light
chain of factor VIII comprises a binding site for low density lipoprotein
receptor-related protein. J Biol Chem 1999; 274: 23734–9.
30 Dasgupta S, Navarrete AM, Bayry J, Delignat S, Wootla B, Andre S,
Christophe O, Nascimbeni M, Jacquemin M, Martinez-Pomares L,
Geijtenbeek TB, Moris A, Saint-Remy JM, Kazatchkine MD, Kaveri
SV, Lacroix-Desmazes S. A role for exposed mannosylations in pre-
sentation of human therapeutic self-proteins to CD4+T lymphocytes.
Proc Natl Acad Sci U S A 2007; 104: 8965–70.
31 van der Laan LJ, Kangas M, Dopp EA, Broug-Holub E, Elomaa O,
Tryggvason K, Kraal G.Macrophage scavenger receptorMARCO: in
vitro and in vivo regulation and involvement in the anti-bacterial host
defense. Immunol Lett 1997; 57: 203–8.
32 Jones C, Virji M, Crocker PR. Recognition of sialylated meningo-
coccal lipopolysaccharide by siglecs expressed on myeloid cells leads to
enhanced bacterial uptake. Mol Microbiol 2003; 49: 1213–25.
33 Delputte PL, Nauwynck HJ. Porcine arterivirus infection of alveolar
macrophages is mediated by sialic acid on the virus. J Virol 2004; 78:
8094–101.
Allo-immunization to therapeutic FVIII 1823
� 2009 International Society on Thrombosis and Haemostasis






![NK Cells Mediate Increase of Phagocytic Activity but Not of Proinflammatory Cytokine (Interleukin6 [IL6], Tumor Necrosis Factor Alpha, and IL12) Production Elicited in Splenic Macrophages](https://static.fdokumen.com/doc/165x107/632147240c12e1161503b7d3/nk-cells-mediate-increase-of-phagocytic-activity-but-not-of-proinflammatory-cytokine.jpg)













