
Soleus H-reflex phase-dependent modulation is preserved during stepping within a robotic exoskeleton
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Soleus H-reflex phase-dependent modulation is preserved during stepping within a robotic exoskeleton

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Soleus H-reflex phase-dependent modulation is preserved during steppingwithin a robotic exoskeleton
Maria Knikou a,b,c,⇑, Nupur Hajela a,b, Chaithanya K. Mummidisetty a, Ming Xiao a,b, Andrew C. Smith a
a Sensory Motor Performance Program, Rehabilitation Institute of Chicago, Chicago, IL, USAb Department of Physical Medicine and Rehabilitation, Northwestern University, Chicago, IL, USAc The Graduate Center and Dept. of Physical Therapy, City University of New York/College of Staten Island, Staten Island, NY, USA
a r t i c l e i n f o
Article history:Accepted 14 December 2010Available online 14 January 2011
Keywords:LokomatH-reflex modulationHuman walkingRobotic gaitSpinal circuitsStepping
h i g h l i g h t s
� The soleus H-reflex is modulated in a phase-dependent manner during stepping within a roboticexoskeleton in healthy humans.� Body unloading and decreased EMG amplitude of ankle extensors are not key factors for the soleusH-reflex phasic excitability to be manifested.� Spinal interneuronal circuits involved in the phasic H-reflex excitability are engaged in a physiologicalmanner during robotic assisted stepping.� Robotic assisted stepping utilized for rehabilitation of walking in neurological disorders will not engagelocomotor spinal circuits in a pathological manner.
a b s t r a c t
Objective: To investigate to what extent the phase-dependent modulation of the soleus H-reflex is pre-served when bilateral leg movements are electromechanically driven by a robotic exoskeleton at differentlevels of body weight support (BWS) in healthy subjects.Methods: The soleus H-reflex was elicited by posterior tibial nerve stimulation with a 1-ms single pulseat an intensity that the M-waves ranged from 4% to 9% of the maximal M-wave across subjects. Stimuliwere randomly dispersed across the step cycle which was divided into 16 equal bins. At each bin, a max-imal M-wave was elicited 100 ms after the test H-reflex and was used to normalize the associated M-wave and H-reflex. Electromyographic (EMG) activity from major hip, knee, and ankle muscles wasrecorded with surface bipolar electrodes. For each subject and muscle, the integrated EMG profile wasestablished and plotted as a function of the step cycle phases. The H-reflex gain was determined asthe slope of the relationship between the H-reflex and soleus EMG amplitudes at 100 ms before the H-reflex for each bin.Results: During robotic assisted stepping, the phase-dependent soleus H-reflex modulation pattern waspreserved and was similar at 25% and 50% BWS, a linear relationship between soleus H-reflex amplitudeand background activity was found, and the reflex gain did not change with alterations of the BWS level.EMG amplitudes were smaller at 50% compared to 25% BWS.Conclusions: Body unloading, decreased EMG amplitude of ankle extensors, and reduced ankle movementare not key factors for the soleus H-reflex phasic excitability to be manifested.Significance: Robotic devices are utilized for rehabilitation of gait in neurological disorders. Based on ourfindings, spinal interneuronal circuits involved in the phase-dependent modulation of the soleus H-reflexwill be engaged in a physiological manner during robotic assisted stepping in neurological disorders.� 2010 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights
reserved.
1. Introduction
During rhythmic motor tasks, sensory afferent feedback refinesmotor output to suit environmental demands. A plethora of studieshave provided evidence towards a sensory, reflex-mediated, regu-lation of locomotion that involves largely extensor muscle and
1388-2457/$36.00 � 2010 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.clinph.2010.12.044
⇑ Corresponding author. Address: Department of Physical Therapy, GraduateCenter of CUNY, 2800 Victory Boulevard, 5N-207, Staten Island, NY 10314, USA. Tel.:+1 718 982 3316; fax: +1 718 982 2984.
E-mail addresses: [email protected], [email protected](M. Knikou).
Clinical Neurophysiology 122 (2011) 1396–1404
Contents lists available at ScienceDirect
Clinical Neurophysiology
journal homepage: www.elsevier .com/locate /c l inph

Author's personal copy
plantar cutaneous afferents mediating load, and muscle afferentsmediating changes of muscle length (see reviews of Dietz andDuysens, 2000; Rossignol et al., 2006; Grillner, 2006; Knikou,2010a). In decerebrate acutely spinalized cats, stimulation of groupI ankle extensor afferents during fictive locomotion prolongedextensor nerve activity when delivered during an extensor burstand terminated flexor nerve activity when delivered during a flexorburst (Conway et al., 1987). It was later shown that the locomotorrhythm could be entrained by rhythmic contractions of ankleextensor muscles, hip extension and unloading are necessary forswing phase initiation while excitatory actions of group I afferentsare distributed to all extensor muscles (Angel et al., 1996; Grillnerand Rossignol, 1978; Guertin et al., 1995; Pearson et al., 1992;Whelan et al., 1995). In addition, cutaneous nerves and afferentsof the foot were shown to have similar effects to those describedfor group I afferents promoting correct foot placement andweight bearing during the stance phase (Bouyer and Rossignol,2003).
Ample evidence suggests that spinal reflex circuits integratinginformation from receptors registering stretch or load regulate hu-man locomotion. Their behavioral relevance is supported by theirsusceptibility to a task-, phase-, and use-dependent modulation(Zehr and Stein, 1999). In particular, the short latency ankle orquadriceps extensor reflexes (stretch or H-reflex) are progressivelyincreased from mid to late stance and significantly depressed oreven abolished during the swing phase of gait (Sinkjær et al.,1996; Mrachacz-Kersting et al., 2004; Capaday and Stein, 1986;Crenna and Frigo, 1987; Larsen et al., 2006; Dietz et al., 1990).The reflex facilitation during the stance phase may be partly re-lated to group I facilitatory actions exerted between ankle exten-sors (Faist et al., 2006). Given that the amount of loading and/orunloading is directly related to the motor output amplitude (Fouadet al., 2001; Sinkjær et al., 2000; Dietz and Colombo, 1996) and thatthe soleus H-reflex amplitude changes in parallel with the soleusEMG activity, one would expect that under conditions of reducedloading the soleus H-reflex modulation will be altered. However,a phase-dependent modulation of the soleus H-reflex has been re-ported during robotic air stepping (Kamibayashi et al., 2010), at 0%,25% and 50% body weight support (BWS) treadmill walking with-out robotic or manual leg guidance (Knikou et al., 2009), rhythmicone-legged step ascending and descending (Knikou, 2011), passivestepping movements (Brooke et al., 1995), and active pedalling(Boorman et al., 1992). Under these conditions, hip proprioceptorsmediating muscle stretch might have provided the gating mecha-nism for adaptation of the synaptic efficacy of the soleus Ia affer-ents (Knikou and Rymer, 2002a,b).
Body weight supported treadmill walking was introduced as ameans of rehabilitation of human walking after spinal cord lesions(Barbeau et al., 1987; Dobkin et al., 2003), based largely on evi-dence from spinalized cats (Barbeau and Rossignol, 1987; Edger-ton et al., 1997; Belanger et al., 1996; Rossignol et al., 1999).During this therapeutic intervention, BWS and manual assistanceis provided by therapists based on patient’s needs. This led to thedevelopment of a sophisticated exoskeleton system (Lokomat�),where manual assistance by therapists is replaced by electrome-chanically driven bilateral leg braces, leg movement patterns oc-cur in a pre-determined trajectory (Colombo et al., 2000), theankle movement is assisted by the foot lifters, and trunk rotationsare minimal. Accordingly, the objective of this study was to estab-lish the modulation pattern of the soleus H-reflex across multiplephases of the step cycle in healthy humans during robotic assistedstepping at BWS levels commonly utilized for rehabilitation ofwalking in neurological disorders while carefully controlling forchanges in the amplitude of the maximal M-wave across the stepcycle. Part of this study has been published in abstract form(Smith et al., 2011).
2. Materials and methods
The experimental protocol received Institutional Review Board(IRB) approval from the Northwestern University and CUNY Col-lege of Staten Island IRB committees. All procedures were con-ducted in compliance with the Declaration of Helsinki and awritten consent was obtained from each subject prior to study par-ticipation. Thirteen healthy subjects (5 male, 8 female) with agesranging from 21 to 51 (30.5 ± 11.4) years participated in the study.Their daily physical activities ranged from moderate to vigorous.No subject reported low back pain or any other type of neuromus-cular disorder.
2.1. EMG recordings
In all subjects, following standard skin preparation procedures,bipolar differential surface electrodes of fixed inter-electrode dis-tance (MA-411-002, Motion Lab Systems Inc., Baton Rouge, LA,USA) were used to record EMG activity from the soleus (SOL), med-ial gastrocnemius (MG), tibialis anterior (TA), peroneus longus (PL),medial hamstrings (MH), vastus lateralis (VL), rectus femoris (RF),and hip adductor gracilis (GRC) muscles from both legs. All EMGsignals were filtered with a cut-off frequency of 10–1000 Hz andsampled at 2000 Hz using a data acquisition card (NI PCI-6225,National Instruments, Austin, TX).
2.2. Robotic assisted stepping
A driven gait orthosis (DGO) system (Lokomat�, Hocoma, Swit-zerland) was utilized in this study for robotic assisted stepping. In-depth details on the Lokomat can be found elsewhere (Colomboet al., 2000; Jezernik et al., 2003). Briefly, the hip, knee, and anklejoint movements of each leg were assisted through separate posi-tion controllers implemented in a computer-based real time sys-tem while stepping on a moving treadmill (ADAL, AndrezieuxBoutheon, France). Subjects were fixed into the DGO using strapsat the waist, thighs, and shanks, which were adjusted at differentbody segments, allowing for adaptable fitting. Both feet were en-closed in foot lifters (using elastic straps and springs), which pro-moted ankle dorsiflexion during the swing phase of gait. TheDGO system was fixed to the treadmill through a parallel frame,while compensation for the weight of the DGO was also provided.Body weight was removed through an upper body harness thatwas connected to the orthotic system. Subjects were instructedto walk with the robotic device, and not to oppose the leg guidanceprovided by the DGO.
2.3. Elicitation and recording protocol of the soleus H-reflex duringrobotic assisted stepping
The soleus H-reflex was evoked according to methods previ-ously employed in healthy subjects during walking on a motorizedtreadmill at different BWS levels without leg assistance (Knikouet al., 2009). Square pulse stimuli of 1-ms duration were deliveredby a custom-built constant current stimulator to the right posteriortibial nerve. With subjects seated, a stainless steel plate of 4 cm2 indiameter was secured proximal to the patella. A hand-held mono-polar stainless steel head electrode was used to establish the mostoptimal stimulation site for the posterior tibial nerve, and corre-sponded to the site that the M-wave had a similar shape to thatof the H-reflex, and at the lowest stimulus intensity an H-reflexcould be evoked without an M-wave. The hand-held electrodewas then replaced by a pre-gelled disposable electrode (N-10-A;Medicotest, Ølstykee, Denmark), which was maintained under con-stant pressure throughout the experiment.
M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404 1397

Author's personal copy
Then, each subject came to a standing position on the tread-mill, and wore an upper body harness that was connected tooverhead pulleys. Thigh and shank segments of the robotic devicewere adjusted depending on each subject’s leg length and diam-eter, and both feet were secured into the foot lifters. The soleusH-reflex and M-wave recruitment curves (approximately 80 stim-uli in total delivered once every 5 s) were constructed duringstanding.
During walking, the orientation of the recording EMG electrodewith respect to the underlying muscle fibers may vary because ofchanges in the muscle architecture, which in turn may affect the
H-reflex amplitude (Gerilovsky et al., 1989). Further, flexion ofthe knee joint during the swing phase of gait moves the stimulat-ing electrode away from the tibial nerve, while extension duringthe stance phase has the opposite effect. Thus, appropriate adjust-ments of the stimulation intensity are required across the phases ofthe step cycle. Lastly, one of the most important factors in H-reflexstudies is the amplitude of the M-wave and H-reflex as a percent-age of the maximal M-wave (Mmax) (Knikou, 2008). To counteractthe aforementioned limitations, we utilized a custom-builtconstant current stimulator in which the stimulus intensity waselectronically controlled and adjusted, a computerized task that
Fig. 1. EMG activity during robotic assisted stepping. Raw EMG activity of soleus (SOL), medialis gastrocnemius (MG), tibialis anterior (TA), peroneus longus (PL), medialhamstrings (MH), vastus lateralis (VL), rectus femoris (RF), gracilis (GRC), and foot contact (as detected by the foot switch; FS) from both legs during stepping at 25% (A) and50% (B) BWS levels at 0.83 m s�1. Dashed lines in the left panels identify the approximate duration of the stance phase as detected by the foot switch. For each muscle at bothBWS levels, the normalized EMG average as a function of the step cycle is shown in the right panels. Heel strike corresponds to the zero normalized step cycle.
1398 M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404
Author's personal copy
cannot be done accurately with commercially available stimulators(Knikou, personal observations). Further, a supramaximal stimuluswas delivered to the posterior tibial nerve 100 ms after theH-reflex (Dyhre-Poulsen and Simonsen, 2002; Knikou et al.,2009). The customized LabView software measured the peak-to-peak amplitude of the un-rectified M-wave and Mmax duringassisted stepping and used a self-teaching algorithm to adjustthe stimulus intensity at each bin. This was based on the amplitudeof the M-wave as a percentage of the Mmax (and with respect tostimulation intensities evoking responses on the ascending limbof the recruitment curve) which was set to range between 4%and 9% of the Mmax (Knikou, 2008; Knikou et al., 2009). Althoughthe Mmax amplitude may vary across different phases of the stepcycle (Ferris et al., 2001; Simonsen and Dyhre-Poulsen, 1999), theMmax was elicited after the test H-reflex so to obtain moreaccurate and effective stimulations of the posterior tibial nerve(Simonsen and Dyhre-Poulsen, 2010).
Stimulation of the tibial nerve was triggered based on the signalfrom the ipsilateral foot switch (MA153, Motion Lab Systems Inc.,Baton Rouge, LA, USA). In all subjects, stimuli were deliveredrandomly across the step cycle, which was divided into 16 equalbins. Bin 1 corresponds to heel strike. Bins 8, 9, and 16 correspondapproximately to stance-to-swing transition, swing phaseinitiation, and swing-to-stance transition, respectively. H-reflexeswere recorded at 25% and 50% BWS levels.
2.4. Data analysis
All EMG signals from steps (average of 100 to 200 steps acrosssubjects) where the posterior tibial nerve was not stimulated werefull-wave rectified, high-pass filtered at 20 Hz, and low-pass fil-
tered at 500 Hz. Linear envelopes were obtained at 20 Hz low-passfilter after full wave rectification, and grouped across subjects sep-arately for stepping at 25% and 50% BWS. A pair-wise comparisonwas conducted for the ipsilateral and contralateral limb muscles toestablish statistically significant differences on the EMGs for step-ping at both BWS levels.
The background activity of the ipsilateral SOL and TA musclesfor each bin was estimated from the mean rectified and filteredEMG (high-pass filtered at 20 Hz, rectified, and low-pass filteredat 400 Hz) at 100 ms before posterior tibial nerve stimulation for60-ms duration. For each bin, subject, and BWS level, the mean sizeof the soleus H-reflex (% of Mmax) was plotted on the y-axisagainst the normalized SOL background activity on the x-axis,and a linear least-square regression was fitted to the data. Then,the slope of the relationship between soleus H-reflex and SOLbackground activity from the regression analysis which representsthe reflex gain (Capaday and Stein, 1986, 1987; Ferris et al., 2001)and the y-intercept were grouped based on the level of BWS for allsubjects. Linear regression slopes and y-intercepts at 25% and 50%BWS of assisted stepping were compared with a paired t-test.
The peak-to-peak amplitude of the H-reflex, M-wave, andMmax evoked at each bin of the step cycle were measured offline.The M-wave and H-reflex recorded at each bin were then ex-pressed as a percentage of the associated Mmax. H-reflexes wereaccepted only when the M-waves ranged from 4% to 9% of the cor-responding Mmax. Across subjects, from approximately 200–500elicited H-reflexes at each BWS level a total of 64–140 wereaccepted. The mean amplitude of the accepted normalized M-waveand H-reflex at each bin from each subject were grouped depend-ing on the BWS level and statistical significant differences wereestablished with a Wilcoxon rank sum test. In all statistical tests,
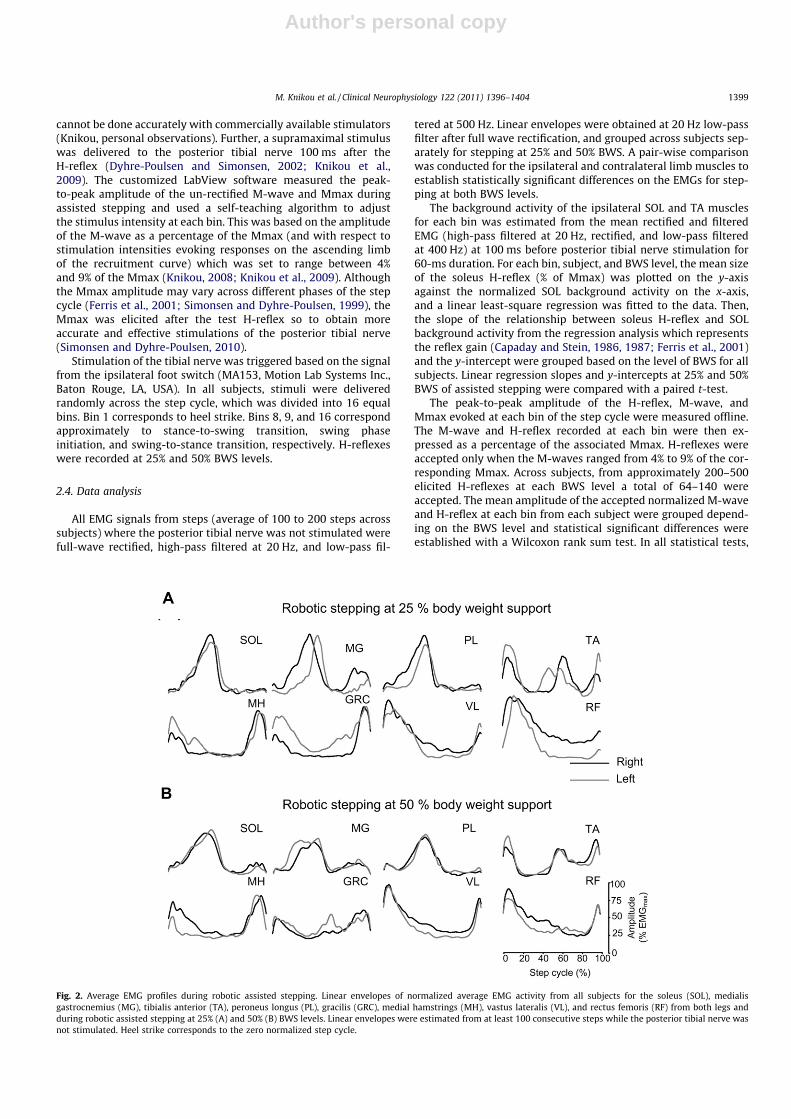
Fig. 2. Average EMG profiles during robotic assisted stepping. Linear envelopes of normalized average EMG activity from all subjects for the soleus (SOL), medialisgastrocnemius (MG), tibialis anterior (TA), peroneus longus (PL), gracilis (GRC), medial hamstrings (MH), vastus lateralis (VL), and rectus femoris (RF) from both legs andduring robotic assisted stepping at 25% (A) and 50% (B) BWS levels. Linear envelopes were estimated from at least 100 consecutive steps while the posterior tibial nerve wasnot stimulated. Heel strike corresponds to the zero normalized step cycle.
M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404 1399

Author's personal copy
statistically significant differences were established at P < 0.05. Re-sults are presented as mean values along with the standard error ofthe mean (SEM).
3. Results
3.1. EMG activation pattern during robotic assisted stepping
The treadmill speed was adjusted for each subject depending onhow comfortable they could step within the DGO system, and ran-ged from 0.61 to 0.83 (0.8 ± 0.07) m s�1 across subjects. For thesetreadmill speeds, the average step cycle duration was 1.46 ± 0.03and 1.42 ± 0.01 s at 25% and 50% of BWS, respectively. The meanduration of the stance and swing phases were 0.87 ± 0.02 and0.58 ± 0.03 s at 25% and 0.8 ± 0.02 and 0.62 ± 0.02 s at 50% BWS.No statistically significant differences of the aforementioned stepparameters were found between 25% and 50% BWS (P > 0.05).
Fig. 1 depicts raw EMGs from both legs (left panels) and linearEMG envelopes (right panels) for a representative subject duringstepping at 25% and 50% BWS at 0.83 m s�1. The linear EMG enve-
lopes are normalized (to the maximal locomotor associated EMG)averages of at least 100 consecutive steps, while no stimulationwas delivered, and plotted with respect to the normalized stepcycle with zero corresponding to heel strike. The main finding fromthese EMGs was that robotic assisted stepping significantlyaffected the EMG profile pattern as a function of the step cycle,with the average mean amplitude reduced at 50% BWS when com-pared to 25% BWS. Fig. 2A and B summarize the normalized linearEMG envelopes across subjects during robotic assisted stepping at25% and 50% BWS, respectively. The pooled average EMG profilewas found to be significantly different between 25% and 50%BWS for all but right MH muscle (P < 0.05). Further, a statisticallysignificant difference was found between the left and right EMGsfor both levels of body weight unloading, except for the SOL at25% BWS and all ankle muscles at 50% BWS.
3.2. Soleus H-reflex modulation pattern during robotic assistedstepping
In Fig. 3A and B, the average normalized amplitudes of the so-leus H-reflex and M-wave during assisted stepping at 25% and50% BWS are indicated. The soleus H-reflex modulation patternat constant M-wave amplitudes remained unchanged at both lev-els of BWS during robotic assisted stepping. The established soleusH-reflex amplitudes during assisted stepping were not affected bythe normalization factor (e.g. walking Mmax or standing Mmax;data not shown graphically), in line with previous reports(Simonsen and Dyhre-Poulsen, 2010). A Wilcoxon rank-sum t-testconducted at each bin separately showed that the soleus H-reflexwas statistically significantly different between 25% and 50%BWS only at bin 8 (P = 0.045). The amplitude of the ‘‘walking’’Mmax normalized to the Mmax recorded with subjects standingas a function of the step cycle is indicated in Fig. 3C.
The overall normalized amplitudes of the ipsilateral SOL and TAmuscles at 100 ms before posterior tibial nerve stimulation for atotal duration of 60 ms for all bins of the step cycle are illustratedin Fig. 4. These values are indicated for 25% BWS (Fig. 4A and B) and50% BWS (Fig. 4E and F). A t-test conducted for each bin separatelyshowed that the SOL background activity was not statistically sig-nificant different between 25% and 50% BWS (P > 0.05), while atbins 2 and 13, the TA background activity was enhanced at 25%BWS when compared to 50% BWS.
The overall average of the SOL/TA ratios for the ipsilateral leg forboth BWS level is indicated in Fig. 4C and G, respectively. The SOL/TA ratio was above 1 throughout the stance phase, suggesting thatassisted ankle movement by the foot lifters did not affect theantagonistic excitation pattern of the ankle muscles. Nonetheless,the SOL/TA ratio started to decrease before the end of late stanceat 50% BWS (see bins 7–8 in Fig. 4G). Lastly, the soleus H-reflexwas modulated in parallel with the SOL background activity(Fig. 4D and H). The soleus H-reflex gain was found not to vary sig-nificantly at 25% and 50% of BWS reaching overall amplitudes of79.06 ± 19.57 and 68.19 ± 13.1 at 25% and 50% BWS, respectively(paired t-test, P = 0.32). The y-intercept of the linear regression be-tween the H-reflex and the SOL background activity reached over-all amplitudes of �1.22 ± 3.07 and 1.29 ± 3.56 at 25% and 50% BWS,respectively, and were found not to be statistically significantly dif-ferent between BWS levels.
4. Discussion
The principal finding of this study is that the phase-dependentmodulation of the soleus H-reflex, commonly observed during overground walking or on a motorized treadmill at full body weightbearing, was preserved during robotic assisted stepping and was
Fig. 3. Soleus H-reflex modulation pattern during robotic assisted stepping. (A andB) The overall amplitude of the soleus H-reflex and the M-wave as a percentage ofthe ‘‘walking’’ maximal M-wave as a function of the step cycle during roboticassisted stepping at 25% and 50% BWS. (C) The ‘‘walking’’ Mmax normalized to the‘‘standing’’ Mmax as a function of the step cycle at 25% and 50% BWS. Bin 1corresponds to heel strike. Bins 8, 9, and 16 correspond approximately to stance-to-swing transition, swing phase initiation, and swing-to-stance transition, respec-tively. Error bars represent the SEM.
1400 M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404

Author's personal copy
similar to the pattern we have previously reported at equivalentBWS levels without robotic or manual leg guidance in healthy sub-jects (Knikou et al., 2009). Our current finding is in line with previ-ous reports in humans without BWS (Capaday and Stein, 1986;Crenna and Frigo, 1987; Dietz et al., 1990). Specifically, the soleus
H-reflex was enhanced progressively from mid to late stance, whilesoleus H-reflex depression started at swing phase initiation and re-mained during the swing phase regardless of the BWS level, whilethe H-reflex during the swing to stance transition (bin 16) reachedamplitudes similar to those observed at heel strike (bin 1) (Fig. 3A).
Fig. 4. The mean normalized soleus (SOL) and tibialis anterior (TA) EMG background activity as a function of the step cycle are indicated in A and B, while the overall ratio ofsoleus and tibialis anterior (SOL/TA) is indicated in C. The ratio equal to 1 indicates that the SOL muscle activity was equal to that of the TA. Values above or below 1 indicatecases of an increased or decreased SOL muscle activity against the antagonist TA muscle. The amplitude of the soleus H-reflex recorded at each bin from all subjects is plottedagainst the SOL EMG background activity. In this graph, the 16 points correspond to the 16 bins of the step cycle. The step cycle was broken into 16 equal bins, and each datapoint represents the average for a bin from all healthy subjects. Bin 1 corresponds to heel strike. Bins 8, 9, and 16 correspond approximately to stance-to-swing transition,swing phase initiation, and swing-to-stance transition, respectively. Error bars denote the SEM.
M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404 1401

Author's personal copy
Possible neural mechanisms accounting for soleus H-reflexdepression during the swing phase include reciprocal inhibitionfrom the corresponding antagonist muscles, recurrent inhibition,hyperpolarization of soleus a motoneurones, and a tonic increasein presynaptic inhibition of group Ia afferent terminals projectingmonosynaptically to soleus a motoneurones (Lavoie et al., 1997;Petersen et al., 1999; Pratt and Jordan, 1987; Capaday and Stein,1987; Gossard et al., 1990; Faist et al., 1996; Gosgnach et al.,1998; see Fig. 1 in Knikou, 2010a). Cyclic modulation of presynap-tic inhibition acting on Ia afferent terminals may partially accountfor the soleus H-reflex modulation in stance as well as swingphases (Faist et al., 1996; Edamura et al., 1991). This is supportedby more direct results obtained during fictive locomotion in adultcats, and specifically by the fact that primary afferents weredepolarized more during the swing phase and less during thestance phase (Gossard et al., 1991), primary afferent depolarizationdecreased group Ia afferent transmission by presynaptic inhibition(Gossard, 1996), and that presynaptic inhibition of monosynaptictransmission induced by muscle afferents was modulated in aphase-dependent manner (Menard et al., 2003).
The antagonistic TA muscle was active in a reciprocal pattern tothat observed in the SOL muscle (Fig. 2). Further, the SOL/TA ratiowas equal to, or less than one at bins 9 and 10 during 25% and 50%BWS, respectively (Fig. 4C and G). The SOL/TA relationship may bedriven by cortical neural circuits on the basis that Ia inhibitoryinterneurones are prone to cortical control (Nielsen et al., 1995;Petersen et al., 1998). On the basis that the soleus H-reflex depres-sion cannot be reversed with SOL voluntary muscle contractionduring the swing phase (Yang and Whelan, 1993), the soleusH-reflex gain was found not to vary as a function of BWS level, inline with previous reports in humans (Ferris et al., 2001; Knikouet al., 2009), background excitability of soleus motoneurones andactivation of antagonists may be considered as minor contributorsto the phasic reflex excitability.
The phasic activity of the soleus motoneurones during assistedstepping may be driven by movement-related afferent feedback orit may reflect embedded synaptic events or intrinsic properties ofthe a-motoneurones tightly coupled with the motor task. The lat-ter constitutes a rhetorical approach given the lack of or little evi-dence (see Rothwell et al., 1982), while there is ample evidence infavor of the former. We have recently reported that plantar cutane-ous afferents normalized the modulation pattern of the soleus H-reflex and TA flexion reflex in people with a chronic spinal cord in-jury at varying levels of BWS during manually assisted stepping(Knikou, 2010b). Because plantar cutaneous afferents subtract pre-synaptic inhibition of group I afferents during fictive locomotion(Menard et al., 2002), these afferents are good candidates for gatingH-reflex excitability in the current experiments. Further, plantarcutaneous afferents interact with hip proprioceptors to adjust so-leus H-reflex excitability, while imposed rhythmic hip angle oscil-lations, that change the length of hip flexors and extensors,modulate the soleus H-reflex in a phase-dependent manner (Kni-kou and Rymer, 2002a,b). This phenomenon has been reportedduring active but not passive bilateral pedalling with the H-reflexto be facilitated during the downstroke and depressed during theupstroke pedal cycles, respectively (Boorman et al., 1992). Duringpassive movements of the limbs resembling that of walking (ro-botic air stepping; 100% BWS), the soleus H-reflex amplitude wasdecreased during the flexion phase when compared to the reflexesrecorded during the extension phase and was similar to that re-corded at 40% BWS (Kamibayashi et al., 2010). The overall H-reflexmodulation pattern was, however, different from what we ob-served here (compare Fig. 3A with Fig. 4. in Kamibayashi et al.,2010), with absent progressively reflex facilitation from mid to latestance phase, and pronounced reflex depression during the swingphase. Differences between the observed soleus H-reflex modula-
tion during robotic air stepping and our findings can be attributedto differences in the methodology employed.
Based on the aforementioned findings, phasic changes of H-re-flex excitability in absence of upright posture and weight bearingas well as under conditions of reduced body loading during as-sisted stepping may be affected or regulated by sensory inputsfrom receptors responding to stretch. At this point, it should benoted that in the current study, unloading was conducted by bodyweight removal and not by removal of actual force loads acting onthe leg joints. In this respect, sudden removal of ground in intactcats did not modify the ankle extensor EMG activity that occurs70 ms before foot contact (Gorassini et al., 1994), suggesting thatthe extensor EMG activation pattern for this part of the step cycleis centrally generated, while flexor responses to loss of ground sup-port were delayed in spinalized animals (Hiebert et al., 1994), sug-gesting of a supraspinal control during the swing phase.
4.1. Muscle activity during robotic assisted stepping
In the decerebrate walking cat, 35% of the ankle extensors force(related to ankle joint stiffness) produced during stance has beenattributed to reflexes due to muscle length changes (Stein et al.,2000), similar to the range (30–60% in the soleus muscle at earlystance phase) reported in humans (Sinkjær et al., 1996). Decreasedefferent motor output has been described during reduced limbloading by imposed ankle extension, body immersion in water,air stepping, and bodyweight unloading during treadmill walkingin humans (Sinkjær et al., 2000; Dietz and Colombo, 1996; Dietzet al., 2002; Finch et al., 1991), with EMG amplitude dependingequally on the speed of walking (Hof et al., 2002; Yang and Winter,1985). In this study, the EMG profiles were significantly different at50% BWS when compared to 25% BWS. It has been reported thathigher BWS levels (50–75%) are related to abnormal EMG and kine-matic patterns during stepping (Finch et al., 1991; van Hedel et al.,2006; Threlkeld et al., 2003). The decreased EMG activity and espe-cially that of the TA, might be related to the foot lifters placed onthe subjects forefeet (which assisted ankle dorsiflexion for toeclearance during swing) and to the slow walking speeds (0.61–0.83 m s�1) utilized in this study. A previous study also attributeddecreased EMG activity in the MG and TA muscles to the roboticexoskeleton foot-lifters (Hidler and Wall, 2005). Further, therewas a clear lack of knee extensor activation during the pre- andearly swing phases (Murray et al., 1984), because knee extensoractivation might have been compensated for and reduced by therobotic exoskeleton system.
Slow treadmill speeds increase the duration of the step cycle,mostly by prolonging the duration of the stance phase (Finchet al., 1991). However, opposite results have also been reported,where treadmill walking at a constant speed of 1.25 m s�1 at 50%and 70% BWS reduced the stance phase and double limb support(Threlkeld et al., 2003). In this study, the increased duration ofstance and swing phases (compared to normal walking) as wellas step cycle duration may be attributed to the slow walking speedas well as to the intervention utilized in this study. Along with anincreased BWS level, an increased stride frequency has also beenshown to be related to decreased overall mean EMG activity, whileutilizing a robotic exoskeleton (Klarner et al., 2010).
4.2. Conclusion
Walking in humans is accomplished through complex interac-tions of neural circuits spanning different levels of the central ner-vous system, though the task appears rather simple at first glance.Robotic devices for recovery of walking in neurological disordersare utilized in rehabilitation centers throughout the world. Currentfindings indicate that spinal interneuronal circuits involved in the
1402 M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404

Author's personal copy
soleus H-reflex phase-dependent modulation pattern are engagedin a physiological manner during robotic assisted stepping, andthat sensory afferent feedback related to body loading, decreasedEMG amplitude of ankle extensors, and reduced ankle movementare not critical for manifestation of this reflex behavior.
Acknowledgements
We are grateful to all subjects for their voluntary participationand thank Drs. W. Zev Rymer, Yasin Y. Dhaher, and Erik B. Simon-sen for their help and support. This work was supported by a grantfrom the New York State Spinal Cord Injury Research Trust Fund,Department of Health (NYSDOH, Wadsworth Center) ContractC023690 and The Craig H. Neilsen Foundation, and was conductedat the Rehabilitation Institute of Chicago.
References
Angel MJ, Guertin P, Jimenez I, McCrea DA. Group I afferents evoke disynaptic EPSPsin cat hindlimb extensor motoneurones during fictive locomotion. J PhysiolLond 1996;494:851–61.
Barbeau H, Rossignol S. Recovery of locomotion after chronic spinalization in theadult cat. Brain Res 1987;412:84–92.
Barbeau H, Wainberg M, Finch L. Description and application of a system forlocomotor rehabilitation. Med Biol Eng Comput 1987;25:341–4.
Belanger M, Drew T, Provencher J, Rossignol S. A comparison of treadmilllocomotion in adult cats before and after spinal transection. J Neurophysiol1996;76:471–91.
Boorman G, Becker WJ, Morrice BL, Lee RG. Modulation of the soleus H-reflex duringpedalling in normal humans and in patients with spinal spasticity. J NeurolNeurosurg Psychiatry 1992;55:1150–6.
Bouyer LJ, Rossignol S. Contribution of cutaneous inputs from the hindpaws to thecontrol of locomotion. II: Spinal cats. J Neurophysiol 2003;90:3640–53.
Brooke JD, Cheng J, Misiaszek JE, Lafferty K. Amplitude modulation of the soleus H-reflex in the human during active and passive stepping movements. JNeurophysiol 1995;73:102–11.
Capaday C, Stein RB. Amplitude modulation of the soleus H-reflex in the humanduring walking and standing. J Neurosci 1986;6:1308–13.
Capaday C, Stein RB. Difference in the amplitude of the human soleus H-reflexduring walking and running. J Physiol Lond 1987;392:513–22.
Colombo G, Joerg M, Schreier R, Dietz V. Treadmill training of paraplegic patientsusing a robotic orthosis. J Rehabil Res Dev 2000;37:693–700.
Conway BA, Hultborn H, Kiehn O. Proprioceptive input resets central locomotorrhythm in the spinal cat. Exp Brain Res 1987;68:643–56.
Crenna P, Frigo C. Excitability of the soleus H-reflex arc during walking and steppingin man. Exp Brain Res 1987;66:49–60.
Dietz V, Colombo G. Effects of body immersion on postural adjustments tovoluntary arm movements in humans: role of load receptor input. J PhysiolLond 1996;497:849–56.
Dietz V, Duysens J. Significance of load receptor input during locomotion: a review.Gait Posture 2000;11:102–10.
Dietz V, Faist M, Pierrot-Deseilligny E. Amplitude modulation of the quadriceps H-reflex in the human during the early stance phase of gait. Exp Brain Res1990;79:221–4.
Dietz V, Muller R, Colombo G. Locomotor activity in spinal man: significance ofafferent input from joint and load receptors. Brain 2002;125:2626–34.
Dobkin BH, Apple D, Barbeau H, Basso M, Behrman A, Deforge D, Ditunno J, DudleyG, Elashoff R, Fugate L, Harkema S, Saulino M, Scott M. Methods for arandomized trial of weight-supported treadmill training versus conventionaltraining for walking during inpatient rehabilitation after incomplete traumaticspinal cord injury. Neurorehabil Neural Repair 2003;17:153–67.
Dyhre-Poulsen P, Simonsen EB. H reflexes recorded during locomotion. Adv ExpMed Biol 2002;508:377–83.
Edamura M, Yang JF, Stein RB. Factors that determine the magnitude and timecourse of human H-reflexes in locomotion. J Neurosci 1991;11:420–7.
Edgerton VR, Roy RR, de Leon RD, Tillakaratne N, Hodgson JA. Does motor learningoccur in the spinal cord? The Neuroscientist 1997;3:287–94.
Faist M, Dietz V, Pierrot-Deseilligny E. Modulation, probably presynaptic in origin,of monosynaptic Ia excitation during human gait. Exp Brain Res1996;109:441–9.
Faist M, Hoefer C, Hodapp M, Dietz V, Berger W, Duysens J. In humans Ib facilitationdepends on locomotion while suppression of Ib inhibition requires loading.Brain Res 2006;1076:87–92.
Ferris DP, Aagaard P, Simonsen EB, Farley CT, Dyhre-Poulsen P. Soleus H-reflex gainin humans walking and running under simulated reduced gravity. J PhysiolLond 2001;530:167–80.
Finch L, Barbeau H, Arsenault B. Influence of body weight support on normal humangait: development of a gait retraining strategy. Physical Therapy 1991;71:842–55.
Fouad K, Bastiaanse CM, Dietz V. Reflex adaptations during treadmill walking withincreased body load. Exp Brain Res 2001;137:133–40.
Gerilovsky L, Tsventinov P, Trenkova G. Peripheral effects on the amplitude ofmonopolar and bipolar H-reflex potentials from the soleus muscle. Exp BrainRes 1989;76:173–81.
Gorassini MA, Prochazka A, Hiebert GW, Gauthier MJA. Corrective responses to lossof ground support during walking. I. Intact cats. J Neurophysiol 1994;71:603–10.
Gosgnach S, Quevedo J, Fedirchuk B, McCrea D. Tonic presynaptic reduction ofmonosynaptic Ia EPSPs during fictive locomotion. Ann NY Acad Sci1998;860:505–7.
Gossard JP, Cabelguen JM, Rossignol S. Phase-dependent modulation of primaryafferent depolarization in single cutaneous primary afferents evoked byperipheral stimulation during fictive locomotion in the cat. Brain Res1990;537:14–23.
Gossard JP, Cabelguen JM, Rossignol S. An intracellular study of muscle primaryafferents during fictive locomotion in the cat. J Neurophysiol 1991;65:914–26.
Gossard JP. Control of transmission in muscle group IA afferents during fictivelocomotion in the cat. J Neurophysiol 1996;76:4104–12.
Grillner S, Rossignol S. On the initiation of the swing phase of locomotion in chronicspinal cats. Brain Res 1978;146:269–77.
Grillner S. Biological pattern generation: the cellular and computational logic ofnetworks in motion. Neuron 2006;52:751–66.
Guertin P, Angel MJ, Perreault MC, McCrea DA. Ankle extensor group I afferentsexcite extensors throughout the hindlimb during fictive locomotion in the cat. JPhysiol Lond 1995;487:197–209.
Hidler JM, Wall AE. Alterations in muscle activation patterns during robotic-assistedwalking. Clin Biomech 2005;20:184–93.
Hiebert GW, Gorassini MA, Jiang W, Prochazka A, Pearson KG. Corrective responsesto loss of ground support during walking. II: Comparison of intact and chronicspinal cats. J Neurophysiol 1994;71:611–22.
Hof AL, Elzinga H, Grimmius W, Halbertsma JP. Speed dependence of averaged EMGprofiles in walking. Gait Posture 2002;16:78–86.
Jezernik S, Colombo G, Keller T, Frueh H, Morani M. Robotic orthosis Lokomat: arehabilitation and research tool. Neuromodulation 2003;6:108–15.
Kamibayashi K, Nakajima T, Fujita M, Takahashi M, Ogawa T, Akai M, Nakazawa K.Effect of sensory inputs on the soleus H-reflex amplitude during robotic passivestepping in humans. Exp Brain Res 2010;202:385–95.
Klarner T, Chan HK, Wakeling JM, Lam T. Patterns of muscle coordination vary withstride frequency during weight assisted treadmill walking. Gait Posture2010;31:360–5.
Knikou M. The H-reflex as a probe: pathways and pitfalls. J Neurosci Methods2008;171:1–12.
Knikou M. Neural control of locomotion and training-induced plasticity after spinaland cerebral lesions. Clin Neurophysiol 2010a;121:1655–68.
Knikou M. Plantar cutaneous afferents normalize the reflex modulation patternsduring stepping in chronic human spinal cord injury. J Neurophysiol2010b;103:1304–14.
Knikou M. Soleus H-reflex phase-dependent modulation during one-legged footreaching and withdrawal in standing humans. Neurosci. Lett. 2011;487:305–9.
Knikou M, Angeli CA, Ferreira CK, Harkema SJ. Soleus H-reflex modulation duringbody weight support treadmill walking in spinal cord intact and injuredsubjects. Exp Brain Res 2009;193:397–407.
Knikou M, Rymer WZ. Effects of changes in hip joint angle on H-reflex excitability inhumans [Erratum in Exp Brain Res 2002;144:558]. Exp Brain Res 2002a;143:149–59.
Knikou M, Rymer WZ. Hip angle induced modulation of H reflex amplitude, latencyand duration in spinal cord injured humans. Clin Neurophysiol 2002b;113:1698–708.
Larsen B, Mrachacz-Kersting N, Lavoie BA, Voigt M. The amplitude modulation ofthe quadriceps H-reflex in relation to the knee joint action during walking. ExpBrain Res 2006;170:555–66.
Lavoie BA, Devanne H, Capaday C. Differential control of reciprocal inhibition duringwalking versus postural and voluntary motor tasks in humans. J Neurophysiol1997;78:429–38.
Menard A, Leblond H, Gossard JP. Sensory integration in presynaptic inhibitorypathways during fictive locomotion in the cat. J Neurophysiol 2002;88:163–71.
Menard A, Leblond H, Gossard JP. Modulation of monosynaptic transmission bypresynaptic inhibition during fictive locomotion in the cat. Brain Res2003;964:67–82.
Mrachacz-Kersting N, Lavoie BA, Andersen JB, Sinkjær T. Characterisation of thequadriceps stretch reflex during the transition from swing to stance phase ofhuman walking. Exp Brain Res 2004;159:108–22.
Murray MP, Mollinger LA, Gardner GM, Sepic SB. Kinematic and EMG patternsduring slow, free, and fast walking. J Orthop Res 1984;2:272–80.
Nielsen J, Petersen N, Ballegaard M. Latency of effects evoked by electrical andmagnetic brain stimulation in lower limb motoneurones in man. J Physiol Lond1995;484:791–802.
Pearson KG, Ramirez JM, Jiang W. Entrainment of locomotor rhythm by group Ibafferents from ankle extensor muscles in spinal cats. Exp Brain Res1992;90:557–66.
Petersen N, Christensen LOD, Sinkjær T, Morita H, Nielsen J. Evidence suggesting atranscortical pathway from muscle afferents to tibialis anterior motoneuronesin man. J Physiol Lond 1998;512:267–76.
M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404 1403

Author's personal copy
Petersen N, Morita H, Nielsen J. Modulation of reciprocal inhibition betweenankle extensors and flexors during walking in man. J Physiol Lond 1999;520:605–19.
Pratt CA, Jordan LM. Ia inhibitory interneurons and Renshaw cells as contributorsto the spinal mechanisms of fictive locomotion. J Neurophysiol 1987;57:56–71.
Rossignol S, Drew T, Brustein E, Jiang W. Locomotor performance and adaptationafter partial or complete spinal cord lesions in the cat. Prog Brain Res1999;123:349–65.
Rossignol S, Dubuc R, Gossard JP. Dynamic sensorimotor interactions in locomotion.Physiol Rev 2006;86:89–154.
Rothwell JC, Traub MM, Day BL, Obeso JA, Thomas PK, Marsden CD. Manual motorperformance in a deafferented man. Brain 1982;105:515–42.
Simonsen EB, Dyhre-Poulsen P. Amplitude modulation of the human soleus H-reflexduring walking and running. J Physiol Lond 1999;513:929–39.
Simonsen EB, Dyhre-Poulsen P. Test–retest reliability of the soleus H-reflexexcitability measured during human walking. Hum Mov Sci 2010.doi:10.1016/j.humov.2010.02.009.
Sinkjær T, Andersen JB, Ladouceur M, Christensen LO, Nielsen JB. Major role forsensory feedback in soleus EMG activity in the stance phase of walking in man. JPhysiol Lond 2000;523:817–27.
Sinkjær T, Andersen JB, Larsen B. Soleus stretch reflex modulation during gait inhumans. J Neurophysiol 1996;76:1112–20.
Smith CA, Mummidisetty CK, Hajela N, Xiao M, Knikou M. On the behavior of humanspinal reflex circuits during robotic assisted stepping. APTA Annual Conference,National Harbor, MD, USA, 2011.
Stein RB, Misiaszek JE, Pearson KG. Functional role of muscle reflexes for forcegeneration in the decerebrate walking cat. J Physiol Lond 2000;525:781–91.
Threlkeld AJ, Coooper LD, Monger BP, Craven AN, Haupt HG. Temporospatial andkinematic gait alterations during treadmill walking with body weightsuspension. Gait Posture 2003;17:235–45.
van Hedel HJ, Tomatis L, Muller R. Modulation of leg muscle activity and gaitkinematics by walking speed and bodyweight unloading. Gait Posture2006;24:35–45.
Whelan PJ, Hiebert GW, Pearson KG. Stimulation of the group I extensorafferents prolongs the stance phase in walking cats. Exp Brain Res1995;103:20–30.
Yang JF, Whelan PJ. Neural mechanisms that contribute to cyclical modulation ofthe soleus H-reflex in walking in humans. Exp Brain Res 1993;95:547–56.
Yang JF, Winter DA. Surface EMG profiles during different walking cadences inhumans. Electroencephalog Clin Neurophysiol 1985;60:485–91.
Zehr EP, Stein RB. What functions do reflexes serve during human locomotion? ProgNeurobiol 1999;58:185–205.
1404 M. Knikou et al. / Clinical Neurophysiology 122 (2011) 1396–1404




















