
Identification of palearctic coregonid fish species using mtDNA and allozyme genetic markers
21
Journal of Fish Biology (2000) 57 (Supplement A), 51–71 doi:10.1006/jfbi.2000.1608, available online at http://www.idealibrary.com on Identification of palearctic coregonid fish species using mtDNA and allozyme genetic markers D. V. P*†‡, N. Y.G*, K. I. A*, Y. P. A* J. W. B† *Vavilov Institute of General Genetics, 3 Gubkin St., GSP-1, Moscow 117809, Russia and †Department of Wildlife and Fisheries Sciences, Texas A&M University, College Station TX, 77843-2258, U.S.A. Two types of molecular genetic markers were used for genetic identification of species and local stocks of palearctic coregonids (Coregonidae, Salmoniformes, Teleostei). Seven nominate species of whitefishes and ciscoes Coregonus spp. of Eurasia Arctic Sea basin and inconnu Stenodus leucichthys nelma represented by specimens from North America were studied. Using restriction analysis of PCR-amplified products of the ND-1 gene of mitochondrial DNA (mtDNA) and allelic composition at several allozyme loci discrimination was successful between C. lavaretus pidschian Siberian whitefish, C. nasus broad whitefish, C. autumnalis Arctic cisco, C. migratorius Baikal omul, C. peled peled, and C. sardinella least cisco. Muksun C. muksun was indistinguishable from Siberian whitefish. Creatine kinase (CK) isozyme patterns and Rsa I restriction patterns of ND-1 were the most effective markers allowing discrimination among species. Intra-specific differentiation in mtDNA was found in all species but was much less pronounced than inter-species variation. In several specimens composite haplotypes typical of another species were found that reflect probable gene introgression by hybridization. A combination of mtDNA and nuclear genetic markers is suggested for reliable identification of both typical species representatives and hybrids. 2000 The Fisheries Society of the British Isles Key words: coregonid; Coregonus; mtDNA; isozyme; identification; species. INTRODUCTION Coregonid fishes (Coregonidae, Salmoniformes) are core elements of estuarine and freshwater ecosystems of the Arctic Sea basin (Reshetnikov, 1980; Novikov et al., 2000), especially in Siberia and North-east European Russia. Along with Arctic charr Salvelinus alpinus (L.) and Arctic grayling Thymallus arcticus (Pallas), coregonids play the principal role in food chains of the tundra zone and have great economic importance in subsistence and commercial fisheries. Besides its practical importance, the identification and differentiation of species and local stocks of coregonid fishes is important for understanding the evolu- tionary mechanisms of speciation at high latitudes and the development of Arctic aquatic faunas. Phenotypic identification of coregonid species is complicated by high plasticity of body features and meristic characters (Lindsey, 1981). Existing keys are often misleading or provide ambiguous diagnoses especially for individuals with intermediate values in traits that are supposed to be discrimi- native. It is not unusual to encounter situations in which individuals possess a combination of character states thought to be typical of different species. Greater efficiency and reliability of morphological identification of coregonids ‡Author to whom correspondence should be addressed. Tel.: (095) 135 5067; fax: (095) 132 8962, email: [email protected] 51 0022–1112/00/57A051+21 $35.00/0 2000 The Fisheries Society of the British Isles
Transcript of Identification of palearctic coregonid fish species using mtDNA and allozyme genetic markers

Journal of Fish Biology (2000) 57 (Supplement A), 51–71doi:10.1006/jfbi.2000.1608, available online at http://www.idealibrary.com on
Identification of palearctic coregonid fish species usingmtDNA and allozyme genetic markers
D. V. P*†‡, N. Y. G*, K. I. A*,Y. P. A* J. W. B†
*Vavilov Institute of General Genetics, 3 Gubkin St., GSP-1, Moscow 117809, Russiaand †Department of Wildlife and Fisheries Sciences, Texas A&M University,
College Station TX, 77843-2258, U.S.A.
Two types of molecular genetic markers were used for genetic identification of species and localstocks of palearctic coregonids (Coregonidae, Salmoniformes, Teleostei). Seven nominatespecies of whitefishes and ciscoes Coregonus spp. of Eurasia Arctic Sea basin and inconnuStenodus leucichthys nelma represented by specimens from North America were studied. Usingrestriction analysis of PCR-amplified products of the ND-1 gene of mitochondrial DNA(mtDNA) and allelic composition at several allozyme loci discrimination was successfulbetween C. lavaretus pidschian Siberian whitefish, C. nasus broad whitefish, C. autumnalis Arcticcisco, C. migratorius Baikal omul, C. peled peled, and C. sardinella least cisco. Muksun C.muksun was indistinguishable from Siberian whitefish. Creatine kinase (CK) isozyme patternsand Rsa I restriction patterns of ND-1 were the most effective markers allowing discriminationamong species. Intra-specific differentiation in mtDNA was found in all species but was muchless pronounced than inter-species variation. In several specimens composite haplotypes typicalof another species were found that reflect probable gene introgression by hybridization. Acombination of mtDNA and nuclear genetic markers is suggested for reliable identification ofboth typical species representatives and hybrids. � 2000 The Fisheries Society of the British Isles
Key words: coregonid; Coregonus; mtDNA; isozyme; identification; species.
‡Author to whom correspondence should be addressed. Tel.: (095) 135 5067; fax: (095) 132 8962, email:[email protected]
INTRODUCTION
Coregonid fishes (Coregonidae, Salmoniformes) are core elements of estuarineand freshwater ecosystems of the Arctic Sea basin (Reshetnikov, 1980; Novikovet al., 2000), especially in Siberia and North-east European Russia. Along withArctic charr Salvelinus alpinus (L.) and Arctic grayling Thymallus arcticus(Pallas), coregonids play the principal role in food chains of the tundra zone andhave great economic importance in subsistence and commercial fisheries.Besides its practical importance, the identification and differentiation of speciesand local stocks of coregonid fishes is important for understanding the evolu-tionary mechanisms of speciation at high latitudes and the development of Arcticaquatic faunas. Phenotypic identification of coregonid species is complicated byhigh plasticity of body features and meristic characters (Lindsey, 1981). Existingkeys are often misleading or provide ambiguous diagnoses especially forindividuals with intermediate values in traits that are supposed to be discrimi-native. It is not unusual to encounter situations in which individuals possess acombination of character states thought to be typical of different species.Greater efficiency and reliability of morphological identification of coregonids
51
0022–1112/00/57A051+21 $35.00/0 � 2000 The Fisheries Society of the British Isles

52 . . .
can be achieved by careful prior analysis of traits in large samples to selectdiagnostic ones (Lindsey, 1962), but this was not always the case when theexisting identification keys for coregonids were created. Discrimination andphylogenetic inferences based on anatomical, especially osteological, traits ofcoregonids have been successful (GTsowska, 1960; Shaposhnikova, 1968, 1970;Smith & Todd, 1992; Chereshnev, 1994). However, these traits are not useful forfield identification. Given these problems, genetic markers such as isozyme lociand mitochondrial DNA (mtDNA) have been employed widely in populationand phylogenetic studies of coregonids. However, until recently most studiesfocused on populations inhabiting North America, Central Europe andFennoscandia (Bernatchez & Dodson, 1991, 1994; Bernatchez et al., 1991; Reistet al., 1998; Vuorinen et al., 1998; Douglas et al., 1999). Genetic data oncoregonids from Russian territories are relatively scarce (Bodaly et al., 1991,1994; Ermolenko, 1992; Bernatchez & Dodson, 1994) and represent poorlythe vast ranges of these fishes in Siberia and North European Russia. Identitiesof European, Siberian and North American coregonid forms known underthe same scientific and common names are often questionable. Even theconspecificity of populations of some Russian coregonids from different parts ofthe species ranges needs to be tested by genetic data.
This paper presents information about genetic identification of coregonidspecies distributed in the Russian part of the Arctic Sea basin and continues aseries of publications on the population genetic structure and phylogeny ofPalearctic (Politov & Gordon 1998; Politov et al., 2000) and Nearctic (Bickhamet al., 1989, 1997; Morales et al., 1993) coregonids.
MATERIALS AND METHODS
Tissue samples were obtained during fieldwork conducted in 1994–1998 (Table I).Most samples were collected during the Swedish–Russian Expedition ‘ Tundra Ecology-94 ’ (Grønlund & Melander, 1995; Novikov et al., 1995; Politov et al., 2000; Novikovet al., 2000). Baikal omul Coregonus migratorius (Georgi) and Baikal whitefishC. lavaretus baicalensis Dybowski were collected from the southern coast of BolshoiUshkaniy Island in the east central part of Baikal Lake (Politov et al., 2000). PeledC. peled Gmelin was collected from Turukhan River (a tributary of the Yenisey River,Table I). In addition, several specimens of circumpolarly distributed inconnu Stenodusleucichthys nelma Guldenstadt were collected from the North American Arctic coastbasin (the same sub-species is known from the Palearctic rivers).
Specimens were identified in the field using traditional morphological keys by Berg(1948) and Reshetnikov (1980). Taxonomy follows Reshetnikov (1998), except forBaikal omul, that is treated here as a separate species according to its first description.This is supported by osteological (Shaposhnikova, 1970) and previous isozyme (Politovet al., 2000) data. Reference specimens of each species from several localities were fixedin formalin and later deposited in collections of the Department of Ichthyology, MoscowState University and the Swedish Museum of Natural History (Stockholm). Tissuestaken for genetic analysis (skeletal muscle, heart, liver and eye) are stored frozen at�80� C at the Laboratory of Population Genetics, Vavilov Institute of General Genetics(Moscow, Russia).
Isozyme loci nomenclature followed Shaklee et al. (1990). Interpretation of isozymepatterns with respect to number of coding loci and enzyme sub-unit structure generallycorresponded to previous reports for coregonids (Vuorinen, 1984; Vuorinen & Piironen,1984). Other details of the isozyme techniques used are to be found in Politov et al.(1998, 2000).

53
T I. Studied samples of coregonid fishes
Species/location Sample size* Dates ofcollection
Arctic cisco—Coregonus autumnalis Pallas n=64 (63)W. to Pechora River delta (68�48�40� N; 53�42� E) 2 (2) August 1994W. Yamal Peninsula (70�20� N; 67�50� E) 3 (3) June and
August 1994N. Yamal Peninsula (72�51�30� N; 70�55� E) 8 (8) August 1994E. to Yana River mouth (72�24� N; 140�57�30� E) 50 (49) August 1994Indigirka River delta (71�26�30� N; 149�45� E) 1 (1) July 1994
Least cisco—Coregonus sardinella Valenciennes n=67 (17)W. to Pechora River delta (68�48�40�; N 53�42� E) 3 (4) June–August
1994W. Yamal Peninsula (70�20� N; 67�50� E) 16 (4) June and
August 1994N. Yamal Peninsula (72�51�30� N; 70�55� E) 3 (1) August 1994Kotelny Island (75�03�30� N; 140�10� E) 9 (1) July–August
1994Indigirka River delta (71�26�30� N; 149�45� E) 32 (4) July 1994Kolyma River delta (69�32�30� N; 160�42� E) 4 (3) July 1994
Peled—Coregonus peled Gmelin n=0 (12)Yenisey River basin (65�47� N; 87�56� E) 0 (12) July 1998
Muksun—Coregonus muksun Pallas n=47 (17)W. Yamal Peninsula (70�20� N; 67�50� E) 36 (8) June and
August 1994Indigirka River delta (71�26�30� N; 149�45� E) 7 (6) July 1994Kolyma River delta (69�32�30� N; 160�42� E) 4 (4) July 1994
Broad whitefish—Coregonus nasus Pallas n=106 (33)W. Yamal Peninsula (70�20� N; 67�50� E) 15 (8) June and
August,1994Olenekskii Bay (73�28�30�N 118�01� E) 1 (1) July 1994E. to Yana River mouth (72�24� N; 140�57�30� E) 11 (10) August 1994Indigirka River delta (71�26�30� N; 149�45� E) 23 (7) July 1994Kolyma River delta (69�32�30� N; 160�42� E) 60 (7) July 1994
Siberian whitefish—Coregonus lavaretus pidschian Pallas n=100 (20)W. to Pechora River delta (68�48�40�; N 53�42� E) 27 (5) August 1994W. Yamal Peninsula (70�20� N; 67�50� E) 17 (4) June and
August 1994Indigirka River delta (71�26�30� N; 149�45� E) 31 (4) July 1994Kolyma River delta (69�32�30� N; 160�42� E) 25 (8) July 1994
Baikal Lake whitefish—Coregonus lavaretusbaicalensis Dybowski
n=0 (2)
Lake Baikal (53�50� N; 108�37�) 0 (2) August 1995
Baikal omul—Coregonus migratorius (Georgi) n=42 (40)Lake Baikal (53�50� N; 108�37�) 42 (40) August 1995
Inconnu—Stenodus leucichthys nelma Guldenstadt n=0 (4)MacKenzie River basin, Canada (66�58� N; 135�08�W) 0 (4) August 1988
*Numbers of fish analysed by isozymes, sample sizes for mtDNA analysis are given in parentheses.Sample sizes for each species are given according to field morphological diagnoses and may differ from
those inferred from genetic data.

54 . . .
Deep-frozen or ethanol preserved tissues were used for DNA analysis. Prior to DNAextraction frozen tissues (heart, skeletal muscle or liver) were transferred to 20% DMSOin saturated NaCl as recommended by Maiers et al. (1998) to enhance DNA recovery.Total DNA was extracted by lysis of tissues for 1 h at 60� C in 0·65 ml extraction buffer(540 �l 0·1 Tris pH 7·8, 70 �l 10% SDS, 40 �l 1% proteinase K). One extraction withphenol: chloroform: isoamyl alcohol (24:24:1) and one extraction with methylenechloride: isoamyl alcohol (24:1) were followed by ethanol precipitation of the aqueousphase, centrifugation for 5 min at 12 000 rpm and resuspension of the DNA pellet in100 �l of TE buffer (Sambrook et al., 1989). Polymerase chain reaction (PCR) was usedto amplify the NADH-dehydrogenase-1 gene of the mtDNA together with flankingregions using primers LGL381: 5�-ACCCCGCCTGTTTACCAAAAACAT-3� andLGL563: 5�-GGTTCATTAGTGAGGGAAGG-3� (Cronin et al., 1993). PCR reactionswere performed in 60 �l volumes using a Perkin-Elmer DNA Thermal Cycler (PerkinElmer Applied Biosystems, Foster City, CA). PCR was carried out in 32 cycles ofdenaturation at 94� C for 1 min, annealing at 52� C for 1 min and extension at 72� C for1 min 45 s. PCR product (hereafter named ‘ ND-1 fragment ’) was c. 2100 bp in length.Aliquots (2·5 �l) of these products were cut by 18 restriction enzymes (Ase I, Ava II, BsaJI, Bsp1286 I, BstN I, BstU I, Dde I, Dpn II, Hae III, Hinc II, Hinf I, Hha I, Hph I, MspI, Nci I, Rsa I, Ssp I, Taq I). Restriction digests were incubated for at least 2 h underconditions specified by the manufacturer (New England BioLabs, Beverly, MA) and runon 2–2·5% agarose gels, stained with ethidium bromide and screened under UV light.Fragment size was determined using GIBCO BRL 100 bp DNA size marker (LifeTechnologies, Gaithersburg, MD). Restriction banding patterns were transformed into amatrix of presence or absence of particular cutting sites. Numbers of non-sharedrestriction sites (hereafter called pairwise differences) were counted among haplotypesand a minimum-spanning network was constructed (Schneider et al., 2000).
RESULTS AND DISCUSSION
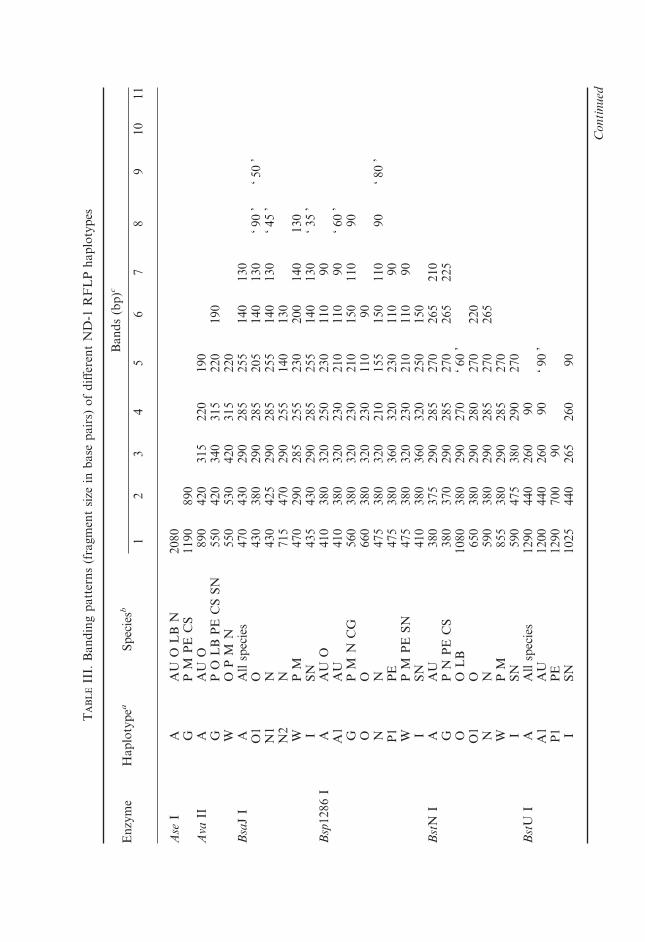
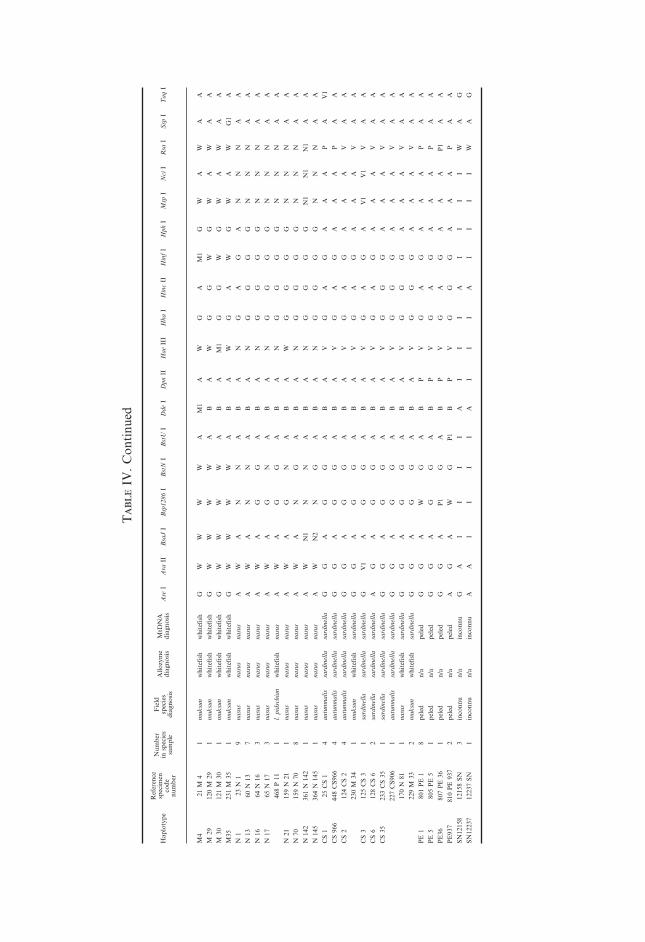
Allelic frequencies at polymorphic allozyme loci are given in Table II.Restriction fragment length polymorphism (RFLP) patterns of ND-1 mtDNAhaplotypes are summarized in Table III. Their distributions among differentspecies and species diagnoses based on different types of data are presented inTable IV.
GENERA COREGONUS AND STENODUSFour specimens of Stenodus leucichthys nelma were analyzed by mtDNA only.
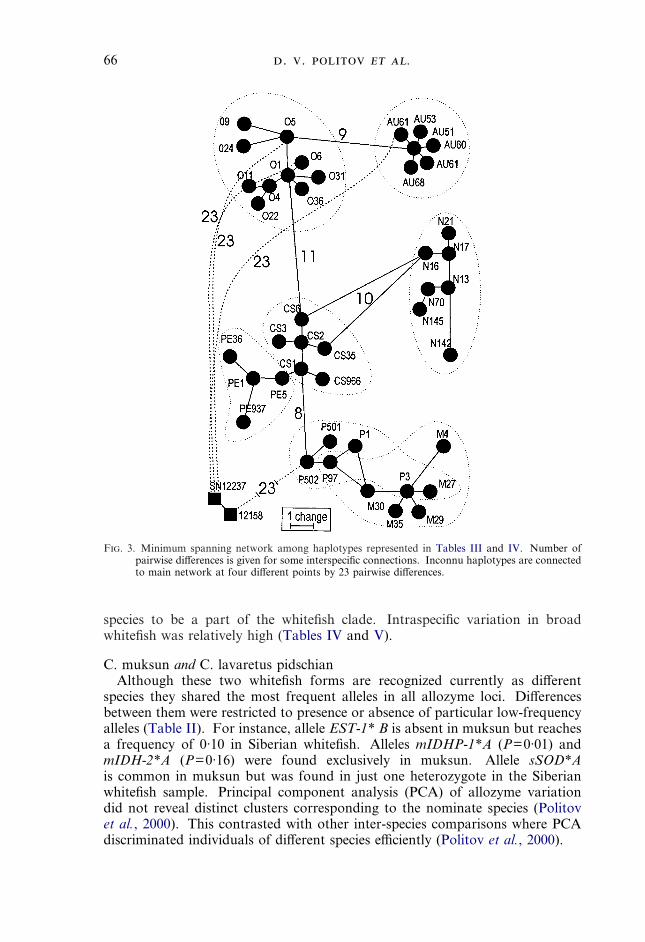
Unique restriction patterns were produced with nine enzymes (Table IV). Theinconnu Nci I pattern occurred only in a rare Baikal omul haplotype O31. Witha minimum of 23 pairwise differences with the closest Coregonus (Table IV),inconnu was the most easily identifiable among the species studied. All geneticstudies have shown inconnu to be closer to Coregonus than the third coregonidgenus Prosopium (Hamada et al., 1998). However, estimations of its positionwithin this clade have been inconsistent. For example, S. leucichthys wassuggested to be the sister taxon to: (1) all species of Coregonus (Ermolenko, 1992;Vuorinen et al., 1998), (2) C. autumnalis (Pallas)–C. artedii (Lesueur) speciescomplex (Bernatchez et al., 1991), (3) all Coregonus excluding C. albula (L.)(Sajdak & Phillips, 1997) and (4) C. albula–C. sardinella (Valenciennes)–C. peledspecies complex (Lockwood et al., 1993; Reist et al., 1998). The observed highgenetic differentiation of inconnu makes it possible to use it as an outgroup forphylogenetic reconstructions among species of genus Coregonus.

55
SUBGENERA COREGONUS AND LEUCICHTHYSNo alleles or mtDNA haplotypes delimited the commonly recognized sub-
genera Coregonus (whitefishes) and Leucichthys (ciscoes). Instead, at least threemajor groups of species were observed. Allozymes indicate equal levels ofdivergence among least cisco C. sardinella, Arctic cisco C. autumnalis andwhitefishes (Politov et al., 2000). Virtually the same result was obtained byanalysis of the mtDNA ND-1 fragment (Table V). Arctic cisco and least ciscohaplotypes were even more differentiated (a minimum of 15 pairwise differences)than least cisco and whitefishes (11–13). Therefore, the traditional sub-divisionof the genus Coregonus into sub-genera based on mouth position (sub-terminalin whitefishes and terminal or supra-terminal in ciscoes) is not supported (TablesII–V). Previous isozyme (Bodaly et al., 1991, 1994; Ermolenko, 1992; Vuorinenet al., 1998) and DNA-based (Bernachez et al., 1991; Reist et al., 1998) studiesalso have not found evidence for the validity of these sub-genera.
IDENTIFICATION OF SPECIES IN THE GENUS COREGONUS
C. sardinellaLeast cisco can be identified by a combination of the following allozymealleles: fixed AH-1*B, AH-2*C, nearly fixed alleles GPI-A1* and PGM-1*, andalso private alleles at polymorphic frequencies in CK-1,2* (Fig. 1) and someother loci (Table II). Siberian populations have a unique ND-1 restrictionpattern with Rsa I (Fig. 2), while European least cisco shares type P with peled(differing in Dpn II). Six different haplotypes were observed among 17 studiedleast cisco individuals (Table IV).
C. peledPeled was not analysed by allozymes. However, mtDNA data provide enough
resolution to discriminate it clearly from other Coregonus species. Peled has aunique Dpn II pattern and shares Rsa I P type (Fig. 2) only with European leastcisco. Peled differs from Arctic cisco, Baikal omul and whitefishes by severalother restriction enzymes (Table IV), being most closely related to least cisco(Table V, Fig. 3). All restriction sites except for Dpn II were shared betweenpeled haplotype P5 and the closest least cisco haplotype CS966 (Tables IV, V;and Fig. 3). Earlier genetic studies also demonstrated that peled is phylogeneti-cally close to least cisco (Bernatchez et al., 1991; Bodaly et al., 1994; Vuorinenet al., 1998) and to its closest relative from Europe, vendace C. albula (Reistet al., 1998). Four haplotypes were found among 12 peled specimens (Tables IV,V, Fig. 3).
C. autumnalisArctic cisco has CK-1,2*A allele at creatine kinase major isoloci (Fig. 1)
that differentiates it reliably from other species except for Baikal omul. Arcticcisco IDH-4*B allele otherwise occurs only in Baikal omul and broad whitefishC. nasus (Pallas) at very low frequencies. Arctic cisco can also be identified byprivate alleles mMEP-1,2*A and GPI-B1*E, and by high (0·98) frequency ofPGM-1*A.
Restriction enzymes Hae III, Hinf I and Rsa I (Fig. 2) produced patternsspecific to Arctic cisco. A minimum of nine pairwise differences has been found
56 . . .
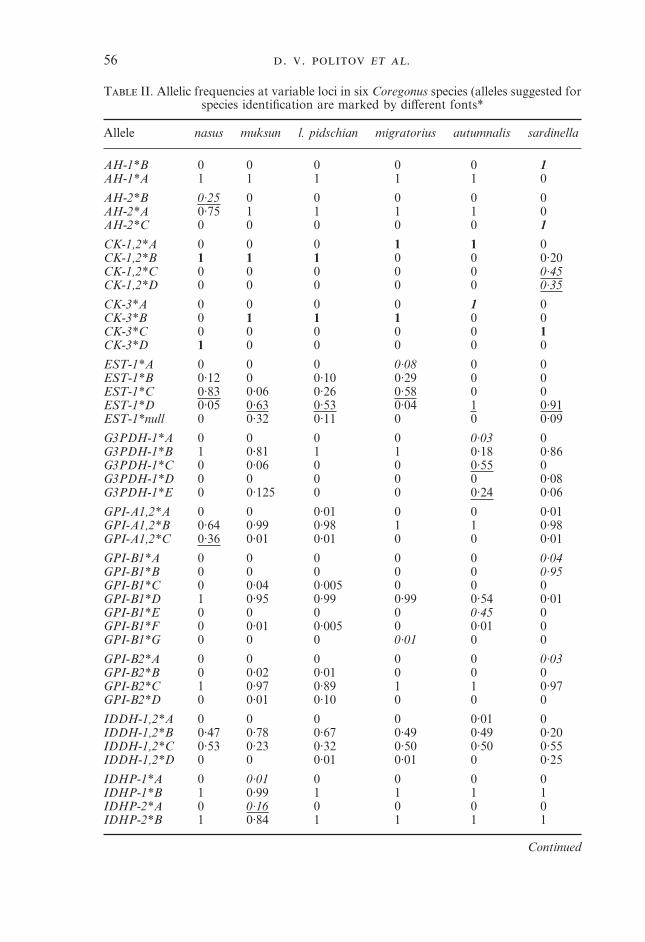
T II. Allelic frequencies at variable loci in six Coregonus species (alleles suggested forspecies identification are marked by different fonts*
Allele nasus muksun l. pidschian migratorius autumnalis sardinella
AH-1*B 0 0 0 0 0 1AH-1*A 1 1 1 1 1 0
AH-2*B 0·25 0 0 0 0 0AH-2*A 0·75 1 1 1 1 0AH-2*C 0 0 0 0 0 1
CK-1,2*A 0 0 0 1 1 0CK-1,2*B 1 1 1 0 0 0·20CK-1,2*C 0 0 0 0 0 0·45CK-1,2*D 0 0 0 0 0 0·35
CK-3*A 0 0 0 0 1 0CK-3*B 0 1 1 1 0 0CK-3*C 0 0 0 0 0 1CK-3*D 1 0 0 0 0 0
EST-1*A 0 0 0 0·08 0 0EST-1*B 0·12 0 0·10 0·29 0 0EST-1*C 0·83 0·06 0·26 0·58 0 0EST-1*D 0·05 0·63 0·53 0·04 1 0·91EST-1*null 0 0·32 0·11 0 0 0·09
G3PDH-1*A 0 0 0 0 0·03 0G3PDH-1*B 1 0·81 1 1 0·18 0·86G3PDH-1*C 0 0·06 0 0 0·55 0G3PDH-1*D 0 0 0 0 0 0·08G3PDH-1*E 0 0·125 0 0 0·24 0·06
GPI-A1,2*A 0 0 0·01 0 0 0·01GPI-A1,2*B 0·64 0·99 0·98 1 1 0·98GPI-A1,2*C 0·36 0·01 0·01 0 0 0·01
GPI-B1*A 0 0 0 0 0 0·04GPI-B1*B 0 0 0 0 0 0·95GPI-B1*C 0 0·04 0·005 0 0 0GPI-B1*D 1 0·95 0·99 0·99 0·54 0·01GPI-B1*E 0 0 0 0 0·45 0GPI-B1*F 0 0·01 0·005 0 0·01 0GPI-B1*G 0 0 0 0·01 0 0
GPI-B2*A 0 0 0 0 0 0·03GPI-B2*B 0 0·02 0·01 0 0 0GPI-B2*C 1 0·97 0·89 1 1 0·97GPI-B2*D 0 0·01 0·10 0 0 0
IDDH-1,2*A 0 0 0 0 0·01 0IDDH-1,2*B 0·47 0·78 0·67 0·49 0·49 0·20IDDH-1,2*C 0·53 0·23 0·32 0·50 0·50 0·55IDDH-1,2*D 0 0 0·01 0·01 0 0·25
IDHP-1*A 0 0·01 0 0 0 0IDHP-1*B 1 0·99 1 1 1 1IDHP-2*A 0 0·16 0 0 0 0IDHP-2*B 1 0·84 1 1 1 1
Continued

57
T II. Continued
Allele nasus muksun l. pidschian migratorius autumnalis sardinella
IDHP-4*A 0 0 0 0 0 0·01IDHP-4*B 0·95 1 1 0·97 0 0·99IDHP-4*C 0·05 0 0 0·03 0·95 0IDHP-4*D 0 0 0 0 0·05 0
mMEP-1,2*A 0 0 0 0 0·27 0mMEP-1,2*B 0·38 0 0 0 0 0mMEP-1,2*C 0·62 1 1 1 0·73 1
PGDH*A 0 0 0 0 0·02 0PGDH*B 0·95 1 1 0·16 0·94 1PGDH*C 0·05 0 0 0·84 0·04 0
PGM-1*A 0·005 0 0·015 0 0·98 0PGM-1*B 0·005 0 0 0 0 0PGM-1*C 0·99 0·90 0·94 0·88 0·01 0·01PGM-1*D 0 0·10 0·02 0·12 0 0PGM-1*E 0 0 0·02 0 0 0PGM-1*F 0 0 0·005 0 0·01 0·97PGM-1*G 0 0 0 0 0 0·01PGM-1*H 0 0 0 0 0 0·01
PGM-2*A 0 0 0 0·86 0 0PGM-2*B 0 0·30 0·27 0·14 0·01 0PGM-2*C 1 0·70 0·73 0 0·99 0·98PGM-2*D 0 0 0 0 0 0·02
sSOD*A 0·04 0·14 0·01 0 0 0·21sSOD*B 0·96 0·86 0·99 1 1 0·79
*Fixed private alleles are given in bold italics; other private alleles in italics; fixed alleles specific to agroup of species in bold; alleles at polymorphic frequencies (>0·1) that are also useful for speciesidentification are underlined.
between Arctic cisco haplotypes and haplotypes of other species making thisspecies most differentiated and easily identified among Coregonus. Arctic ciscohad the least pronounced intraspecific variation in mtDNA with 58 out of 64individuals represented by a single haplotype A1 and an average of two pairwisedifferences within species (Table V).
C. migratoriusArctic cisco and Baikal omul are fixed for the same allele at the major creatine
kinase isoloci CK-1,2* (Fig. 1) that was not observed in any other species.However, these two forms can be differentiated by position of a minor CK3 band(Fig. 1), by alleles EST-1*C and PGM-1*D (not found in Arctic cisco), andPGDH*C and PGM-1*C (rare in Arctic cisco), IDH-4*C (rare in Baikal omul),G3PDH-1*C and GPI-B1*E (absent in Baikal omul). Baikal omul has aspecies-specific allele (frequency 0·86) at PGM-2*A.
Five restrictases produced different RFLP patterns in Arctic cisco and Baikalomul. In addition to Hae III, Hinf I and Rsa I (Fig. 2) A haplotypes which arespecific to C. autumnalis, BstN I and Hph I A haplotypes are absent in Baikalomul. Alternatively, O and O1 patterns produced by BstN I are specific to
T
III.
Ban
ding
patt
erns
(fra
gmen
tsi
zein
base
pair
s)of
diff
eren
tN
D-1
RF
LP
hapl
otyp
es
Enz
yme
Hap
loty
pea
Spec
iesb
Ban
ds(b
p)c
12
34
56
78
910
11
Ase
IA
AU
OL
BN
2080
GP
MP
EC
S11
9089
0A
vaII
AA
UO
890
420
315
220
190
GP
OL
BP
EC
SSN
550
420
340
315
220
190
WO
PM
N55
053
042
031
522
0B
saJ
IA
All
spec
ies
470
430
290
285
255
140
130
O1
O43
038
029
028
520
514
013
0‘9
0’
‘50
’N
1N
430
425
290
285
255
140
130
‘45
’N
2N
715
470
290
255
140
130
WP
M47
029
028
525
523
020
014
013
0I
SN43
543
029
028
525
514
013
0‘3
5’
Bsp
1286
IA
AU
O41
038
032
025
023
011
090
A1
AU
410
380
320
230
210
110
90‘6
0’
GP
MN
CG
560
380
320
230
210
150
110
90O
O66
038
032
023
011
090
NN
475
380
320
210
155
150
110
90‘8
0’
P1
PE
475
380
360
320
230
110
90W
PM
PE
SN47
538
032
023
021
011
090
ISN
410
380
360
320
250
150
Bst
NI
AA
U38
037
529
028
527
026
521
0G
PN
PE
CS
380
370
290
285
270
265
225
OO
LB
1080
380
290
270
‘60
’O
1O
650
380
290
280
270
220
NN
590
380
290
285
270
265
WP
M85
538
029
028
527
0I
SN59
047
538
029
027
0B
stU
IA
All
spec
ies
1290
440
260
90A
1A
U12
0044
026
090
‘90
’P
1P
E12
9070
090
ISN
1025
440
265
260
90
Con
tinu
ed

T
III.
Con
tinu
ed
Enz
yme
Hap
loty
pea
Spec
iesb
Ban
ds(b
p)c
12
34
56
78
910
11
Dde
IB
AU
+A
llsp
ecie
s61
039
026
021
017
517
013
090
AA
U54
039
026
021
017
517
013
090
‘70
’M
1M
610
390
215
210
175
170
130
90‘4
5’
Dpn
IIA
All
spec
ies
795
310
265
200
170
130
9080
‘40
’P
PE
780
310
265
200
170
130
9080
‘40
’‘1
5’
ISN
720
310
265
200
170
130
9080
‘75
’‘4
0’
Hae
III
AA
U60
028
020
018
014
512
090
‘UI
’N
N60
040
518
014
013
512
095
90‘U
I’
OO
LB
P60
050
020
018
013
512
090
‘UI
’O
1O
600
280
220
200
180
135
120
90‘U
I’
VP
EC
S60
040
520
018
013
512
095
90‘U
I’
WP
M60
050
018
014
013
512
090
‘50
’‘U
I’
ISN
600
485
225
180
135
120
90‘U
I’
Hha
IA
AU
OC
S69
060
037
530
0G
PM
NP
EC
S12
9037
530
0I
SN69
037
535
530
024
5H
inc
IIA
AU
OP
MP
EC
SSN
1790
290
GP
NC
S20
80H
inf
IA
AU
995
505
410
170
GO
NP
EC
S99
558
050
5W
PM
580
565
505
430
W1
P56
550
544
543
013
5W
2P
580
565
510
280
150
M1
M99
550
544
513
5V
1C
S71
558
050
528
0I
SN56
550
543
041
017
0H
phI
AA
UN
CS
PE
SN11
4094
0G
OL
BP
MN
1140
880
‘60
’
Con
tinu
ed

T
III.
Con
tinu
ed
Enz
yme
Hap
loty
pea
Spec
iesb
Ban
ds(b
p)c
12
34
56
78
910
11
Msp
IA
AU
OP
CS
SN47
045
029
026
025
019
012
5‘4
5’
WO
PM
730
450
290
250
190
125
‘45
’N
NC
S45
036
029
026
025
019
012
511
0‘4
5’
N1
N45
545
026
025
019
519
011
0‘4
5’
V1
CS
470
375
290
260
250
190
125
‘75
’‘4
5’
ISN
550
470
375
250
190
125
‘75
’‘4
5’
Nci
IA
AU
OP
MP
EC
S12
7081
0I
SNO
810
750
520
O1
O12
7071
010
0N
NO
810
715
555
NN
810
715
345
210
V1
CS
1270
440
370
Rsa
IA
AU
1010
450
365
230
‘25
’O
O10
7045
023
022
511
0O
1O
550
520
450
230
225
110
O2
O77
045
028
023
022
511
0W
PM
550
520
450
350
230
NN
880
520
450
230
NN
650
520
450
230
230
PP
E52
045
036
535
023
017
0V
CS
525
520
455
365
230
Ssp
IA
All
spec
ies
2080
G1
OM
1400
680
Taq
IA
All
spec
ies
660
525
510
215
130
‘40
’O
1O
525
510
495
215
165
130
‘40
’O
2O
740
660
510
130
‘40
’G
PM
660
525
510
150
130
‘65
’‘4
0’
V1
CS
525
510
460
215
200
130
‘40
’
aA
,cha
ract
eris
tic
toA
rcti
cci
sco
orco
mm
onin
alls
peci
es;G
,com
mon
inal
lspe
cies
ifdi
ffer
ent
from
Arc
tic
cisc
o;O
,com
mon
inB
aika
lom
ul;P
,com
mon
inpe
led;
V,
com
mon
inle
ast
cisc
o;W
,co
mm
onin
whi
tefis
hes;
I,co
mm
onin
inco
nnu;
num
bere
dha
plot
ypes
(A1,
O1,
W2
etc.
)ar
ele
ssco
mm
onin
corr
espo
ndin
gsp
ecie
s.bA
U,
Arc
tic
cisc
o;M
,m
uksu
n;N
,br
oad
whi
tefis
h;O
,B
aika
lom
ul;
P,
Sibe
rian
whi
tefis
h;P
E,
pele
d;C
S,le
ast
cisc
o;SN
,in
conn
u.c F
ragm
ents
not
visi
ble
onge
lar
egi
ven
insi
ngle
quot
es.

61
Baikal whitefishC. lavaretus baicalensis has haplotype O1 which is the most frequent haplotype
in Baikal omul and absent from any other whitefishes. This suggests that theBaikal whitefish is a form of omul, C. migratoruis, at least with respect tomtDNA. Further examination of nuclear markers and morphology is needed todetermine whether its sub-terminal mouth, typical to whitefishes, evolvedindependently or through introgression with true whitefishes (e.g. C. lavaretuspidschian, which is also present in Baikal Lake). Sukhanova et al. (2000) haveshown extremely high sequence similarity among Baikal omul and Baikal lakewhitefish in both the cytochrome b gene and the control region of mtDNA.
C. nasus and other whitefishesAs a group, the whitefishes (Coregonus with sub-terminal mouths) possess
allozyme alleles that are alternative to those that distinguish other species.Clustering based on allozyme data revealed a clade that included muksun C.muksun (Pallas), Siberian and broad whitefish that had high bootstrap support.However, mtDNA haplotypes of broad whitefish do not cluster together withmuksun and Siberian whitefish on the MST (Fig. 3), this fact raising a questionabout the monophyletic origin of this group. Introgression also might accountfor closer similarity among whitefishes at allozyme loci than for mtDNAhaplotypes.
Among the other whitefishes, reliable identification is possible only for thebroad whitefish. This species can be differentiated clearly from both C. muksun
Baikal omul. This species can be identified reliably by Hae III O and O1haplotypes and Rsa I O, O1 and O2 haplotypes as well (Fig. 2). The most similarhaplotypes in Arctic cisco (A1) and Baikal omul (O5) differed by nine restrictionsites. For comparison, a minimum of 12 pairwise differences were observedbetween Baikal omul and Siberian whitefish C. lavaretus pidschian Gmelin, 11with least cisco and 13 with peled. It is noteworthy that often Baikal omul istreated currently as a sub-species of C. autumnalis (Berg, 1948; Nikolsky &Reshetnikov, 1970; Reshetnikov, 1980). An alternative classification assumes itsstatus as a full species C. migratorius (GTsowska, 1960; Shaposhnikova, 1968,1970). Present data indicate that it is as distinct genetically in both isozymes andmtDNA (Table II–IV, see also Politov et al., 2000) as other commonlyrecognized coregonid species, so its status as a sub-species of C. autumnalisshould be rejected. Likewise, Sukhanova et al. (2000) found sequences of D-loopand cytochrome b regions of mtDNA in Baikal omul to be much more similar towhitefishes than Arctic cisco. It is noteworthy that Baikal omul was the mosthighly variable species among those studied (Table IV); 10 haplotypes werefound among 40 individuals. Considering these facts, a currently popularhypothesis of the origin of this form from late-Pleistocene Arctic cisco migrantsfrom the Lena River or the Yenisey River seems to be unrealistic. It is veryunlikely that such a great intraspecific diversity appeared in postglacial time.The MST indicates Arctic cisco to be a terminal taxon, and if Baikal omul andArctic cisco do form a monophyletic clade (which is also questionable) it is morelikely that Baikal omul is a pre-Pleistocene ancestor of Arctic cisco rather thanits descendant.

T
IV.
Dis
trib
utio
nof
mtD
NA
hapl
otyp
esam
ong
spec
ies
and
spec
ies
diag
nose
sba
sed
onfie
ldex
amin
atio
n(e
xter
nal
mor
phol
ogy)
and
gene
tic
data
Hap
loty
pe
Ref
eren
cesp
ecim
enco
denu
mbe
r
Num
ber
insp
ecie
ssa
mpl
e
Fie
ldsp
ecie
sdi
agno
sis
Allo
zym
edi
agno
sis
MtD
NA
diag
nosi
sA
seI
Ava
IIB
saJ
IB
sp12
86I
Bst
NI
Bst
UI
Dde
ID
pnII
Hae
III
Hha
IH
inc
IIH
inf
IH
phI
Msp
IN
ciI
Rsa
IS
spI
Taq
I
AU
6845
0A
U68
1au
tum
nalis
autu
mna
lisau
tum
nalis
AA
AA
AA
1B
AA
AA
AA
AA
AA
A
AU
141
AU
158
autu
mna
lisau
tum
nalis
autu
mna
lisA
AA
AA
AB
AA
AA
AA
AA
AA
A
AU
5134
9A
U51
1au
tum
nalis
autu
mna
lisau
tum
nalis
AA
AA
AA
BA
AA
AA
AA
AA
G1
A
AU
5335
1AU
531
autu
mna
lisau
tum
nalis
autu
mna
lisA
AA
AA
AB
AA
AG
AA
AA
AA
A
AU
6039
3A
U60
1au
tum
nalis
autu
mna
lisau
tum
nalis
AA
AA
1A
AB
AA
AA
AA
AA
AA
A
AU
6139
4A
U61
2au
tum
nalis
autu
mna
lisau
tum
nalis
AA
AA
AA
A1
AA
AA
AA
AA
AA
A
O1
701
O1
21m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GA
OO
AB
AO
AA
GG
AA
OA
A
LB
11
l.ba
ikal
ensi
sn/
am
igra
tori
usA
GA
OO
AB
AO
AA
GG
AA
OA
A
O4
704
O4
5m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GA
OO
AB
AO
AA
GG
WA
OA
A
LB
21
l.ba
ikal
ensi
sn/
am
igra
tori
usA
GA
OO
AB
AO
AA
GG
WA
OA
A
O5
705
O5
5m
igra
tori
usm
igra
tori
usm
igra
tori
usA
AA
AO
AB
AO
1A
AG
GA
AO
AA
O6
706
O6
1m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GA
OO
AB
AO
AA
GG
AO
1O
AA
O9
709
O9
2m
igra
tori
usm
igra
tori
usm
igra
tori
usA
AA
AO
1A
BA
O1
AA
GG
AA
O1
AA
O11
711
O11
1m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GA
OO
AB
AO
AA
GG
WA
O2
AA
O22
722
O22
1m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GA
OO
AB
AO
AA
GG
WA
OA
O2
O24
724
O24
1m
igra
tori
usm
igra
tori
usm
igra
tori
usA
WA
AO
AB
AO
1A
AG
GA
NO
AO
2
O31
731
O31
2m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GO
1O
OA
BA
OA
AG
GA
IO
AA
O36
736
O36
1m
igra
tori
usm
igra
tori
usm
igra
tori
usA
GA
OO
AB
AO
AA
GG
AA
OA
O1
P1
46P
12
l.pi
dsch
ian
whi
tefis
hw
hite
fish
GG
WG
WA
BA
OG
AW
1G
WA
WA
A
P3
460
P3
8l.
pids
chia
nw
hite
fish
whi
tefis
hG
WW
WW
AB
AW
GA
WG
WA
WA
A
18M
19
muk
sun
whi
tefis
hw
hite
fish
GW
WW
WA
BA
WG
AW
GW
AW
AA
162
N73
1na
sus
whi
tefis
hw
hite
fish
GW
WW
WA
BA
WG
AW
GW
AW
AA
453
AU
711
autu
mna
lishy
brid
AU
�P
whi
tefis
hG
WW
WW
AB
AW
AA
WG
WA
WA
A
P97
186
P97
5l.
pids
chia
nw
hite
fish
whi
tefis
hG
GW
GW
AB
AO
GA
W2
GW
AW
AG
122
M31
1m
uksu
nw
hite
fish
whi
tefis
hG
GW
GW
AB
AO
GA
W2
GW
AW
AG
P50
123
4P
501
1l.
pids
chia
nw
hite
fish
whi
tefis
hG
GW
GW
AB
AW
2G
AW
2G
WA
WA
G
P50
223
5P
502
1l.
pids
chia
nw
hite
fish
whi
tefis
hG
GW
GW
AB
AO
GA
W2
AW
AW
AG
M27
257
P52
41
l.pi
dsch
ian
whi
tefis
hw
hite
fish
GW
WW
WA
BA
WG
AW
AW
AW
AA
118
M27
2m
uksu
nw
hite
fish
whi
tefis
hG
WW
WW
AB
AW
GA
WA
WA
WA
A
226
CS9
051
autu
mna
lissa
rdin
ella
whi
tefis
hG
WW
WW
AB
AW
GA
WA
WA
WA
A
Con
tinu
ed
T
IV.
Con
tinu
ed
Hap
loty
pe
Ref
eren
cesp
ecim
enco
denu
mbe
r
Num
ber
insp
ecie
ssa
mpl
e
Fie
ldsp
ecie
sdi
agno
sis
Allo
zym
edi
agno
sis
MtD
NA
diag
nosi
sA
seI
Ava
IIB
saJ
IB
sp12
86I
Bst
NI
Bst
UI
Dde
ID
pnII
Hae
III
Hha
IH
inc
IIH
inf
IH
phI
Msp
IN
ciI
Rsa
IS
spI
Taq
I
M4
21M
41
muk
sun
whi
tefis
hw
hite
fish
GW
WW
WA
M1
AW
GA
M1
GW
AW
AA
M29
120
M29
1m
uksu
nw
hite
fish
whi
tefis
hG
WW
WW
AB
AW
GG
WG
WA
WA
A
M30
121
M30
1m
uksu
nw
hite
fish
whi
tefis
hG
WW
WW
AB
AM
1G
GW
GW
AW
AA
M35
231
M35
1m
uksu
nw
hite
fish
whi
tefis
hG
WW
WW
AB
AW
GA
WG
WA
WG
1A
N1
23N
19
nasu
sna
sus
nasu
sA
WA
NN
AB
AN
GA
GA
NN
NA
A
N13
60N
137
nasu
sna
sus
nasu
sA
WA
NN
AB
AN
GG
GG
NN
NA
A
N16
64N
163
nasu
sna
sus
nasu
sA
WA
GG
AB
AN
GG
GG
NN
NA
A
N17
65N
173
nasu
sna
sus
nasu
sA
WA
GN
AB
AN
GG
GG
NN
NA
A
468
P11
1l.
pids
chia
nw
hite
fish
nasu
sA
WA
GG
AB
AN
GG
GG
NN
NA
A
N21
159
N21
1na
sus
nasu
sna
sus
AW
AG
NA
BA
WG
GG
GN
NN
AA
N70
159
N70
8na
sus
nasu
sna
sus
AW
AN
GA
BA
NG
GG
GN
NN
AA
N14
236
1N
142
1na
sus
nasu
sna
sus
AW
N1
NN
AB
AN
GG
GG
N1
N1
N1
AA
N14
536
4N
145
1na
sus
nasu
sna
sus
AW
N2
NG
AB
AN
GG
GG
NN
NA
A
CS
125
CS
14
autu
mna
lissa
rdin
ella
sard
inel
laG
GA
GG
AB
AV
GA
GA
AA
PA
V1
CS
966
448
CS9
664
autu
mna
lissa
rdin
ella
sard
inel
laG
GA
GG
AB
AV
GA
GA
AA
PA
A
CS
212
4C
S2
4au
tum
nalis
sard
inel
lasa
rdin
ella
GG
AG
GA
BA
VG
AG
AA
AV
AA
230
M34
1m
uksu
nw
hite
fish
sard
inel
laG
GA
GG
AB
AV
GA
GA
AA
VA
A
CS
312
5C
S3
1sa
rdin
ella
sard
inel
lasa
rdin
ella
GV
1A
GG
AB
AV
GA
GA
V1
V1
VA
A
CS
612
8C
S6
2sa
rdin
ella
sard
inel
lasa
rdin
ella
AG
AG
GA
BA
VG
AG
AA
AV
AA
CS
3523
3C
S35
1sa
rdin
ella
sard
inel
lasa
rdin
ella
GG
AG
GA
BA
VG
GG
AA
AV
AA
227
CS9
061
autu
mna
lissa
rdin
ella
sard
inel
laG
GA
GG
AB
AV
GG
GA
AA
VA
A
170
N81
1na
sus
whi
tefis
hsa
rdin
ella
GG
AG
GA
BA
VG
GG
AA
AV
AA
229
M33
2m
uksu
nw
hite
fish
sard
inel
laG
GA
GG
AB
AV
GG
GA
AA
VA
A
PE
180
1P
E1
8pe
led
n/a
pele
dG
GA
WG
AB
PV
GA
GA
AA
PA
A
PE
580
5P
E5
1pe
led
n/a
pele
dG
GA
GG
AB
PV
GA
GA
AA
PA
A
PE
3680
7P
E36
1pe
led
n/a
pele
dG
GA
P1
GA
BP
VG
AG
AA
AP
1A
A
PE
937
810
PE
937
2pe
led
n/a
pele
dA
GA
WG
P1
BP
VG
GG
AA
AP
AA
SN12
158
1215
8SN
3in
conn
un/
ain
conn
uG
AI
II
IA
II
IA
II
II
WA
G
SN12
237
1223
7SN
1in
conn
un/
ain
conn
uA
AI
II
IA
II
IA
II
II
WA
G

64 . . .
F. 1. Creatine kinase zymogram showing characteristic alleles of CK-1,2* isoloci (A–D) and position ofminor CK3 band in different species (AU, C. autumnalis; O, C. migratorius; P, C. lavaretuspidschian; M, C. muksun; N, C. nasus; CS, C. sardinella).
T V. Mean (below diagonal) and minimum (above diagonal) number of non-sharedrestriction sites among OTUs
Species AU O P+M N CS PE SN
C. autumnalis (AU) 2·00 9 20 20 15 17 23C. migratorius (O) 13·60 3·96 12 15 11 13 23C. l. pidschian+C. muksun (P+M) 22·19 15·50 4·56 12 8 9 23C. nasus (N) 23·96 19·45 16·68 4·39 10 12 31C. sardinella (CS) 17·11 14·50 13·03 14·54 2·76 1 26C. peled (P E) 17·83 15·20 12·80 15·08 5·75 3·50 26S. l. nelma (SN) 25·17 25·10 26·80 33·88 27·17 27·00 1·00
and C. lavaretus pidschian by the absence of the CK� band (Fig. 1), that isotherwise always present in muksun and Siberian whitefish. It has species-specific alleles at ACO-2* and mMDHP-1,2*. Broad whitefish also carries afixed allele C at sMDHP-3,4* isoloci (coding for supernatant malic enzyme),while C. muksun has different fixed alleles at sMDHP-3* and sMDHP-4*, andthe Siberian whitefish is polymorphic for three alleles. Frequencies of theEST-1*B allele and GPI-B1,2*C in broad whitefish are significantly higher thanin other whitefishes.
ND-1 RFLP patterns also separate broad whitefish clearly from othercoregonids. All studied specimens demonstrate a unique haplotype N when thefragment was digested by Hae III and Rsa I (Fig. 2). Patterns produced by MspI and Nci I (N haplotypes, Tables III and IV) are different from all other speciesand occur only as rare variants in least cisco and Baikal omul, respectively. Ingeneral, the distinctness of C. nasus in allozymes and mtDNA agrees well withthe results of comparative morphological (Lindsey, 1962) and karyological(Viktorovsky & Ermolenko, 1982) studies. Earlier allozyme (Bodaly et al., 1991,1994; Ermolenko, 1992; Vuorinen et al., 1998) and mtDNA studies showed this

F
.2.
Res
tric
tion
patt
erns
ofP
CR
-am
plifi
edN
D-1
frag
men
tof
mtD
NA
prod
uced
byR
saI
indi
ffer
ent
core
goni
dsp
ecie
s.H
aplo
type
code
s(t
op)
corr
espo
ndto
Tab
les
IIan
dII
I.Sp
ecie
sby
lane
s:1–
9,C
.mig
rato
rius
;10–
12,C
.lav
aret
uspi
dsch
ian;
13–2
0,C
.muk
sun;
21–2
6,C
.nas
us;2
7–28
,C.p
eled
;29,
C.a
utum
nalis
;30
–31,
C.
sard
inel
la.
66 . . .
species to be a part of the whitefish clade. Intraspecific variation in broadwhitefish was relatively high (Tables IV and V).
F. 3. Minimum spanning network among haplotypes represented in Tables III and IV. Number ofpairwise differences is given for some interspecific connections. Inconnu haplotypes are connectedto main network at four different points by 23 pairwise differences.
C. muksun and C. lavaretus pidschianAlthough these two whitefish forms are recognized currently as different
species they shared the most frequent alleles in all allozyme loci. Differencesbetween them were restricted to presence or absence of particular low-frequencyalleles (Table II). For instance, allele EST-1* B is absent in muksun but reachesa frequency of 0·10 in Siberian whitefish. Alleles mIDHP-1*A (P=0·01) andmIDH-2*A (P=0·16) were found exclusively in muksun. Allele sSOD*Ais common in muksun but was found in just one heterozygote in the Siberianwhitefish sample. Principal component analysis (PCA) of allozyme variationdid not reveal distinct clusters corresponding to the nominate species (Politovet al., 2000). This contrasted with other inter-species comparisons where PCAdiscriminated individuals of different species efficiently (Politov et al., 2000).

67
RFLP analysis revealed three haplotypes (P3, P97 and M27) found in bothspecies and some low frequency haplotypes specific to either muksun or toSiberian whitefish (Table IV). Present results (Fig. 3) correspond well withearlier allozyme (Ermolenko, 1992) and mtDNA (Bernatchez & Dodson, 1994;Reist et al., 1998) studies where muksun clustered among forms belonging to theC. lavaretus group but did not form a sister clade to them.
Identification of muksun and Siberian whitefish was impossible by any genetictechniques employed. Individuals identified morphologically as C. muksun andC. lavaretus pidschian are genetically very similar. Relatively easy morphologicalrecognition of muksun is possible by its higher gill raker counts and wider snout(Berg, 1948). However, these traits, especially gill raker number are distinctiveonly when muksun is compared with sympatric low gill-rakered Siberianwhitefish. Some North European whitefish forms of the C. lavaretus lavaretuscomplex (i.e., conspecific to Siberian whitefish) have counts overlapping withmuksun (e.g. Berg, 1948). At present it is difficult to decide whether the muksunis a distinct species, or just one of the numerous ecotypes of the polymorphicspecies C. lavaretus. Conspecificity and monophyletic origin of muksun popu-lations from different parts of the nominal species’ range should be confirmedfurther. If muksun and Siberian whitefish do represent separate species, theirlow genetic differentiation may indicate that gene flow between them has ceasedrecently.
Interspecific v. intraspecific variationFrom Table V we calculated that, on average, mtDNA haplotypes differed at
3·16 sites within species v. 18·87 sites between species. Muksun and Siberianwhitefishes that share some haplotypes were combined here in a single OTU(operational taxonomic unit). Besides these whitefishes, only peled P5 and leastcisco CS1 had just one non-shared site followed by CS1 v. Siberian whitefishP502 (eight sites), Arctic cisco AU1 v. Baikal omul O5 (nine sites). In all otherpairs the differences were even greater. Judging from this, misidentification ofcoregonid species is very unlikely with the markers used. The above-mentionedexceptions represent cases of recently diverged forms that would requireemployment of additional highly variable genetic markers.
Introgressive hybridization and identification of coregonid speciesIn several cases allozyme and mtDNA diagnoses did not correspond with the
initial morphological ones (Table IV). The least cisco haplotypes CS1 and CS35appeared in muksun specimens 230M34 and 229M33, respectively. Broadwhitefish haplotype N17 was found in the individual identified as a Siberianwhitefish 468P11. Composite genotype P3 was found not only in eight Siberianwhitefish and nine muksun specimens but also in one specimen each identifiedinitially as least cisco, broad whitefish, and Arctic cisco. The Arctic ciscospecimen 453AU71 carried several allozyme alleles typical for whitefishes.
As these examples illustrate, the presence of haplotypes typical for anotherspecies might be caused by mis-identification of species in the field, or byintrogression among species. These two possible causes are not necessarilyindependent, as mis-identification is more likely among individuals withintermediate morphology, which, in turn, reflects probable hybrid origin.

68 . . .
Hybridization is reported frequently among coregonid species (Berg, 1948;Svardson, 1998) and certified by genetic studies (Luczynski et al., 1992;Reist et al., 1992; Bickham et al., 1997). Vertebrate mtDNA has maternalinheritance, and when used alone reflects only partly interspecific gene flow.Employment of different types of genetic markers (especially nuclear ones) isdesired and helpful in inferring both possible parents (Carr et al., 1986; Catheyet al., 1998) and, potentially, generation of the hybrid (F1, F2, different types ofbackcrosses etc.).
CONCLUSIONS
Allozyme loci and RFLP analysis of PCR-amplified ND-1 fragments ofmitochondrial DNA proved to be reliable for the discrimination of coregonidspecies. Creatine kinase zymograms, and restriction digestions with enzymes RsaI and Dpn II discriminated among all the studied Palearctic Coregonus speciesexcept for the muksun–Siberian whitefish pair. The reliability of identificationincreased with more genetic markers employed. Application of at least two typesof markers (e.g. maternally inherited mtDNA and nuclear genes like isozymeloci) is especially helpful in the identification of hybrids. Non-typical mtDNAhaplotypes for a particular species might have originated from another species asa result of past or recent introgression. Taking hybridization into account is ofgreat importance not only theoretically for understanding evolutionary path-ways in fish (Altukhov, 1982; Smith, 1992) but also practically for geneticidentification of coregonids (Bickham et al., 1997).
The genetic techniques used here are proposed not as replacements oftraditional morphological keys, but to assist taxonomists to create new, morepowerful methods of identification. Morphological keys should be based oncarefully selected characters that appear in all typical representatives of par-ticular species. Genetic methods can help to select such representatives that werenot subjected to interspecific crossing. Deviated or intermediate forms such ashybrids can also be revealed by molecular genetic markers, morphologicallyre-examined and included into such key tables as separate items.
Unlike most morphological methods, it is possible to use genetic techniqueswhen whole fish are unavailable. Small pieces of frozen tissue or even tissuepreserved at room temperature in DMSO/sodium chloride solution can be usedsuccessfully for identification. This feature is important when populations andspecies are overexploited and fisheries management requires monitoring ofgenetic resources. Commercial catches, where often samples available are onlyheadless fillets, salted or otherwise preserved fish products, may be subjected togenetic analysis for species composition and proportion of different species in thetotal product.
Further development of powerful new genetic methods of identification willensure differentiation not only among species but perhaps also among localpopulations or stocks.
The authors thank sponsors, organizers and participants of the expedition ‘ TundraEcology–94 ’, the crew of ‘ Akademik Fedorov ’ research vessel, and to two helicoptercrews for their invaluable help during field work; and V. V. Smirnov and L. Yu.Yampolsky for assistance in obtaining and transporting Baikal omul samples. The

69
laboratory treatment of samples was supported by the Russian Foundation for BasicResearch, projects #96-04-49174 and #95-04-12614a, and by the Program for Support ofthe Scientific Schools, grant #96-15-97862. This paper represents contribution #00 of theCenter for Biosystematics and Biodiversity, Texas A&M University.
References
Altukhov, Yu. P. (1982). Biochemical population genetics and speciation. Evolution 36,1168–1181.
Berg, L. S. (1948). Freshwater Fishes of USSR and Adjacent Countries Vol. 1. Moscow–Leningrad: Izdatel’stvo Akademii Nauk USSR (in Russian).
Bernatchez, L. & Dodson, J. J. (1991). Phylogeographic structure in mitochondrial DNAof the lake whitefish (Coregonus clupeaformis) and its relation to Pleistoceneglaciations. Evolution 45, 1016–1035.
Bernatchez, L. & Dodson, J. J. (1994). Phylogenetic relationships among Palearcticand Nearctic whitefish (Coregonus sp.) populations as revealed by mitochondrialDNA variation. Canadian Journal of Fisheries and Aquatic Sciences 51,240–251.
Bernatchez, L., Colombani, F. & Dodson, J. J. (1991). Phylogenetic relationships amongthe subfamily Coregoninae as revealed by mitochondrial DNA restriction analysis.Journal of Fish Biology 39, 283–290.
Bickham, J. W., Carr, S. M., Hanks, B. G., Burton, D. W. & Gallaway, B. J. (1989).Genetic analysis of population variation in the Arctic cisco (Coregonus autumnalis)using electrophoretic, flow cytometric, and mitochondrial DNA restrictionanalyses. Biological Papers of the University of Alaska 24, 112–122.
Bickham, J. W., Patton, J. C., Minzenmayer, S., Moulton, L. L. & Gallaway, B. J. (1997).Identification of Arctic and Bering ciscoes in the Colville River delta, Beaufort SeaCoast, Alaska. In Fish Ecology in Arctic North America (Reynolds, J. B., ed.),pp. 224–228. Bethesda, MD: American Fisheries Society.
Bodaly, R. A., Vuorinen, J., Ward, R. D., Luczynski, M. & Reist, J. D. (1991). Geneticcomparisons of New and Old World coregonid fishes. Journal of Fish Biology 38,37–51.
Bodaly, R. A., Vuorinen, D. A., Reshetnikov, Yu. S. & Reist, J. D. (1994). Geneticrelationships between five species of coregonid species from Siberia. Journal ofIchthyology 34, 117–130.
Carr, S. M., Ballinger, S. W., Derr, J. N., Blankenship, L. H. & Bickham, J. W. (1986).Mitochondrial DNA analysis of hybridization between sympatric whitetailed andmule deer in west Texas. Proceedings of the National Academy of Sciences USA 83,9576–9580.
Cathey, J. C., Bickham, J. W. & Patton, J. C. (1998). Introgressive hybridization andnonconcordant evolutionary history of maternal and paternal lineages in NorthAmerican deer. Evolution 52, 1224–1229.
Chereshnev, I. A. (1994). Comparative craniology of Holarctic omuls. In Biology &Biotechniques of Coregonid Fish Reproduction, pp. 157–160. St. Petersburg:GosNIORKH.
Cronin, M. A., Spearman, W. J., Wilmot, R. L., Patton, J. C. & Bickham, J. W. (1993).Mitochondrial DNA variation in chinook (Oncorhynchus tshawytscha) and chumsalmon (O. keta) detected by restriction enzyme analysis of polymerase chainreaction (PCR) products. Canadian Journal of Fisheries and Aquatic Sciences 50,708–715.
Douglas, M. R., Brunner, P. C. & Bernatchez, L. (1999). Do assemblages of Coregonus(Teleostei: Salmoniformes) in the Central Alpine region of Europe representspecies flocks? Molecular Ecology 8, 589–603.
Ermolenko, L. N. (1992). Genetic divergence in the family Coregonidae. PolskieArchiwum Hydrobiologii 39, 533–539.

70 . . .
GTsowska, M. (1960). Genus Coregonus L. discussed in connection with a newsystematic feature that of shape and proportion of os maxillare and os supramaxil-lare. Annales Zoologici XVIII, 471–513.
Grønlund, E. & Melander, O. (Eds) (1995). Swedish-Russian Tundra Ecology-Expedition—94. A Cruise Report. Stockholm: Swedish Polar Research Secretariat.
Hamada, M., Himberg, M., Bodaly, R. A., Reist, J. D. & Okada, N. (1998).Monophyletic origin of the genera Stenodus and Coregonus as inferred from ananalysis of the insertion of SINEs (short interspersed repetitive elements). Archivfur Hydrobiologie Special Issues Advances in Limnology 50, 383–389.
Lindsey, C. C. (1962). Distinction between the broad whitefish, Coregonus nasus, andother North American coregonids. Journal of the Fisheries Research Board ofCanada 19, 687–714.
Lindsey, C. C. (1981). Stocks are chameleons: plasticity in gill rakers of coregonid fishes.Canadian Journal of Fisheries and Aquatic Sciences 38, 1491–1506.
Lockwood, S. F., Dilinger, R. E. J., Birt, T. P., Green, J. M. & Snyder, T. P. (1993).Phylogenetic relationships among members of the Coregoninae inferred fromdirect sequencing of PCR-amplified mitochondrial DNA. Canadian Journal ofFisheries and Aquatic Sciences 50, 2112–2118.
Luczynski, M., Falkowski, S., Vuorinen, J. & Jankun, M. (1992). Genetic identificationof European whitefish (Coregonus lavaretus), peled (C. peled ) and their hybridsin spawning stocks of ten Polish lakes. Polskie Archiwum Hydrobiologii 39,571–577.
Maiers, L. D., Carmichael, T. J., Reist, J. D. & Bodaly, R. A. (1998). Enhanced recoveryof DNA from frozen fish tissues treated with dimethyl sulphoxide (DMSO).Archiv fur Hydrobiologie Special Issues Advances in Limnology 50, 371–374.
Morales, J. C., Hanks, B. G., Bickham, J. W., Derr, J. N. & Gallaway, B. J. (1993).Allozyme analysis of population structure in Arctic cisco (Coregonus autumnalis)from the Beaufort Sea. Copeia 1993, 863–867.
Nikolsky, G. W. & Reshetnikov, Yu. S. (1970). Systematics of coregonid fishes in theUSSR: intraspecies variability and difficulties in taxonomy. Biology of CoregonidFish (Lindsey, C. C. & Woods, C. S., eds), pp. 251–266. Winnipeg, Manitoba:University of Manitoba Press.
Novikov, G. G., Afanasiev, K. I., Fernholm, B., Levenko, B. A., Makhrov, A. A.,Malinina, T. V., Politov, D. V. & Tseitlin, D. G. (1995). Ichthyological Investi-gation in the Arctic. In Swedish-Russian Tundra Ecology-Expedition—94. A CruiseReport (Grønlund, E. & Melander, O., eds), pp. 387–388. Stockholm: SwedishPolar Research Secretariat.
Novikov, G. G., Politov, D. V., Makhrov, A. A., Malinina, T. V., Afanasiev, K. I. &Fernholm, B. (2000). Freshwater and Estuarine Fishes of the Russian ArcticCoast in Collections of Swedish–Russian Expedition ‘ Tundra Ecology—94 ’.Journal of Fish Biology Supplement 57 (Suppl. A), 158–162.
Politov, D. V. & Gordon, N. Yu. (1998). Taxonomic status of the Baikal omul inview of isozyme analysis data. In Modern Problems in Fish Taxonomy, p. 72.St Petersburg, Russia: Zoological Institute (in Russian).
Politov, D. V., Omelchenko, V. T., Salmenkova, E. A. & Malinina, T. V. (1998). Geneticdifferentiation of Arctic and Far Eastern Chars of the Genus Salvelinus. RussianJournal of Genetics 34, 71–79.
Politov, D. V., Gordon, N. Yu. & Makhrov, A. A. (2000). Genetic identificationand taxonomic relationships of six Siberian Coregonus species. Archiv furHydrobiologie Special Issues–Advances in Limnology (in press).
Reist, J. D., Vuorinen, J. & Bodaly, R. A. (1992). Genetic and morphologicalidentification of coregonid hybrid fishes from Arctic Canada. Polskie ArchiwumHydrobiologii 39, 551–561.
Reist, J. D., Maiers, L. D., Bodaly, R. A., Vuorinen, J. A. & Carmichael, T. J. (1998).The phylogeny of New and Old World coregonine fishes as revealed by sequencevariation in a portion of the d-loop of mitochondrial DNA. Archiv furHydrobiologie Special Issues–Advances in Limnology 50, 323–339.

71
Reshetnikov, Yu. S. (1980). Ecology and Systematics of Coregonid Fish. Moscow: Nauka(in Russian).
Reshetnikov, Yu. S. (Ed.) (1998). Annotated Check-List of Cyclostomata and Fishes of theContinental Waters of Russia. Moscow: Nauka (in Russian).
Sajdak, S. L. & Phillips, B. (1997). Phylogenetic relationships among Coregonus speciesinferred from the DNA sequence of the first internal transcribed spacer (ITS1)of ribosomal DNA. Canadian Journal of Fisheries and Aquatic Sciences 54,1494–1503.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecular Cloning: a LaboratoryManual. Cold Spring Harbor, USA: Cold Spring Harbor Laboratory Press.
Schneider, S., Roessli, D. & Excoffier, L. (2000). Arlequin: A software for populationgenetics data analysis. Ver 2.000. Geneva: Genetics and Biometry Lab., Dept. ofAnthropology, University of Geneva.
Shaklee, J. B., Allendorf, F. W., Morizot, D. C. & Whitt, G. S. (1990). Genenomenclature for protein-coding loci in fish. Transactions of the AmericanFisheries Society 119, 2–15.
Shaposhnikova, G. K. (1968). Comparative morphology of the whitefishes(Coregoninae) from the USSR. In Morphology of Lower Vertebrates. TrudyZoologicheskogo Instituta 36, 207–257 (in Russian).
Shaposhnikova, G. K. (1970). On the taxonomy of whitefishes from the USSR. InBiology of Coregonid Fish (Lindsey, C. C. & Woods, C. S., eds), pp. 195–207.Winnipeg: University of Manitoba Press.
Smith, G. R. (1992). Introgression in fishes: significance for paleontology, cladistics, andevolutionary rates. Systematic Biology 41, 41–57.
Smith, G. R. & Todd, T. D. (1992). Morphological cladistic study of coregonine fishes.Polskie Archiwum Hydrobiologii 39, 479–490.
Sukhanova, L. V., Smirnov, V. V., Smirnova-Zalumi, N. S., Kirilchik, S. V., Griffits, D.& Belikov, S. I. (2000). The taxonomic position of the Lake Baikal omulCoregonus autumnalis migratorius (Georgi) as revealed by sequence analysis ofmtDNA cytochrome b and control region. Archiv fur Hydrobiologie SpecialIssues–Advances in Limnology (in press).
Svardson, G. (1998). Postglacial dispersal and reticulate evolution of Nordic coregonids.Nordic Journal of Freshwater Research 74, 3–32.
Viktorovsky, R. M. & Ermolenko, L. N. (1982). The chromosomal complexes ofCoregonus nasus and C. lavaretus and the problem of the Coregonus karyotypedivergence. Tsitologiya 24, 797–801 (in Russian).
Vuorinen, J. (1984). Electrophoretic expression of genetic variation and duplicate geneactivity in vendace, Coregonus albula (Salmonidae). Hereditas 101, 85–96.
Vuorinen, J. & Piironen, J. (1984). Inheritance and joint segregation of biochemical lociin European whitefish, genus Coregonus. Hereditas 101, 97–102.
Vuorinen, J. A., Bodaly, R. A., Reist, J. D. & Luczynski, M. (1998). Phylogeny of fiveProsopium species with comparisons with other coregonine fishes based on isozymeelectrophoresis. Journal of Fish Biology 53, 917–927.




















