
Volunteer Career Mentor Binder - Aligned Impact Muscatine ...
Upload
univ-bordeauxCategory
view
3download
0
Available online at www.sciencedirect.com
Biochimica et Biophysica Acta 1768 (2007) 2979–3000www.elsevier.com/locate/bbamem
Review
Solid-State 2H NMR spectroscopy of retinal proteins in aligned membranes
Michael F. Brown a,b,c,⁎, Maarten P. Heyn c,d, Constantin Job a, Suhkmann Kim e, Stephan Moltke a,Koji Nakanishi f, Alexander A. Nevzorov g, Andrey V. Struts a,
Gilmar F. J. Salgado c,1, Ingrid Wallat d
a Department of Chemistry, University of Arizona, Tucson, Arizona 85721, USAb Department of Physics, University of Arizona, Tucson, Arizona 85721, USA
c Department of Biochemistry and Molecular Biophysics, University of Arizona, Tucson, Arizona 85721, USAd Biophysics Group, Department of Physics, Freie Universität Berlin, D-14195 Berlin, Germany
e Department of Chemistry and Chemistry Institute for Fundamental Materials, Pusan National University, Busan 609-735, Koreaf Department of Chemistry, Columbia University, New York, New York 10027, USA
g Department of Chemistry, North Carolina State University, Raleigh, North Carolina 27695, USA
Received 8 August 2007; received in revised form 10 October 2007; accepted 10 October 2007Available online 23 October 2007
Abstract
Solid-state 2H NMR spectroscopy gives a powerful avenue to investigating the structures of ligands and cofactors bound to integral membraneproteins. For bacteriorhodopsin (bR) and rhodopsin, retinal was site-specifically labeled by deuteration of the methyl groups followed byregeneration of the apoprotein. 2H NMR studies of aligned membrane samples were conducted under conditions where rotational and translationaldiffusion of the protein were absent on the NMR time scale. The theoretical lineshape treatment involved a static axial distribution of rotating C–C2H3 groups about the local membrane frame, together with the static axial distribution of the local normal relative to the average normal.Simulation of solid-state 2H NMR lineshapes gave both the methyl group orientations and the alignment disorder (mosaic spread) of themembrane stack. The methyl bond orientations provided the angular restraints for structural analysis. In the case of bR the retinal chromophore isnearly planar in the dark- and all-trans light-adapted states, as well upon isomerization to 13-cis in the M state. The C13-methyl group at the“business end” of the chromophore changes its orientation to the membrane upon photon absorption, moving towards W182 and thus driving theproton pump in energy conservation. Moreover, rhodopsin was studied as a prototype for G protein-coupled receptors (GPCRs) implicated inmany biological responses in humans. In contrast to bR, the retinal chromophore of rhodopsin has an 11-cis conformation and is highly twisted inthe dark state. Three sites of interaction affect the torsional deformation of retinal, viz. the protonated Schiff base with its carboxylate counterion;the C9-methyl group of the polyene; and the β-ionone ring within its hydrophobic pocket. For rhodopsin, the strain energy and dynamics of retinalas established by 2H NMR are implicated in substituent control of activation. Retinal is locked in a conformation that is twisted in the direction ofthe photoisomerization, which explains the dark stability of rhodopsin and allows for ultra-fast isomerization upon absorption of a photon.Torsional strain is relaxed in the meta I state that precedes subsequent receptor activation. Comparison of the two retinal proteins using solid-state2H NMR is thus illuminating in terms of their different biological functions.© 2007 Elsevier B.V. All rights reserved.
Keywords: Bacteriorhodopsin; G protein-coupled receptor; Membrane; Proton pump; Solid-state NMR; Retinal; Rhodopsin; Vision
Abbreviations: bR, bacteriorhodopsin; bRall-t, bacteriorhodopsin with all-trans, 15-anti retinal chromophore; bR13-c, bacteriorhodopsin with 13-cis, 15-syn retinalchromophore; CD, circular dichroism; EFG, electric field gradient; GPCR, G protein-coupled receptor; M, bacteriorhodopsin M intermediate with 13-cis, 15-antichromophore; MAS, magic-angle spinning; MD, molecular dynamics; meta I, metarhodopsin I; meta II, metarhodopsin II; NMR, nuclear magnetic resonance; PAS,principal axes system; PDB, Protein Data Bank; POPC, 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine; PSB, protonated Schiff base; PYP, photoactive yellowprotein; RMSD, root mean square deviation; RQC, residual quadrupolar coupling⁎ Corresponding author. Department of Chemistry, University of Arizona, Tucson, Arizona 85721, USA. Fax: +1 520 621 8407.E-mail address: [email protected] (M.F. Brown).
1 Present address: Faculté de Pharmacie, Université René Descartes, Paris 75270, France.
0005-2736/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.bbamem.2007.10.014

2980 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
Contents
1. Introduction and motivation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29802. Deuterium NMR of retinal proteins in oriented membrane bilayers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2981
2.1. Angular and positional restraints are related to biomolecular structure and dynamics . . . . . . . . . . . . . . . . . . . . 29812.2. Non-aligned samples establish principal values for nuclear spin interactions . . . . . . . . . . . . . . . . . . . . . . . . 29812.3. Oriented samples yield principal axes of spin interaction tensors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2982
3. Theoretical NMR lineshape analysis for static uniaxial distribution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29833.1. Angular restraints are obtained from 2H NMR spectroscopy of aligned membrane samples. . . . . . . . . . . . . . . . . 29833.2. Example of oriented purple membranes containing bacteriorhodopsin . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29843.3. Global fitting of experimental tilt series of 2H NMR spectra yields bond orientations and alignment disorder of membranes . 2985
4. Application of solid-state 2H NMR to light-driven proton pump bacteriorhodopsin . . . . . . . . . . . . . . . . . . . . . . . . 29864.1. Solid-state 2H NMR shows changes in retinylidene chromophore in photocycle intermediates of bacteriorhodopsin . . . . 2986
4.1.1. Light-adapted bacteriorhodopsin (13-trans, 15-anti) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29874.1.2. Dark-adapted bacteriorhodopsin (13-cis, 15-syn and all-trans, 15-anti) . . . . . . . . . . . . . . . . . . . . . . . 29874.1.3. The M intermediate (13-cis, 15-anti) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2988
4.2. Solid-state NMR results are used for structural refinement of photocycle intermediates of bacteriorhodopsin. . . . . . . . 29904.3. Conformational changes are localized to the Schiff base end of retinal chromophore in bR photocycle. . . . . . . . . . . 2991
5. Application of solid-state 2H NMR to G protein-coupled receptor rhodopsin. . . . . . . . . . . . . . . . . . . . . . . . . . . . 29925.1. Solid-state 2H NMR of rhodopsin in oriented membranes gives angular restraints for structural analysis . . . . . . . . . . 29925.2. Analysis of 2H NMR data reveals structural changes upon bleaching of rhodopsin . . . . . . . . . . . . . . . . . . . . . 2993
5.2.1. Torsional twisting of retinal chromophore in dark-adapted state (11-cis) . . . . . . . . . . . . . . . . . . . . . . 29935.2.2. Conformational relaxation in metarhodopsin I state (all-trans) . . . . . . . . . . . . . . . . . . . . . . . . . . . 2994
5.3. Combining 2H NMR data with molecular dynamics simulations further illuminates rhodopsin function . . . . . . . . . . 29956. Comparison of retinal proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29957. Experimental procedures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2996Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2996References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2997
1. Introduction and motivation
Deuterium NMR spectroscopy yields a powerful avenue forinvestigating the structure and dynamics of integral membraneproteins. Membrane receptors, transporters, and ion channelsconstitute important therapeutic targets and are often refractoryto crystallographic analysis [1]. Although crystallography isoften viewed as the biophysical method of choice for structuralstudies, there are great difficulties in using this approach toaccurately describe the functional changes of membrane pro-teins [2]. For the retinal proteins bacteriorhodopsin (bR) andrhodopsin, the atomic displacements upon light absorption areeither small [2,3] or lead to disruption of the crystal [4]. Thecrystalline lattice resists large-scale conformational changes thatcan prevent the significant accumulation of the intermediates ofinterest [2,3,5]. A complementary approach is to use solid-stateNMR spectroscopy and to study the local conformation changesin a native-like membrane environment [6–11], where proteinfunction is preserved [12–15]. Here, we describe 2H NMRstudies of membranes containing the proton pump bR or rho-dopsin as a canonical G protein-coupled receptor (GPCR). Inboth cases the retinal chromophore was site-specifically labeledby deuteration of the methyl groups through total organicsynthesis. Solid-state NMR methods involved 2H NMR spec-troscopy of aligned membranes as a function of the orientationto the magnetic field, together with theoretical lineshape simu-lations. Analysis of the solid-state 2H NMR lineshapes entailedmapping the geometrical distribution of the molecules into thefrequency distribution of the 2H NMR spectra, using either a
closed form or Monte Carlo treatment [16]. From the methylbond angles the conformation and orientation of the retinalcofactor within the protein binding cavity could be established.
An important aspect is that solid-state NMR studies of staticaligned samples can reveal structural knowledge for non-crystalline biological systems, as in the case of biomembranes.Our work has investigated the 2H NMR spectra of planar-supported membranes versus the tilt angle to the magnetic fieldunder conditions where rotational and translational diffusion ofthe protein is effectively quenched. The problem of simulatingthe spectral lineshapes of uniaxially oriented immobile samplesarises in solid-state NMR of various biological systems,including integral membrane proteins [7–9,17,18] and macro-scopically aligned biopolymer fibers [19,20]. In each case, astatic distribution of the coupling tensors for NMR spininteractions (quadrupolar or dipolar couplings, chemical shifts)about a preferred axis is applicable. The oriented membranesinvolve substantial alignment disorder, and thus a propertreatment of the mosaic spread is vital to determining accuratebond angle restraints. For the retinal proteins bR and rhodopsin,global fitting of the experimental tilt series of 2H NMR spectrato a theoretical lineshape analysis for a static uniaxial dis-tribution gave the retinylidene bond orientations to themembrane normal. In this way, 2H NMR spectroscopy of thealigned membranes has allowed us to investigate light-inducedchanges in various photointermediate states that are coupled totheir photochemistry and biological activity.
As a first application, bR provides a representative examplefor membrane transporters, due to its function as a light-driven

Fig. 1. Powder-type 2H NMR spectra for rhodopsin with 2H-labeled retinal in thedark state indicate rotating methyl groups with large order parameters. (a)–(c)Experimental 2H NMR spectra for 11-Z-[9-C2H3]-retinylidene rhodopsin, i.e.with 11-cis retinal deuterated at the C9-methyl group, in gel-phase POPCmembranes (1:50 molar ratio). (d) Theoretical 2H NMR spectrum for randomlyoriented C–C2H3 groups undergoing rapid 3-fold rotation on the 2H NMR timescale (b (3χQ/8)
−1≈10 μs). (e),(f) Representative 2H NMR spectra for dark-state11-Z-[5-C2H3]-retinylidene rhodopsin and 11-Z-[13-C
2H3]-retinylidene rhodop-sin, i.e. with 11-cis retinal deuterated at the C5-methyl (yellow) or C13-methylgroup (red), respectively, in POPC membranes (1:50). Theoretical 2H NMRspectra for C–C2H3 groups undergoing axial rotation (continuous color lines) aresuperimposed on the experimental spectra, with residuals below. Adapted withpermission from Ref. [77].
2981M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
proton pump [2,13,21]. A nearly planar structure was found inthe dark- and light-adapted states, with an additional light-induced change upon 13-cis isomerization in the M photo-intermediate. According to 2H NMR a light-induced switchoccurs in the N–H bond orientation of the retinal chromophorefrom the extracellular side to the cytoplasmic side of the mem-brane in the proton pump mechanism [8]. Moreover rhodopsinwas studied as a prototype for GPCRs in cellular membranes[22]. Most striking, the 2H NMR structure of 11-cis retinal inthe dark state revealed torsional twisting of the polyene chainand the β-ionone ring, in direct contrast to bR. For rhodopsinthe retinal cofactor is locked within the binding pocket with anegative pre-twist about the C11_C12 double bond, whichexplains its dark-state stability as well as its rapid photo-chemistry, and also indicates the trajectory of the light-induced11-cis to trans isomerization. According to 2H NMR, the retinalstrain is progressively relaxed in forming the pre-activated metaI state. Conformational differences of the same retinal moleculein the two retinal proteins are instructive with regard to theirdistinctive biological mechanisms of action.
2. Deuterium NMR of retinal proteins in orientedmembrane bilayers
In solid-state NMR spectroscopy one observes couplingtensors which manifest the nuclear or electronic interactions ofinterest [23,24]. The nuclear spin interactions encompass thechemical shift, the magnetic dipolar coupling (direct orindirect), and the electric quadrupolar coupling. Physicalinteractions are represented by second-rank coupling tensorsusing either a Cartesian or spherical basis. The principal valuesare related to the strength of the interactions, and the principalaxes are connected with the geometry. Apart from differentprincipal values and principal axes, the various tensorial inter-actions all transform in exactly the same way [25]. Spininteractions are most generally formulated in terms ofirreducible tensors and tensor operators; the same models areapplicable and only the coupling mechanisms differ [23–25].Together, the NMR lineshapes and relaxation times provide aunified view of both the structure and dynamics [25]. Theequilibrium structure is related to the static or residual tensorialinteractions as obtained from the NMR lineshape. Fluctuationsof the tensor with respect to the laboratory frame give anadditional time-dependent perturbation that is manifested bythe nuclear spin relaxation. In this article, we focus mainly onequilibrium structures as investigated by solid-state 2H NMRlineshapes.
2.1. Angular and positional restraints are related tobiomolecular structure and dynamics
The various solid-state NMR methods generally fall into twocategories. First, dipolar recoupling experiments with unor-iented samples under magic-angle spinning (MAS) allowdistances to be obtained [26–36], and moreover relative tensororientations can yield molecular torsion angles [37–39].Alternatively, solid-state NMR of aligned membranes [40,41]
can be used to directly study the orientations of membraneconstituents [6,7,9,42,43]. The strategy is well illustrated byinvestigations of small peptides in membrane bilayers [42–52]as well as membrane proteins [6–9,17,53] and lipid bilayers[54,55]. Here, one determines the angles of the nuclear spininteraction tensors with respect to an axis of macroscopic align-ment, e.g. the normal to the membrane surface [7,9,16,17,43,49]. Analogous information is in principle obtained for orientedsamples rotating under magic angle conditions [18,56]. In factcombination of angular [7,9,17,43,49] and positional [29,57]restraints in solid-state NMR is the counterpart of measuringresidual dipolar couplings (RDCs) and nuclear Overhauserenhancements (NOEs) in solution NMR [58–61]. The angularand distance restraints are less in solid-state NMR; yet they areknown to higher precision. In either case, the nuclear positionsand bond orientations can be used for structure elucidation.
2.2. Non-aligned samples establish principal values fornuclear spin interactions
In what follows, we consider the specific case of 2H NMR ofintegral membrane proteins, e.g. transporters or receptors. For 2HNMR experiments with retinal proteins such as rhodopsin or bR,

Fig. 2. Solid-state NMR yields structural information for ligands bound tointegralmembrane proteins. (a) Illustration of nuclear spin energy levels obtainedby solving the Schrödinger equation for an 2H system [25]. Degeneracy of thesingle-quantum transitions due to the Zeeman interaction (HZ) is removed by theorientation-dependent quadrupolar coupling (HQ). The frequency separationbetween the single-quantum quadrupolar frequencies is the (residual) quad-rupolar couplingΔνQ. (b) As an illustration we show the retinal chromophore ofbacteriorhodopsin in membranes as investigated by solid-state 2H NMRspectroscopy. Geometry of the tilt experiments is presented for a static uniaxialdistribution of aligned bilayers. For a given methyl group, θB is the angle of theC–C2H3 bond axis to the local membrane normal n, with static rotationalsymmetry given by the azimuthal angle ϕ. Alignment disorder is described bythe angle θ′ of n relative to the average membrane normal n0, and is likewiseuniaxially distributed as characterized by ϕ′. Next, θ is the tilt angle from n0 tothe main magnetic field B0, about which there is cylindrical symmetry. Lastly, θ″and ϕ″ are the angles for overall transformation from n to B0 (not shown) [20].
2982 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
the different methyl groups of the cofactor are specificallydeuterated (Fig. 1). This isotopic labeling is an isomorphicreplacement that does not change the chemistry of thephotoisomerization. The 2H NMR method allows one to studythe orientation of a single labeled group in an oriented sample. Itcan be used to study local structural changes and is complemen-tary to X-ray diffraction [2,62–64]. In the latter method, everyatom contributes to the intensity of every reflection and the Rfactor is minimized globally; by contrast in 2H NMR the mini-mization is conducted only for the functional group of interest.
For rhodopsin [9,18,65] or bR [6,66,67] organic synthesis[68–71] has been applied extensively to 13C and 2H labeling ofretinal for solid-state NMR applications. Fig. 1(a)–(c) showsillustrative 2H NMR spectra of 11-Z-[9-C2H3]-retinal bound torhodopsin in random (non-aligned) POPC membrane disper-sions (so-called powder-type samples) over a temperature rangefrom −30 °C to −150 °C, below the lipid order–disordertransition temperature (Tm=−5 °C for POPC). The lowesttemperature studied, T=−150 °C, is close to the cryogenictemperatures employed in X-ray crystallography of rhodopsin[63,64,72,73]. Below Tm the 2H NMR spectra correspond to thelipid gel state, where rotational and translational diffusion ofboth the protein [74,75] and lipids are diminished on the 2HNMR time scale. The 2H NMR spectra were simulated using thetheoretical expression for a Pake doublet due to a sphericaldistribution of electric field gradient (EFG) principal axes [25].Below the powder pattern, the residuals between fit and data areplotted. In Fig. 1(d) the spectral probability density p(ξ±) for thetwo spin I=1 spectral branches is graphed against the reducedfrequencies ξ±. Additional powder-type
2H NMR spectra of 11-Z-[5-C2H3]-retinylidene rhodopsin and 11-Z-[13-C2H3]-retiny-lidene rhodopsin in POPC membranes (1:50) are shown in Fig.1(e) and (f), respectively.
An immediate conclusion is that the quadrupolar coupling(EFG tensor) is motionally averaged versus the static EFGcoupling tensor for an aliphatic C–2H bond. The averaging isdue to rapid 3-fold rotation of the methyl groups on the 2HNMR time scale (b10 μs) at all temperatures studied, down to−150 °C. For if there was a large off-axial motion of the methylgroups, then the residual quadrupolar coupling (RQC) would besignificantly less, as seen for liquid-crystalline lipid bilayers[25,76]. As the observed coupling is near the theoretical valuefor methyl 3-fold rotation, we conclude that there is noadditional rotation of the protein on the 2H NMR time scale inthe gel-state lipid bilayer. This conclusion is in agreement withprevious spin-label EPR studies of rhodopsin rotationaldiffusion [74,75]. Comparison of the RQC to the theoreticalvalue for a rotating C–C2H3 group indicates that the orderparameter for the methyl 3-fold axis is SC3
=38/41.75≈0.9 foreach for the three methyl groups [77], with off-axial fluctuationsof ≈15°. (A value of χQ=167 kHz is assumed for the staticquadrupolar coupling constant.) Despite the phenomenal dark-state stability of retinal it has appreciable mobility within therhodopsin binding pocket, with rapidly rotating methyls andrestricted off-axial fluctuations. Yet knowledge of the methylorientation within the membrane frame is unavailable fromstudies of non-aligned membrane dispersions. The same is true
for dipolar recoupling experiments under MAS, which providedistances but not angles with respect to the membrane.
2.3. Oriented samples yield principal axes of spin interactiontensors
The preparation of well-aligned membranes containingproteins is the counterpart of obtaining 3D crystals in X-raycrystallography [3,5,64,72,78,79]. Orientation of purple mem-branes containing bR was achieved by slowly drying themembrane suspension on glass slides [7,8,17]. Analogously,rhodopsin was aligned on planar substrates by isopotential spin-dry centrifugation of the membranes onto glass slides [9,41].Now for 2H NMR we shall confine our discussion to an axiallysymmetric EFG tensor. The quadrupolar frequencies of the twoI=1 spectral branches depend on the orientation of the C–2Hbond (the principal EFG tensor axis) to the main magnetic fieldB0. The coupling tensor is molecule fixed, while the nuclearspin angular momentum (magnetic moment) is quantized in thelaboratory system. The Schrödinger equation for the quad-rupolar interactions can be formulated in terms of the anglesbetween the EFG principal axes system (PAS) and the B0 frame.

2983M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
Solving the Schrödinger equation for the first-order quadrupolarenergies [25] then yields two single-quantum transitions, cf.Fig. 2(a). However, the orientational distribution of the couplingtensor with respect to the laboratory system is unknown and isneeded to describe the molecular geometry of anisotropicbiomembrane systems. Generally, the transformation from onecoordinate system to another is accomplished by a standardsimilarity transformation in terms of a Cartesian representation,as in elementary physics texts. But for multiple transformationsof coordinates, such an approach rapidly gets out of hand.Moreover, the physical significance of the rotations and thecorrespondence to spectroscopic observables (like order para-meters) tend to become lost. A better way is to formulate thenuclear spin interactions with an irreducible representation, i.e.using spherical tensor operators. This allows one to transformstep-wise from the molecule to the membrane to the laboratoryframe in a straightforward manner [25]. Essentially one uses themachinery developed for angular momentum theory in nuclearphysics as a conceptual tool [80,81].
As an illustration, we consider in Fig. 2(b) an alignedmembrane sample where the aim is to investigate the structureof retinal in the binding pocket of the protein. Retinal is labeledwith 2H at the methyl groups in the case of bR [7]. Our strategyis to decompose the overall angle of the C–C2H3 bond to themagnetic field in terms of various angles that characterize thebiomembrane geometry. The step-wise transformation ofcoordinates is accomplished using closure as illustrated byFig. 2(b). First we transform from the PAS of the residual EFGtensor of the rotating C–C2H3 groups to the frame of the localmembrane normal, about which there is a static uniaxial dis-tribution below the lipid Tm; next we transform from the localframe that is likewise axially distributed about the averagenormal to the aligned membrane stack; and last we transformfrom the average frame to the laboratory axis system defined bythe external magnetic field. Hence the lineshape analysisinvolves two static uniaxial distributions. Only static distribu-tions are considered, and motional rates appear only through theintrinsic linebroadening in the simulations.
3. Theoretical NMR lineshape analysis for static uniaxialdistribution
The probability distribution for the bond orientations inclosed form is of potential interest for solid-state NMRspectroscopy of integral membrane proteins [7,18] and peptides[47] involving dipolar, quadrupolar, and chemical shift interac-tions. For the case of aligned biomolecules our general aim is todetermine the coupling tensor orientation within the alignmentframe, e.g. the quadrupolar or dipolar coupling. One can thenuse the angular restraints for structure determination. But hy-drated membranes containing proteins or peptides can involvesubstantial alignment disorder, i.e., the individual membranenormals are not all completely aligned. They are distributedaround the glass plate normal, and the lineshape consists ofinhomogeneously broadened lines; this distribution is known asthe mosaic spread of the sample. An assumption must then bemade about the shape of the distribution in order to determine the
bond orientation with respect to the membrane normal from the2H NMR lineshape. An important aspect is the correct treatmentof the distribution of local membrane normals with respect to theaverage alignment axis. Previous treatments have neglected the3D nature of the mosaic spread of the aligned membranes, whichcan lead to different calculated results. In general, we canconsider two limiting regimes. First, in solution NMR theweakly oriented limit is applicable for molecules due to presenceof bicelles, stretched gels, and so forth. In this case the weakalignment order is introduced as a perturbation of the isotopicNMR spectrum [59,60]. Second, in solid-state NMR the stronglyoriented limit is appropriate, as in the case of macroscopicallyaligned membranes. Here the disorder (mosaic spread) is nowintroduced as a perturbation [16]. The derived lineshape functionyields the correct powder-type limit, and has been tested bysimulating a tilt series of 2H NMR spectra of purple membranescontaining bR with a specifically deuterated C1R-methyl groupof the retinal β-ionone ring [7,16]. For bR or rhodopsin, therelative orientations of the quadrupolar coupling tensors due topairs of the methyl groups then allow us to calculate effectivetorsional angles between the different planes of unsaturation ofthe retinal chromophore.
3.1. Angular restraints are obtained from 2H NMRspectroscopy of aligned membrane samples
At this point we recall that the quadrupolar interaction de-pends on the angle θ between the EFG and the magnetic field[25]. For a random angular distribution of the coupling tensorover the surface of a unit sphere, the NMR lineshape cor-responds to a Pake doublet [25]. Fig. 3(a) shows that the weaksingularities correspond to the largest principal value of thecoupling tensor aligned perpendicular to the external magneticfield (θ=90°); and the shoulders represent the parallel orienta-tion (θ=0°). The reason is that only a single orientationcorresponds to θ=0° (pole) whereas for θ=90° the values areazimuthally distributed (equator). The angular dependence ofthe quadrupolar frequencies for the two spectral branches isshown at the bottom of Fig. 3(a). (For the more general situationof a non-axially symmetric tensor the spectral discontinuitiescorrespond to each of the three principal axes pointing along themagnetic field direction.)
Now for the more interesting case of a semi-random distri-bution, the orientational probability is nonuniform and therebymanifests the geometry of the sample. (By a semi-randomdistribution we mean that the molecules have a preferentialalignment relative to some direction in the sample.) One suchexample is a static uniaxial distribution [16,25,82], whichoccurs in aligned samples of membrane proteins [16], or drawnor aligned biopolymers such as DNA fibers [20]. Effectively,one has a two-dimensional powder-type distribution in the planeperpendicular to the axis of symmetry; whereas the probabilitydensity is nonuniform elsewhere. This is illustrated at the left ofFig. 3(b), where the largest tensor principal axis is distributed onthe rim of a cone and the orientation to B0 is designated as θ.For a perfectly aligned membrane sample, all bonds lie on therim of a cone around the orientation axis, whose semi-angle is

Fig. 3. Sample geometry is related to nuclear spin interaction tensors in solid-state NMR spectroscopy. (a) Case of random distribution of coupling tensor principalsymmetry axes. The 2H NMR spectrum for the two spectral branches of an I=1 spin system is graphed as a function of the reduced frequency ξ± for the EFG tensorprincipal axes distributed over the surface of a sphere. The probability distribution p(ξ±) scales as 1 / |cosθ|, where θ is the angle between the EFG tensor principalz-axis (assumed axially symmetric) and the static external magnetic field B0. Weak singularities are evident with intensity maxima at θ=90° (equator) and intensityextending to a minimum at θ=0° (poles). (b) Case of semi-random distribution of coupling tensor principal symmetry axes, where the positions of the spectral intensitymaxima for an individual spectral branch of the I=1 nucleus are depicted. A static uniaxial distribution is considered in which the coupling tensor z-axes are confinedto the rim of a cone. Due to inversion and reflection symmetry only the distribution for a quartersphere is needed. The probability distribution p(ξ±) scales as 1 / |cosθ|[cos(θ+θB)−cosθ]1/2[cosθ−cos(θ−θB)]1/2, where θ is the tilt angle of the alignment axis to the main magnetic field B0 and θB is the bond orientation to the alignmentaxis. At the right an axially symmetric spectrum is obtained for zero tilt; with increasing tilt angle the spectral features correspond to the sum and difference of θ andθB. At a sufficiently large tilt angle a further weak singularity becomes evident at θ=90°, as in the case of a random distribution. (c) Example of simulated lineshape fora uniaxial static distribution of methyl groups undergoing 3-fold axial rotation, e.g. in the case of an aligned membrane protein. The two spectral branches of the I=12H nucleus are depicted at the bottom with the resultant spectrum at top.
2984 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
between the bond and the membrane normal. If the alignmentaxis is parallel to the main external magnetic field B0, then onlya single angle to the magnetic field occurs; and for eachindividual spectral branch there is but one transition.
Unfortunately, there is a potential complication in using onlythe zero-tilt 2H NMR spectrum to calculate the angle of themethyl rotor axis to the alignment axis (membrane normal in ourcase). The ambiguity lies in the fact that only the absolute valueof the quadrupolar splitting ΔνQ is measured, which isproportional to |P2(cosθ)|, where P2 is the second Legendrepolynomial [25,83,84]. Two solutions are then obtained whenthe P2 curve falls between −1/2 and +1/2, i.e. when 35.3≤θ≤90°. But if the alignment axis is tilted away fromB0, then themaxima in the probability distribution shift to the sum anddifference of the alignment (tilt) angle θ with the cone semi-angle θB. The distribution of bonds with respect to the magneticfield is then characteristic of the cone angle, and allows one todistinguish the positive and negative solutions [7,8,13,16].Moreover, at sufficiently large tilt angle or cone semi-angle, theangular distribution will cross the xy-plane and there is anadditional weak singularity at θ=90°, as for a random dis-tribution, cf. Fig. 3(b). Thus, at large tilt angles there will alwaysbe a component at 90° that is independent of the alignment axis[16,25,82]. Assuming an axially symmetric EFG tensor, we thenobserve a maximum of three peaks for each of the two spectralbranches, corresponding to six lines for the NMR spectrum of anI=1 nucleus such as 2H (or 14N). There are three discontinuitiesfor each spectral branch, i.e. at θ=θ±θB, 90°. A further aspect isthat due to the even parity of the rank-2 coupling tensor, i.e. theinversion symmetry [80,81], it suffices to only consider theorientations for a quartersphere. Lastly, Fig. 3(c) shows a
representative simulated 2H NMR lineshape; where as a generalrule the experimental quadrupolar lineshape reflects theprobability density of all bond orientations in the sample.
We have previously worked out the theory of the 2H NMRlineshapes for such a uniaxial, immobile distribution [16].Either an analytical solution in mathematical closed form or anumerical Monte Carlo procedure can be used to simulate thelineshape. The latter in principle allows for any arbitrarydistribution to be included [7,16]. The frequency separation ofthe single-quantum transitions (quadrupolar splitting) is givenby ΔνQ=ΔνQ
powder (3cos2θ−1), where θ is the angle betweenthe C–C2H3 bond axis and the magnetic field B0, which mayvary due to rotation about the membrane normal for non-zerotilt. Here ΔνQ
powder =3/4⟨χQ⟩ is the splitting of the two linesfor θ=90° in the case of a non-oriented powder-type sample.The residual quadrupolar coupling constant ⟨χQ⟩ is averaged bythe methyl group rotation, and depends on the static couplingconstant χQ (=167 kHz), the tetrahedral geometry of the methylgroup, and the order parameter SC3
for the off-axial motion [77].Its value is ≈40 kHz for a rotating methyl group in the fastmotional limit that applies to our samples [7,67,77]. To obtainthe correct C–C2H3 bond orientation, it is necessary to simulatea tilt series of angular-dependent 2H NMR spectra for |ΔνQ|≤|ΔνQ
powder| as noted above [7–9,16,77]. Further description ofthe mathematical treatment can be found elsewhere [7,8,16].
3.2. Example of oriented purple membranes containingbacteriorhodopsin
As a specific example, a tilt series of 2H NMR spectra isshown in Fig. 4 for the case of bR in oriented purple
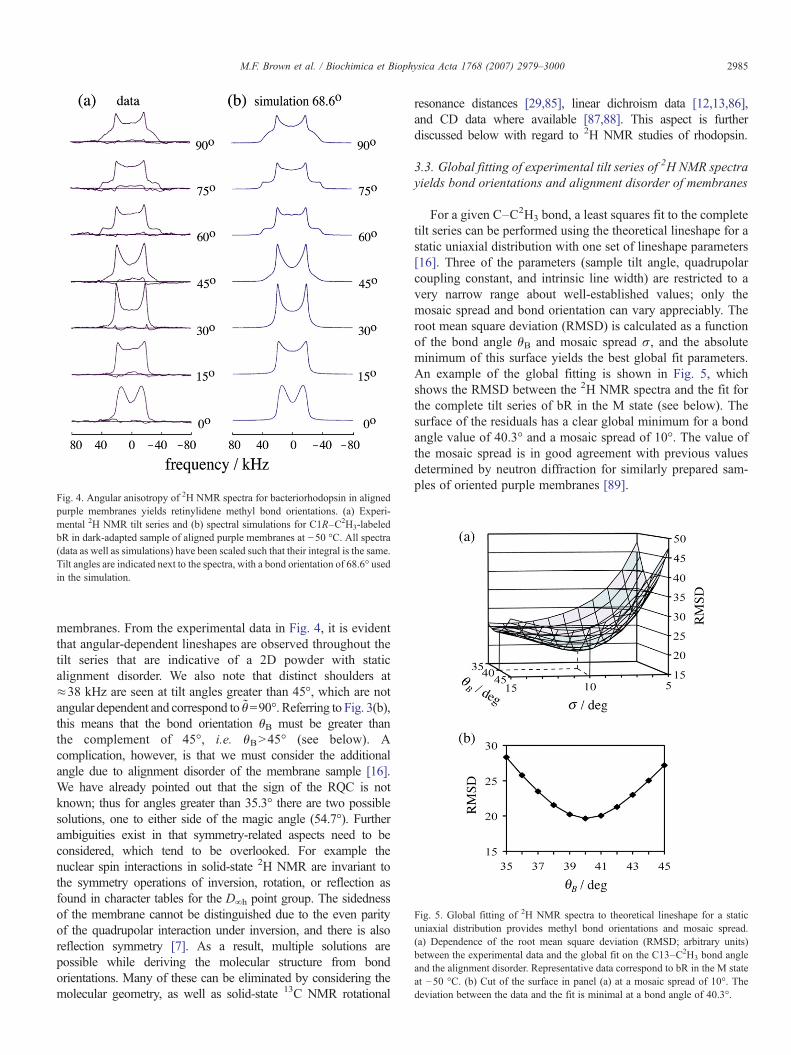
Fig. 4. Angular anisotropy of 2H NMR spectra for bacteriorhodopsin in alignedpurple membranes yields retinylidene methyl bond orientations. (a) Experi-mental 2H NMR tilt series and (b) spectral simulations for C1R–C2H3-labeledbR in dark-adapted sample of aligned purple membranes at −50 °C. All spectra(data as well as simulations) have been scaled such that their integral is the same.Tilt angles are indicated next to the spectra, with a bond orientation of 68.6° usedin the simulation.
Fig. 5. Global fitting of 2H NMR spectra to theoretical lineshape for a staticuniaxial distribution provides methyl bond orientations and mosaic spread.(a) Dependence of the root mean square deviation (RMSD; arbitrary units)between the experimental data and the global fit on the C13–C2H3 bond angleand the alignment disorder. Representative data correspond to bR in the M stateat −50 °C. (b) Cut of the surface in panel (a) at a mosaic spread of 10°. Thedeviation between the data and the fit is minimal at a bond angle of 40.3°.
2985M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
membranes. From the experimental data in Fig. 4, it is evidentthat angular-dependent lineshapes are observed throughout thetilt series that are indicative of a 2D powder with staticalignment disorder. We also note that distinct shoulders at≈38 kHz are seen at tilt angles greater than 45°, which are notangular dependent and correspond to θ=90°. Referring to Fig. 3(b),this means that the bond orientation θB must be greater thanthe complement of 45°, i.e. θBN45° (see below). Acomplication, however, is that we must consider the additionalangle due to alignment disorder of the membrane sample [16].We have already pointed out that the sign of the RQC is notknown; thus for angles greater than 35.3° there are two possiblesolutions, one to either side of the magic angle (54.7°). Furtherambiguities exist in that symmetry-related aspects need to beconsidered, which tend to be overlooked. For example thenuclear spin interactions in solid-state 2H NMR are invariant tothe symmetry operations of inversion, rotation, or reflection asfound in character tables for the D∞h point group. The sidednessof the membrane cannot be distinguished due to the even parityof the quadrupolar interaction under inversion, and there is alsoreflection symmetry [7]. As a result, multiple solutions arepossible while deriving the molecular structure from bondorientations. Many of these can be eliminated by considering themolecular geometry, as well as solid-state 13C NMR rotational
resonance distances [29,85], linear dichroism data [12,13,86],and CD data where available [87,88]. This aspect is furtherdiscussed below with regard to 2H NMR studies of rhodopsin.
3.3. Global fitting of experimental tilt series of 2H NMR spectrayields bond orientations and alignment disorder of membranes
For a given C–C2H3 bond, a least squares fit to the completetilt series can be performed using the theoretical lineshape for astatic uniaxial distribution with one set of lineshape parameters[16]. Three of the parameters (sample tilt angle, quadrupolarcoupling constant, and intrinsic line width) are restricted to avery narrow range about well-established values; only themosaic spread and bond orientation can vary appreciably. Theroot mean square deviation (RMSD) is calculated as a functionof the bond angle θB and mosaic spread σ, and the absoluteminimum of this surface yields the best global fit parameters.An example of the global fitting is shown in Fig. 5, whichshows the RMSD between the 2H NMR spectra and the fit forthe complete tilt series of bR in the M state (see below). Thesurface of the residuals has a clear global minimum for a bondangle value of 40.3° and a mosaic spread of 10°. The value ofthe mosaic spread is in good agreement with previous valuesdetermined by neutron diffraction for similarly prepared sam-ples of oriented purple membranes [89].

Fig. 6. Carbon numbering for the retinylidene chromophore of bacteriorho-dopsin. The chromophore is illustrated with a specifically deuterated C13-methyl group. The three isomers are the three structures investigated by solid-state deuterium NMR. (a) The all-trans conformer bRall-t occurs in the activeform of bacteriorhodopsin (light-adapted state). (b) The 13-cis, 15-synconformer occurs in the dark-adapted state in a 2:1 mixture of bR13-c andbRall-t. (c) In the M state the configuration is 13-cis, 15-anti and the Schiff baseis deprotonated.
2986 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
In addition, Fig. 5(b) shows a cut of the surface of Fig. 5(a) ata constant mosaic spread of 10°, and indicates that the minimumis steep and very well defined. The simulation is very sensitiveto the bond orientation, which is highly correlated with thequadrupolar coupling constant. The error of the determinedangle can therefore only be estimated to be about ±2°. Values ofthe mosaic spread σ are 7.2° at 20 °C and 8.7° at −50 °C.Referring back to Fig. 2, we see that mosaic spread is definedfor the angle θ′ and is therefore a property of the sample.However, the distribution of the local membrane normal in thelaboratory system also depends on the tilt angle θ. While themembrane normals are distributed in a Gaussian manner in theframe of the average normal, it is not so in the laboratory frame[16]. As a result, in the case of a uniaxial distribution it isincorrect to simply add a Gaussian-weighted series ofsubspectra at varying tilt angles.
4. Application of solid-state 2H NMR to light-drivenproton pump bacteriorhodopsin
We then applied these methodologies to bR as a paradigm forlight-driven ion (proton) pumps in archea. The bR moleculecomprises seven transmembrane helices, where the retinalcofactor is all-trans retinal in the light-adapted state, andundergoes isomerization to 13-cis in the proton pump mechan-ism. Solid-state 2H NMR of aligned purple membranes hastested the hypothesis that specific methyl sites within the retinalbinding pocket are implicated in the proton pump mechanism.This work has established the retinal structure in both the light-adapted and the dark-adapted states, as well as the activated Mstate in the photocycle of bR. In these states retinal adopts threedifferent conformations: a 2:1 mixture of 13-cis, 15-syn and all-trans, 15-anti (dark-adapted ground state); all-trans, 15-anti(cryotrapped light-adapted state); and 13-cis, 15-anti(M intermediate) at −50 °C. Fig. 6 shows the isomericconfiguration, carbon numbering, and site of 2H labeling ofall three chromophore states investigated. (Bacteriorhodopsinwith an all-trans, 15-anti retinal chromophore is denoted asbRall-t, with a 13-cis, 15-syn retinal chromophore as bR13-c, andthe M intermediate with a 13-cis, 15-anti chromophore as M.) Inthe wild-type samples, the lifetime of the M state was increasedby applying a guanidine hydrochloride solution at pH 9.5, andin the D96A mutant by raising the pH. The orientations of fourof the five methyl bonds of the retinylidene chromophore wereexamined in the dark-adapted state [7,17]. Light-inducedorientational changes of the C1R-, C5-, and C9-methyl bondshave also been investigated in the late M state [7,8]. However,of the methyl bonds of the chromophore of bR, the C13–C2H3
bond is expected to be most sensitive to isomerization about theadjacent C13_C14 double bond during light–dark adaptation,and during the photocycle. Therefore, we have focused on thisparticular site. We detected substantial differences in the orien-tation of the C13–C2H3 bond in all three states bRall-t, bR13-c,and M by 2H solid-state NMR with oriented purple membranes.Moreover, we detected for the first time structural changes ofthis bond orientation between the bRall-t and bR13-c isomers thatoccur during light–dark adaptation.
4.1. Solid-state 2H NMR shows changes in retinylidenechromophore in photocycle intermediates of bacteriorhodopsin
The molecular mechanism of the light-driven proton pumpwas investigated through studies of the photocycle intermedi-ates of bR. What are the changes in the retinal chromophore ofbR that underlie its activity as a light-driven (photosynthetic)H3O
+ pump? In this regard, application of FTIR [90–93] andsolid-state NMR [7,28,39,56] spectroscopy together with X-raycrystallography [2,21] gives results that can be combined toprovide mechanistic insight. Studies of bR single crystals haveyielded structures for several of the photointermediates of bR(dark-adapted, K, L, M1, M2, and M2′ states) [2]. Yet despite thisimpressive progress the molecular mechanism by which H3O
+
ion transport occurs remains obscure [2,92–94]. The retinalchromophore and its binding pocket clearly define a domain ofgreat functional significance, and solid-state NMR can gobeyond X-ray diffraction to locally refine the structure of retinalbound to bR. Accurate distance (13C-to-13C, 13C-to-15N) andangular (C–C2H3) constraints provide high-resolution localstructural information about the chromophore and amino acidsin its immediate environment [7,8,95,96]. Thus our objectivewas to contribute to understanding the mechanism of the protonpump in atomic detail.

2987M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
Purple membranes containing wild-type or mutant D96Abacteriorhodopsin were regenerated with all-trans retinaldeuterated at the C1R-, C5-, C9-, or C13-methyl groups [7,8],i.e. all-E-[1R-C2H3]-, all-E-[5-C
2H3]-, all-E-[9-C2H3]-, or all-
E-[13-C2H3]-retinals. The experiments were performed at−50 °C with planar-supported purple membranes in the absenceor presence of guanidine hydrochloride (GuHCl), pH 9.5 and85% relative humidity. The NMR structure of retinal within thebR binding pocket is consistent with a ≈ planar retinal con-formation over most of the polyene that is in the light-adaptedand dark-adapted states, as well as the M intermediate of theproton pump. Here we mainly focused on the “business end” ofthe chromophore, particularly the C13-methyl group toward theSchiff base end. While only minor differences were detectedbetween dark- and light-adapted states, all four methyl anglesincreased in the M state. An upwards movement of the C13-methyl towards W182 (cytoplasmic direction) was inducedupon formation of the 13-cis isomer in the activated M state.This is the first 2H NMR report of such a structural differencebetween the dark- and light-adapted isomers of the chromo-phore in the ground state of bacteriorhodopsin.
4.1.1. Light-adapted bacteriorhodopsin (13-trans, 15-anti)For the light-adapted state with an all-trans, 15-anti
chromophore, there is good agreement between the X-raydiffraction and 2H NMR results for all the methyl bonds exceptthe C13–C2H3 bond nearest the Schiff base (for a review seeRef. [13]). In the latter case the average value of the C13-methylorientation over four different X-ray structures deposited in thePDB is 21.6° whereas the previous 2H NMR result is θB=32°[17]. Since the error in these measurements is approximately ±3°[13], a discrepancy seems to exist. We therefore conductedadditional 2H NMR measurements in the light-adapted state to
Fig. 7. Angular-dependent 2H NMR spectra for bacteriorhodopsin in light-adapted sNMR tilt series spectra (left) and their best least squares fit (right) for light-adapted baat −50 °C. The best global fit of the tilt series was obtained with a bond orientation opanel. (b) Comparison of the 2H NMR tilt series spectra for the light-adapted state wX-ray diffraction and a mosaic spread of 10° (right panel). The deviations between dathis value for the bond orientation.
shed light on the possible source of this discrepancy. Here bRwas trapped in the light-adapted ground state with the results ofthe tilt series presented in Fig. 7. The data of Fig. 7 were analyzedas described above [7] and a single species (all-trans, 15-anti)was assumed. The residuals showed a global minimum with abond angle of θB=34.2° and a mosaic spread of 10°. This valuefor the bond angle is in excellent agreement with the previousvalue of 32° obtained by 2H NMR with a similarly preparedsample in the absence of GuHCl [17]. However, the valuesobtained in X-ray diffraction experiments with light-adaptedcrystals are consistently smaller. Fig. 7 shows that it is notpossible to simulate the NMR spectra with a bond orientation of21.6° corresponding to the results of X-ray crystallography. Inthe crystallographic work, the known high-resolution structuresof all-trans retinal model compounds were used as an inputmodel in the refinement. This may possibly lead to a biastowards this structure. Interestingly, our value for the bR13-c
isomer of 24.7° obtained from the dark-adapted spectra is inreasonable agreement with the X-ray value of 21.5° for the bRall-t
isomer in light-adapted crystals.
4.1.2. Dark-adapted bacteriorhodopsin (13-cis, 15-syn andall-trans, 15-anti)
The dark-adapted state of bacteriorhodopsin consists of anequilibrium between two isomeric states of the chromophore:the 13-cis, 15-syn isomer and the all-trans, 15-anti isomer(Fig. 6). The fractional populations of the two isomers are 2:1 atroom temperature [97]. The 13-cis, 15-syn form does not pumpprotons and has a very different photocycle which lacks an Mintermediate. Structural differences between these two forms ofbR are thus to be expected. Recently, a small difference of 0.3 Åwas detected in the distance between the C14 carbon of thechromophore and the indole nitrogen of W86 for the two
tate show differences versus X-ray crystal structure. (a) Comparison of the 2Hcteriorhodopsin deuterium labeled in the C13-methyl group of the chromophoref 34.2° for the C13–C2H3 bond. The least squares residuals are shown in the leftith a simulation using as parameters a bond orientation of 21.6° obtained fromta and simulation on the left clearly indicate that the results cannot be fitted with

2988 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
isomers of the dark-adapted state using solid-state NMRspectroscopy [95]. Concerning the dark state, representative2H NMR spectra are presented in Fig. 4 for the C1R-methylgroups. Similar tilt series have been acquired for the C5- andC9-methyl groups of bR in the dark state [8]. The 2H NMRspectra of samples with a deuterated C13–C2H3 group may bethe most sensitive to structural changes due to isomerizationaround the adjacent C13_C14 bond in light–dark adaptation.With deuterated methyl groups further away from theC13_C14 bond, only small orientational changes weredetected in light–dark adaptation by solid-state 2H NMR [8].
Here bR was trapped in the dark-adapted ground state at−50 °C with the results of the tilt series presented in Fig. 8(a).Since the bond orientation in the all-trans, 15-anti configura-tion is known from our analysis of the light-adapted state(above), the dark-adapted data were analyzed as follows. Theobserved NMR spectra are a superposition of the 13-cis, 15-synand all-trans, 15-anti basis spectra with 2:1 weights. With thespectrum of the all-trans, 15-anti component fixed, two-component fits were performed as before to find the best bondorientation for the 13-cis, 15-syn species. The results in Fig. 8(a)show that the best fit was obtained for a bond angle of 24.7°for the 13-cis, 15-syn configuration. Moreover, comparison ofFigs. 7 and 8(a) indicates that there are major differencesbetween the NMR spectra in the light- and dark-adapted statescorresponding to a change in the C13-methyl bond angle from34.2° to 24.7°. To establish the reproducibility of the dark-adapted spectra the data were collected in the sequence dark-adapted, light-adapted, and dark-adapted. Repeating the experi-
Fig. 8. Solid-state 2H NMR spectroscopy of aligned purple membranes yields stru(a) Comparison of the dark-adapted 2H NMR tilt series (data) and their best leaschromophore at −50 °C. The best global fit of the tilt series was obtained with a bonddetails of the data analysis see text. The least squares residuals are shown in the left pabR molecule as derived from 2H NMR orientational constraints. Structure correspondis consistent with linear dichroism results; whereas that in (b) differs significantly [7].with permission from Ref. [7].
ments in the dark-adapted state reproduced the results of theinitial experiments.
A further point of interest is that for bR the retinal chro-mophore is nearly planar; hence the conformation and orien-tation are described by a model with tilt and roll angles for asingle plane of unsaturation [7]. The plane is defined by twoangular restraints and has even parity, giving four possibleorientations. Introducing restraints from additional biophysicalmethods enables the roll and tilt angles to be determined [7].The results are summarized in Table 1. As a specific example,Fig. 8(b) shows two possible orientations of all-trans retinalwithin the binding pocket of the dark-adapted state of the bRmolecule that are consistent with the 2HNMR data [7]. Howeveronly the top structure agrees with linear dichroism results [13].The calculated roll angle is 24° which means that the averageretinal plane is almost perpendicular to the membrane plane.This value is in good agreement with the conclusions from lineardichroism of the hydrogen-out-of-plane (HOOP) modes in FTIRspectroscopy, and is generally consistent with the notion that theplane of the chromophore is nearly although not quiteperpendicular to the membrane plane [13].
4.1.3. The M intermediate (13-cis, 15-anti)The orientations of the C5-, C9-, and C13-methyl bonds of
the retinylidene chromophore of bacteriorhodopsin have alsobeen investigated in the M photointermediate using solid-state2H NMR. The M intermediate has a 13-cis, 15-anticonformation and a deprotonated Schiff base (Fig. 6). Directstructural evidence for a reorientation of the C9–C2H3 bond
cture for retinylidene chromophore of bacteriorhodopsin in dark-adapted state.t squares fit (right) for bR deuterium labeled in the C13-methyl group of theangle of 24.7° for the C13–C2H3 bond of the 13-cis, 15-syn isomer. For furthernel. (b) Three-dimensional rendering of two possible orientations of retinal in thes to all-trans, 15-anti isomer of the dark-adapted state. Only the orientation in (a)Orientation (a) is also consistent with the X-ray crystal structure [143]. Adapted

2989M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
in the M intermediate has been obtained from 2H NMRmeasurements with wild-type bR [8]. These experimentsshowed that the angle between this bond and the membranenormal increased by 7.5° in M, consistent with a movementof the polyene chain in the direction of the indole ring ofW182. Here we report a substantial reorientation of the C13-methyl group near the Schiff base end of retinal in light–darkadaptation, as well as formation of the M state. The stericinteraction between the C13-methyl group and the side chainof W182 in helix F may thus be an essential element in thecoupling between the chromophore and the protein leading tothe tilt of helix F.
Large differences in the 2H NMR spectra were observed fortheM state versus the dark- and light-adapted forms. In Fig. 9 wedisplay the difference 2H NMR spectra of bR deuterium labeledin the C13-methyl group between the M, light-adapted, anddark-adapted states at a membrane tilt angle of 15°. The topshows a comparison of M and dark-adapted spectra; the middlecomparesM and light-adapted spectra; and the bottom comparesthe light-adapted and dark-adapted spectra. For light- and dark-adapted states the lineshapes are clearly different from those forthe M intermediate, in particular at tilt angles of 0° (not shown)and 15°. For a tilt angle of 15° the difference 2HNMR spectra forbacteriorhodopsin clearly indicate that the C13-methyl angleincreases in the sequence darkb lightbMstate. In theM state theC13-methyl bond angle with respect to the membrane normalwas found to increase by ≈7°, as confirmed subsequently in the
Table 1Summary of structural analysis of retinal conformation in bacteriorhodopsin and rho
Bacteriorhodopsin (−50°C)
Dark-adapted Light-adapted
Methyl bond orientation (θB)/dega
C1R 69±2 b 70±3 c
C5 37±2 d 36±3 c
C9 40±3 d 40±3 c
C13 25±3 g 34±3 g
(Roll, Tilt) angles/deg h
C1 to C7 (A plane) – (24, 74)C6 to C12 (B plane) – (24, 74)C12 to C15 (C plane) – (24, 74)
Torsion angle/deg i
C5_C6–C7_C8 180 180C10–C11_C12–C13 0 0
C11_C12–C13_C14 0 0
a Bond orientations represent angle between the retinylidene C–CH3 axis and locaθB cannot be distinguished from its supplement 180°−θB.b Ref. [7].c Ref. [8].d Ref. [6].e Ref. [77].f Ref. [11].g This work.h Roll and tilt angles are calculated for the plane containing the C5-to-N axis for bR
(C plane) for rhodopsin [9].i For bR retinal is assumed to be planar all-trans [7]. For rhodopsin constraints
dichroism data where available [77]. The simple three-plane model considers only Cincludes torsional twisting about the C11_C12 double bond.
X-ray crystal structure [98]. As a result we conclude that we havestructural changes towards the Schiff base end of thechromophore. The substantial differences in the 2H NMRspectra are due to a reorientation of the C13-methyl bond at the“business end” of the retinal, adjacent to the site of trans–cisisomerization.
Next, let us consider the tilt series of 2H NMR spectraacquired for bR deuterated at the C13-methyl group of thebound retinal ligand in the M state. Representative 2H NMRspectra are shown in Fig. 10(a) for bR trapped in M in thepresence of GuHCl at −50 °C. Compared to our previous 2HNMR spectra for the other methyl groups of M trapped inGuHCl-treated films at high pH [8], the present spectra are ofhigher quality. They show moreover a stronger dependence onthe tilt angle reflecting the smaller mosaic spread (σ≈10°versus ≈13.5°). This is due to a number of significant improve-ments in sample preparation. For the quadrupolar couplingconstant and the line broadening, the same values were used forthe M state as for the light-adapted and dark-adapted states. It isapparent from Fig. 10(a) that the spectra (unsymmetrized) arestrongly dependent on the tilt angle, as expected for an orientedsample. The results of the data analysis are summarized inTable 1. It was assumed that only one population was present(13-cis, 15-anti). This assumption is further justified by the factthat at the conclusion of the 2H NMR experiments visual in-spection showed that the sample was still completely bleached.The quality of the fit can be judged best by the residuals
dopsin
Rhodopsin
M state Dark-adapted (−150 °C) Meta I (−100 °C)
73±3 c – –39±3 c 70±3 e 72±4 f
47±3 c 52±3 e 53±3 f
40±3 g 68±2 e 59±3 f
(24, 79) (−180, 15) (−126, 31)(24, 79) (46, 62) (50, 71)(24, 79) (−115, 119) (simple) (56, 68)
(−112, 99) (extended)
180 −65 e ∓32 f
0 0 (simple) e 0−25 (extended)
0 147 (simple) e ±173 f
176 (extended)
l normal to membrane surface. Due to even parity of the quadrupolar interaction
[7], or the C2-to-C6 axis (A plane), C6-to-C12 axis (B plane), or C12-to-N axis
include solid-state 13C NMR dipolar couplings, linear dichroism, and circular6–C7 and C12–C13 dihedral angles; while the extended three-plane model also

Fig. 10. Solid-state 2H NMR spectroscopy reveals conformational changes ofretinal chromophore during the bacteriorhodopsin photocycle. (a) Experimental2H NMR tilt series (left) and the corresponding least squares fit (right) for bR2H-labeled in the C13-methyl group of retinal and trapped in theM intermediateat −50 °C. The tilt angles are indicated between the spectra and their best fits.All the spectra (data as well as simulations) have been scaled such that theirintegrals are the same. The residuals between data and fit are also plotted in theleft panel. The best global fit of the complete tilt series (shown on the right) wasobtained with a bond angle of 40.3° for the C13–C2H3 bond. (b) Retinalorientation in the M state (13-cis, shaded atoms) and the light-adapted state (all-trans, white atoms) of bR obtained from 2H NMR constraints for the C1R-methyl and C5-methyl bonds. The C5-to-N vector characterizes the molecularlong axis and is close to the electronic transition dipole moment in the all-transconformer. Its inclination with respect to the membrane normal is designated bythe angle αP and has a value of 74° in the ground state and 79° in the Mintermediate; this change is largely due to the all-trans to 13-cis isomerization.The C5 atom is assumed to have the same coordinates in both states. Adaptedwith permission from Ref. [8].
Fig. 9. Difference 2H NMR spectra for bacteriorhodopsin deuterium labeled inthe C13–C2H3 bond indicate conformational variation of retinal cofactorbetween the M, light-adapted, and dark-adapted states. (Top) Comparison of Mand dark-adapted spectra; (middle) comparison of M and light-adapted spectra;and (bottom) comparison of light-adapted and dark-adapted spectra. In each casethe first spectrum has the bold black color and the second spectrum the graycolor; where the tilt angle is 15°. The differences between the normalized spectraare plotted below each pair of spectra.
2990 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
between data and fit which is the quantity minimized in the leastsquares fit, cf. Fig. 10(a).
It is currently believed that during the lifetime of M, theaccessibility and pK of the Schiff base change from a state inwhich a proton has just been released to the extracellular side ofthe membrane to a state with higher pK, in which the Schiff baseis ready to pick up a proton from D96 on the cytoplasmic side ofthe membrane. This so-called reprotonation switch would re-quire at least two sequential M intermediates. There is con-sensus that under our conditions an M substrate accumulatesthat is very similar to the late M state that was first characterizedin the mutant D96N [99]. Strong evidence for the occurrence oftwo sequential M states was recently obtained from solid-stateNMR with wild-type samples in 0.3 M GuHCl at pH 10.0 [100].Illumination at −60 or −10 °C led to M states with Schiff base15N chemical shifts that differed by 7 ppm. The initial M stateproduced by illumination at the lower temperature decayedupon warming to the late M state. This transition was ac-companied by a substantial protein conformational change andan increase in the pK of the Schiff base [100]. Apparently, theSchiff base becomes more disposed to reprotonation in theinitial M to late M transition. Since we produced M byillumination of our GuHCl-treated samples at +5°C, we havepresumably also prepared this late M “post switch” state, inwhich helix F is tilted. A direct comparison of the retinalorientation in the M state to the light-adapted state is provided inFig. 10(b).
4.2. Solid-state NMR results are used for structural refinementof photocycle intermediates of bacteriorhodopsin
To recapitulate at this point, by considering the retinalchromophore to be almost planar in the dark- and light-adapted

2991M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
states its orientation can be defined by the roll and tilt anglesversus the membrane [7]. In this way we were able to establishits conformation and orientation within the bR binding cavity.For the dark- and light-adapted states, the data indicate thatthe methyl groups on the polyene chain point toward thecytoplasmic side of the membrane; whereas the N–H bond ofthe Schiff base points to the extracellular side, i.e., toward theside of proton release in the pump pathway. The tilt angle for theC5-to-N axis was found to increase by ≈5° in the M state;whereas the roll angle remained about the same (Table 1). Wethus confirmed that the Schiff base end of the chromophoreswitches its orientation relative to the membrane surface uponisomerization from all-trans to 13-cis in the transition to the Mstate.
Several high-resolution X-ray diffraction models of thestructure of bR have appeared for the light-adapted ground state[62,101], the dark-adapted state [102], for the low temperatureK intermediate [103], for the L intermediate [104], for the earlyM intermediate [105], and for the late M intermediate [98,106,107]. In particular, crystal structures have been determined forthe light-adapted state with an all-trans, 15-anti chromophore,as well as the dark-adapted state with a 13-cis, 15-syn isomer ofretinal, i.e. the bR13-c conformer. For the light-adapted state, theresults for the C13–C2H3 bond orientation fall in a rathernarrow range from 19.0° to 23.2° with an average of 21.5°. Forthe C9–C2H3 bond the values vary more, between 33.8° and41.3° with an average of 36.9°. The C5–C2H3 bond is in therange from 33.4° to 39.8° with an average of 35.8°. Thesevalues underscore that the polyene chain is curved yielding thesmallest angle for the C13–C2H3 bond. This in-plane curvatureis also a feature of retinal model compounds with a protonatedSchiff base [108], i.e. in the absence of interactions with abinding pocket. The agreement between the 2H NMR and X-rayresults for the light-adapted state is excellent for the C5–C2H3
and C9–C2H3 bonds, taking into account the errors of 2–3° foreach method. It should be mentioned that for the C5–C2H3 andC9–C2H3 bonds no significant differences were obtained be-tween the light- and dark-adapted 2H NMR spectra [8]. Yet forthe C13–C2H3 bond of the light-adapted state, there appears tobe a real discrepancy. Our 2H NMR value of 34.2° cannot bereconciled with the average X-ray value of 21.5° obtained withlight-adapted bR crystals. Possible reasons for this differencehave been discussed above.
Now in the M intermediate the averaged values from X-raycrystallography for the C5- and C13-methyl bond orientationsare 38.2° and 43.7°, in excellent agreement with the cor-responding 2H NMR results (Table 1). For the C13–C2H3 bondangle there is no significant difference between the X-raydiffraction results for early M (mutant E204Q) and late M(mutant D96N and wild-type). The four X-ray measurementsaverage to 38.0°, which is in excellent agreement with our 2HNMR result of 40.3° for the C13-methyl group (Fig. 10). Hencethe increased bond angle for the C13-methyl group in M thatwe detected here by 2H NMR is in complete agreement withthe X-ray diffraction results. Our previous observation of asimilar angular increase for the C9–C2H3 bond also fits verywell with the late M structure [98]. The X-ray and 2H NMR
experiments for the C13-methyl group both indicate asignificant increase in the angle between the light-adaptedstate and M, but the change is considerably larger for thediffraction method (16–18° versus 6°). As discussed above, thedifferent values for the light-adapted angles are responsible forthis difference.
One of the most interesting questions is at what point in thephotocycle these angular changes occur. Several lines ofevidence using spectroscopic methods have shown that themethyl groups of the chromophore occupy specific sites in thebinding pocket that are important in the photocycle [90,109].The results from X-ray diffraction experiments with bR trappedin the low temperature K intermediate [103] and in the Lintermediate [104] indicate that already in the earlier K (13-cis,15-anti) and L (13-cis, 15-anti) intermediates the orientation ofthe C13–C2H3 bond is 39.6° and 37.6° respectively, i.e. closeto the 2H NMR value of 40.3° in the M state. Hence thedeprotonation in the L to M transition does not seem to affectmuch the chromophore structure as judged by the methyl bondorientations. Early neutron diffraction and linear dichroismwork had already indicated that the C5-to-C13 part of thepolyene chain tilts up (towards the cytoplasmic side) in M dueto the kink in the chain introduced by the C13_C14isomerization [13]. X-ray diffraction experiments on the lateM intermediate of the mutant D96N clearly reveal a movementof the C13-methyl group by 1.3 Å towards the cytoplasmic sideof the membrane [98]. The indole ring of W182 is displacedby 1.5 Å in the same direction, as if being pushed up by themovement of the C13-methyl group [98]. The W182 residue isin helix F, which undergoes a major structural change in M[110].
4.3. Conformational changes are localized to the Schiff baseend of retinal chromophore in bR photocycle
All of these observations taken together lead to a consistentmolecular picture for the proton pump. In the case of bR theretinal chromophore is nearly planar in the light- and dark-adapted states, as well upon isomerization to 13-cis in the Mstate. The isomerization from all-trans, 15-anti to 13-cis, 15-anti gives a local change at the site of the Schiff base whichswitches the N–H orientation from one side of the membrane tothe other. This light-induced molecular motion actively trans-ports H3O
+ ions against the electrochemical potential gradient,leading to energy conservation involving Schiff base deproto-nation in the M state. In the dark-adapted state with a 2/3population in the bR13-c configuration, differences with thebRall-t conformer are expected since protons are only pumped inthe latter form. Further evidence for subtle structural differencesbetween these two isomers in the dark-adapted state comes fromhigh-resolution solid-state NMR experiments [95,96]. More-over, the change in the C13–C2H3 angle of 9° between bR13-c
and bRall-t is larger than the change of 6° in the transition fromlight-adapted to M. The curved polyene chain in the light-adapted state changes its conformation in M, thereby pushingon the indole side chain of W182 of helix F. This couplingbetween chromophore and binding pocket may well be the

Fig. 11. Orientation-dependent 2H NMR spectra for aligned rhodopsin/POPC(1:50) recombinant membranes provide angular restraints for retinylidene ligandin the dark state. (a)–(c) 2H NMR spectra for 11-Z-[5-C2H3]-retinylidenerhodopsin (blue), 11-Z-[9-C2H3]-retinylidene rhodopsin (magenta), and 11-Z-[13-C2H3]-retinylidene rhodopsin (green) at pH 7 and T=−150 °C. Theoreticallineshapes for an immobile uniaxial distribution (solid lines) are superimposedon the experimental 2H NMR spectra. Note that characteristic lineshape changesare observed as a function of the tilt angle, which manifest the different methylbond orientations with respect to the membrane frame. Reproduced withpermission from Ref. [77].
2992 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
essential step in triggering the protein conformational change inlate M that involves helix F.
To summarize, we have established that the changes uponforming the 13-cis isomer in the M state mainly involve the“business end” of the chromophore nearest to the Schiff base.The C13-methyl of the chromophore changes its orientation tothe membrane upon photon absorption, moving towards W182,thus driving the proton pump in energy conservation. Whileonly minor differences were detected between dark- and light-adapted states, the C5–, C9–, and C13–C2H3 angles all in-creased in the M state. The C9–C2H3 bond showed the largestorientational change of 7° in M. Changes of the C13-methylgroup orientation illuminate differences among the light-adapted, dark-adapted, and M states. The various states aredistinct and the conformation at the “business end” of thechromophore as manifested by the C13-methyl orientation isdifferent in each case.
5. Application of solid-state 2H NMR to G protein-coupledreceptor rhodopsin
The visual pigment rhodopsin is an example of a G protein-coupled receptor, where the ligand is 11-cis retinal. The acti-vation of rhodopsin in the visual process and the X-ray crystalstructure [63,64,72,73,111] have been reviewed [112–118].Rhodopsin has seven transmembrane helices with a ligand-binding pocket in its center, and an additional surface helix towhich post-translational lipid modifications are attached. Herethe retinal chromophore functions as a light-activated switch,and generates a signaling state in a time-ordered sequence that isthermodynamically irreversible. For rhodopsin, the site-directed2H NMR approach was used for structural analysis of retinalwithin its binding cavity in the dark-adapted (11-cis) and pre-activated meta I (all-trans) states. The meta I and meta II statesof rhodopsin correspond to the low-affinity and high-affinityforms of ligand-activated GPCRs. Three sites of interactionwithin the binding cavity are implicated, viz. the protonatedSchiff base and its associated counterion; the polyene chain withits C9-methyl group; and last the β-ionone ring which is in ahydrophobic pocket. Our hypothesis is that specific methylbinding sites are necessary for rhodopsin pigment formation andphotoreceptor activation [69]. The interaction sites are probedby 2H NMR of the C5-, C9-, and C13-methyl groups. Solid-state 2H NMR of the planar-supported bilayers provided theorientation of retinal as well as its conformation within theprotein binding pocket. Differences in the retinal conformationand orientation as revealed by solid-state 2H NMR spectroscopyare significant for explaining the mechanism of action of bothbacteriorhodopsin and rhodopsin.
5.1. Solid-state 2H NMR of rhodopsin in oriented membranesgives angular restraints for structural analysis
How can the residual quadrupolar couplings (RQCs) in 2HNMRyield insight into the retinal structure bound to rhodopsin?Rhodopsin poses a challenging application of the 2H NMRtechnology due to the larger molar mass than bR, with appre-
ciable extracellular domains that can affect membrane align-ment. The bleached apoprotein was regenerated with 11-Z-[5-C2H3]-, 11-Z-[9-C
2H3]-, and 11-Z-[13-C2H3]-retinals, i.e. 11-cis retinal deuterated at the C5-, C9-, or C13-methyl groups byorganic total synthesis [119]. It was then recombined withPOPC followed by alignment of the membrane bilayers onplanar glass substrates by isopotential ultracentrifugation[9,120]. From a tilt series of the oriented sample in the magneticfield and an analysis of the 2H NMR lineshapes, the anglesbetween the individual C–C2H3 bonds and the membranenormal could be determined, even in the presence of a sub-stantial degree of orientational disorder. We measured a tiltseries of 2H NMR spectra below the order–disorder transitiontemperature (Tm) of the membrane lipid bilayers. Below Tmrhodopsin is in a state of substantially reduced mobility, due tothe presence of gel-state lipids, as shown by freeze-fractureelectron microscopy (EM) studies [121] and spin-label EPRspectroscopy [74,75]. Our 2H NMR studies show that theretinylidene methyl groups are all rotating at temperatures downto at least −150 °C [9,67], thus providing an efficient relaxationmechanism; long recycle delays are unnecessary. Fig. 11 showsthe complete tilt series of angular-dependent 2H NMR spectraacquired for rhodopsin in the dark state containing retinal

Fig. 12. Global fitting of 2H NMR spectra for 11-cis retinal in the dark state ofrhodopsin gives methyl bond orientations and mosaic spread of alignedmembranes. (a)–(c) RMSD of calculated versus experimental 2H NMR spectrafor retinal deuterated at C5-, C9-, or C13-methyl groups, respectively and(d)–(f) cross-sections through hypersurfaces. Distinct minima are found in thebond orientation θB and mosaic spread σ of aligned membranes. Reproducedwith permission from Ref. [77].
2993M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
deuterated at the C5-, C9-, or C13-methyl groups in the POPCmembranes at T=−150 °C. Characteristic lineshape changesare observed as a function of the tilt angle, which differ forthe various deuterated methyl positions, due to the differentC–C2H3 bond orientations with respect to the membrane plane.
Some qualitative deductions can be made simply from theangular dependence of the 2H NMR spectra. From the ap-pearance of the θ=90° spectral shoulders we can say that thebond orientation θB must be greater than ≈30–45° in each case[16,17], cf. Fig. 3(b). However due to the alignment disorderone must simulate the lineshapes to obtain more accurateinformation. Theoretical simulations in Fig. 11 assume a staticuniaxial distribution [16] of the rhodopsin molecules withrotating methyl groups, and are superimposed on the experi-mental 2H NMR data. Similar results were obtained attemperatures of T=−60° and −30 °C [119]. Simultaneousfitting of the 2H NMR lineshapes gives the orientations of thevarious methyl C–C2H3 axes relative to the bilayer normal(Table 1), together with the mosaic spread of aligned samples.In accord with results for powder-type samples (Fig. 1), weobtained off-axial order parameters for the rotating methylgroups of SC3
≈0.9. Excellent agreement between the experi-mental and simulated lineshapes is found in all cases, asdemonstrated by the flat residuals (not shown).
The results of global fitting of 2H NMR lineshapes areshown in Fig. 12 (left) for the C5-, C9-, or C13-methyl groupsof retinal bound to rhodopsin in POPC membranes in the darkstate. The RMSD of the calculated 2H NMR spectra from theexperimental data for the entire tilt series is graphed as afunction of θB and σ, and differs significantly for the various2H-labeled sites. In Fig. 12 (right) we display cross-sectionsthrough the RMSD surfaces at the global minimum for the C5-,C9-, and C13-methyl groups, respectively. Accurate bondorientations θB of the 2H-labeled methyl groups are obtained,despite the appreciable mosaic spread (σ=18–21°). Values forthe mosaic spread obtained from the 2H NMR lineshapesimulations are larger than for aligned purple membranescontaining bR [7]. This may be due to the fact that rhodopsin(Mr=40 kDa) is a larger molecule than bR (Mr =26 kDa), withsubstantial extramembranous domains [122]. We determinedvalues of the C–C2H3 bond orientations of 70±3°, 52±3°,and 68±2° for the C5-, C9-, and C13-methyl groups ofretinal in the dark state of rhodopsin (Table 1) [9]. Theseresults are in excellent agreement with the 2.2 Å X-raystructure of rhodopsin [63].
5.2. Analysis of 2H NMR data reveals structural changes uponbleaching of rhodopsin
For rhodopsin, the 2H NMR lineshape analysis had as its aimto determine the conformation of the retinal ligand and toinvestigate distortion of the chromophore linked to itsphotochemistry. What is the 3D structure and dynamics of theretinal chromophore in rhodopsin as viewed by 2H NMR? Foranalysis of the retinal conformation, we applied a simple modelwith three planes of unsaturation [9,123] (Fig. 13), encompass-ing the β-ionone ring and the polyene chain to either side of the
C12–C13 bond [9,69,88,123]. The relative orientations of theC5-, C9-, and C13-methyl groups specify the C6–C7 and C12–C13 dihedral angles, as shown in Fig. 13. Each plane(designated A, B, or C) is defined by two vectors, and requirestwo angular constraints (degrees of freedom) to define its spatialorientation (yielding a total of four angular constraints for threeplanes with shared bonds). Thus one must introduce restraintsfrom linear dichroism studies [86]; yet unique roll and tilt anglesare unobtainable. Rather, four (4) solutions are found for eachplane due to the even parity, giving 4×4=16 combinations foreach torsion angle connecting adjacent planes. It follows thatthe results of additional biophysical methods are needed toobtain the unique solutions.
5.2.1. Torsional twisting of retinal chromophore indark-adapted state (11-cis)
For structural analysis, it is most accurate to combine angularand distance restraints, as in solution NMR spectroscopy [61],using 13C-to-13C distances from rotational resonance 13C NMRstudies [85]. Circular dichroism (CD) data are also available forthe dark state of rhodopsin, which yield information on theenantiomeric selectivity of the protein binding pocket [87,88].The most plausible physical solution is shown in Fig. 13 andyields a positively twisted, 12-s-trans conformation, where the

Fig. 14. Solid-state NMR provides the structure of retinal ligand of the canonicalGPCR rhodopsin. Vertical direction corresponds to membrane normal; theextracellular side is up and cytoplasmic side is down. The 2H NMR structure of11-cis retinal in the dark state (green) is compared to retinal structure in the pre-activated metarhodopsin I state (red). Figure prepared with PyMOL [http://pymol.sourceforge.net/].
Fig. 13. Solid-state 2H NMR spectroscopy yields conformation and orientationof 11-cis retinal ligand in the dark state of rhodopsin. Retinal is described bythree planes of unsaturation (designated A, B, C). (a) Simple three-plane modelwith torsional twisting only about C6–C7 and C12–C13 bonds. (b) Extendedthree-plane model with additional pre-twisting about C11_C12 double bond.Reproduced with permission from Ref. [77].
2994 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
C13- and C9-methyl groups point oppositely from the mem-brane plane, and the retinylidene β-ionone ring is negativelytwisted 6-s-cis. In Fig. 13(a) a simple three-plane model isassumed which gives dihedral angles of χ9,13=+147±4 andχ5,9=−65±6° (Table 1). The calculated roll and tilt angles forthe planes of unsaturation are also included in Table 1. Analysisof the conformation of retinal in rhodopsin from the 2H NMRdata indicates that the chromophore is highly non-planar, incontrast to bR. Compared to bR the roll angle of the central partof the polyene from C6 to C12 is increased, with the torsionaltwisting affecting both the β-ionone ring and the retinal Schiffbase (Table 1). By contrast, for bR the all-trans retinal is nearlyplanar and the β-ionone ring is 6-s-trans [7]. The 11-cis-retinylidene structure derived from 2H NMR can be furtherrefined by inserting it into the binding pocket of the X-raystructure of rhodopsin in the dark state (2.2 Å) [63]. Yet due tosteric clashes a simple model is not readily accommodatedwithin the rhodopsin binding cavity. As indicated in Fig. 13(b),an extended three-plane model can then be introduced to allowfor pre-twisting of the C11_C12 cis-double bond in the direc-tion of the isomerization [77]. The positive helical twist aboutthe C12–C13 bond is in agreement with CD and bioorganicstudies of locked retinoids [88] and with our previous results[9]. Pre-twisting about the 11-cis-double bond is consistent withX-ray [63] and quantum-mechanical calculations [124] and cangovern the chromophore movements upon photoexcitation,thereby affecting the ultra-fast reaction dynamics [77,125].
Next, Fig. 14 shows the structure of retinal obtained by 2HNMR [77] inserted into the binding pocket of the rhodopsin X-ray structure [63]. Occupation of specific methyl sitescontributes to the dihedral twisting of the retinal chromophorein the dark state of rhodopsin. Torsional twisting of the polyenestems from localization of the β-ionone ring within itshydrophobic pocket at one end of the chromophore, togetherwith the salt bridge of the retinylidene Schiff base at the otherend, and the C9-methyl positioned between T118 and W268.The absolute sense of the helical twists of the 11-cis, 12-s-transand 6-s-cis conformations manifest the chiral selectivity of thebinding site, which is related to the trajectory of the 11-cis totrans photoisomerization. The retinal conformation is lockedwithin the binding pocket, which accounts for its phenomenaldark stability. The distance from the C13-methyl group to theclosest carbon atom of the W265 indole ring is ≈4 Å,suggesting that interactions of retinal with W265 help stabilizethe dark-state conformation [34]. Moreover, the configurationof the –C_NH+– bond of the protonated Schiff base is antiand its hydrogen points oppositely from the C13-methyl group,i.e. toward the extracellular side in the direction of thecounterion E113.
5.2.2. Conformational relaxation in metarhodopsin I state(all-trans)
What are the changes in the conformation and orientation ofthe retinal chromophore of rhodopsin that are coupled to lightactivation of the receptor? At present X-ray structures areavailable for bleached rhodopsin in the bathorhodopsin andlumirhodopsin states [5], and an electron density map for meta I

2995M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
from electron crystallography [126]. The next step was tomeasure the tilt series of 2H NMR spectra for aligned mem-branes containing rhodopsin in the all-transmeta I state. The 2HNMR lineshape analysis for meta I gave the orientation θB ofthe various methyl bonds as summarized in Table 1. NMRstructures were calculated for meta I in terms of a three-planemodel as described [11]. Besides the C5-, C9-, and C13-methylbond orientations, the electronic transition dipole momentobtained from linear dichroism was introduced as a furtherorientational restraint [127]. Some of the NMR structures wereeliminated based on electron crystallography data indicatingthat in the meta I state the β-ionone ring remains in its groundstate position [77,119,126]. The calculated meta I structureswere further restrained by inserting them into the dark-staterhodopsin structure [63]. It was assumed that the shape of theretinal binding cavity was nearly the same in meta I as in thedark state, in agreement with electron crystallography [126].The structure with χ5,9=−32°, and χ9,13=173° fits best withinthe binding cleft of the dark-state X-ray structure, having itsonly close contact with the side chain of W265. Although thepolyene chain is relaxed in meta I, the β-ionone ring retains itstorsional strain as in the dark state.
One can then address how this conformational distortion ispropagated through the protein allosteric network, leading tounfolding of the cytoplasmic loops and activation of thereceptor [122]. In Fig. 14 we compare the 2H NMR structureproposed for all-trans retinal in the meta I state (red), as com-pared to the NMR structure of 11-cis retinal in the dark-adaptedstate of rhodopsin (green). The 11-cis to all-trans isomerizationof retinal is in the negative direction due to the helicity of theC11_C12 bond in the dark state, together with the sterichindrance within the binding pocket [125]. Assuming a three-plane model, the Schiff base hydrogen would switch fromfacing the extracellular side in the dark state to the cytoplasmicside in meta I. This may have implications for a (possiblycomplex) counterion switch [128–130], as discussed below.Steric hindrance between trans retinal and W265 can beinvolved with triggering the transition to the active meta II state.Thus our work provides a basis for investigating the changes inretinal conformation and orientation giving the signaling meta IIstate in visual excitation.
5.3. Combining 2H NMR data with molecular dynamicssimulations further illuminates rhodopsin function
An additional aspect entails conceptual integration of theexperimental 2H NMR findings with molecular dynamics (MD)simulations [122,130–132]. Large-scale MD simulations wereused to simulate the chromophore environment in both the darkstate [132] as well as the pre-activated meta I state [130]. In thedark state, a total of 23 independent, 100-ns all-atom MDsimulations were analyzed for rhodopsin embedded in a lipidbilayer in the microcanonical (N,V,E) ensemble. It was foundthat the polyene chain of retinal is rigidly locked into a single,twisted conformation, consistent with its function as an inverseagonist in the dark state. By contrast, the β-ionone ring ismobile within its binding pocket; the cavity is sufficiently large
to enable structural heterogeneity. It was found that the β-ionone ring occupies two distinct conformations in the darkstate, contrary to previous assumptions. The β-ionone ring canrotate relative to the polyene chain, thereby populating bothpositively and negatively twisted 6-s-cis enantiomers. Moreoverthe influences of 11-cis to trans isomerization of the retinalhave been investigated through ultra-long simulations ofrhodopsin with trajectories of 1000–2000 ns [130]. In themeta I state, analysis of the MD and NMR data gives insightsinto the counterion switch mechanism that stabilizes the pro-tonated Schiff base (PSB). Deprotonation of the retinal PSB isthermodynamically the most important event that characterizesthe subsequent activated meta II state. Comparison of thesimulated 2H NMR spectra with experimental data supports acomplex counterion mechanism in which both E113 and E181stabilize the retinal PSB in the meta I state prior to forming theactivated meta II state of rhodopsin [130].
6. Comparison of retinal proteins
Rapid light-induced isomerization around a specific doublebond of the chromophore is a common feature in the mechanismof many photoreceptors (rhodopsin, phytochrome, PYP,sensory rhodopsin) and light-driven pumps (bacteriorhodopsin,halorhodopsin). The photoisomerization leads to a change in theconfiguration and orientation of the chromophore in its bindingpocket, and is essential for the coupling between the chromo-phore and the protein. In a series of slower thermal transitions,the altered chromophore structure drives the protein into theactive state (signaling or transport). The structural switch thatwas originally localized in the chromophore leads on a slowertime scale to global structural changes in parts of the protein thatare far away from the binding pocket (e.g. the cytoplasmic loopdomain in the meta II state of rhodopsin or the signal trans-duction domain in phytochrome).
This raises the following questions. How is the structuralchange of the chromophore transmitted to the protein? What isthe mechanism of this chromophore–protein interaction(coupling)? Active reorientational motions of the chromophoremost likely play an important role in these processes.Structural evidence for a major transient chromophorereorientation has been obtained for the photoactive yellowprotein (PYP) in which the chromophore changes itsorientation by approximately 60° in the blue-shifted I2intermediate [133]. But already much smaller movements areexpected to induce significant effects. In the retinal proteins,for example, even a small movement of the protonated Schiffbase will lead to major changes in electrostatic interactionsand pK values. In this regard 2H NMR studies of the tworetinal proteins are illuminating both in terms of theirsimilarities and differences, which are connected with theirrather different biological functions. For rhodopsin and bR thestructural changes triggered by the photoisomerization maywell be mediated by steric interactions between the protrudingmethyl groups of the chromophore and amino acid side chainsin the binding pocket [29,62,109,134–137]. The angularrestraints from 2H NMR for retinal within the binding pocket

2996 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
can then be used in conjunction with X-ray crystallography,where available, to investigate the light-induced changes thatlead to biological function, either proton pumping in the caseof bR, or amplified visual excitation in the case of rhodopsin.Additional 2H NMR investigations of their photointermediatesare clearly desirable.
For bR the retinal cofactor is almost planar from the β-iononering (apart from ring puckering) to the C15 atom of the polyene[7]. Deviations from planarity (mostly less than 15°) mainlyinvolve twisting in the vicinity of the C14–C15 bond near theSchiff base, as indicated by NMR [39], FTIR [93], X-raycrystallography [105], and hybrid quantum-mechanical/mole-cular mechanical calculations [138]. At this point it is not clearto what extent these deviations affect retinal isomerization inbR. Hypothetically, they could be responsible for differencesin the potential energy surfaces and consequently the efficiencyand selectivity of the isomerization. Yet it is known that thephotoisomerization efficiency exhibited in bR is due totolerance of the protein cavity for the atomic rearrangementsin all-trans to 13-cis isomerization [139]. As a result, in thelight-adapted state the retinal orientation is essentially describedby the roll and tilt angles of the nearly planar all-trans chro-mophore [7]. The chromophore plane has a roll angle of 24° andthe long axis is inclined by 16° to the membrane plane. Retinalisomerization from all-trans to 13-cis in the dark-adapted or Mstate keeps this approximate planarity, with a decrease in theinclination of the C5-to-N axis from 16° in the light-adaptedstate to 11° in the M state; while the C5-to-C13 axis moves outof the membrane plane by 4–5° [8]. Hence the structuralchanges mainly occur at the “business end” of the retinalmolecule near the Schiff base linkage. All-trans to 13-cisisomerization switches the orientation of the N–H bond fromthe extracellular side to the cytoplasmic side and is implicated inthe mechanism of the proton pump. For bR strong indirectevidence for a steric interaction between the C9- and C13-methyl groups of the chromophore and the indole ring ofW182 in the photocycle has been obtained from spectroscopicmethods employing mutants and retinal analogs [109,134].
By contrast in rhodopsin the most intriguing aspect comparedto bR is the torsional twisting of the ligand, involving both the β-ionone ring [9,30] and the polyene chain [77]. The significancefor rhodopsin is that retinal distortion is implicated in its ultra-fastisomerization and high quantum yield. In the dark-adapted statethe retinal cofactor is 11-cis and the polyene is highly distorted, asshown by resonance Raman spectroscopy [140], X-ray crystal-lography [63], 2H NMR [77], and quantum-mechanical calcula-tions [124,125]. The NMR conformational analysis for rhodopsinis more challenging than for bR due to the additional degrees offreedom, as well as the greater difficulty of preparing alignedspecimens. We have assumed an analysis in terms of three planesof unsaturation, corresponding to three different sites ofinteraction of the retinal ligand with the protein binding cavity.In comparison to bR, the angle the central polyene plane makes tothe membrane plane is less, with substantial twisting affecting theβ-ionone ring and the Schiff base. Moreover, combining thesolid-state 2H NMR data with the X-ray crystal structure for dark-state rhodopsin suggests that an extended three-plane model may
be appropriate, including the torsional twist about the activatedC11_C12 bond [77]. Hence the C11_C12 double bond is pre-twisted within the rhodopsin binding pocket in the direction of theisomerization. The resulting 2H NMR structure explains its dark-state stability and indicates the direction of the ultra-fastphotoisomerization. Activation of the receptor involves sequen-tial relief of the torsional strain of the ligand (frustration) and iscoupled to interactions among the protein, and the surroundinglipid bilayer, which are all involved in the activation mechanismof rhodopsin [15].
In closing, for both bacteriorhodopsin and rhodopsininteractions of retinal with the protein binding cavitydetermine the isomerization pathway selectivity and effi-ciency. As a result the same ligand molecule undergoes vastlydifferent pathways of isomerization in these retinal proteins.Despite the great deal of experimental data, many aspects ofthe cofactor interactions with the protein are still unknown,and further studies are required to fully illuminate theirfunctioning. Within this context, solid-state NMR spectro-scopy can be expected to play a continued prominent roletogether with other complementary spectroscopic and compu-tational approaches.
7. Experimental procedures
Solid-state 2HNMR spectroscopy employed Bruker AMX-300 and AMX-500spectrometers equipped with locally-built high-power probes that included high-voltage capacitors and a horizontal solenoidal inductor (1.8–4.2 μs 90° pulsesdepending on the radiofrequency coil diameter and frequency) [9,54,55]. Aquadrupolar echo sequence with composite pulses was applied with appropriatephase-cycling to accumulate the 2HNMR signals. Bacteriorhodopsinwas obtainedfromHalobacterium salinarium strain ET1001 and was regenerated with synthetic2H-labeled all-trans retinal followed by alignment of the purple membranes [7].Well-ordered films were obtained by slowly drying a purple membrane suspensionon glass (or quartz or mylar) supports [7]. For the dark-adapted state, the samplewas first brought to room temperature and then cryotrapped at−50 °C. TheM statewas formed by illuminating GuHCl-treated films of wild-type bR at pH 10.5 withyellow light (550 nm band-pass filter) at 5 °C, which caused the purple films tobleach to a bright yellow, and it was cryotrapped at −78 °C [8]. Light adaptationmade use of the fact that bR molecules that are photoreversed from the M state byblue light [142] have an all-trans chromophore [8], which were cryotrapped at−78 °C. Bovine rhodopsin was prepared with 2H-labeled 11-cis retinal and POPCrecombinant membranes were aligned on planar glass slides by isopotential spin-dry centrifugation [9,41]. Metarhodopsin I was formed in POPCmembranes at pH6 at 2 °C by exposure to green light (550 nm high-pass filter) and then cryotrappedat −78 °C. Extensive studies of the photochemical behavior of rhodopsin indifferent membrane environments were conducted to establish conditions fortrapping the photointermediates [14,15,141].
Acknowledgements
Research described in this article was supported by NIHgrants EY 12049 (M. F. B.) and GM 36564 (K. N.), NSF grantCHE-607917 (M.F.B.), and by Deutsche Forschungsge-meinschaft grant Sfb 498 (M. P. H.). We are especially gratefulto our colleagues and laboratory members for their manycontributions to this work. Organic synthesis of retinal benefitedfrom the collaboration of S. Krane, N. Fujioka, N. Sakai, andK. Tanaka whose efforts are warmly acknowledged. We alsoexpress our appreciation to R. Bogomolni andW. L. Hubbell forstimulating discussions.

2997M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
References
[1] A. Watts, Solid-state NMR in drug design and discovery for membrane-embedded targets, Nat. Rev. Drug Disc. 4 (2005) 555–568.
[2] J.K. Lanyi, Bacteriorhodopsin, Annu. Rev. Physiol. 66 (2004) 665–688.[3] D. Salom, D.T. Lodowski, R.E. Stenkamp, I. Le Trong, M. Golczak, B.
Jastrzebska, T. Harris, J.A. Ballesteros, K. Palczewski, Crystal structureof a photoactivated deprotonated intermediate of rhodopsin, Proc. Natl.Acad. Sci. U. S. A. 103 (2006) 16123–16128.
[4] T. Okada, K. Palczewski, Crystal structure of rhodopsin: implications forvision and beyond, Curr. Op. Struct. Biol. 11 (2001) 420–426.
[5] H. Nakamichi, T. Okada, Local peptide movement in the photoreactionintermediate of rhodopsin, Proc. Natl. Acad. Sci. U. S. A. 103 (2006)12729–12734.
[6] A.S. Ulrich, I. Wallat, M.P. Heyn, A. Watts, Re-orientation of retinal inthe M-photointermediate of bacteriorhodopsin, Nat. Struct. Biol. 2 (1995)190–192.
[7] S. Moltke, A.A. Nevzorov, N. Sakai, I. Wallat, C. Job, K. Nakanishi,M.P. Heyn, M.F. Brown, Chromophore orientation in bacteriorho-dopsin determined from the angular dependence of deuterium NMRspectra of oriented purple membranes, Biochemistry 37 (1998)11821–11835.
[8] S.M. Moltke, I. Wallat, N. Sakai, K. Nakanishi, M.F. Brown, M.P. Heyn,The angles between the C1-, C5-, and C9-methyl bonds of the retinylidenechromophore and the membrane normal increase in the M intermediate ofbacteriorhodopsin: direct determination with solid-state 2H NMR,Biochemistry 38 (1999) 11762–11772.
[9] G.F.J. Salgado, A.V. Struts, K. Tanaka, N. Fujioka, K. Nakanishi, M.F.Brown, Deuterium NMR structure of retinal in the ground state ofrhodopsin, Biochemistry 43 (2004) 12819–12828.
[10] A.B. Patel, E. Crocker, M. Eilers, A. Hirshfeld, M. Sheves, S.O. Smith,Coupling of retinal isomerization to the activation of rhodopsin, Proc.Natl. Acad. Sci. U. S. A. 101 (2004) 10048–10053.
[11] G.F.J. Salgado, A.V. Struts, K. Tanaka, S. Krane, K. Nakanishi, M.F.Brown, Solid-state 2H NMR structure of retinal in metarhodopsin I,J. Am. Chem. Soc. 128 (2006) 11067–11071.
[12] B. Borucki, H. Otto, M.P. Heyn, Reorientation of the retinylidenechromophore in the K, L, and M intermediates of bacteriorhodopsin fromtime-resolved linear dichroism: resolving kinetically and spectrally over-lapping intermediates of chromoproteins, J. Phys. Chem. B 103 (1999)6371–6383.
[13] M.P. Heyn, B. Borucki, H. Otto, Chromophore reorientation during thephotocycle of bacteriorhodopsin: experimental methods and functionalsignificance, Biochim. Biophys. Acta 1460 (2000) 60–74.
[14] A.V. Botelho, N.J. Gibson, Y. Wang, R.L. Thurmond, M.F. Brown, Con-formational energetics of rhodopsin modulated by nonlamellar forminglipids, Biochemistry 41 (2002) 6354–6368.
[15] A.V. Botelho, T. Huber, T.P. Sakmar, M.F. Brown, Curvature and hydro-phobic forces drive oligomerization and modulate activity of rhodopsin inmembranes, Biophys. J. 91 (2006) 4464–4477.
[16] A.A. Nevzorov, S. Moltke, M.P. Heyn, M.F. Brown, Solid-state NMRline shapes of uniaxially oriented immobile systems, J. Am. Chem. Soc.121 (1999) 7636–7643.
[17] A.S. Ulrich, A. Watts, I. Wallat, M.P. Heyn, Distorted structure of theretinal chromophore in bacteriorhodopsin resolved by 2H-NMR,Biochemistry 33 (1994) 5370–5375.
[18] G. Gröbner, I.J. Burnett, C. Glaubitz, G. Choi, A.J. Mason, A. Watts,Observations of light-induced structural changes of retinal withinrhodopsin, Nature (London) 405 (2000) 810–813.
[19] R.R.Vold, Nucleic acid flexibility and dynamics: deuteriumNMR, in: D.M.Grant, R.K. Harris (Eds.), Encyclopedia of Nuclear Magnetic Resonance,Wiley, New York, 1996, pp. 3314–3320.
[20] A.A. Nevzorov, S. Moltke, M.F. Brown, Structure of the A-form and B-form of DNA from deuterium NMR line shape simulation, J. Am. Chem.Soc. 120 (1998) 4798–4805.
[21] H. Luecke, J.K. Lanyi, Structural clues to the mechanism of ion pumpingin bacteriorhodopsin, Adv. Prot. Chem. 63 (2003) 111–130.
[22] F. Fanelli, P.G. De Benedetti, Computational modeling approaches tostructure-function analysis of G protein-coupled receptors, Chem. Rev.105 (2005) 3297–3351.
[23] U. Häberlen, High Resolution NMR in Solids: Selective Averaging,Academic Press, New York, 1976.
[24] H.W. Spiess, Rotation of molecules and nuclear spin relaxation, in: P.Diehl, E. Fluck, R. Kosfeld (Eds.), NMR Basic Principles and Progress,Springer-Verlag, Heidelberg, 1978, pp. 55–214.
[25] M.F. Brown, Membrane structure and dynamics studied with NMRspectroscopy, in: K.M. Merz Jr., B. Roux (Eds.), Biological Membranes:A Molecular Perspective from Computation and Experiment, Birkhäuser,Basel, 1996, pp. 175–252.
[26] F. Creuzet, A. McDermott, R. Gebhard, K. van der Hoef, M.B. Spijker-Assink, J. Herzfeld, J. Lugtenburg, M.H. Levitt, R.G. Griffin, Deter-mination of membrane protein structure by rotational resonance NMR:bacteriorhodopsin, Science 251 (1991) 783–786.
[27] L.K. Thompson, A.E. Mcdermott, J. Raap, C.M. Vanderwielen, J.Lugtenburg, J. Herzfeld, R.G. Griffin, Rotational resonance NMR studyof the active site structure in bacteriorhodopsin: conformation of theSchiff base linkage, Biochemistry 31 (1992) 7931–7938.
[28] A.E. McDermott, F. Creuzet, R. Gebhard, K. van der Hoef, M.H. Levitt,J. Herzfeld, J. Lugtenberg, R.G. Griffin, Determination of internucleardistances and the orientation of functional groups by solid-state NMR:rotational resonance study of the conformation of retinal in bacteriorho-dopsin, Biochemistry 33 (1994) 6129–6136.
[29] P.J.E. Verdegem, P.H.M. Bovee-Geurts, W.J. de Grip, J. Lugtenburg,H.J.M. de Groot, Retinylidene ligand structure in bovine rhodopsin,metarhodopsin-I, and 10-methylrhodopsin from internuclear distancemeasurements using 13C-labeling and 1-D rotational resonance MASNMR, Biochemistry 38 (1999) 11316–11324.
[30] P.J.R. Spooner, J.M. Sharples, S.C. Goodall, H. Seedorf, M.A. Verhoeven,J. Lugtenburg, P.H.M. Bovee-Geurts, W.J. DeGrip, A. Watts, Conforma-tional similarities in the β-ionone ring region of the rhodopsin chromo-phore in its ground state and after photoactivation to the metarhodopsin-Iintermediate, Biochemistry 42 (2003) 13371–13378.
[31] M. Carravetta, X. Zhao, O.G. Johannessen, W.C. Lai, M.A.Verhoeven, P.H.M. Bovee-Geurts, P.J.E. Verdegem, S. Kiihne, H.Luthman, H.J.M. de Groot, W.J. deGrip, J. Lugtenburg, M.H. Levitt,Protein-induced bonding perturbation of the rhodopsin chromophoredetected by double-quantum solid-state NMR, J. Am. Chem. Soc. 126(2004) 3948–3953.
[32] T. Fujiwara, Y. Todokoro, H. Yanagishita, M. Tawarayama, T. Kohno, K.Wakamatsu, H. Akutsu, Signal assignments and chemical-shift structuralanalysis of uniformly 13C, 15N-labeled peptide, mastoparan-X, by multi-dimensional solid-state NMR under magic-angle spinning, J. Biomol.NMR 28 (2004) 311–325.
[33] O.C. Andronesi, S. Becker, K. Seidel, H. Heise, H.S. Young, M. Baldus,Determination of membrane protein structure and dynamics by magic-angle-spinning solid-state NMR spectroscopy, J. Am. Chem. Soc. 127(2005) 12965–12974.
[34] E. Crocker, M. Eilers, S. Ahuja, V. Hornak, A. Hirshfeld, M. Sheves, S.O.Smith, Location of Trp265 in metarhodopsin II: implications for theactivation mechanism of the visual receptor rhodopsin, J. Mol. Biol. 357(2006) 163–172.
[35] J.L. Lorieau, A.E. McDermott, Conformational flexibility of a micro-crystalline globular protein: order parameters by solid-state NMRspectroscopy, J. Am. Chem. Soc. 128 (2006) 11505–11512.
[36] Y. Todokoro, I. Yumen, K. Fukushima, S.-W. Kang, J.-S. Park, T. Kohno,K. Wakamatsu, H. Akutsu, T. Fujiwara, Structure of tightly membrane-bound mastoparan-X, a G-protein-activating peptide, determined bysolid-state NMR, Biophys. J. 91 (2006) 1368–1379.
[37] X. Feng, P.J.E. Verdegem, Y.K. Lee, D. Sandström, M. Edén, P. Bovee-Geurts, W.J. deGrip, J. Lugtenburg, H.J.M. deGroot, M.H. Levitt, Directdetermination of a molecular torsional angle in the membrane proteinrhodopsin by solid-state NMR, J. Am. Chem. Soc. 119 (1997)6853–6857.
[38] C.M. Rienstra, M. Hohwy, L.J. Mueller, C.P. Jaroniec, B. Reif, R.G.Griffin, Determination of multiple torsion-angle constraints in U–13C,

2998 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
15N-labeled peptides: 3D 1H–15N–13C–1H dipolar chemical shiftNMR spectroscopy in rotating solids, J. Am. Chem. Soc. 124 (2002)11908–11922.
[39] J.C. Lansing, M. Hohwy, C.P. Jaroniec, A.F.L. Creemers, J. Lugtenburg,J. Herzfeld, R.G. Griffin, Chromophore distortions in the bacteriorho-dopsin photocycle: evolution of the H–C14–C15–H dihedral anglemeasured by solid-state NMR, Biochemistry 41 (2002) 431–438.
[40] T.P. Trouard, T.M. Alam, M.F. Brown, Angular dependence of deuteriumspin-lattice relaxation rates of macroscopically oriented dilaurylpho-sphatidylcholine in the liquid-crystalline state, J. Chem. Phys. 101 (1994)5229–5261.
[41] G. Gröbner, A. Taylor, P.T.F. Williamson, G. Choi, C. Glaubitz, J.A.Watts, W.J. deGrip, A. Watts, Macroscopic orientation of natural andmodel membranes for structural studies, Anal. Biochem. 254 (1997)132–138.
[42] M.F. Mesleh, S. Lee, G. Veglia, D.S. Thiriot, F.M. Marassi, S.J. Opella,Dipolar waves map the structure and topology of helices in membraneproteins, J. Am. Chem. Soc. 125 (2003) 8928–8935.
[43] A.A. Nevzorov, S.J. Opella, Structural fitting of PISEMA spectra ofaligned proteins, J. Magn. Res. 160 (2003) 33–39.
[44] Q. Teng, L.K. Nicholson, T.A. Cross, Experimental determination oftorsion angles in the polypeptide backbone of the gramicidin A channelby solid state nuclear magnetic resonance, J. Mol. Biol. 218 (1991)607–619.
[45] R.R. Ketchem, W. Hu, T.A. Cross, High-resolution conformation ofgramicidin A in a lipid bilayer by solid-state NMR, Science 261 (1993)1457–1460.
[46] C.H.Wu,A.Ramamoorthy, S.J. Opella, High-resolution heteronuclear dipolarsolid-state NMR spectroscopy, J. Magn. Reson. A 109 (1994) 270–272.
[47] R.Q. Fu, T.A. Cross, Solid-state nuclear magnetic resonance investigationof protein and polypeptide structure, Annu. Rev. Biophys. Biomol. Struct.28 (1999) 235–268.
[48] S.J. Opella, F.M. Marassi, J.J. Gesell, A.P. Valente, Y. Kim, M. Oblatt-Montal, M. Montal, Structures of the M2 channel-lining segments fromnicotinic acetylcholine and NMDA receptors by NMR spectroscopy, Nat.Struct. Biol. 6 (1999) 374–379.
[49] J. Wang, J. Denny, C. Tian, S. Kim, Y. Mo, F. Kovacs, Z. Song, K.Nishimura, Z. Gan, R. Fu, J.R. Quine, T.A. Cross, Imaging membraneprotein helical wheels, J. Magn. Reson. 144 (2000) 162–167.
[50] S.J. Opella, C. Ma, F.M. Marassi, Nuclear magnetic resonance ofmembrane-associated peptides and proteins, Meth. Enzymol. 339 (2001)285–313.
[51] K.A. Henzler-Wildman, G.V. Martinez, M.F. Brown, A. Ramamoorthy,Perturbation of the hydrophobic core of lipid bilayers by the humanantimicrobial peptide, LL-37, Biochemistry 43 (2004) 8459–8469.
[52] A. Ramamoorthy, Y.F. Wei, D.K. Lee, PISEMA solid-state NMRspectroscopy, Annu. Rep. NMR Spectr. 52 (2004) 1–52.
[53] A.S. Ulrich, M.P. Heyn, A. Watts, Structure determination of thecyclohexene ring of retinal in bacteriorhodopsin by solid-state deuteriumNMR, Biochemistry 31 (1992) 10390–10399.
[54] A.A. Nevzorov, T.P. Trouard, M.F. Brown, Lipid bilayer dynamics fromsimultaneous analysis of orientation and frequency dependence ofdeuterium spin-lattice and quadrupolar order relaxation, Phys. Rev. E58 (1998) 2259–2281.
[55] T.P. Trouard, A.A. Nevzorov, T.M. Alam, C. Job, J. Zajicek, M.F. Brown,Influence of cholesterol on dynamics of dimyristoylphosphatidylcholineas studied by deuterium NMR relaxation, J. Chem. Phys. 110 (1999)8802–8818.
[56] C. Glaubitz, I.J. Burnett, G. Gröbner, A.J. Mason, A. Watts, Deuterium-MAS NMR spectroscopy on oriented membrane proteins: applications tophotointermediates of bacteriorhodopsin, J. Am. Chem. Soc. 121 (1999)5787–5794.
[57] A.E. McDermott, Structural and dynamic studies of proteins by solid-state NMR spectroscopy: rapid movement forward, Curr. Op. Struct.Biol. 14 (2004) 554–561.
[58] G.M. Clore, A.M. Gronenborn, New methods of structure refinement formacromolecular structure determination by NMR, Proc. Natl. Acad. Sci.U. S. A. 95 (1998) 5891–5898.
[59] A. Bax, Weak alignment offers new NMR opportunities to study proteinstructure and dynamics, Protein Sci. 12 (2003) 1–16.
[60] J.H. Prestegard, K.L. Mayer, H. Valafar, G.C. Benison, Determination ofprotein backbone structures from residual dipolar couplings, Meth.Enzymol. 394 (2005) 175–209.
[61] J. Cavanaugh, W.J. Fairbrother, A.G. Palmer III, N.J. Skelton, M. Rance,Protein NMR Spectroscopy: Principles and Practice, Second Edition,Academic Press, New York, 2006.
[62] H. Luecke, B. Schobert, H.-T. Richter, J.-P. Cartailler, J.K. Lanyi, Struc-ture of bacteriorhodopsin at 1.55 Å resolution, J. Mol. Biol. 291 (1999)899–911.
[63] T. Okada, M. Sugihara, A.-N. Bondar, M. Elstner, P. Entel, V. Buss, Theretinal conformation and its environment in rhodopsin in light of a new2.2 Å crystal structure, J. Mol. Biol. 342 (2004) 571–583.
[64] J. Li, P.C. Edwards, M. Burghammer, C. Villa, G.F.X. Schertler, Structureof bovine rhodopsin in a trigonal crystal form, J. Mol. Biol. 343 (2004)1409–1438.
[65] S.O. Smith, I. Palings, M.E. Miley, J. Courtin, H. de Groot, J. Lugtenberg,R.A. Mathies, R.G. Griffin, Solid-state NMR studies of the mechanism ofthe opsin shift in the visual pigment rhodopsin, Biochemistry 29 (1990)8158–8164.
[66] G.S. Harbison, S.O. Smith, J.A. Pardoen, J.M.L. Courtin, J. Lugtenburg,J. Herzfeld, R.A. Mathies, R.G. Griffin, Solid-state 13C NMR detection ofa perturbed 6-s-trans chromophore in bacteriorhodopsin, Biochemistry24 (1985) 6955–6962.
[67] V. Copié, A.E. McDermott, K. Beshah, J.C. Williams, M. Spyker-Assink,R.T. Gebhard, J. Lugtenberg, J. Herzfeld, R.G. Griffin, Deuterium solid-state NMR studies of methyl group dynamics in bacteriorhodopsin andretinal model compounds: evidence for a 6-s-trans chromophore in theprotein, Biochemistry 33 (1994) 3280–3286.
[68] J. Lugtenburg, The synthesis of 13C-labeled retinals, Pure Appl. Chem. 57(1985) 753–762.
[69] K. Nakanishi, R. Crouch, Application of artificial pigments to structuredetermination and study of photoinduced transformations of retinalproteins, Isr. J. Chem. 35 (1995) 253–272.
[70] A.Wada, N. Fujioka, Y. Tanaka,M. Ito, A highly stereoselective synthesisof 11Z-retinal using tricarbonyliron complex, J. Org. Chem. 65 (2000)2438–2443.
[71] A.F.L. Creemers, J. Lugtenburg, The preparation of all-trans uniformly13C-labeled retinal via a modular total organic synthetic strategy.Emerging central contribution of organic synthesis toward the structureand function study with atomic resolution in protein research, J. Am.Chem. Soc. 124 (2002) 6324–6334.
[72] K. Palczewski, T. Kumasaka, T. Hori, C.A. Behnke, H. Motoshima, B.A.Fox, I. Le Trong, D.C. Teller, T. Okada, R.E. Stenkamp, M. Yamamoto,M. Miyano, Crystal structure of rhodopsin: a G protein-coupled receptor,Science 289 (2000) 739–745.
[73] T. Okada, Y. Fujiyoshi, M. Silow, J. Navarro, E.M. Landau, Y. Shichida,Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography, Proc. Natl. Acad. Sci. U. S. A. 99 (2002)5982–5987.
[74] A. Kusumi, J.S. Hyde, Spin-label saturation-transfer electron spinresonance detection of transient association of rhodopsin in reconstitutedmembranes, Biochemistry 21 (1982) 5978–5983.
[75] N.J.P. Ryba, D. Marsh, Protein rotational diffusion and lipid/proteininteractions in recombinants of bovine rhodopsin with saturateddiacylphosphatidylcholines of different chain lengths studied by con-ventional and saturation-transfer electron spin resonance, Biochemistry 31(1992) 7511–7518.
[76] H.I. Petrache, S.W. Dodd, M.F. Brown, Area per lipid and acyl lengthdistributions in fluid phosphatidylcholines determined by 2H NMRspectroscopy, Biophys. J. 79 (2000) 3172–3192.
[77] A.V. Struts, G.F.J. Salgado, K. Tanaka, S. Krane, K. Nakanishi, M.F.Brown, Structural analysis and dynamics of retinal chromophore in darkand meta I states of rhodopsin from 2H NMR of aligned membranes,J. Mol. Biol. 372 (2007) 50–66.
[78] D.C. Rees, G. Chang, R.H. Spencer, Crystallographic analyses of ionchannels: lessons and challenges, J. Biol. Chem. 275 (2000) 713–716.

2999M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
[79] D. Fu, A. Libson, L.J.W. Miercke, C. Weitzman, P. Nollert, J. Krucinski,R.M. Stroud, Structure of a glycerol-conducting channel and the basis forits selectivity, Science 290 (2000) 481–486.
[80] M.E. Rose, Elementary Theory of Angular Momentum, Wiley, NewYork, 1957.
[81] D.M. Brink, G.R. Satchler, Angular Momentum, Oxford UniversityPress, London, 1968.
[82] A.S. Ulrich, A. Watts, 2H-NMR lineshapes of immobilized uniaxiallyoriented membrane proteins, Solid State Nucl. Magn. Reson. 2 (1993)21–36.
[83] M.F. Brown, S. Lope-Piedrafita, G.V. Martinez, H.I. Petrache. Solid-statedeuterium NMR spectroscopy of membranes, in: G.A. Webb (Ed.),Modern Magnetic Resonance, Springer, Heidelberg 2006, pp. 245–256.
[84] H.I. Petrache, M.F. Brown, X-ray scattering and solid-state deuteriumnuclear magnetic resonance probes of structural fluctuations in lipidmembranes, in: A.M. Dopico (Ed.), Methods in Molecular Biologyvol. 400, Methods in Membrane Lipids, Humana, Totowa, 2007,pp. 341–353.
[85] P.J.R. Spooner, J.M. Sharples, M.A. Verhoeven, J. Lugtenberg, C.Glaubitz, A. Watts, Relative orientation between the β-ionone ring andthe polyene chain for the chromophore of rhodopsin in native mem-branes, Biochemistry 41 (2002) 7549–7555.
[86] S. Jäger, J.W. Lewis, T.A. Zvyaga, I. Szundi, T.P. Sakmar, D.S. Kliger,Chromophore structural changes in rhodopsin from nanoseconds tomicroseconds following pigment photolysis, Proc. Natl. Acad. Sci.U. S. A. 94 (1997) 8557–8562.
[87] Y. Fujimoto, J. Ishihara, S. Maki, N. Fujioka, T. Wang, T. Furuta, N.Fishkin, B. Borhan, N. Berova, K. Nakanishi, On the bioactiveconformation of the rhodopsin chromophore: absolute sense of twistaround the 6-s-cis bond, Chem. Eur. J. 7 (2001) 4198–4204.
[88] Y. Fujimoto, N. Fishkin, G. Pescitelli, J. Decatur, N. Berova, K.Nakanishi, Solution and biologically relevant conformations of enantio-meric 11-cis-locked cyclopropyl retinals, J. Am. Chem. Soc. 124 (2002)7294–7302.
[89] T. Hauss, S. Grzesiek, H. Otto, J. Westerhausen, M.P. Heyn,Transmembrane location of retinal in bacteriorhodopsin by neutrondiffraction, Biochemistry 29 (1990) 4904–4913.
[90] Y. Yamazaki, J. Sasaki, M. Hatanaka, H. Kandori, A. Maeda, R.Needleman, T. Shinada, K. Yoshihara, L.S. Brown, J.K. Lanyi,Interaction of tryptophan-182 with the retinal 9-methyl group in theL intermediate of bacteriorhodopsin, Biochemistry 34 (1995)577–582.
[91] A. Maeda, J.E. Morgan, R.B. Gennis, T.G. Ebrey, Water as a cofactor inthe unidirectional light-driven proton transfer steps in bacteriorhodopsin,Photochem. Photobiol. 82 (2006) 1398–1405.
[92] F. Garczarek, K. Gerwert, Functional waters in intraprotein protontransfer monitored by FTIR difference spectroscopy, Nature 439 (2006)109–112.
[93] N. Mizuide, M. Shibata, N. Friedman, M. Sheves, M. Belenky, J.Herzfeld, H. Kandori, Structural changes in bacteriorhodopsin followingretinal photoisomerization from the 13-cis form, Biochemistry 45 (2006)10674–10681.
[94] J.M.J. Swanson, C.M. Maupin, H. Chen, M.K. Petersen, J. Xu, Y. Wu,G.A. Voth, Proton solvation and transport in aqueous and biomolecularsystems: insights from computer simulations, J. Phys. Chem. B 111(2007) 4300–4314.
[95] M. Helmle, H. Patzelt, A. Ockenfels, W. Gärtner, D. Oesterhelt, B.Bechinger, Refinement of the geometry of the retinal binding pocket indark-adapted bacteriorhodopsin by heteronuclear solid-state NMRdistance measurements, Biochemistry 39 (2000) 10066–10071.
[96] J.M. Griffiths, A.E. Bennett, M. Engelhard, F. Siebert, J. Raap, J.Lugtenburg, J. Herzfeld, R.G. Griffin, Structural investigation of theactive site in bacteriorhodopsin: geometric constraints on the roles ofAsp-85 and Asp-212 in the proton-pumping mechanism from solid stateNMR, Biochemistry 39 (2000) 362–371.
[97] P. Scherrer, M.K. Mathew, W. Sperling, W. Stoeckenius, Retinal isomerratio in dark-adapted purple membrane and bacteriorhodopsin monomers,Biochemistry 28 (1989) 829–834.
[98] H. Luecke, B. Schobert, H.T. Richter, J.P. Cartailler, J.K. Lanyi,Structural changes in bacteriorhodopin during ion transport at 2Angstrom resolution, Science 286 (1999) 255–260.
[99] F.M.H. Betancourt, R.M. Glaeser, Chemical and physical evidence formultiple functional steps comprising the M state of the bacteriorhodopsinphotocycle, Biochim. Biophys. Acta 1460 (2000) 106–118.
[100] J.G. Hu, B.Q. Sun, M. Bizounok, M.E. Hatcher, J.C. Lansing, J. Raap,P.J.E. Verdegem, J. Lugtenburg, R.G. Griffin, J. Herzfeld, Early and lateM intermediates in the bacteriorhodopsin photocycle: a solid-state NMRstudy, Biochemistry 37 (1998) 8088–8096.
[101] E. Pebay-Peyroula, G. Rummel, J.P. Rosenbusch, E.M. Landau, X-raystructure of bacteriorhodopsin at 2.5 angstroms from microcrystals grownin lipid cubic phases, Science 277 (1997) 1676–1681.
[102] T. Nishikawa, M. Murakami, T. Kouyama, Crystal structure of the 13-cisisomer of bacteriorhodopsin in the dark-adapted state, J. Mol. Biol. 352(2005) 319–328.
[103] K. Edman, P. Nollert, A. Royant, H. Belrhali, E. Pebay-Peyroula, J.Hajdu, R. Neutze, E.M. Landau, High-resolution X-ray structure of anearly intermediate in the bacteriorhodopsin photocycle, Nature 401(1999) 822–826.
[104] A. Royant, K. Edman, T. Ursby, E. Pebay-Peyroula, E.M. Landau, R.Neutze, Helix deformation is coupled to vectorial proton transport in thephotocycle of bacteriorhodopsin, Nature 406 (2000) 645–648.
[105] H. Luecke, B. Schobert, J.-P. Cartailler, H.-T. Richter, A. Rosengarth, R.Needleman, J.K. Lanyi, Coupling photoisomerization of retinal to direc-tional transport in bacteriorhodopsin, J. Mol. Biol. 300 (2000)1237–1255.
[106] H.J. Sass, G. Büldt, R. Gessenich, D. Hehn, D. Neff, R. Schlesinger,J. Berendzen, P. Ormos, Structural alterations for proton translocationin the M state of wild-type bacteriorhodopsin, Nature 406 (2000)649–653.
[107] K. Takeda, Y. Matsui, N. Kamiya, S. Adachi, H. Okumura, T. Kouyama,Crystal structure of the M intermediate of bacteriorhodopsin: allostericstructural changes mediated by sliding movement of a transmembranehelix, J. Mol. Biol. 341 (2004) 1023–1037.
[108] B.D. Santarsiero, M.N.G. James, M. Mahendran, R.F. Childs, Crystal-structure of N-methyl-N-phenylretinal iminium perchlorate: a structuralmodel for the bacteriorhodopsin chromophore, J. Am. Chem. Soc. 112(1990) 9416–9418.
[109] S. Hashimoto, K. Obata, H. Takeuchi, R. Needleman, J.K. Lanyi,Ultraviolet resonance Raman spectra of Trp-182 and Trp-189 inbacteriorhodopsin: novel information on the structure of Trp-182 andits steric interaction with retinal, Biochemistry 36 (1997) 11583–11590.
[110] S. Subramaniam, R. Henderson, Molecular mechanism of vectorialproton translocation by bacteriorhodopsin, Nature 406 (2000) 653–657.
[111] D.C. Teller, T. Okada, C.A. Behnke, K. Palczewski, R.E. Stenkamp,Advances in determination of a high-resolution three-dimensional struc-ture of rhodopsin, a model of G-protein-coupled receptors (GPCRs),Biochemistry 40 (2001) 7761–7772.
[112] M.F. Brown, Influence of non-lamellar forming lipids on rhodopsin, Curr.Top. Membr. 44 (1997) 285–356.
[113] T. Ebrey, Y. Koutalos, Vertebrate photoreceptors, Prog. Ret. Eye Res. 20(2001) 49–94.
[114] T. Okada, O.P. Ernst, K. Palczewski, K.P. Hofmann, Activation ofrhodopsin: new insights from structural and biochemical studies, TrendsBiochem. Sci. 26 (2001) 318–324.
[115] T.P. Sakmar, S.T. Menon, E.P. Marin, E.S. Awad, Rhodopsin: insightsfrom recent structural studies, Annu. Rev. Biophys. Biomol. Struct. 31(2002) 443–484.
[116] W.L. Hubbell, C. Altenbach, C.M. Hubbell, H.G. Khorana, Rhodopsinstructure, dynamics, and activation: a perspective from crystallography,site-directed spin labeling, sulfhydryl reactivity, and disulfide cross-linking, Adv. Prot. Chem. 63 (2003) 243–290.
[117] K. Palczewski, G protein-coupled receptor rhodopsin, Annu. Rev.Biochem. 75 (2006) 743–767.
[118] K.D. Ridge, K. Palczewski, Visual rhodopsin sees the light: structureand mechanism of G protein signaling, J. Biol. Chem. 282 (2007)9297–9301.

3000 M.F. Brown et al. / Biochimica et Biophysica Acta 1768 (2007) 2979–3000
[119] K. Tanaka, A.V. Struts, S. Krane, N. Fujioka, G.F.J. Salgado, K.Martínez-Mayorga, M.F. Brown, K. Nakanishi, Synthesis of CD3-labeled11-cis-retinals and applications to solid-state deuterium NMR spectro-scopy of rhodopsin, Bull. Chem. Soc. Japan 80 (2007) 2177–2184.
[120] G. Gröbner, G. Choi, I.J. Burnett, C. Glaubitz, P.J.E. Verdegem, J.Lugtenberg, A. Watts, Photoreceptor rhodopsin: structural and conforma-tional study of its chromophore 11-cis retinal in oriented membranes bydeuterium solid state NMR, FEBS Lett. 422 (1998) 201–204.
[121] Y.S. Chen, W.L. Hubbell, Temperature- and light-dependent structuralchanges in rhodopsin-lipid membranes, Exp. Eye Res. 17 (1973) 517–532.
[122] T. Huber, A.V. Botelho, K. Beyer, M.F. Brown, Membrane model for theGPCR prototype rhodopsin: hydrophobic interface and dynamicalstructure, Biophys. J. 86 (2004) 2078–2100.
[123] N. Fishkin, N. Berova, K. Nakanishi, Primary events in dim light vision: achemical and spectroscopic approach toward understanding protein/chromophore interactions in rhodopsin, Chem. Rec. 4 (2004) 120–135.
[124] M. Sugihara, J. Hufen, V. Buss, Origin and consequences of steric strainin the rhodopsin binding pocket, Biochemistry 45 (2006) 801–810.
[125] J.A. Gascon, V.S. Batista, QM/MM study of energy storage and molec-ular rearrangements due to the primary event in vision, Biophys. J. 87(2004) 2931–2941.
[126] J.J. Ruprecht, T. Mielke, R. Vogel, C. Villa, G.F.X. Schertler, Electroncrystallography reveals the structure of metarhodopsin I, EMBO J. 23(2004) 3609–3620.
[127] M. Chabre, J. Breton, The orientation of the chromophore of vertebraterhodopsin in the “meta” intermediate states and the reversibility of themeta II–meta III transition, Vis. Res. 19 (1979) 1005–1018.
[128] E.C.Y. Yan, M.A. Kazmi, Z. Gani, J.-M. Hou, D. Pan, B.S.W. Chang, T.P.Sakmar, R.A. Mathies, Retinal counterion switch in the photoactivationof the G protein-coupled receptor rhodopsin, Proc. Natl. Acad. Sci.U. S. A. 100 (2003) 9262–9267.
[129] S. Lüdeke, R. Beck, E.C.Y. Yan, T.P. Sakmar, F. Siebert, R. Vogel, Therole of Glu181 in the photoactivation of rhodopsin, J. Mol. Biol. 353(2005) 345–356.
[130] K. Martínez-Mayorga, M.C. Pitman, A. Grossfield, S.E. Feller, M.F.Brown, Retinal counterion switch mechanism in vision evaluated bymolecular simulations, J. Am. Chem. Soc. 128 (2006) 16502–16503.
[131] T. Huber, K. Rajamoorthi, V.F. Kurze, K. Beyer, M.F. Brown, Structure ofdocosahexaenoic acid-containing phospholipid bilayers as studied by ce:sup>H/ce:sup>H NMR and molecular dynamics simulations, J. Am.Chem. Soc. 124 (2002) 298–309.
[132] P.-W. Lau, A. Grossfield, S.E. Feller, M.C. Pitman, M.F. Brown,Dynamic structure of retinylidene ligand of rhodopsin probed bymolecular simulations, J. Mol. Biol. 372 (2007) 906–917.
[133] U.K. Genick, G.E.O. Borgstahl, K. Ng, Z. Ren, C. Pradervand, P.M.Burke, V. Srajer, T.-Y. Teng,W. Schildkamp,D.E.McRee,K.Moffat, E.D.Getzoff, Structure of a protein photocycle intermediate by millisecondtime-resolved crystallography, Science 275 (1997) 1471–1475.
[134] O. Weidlich, B. Schalt, N. Friedman, M. Sheves, J.K. Lanyi, L.S. Brown,F. Siebert, Steric interaction between the 9-methyl group of the retinal andtryptophan 182 controls 13-cis to all-trans reisomerization and protonuptake in the bacteriorhodopsin photocycle, Biochemistry 35 (1996)10807–10814.
[135] G.G. Kochendoerfer, P.J.E. Verdegem, I. van der Hoef, J. Lugtenburg,R.A. Mathies, Retinal analog study of the role of steric interactions inthe excited state isomerization dynamics of rhodopsin, Biochemistry 35(1996) 16230–16240.
[136] R. Vogel, F. Siebert, S. Lüdeke, A. Hirshfeld, M. Sheves, Agonists andpartial agonists of rhodopsin: retinals with ring modifications, Biochem-istry 44 (2005) 11684–11699.
[137] R. Vogel, S. Lüdeke, F. Siebert, T.P. Sakmar, A. Hirshfeld, M. Sheves,Agonists and partial agonists of rhodopsin: retinal polyene methylationaffects receptor activation, Biochemistry 45 (2006) 1640–1652.
[138] S. Hayashi, I. Ohmine, Proton transfer in bacteriorhodopsin:structure, excitation, IR spectra, and potential energy surface analysesby an ab initio QM/MM method, J. Phys. Chem. B 104 (2000)10678–10691.
[139] F. Gai, K.C. Hasson, J.C. McDonald, P.A. Anfinrud, Chemical dynamicsin proteins: the photoisomerization of retinal in bacteriorhodopsin,Science 279 (1998) 1886–1891.
[140] P. Kukura, D.W. McCamant, S. Yoon, D.B. Wandschneider, R.A.Mathies, Structural observation of the primary isomerization in visionwith femtosecond-stimulated Raman, Science 310 (2005) 1006–1009.
[141] N.J. Gibson, M.F. Brown, Lipid headgroup and acyl chain compositionmodulate the MI–MII equilibrium of rhodopsin in recombinantmembranes, Biochemistry 32 (1993) 2438–2454.
[142] S. Dickopf, M.P. Heyn, Evidence for the first phase of the reprotonationswitch of bacteriorhodopsin from time-resolved photovoltage and flashphotolysis experiments on the photoreversal of the M-intermediate,Biophys. J. 73 (1997) 3171–3181.
[143] H. Luecke, H.-T. Richter, J.K. Lanyi, Proton transfer pathways in bacterio-rhodopsin at 2.3 angstrom resolution, Science 280 (1998) 1934–1937.
Copyright © 2022 FDOKUMEN




















