
SPECIES BOUNDARIES AMONG THE TELMATOBIUS (ANURA: LEPTODACTYLIDAE) OF THE LAKE TITICACA BASIN:...
26
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. SPECIES BOUNDARIES AMONG THE TELMATOBIUS (ANURA: LEPTODACTYLIDAE) OF THE LAKE TITICACA BASIN: ALLOZYME AND MORPHOLOGICAL EVIDENCE Author(s): E. Benavides, J. C. Ortiz, J. W. Sites Jr. Source: Herpetologica, 58(1):31-55. 2002. Published By: The Herpetologists' League DOI: 10.1655/0018-0831(2002)058[0031:SBATTA]2.0.CO;2 URL: http://www.bioone.org/doi/full/10.1655/0018-0831%282002%29058%5B0031%3ASBATTA%5D2.0.CO%3B2 BioOne (www.bioone.org ) is an electronic aggregator of bioscience research content, and the online home to over 160 journals and books published by not-for-profit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of SPECIES BOUNDARIES AMONG THE TELMATOBIUS (ANURA: LEPTODACTYLIDAE) OF THE LAKE TITICACA BASIN:...

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries,and research funders in the common goal of maximizing access to critical research.
SPECIES BOUNDARIES AMONG THE TELMATOBIUS (ANURA:LEPTODACTYLIDAE) OF THE LAKE TITICACA BASIN: ALLOZYME ANDMORPHOLOGICAL EVIDENCEAuthor(s): E. Benavides, J. C. Ortiz, J. W. Sites Jr.Source: Herpetologica, 58(1):31-55. 2002.Published By: The Herpetologists' LeagueDOI: 10.1655/0018-0831(2002)058[0031:SBATTA]2.0.CO;2URL:http://www.bioone.org/doi/full/10.1655/0018-0831%282002%29058%5B0031%3ASBATTA%5D2.0.CO%3B2
BioOne (www.bioone.org) is an electronic aggregator of bioscience research content, and the online home to over 160journals and books published by not-for-profit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’sTerms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries orrights and permissions requests should be directed to the individual publisher as copyright holder.

March 2002] 31HERPETOLOGICA
Herpetologica, 58(1), 2002, 31–55� 2002 by The Herpetologists’ League, Inc.
SPECIES BOUNDARIES AMONG THE TELMATOBIUS(ANURA: LEPTODACTYLIDAE) OF THE LAKE TITICACABASIN: ALLOZYME AND MORPHOLOGICAL EVIDENCE
E. BENAVIDES1,2, J. C. ORTIZ1, AND J. W. SITES, JR.2
1Casilla 160-C, Departamento de Zoologıa, Facultad de Naturales y Oceanograficas,Universidad de Concepcion, Concepcion, Chile
2Department of Zoology and M. L. Bean Life Science Museum, Brigham Young University,Provo, UT 84602, USA
ABSTRACT: The frog genus Telmatobius represents the radiation of 49 recognized species con-fined to high elevation aquatic habitats in the Andes Mountains of South America. The genus hasnot been shown to be monophyletic, and issues of species boundaries and relationships are poorlyresolved. In this study, we sampled the four recognized subspecies of T. albiventris from LakeTiticaca, and other species (T. crawfordi, T. culeus, and T. marmoratus) from within the lake andthe greater Titicaca Basin. We used allozyme characters and multivariate assessments of morpho-metric variation to test species boundaries in this complex. Our results suggest that four evolutionaryspecies can be recognized: T. culeus (which includes T. albiventris and T. crawfordi), T. marmoratus,T. affinis halli, and T. sp. nov. Unique combinations of allozyme and morphometric variables candistinguish these, but only the sample of T. culeus is large enough, relative to the proportion offixed characters, to evaluate distinctness of diagnostic characters with a high degree of statisticalconfidence (� � 0.05). Phylogenetic analysis of 10 polymorphic allozyme loci revealed weak supportfor monophyly of Lake Titicaca Telmatobius. All taxa from Lake Titicaca examined here (under thename T. culeus) are interpreted as a single evolutionary species, but differences in body sizes andmicrohabitats provide circumstantial evidence for adaptation along ecological resource gradients.The possibility of ecological speciation among different morphs within the lake is considered, alongwith evidence needed to test this hypothesis.
Key words: Telmatobius; Allozymes; Null alleles; Morphometrics; Species boundaries; Resourceniche polymorphism; Lake Titicaca
THE frog genus Telmatobius Wiegmann,1835 contains 49 currently recognized spe-cies typically found in the Andes MountainRange at altitudes above 2500 m (Lynch,1986). Geographically, the genus extendsfrom northern Ecuador (02� 05� S) tonorthern Argentina (29� 39� S) and Chile(22� 51� S) (Benavides, 1999; Lavilla, per-sonal communication). There is no clearevidence for monophyly of the group, asdiagnosed by hypothesized synapomor-phies (de la Riva, 1994; Formas et al.,1999; Lynch, 1978; Trueb, 1979; the pos-sible synapomorphies suggested by Wiens,
1993, were rejected by Formas et al.,1999), and most species have been as-signed to the genus on a phenetic basisand following typological criteria, increas-ing confusion regarding the identificationof species (e.g., Laurent, 1970; Vellard,1951, 1953, 1960).
Historically, Lake Titicaca was a primarydestination of early expeditions to the cen-tral region of the Andes (Agassiz and Gar-man, 1876; D’orbigny, 1835–1847; Gilson,1939; Pentland, 1848). Specimens collect-ed during those trips have led to the de-scription of four species of Telmatobius

32 [Vol. 58, No. 1HERPETOLOGICA
from the Titicaca Basin. Two of these areprimarily lacustrine species: T. albiventris(Parker, 1940) and T. culeus (Garman,1876), whereas T. marmoratus (Dumeriland Bibron, 1841) and T. crawfordi (Park-er, 1940) are semi-aquatic species inhab-iting streams and shallow lagoons of thegreater Titicaca Basin.
Although Vellard’s thorough revision ofAndean Telmatobius predates the currentphylogenetic paradigm, he recognized thefour above-mentioned species as a single,albeit highly polymorphic, evolutionarylineage (Vellard, 1951, 1953, 1960, 1969).The current taxonomic arrangement isbased on a combination of polymorphicmorphological characters and more stableecological features (Vellard, 1953, 1960).Species of Telmatobius from the TiticacaBasin have also been affiliated to a pre-sumably undifferentiated lowland taxon(2500 m; approximately 250 km east ofLake Titicaca), T. marmoratus hintoni(Parker, 1940), through a number of sub-species that represent intermediatemorphs between each species group, oftenin questionable allopatry. The outcome ofthis arrangement was the rather arbitraryspecies boundaries among all taxa de-scribed, a point that has been repeatedlyemphasized; ‘‘les coupes specifiques etsous-specifiques sont tres difficiles a eta-blir et deporvues de toute valeur reelle’’(‘‘specific and subspecific boundaries arehard to establish, and they lack any realvalue’’; Vellard, 1952). All of the species inthe Titicaca Basin are thought to be partof an albiventris-crawfordi-culeus-mar-moratus complex (Cei, 1986), but withinthis group there has been no rigorous eval-uation of species boundaries.
Within Lake Titicaca, T. albiventris isgeographically restricted to shallow waterareas around the periphery of the LagoGrande (Fig. 1). Initially described as asubspecies of T. culeus, it was later as-signed to specific rank based on ecologicaland morphological differences (Vellard,1951). It is confined to shallow water hab-itats, has a less emarginated toe webbing,possesses a dorsal disc [the thickened dor-sal structure described by Angel, 1926 (pp.108–109) and Parker, 1940 (p. 210)], paler
ventral coloration, and smaller adult sizethan T. culeus (Table 1). In addition to thenominate form, T. albiventris albiventris(Parker, 1940) from the shoreline of theIsla de Taquiri in Lago Pequeno, three ad-ditional subspecies are currently recog-nized: T. a. globulosus Vellard, 1960, isconfined to Playa Copani, in the southernembayment of the lake; T. a. punensis Vel-lard, 1951, applies to a form inhabiting Ba-hıa de Puno; and T. a. parkeri Vellard,1951, is restricted to the shallow LagunaArapa (Fig. 1). Intraspecific diagnosis ofthese subspecies is ambiguous and basedmainly on locality, although size and se-lected body proportions are presumablydifferent between some taxa (Vellard,1960). Furthermore, the subspecies inLago Pequeno are, in all probability, sym-patric (T. a. albiventris and T. a. globulo-sus; Vellard, 1960), and Laguna Arapa andTiticaca Lake may be connected in excep-tionally rainy years (Cei, 1986).
Telmatobius culeus is the most distinctspecies within Lake Titicaca; it was de-scribed on the basis of specimens capturedin deep water far from the shoreline (Gar-man, 1876), and it is characterized by un-usual morphological features presumablyrelated to life in this habitat (Hutchinson,1982; Hutchinson et al., 1976; Macedo,1960). The nominate form is well charac-terized by distinct phenotypic featuressuch as large body size, a large flattenedhead with a round snout, a thick dorsaldisc, smooth skin with a richly vascularizedsurface, and highly developed lateral der-mal folds (bagginess). This form is foundalong the eastern coast of Lake Titicaca,but it extends partially into Lago Pequenothrough the Estrecho de Tiquina (Fig. 1).However, it is absent from shallower areasin the southern basin, and from the La-guna Arapa and the Bahıa de Puno, in thenorthern basin. Other subspecies are pre-sent in rivers flowing into the lake (T. cu-leus fluviatilis, T. culeus dispar) and insome small lagoons elsewhere within thebasin (T. culeus escomeli, T. culeus lacus-tris). More distant from Lake Titicaca, T.culeus exsul was described on the basis ofone specimen, taken from isolated warmsprings near the Rio Yura at 2500 m, about
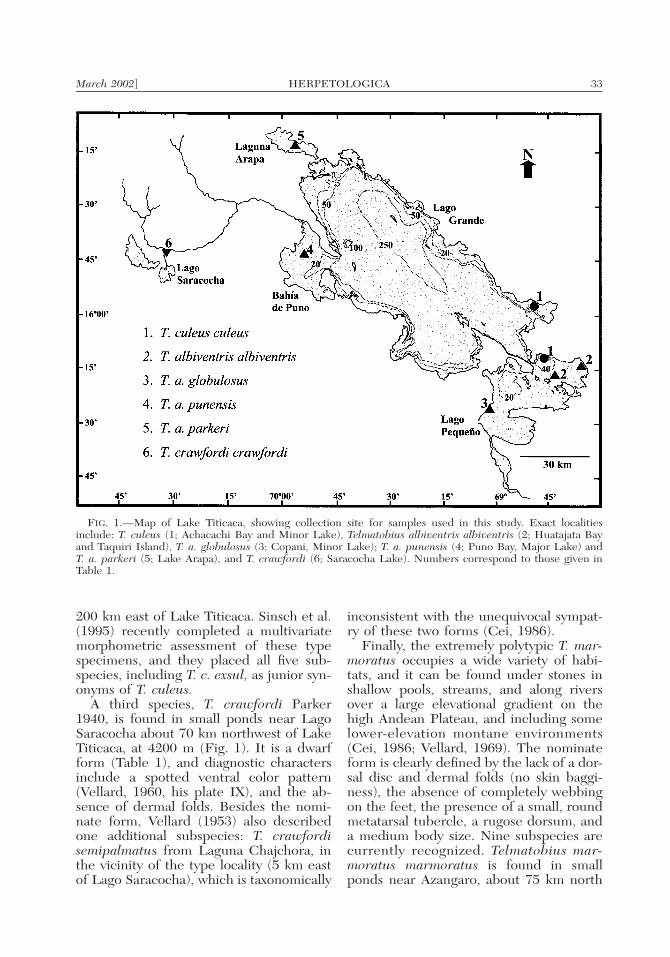
March 2002] 33HERPETOLOGICA
FIG. 1.—Map of Lake Titicaca, showing collection site for samples used in this study. Exact localitiesinclude: T. culeus (1; Achacachi Bay and Minor Lake), Telmatobius albiventris albiventris (2; Huatajata Bayand Taquiri Island), T. a. globulosus (3; Copani, Minor Lake); T. a. punensis (4; Puno Bay, Major Lake) andT. a. parkeri (5; Lake Arapa), and T. crawfordi (6; Saracocha Lake). Numbers correspond to those given inTable 1.
200 km east of Lake Titicaca. Sinsch et al.(1995) recently completed a multivariatemorphometric assessment of these typespecimens, and they placed all five sub-species, including T. c. exsul, as junior syn-onyms of T. culeus.
A third species, T. crawfordi Parker1940, is found in small ponds near LagoSaracocha about 70 km northwest of LakeTiticaca, at 4200 m (Fig. 1). It is a dwarfform (Table 1), and diagnostic charactersinclude a spotted ventral color pattern(Vellard, 1960, his plate IX), and the ab-sence of dermal folds. Besides the nomi-nate form, Vellard (1953) also describedone additional subspecies: T. crawfordisemipalmatus from Laguna Chajchora, inthe vicinity of the type locality (5 km eastof Lago Saracocha), which is taxonomically
inconsistent with the unequivocal sympat-ry of these two forms (Cei, 1986).
Finally, the extremely polytypic T. mar-moratus occupies a wide variety of habi-tats, and it can be found under stones inshallow pools, streams, and along riversover a large elevational gradient on thehigh Andean Plateau, and including somelower-elevation montane environments(Cei, 1986; Vellard, 1969). The nominateform is clearly defined by the lack of a dor-sal disc and dermal folds (no skin baggi-ness), the absence of completely webbingon the feet, the presence of a small, roundmetatarsal tubercle, a rugose dorsum, anda medium body size. Nine subspecies arecurrently recognized. Telmatobius mar-moratus marmoratus is found in smallponds near Azangaro, about 75 km north
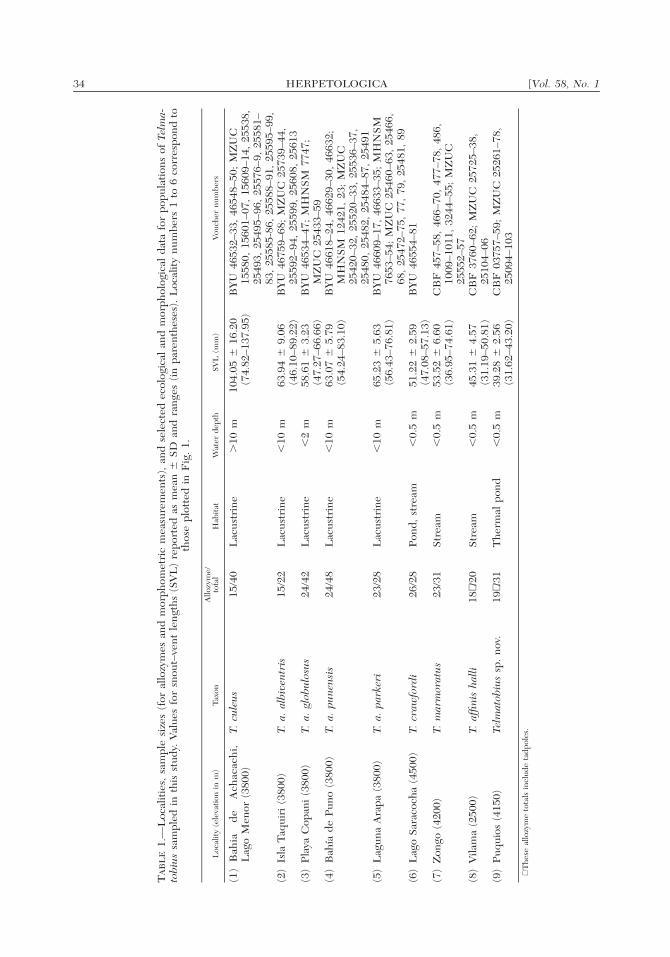
34 [Vol. 58, No. 1HERPETOLOGICA
TA
BL
E1.
—L
ocal
itie
s,sa
mpl
esi
zes
(for
allo
zym
esan
dm
orph
omet
ric
mea
sure
men
ts),
and
sele
cted
ecol
ogic
alan
dm
orph
olog
ical
data
for
popu
lati
ons
ofTe
lma-
tobi
ussa
mpl
edin
this
stud
y.V
alue
sfo
rsn
out–
vent
leng
ths
(SV
L)
repo
rted
asm
ean
�SD
and
rang
es(i
npa
rent
hese
s).
Loc
alit
ynu
mbe
rs1
to6
corr
espo
ndto
thos
epl
otte
din
Fig
.1.
Loc
alit
y(e
leva
tion
inm
)T
axon
Allo
zym
e/to
tal
Hab
itat
Wat
erde
pth
SVL
(mm
)V
ouch
ernu
mbe
rs
(1)
Bah
iad
eA
chac
ach
i,L
ago
Men
or(3
800)
T.cu
leus
15/4
0L
acus
trin
e�
10m
104.
05�
16.2
0(7
4.82
–137
.95)
BY
U46
532–
33,
4654
8–50
;M
ZU
C15
580,
1560
1–07
,15
609–
14,
2553
8,25
493,
2549
5–96
,25
576–
9,25
581–
83,
2558
5-86
,25
588–
91,
2559
5–99
,(2
)Is
laT
aqui
ri(3
800)
T.a.
albi
vent
ris
15/2
2L
acus
trin
e�
10m
63.9
4�
9.06
(46.
10–8
9.22
)B
YU
4675
9–68
;M
ZU
C25
739–
44,
2559
2–94
,25
599,
2560
8,25
613
(3)
Pla
yaC
opan
i(3
800)
T.a.
glob
ulos
us24
/42
Lac
ustr
ine
�2
m58
.61
�3.
23(4
7.27
–66.
66)
BY
U46
534–
47;
MH
NSM
7747
;M
ZU
C25
433–
59(4
)B
ahıa
deP
uno
(380
0)T.
a.pu
nens
is24
/48
Lac
ustr
ine
�10
m63
.07
�5.
79(5
4.24
–83.
10)
BY
U46
618–
24,
4662
9–30
,46
632;
MH
NSM
1242
1,23
;M
ZU
C25
420–
32,
2552
0–33
,25
536–
37,
2548
0,25
482,
2548
4–87
,25
491
(5)
Lag
una
Ara
pa(3
800)
T.a.
park
eri
23/2
8L
acus
trin
e�
10m
65.2
3�
5.63
(56.
43–7
6.81
)B
YU
4660
9–17
,46
633–
35;
MH
NSM
7653
–54;
MZ
UC
2546
0–63
,25
466,
68,
2547
2–75
,77
,79
,25
481,
89(6
)L
ago
Sara
coch
a(4
500)
T.cr
awfo
rdi
26/2
8P
ond,
stre
am�
0.5
m51
.22
�2.
59(4
7.08
–57.
13)
BY
U46
554–
81
(7)
Zon
go(4
200)
T.m
arm
orat
us23
/31
Stre
am�
0.5
m53
.52
�6.
60(3
6.95
–74.
61)
CB
F45
7–58
,46
6–70
,47
7–78
,48
6,10
09–1
011,
3244
–55;
MZ
UC
2555
2–57
(8)
Vila
ma
(250
0)T.
affin
isha
lli18
∗/20
Stre
am�
0.5
m45
.31
�4.
57(3
1.19
–50.
81)
CB
F37
60–6
2;M
ZU
C25
725–
38,
2510
4–06
(9)
Puq
uios
(415
0)Te
lmat
obiu
ssp
.no
v.19
∗/31
The
rmal
pond
�0.
5m
39.2
8�
2.56
(31.
62–4
3.20
)C
BF
0375
7–59
;M
ZU
C25
261–
78,
2509
4–10
3∗
The
seal
lozy
me
tota
lsin
clud
eta
dpol
es.

March 2002] 35HERPETOLOGICA
of Lake Titicaca. Three subspecies arethought to be present in different parts ofLake Titicaca: T. m. angustipes, T. m. ri-parius, and T. m. rugosus, all of which maybe in contact with the widespread T. m.marmoratus. Three other forms are re-stricted to eastern slope drainage of theAndes (i.e., T. m. microcephalus, T. m. pus-tulosus, and T. m. pseudojelski), and an-other, T. m. hintoni, is confined to westerndrainages of the Andes. Finally, T. m. gigasVellard, 1969, was described on the basisof a single female from Huallamarca, Oru-ro, Bolivia, about 175 km south of LakeTiticaca. This taxon shows all typical fea-tures of the deep-water morphs of T. cu-leus of Lake Titicaca, including large bodysize (SVL � 109 mm), flattened head witha rounded snout, well-developed dorsaldisc, and prominent lateral skin folds. Re-cent exploration of the type locality hasconfirmed the continued existence of T. m.gigas (de La Riva, personal communica-tion). Surprisingly, it is a stream-dwellingform found in ponds of 0.6–1.0 m deep,but its real identity cannot yet be ascer-tained.
In summary, neither Parker (1940), Cei(1986), nor Vellard (1953, 1960, 1991)reached firm conclusions regarding thespecies and subspecies status of the frogsof Lake Titicaca. All stressed the extensivemorphological variation and the lack ofdistinct taxonomic boundaries among thewidely intergrading populations of Telma-tobius from the Titicaca Basin and sur-rounding region. In addition, the descrip-tion of presumably ‘‘transitional’’ morphssuch as T. a. parkeri (thought to representan intermediate morph between T. a. al-biventris and T. culeus; Vellard, 1953), andT. m. angustipes and T. m. rugosus, (con-sidered intermediate between T. culeusand T. marmoratus; Vellard, 1953), ren-ders even more difficult the interpretationof species boundaries among populationsin Lake Titicaca.
Species of Telmatobius are generallyconservative with respect to morphologicalcharacters, and adaptation to a fully aquat-ic life style has probably resulted in con-vergence in features such as coloration,general morphology, and behavior. This
makes it difficult to find useful charactersfor sorting out species other than size, pre-sumably body proportions, and to a certainextent, osteological characters. Trueb(1979) has shown that most of the mor-phological characters upon which speciesand subspecies diagnoses were made aresexually, seasonally, or ontogenetically var-iable, and as a rule, ambiguous descrip-tions, discordant patterns of intra- and in-terspecific variation, and inadequate diag-noses are typical for all but the most re-cently described taxa of Telmatobius (Salasand Sinsch, 1996; Trueb, 1979; Wiens,1993).
In this context, the aim of this study isto test species boundaries in the complete-ly lacustrine taxa of T. albiventris and se-lected other taxa of the albiventris-craw-fordi-culeus-marmoratus ‘‘complex’’, byextensive populational sampling, and com-parisons of multiple samples with speci-mens from type localities. We used allo-zyme characters and a multivariate assess-ment of morphometric variables, and weadopted the evolutionary species concept(Frost and Hillis, 1990; Frost and Kluge,1994; Wiley, 1978) as the philosophicaljustification for species recognition.
MATERIALS AND METHODS
Collection of Specimens
Samples (n � 290 frogs) from nine pop-ulations representing six putative speciesof Telmatobius were collected betweenAugust 1997 and February 1998. Popula-tions from Lake Titicaca (14� 17� S, 68� 71�W) were collected from type localities forall recognized taxa except T. marmoratus(Table 1). One distant population of T. m.marmoratus was, however, collected from75 km southeast of Lake Titicaca at 4200m on the road to Valle de Zongo. Two oth-er taxa were used as outgroups for theLake Titicaca populations. Telmatobius af-finis halli, previously confused with T. halliNoble, 1938 (Benavides, 1999), was col-lected from Arroyo Vilama (22� 51� 432 Sto 68� 23� 256 W; 2600 m), near San Pe-dro de Atacama (Chile). A second popu-lation, T. sp. nov, was collected from asmall, warm spring at Puquios (21� 00� 42

36 [Vol. 58, No. 1HERPETOLOGICA
TABLE 2.—Enzymes, enzyme commission numbers (following recommendations of Murphy et al., 1996),enzyme abbreviations, number of putative loci, buffer systems (1. TBE-NAD, pH 9.1; 2. Tris-Citric, pH 7.0;
3. Lithium hydroxide, pH 8.0), and tissue sources (L � liver, M � skeletal muscle) used in this study.
Enzyme (E.C. number) Abbreviations No. of loci Buffer Tissue
Iditol dehydrogenase (1.1.1.14)Glucose-6-phosphate isomerase (5.3.1.9)Phosphoglucomutase (5.4.2.2)Glycerol-3-phosphate dehydrogenase (1.1.1.8)Malate dehydrogenase (1.1.1.37)
IDDHGPIPGMG3PDH∗MDH
11122
21322
MM, LM, LMM, L
Superoxide dismutase (1.15.1.1)Malate dehydrogenase (NADP) (1.1.1.40)Aconitate hydratase (4.2.1.3)Lactate dehydrogenase (1.1.1.27)Esterase (3.1.1.—)
SODMDHPACOHLDHEST
22222
3222
2, 3
MMMM, LL
Leucine-aminopeptidase (3.4.11.1)Aspartate aminotransferase (2.6.1.1)Alcohol dehydrogenase (1.1.1.1)Phosphogluconate dehydrogenase (1.1.1.44)Isocitrate dehydrogenase (1.1.1.42)
LAPAATADHPGDHIDH
22112
31, 2
12
2, 3
MLMMM, L
∗ This locus was fixed for a duplication in all samples, and displayed the expected three banded ‘‘heterozygote’’ isozyme pattern (see details inSites and Murphy, 1991).
S to 68� 23� 256 W; 4150 m approximate-ly) 15 km northwest of Ollague, on the Bo-livian border; this taxon is currently beingdescribed by Benavides et al. (unpub-lished). The two last localities are part ofthe Provincia El Loa, II Region de Anto-fagasta, Chile. Most localities are shown inFig. 1, and all are described in Table 1,along with some relevant ecological andmorphological observations. Voucher spec-imens are cataloged in the Coleccion Bo-liviana de Fauna (CBF), Museo de His-toria Natural Universidad Nacional Mayorde San Marcos (MHNSM), Museo deZoologıa de la Universidad de Concepcion(MZUC), and M. L. Bean Life ScienceMuseum, Brigham Young University(BYU).
Protein ElectrophoresisSamples of liver and skeletal muscle
were extracted from a subsample (n �187) of freshly killed specimens and storedat �80 C. Small pieces of tissue wereground manually and diluted 1:1 (muscle)or 1:3 (liver) with 0.0099 M Tris-0.018 MEDTA, pH 6.8 plus 1% NADP (w/v). Ho-mogenates were centrifuged at 15,000 rpmfor 5 min, and supernatant was stored forlater use. The tissue extracts were analyzedby standard techniques of horizontalstarch gel electrophoresis (Murphy et al.,1996; Selander et al., 1971). Three buffer
systems were used: (1) TBE-NAD 9.1(electrode and gel): 175 mM Tris-17.5 mMboric acid, 2.75mM EDTA plus NAD 100mg (gel), 60 mg (electrode); (2) TC-7,electrode: 0.135 M Tris, 0.043 M citricacid, gel: 1:9 electrode buffer; and (3)LiOH, electrode: 0.06 M lithium hydrox-ide, 0.3 M boric acid, pH. 8.1; gel: 0.03 MTris-0.05 M citric acid, pH 8.5, 1:9 elec-trode; and procedures for staining fol-lowed Richardson et al. (1986), Morizotand Schmidt (1990), and Murphy et al.(1996). All enzymes, tissue sources, andbuffer systems are listed in Table 2.
Population Genetic AnalysisElectromorphs of any given locus were
considered homologous if they had thesame mobility. Loci were numbered fromanode to cathode and electromorphs werelabeled a, b, c, etc., in order of increasinganodal mobility. Individual genotypes wereused to calculate electromorph frequen-cies, proportion of polymorphic loci, av-erage heterozygosities, mean number ofelectromorphs per polymorphic locus, andgenetic identity and distance coefficients(Nei, 1978). We then tested goodness offit to Hardy-Weinberg proportions bymeans of a U exact test with heterozygotedeficiency as the alternative hypothesis(Rousset and Raymond, 1995); the signif-icance of this test was evaluated by a Mar-

March 2002] 37HERPETOLOGICA
kov chain method (Guo and Thompson,1992). We estimated linkage disequilibri-um using Fisher’s exact test (Garnier-Gereand Dillman, 1992), and for those popu-lations interpreted as possibly conspecific(see criteria below), we tested genetic dif-ferentiation by using an exact probabilitychi-square test (Raymond and Rousset,1995a). We also calculated the F statistic(� � P) of Weir and Cockerman (1984),and from this we estimated the number ofmigrants per generation using the privateallele method of Barton and Slatkin(1986). We implemented all calculationswith the GENEPOP-3.1b (Raymond andRousset, 1995b) and GDA-1.0 (Lewis andZaykin, 1997; see http://lewis.eeb.uconn.edu/lewishome/gda.html) computer pro-grams.
Phylogenetic AnalysisWe performed a phylogenetic analysis of
all samples, based on the allozyme matrixin which four loci with ‘‘fixed’’ alternativestates (Mdh-2, Est-2, Acoh-2, and G3pdh–1 in Table 3) were entered into a PAUPmatrix as characters, and with alternativeelectromorphs coded as unordered states.The remaining six polymorphic loci werecoded by transforming the electromorphfrequencies of each locus into Manhattandistances, which served as weighted valuesin a step matrix for each of these loci (daSilva and Sites, 2001; Wiens, 1995). Thisapproach incorporates frequency infor-mation and permits the use of multistatecharacters, and frequency-based methods(distance, likelihood, or parsimony) appearto provide the most accurate estimates ofphylogeny (Wiens, 2000; Wiens and Ser-vedio, 1997, 1998).
Morphometric Data AnalysisWe used 17 morphometric variables to
quantify morphological differentiationamong and within Lake Titicaca Basinsamples. The two distinct southern taxa, T.affinis halli and T. sp. nov., were used ascomparative groups in building principalcomponents and discriminant functions.We assessed sexual maturity on the basisof the development of nuptial excrescenc-es in males. Measurements of external fea-
tures were taken with a digital Mitutoyocaliper to the nearest 0.05 mm, and ab-breviations used throughout are: SVL(snout–vent length); TIBL (tibia length);FOOT (foot length); HLEN (head length);HWID (head width); LJW (lower jawwidth); SED (snout–eye distance, anterioredge of eye to tip of snout); IOD (inter-orbital distance); ENOS (eye–nostril dis-tance, anterior corner of eye to posterioredge of naris); IND (internarial distance);EYE (eye diameter); HNDL (handlength); RDL (radioulnar length, flexed el-bow to proximal edge of outer palmar tu-bercle); FEML (femur length, from ventto flexed knee); HUML (humerus length,from the body central axis to flexed el-bow); NMD (naris–mouth distance), andWEBL (web length, proximal edge of theinner metatarsal tubercle to distal edge ofthe III–IV toe web). We measured all bi-lateral characters on the right side. Allmeasurements are illustrated in Fig. 2.
Because selection presumably acts uponwhole phenotypes and not on single char-acters, we chose a multivariate approachto assess size and shape variation separate-ly. Three procedures, Multivariate Analysisof Variance (MANOVA), Principal Com-ponent Analysis (PCA), and DiscriminantFunction Analysis (DFA) were used to an-swer three questions regarding morpho-metric variation among nine samples ofTelmatobius. First, are samples a priori as-signed to named taxa morphometricallydistinct? Second, are taxa recognized by al-lozymic criteria also morphometrically dif-ferentiated? Finally, upon which charac-ters is this differentiation based? To an-swer these questions, we assessed patternsof morphometric variation in a hierarchicaldesign that was suggested by the existingtaxonomy and results of the allozyme anal-yses. First, variation within and amongsubspecies of T. albiventris was summa-rized by MANOVA and PCA of non-trans-formed variables. Second, variation withinand among samples of T. albiventris, T.crawfordi, and T. culeus was assessed byPCA of transformed variables. Third, var-iation within and among Lake Titicaca spe-cies (i.e., T. albiventris, T. crawfordi, andT. culeus), versus Lake Titicaca Basin spe-
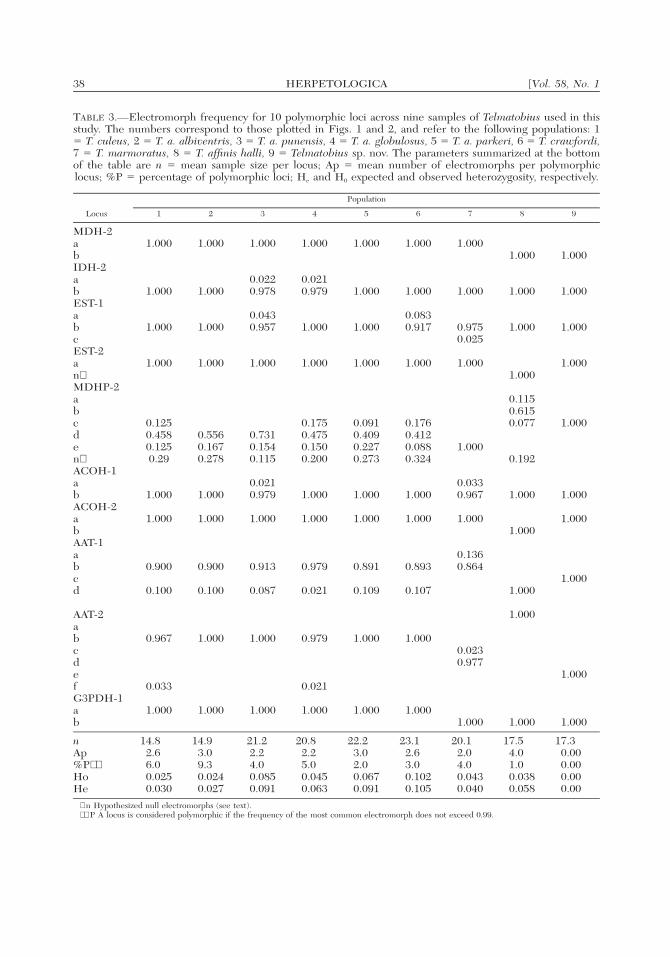
38 [Vol. 58, No. 1HERPETOLOGICA
TABLE 3.—Electromorph frequency for 10 polymorphic loci across nine samples of Telmatobius used in thisstudy. The numbers correspond to those plotted in Figs. 1 and 2, and refer to the following populations: 1� T. culeus, 2 � T. a. albiventris, 3 � T. a. punensis, 4 � T. a. globulosus, 5 � T. a. parkeri, 6 � T. crawfordi,7 � T. marmoratus, 8 � T. affinis halli, 9 � Telmatobius sp. nov. The parameters summarized at the bottomof the table are n � mean sample size per locus; Ap � mean number of electromorphs per polymorphiclocus; %P � percentage of polymorphic loci; He and H0 expected and observed heterozygosity, respectively.
Locus
Population
1 2 3 4 5 6 7 8 9
MDH-2abIDH-2ab
1.000
1.000
1.000
1.000
1.000
0.0220.978
1.000
0.0210.979
1.000
1.000
1.000
1.000
1.000
1.000
1.000
1.000
1.000
1.000EST-1abcEST-2an∗
1.000
1.000
1.000
1.000
0.0430.957
1.000
1.000
1.000
1.000
1.000
0.0830.917
1.000
0.9750.025
1.000
1.000
1.000
1.000
1.000
MDHP-2abcden∗
0.1250.4580.1250.29
0.5560.1670.278
0.7310.1540.115
0.1750.4750.1500.200
0.0910.4090.2270.273
0.1760.4120.0880.324
1.000
0.1150.6150.077
0.192
1.000
ACOH-1abACOH-2ab
1.000
1.000
1.000
1.000
0.0210.979
1.000
1.000
1.000
1.000
1.000
1.000
1.000
0.0330.967
1.000
1.000
1.000
1.000
1.000
AAT-1abcd
0.900
0.100
0.900
0.100
0.913
0.087
0.979
0.021
0.891
0.109
0.893
0.107
0.1360.864
1.0001.000
AAT-2abcdef
0.967
0.033
1.000 1.000 0.979
0.021
1.000 1.0000.0230.977
1.000
1.000
G3PDH-1ab
1.000 1.000 1.000 1.000 1.000 1.0001.000 1.000 1.000
nAp%P∗∗HoHe
14.82.66.00.0250.030
14.93.09.30.0240.027
21.22.24.00.0850.091
20.82.25.00.0450.063
22.23.02.00.0670.091
23.12.63.00.1020.105
20.12.04.00.0430.040
17.54.01.00.0380.058
17.30.000.000.000.00
∗ n Hypothesized null electromorphs (see text).∗∗ P A locus is considered polymorphic if the frequency of the most common electromorph does not exceed 0.99.
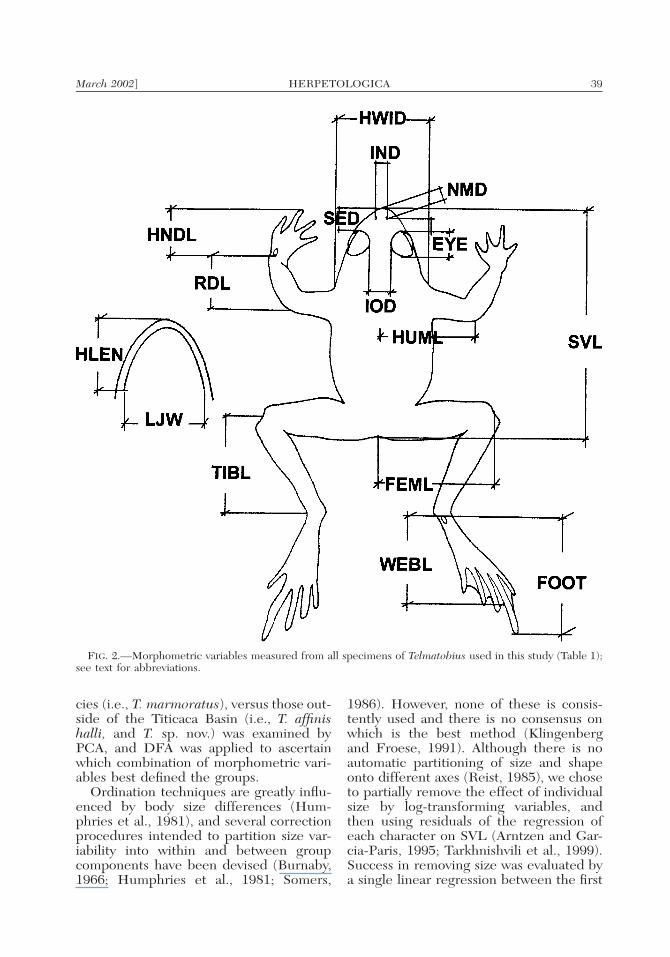
March 2002] 39HERPETOLOGICA
FIG. 2.—Morphometric variables measured from all specimens of Telmatobius used in this study (Table 1);see text for abbreviations.
cies (i.e., T. marmoratus), versus those out-side of the Titicaca Basin (i.e., T. affinishalli, and T. sp. nov.) was examined byPCA, and DFA was applied to ascertainwhich combination of morphometric vari-ables best defined the groups.
Ordination techniques are greatly influ-enced by body size differences (Hum-phries et al., 1981), and several correctionprocedures intended to partition size var-iability into within and between groupcomponents have been devised (Burnaby,1966; Humphries et al., 1981; Somers,
1986). However, none of these is consis-tently used and there is no consensus onwhich is the best method (Klingenbergand Froese, 1991). Although there is noautomatic partitioning of size and shapeonto different axes (Reist, 1985), we choseto partially remove the effect of individualsize by log-transforming variables, andthen using residuals of the regression ofeach character on SVL (Arntzen and Gar-cia-Paris, 1995; Tarkhnishvili et al., 1999).Success in removing size was evaluated bya single linear regression between the first

40 [Vol. 58, No. 1HERPETOLOGICA
principal component, PC I (the dependentvariable), and SVL (the independent vari-able). Fit of the data to critical assump-tions was assessed by two methods. Datadistribution was checked by univariatenormal distribution tests, which can betaken as partial proof of normal multivar-iate distribution. Violations of homosce-dasticity were checked with Bartlett’s (uni-variate) and Box M (multivariate) tests. Allstatistical procedures were performed us-ing STATISTICA 5.1 (StatSoft, 1995).
Species DiagnosisMethodological aspects of testing spe-
cies boundaries have received little atten-tion, in spite of considerable focus on thespecies–concept problem (Frost and Hil-lis, 1990; Frost and Kluge, 1994; Wiens,1999). Although differences among spe-cies concepts are often rooted in differentontological views, de Queiroz (1998) sug-gested that all species concepts agree fun-damentally on what species are (distinctevolutionary entities), but differ in criteriafor their recognition. Sites and Crandall(1997) suggested greater clarity would beachieved if investigators would provide apriori operational criteria by which speciesboundaries may be tested as a hypothesis,and empirically accepted or rejected.
From an operational perspective, sev-eral authors have emphasized that the is-sue of deciding which characters are poly-morphic and which are ‘‘fixed’’ is certainlythe most relevant, because some authorsconsider only fixed characters to be prop-erly diagnostic of species boundaries (Da-vis and Nixon, 1992). Recently, Wiens andServedio (2000) demonstrated that docu-menting true fixation of a character re-quires sampling every individual in a spe-cies, while arguing probabilistically that acharacter is present only at a low frequen-cy only requires sampling many individu-als. In order to overcome this limitation,Wiens and Servedio (2000) described amore realistic procedure, which allowscharacters to be diagnostic for species ifthey can be shown to be below a certainfrequency cut-off with a high degree ofstatistical confidence. In this approach, theH0 states that low frequency character
states are present at a frequency greaterthan p (the pre-determined cut-off) evenif they are undetected. Rejection of the Ho
suggests that one or more characters arebelow the frequency cut-off, which is tak-en as evidence of reduced or absent geneflow. We employ this approach here, usinga character state frequency cut-off of 10%,with a 5% confidence interval (� � 0.05),for a statistically supported delimitation ofspecies boundaries. In the discussion, wereturn to this issue with respect to caveatsand potential limitations of the method.
In the absence of a statistical test, wethink diagnosis of independent evolution-ary trajectories is strongest if it is based ona concordant pattern of differentiation inunlinked markers (Avise and Ball, 1990;Avise and Wollenberg, 1997), and this cri-terion is applied here. Therefore, we ex-plicitly diagnose taxa as species by thepresence of ‘‘fixed’’ electromorphic differ-ences based on the Wiens-Servedio test,and also the presence of unique electro-morphic combinations (non-fixed privatealleles) if they are concordant with otherdiagnostic markers, and/or morphometricdifferences (see Gergus, 1998; Mink andSites, 1996, for non-statistical applicationsof a concordance approach).
RESULTS
Allozymes: Patterns of Variation
We examined 15 enzymes encoded by25 presumptive loci (Table 2), and 10 lociwere variable within or between popula-tions (Mdh-2, Idh-2, Est-1, Est-2, Mdhp-2, Acoh-1, Acoh-2, Aat-1, Aat-2, G3pdh–1). The electromorph frequencies for thevariable loci, percentage of polymorphicloci, observed and expected heterozygosi-ty, and mean number of electromorphs perpolymorphic locus are shown in Table 3.Of the 10 variable loci, four (Mdh-2, Est-2, Acoh-2, and G3pdh–1) revealed among-sample variation in the form of seeminglyfixed, alternative electromorphs. One ofthese, Est-2, is hypothesized to be fixed fora null electromorph (Katoh and Foltz,1989) in T. affinis halli (sample 8), becauseno product was ever resolved for any ofthe 18 individuals included in this sample

March 2002] 41HERPETOLOGICA
FIG. 3.—Isozyme patterns showing homozygotes for a presumed null electromorph at the Est-2 locus inT. affins halli, and normal enzyme phenotypes in Telmatobius sp. nov. and T. culeus; the ‘‘normal’’ patternseen in these two species was resolved in all other samples.
FIG. 4.—Isozyme patterns showing monomorphic Mdhp-1 locus, and hypothesized null electromorphs (N)and genotypes scored at polymorphic Mdhp-2 locus. All individuals represent samples of the culleus group.
(Fig. 3). We observed consistent expres-sion of the Est-1 locus in this sample, andan intensely stained product for the Est-2locus in all samples from the eight otherlocalities. The fact that all tissue sampleswere treated the same way, and that thesamples of T. affinis halli showed clear ex-pression of all other enzyme products,strongly suggests that the Est-2 null pat-tern in T. affinis halli is not a manifestationof enzyme degradation.
The remaining six loci were variable atthe level of within-population sample poly-morphism, and some populations are char-acterized by unique electromorphs. Whenthese six loci were tested for conformance
to Hardy-Weinberg expectations, all werefound to have genotypic ratios consistentwith Hardy-Weinberg equilibrium exceptthe Mdhp-2 locus. Examination of the iso-zyme patterns at this locus revealed manypopulation samples with a high proportionof alternative homozygotes, and no hetero-zygotes. If a null electromorph is postulat-ed again, as for Est-2 (see also Foltz, 1986;Gaffney et al., 1990; Gardner, 1992), andthe genotypes scored so that the lightbands are heterozygous for the null (Fig.4), then all the population samples but oneconform to Hardy–Weinberg expectations(the exception being T. a. globulosus, P �0.036, in the direction of heterozygote de-

42 [Vol. 58, No. 1HERPETOLOGICA
ficiency). Because the alternative interpre-tation would require postulation of ex-treme negative heterosis (selection againstthe heterozygote) at this locus only, whileall other loci conform to random matingexpectations, we retain the Mdhp-2 locusin our analysis under the assumption thata null electromorph is segregating in mostpopulations (Table 3).
Observed values of heterozygosityranged from 0.058 for pooled populationsof the T. culeus group (T. albiventris, T.culeus, and T. crawfordi, nos. 1–6 in Table3; n � 129), to 0.043 in T. m. marmoratus(n � 22), 0.038 in T. affinis halli (n � 18),and 0.00 in T. sp. nov. (n � 19). The per-centage of polymorphic loci was 4.8 for theT. culeus group, 4.0 for T. m. marmoratus,1.0 for T. affinis halli, and 0.00 in T. sp.nov. (Table 3). A test of association be-tween genotypes across all populations ofthe culeus ‘‘group’’ revealed three cases ofsignificant linkage disequilibrium (Acoh-1� Idh-2, Est-1 � Aat-1, and Aat-1 �Mdhp-2; P � 0.05); no significant associ-ation between genotypes was found forany combination of polymorphic loci in T.marmoratus.
Populations of the T. culeus group shareat least one electromorph for each of the10 variable loci; this group shares electro-morphs at eight variable loci with T. mar-moratus, five with T. affinis halli, and fivewith T. sp. nov. (Table 3). The three latterspecies share one seemingly ‘‘fixed’’ differ-ence (G3pdh–1bb) distinguishing themfrom the culeus group. Finally, T. affinishalli and T. sp. nov. are fixed for a secondunique electromorph (Mdh-2bb). At a finerlevel of resolution, T. m. marmoratus is di-agnosed by non-fixed but exclusive ( � pri-vate) electromorphs in three loci (Aat-1a,Aat-2cd, and Est-1c). Alternatively, threefixed electromorphs (Est-2nn, Acoh-2bb,Aat-2aa) and polymorphism for two otherprivate electromorphs at one locus (Mdhp-2a,b) diagnose T. affinis halli. Lastly, fixationfor unique electromorphs at two loci (Aat-1cc and Aat-2ee) diagnoses T. sp. nov.
Indices of Nei’s pairwise distance (D)and identity (I) values reflect very low lev-els of divergence between all combinationsof the samples from within Lake Titicaca
(Benavides, 1999; data not shown). For ex-ample, values of D range from 0.0–0.02within the lake, while including the threetaxa taken from outside of the lake increas-es the range of values from 0.31 (T. m.marmoratus–T. a. albiventris) to 0.67 (T.m. marmoratus–T. sp. nov.). Given the lowlevels of divergence among the recognizedtaxa in Lake Titicaca, it is not surprisingthat the six ‘‘culeus group’’ populations alsoshowed no significant allelic differentiation( 2
12 � 12.38, P � 0.41). Consequently, thevalue of � � P (Fst) was quite low (� � P� 0.0017) and the number of migrants, af-ter correction for sample size, was esti-mated as Nm � 12.29 per generation.
Allozymes: Statistical Tests of ‘‘Fixation’’The application of the Wiens–Servedio
test revealed two things, on the basis ofthe total number of characters resolved inthis study (25 loci). First, the number ofseemingly ‘‘fixed’’ characters and the meansample size per locus screened (row N inTable 3), suggests that the T. culeus group(samples 1–6 in Table 3) can be inferredto have at least one character state with afrequency �10% at a confidence level of� � 0.05 (P � 0.00063; see Appendix A ofWiens and Servedio, 2000). However, thistest does not identify which character thismight be, thus either electromorph Aat-2b
or G3pdh–1a could be diagnostic. Second,application of the same test to the threetaxa outside of the lake (T. m. marmoratus,T. affinis halli, and T. sp. nov) revealed thatsample sizes are not large enough to ruleout the possibility of alternative allelespresent at a frequency of 10% or more, inthose loci that appear diagnostic (i.e.,‘‘fixed’’ in Table 3) for each species.
Phylogenetic Analysis of AllozymeCharacters
Figure 5 shows the strict consensus ofstep-matrix maximum parsimony searchesbased on the 10 isozyme loci, and rootedalternatively with T. affinis halli or T. sp.nov. Both topologies recovered a mono-phyletic Lake Titicaca clade, albeit withweak bootstrap support, and with T. mar-moratus as the first outgroup. If this to-pology is retained with stronger support,

March 2002] 43HERPETOLOGICA
FIG. 5.—Strict consensus of 945 equally parsimo-nious trees from step-matrix weighted analysis of al-lozyme data (TL � 548, CI � 0.81, RI � 0.75); num-bers on internal nodes are bootstrap values based on1000 pseudoreplicates.
as future studies add more characters andsample other species of Telmatobius on thehigh Andean plateau, then the possibilityof nonmonophyly of the Lake Titicacaforms can be rejected with greater confi-dence.
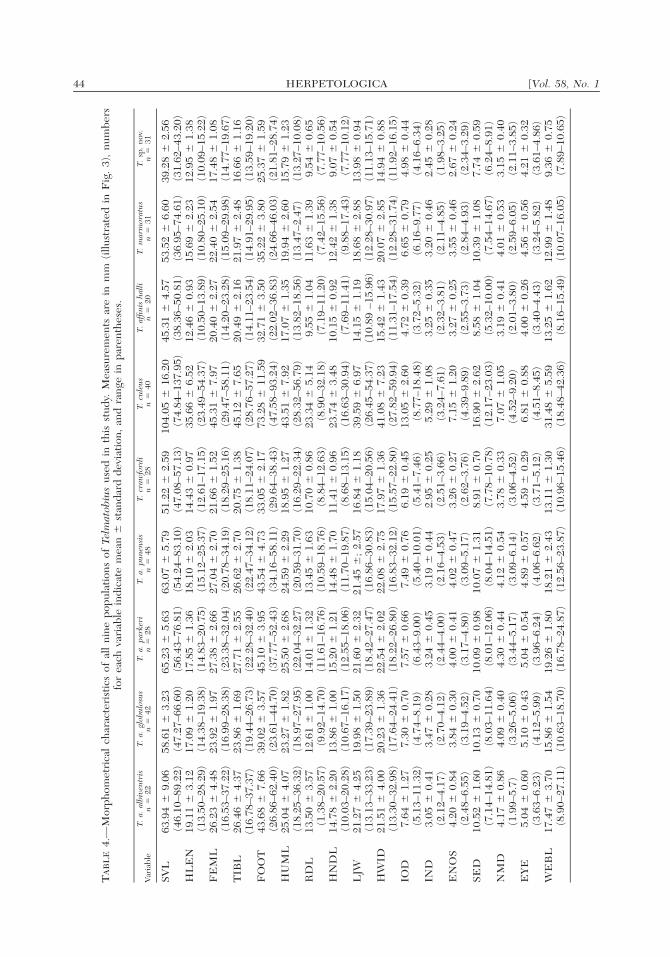
Hierarchical Analysis of MorphometricVariation
The summary statistics for all morpho-metric variables taken from the nine sam-ples of Telmatobius are shown in Table 4.The first hierarchical grouping assessedvariation among the subspecies of T. albi-ventris (T. a. albiventris, T. a. globulosus,T. a. punensis, and T. a. parkeri) by MAN-OVA, and revealed significant differenceswith non-transformed variables (Wilks’Lambda � 0.199, F51,355.087 � 5.00, P �0.001). This result is supported by a Kol-mogorov-Smirnov test for each character,which showed that only the variableENOS differed significantly from normal-ity (P � 0.01). However, homogeneity ofvariance-covariance matrices is not sup-ported by the Box M test (M � 969.3820, 2
459 � 737.2872, P � 0.0001), and a Bart-lett’s test showed homogeneity only in theEYE variable ( 2
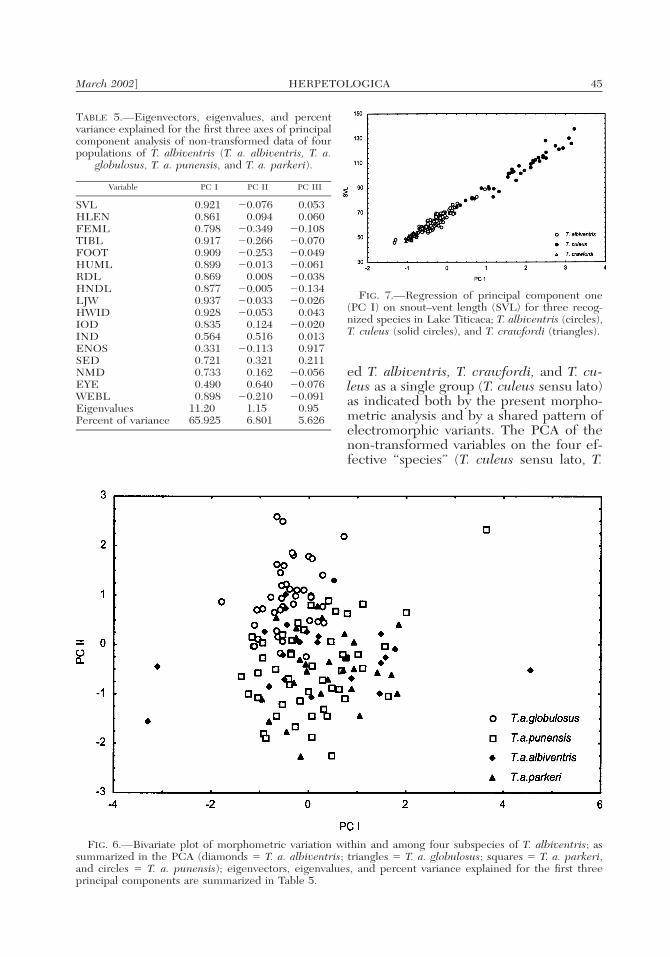
3 � 4.084, P � 0.252).Therefore the results of the PrincipalComponent Analysis should be preferred.The first three components explained78.35% of the variation of the original var-
iables (Table 5), and a scree test of the PCsindicated that only the two first compo-nents contain nontrivial information. Thefirst principal component is largely a sizeaxis, with most variables showing high pos-itive loadings, while PC II is a shape axis,with positive loadings (albeit low in manycases) for IOD, IND, SED, NMD, andEYE, and negative loadings for FEML,TIBL, FOOT, ENOS, and WEBL (Table5). All four subspecies of T. albiventrisshowed extensive overlap when scoreswere plotted in a bivariate space along PCI and PC II (Fig. 6), and were thereforegrouped together in the next round ofanalysis.
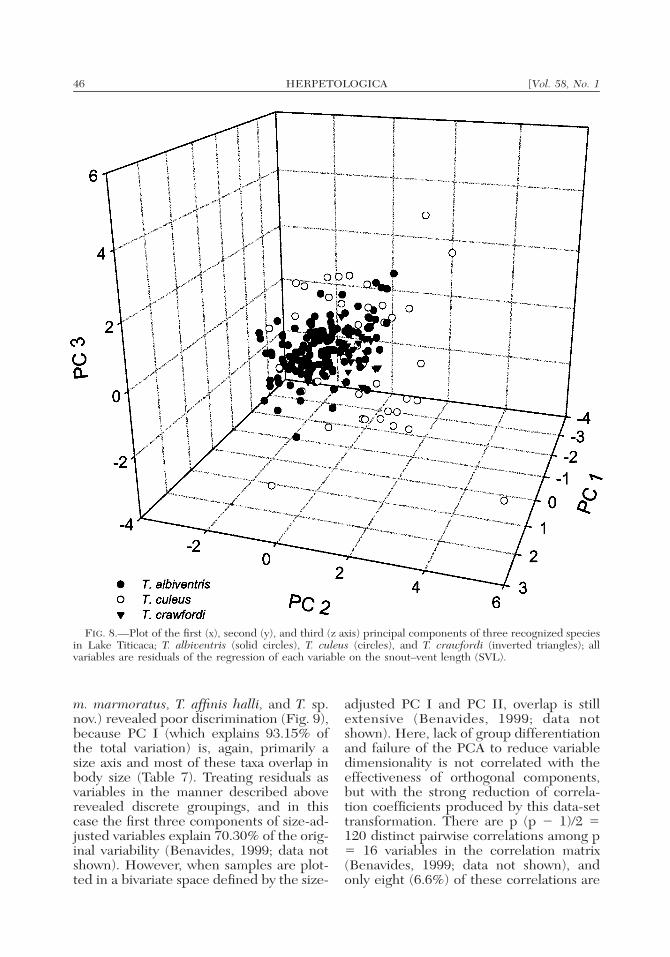
A second hierarchical grouping includedthe four subspecies of T. albiventris as asingle sample, T. crawfordi, and T. culeus.The three samples are consistently differ-ent in SVL (Kruskall-Wallis ANOVA, H �135.912, df � 2; P � 0.0001; Table 1, Fig.7). However, a preliminary bivariate ap-proach on the regression of each log-trans-formed variable against log-SVL showedheterogeneity of slopes in only four vari-ables (HUML, IOD, SED, and WEBL; P� 0.05; data not shown, see Benavides,1999), suggesting a common growth pat-tern for the remaining variables. On theother hand, variation of non-transformedvariables assessed by PCA was largely ex-plained by variation in overall body size(Fig. 8; R2 � 89.1%), which distinguishedthe three taxa. The use of residuals as var-iables eliminates the influence of SVL,while preserving the assumption of nor-mality in all but four (HLEN, FEML,HUML, and ENOS) adjusted characters.High positive loadings of FEML, TIBL,and FOOT, indicated the strong influenceof limb proportions on PC I. Positive load-ings of HLEN, LJW, and HWID, coupledwith negative loadings of FOOT andWEBL on PC II, revealed shape differ-ences (Table 6). Nevertheless, when thefirst three PCs were used to summarizemorphometric variation, all three taxa col-lapsed into a single group in multivariatespace (Fig. 8). This pattern suggests an ab-sence of major differences in proportion orshape among these taxa.
In a third hierarchical design, we treat-
44 [Vol. 58, No. 1HERPETOLOGICA
TA
BL
E4.
—M
orph
omet
rica
lch
arac
teri
stic
sof
all
nine
popu
lati
ons
ofTe
lmat
obiu
sus
edin
this
stud
y.M
easu
rem
ents
are
inm
m(i
llust
rate
din
Fig
.3)
,nu
mbe
rsfo
rea
chva
riab
lein
dica
tem
ean
�st
anda
rdde
viat
ion,
and
rang
ein
pare
nthe
ses.
Var
iabl
eT.
a.al
bive
ntri
sn
�22
T.a.
glob
ulos
usn
�42
T.a.
park
eri
n�
28T.
a.pu
nens
isn
�48
T.cr
awfo
rdi
n�
28T.
cule
usn
�40
T.af
finis
halli
n�
20T.
mar
mor
atus
n�
31T.
sp.
nov.
n�
31
SVL
HL
EN
FE
ML
63.9
4�
9.06
(46.
10–8
9.22
)19
.11
�3.
12(1
3.50
–28.
29)
26.2
3�
4.48
(16.
53–3
7.22
)
58.6
1�
3.23
(47.
27–6
6.60
)17
.09
�1.
20(1
4.38
–19.
38)
23.9
2�
1.97
(16.
99–2
8.38
)
65.2
3�
5.63
(56.
43–7
6.81
)17
.85
�1.
36(1
4.83
–20.
75)
27.3
8�
2.66
(23.
38–3
2.04
)
63.0
7�
5.79
(54.
24–8
3.10
)18
.10
�2.
03(1
5.12
–25.
37)
27.0
4�
2.70
(20.
78–3
4.19
)
51.2
2�
2.59
(47.
08–5
7.13
)14
.43
�0.
97(1
2.61
–17.
15)
21.6
6�
1.52
(18.
29–2
5.16
)
104.
05�
16.2
0(7
4.84
–137
.95)
35.6
6�
6.52
(23.
49–5
4.37
)45
.31
�7.
97(2
9.47
–58.
11)
45.3
1�
4.57
(38.
36–5
0.81
)12
.46
�0.
93(1
0.50
–13.
89)
20.4
0�
2.27
(14.
20–2
3.28
)
53.5
2�
6.60
(36.
95–7
4.61
)15
.69
�2.
23(1
0.80
–25.
10)
22.4
0�
2.54
(15.
09–2
9.98
)
39.2
8�
2.56
(31.
62–4
3.20
)12
.95
�1.
38(1
0.09
–15.
22)
17.4
8�
1.08
(14.
77–1
9.67
)T
IBL
FO
OT
HU
ML
26.4
6�
4.37
(16.
78–3
7.37
)43
.68
�7.
66(2
6.86
–62.
40)
25.0
4�
4.07
(18.
25–3
6.32
)
23.8
6�
1.69
(19.
44–2
6.73
)39
.02
�3.
57(2
3.61
–44.
70)
23.2
7�
1.82
(18.
97–2
7.95
)
27.7
1�
2.55
(22.
28–3
2.40
)45
.10
�3.
95(3
7.77
–52.
43)
25.5
0�
2.68
(22.
04–3
2.27
)
26.6
2�
2.70
(22.
47–3
4.12
)43
.54
�4.
73(3
4.16
–58.
11)
24.5
9�
2.29
(20.
59–3
1.70
)
20.7
5�
1.38
(18.
11–2
4.07
)33
.05
�2.
17(2
9.64
–38.
43)
18.9
5�
1.27
(16.
29–2
2.34
)
45.1
2�
7.65
(28.
76–5
7.27
)73
.28
�11
.59
(47.
58–9
3.24
)43
.51
�7.
92(2
8.32
–56.
79)
20.4
9�
2.16
(14.
11–2
3.54
)32
.71
�3.
50(2
2.02
–36.
83)
17.0
7�
1.35
(13.
82–1
8.56
)
21.9
7�
2.48
(14.
91–2
9.95
)35
.22
�3.
80(2
4.66
–46.
03)
19.9
4�
2.60
(13.
47–2
.47)
16.6
6�
1.16
(13.
59–1
9.20
)25
.37
�1.
59(2
1.81
–28.
74)
15.7
9�
1.23
(13.
27–1
0.08
)R
DL
HN
DL
LJW
13.5
0�
3.57
(1.3
8–20
.57)
14.7
8�
2.20
(10.
03–2
0.28
)21
.27
�4.
25(1
3.13
–33.
23)
12.6
1�
1.00
(9.9
2–14
.70)
13.8
6�
1.00
(10.
67–1
6.17
)19
.98
�1.
50(1
7.39
–23.
89)
14.0
1�
1.32
(11.
61–1
6.76
)15
.20
�1.
21(1
2.55
–18.
06)
21.6
0�
2.32
(18.
42–2
7.47
)
13.4
5�
1.63
(10.
59–1
8.76
)14
.48
�1.
70(1
1.70
–19.
87)
21.4
5�
;2.
57(1
6.86
–30.
83)
10.7
0�
0.86
(8.8
4–12
.63)
11.4
1�
0.96
(8.6
8–13
.15)
16.8
4�
1.18
(15.
04–2
0.56
)
23.3
4�
5.14
(8.9
0–32
.18)
23.7
4�
3.48
(16.
63–3
0.94
)39
.59
�6.
97(2
6.45
–54.
37)
9.55
�1.
04(7
.19–
11.2
0)10
.15
�0.
92(7
.69–
11.4
1)14
.15
�1.
19(1
0.89
–15.
96)
11.6
3�
1.39
(7.4
2–15
.56)
12.4
2�
1.38
(9.8
8–17
.43)
18.6
8�
2.88
(12.
28–3
0.97
)
9.54
�0.
65(7
.77–
10.5
6)9.
07�
0.54
(7.7
7–10
.12)
13.9
8�
0.94
(11.
13–1
5.71
)H
WID
IOD
IND
21.5
1�
4.00
(13.
30–3
2.98
)7.
64�
1.27
(5.1
3–11
.32)
3.05
�0.
41(2
.12–
4.17
)
20.2
3�
1.36
(17.
64–2
4.41
)7.
30�
0.70
(4.7
4–8.
19)
3.47
�0.
28(2
.70–
4.12
)
22.5
4�
2.02
(18.
22–2
6.80
)7.
57�
0.66
(6.4
3–9.
00)
3.24
�0.
45(2
.44–
4.00
)
22.0
8�
2.75
(16.
83–3
2.12
)7.
49�
0.76
(5.4
0–10
.01)
3.19
�0.
44(2
.16–
4.53
)
17.9
7�
1.36
(15.
57–2
2.80
)6.
19�
0.45
(5.4
1–7.
46)
2.95
�0.
25(2
.51–
3.66
)
41.0
8�
7.23
(27.
82–5
9.94
)13
.05
�2.
60(8
.77–
18.4
8)5.
29�
1.08
(3.2
4–7.
61)
15.4
2�
1.43
(11.
31–1
7.54
)4.
72�
0.39
(3.7
2–5.
32)
3.25
�0.
35(2
.32–
3.81
)
20.0
7�
2.85
(12.
28–3
1.74
)6.
65�
0.79
(6.1
6–9.
77)
3.20
�0.
46(2
.11–
4.85
)
14.9
4�
0.88
(11.
92–1
6.15
)4.
98�
0.44
(4.1
6–6.
34)
2.45
�0.
28(1
.98–
3.25
)E
NO
S
SED
NM
D
4.20
�0.
84(2
.48–
6.55
)10
.52
�1.
60(7
.14–
14.8
1)4.
17�
0.86
(1.9
9–5.
7)
3.84
�0.
30(3
.19–
4.52
)10
.13
�0.
70(8
.03–
11.6
4)4.
09�
0.40
(3.2
6–5.
06)
4.00
�0.
41(3
.17–
4.80
)10
.09
�0.
98(8
.01–
12.0
6)4.
30�
0.44
(3.4
4–5.
17)
4.02
�0.
47(3
.09–
5.17
)10
.07
�1.
31(8
.04–
14.5
1)4.
12�
0.54
(3.0
9–6.
14)
3.26
�0.
27(2
.62–
3.76
)8.
91�
0.70
(7.7
8–10
.78)
3.78
�0.
33(3
.06–
4.52
)
7.15
�1.
20(4
.39–
9.89
)16
.90
�2.
62(1
2.17
–23.
03)
7.07
�1.
05(4
.52–
9.20
)
3.27
�0.
25(2
.55–
3.73
)8.
58�
1.04
(5.3
2–10
.00)
3.19
�0.
41(2
.01–
3.80
)
3.55
�0.
46(2
.84–
4.93
)10
.39
�1.
08(7
.54–
14.6
7)4.
01�
0.53
(2.5
9–6.
05)
2.67
�0.
24(2
.34–
3.29
)7.
74�
0.59
(6.2
4–8.
91)
3.15
�0.
40(2
.11–
3.85
)E
YE
WE
BL
5.04
�0.
60(3
.63–
6.23
)17
.47
�3.
70(8
.90–
27.1
1)
5.10
�0.
43(4
.12–
5.99
)15
.86
�1.
54(1
0.63
–18.
70)
5.04
�0.
54(3
.96–
6.24
)19
.26
�1.
80(1
6.78
–24.
87)
4.89
�0.
57(4
.06–
6.62
)18
.21
�2.
43(1
2.56
–23.
87)
4.59
�0.
29(3
.71–
5.12
)13
.11
�1.
30(1
0.96
–15.
46)
6.81
�0.
88(4
.51–
8.45
)31
.48
�5.
59(1
8.48
–42.
36)
4.00
�0.
26(3
.40–
4.43
)13
.25
�1.
62(8
.16–
15.4
9)
4.56
�0.
56(3
.24–
5.82
)12
.99
�1.
48(1
0.07
–16.
05)
4.21
�0.
32(3
.61–
4.86
)9.
36�
0.75
(7.8
9–10
.65)
March 2002] 45HERPETOLOGICA
TABLE 5.—Eigenvectors, eigenvalues, and percentvariance explained for the first three axes of principalcomponent analysis of non-transformed data of fourpopulations of T. albiventris (T. a. albiventris, T. a.
globulosus, T. a. punensis, and T. a. parkeri).
Variable PC I PC II PC III
SVLHLENFEMLTIBLFOOT
0.9210.8610.7980.9170.909
�0.0760.094
�0.349�0.266�0.253
0.0530.060
�0.108�0.070�0.049
HUMLRDLHNDLLJWHWID
0.8990.8690.8770.9370.928
�0.0130.008
�0.005�0.033�0.053
�0.061�0.038�0.134�0.026
0.043IODINDENOSSEDNMD
0.8350.5640.3310.7210.733
0.1240.516
�0.1130.3210.162
�0.0200.0130.9170.211
�0.056EYEWEBLEigenvaluesPercent of variance
0.4900.898
11.2065.925
0.640�0.210
1.156.801
�0.076�0.091
0.955.626
FIG. 7.—Regression of principal component one(PC I) on snout–vent length (SVL) for three recog-nized species in Lake Titicaca; T. albiventris (circles),T. culeus (solid circles), and T. crawfordi (triangles).
FIG. 6.—Bivariate plot of morphometric variation within and among four subspecies of T. albiventris; assummarized in the PCA (diamonds � T. a. albiventris; triangles � T. a. globulosus; squares � T. a. parkeri,and circles � T. a. punensis); eigenvectors, eigenvalues, and percent variance explained for the first threeprincipal components are summarized in Table 5.
ed T. albiventris, T. crawfordi, and T. cu-leus as a single group (T. culeus sensu lato)as indicated both by the present morpho-metric analysis and by a shared pattern ofelectromorphic variants. The PCA of thenon-transformed variables on the four ef-fective ‘‘species’’ (T. culeus sensu lato, T.
46 [Vol. 58, No. 1HERPETOLOGICA
FIG. 8.—Plot of the first (x), second (y), and third (z axis) principal components of three recognized speciesin Lake Titicaca; T. albiventris (solid circles), T. culeus (circles), and T. crawfordi (inverted triangles); allvariables are residuals of the regression of each variable on the snout–vent length (SVL).
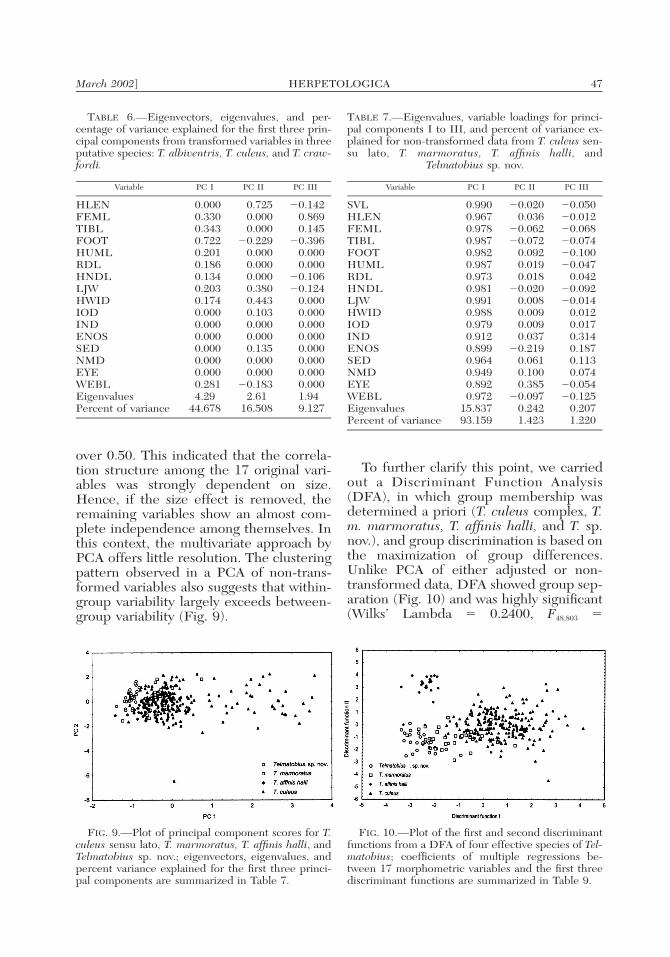
m. marmoratus, T. affinis halli, and T. sp.nov.) revealed poor discrimination (Fig. 9),because PC I (which explains 93.15% ofthe total variation) is, again, primarily asize axis and most of these taxa overlap inbody size (Table 7). Treating residuals asvariables in the manner described aboverevealed discrete groupings, and in thiscase the first three components of size-ad-justed variables explain 70.30% of the orig-inal variability (Benavides, 1999; data notshown). However, when samples are plot-ted in a bivariate space defined by the size-
adjusted PC I and PC II, overlap is stillextensive (Benavides, 1999; data notshown). Here, lack of group differentiationand failure of the PCA to reduce variabledimensionality is not correlated with theeffectiveness of orthogonal components,but with the strong reduction of correla-tion coefficients produced by this data-settransformation. There are p (p � 1)/2 �120 distinct pairwise correlations among p� 16 variables in the correlation matrix(Benavides, 1999; data not shown), andonly eight (6.6%) of these correlations are
March 2002] 47HERPETOLOGICA
TABLE 6.—Eigenvectors, eigenvalues, and per-centage of variance explained for the first three prin-cipal components from transformed variables in threeputative species: T. albiventris, T. culeus, and T. craw-fordi.
Variable PC I PC II PC III
HLENFEMLTIBLFOOTHUML
0.0000.3300.3430.7220.201
0.7250.0000.000
�0.2290.000
�0.1420.8690.145
�0.3960.000
RDLHNDLLJWHWIDIOD
0.1860.1340.2030.1740.000
0.0000.0000.3800.4430.103
0.000�0.106�0.124
0.0000.000
INDENOSSEDNMDEYE
0.0000.0000.0000.0000.000
0.0000.0000.1350.0000.000
0.0000.0000.0000.0000.000
WEBLEigenvaluesPercent of variance
0.2814.29
44.678
�0.1832.61
16.508
0.0001.949.127
TABLE 7.—Eigenvalues, variable loadings for princi-pal components I to III, and percent of variance ex-plained for non-transformed data from T. culeus sen-su lato, T. marmoratus, T. affinis halli, and
Telmatobius sp. nov.
Variable PC I PC II PC III
SVLHLENFEMLTIBLFOOT
0.9900.9670.9780.9870.982
�0.0200.036
�0.062�0.072
0.092
�0.050�0.012�0.068�0.074�0.100
HUMLRDLHNDLLJWHWID
0.9870.9730.9810.9910.988
0.0190.018
�0.0200.0080.009
�0.0470.042
�0.092�0.014
0.012IODINDENOSSEDNMD
0.9790.9120.8990.9640.949
0.0090.037
�0.2190.0610.100
0.0170.3140.1870.1130.074
EYEWEBLEigenvaluesPercent of variance
0.8920.972
15.83793.159
0.385�0.097
0.2421.423
�0.054�0.125
0.2071.220
FIG. 9.—Plot of principal component scores for T.culeus sensu lato, T. marmoratus, T. affinis halli, andTelmatobius sp. nov.; eigenvectors, eigenvalues, andpercent variance explained for the first three princi-pal components are summarized in Table 7.
FIG. 10.—Plot of the first and second discriminantfunctions from a DFA of four effective species of Tel-matobius; coefficients of multiple regressions be-tween 17 morphometric variables and the first threediscriminant functions are summarized in Table 9.
over 0.50. This indicated that the correla-tion structure among the 17 original vari-ables was strongly dependent on size.Hence, if the size effect is removed, theremaining variables show an almost com-plete independence among themselves. Inthis context, the multivariate approach byPCA offers little resolution. The clusteringpattern observed in a PCA of non-trans-formed variables also suggests that within-group variability largely exceeds between-group variability (Fig. 9).
To further clarify this point, we carriedout a Discriminant Function Analysis(DFA), in which group membership wasdetermined a priori (T. culeus complex, T.m. marmoratus, T. affinis halli, and T. sp.nov.), and group discrimination is based onthe maximization of group differences.Unlike PCA of either adjusted or non-transformed data, DFA showed group sep-aration (Fig. 10) and was highly significant(Wilks’ Lambda � 0.2400, F48,803 �

48 [Vol. 58, No. 1HERPETOLOGICA
TABLE 8.—Discriminating analysis classification matrix for non-transformed data of four effectivespecies of Telmatobius.
Actual speciesNumberof cases
% classifiedcorrectly
Predicted classification
1 2 3 4
1. T. sp. nov.2. T. m. marmoratus3. T. affinis halli4. T. culeus
313120
208
100.000074.193695.000097.5962
31111
02303
00
191
070
203
TABLE 9.—Coefficients from multiple regressions be-tween each of the first three discriminant functions(DF) and 17 morphometric variables from four ef-fective ‘‘species’’ of Telmatobius. Loadings for char-acters with the highest discriminatory power for each
DF are typed in bold face.
Variable DF I DF II DF III
SVLHLENFEMLTIBLFOOT
3.64758�1.18222
0.05262�1.20847
0.79378
0.314930.92335
�0.211151.483041.45396
�2.755192.351550.200480.68699
�1.05496HUMLRDLHNDLLJWHWID
�0.37061�1.07674
1.33202�0.17001
�1.78716
�0.81803�1.63040�0.97730�0.55040�1.78262
2.470790.40403
�1.31428�0.22812�1.24335
IODINDENOSSED
1.79383�0.71357�0.07465�1.14034
�2.052931.897040.238650.00576
0.11156�0.37643
0.28968�2.04936
NMDEYEWEBL
0.187220.39217
�0.08126
�0.37437�0.029342.31233
0.057400.686521.87503
10.30992, P � 0.0001). Discriminationfunctions were most successful in correctlyassigning individuals to their groups basedon non-transformed data. Despite largewithin-group variability, misclassificationof T. culeus sensu lato occurs in less than3.0% of the cases, whereas T. m. marmor-atus was more often wrongly classified ei-ther as T. culeus (22.5%) or T. sp. nov.(3.22%). Erroneous classification was non-existent in T. sp. nov. and low in T. affinishalli (5.0%; Table 8). Although DFA is ana posteriori classification procedure, it nev-ertheless provides an indication of the ex-tent of variable influence on group dis-crimination. Table 9 summarizes regres-sion coefficients of each variable on thefirst three discriminant functions, suggest-ing that SVL, HWID, and IOD are themost effective variables in discriminatinggroups along DF I, and RDL, HWID,
IOD, and WEBL along DF II. It is note-worthy that although DF I is strongly in-fluenced by body size differences amongtaxa (regression coefficient � 3.647), load-ings from DFA calculated for DF I andDF II had positive and negative values, in-dicating that both axes summarize shaperather than size discrimination alone (Ta-ble 9). Most importantly, our results showthat even though overlap is considerableamong some groups, each species occupiesa fairly well defined area in the discrimi-nant space of 17 measured variables (Fig.10).
DISCUSSION
Species Diagnosis and Statistical Tests
The main objective of this study was totest taxonomic boundaries of the recog-nized subspecies of T. albiventris, and oth-er species within and outside of the LakeTiticaca Basin. The absence of either iso-zyme or morphometric characters for dis-criminating among the named subspeciesof T. albiventris was anticipated, but thelack of discrimination between this and theother two species from Lake Titicaca wasunexpected. This is because differences inadult body sizes would appear to providea mechanism for reproductive isolation, atleast between the large T. culeus (averageSVL � 104.5 mm), and the smaller T.crawfordi (average SVL � 51.22 mm) andT. albiventris (average SVL � 62.71 mm).However, as long as allopatry and verticalisolation along depth gradients cannot besafely argued, and estimates of populationstructure reveal high gene flow (popula-tions could be interconnected by gene flowaround the lake’s perimeter, and along riv-er courses), we conclude that the threeformer species conform to a single evolu-tionary lineage in Lake Titicaca. This con-

March 2002] 49HERPETOLOGICA
clusion is conditional given several caveats(see next section), but further study de-signed to disentangle historical from on-going demographic components of popu-lation structure will ultimately be neededto clarify this point.
At the most inclusive hierarchical level,both allozyme and morphometric dataprovide corroborating evidence for diag-nostic differences among at least four spe-cies: the T. culeus group, T. m. marmora-tus, T. affinis halli, and Telmatobius sp.nov. However, Wiens (1999) has revisitedthe issue of species delimitation and char-acter polymorphism, and he pointed outthat unless ‘‘fixed’’ characters are surveyedfrom the entire population, they cannot beconsidered as truly invariant characters.This is one of the requirements of the PAAaggregation method (Davis and Nixon,1992), and as shown by Wiens and Ser-vedio (2000), determining absolute fixationof a character with a conventional level ofstatistical confidence (� � 0.05) is almostunattainable. Our data suggest that usinga frequency cutoff of 10%, while maintain-ing the � � 0.05 degree of statistical con-fidence, will not be feasible in empiricalstudies of populations characterized by lowlevels of character divergence. In this case,only a single locus of 25 screened was‘‘fixed’’ between the Lake Titicaca samplesand T. marmoratus (Table 3), and our sam-ple sizes of both characters and specimens(Table 1) are probably within the range ofthose used in most conventional studies ofgeographic variation, metapopulationstructure, and species boundaries. How-ever, all but the largest sample (the com-bined T. culeus group; n � 127) are toosmall to achieve conventional statisticalpower, primarily because the Wiens-Ser-vedio test is sensitive to both sample sizesand the number of fixed character differ-ences (sample sizes of 25 individuals areadequate if populations are distinguishedby at least seven fixed differences; Wiensand Servedio, 2000). We stress that thislimitation is not restricted to the Wiens-Servedio test, because recently derivedspecies will be the hardest to diagnose byany criterion (Avise, 2000).
Beyond the sampling limitation, T. mar-
moratus can be unambiguously separatedfrom all samples of culeus in a nonstatis-tical sense by a single diagnostic locus(Aat-2cd), as well as two private, low fre-quency electromorphs at two other loci(Est-1c and Aat-1a). Differentiation is evenstronger for T. affinis halli and T. sp. nov.by the criteria of character diagnosabilityand allopatry. Telmatobius sp. nov. is di-agnosed by unique ‘‘fixed’’ electromorphsat Aat-1cc and Aat-2dd, while T. affinis halliis ‘‘fixed’’ for unique electromorphs at Est-2nn, Acoh-2bb, Aat-2aa, and segregates forthe unique electromorphs Mdhp-2ab. Tel-matobius marmoratus, T. affinis halli, andT. sp. nov. can therefore be unambiguouslydiagnosed on the basis of unique concor-dant allozyme character states. Thus, con-cordance among presumably independentmarkers still can be taken as strong evi-dence of long-term isolation (i.e., sufficienttime for these lineages to accumulate nov-el electromorphs; Avise, 2000). We rec-ognize that in terms of statistical power, wecannot make a strong case that these elec-tromorphs are strictly unique to the spe-cies that they appear to diagnose (i.e., theymight be present at low frequencies inother species), but the evidence providedby allozymes is also supported by the as-sessment of morphometric variability. TheDFA identified the same four groups ofTelmatobius as those identified by diag-nostic electromorphs (Fig. 10), and inmost studies based on modest sample siz-es, evidence from concordant patterns ofvariation across different data sets willcontinue to provide a valid assessment ofspecies boundaries.
Telmatobius in Lake Titicaca: NichePolymorphism and Ecological Speciation
Although we interpret three namedforms within Lake Titicaca Basin as con-specific, the differentiation in body sizemorphs along a depth gradient (Table 1)appears to be similar to that for severalfreshwater fish species flocks, in whichecological and trophic specializations cor-relate strongly with phenotypic diver-gence, but not genetic structure (Lu andBernatchez, 1999; Nagel and Schluter,1998; Rundle et al., 2000). Many of these

50 [Vol. 58, No. 1HERPETOLOGICA
examples in fishes provide evidence forecologically driven adaptive divergence, inwhich phenotypic divergence among pop-ulations within a lake is promoted by re-laxed resource competition and use of al-ternative niches (Lu and Bernatchez,1999; Schluter, 1998). If reproductive iso-lation follows as a consequence of thesame forces causing phenotypic and eco-logical divergence, ecological speciationmay result in the absence of allopatry(Schluter 1996a,b).
In other words, resource-based diver-gent natural selection may play a majorrole in the evolution of reproductive iso-lation over very short geographic distances(Schluter, 1998), and recent empirical ev-idence has also suggested that this kind ofdivergence, or life history shift, may occurover extremely short time frames (Bridleand Jiggins, 2000; Orr and Smith, 1998;Reznick et al., 1990). Moreover, severalkey predictions dealing with the model ofsympatric speciation have been successful-ly tested. First, phenotypic differencesmay accumulate between populations indifferent but adjoining habitats despitecontinued gene flow (Blondel et al., 1999;Lu and Bernatchez, 1999; Schneider et al.,1999; Smith et al., 1997). Second, distinctecotypes do have differential fitness andhybrids often show reduced fitness and vi-ability (Nagel and Schluter, 1995; Schluter,1993). Finally, assortative mating may leadto reinforcement mechanisms among eco-logically differentiated populations in theface of gene flow (Nagel and Schluter,1995; Saetre et al., 1997).
The pattern of widely shared electro-morph frequencies among Telmatobius‘‘ecotypes’’ in Lake Titicaca Basin, whichwe take here as evidence of interconnec-tedness by gene flow, is also consistentwith the hypothesis of a recent origin andongoing ecological speciation among thesepopulations. Speciation in Telmatobius haslong been regarded as a rapid adaptiveprocess, facilitated by colonization of newand unoccupied niches in the recently up-lifted Andes (Cei, 1986; Lynch, 1986).However, the mechanisms underlying theprocesses of speciation by Telmatobius arenot well understood, and current Andean
amphibian diversity cannot be predictedfrom a purely geographic perspective(Lynch, 1986). However, two lines of evi-dence seem to support, the possibility ofdifferential niche use (Skulason and Smith,1995; Smith and Skulason, 1996) by adultanurans, and ongoing processes of ecolog-ical speciation.
First, the Titicaca Basin is geologicallyrelatively young, on the order of about 3.0� 106 yr old (Dejoux, 1994; Lavenu, 1981),and Lake Titicaca has had a very recenthistory of dramatic oscillations in waterand salinity-levels (approximately the last20,000 yr; Mourguiart et al., 1995a,b;Wirrmann et al., 1991). Such changes havepromoted several widespread extinctionand recolonization events, as shown by de-tailed palynological evidence (Dejoux,1994; Wirrmann et al., 1991) and by theactual composition of the aquatic macro-flora in Lake Titicaca when compared tothe rest of the basin (Ybert, 1991). Massivecyclic turnovers in the lake macrophytecommunity were likely to have heavily im-pacted the animal communities, and mayhave contributed to repeated extinctionand recolonization events of unoccupiedniches; the most recent such turnover mayhave occurred as recently as 3000 yr ago(Ybert, 1991). In parallel to Telmatobius,the native fish genus Orestias Valenci-ennes, 1839 (Cyprinodontiformes: Cy-pronodontidae) displays the very same pat-tern of extensive phenotypic plasticity(Villwock, 1986) and differential habitatuse (littoral, pelagic, and benthic; Lauzan-ne, 1991). Furthermore, the taxonomy ofthe species in Lake Titicaca is controver-sial, and the lake is unexpectedly rich interms of number of fish species (Lauzan-ne, 1982, 1991, recognized 24 specieswhile Parenti, 1984, recognized 28).
Second, regardless of the absolute ageof the T. culeus group, we think that theconditions described above for the ‘‘re-source-based divergence’’ version of eco-logical speciation (Schluter 1996a,b) maybe met by the ecotypes of Telmatobius inLake Titicaca. The large frogs are confinedto deep water (they are collected by fish-ermen with nets), while the early-matur-ing, small-bodied morphs are restricted to

March 2002] 51HERPETOLOGICA
shallow embayments (Table 1). This is cor-relational evidence only, and given thatsome amphibians are characterized by dis-assortative mating, and virtually all haveindeterminate growth (Duellman andTrueb, 1986; Castellano and Giacoma,1998), such a pattern could be explainedby two non-exclusive alternatives. If ourinterpretation of conspecificity of the LakeTiticaca populations is correct, then T. cu-leus is characterized by extreme phenotyp-ic plasticity and niche polymorphism (Sku-lason and Smith, 1995; Smith and Skula-son, 1996), with low heritability for bodysize. This hypothesis can be tested by com-mon garden experiments designed to es-timate heritabilities of different body sizes,and studies of ecological performance ofthe morphotypes and their intermediates,in different microhabitats. One circum-stantial line of evidence, suggesting highheritability of body size, is the recent re-discovery of a large-bodied T. marmoratusgigas (holotype SVL � 109.0 mm) in veryshallow streams 150 km south of Lake Ti-ticaca (de la Riva, personal communica-tion). This taxon is a probable relict froman ancient lake, and its restriction to shal-low streams suggests that gigantism is notreadily lost despite a shift to dramaticallydifferent ecological conditions.
Another possibility is that the differentsize morphs of T. culeus represent inde-pendent colonization events into Lake Ti-ticaca by previously differentiated lineag-es, in which case the present niche differ-ences are unrelated to in situ adaptive dif-ferentiation among conspecific populationsalong resource gradients. This interpreta-tion, however, would require sufficientpost-colonization hybridization and intro-gression to homogenize electromorph fre-quencies across all ecotypes. This hypoth-esis could be falsified by a phylogeneticstudy demonstrating unambiguous mono-phyly of the Lake Titicaca populations andextreme recency of the clade’s origin with-in the lake (i.e., corroboration of the pat-tern shown in Fig. 5). Populations cur-rently inhabiting the lake could yet rep-resent a monophyletic assemblage if allpreviously differentiated forms themselvesformed a clade prior to colonization of
Lake Titicaca, but in this case we wouldexpect some evidence for older allopatricdivergence in taxa outside of the lake.
At a more general level, the study ofspeciation processes and patterns ultimate-ly centers on disentangling evidence basedon correlation from that based on causa-tion. Under the assumptions of habitat var-iability, open niches, relaxation of inter-specific competition, and strong selection(Smith and Skulason, 1996), equivalentevolutionary outcomes, or parallel specia-tion, has been cited as the strongest cor-relational argument of speciation by ad-aptation or habitat shift (Losos et al., 1998;Rundle et al., 2000; Schluter, 1996a,b). Inthis context, studies by Sinsch and Juraske(1995) and Sinsch et al. (1995) are relevantto this work. These investigators taxonom-ically reassessed the two recognized spe-cies of the genus Batrachophrynus (B.brachydactylus and B. macrostomus) andshowed that significant differences in hab-itat between the small stream-dwelling B.brachydactylus (SVL � 56.9; n � 53) andthe larger lake-dwelling B. macrostomus(SVL � 131.5; n � 13) are positively cor-related with morphometric differentiation(Sinsch et al., 1995). However, an allozymesurvey revealed the lowest pairwise genet-ic distance ever recorded for two recog-nized species of amphibians [Nei (1978) D� 0.031; Sinsch and Juraske, 1995]. Fur-thermore, both species of Batrachophry-nus can occur in sympatry and apparentlydo not interbreed (Sinsch and Juraske,1995). Whether this example of microgeo-graphic phenotypic differentiation is adap-tive remains to be demonstrated (but seeBlondel et al., 1999; Lambrechts et al.,1997; Schilthuizen, 2000), but it does sug-gest a common pattern of recent morpho-logical differentiation in high Andean pop-ulations of telmatobiine frogs (for anotherpossible example, see Laurent, 1973, 1977,and Cei, 1986).
In summary, although the paradigm ofPleistocene changes and speciation hasbeen taken as the most general model ofspeciation in many groups, many lines ofevidence are suggesting that the origin ofliving species may either pre-date thePleistocene (Klicka and Zink, 1997) or be

52 [Vol. 58, No. 1HERPETOLOGICA
due to recent divergence along ecologicalgradients (Schilthuizen, 2000; Schneideret al., 1999; Smith et al., 1997). We suggestthat a relatively continuous history of pre-and post-Pleistocene speciation for Tel-matobius, based both on ecological oppor-tunity and vicariant events, may better ex-plain the radiation of the genus in the highAndes.
RESUMEN
Las diversidad especıfica del genero Tel-matobius es el resultado de una radiacionadaptativa que actualmente alcanza a 49especies, confinadas en ambientes acuati-cos de altura en la Cordillera de los Andes.El genero carece de una hipotesis filoge-netica que sustente su monofilia y en gen-eral, las relaciones y lımites interespecıfi-cos han sido escasamente abordados. Eneste trabajo, colectamos las cuatro subes-pecies reconocidas de T. albiventris delLago Titicaca, mas ejemplares asignados aT. culeus (Lago Titicaca), T. crawfordi y T.marmoratus de la cuenca circundante. Es-tas poblaciones han sido comparadas me-diante caracteres aloenzimaticos y a travesde un estudio multivariado de variablesmorfometricas para determinar su estatusespecıfico. La delimitacion propuesta estabasada en la utilizacion de un concepto ev-olutivo de especie y en el uso de un cri-terio de concordancia entre caracteres in-dependientes como metodo explıcito dereconocimiento de especies evolutivas.Nuestros resultados fundamentan el re-conocimiento de cuatro especies evoluti-vas; T. culeus (que incluye a T. albiventrisy T. crawfordi), T. marmoratus, T. affinishalli y Telmatobius sp. nov. Los cuatro taxaanteriores pueden ser discriminados porcombinaciones unicas de caracteres mor-fometricos y aloenzimaticos. Sin embargo,solo T. culeus cuenta con un tamano mues-tral suficiente en relacion al numero decaracteres fijos como para evaluar con ungrado de confianza significativo (� � 0.05)los caracteres diagnostico propuestos. Lostres taxa considerados como parte de unasola especie evolutiva muestran diferenciassignificativas en la talla corporal y micro-habitat que pueden ser interpretadas comoevidencia circunstancial de adaptacion a
un gradiente ecologico continuo. Final-mente, se discute la posibilidad de un fen-omeno de especiacion ecologica y las pred-icciones que deben ser testeadas paracomprobar tal hipotesis.
Acknowledgments.—The first author is indebted toJ. H. Cordoba at the Museo de Historia NaturalUniversidad Nacional Mayor de San Marcos(MHNSM); J. Aparicio, M. E. Perez, and C. Cortezat the Coleccion Boliviana de Fauna (CBF); P. Riffo,M. Contreras, and E. Salazar from the Museo deZoologıa de la Universidad de Concepcion (MZUC);E. Lavilla, at the Instituto ‘‘Miguel Lillo’’ of Tucumanprovided rare literature and gave us thoughtful in-sights. J. Wiens of the Carnegie Museum and J. Fetz-ner at BYU made crucial contributions. K. Crandall(BYU) facilitated this work. We also appreciated com-ments on this paper from D. Adams (Iowa State Uni-versity). For assistance in field trips, the first authoris grateful to E. Benavides, H. Salazar, N. Arapa andson, and several anonymous fishermen. Dr. L. Lau-zanne at ORSTOM-BOLIVIA kindly helped withtransportation and logistics. Financial support wasprovided by FONDECYT sectorial BIOMA n�5960021.
LITERATURE CITED
AGASSIZ, A., AND S. W. GARMAN. 1876. Explorationof Lake Titicaca. Bulletin of the Museum of Com-parative Zoology at Harvard College 3:273–349.
ANGEL, F. 1926. Sur un batracien nouveau du Perouappartennat au genre Telmatobius. Annales des Sci-ences Naturelles-X-Zoologie VI:107–111
ARNTZEN, J. W., AND M. GARCıA-PARIS. 1995. Mor-phological and allozyme studies of midwife toads(genus Alytes), including the description of twonew taxa from Spain. Contributions to Zoology 65:5–34.
AVISE, J. C. 2000. Phylogeography: the History andFormation of Species. Harvard University Press,Cambridge, Massachusetts, U.S.A.
AVISE, J. C., AND R. M. BALL. 1990. Principles ofgenealogical concordance in species concepts andbiological taxonomy. Oxford Surveys in Evolution-ary Biology 7:45–67.
AVISE, J. C., AND K. WOLLENBERG. 1997. Phyloge-netics and the origin of species. Proceedings of theNational Academy of Sciences USA 94:7748–7755.
BARTON, N. H., AND M. SLATKIN. 1986. A quasi equi-librium theory of the distribution of rare alleles ina subdivided population. Heredity 56:409–415.
BENAVIDES, E. 1999. Situacion taxonomica de Tel-matobius albiventris (Anura: Leptodactylidae) ycomplejos relacionados. Un enfoque morfologico ymolecular. M. S. Thesis, Universidad de Concep-cion, Concepcion, Chile.
BLONDEL, J., P. C. DIAS, P. PERRET, M. MAISTRE,AND M. LAMBRECHTS. 1999. Selection-based bio-diversity at a small spatial scale in a low-dispersinginsular bird. Science 285:1399–1402.
BRIDLE, J. R., AND D. JIGGINS. 2000. Adaptive dy-

March 2002] 53HERPETOLOGICA
namics: is speciation too easy? Trends in Ecologyand Evolution 15:225–226.
BURNABY, T. P. 1966. Growth-invariant discriminantfunctions and generalized distances. Biometrics 22:96–110.
CASTELLANO, S., AND C. GIACOMA. 1998. Morpho-logical variation of the green toad Bufo viridis, inItaly: a test of causation. Journal of Herpetology 32:540–550.
CEI, J. M. 1986. Speciation and adaptive radiation inAndean Telmatobius frogs. Pp. 374–386. In F. Vuil-leumier and M. Monasterio (Eds.), High AltitudeTropical Biology. Oxford University Press, Oxford,U.K.
DAVIS, J. I., AND K. C. NIXON. 1992. Populations,genetic variation, and the delimitation of phyloge-netic species. Systematic Biology 41:421–435.
DA SILVA, N. J., AND J. W. SITES, JR. 2001. Phylogenyof South America triad coral snakes (Elapidae; Mi-crurus) based on molecular characters. Herpeto-logica 57:1–22.
DEJOUX, C. 1994. Lake Titicaca. Speciation in ancientlakes. Schweizerbartsche verlagsbuchhandl, Stutt-gart 44:35–42.
DE LA RIVA, I. 1994. Description of a new species ofTelmatobius from Bolivia (Amphibia: Anura: Lep-todactylidae). Graellsia 50:161–164.
DE QUEIROZ, K. 1998. The general lineage conceptof species, species criteria, and the process of spe-ciation: a conceptual unification and terminologicalrecommendation. Pp. 57–75. In D. Howard and S.H. Berlocher (Eds.), Endless Forms: Species andSpeciation. Oxford University Press, New York,New York, U.S.A.
D’ORBIGNY, A. 1835–1847. Voyages dans l’AmeriqueMeridionale. Pitois-Levrault et Cie., Paris, France.
DUELLMAN, W. E., AND L. TRUEB. 1986. Biology ofAmphibians. McGraw-Hill Book Company, NewYork, New York, U.S.A.
DUMERIL, A. M. C., AND G. BIBRON. 1841. Erpeto-logie Generale ou Histoire Naturelle Complete desReptiles. Roret, Paris, France.
FOLTZ, D. W. 1986. Null alleles as a possible causeof heterozygote deficiencies in the oyster Crassos-trea virginica and other bivalves. Evolution 40:869–870.
FORMAS, R. J., I. NORTHLAND, J. CAPETILLO, NUNEZ,C. C. CUEVAS, AND L. M. BRIEVA. 1999. Telmato-bius dankoi., una nueva especie de rana acuaticadel norte de Chile (Leptodactylidae). Revista Chi-lena de Historia Natural 72:427–445.
FROST, D. R., AND D. M. HILLIS. 1990. Species inconcept and practice: herpetological applications.Herpetologica 46:87–104.
FROST, D. R., AND A. G. KLUGE. 1994. A consider-ation of epistemology in systematic biology withspecial reference to species. Cladistics 19:259–294.
GAFFNEY, P. M., T. M. SCOTT, R. K. KOEHN, AND W.J. DIEHL. 1990. Interrelationships of heterozygote,growth rate and heterozigote deficiencies in thecoot clam, Mulinia lateralis. Genetics 124:687–699.
GARDNER, J. P. A. 1992. Null alleles and heterozygotedeficiencies among mussels (Mytilus edulis and M.
galloprovincialis) of two sympatric populations.Malacologia 34:99–106.
GARMAN, S. W. 1876. I. Fishes and reptiles. Pp. 274–278. In A. Agassiz and S. W. Garman (Eds.), Ex-ploration of Lake Titicaca. Bulletin of the Museumof Comparative Zoology, Cambridge no.11, 19(1).
GARNIER-GERE, P., AND C. DILLMANN. 1992. A com-puter program for testing pairwaise linkage dis-equilibrium in subdivided populations. Journal ofHeredity 83:239.
GERGUS, E. W. A. 1998. Systematics of the Bufo mi-croscaphus complex: allozymic evidence. Herpeto-logica 54:317–325.
GILSON, H. C. 1939. 1. Description of the Expedi-tion. In The Percy Sladen Trust Expedition to LakeTiticaca in 1937. Transactions of the Linnean So-ciety, London, ser 3, 1:1–20.
GUO, S. W., AND E. A. THOMPSON. 1992. Performingthe exact test of Hardy-Weinberg proportion formultiple alleles. Biometrics 48:361–372.
HUMPHRIES, J. M., F. L. BOOKSTEIN, B. CHERNOFF,G. R. SMITH, R. L. ELDER, AND S. G. POSS. 1981.Multivariate discrimination by shape in relation tosize. Systematic Zoology 30:291–308.
HUTCHINSON, V. H. 1982. Physiological ecology ofthe telmatobiid frogs of the Lake Titicaca. NationalGeographic Society Research Report 14:357–361.
HUTCHINSON, V. H., H. B. HAINES, AND G. ENG-BRETSON. 1976. Aquatic life at high altitude: respi-ratory adaptations in the Lake Titicaca frog, Tel-matobius culeus. Respiration Physiology 27:115–129.
KATOH, M., AND D. W. FOLTZ. 1989. Biochemicalevidence for the existence of a null allele at theleucine aminopeptidase-2 (Lap-2) locus in the oys-ter Crassostrea virginica (Gmelin). Genome 32:687–690.
KLICKA, J., AND R. M. ZINK. 1997. The importanceof recent ice ages in speciation: a failed paradigm.Science 277:1666–1669.
KLINGENBERG, C. P., AND R. FROESE. 1991. A mul-tivariate comparison of allometric growth patterns.Systematic Zoology 40:410–419.
LAMBRECHTS, M. M., J. BLONDEL, M. MAISTRE, ANDP. PERRET. 1997. A single response mechanism isresponsible for evolutionary adaptive variation in abird’s laying date. Ecology 94:5135–5155.
LAURENT, R. F. 1970. Contribucion a la biometrıa dealgunas especies argentinas del genero Telmato-bius. Acta Zoologica Lilloana XXV(21):279–302.
. 1973. Nuevos datos sobre el genero Telma-tobius en el noroeste argentino, con descripcion deuna nueva especie en la sierra del Manchao. ActaZoologica Lilloana 30:163–187.
. 1977. Contribucion al conocimiento del ge-nero Telmatobius Wiegmann (4a Nota). Acta Zool-ogica Lilloana 10:189–206.
LAUZANNE, L. 1982. Les Orestias (Pisces, Cyprino-dontidae) du Petit Lac Titicaca. Revue de Hidro-biologie Tropicale 15:39–70.
. 1991. VI.5a Especies nativas, los Orestias.Pp. 409–424. In C. Dejoux and A. Iltis (Eds), ElLago Titicaca. Sıntesis del conocimiento limnolo-gico actual, ORSTOM-HISBOL, La Paz, Bolivia.

54 [Vol. 58, No. 1HERPETOLOGICA
LAVENU, A. 1981. Origine et evolution neotectoniquedu Lac Titicaca. Revue de Hydrobiologie Tropicale14:289–297.
LOSOS, J. B., T. R. JACKMAN, A. LARSON, K. DE QUEI-ROZ, AND L. RODRIGUEZ-SCHETTINO. 1998. Con-tingency and determinism in replicated adaptiveradiations of island lizards. Science 279:2115–2118.
LU, G., AND L. BERNATCHEZ. 1999. Correlated tro-phic specialization and genetic divergence in sym-patric lake whitefish ecotypes (Coregonus clupea-formis): support for the ecological speciation hy-pothesis. Evolution 53:1491–1505.
LYNCH, J. D. 1978. A re-assessment of the telmato-biine leptodactyloid frogs of Patagonia. OccasionalPapers of the Museum of Natural History, Univer-sity of Kansas 72:1–57.
. 1986. Origins of the high Andean herpeto-logical fauna. Pp. 478–499. In F. VUILLEUMIERAND M. MONASTERIO (Eds.), High Altitude Trop-ical Biogeography. Oxford University Press, Oxford,U.K.
MACEDO, H. DE. 1960. Vergleichende untersunchun-gen an arten der gattung Telmatobius (Amphibia:Anura). Zeitschrift fur wissenschaftliche zoologie163:355–396.
MINK, D. G., AND J. W. SITES, JR. 1996. Species lim-its, phylogenetics relationships, and origins of vivi-parity in the Scalaris complex of the lizard genusSceloporus (Phrynosomatidae: Sauria). Herpetolo-gica 52:551–571.
MORIZOT, D. C., AND M. E. SCHMIDT. 1990. Starchgel electrophoresis and histochemichal visualiza-tion of proteins. Pp. 23–80. In D. H. Whitmore(Ed.), Electrophoretic and Isoelectric FocusingTechniques in Fisheries Management. CRC Press,Boca Raton, Florida, U.S.A.
MOURGUIART, P., J. ARGOLLO, P. CARBONEL, E. CO-RREGE, AND D. WIRRMANN. 1995b. El Lago Titi-caca durante el Holoceno: una historia compleja.Pp. 173–188. In J. Argollo and P. Mourguiart(Eds.), Cambios Cuaternarios en America del Sur.ORSTOM-HISBOL, La Paz, Bolivia.
MOURGUIART, P., J. ARGOLLO, AND D. WIRRMANN.1995a. Evolucion del Lago Titicaca desde 25 000anos bp. Pp. 157–171. In J. Argollo and P. Mour-guiart (Eds.), Cambios Cuaternarios en Americadel Sur. ORSTOM-HISBOL, La Paz, Bolivia.
MURPHY, R. W., J. W. SITES, JR., D. G., BUTH, ANDC. H. HAUFLER. 1996. Proteins: isozyme electro-phoresis. Pp. 51–120. In D. M. Hillis, C. Moritz,and B. K. Mable (Eds.), Molecular Systematics,2nd ed. Sinauer, Sunderland, Massachusetts,U.S.A.
NAGEL, L., AND D. SCHLUTER. 1998. Body size, nat-ural selection, and speciation in sticklebacks. Evo-lution 52:209–218.
NEI, M. 1978. Estimation of average heterozygosityand genetic distance from a small number of in-dividuals. Genetics 23:341–369.
ORR, M. R., AND T. B. SMITH. 1998. Ecology andspeciation. Trends in Ecology and Evolution 13:502–506.
PARENTI, L. R. 1984. A taxonomic revision of the An-dean killifish genus Orestias (Cyprinodontiformes,
Cyprinodontidae). Bulletin of the American Mu-seum of Natural History 178:107–214.
PARKER, H. W. 1940. Percy Sladen Trust Expeditionto Lake Titicaca under the leadership of Mr. H.Cary Gilson, M. A.—XII—Amphibia. Transactionsof the Linnean Society, London 3:203–216.
PENTLAND, J. B. 1848. The Laguna of Titicaca andthe valleys of Yukay, Collao and Desaguadero inPeru and Bolivia, from geodesic and astronomicalobservations made in the years 1827 and 1828,1837 and 1838. British Admiralty Chart No.1868,London, U.K.
RAYMOND, M., AND F. ROUSSET. 1995a. An exact testfor population differentiation. Evolution 49:1280–1283.
. 1995b. GENEPOP (version 1.2.): populationgenetics software for exact test and ecumenicism.Journal of Heredity 86:248–249.
REIST, J. D. 1985. An empirical evaluation of severalunivariate methods that adjust for size variation inmorphometric data. Canadian Journal of Zoology63:1429–1435.
REZNICK, D. A., H. BRYGA, AND J. A. ENDLER. 1990.Experimentally induced life-history evolution in anatural population. Nature 346:357–359.
RICHARDSON, B. J., P. R. BAVERSTOCK, AND M. AD-AMS. 1986. Allozyme Electrophoresis. A Handbookfor Animal Systematics and Population Structure.Academic Press, Sydney, Australia.
ROUSSET, F., AND M. RAYMOND. 1995. Testing het-erozygote excess and deficiency. Genetics 140:1413–1419.
RUNDLE, H. D., L. NAGEL, J. W. BOUGHMAN, ANDD. SCHLUTER. 2000. Natural selection and parallelspeciation in sympatric sticklebacks. Science 287:306–308.
SAETRE, G. P., T. MOUM, S. BURES, M. KRAL, M.ADAMJAN, AND J. MORENO. 1997. A sexually se-lected character displacement in flycatchers rein-forces premating isolation. Nature 387:589–592.
SALAS, A. W., AND U. SINSCH. 1996. Two new Tel-matobius species (Leptodactylidae, Telmatobiinae)of Ancash, Peru. Alytes 14:1–26.
SCHILTHUIZEN, M. 2000. Ecotone: speciation-prone.Trends in Ecology and Evolution 15:130–131.
SCHLUTER, D. 1993. Adaptive radiation in stickle-backs: size, shape and habitat use efficiency. Ecol-ogy 74:699–709.
. 1996a. Ecological speciation in postglacialfishes. Philosophical Transactions of the Royal So-ciety of London Series B 351:807–814
. 1996b. Ecological causes of adaptive radia-tion. American Naturalist 148:S40-S64.
. 1998. Ecological causes of speciation. Pp.114–129 In D. Howard, and S. H. Berlocher(Eds.), Endless Forms. Species and Speciation. Ox-ford University Press, New York, New York, U.S.A.
SCHNEIDER, C. J., T. B. SMITH, B. LARISON, AND C.MORITZ. 1999. A test of alternative models of di-versification in tropical rainforests: ecological gra-dients vs. rainforest refugia. Proceedings of the Na-tional Academy of Sciences USA 24:13869–13873.
SELANDER, R. K., M. H. SMITH, S. J. YANG, W. E.JOHNSON, AND J. B. GENTRY. 1971. Biochemichal

March 2002] 55HERPETOLOGICA
polymorphism and systematics in the genus Pero-myscus. I. Variation in the old-field mouse (Pero-myscus polionotus). Studies in Genetics VI, Uni-versity of Texas Publication 7103:49–90.
SINSCH, U., AND N. JURASKE. 1995. Reassessment ofcentral Peruvian Telmatobiinae (genera Batracho-phrynus and Telmatobius). II. Allozymes and phy-logenetics relationships. Alytes 13:52–66.
SINSCH, U., A. W. SALAS, AND V. CANALES. 1995. Re-assessment of central Peruvian Telmatobiinae (gen-era Batrachophrynus and Telmatobius). I. Mor-phometry and classification. Alytes 13:14–44.
SITES, J. W., JR., AND K. A. CRANDALL. 1997. Testingspecies boundaries in biodiversity studies. Conser-vation Biology 11:1289–1297.
SITES, J. W., JR., AND R. W. MURPHY. 1991. Isozymeevidence for independently derived, duplicateG3PDH loci among squamate reptiles. CanadianJournal of Zoology 69:2381–2396.
SKULASON, S., AND T. S. SMITH. 1995. Resource poly-morphism in vertebrates. Trends in Ecology andEvolution 10:366–370
SMITH, T. B., AND S. SKULASON. 1996. Evolutionarysignificance of resource polymorphisms in fishes,amphibians, and birds. Annual Review of Ecologyand Systematics 27:111–133.
SMITH, T. B., R. K. WAYNE, D. J. GIRMAN, AND M.W. BRUFORD. 1997. A role for ecotones in gener-ating rainforest biodiversity. Science 276:1855–1857.
SOMERS, K. M. 1986. Multivariate allometry and re-moval of size with principal components analysis.Systematic Zoology 35:359–368.
STATSOFT. 1995. STATISTICA for Windows. Stat-Soft, Inc., Tulsa, Oklahoma, U.S.A.
TARKHNISHVILI, D. N., J. W. ARNTZEN, AND R. S.THORPE. 1999. Morphological variation in brownfrogs from the Caucasus and the taxonomy of theRana macrocnemis group. Herpetologica 55:406–417.
TRUEB, L. 1979. Leptodactylid frogs of the genus Tel-matobius in Ecuador with the description of a newspecies. Copeia 1979:714–733.
VELLARD, J. 1951. Estudios sobre batracios andinos.I. El Grupo Telmatobius y formas afines. Memoriasdel Museo de Historia Natural, Javier Prado 1:1–89 viii pl.
. 1952. Adaptation des batraciens a la vie agrande hauteur dans les Andes. Bulletin de la So-ciete Zoologique de France 77:169–187.
. 1953. Estudios sobre batracios andinos. II.El Grupo Marmoratus y formas afines. Memoriasdel Museo de Historia Natural. Javier Prado 2:1–53 vi pl.
. 1960. Estudios sobre batracios andinos. VI.Notas complementarias sobre Telmatobius. Me-
morias del Museo de Historia Natural. Javier Prado10:1–19 xiv pl.
. 1969. Les Telmatobius du groupe marmor-atus (Amphibia). Bulletin du Museum NationalD’histoire Naturelle 2eme ser. 40:1110–1113.
. 1991. Los Batracios. Pp. 453–462. In C. De-joux, and A. Iltis (Eds.), El Lago Titicaca. Sıntesisdel Conocimiento Actual. ORSTOM-HISBOL, LaPaz, Bolivia.
VILLWOCK, W. 1986. Speciation and adaptive radia-tion in Andean Orestias fishes. Pp. 387–403. In F.Vuilleumier and M. Monasterio (Eds.), High Alti-tude Tropical Biology. Oxford University Press, Ox-ford, U.K.
WEIR, B. S., AND C. C. COCKERMAN. 1984. Estimat-ing F-statistics for the analysis of population struc-ture. Evolution 38:1358–1370.
WIENS, J. J. 1993. Systematics of the leptodactylidfrog genus Telmatobius in the Andes of northernPeru. Occasional Papers of the Museum of NaturalHistory, University of Kansas 162:1–76.
. 1995. Polymorphic characters in phylogenet-ic systematics. Systematic Biology 44:482–500.
. 1999. Polymorphism in systematics and com-parative biology. Annual Review of Ecology andSystematics 30:327–362.
. 2000. Reconstructing phylogenies from al-lozyme data: comparing method performance withcongruence. Biological Journal of the Linnean So-ciety 70:613–632.
WIENS, J. J., AND M. R. SERVEDIO. 1997. Accuracyof phylogenetic analysis including and excludingpolymorphic characters. Systematic Biology 46:332–345.
. 1998. Phylogenetic analysis and intraspecificvariation: performance of parsimony, distance, andlikelihood methods. Systematic Biology 47:228–253.
. 2000. Species delimitation in systematics: in-ferring diagnostic differences between species.Proceedings of the Royal Society, London. B, Bi-ological Sciences 267:631–636.
WILEY, E. O. 1978. The evolutionary species conceptreconsidered. Systematic Zoology 27:17–26
WIRRMANN, D., J. P. YBERT, AND P. MOURGUIART.1991. Una evaluacion paleohidrologica de 20 000anos. Pp. 37–45. In C. Dejoux, and A. Iltis (Eds.),El Lago Titicaca. Sıntesis del Conocimiento Actual.ORSTOM-HISBOL, La Paz, Bolivia.
YBERT, J. P. 1991. Los paisajes lacustres antiguos se-gun el analisis palinologico. Pp. 69–82. In C. De-joux and A. Iltis (Eds.), El Lago Titicaca. Sıntesisdel Conocimiento Actual. ORSTOM-HISBOL, LaPaz, Bolivia.
Accepted: 24 February 2001Associate Editor: Joseph Mendelson III




















