
Smooth Muscle Cell-Specific Knockout of Androgen Receptor: A New Model for Prostatic Disease
11
Smooth Muscle Cell-Specific Knockout of Androgen Receptor: A New Model for Prostatic Disease Michelle Welsh, Lindsey Moffat, Alan McNeilly, David Brownstein, Philippa T. K. Saunders, Richard M. Sharpe, and Lee B. Smith Medical Research Council Centre for Reproductive Health (M.W., L.M., A.M., P.T.K.S., R.M.S., L.B.S.), and University of Edinburgh (D.B.), The Queen’s Medical Research Institute, Edinburgh, EH16 4TJ, Scotland, United Kingdom Androgen-driven stromal-epithelial interactions play a key role in normal prostate development and function as well as in the progression of common prostatic diseases such as benign prostatic hyperplasia and prostate cancer. However, exactly how, and via which cell type, androgens mediate their effects in the adult prostate remains unclear. This study investigated the role for smooth muscle (SM) androgen signaling in normal adult prostate homeostasis and function using mice in which androgen receptor was selectively ablated from prostatic SM cells. In adulthood the knock- out (KO) mice displayed a 44% reduction in prostate weight and exhibited histological abnormal- ities such as hyperplasia, inflammation, fibrosis, and reduced expression of epithelial, SM, and stem cell identify markers (e.g. p63 reduced by 27% and Pten by 31%). These changes emerged beyond puberty and were not explained by changes in serum hormones. Furthermore, in response to exogenous estradiol, adult KO mice displayed an 8.5-fold greater increase in prostate weight than controls and developed urinary retention. KO mice also demonstrated a reduced response to castration compared with controls. Together these results demonstrate that prostate SM cells are vital in mediating androgen-driven stromal-epithelial interactions in adult mouse prostates, de- termining cell identity and function and limiting hormone-dependent epithelial cell proliferation. This novel mouse model provides new insight into the possible role for SM androgen action in prostate disease. (Endocrinology 152: 3541–3551, 2011) T he prostate, like much of the male reproductive tract, depends on androgen-driven stromal-epithelial inter- actions for normal structure and function (1–5) but the underlying mechanisms remain unclear. Classical exper- iments by Cunha and and Donjacour (5) and Chung and Cunha (6) have highlighted the direct role for androgen action via the stromal cells in inducing normal prostate development. However, much of the evidence for this comes from tissue recombination studies using neonatal stroma and epithelium; therefore, understanding of the role for stromal androgen action in the normal adult pros- tate remains limited. Perturbation of these androgen- driven stromal-epithelial interactions is also implicated in prostatic diseases such as benign prostatic hyperplasia and prostate cancer (PCa) (7, 8); therefore, greater under- standing of the role for stromal AR in normal adult pros- tate homeostasis could uncover new insight into how an- drogens mediate prostate disease and so provide new opportunities for intervention. Although this is challeng- ing to address directly in humans, the advent of genetic modification in mice provides an exciting tool with which to advance our fundamental understanding of prostate development and function. The mouse prostate consist of ducts and glands lined by a secretory epithelial layer that lies above a layer of p63- positive basal cells (9), which have been proposed to con- tain one of the two stem cell populations identified in the mouse prostate (10 –13). Prostatic ducts are surrounded ISSN Print 0013-7227 ISSN Online 1945-7170 Printed in U.S.A. Copyright © 2011 by The Endocrine Society doi: 10.1210/en.2011-0282 Received March 14, 2011. Accepted June 17, 2011. First Published Online July 5, 2011 Abbreviations: AP, Anterior prostate; AR, androgen receptor; DLP, dorsolateral prostate; E2, estradiol; GFP, green fluorescent protein; KO, knockout; MH, myosin-heavy chain promoter; PCa, prostate cancer; PTM-ARKO, SM cell AR knockout; SM, smooth muscle; SMA, smooth muscle actin; SV, seminal vesicles; T, testosterone; VP, ventral prostate. REPRODUCTION-DEVELOPMENT Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3541 The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.
Transcript of Smooth Muscle Cell-Specific Knockout of Androgen Receptor: A New Model for Prostatic Disease

Smooth Muscle Cell-Specific Knockout of AndrogenReceptor: A New Model for Prostatic Disease
Michelle Welsh, Lindsey Moffat, Alan McNeilly, David Brownstein,Philippa T. K. Saunders, Richard M. Sharpe, and Lee B. Smith
Medical Research Council Centre for Reproductive Health (M.W., L.M., A.M., P.T.K.S., R.M.S., L.B.S.),and University of Edinburgh (D.B.), The Queen’s Medical Research Institute, Edinburgh, EH16 4TJ,Scotland, United Kingdom
Androgen-driven stromal-epithelial interactions play a key role in normal prostate developmentand function as well as in the progression of common prostatic diseases such as benign prostatichyperplasia and prostate cancer. However, exactly how, and via which cell type, androgens mediatetheir effects in the adult prostate remains unclear. This study investigated the role for smoothmuscle (SM) androgen signaling in normal adult prostate homeostasis and function using mice inwhich androgen receptor was selectively ablated from prostatic SM cells. In adulthood the knock-out (KO) mice displayed a 44% reduction in prostate weight and exhibited histological abnormal-ities such as hyperplasia, inflammation, fibrosis, and reduced expression of epithelial, SM, and stemcell identify markers (e.g. p63 reduced by 27% and Pten by 31%). These changes emerged beyondpuberty and were not explained by changes in serum hormones. Furthermore, in response toexogenous estradiol, adult KO mice displayed an 8.5-fold greater increase in prostate weight thancontrols and developed urinary retention. KO mice also demonstrated a reduced response tocastration compared with controls. Together these results demonstrate that prostate SM cells arevital in mediating androgen-driven stromal-epithelial interactions in adult mouse prostates, de-termining cell identity and function and limiting hormone-dependent epithelial cell proliferation.This novel mouse model provides new insight into the possible role for SM androgen action inprostate disease. (Endocrinology 152: 3541–3551, 2011)
The prostate, like much of the male reproductive tract,depends on androgen-driven stromal-epithelial inter-
actions for normal structure and function (1–5) but theunderlying mechanisms remain unclear. Classical exper-iments by Cunha and and Donjacour (5) and Chung andCunha (6) have highlighted the direct role for androgenaction via the stromal cells in inducing normal prostatedevelopment. However, much of the evidence for thiscomes from tissue recombination studies using neonatalstroma and epithelium; therefore, understanding of therole for stromal androgen action in the normal adult pros-tate remains limited. Perturbation of these androgen-driven stromal-epithelial interactions is also implicated inprostatic diseases such as benign prostatic hyperplasia and
prostate cancer (PCa) (7, 8); therefore, greater under-standing of the role for stromal AR in normal adult pros-tate homeostasis could uncover new insight into how an-drogens mediate prostate disease and so provide newopportunities for intervention. Although this is challeng-ing to address directly in humans, the advent of geneticmodification in mice provides an exciting tool with whichto advance our fundamental understanding of prostatedevelopment and function.
The mouse prostate consist of ducts and glands lined bya secretory epithelial layer that lies above a layer of p63-positive basal cells (9), which have been proposed to con-tain one of the two stem cell populations identified in themouse prostate (10–13). Prostatic ducts are surrounded
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2011 by The Endocrine Societydoi: 10.1210/en.2011-0282 Received March 14, 2011. Accepted June 17, 2011.First Published Online July 5, 2011
Abbreviations: AP, Anterior prostate; AR, androgen receptor; DLP, dorsolateral prostate;E2, estradiol; GFP, green fluorescent protein; KO, knockout; MH, myosin-heavy chainpromoter; PCa, prostate cancer; PTM-ARKO, SM cell AR knockout; SM, smooth muscle;SMA, smooth muscle actin; SV, seminal vesicles; T, testosterone; VP, ventral prostate.
R E P R O D U C T I O N - D E V E L O P M E N T
Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3541
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

by a thin smooth muscle (SM) layer and an outer hetero-geneous stromal layer. From puberty, under the control ofandrogens, luminal epithelial cells secrete several well-characterized products that contribute to the seminalfluid, such as serine peptidase inhibitor Kazal type 3(Spink3) and prostate � defensin 1 (Pbd1/Defb37) (14).Androgens modulate gene transcription by binding to theandrogen receptor (AR), which is expressed in prostaticstromal and epithelial cells, but not in the basal stem cells(15, 16). Adult prostates also express both estrogen re-ceptors (ER) � and � (17), and disruption of the androgen-estrogen balance in mice leads to abnormal prostate his-tology and carcinoma (18). Interestingly, androgens arethought to mediate different effects in each cell compart-ment; for example, prostatic epithelial AR is essential forthe epithelial secretory function observed in the prostate ofmature mice (19) and inhibits proliferation (20) whereasstroma AR is thought to determine the fate of the epithe-lium, regulate epithelial cell apoptosis, and promote pro-liferation (21, 22). However, because prostatic stroma iscomposed of many cell types including immune cells andfibroblasts, exactly which cell types mediate these effectsis unclear. For example, it is unknown whether the epi-thelium relies on interactions with the entire stromal com-partment, or whether signaling from a subset of thestroma, such as the SM cells or the stromal fibroblasts, issufficient to mediate normal epithelial cell identity andfunction. Recent generation of a SM cell AR knockout(KO) mouse model [termed PTM-ARKO (23)] has pro-vided an exciting opportunity to investigate the role forSM-specific androgen action in normal adult prostatefunction as well as its possible role in prostate disease.
Materials and Methods
Breeding of transgenic miceMice in which AR has been ablated from the SM cells of the
testis and seminal vesicles (SV) have been described by us pre-viously (23, 24), using male mice expressing Cre recombinaseand green fluorescent protein (GFP), both under the control of aSM myosin-heavy chain promoter (MH) (25). These mice alsoexpressed Cre recombinase in the SM cells of the prostate, there-fore providing a novel opportunity to investigate the role forandrogen action in the prostate SM. The Cre-positive (ARflox
positive) male offspring are termed PTM-ARKO or KO, whereasthe ARflox positive Cre negative male littermates were used ascontrols (termed control), as previously reported (23). All micewere bred under standard conditions of care and use under li-censed approval from the United Kingdom Home Office. Studmale MH mice and PTM-ARKO male offspring were genotypedfor the presence of Cre using PCR as previously described (23).
Recovery of reproductive tissuesMale mice were culled at embryonic day (e)17.5 and at post-
natal days (d) 35, 50, 100, and 200, and prostates were recov-ered, weighed, and processed as previously reported (23). Eachprostate lobe [ventral (VP), anterior (AP), and dorsolateral(DLP)] was isolated and weighed separately. The Bar Harborconsensus report (26) states that no mouse prostate lobe is morerelevant to human prostate disease then another; therefore theVP was examined in further detail as a representative lobe. VPwere stained with hematoxylin and eosin and examined for his-tological abnormalities such as hyperplasia, edema, and hyper-trophy, by an expert mouse pathologist.
Hormone implant studyControl (n � 5) and PTM-ARKO (n � 5) male mice (d130–
d150) were implanted with a 1-cm SILASTIC implant (DowCorning Corp., Midland, MI) containing 100 mg testosterone(T): 10 mg estradiol (E2) (Sigma-Aldrich, Poole, UK), as previ-ously reported in PTM-ARKO (24); this treatment regime wasoriginally reported by Ricke et al. (27). Empty sham implantswere used as a treatment control. Implants were left in situ for 13wk after which mice were culled, blood was collected, and VPwere recovered.
Castration studiesBoth testes were surgically removed from adult control (n �
9) and PTM-ARKO (n � 4) male mice (d 130–d 150) underisofluorane. After 7 d, mice were culled and VP were recovered.
Immunohistochemical analysisMice were examined by immunohistochemistry for the pres-
ence of Cre in VP SM cells, identified by immunoexpression of�-smooth muscle actin (SMA), using a previously publishedmethod (23, 24). Fluorescent immunostained sections weremounted in Mowiol mounting medium (Calbiochem, San Diego,CA) and fluorescent images were captured using a Zeiss LSM510 Meta Axiovert 100M confocal microscope (Carl Zeiss Ltd.,Welwyn, UK). Adult VP from PTM-ARKO and control maleswere also stained for green fluorescent protein as a marker forCre expression, e-cadherin, SMA, pan-cytokeratin, phosphohi-stone H3, Sox9, and p63. Sections were deparaffinized and re-hydrated and antigen was retrieved as previously detailed (24)before using a specific polymer high-contrast program on aBond-X automated immunostaining machine (Vision Biosys-tems, Newcastle, UK). Optimal antibody concentrations are de-tailed in supporting information Table 1. All kits were purchasedfrom Vision Biosystems, and images were captured using a Provismicroscope (Olympus Optical Co., Tokyo, Japan) equippedwith a Kodak DCS330 camera (Eastman Kodak, Rochester,NY). To ensure reproducibility of results, representative VP fromat least three animals at each age were used, and sections fromPTM-ARKO and control littermates were processed in parallelon the same slide on at least two occasions. Appropriate negativecontrols were included to ensure that any staining observed wasspecific. All antibodies used showed only minor nonspecificstaining.
Quantification of VP cell compositionThe proportion of VP composed of epithelium, SM, stroma,
and lumen was quantified stereologically in d 100 control and
3542 Welsh et al. Smooth Muscle AR in Prostate Disease Endocrinology, September 2011, 152(9):3541–3551
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

PTM-ARKO VP immunostained for SMA by grid point countingusing stereology equipment and sampling procedures previouslydescribed (23, 24). SM thickness and epithelial cell height werealso quantified in d 100 control and KO VP, as previously de-scribed (24).
Analysis of cell mitosis and apoptosis in VPVP from control and KO adults were immunostained for
phosphohistone H3 and positive or negative mitotic cells werecounted in the epithelial compartment, as previously described(24). The number of positive cells in the epithelial compartmentwas then divided by the total number of epithelial cells visible insection to calculate the epithelial proliferation index. Apoptoticcells were identified in d 12 and d 100 VP from PTM-ARKO andcontrol mice using terminal deoxynucleotide transferase-medi-ated dUTP nick end labeling, as previously described (28). Quan-tification was deemed unnecessary because few apoptotic cellswere identified at either age.
Determination of deletion of AR exon 2RNA was isolated and quantified from frozen VP, and cDNA
was prepared, as previously described (23). RT-PCR was per-formed as previously described (23) on cDNA synthesized fromVP from PTM-ARKO and controls. Testes from ARKO micewere used as a positive control for AR deletion because ARKOmice have no prostate.
Quantitative analysis of gene expressionQuantitative PCR was performed on d 100 PTM-ARKO and
control VP for the genes listed in Table 2, using previously de-scribed methodology (23). The expression of each gene was re-
lated to 18S, an internal positive control, and expression wascorrected for proportional changes in cell compartments.
Statistical analysisData were analyzed using GraphPad Prism version 5 (Graph
Pad Software Inc., San Diego, CA) using a one-tailed unpaired ttest, one-way ANOVA or two-way ANOVA, as appropriate,followed by Bonferroni post hoc tests. Values are expressed asmean � SEM. Normality was confirmed using KS normality test.
Results
Identification of SM AR deletion in VP fromPTM-ARKO mice
We have already described mice with AR ablation fromthe SM cells of the testis and SV (23, 24); these mice ex-press Cre recombinase and GFP under the control of a SMmyosin-heavy chain promoter (25). Within the prostate,GFP (Fig. 1A, arrow) (and therefore Cre) was expressedexclusively in the SM layer in KO mice at d 100. RT-PCRfor AR exon 2 revealed that control prostates only ex-pressed the full-length [wild-type (WT)] AR transcriptwhereas KO prostates expressed both the WT and thesmaller KO AR transcripts, demonstrating that AR wasdeleted from a proportion of prostate cells (Fig. 1B).ARKO testis cDNA, the positive control, only expressedthe smaller KO transcript because AR had been deleted
TABLE 1. Immunohistochemistry antibody details
Antibody Antibody source Dilution Detection system
SMA/ARSMA Sigma 1:1000 Tyramide 488AR Santa Cruz 1:50 Goat antirabbit Alexa 546
GFP Abcam 1:100 Bond automated polymer systemCytokeratin Sigma 1:1000 Bond automated polymer systemSMA Sigma 1:4000 Bond automated polymer systemE-cadherin BD Bioscience 1:400 Bond automated polymer systemSox9 Chemicon 1:4000 Bond automated polymer systemp63 Santa Cruz 1:1200 Bond automated polymer system
TABLE 2. Taqman primer details
Gene Forward primer Reverse primer
Cytokeratin 18 (CK18) agatgacaccaacatcacaagg cttccagaccttggacttcctSox9 cagcaagactctgggcaag tccacgaagggtctcttctcNkx3.1 cgactgaacccgagtctgat aatcacctgagtgtgagagaaggPten aggcacaagaggccctagat ctgactgggaattgtgactccCytokeratin 14 (CK14) atcgaggacctgaagagcaa tcgatctgcaggaggacattDesmin gccacctaccggaagctact gcagagaaggtctggataggaaSmooth muscle actin (SMA) actctcttccagccatctttca ataggtggtttcgtggatgcProbasin tcatcctcctgctcacactg gctaagtaaattgtttgccaaggSpermine binding protein (SBP) gcatgcggatcttcttaagtgt catcagtccagttactgccaaaDefensin �2 (Pros Def) tgctgcctccttttctcatatac tcaagttctgcttcgtatccaaSerine peptidase inhibitor Kazal type 3 (Spink3) cacccagatcttcgacaatg taaactcagcagggccaaagCyclooxygenase 1 (Cox-1) cctctttccaggagctcaca tcgatgtcaccgtacagctc
Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3543
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

from all cells (Fig. 1B). Immunohistochemistry demon-strated that AR was absent in most SM cells in KO pros-tates (identified by SM actin expression) (Fig. 1C). ARcontinued to be expressed in the prostate epithelial andnon-SM stromal cells in KO as in controls (Fig. 1C). De-letion of AR can be identified in the PTM-ARKO urogen-ital sinus from e 17.5 (24). Serum T and E2 concentrationsare normal at all ages, based on measurements in a total of93 mice measured in five different assays (23, 24). External
sexual development was normal in these males (23, 24),and their reproductive tracts formed normally.
Prostate weight, morphology, and histologyAP, DLP, and VP weights were all found to be reduced
at d 200 (Fig. 2A) in KO males. Because the Bar Harborconsensus report (26), states that no mouse prostate lobeis more relevant to human prostate disease then another,further detailed examination was focused upon the ventralprostate as a representative lobe. Further investigation re-vealed that VP weights were significantly reduced at d 50and d 100, but not at d 35, compared with controls (Fig.2B). Note that there was no significant reduction in VPweight in adult males expressing either MH Cre or ARflox
alone compared with MH Cre-negative, ARflox-negativemales (P � 1.0 and P � 0.73, respectively); this demon-strates that the phenotype observed in the KO animalsresults from AR ablation rather than from expression ofCre recombinase or ARflox per se. Histological abnormal-ities and atrophy were seen in the stromal and epithelialcells in adult KO VP, but not in controls (Fig. 2C andSupplemental Fig. 1A published on The Endocrine Soci-ety’s Journals web site at http://end.endojournals.org).The phenotype of individual acini varied within each KOVP, as in controls, but all KO VPs were similarly affected,displaying signs of elongated epithelial folding, epithe-lial cell hypertrophy and intracellular edema, and dif-fuse epithelial cell hyperplasia leading to overcrowdingand desquamation (d 100 in Figs. 2C and 3B and d 35in Supplemental Fig. 1A). This hyperplasia was con-firmed quantitatively at d 100 (Fig. 3A) but not at d 35(Supplemental Fig. 1B). Epithelial cell height was un-changed in d 100 KO VP compared with controls (Fig. 3B)but lumens were smaller in KO VP (d 100 in Fig. 2C andd 35 in Supplemental Fig. 1); this was confirmed quanti-tatively at d 100 (Fig. 3A) but not at d 35 (SupplementalFig. 1B). Stromal fibrosis was observed in d 100 KO VP,characterized by eosinophilic staining around fibroblasts(Fig. 2C). Diffuse stromal hyperplasia was evident in KOVP (d 100 in Fig. 2C and d 35 in Supplemental Fig. 1) andwas confirmed quantitatively at d 100 (Fig. 3A), but not atd 35 (Supplemental Fig. 1B); this was mostly composed offibroblasts and immune cells. However, the proportion ofSM in KO VP was unchanged compared with controls atd 100 (Fig. 3, A and C) and d 35 (Supplemental Fig. 1B).Considerable infiltration of neutrophils, leukocytes, andmonocytes was observed in the glands and stroma of mostKO VP at d 35 and d 100 (Supplemental Fig. 1, asterisk;and Fig. 2C). The epithelial mitotic index did not differsignificantly in KO VPs at d100 compared with controls(data not shown) and few apoptotic cells were identified inKO or control VP.
FIG. 1. Characterization of Cre recombinase expression in adult PTM-ARKO VP. A, GFP (expression of which is also driven by the smMH-promoter) was expressed in the SM cells, but not in the outer stromalcells, in d 100 KO VP (brown, arrow). GFP was not detected in any cellsin control VP. B, Deletion of AR in d 100 VP was determined using RT-PCR spanning exon 2. Only the larger WT band was seen in control VPwhereas the smaller KO band was seen in ARKO testis positive controlcDNA. Both bands were identified in KO VP, showing deletion of AR ina proportion of cells. C, Immunohistochemistry for SMA (green) andAR (red) showed that AR was not expressed in KO VP SM cells (arrow),but was expressed in epithelial (*) and non-SM stromal cells in KO andcontrol VP and in smooth muscle in control VP.
3544 Welsh et al. Smooth Muscle AR in Prostate Disease Endocrinology, September 2011, 152(9):3541–3551
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

Expression of VP cell identity markersE-cadherin was expressed in all luminal epithelial cells
in control VP at d 100 whereas in KO VP, E-cadherinexpression was absent from some acini (Fig. 4 A). Sox9was expressed in luminal and basal epithelial cells in d 100control VP but was absent from some luminal epithelialcells in d 100 KO VP (Fig. 4B). p63 was expressed in basalepithelial cells in control and KO VP at d 35 and d 100 (Fig.4, C and D), but significantly fewer p63 positive cells wereevident in KO VP at d 100 (Fig. 4, D and E) but not at d35 (Fig. 4C). Similarly, a significant reduction in relativemRNA expression of luminal epithelial cell markers cy-tokeratin 18 (CK18), Sox9, Nkx3.1, and Pten and thebasal epithelial cell marker cytokeratin 14 was found in d100 KO VP compared with controls (Fig. 4F). RelativemRNA expression of the SM cell genes desmin and SMA
was also significantly reduced in d 100 KO VP (Fig. 4F).Notably there was no change in expression of any of thesemarkers at d 35 (Supplemental Fig. 1C). Relative mRNAexpression of ER� or � was similar in d 100 KO andcontrol VP (data not shown).
Expression of genes encoding epithelial secretoryproteins
mRNA expression of Defensin �2 (Pros Def), serinepeptidase inhibitor Kazal type 3 (Spink3), and Cycloox-ygenase 1 (Cox-1) was significantly reduced in d 100 KOVP, compared with controls (Fig. 4F); conversely, Pro-basin and spermine-binding protein (SBP) mRNA ex-pression was unchanged (Fig. 4F). Expression of thesegenes was not altered in KO VP at d 35 (SupplementalFig. 1C).
FIG. 2. Gross morphology and histology of PTM-ARKO prostates. A, Quantification of prostate lobe weights at d 200. B, Quantification of VPweight in control and KO mice at d 35–d 200. Values are means � SEM (n � 3–14 mice). *, P � 0.05; **, P � 0.01, compared with controls. C,Hematoxylin and eosin (H&E) staining of d 100 KO and control VP. Histological abnormalities can be seen in KO VP with epithelial cells (E)detaching from the basement membrane (*) and sloughing off into the lumens (L). Hyperplasia of the stroma (S) is also obvious (arrow).
Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3545
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.
Effect of treatment withexogenous T and E2
To assess the prostate response to es-trogens without causing involution, wetreated adult mice with exogenous Tplus estradiol (T�E2) for 13 wk, as pre-viously described (24). All comparisonswere made to age-matched control andKO males treated with ‘empty’ shamimplants for 13 wk. T�E2 treatmentresulted in normal serum T concentra-tions and 400-fold increased serumE2 concentrations, compared with un-treated mice (24). VP recovered fromT�E2-treated controls and KO dis-played a significant increase in weightcompared with VP from untreatedmales. Interestingly, VP from T�E2-treated KO males increased by 213%and became significantly larger than VPfrom T�E2 -treated control males,which only increased by 25% (Fig. 5A).All T�E2-treated KO males developedmarked urinary retention, resulting intwo deaths; no T�E2-treated controlsdeveloped urinary retention. VP fromT�E2-treated control and KO miceboth displayed stromal and epithelialhyperplasia (Fig. 5B) but this was morepronounced in KO. Epithelial hyper-trophy was also seen in T�E2-treatedcontrol and KO VP, but this was moredramatic in KO. Squamous metaplasiawas seen in T�E2-treated control andKO VP (Fig. 5B).
Prostatic response to castrationSeven days after castration, VP weight
was reduced by 63% in control malesand by 51% in KOs, resulting in sim-ilar sized VP in both controls (0.0035g � 0.004) and KO (0.0028 g �0.004).
Discussion
It is well established that stromal an-drogen action is vital for normal pros-tate development, but its role in adultprostate homeostasis, and thus pros-tatic diseases such as PCa and BPH, is
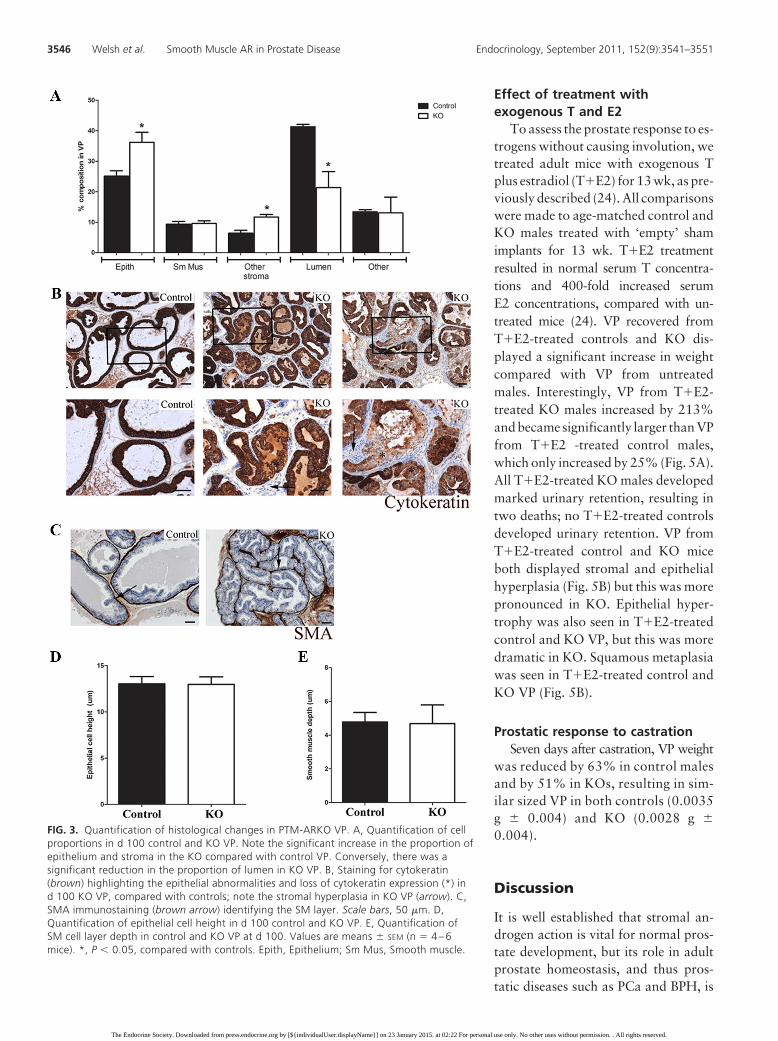
FIG. 3. Quantification of histological changes in PTM-ARKO VP. A, Quantification of cellproportions in d 100 control and KO VP. Note the significant increase in the proportion ofepithelium and stroma in the KO compared with control VP. Conversely, there was asignificant reduction in the proportion of lumen in KO VP. B, Staining for cytokeratin(brown) highlighting the epithelial abnormalities and loss of cytokeratin expression (*) ind 100 KO VP, compared with controls; note the stromal hyperplasia in KO VP (arrow). C,SMA immunostaining (brown arrow) identifying the SM layer. Scale bars, 50 �m. D,Quantification of epithelial cell height in d 100 control and KO VP. E, Quantification ofSM cell layer depth in control and KO VP at d 100. Values are means � SEM (n � 4 – 6mice). *, P � 0.05, compared with controls. Epith, Epithelium; Sm Mus, Smooth muscle.
3546 Welsh et al. Smooth Muscle AR in Prostate Disease Endocrinology, September 2011, 152(9):3541–3551
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

less clear. Furthermore, exactly how and via which stro-mal cell type androgens act has also remained unclear. Thecurrent study investigated the role for SM androgen actionin mediating normal adult prostate function, cell identity,and homeostasis using mice in which prostatic SM AR wasablated. These mice had significantly smaller prostates inadulthood, which exhibited histological abnormalities, al-tered gene expression, inflammation, and fibrosis. Thesechanges could not be explained by changes in serum T orE2 concentrations. Furthermore, KO prostates displayed
exaggerated epithelial cell proliferationin response to high concentrations ofexogenous E2, resulting in urinary re-tention in all KO. These results demon-strate that AR signaling via the SM isvital in mediating normal androgen-driven stromal-epithelial interactionsin the adult prostate, determining pros-tate homeostasis, cell identity, andfunction, and for limiting hormone-dependent prostatic-epithelial cellproliferation. These changes mayprovide some explanation for the rolefor SM androgen action in facilitatingprostate disease development and/orprogression and thus highlights an-drogen-dependent SM signaling as apossible new therapeutic target inprostate disease.
We have previously shown that ARis ablated from SM cells of the male re-productive tract from e 17.5 in KO mice(23, 24). As with the SV (24), this modeloffers a unique opportunity to investi-gate androgen action in postnatal SMcells, without preventing prostate for-mation or its prepubertal growth. Ab-sence of any effect on formation is prob-ably due to Cre recombinase not beingexpressed until after the prostate has al-ready been programmed to form (29).Furthermore, serum hormone concen-trations and AR expression in prostaticnon-SM cells were normal in these KO,so any prostatic phenotype must resultfrom ablation of SM AR. Control pros-tates greatly increase in weight between d35 and d 200 but PTM-ARKO prostatesdid not, resulting in adult KO prostatesbeing approximately 50–60% of nor-mal adult weight. In contrast, ablationof epithelial AR (PEARKO mice) onlycaused a 20% reduction in prostate
weight (19). Interestingly, castration of PTM-ARKOresulted in a smaller reduction in prostate weight thanin controls. This is consistent with tissue recombinationstudies, which suggested that stromal, not epithelial,AR is important in mediating the castration response(30). Together these studies show that SM AR is essen-tial for normal androgen-driven growth of the postpu-bertal prostate, with epithelial AR appearing to be lesscritical.
FIG. 4. Gene/protein expression in d 100 PTM-ARKO VP. A, E-cadherin (brown) wasexpressed in control d 100 VP (arrow) but was absent from some areas of epithelium in d 100KO VPs (arrowhead). A similar difference and pattern was observed for Sox 9 (B) C and D,p63 (brown) was expressed in the basal cells (arrow) in control and KO VP at d 35 (C)and d 100 (D), but fewer p63-positive cells were evident in KO VP at d 100 (arrow). E,Quantification of the number of p63-positive cells in d 100 control and KO VP. F, RelativemRNA expression of luminal epithelial, basal epithelial, and SM cell identity markers andepithelial cell secretory products in control and KO VP at d 100. Values are means � SEM (n �4 mice). *, P � 0.05; **, P � 0.01; ***, P � 0.001 compared with controls.
Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3547
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

The adult prostate is normally growth quiescent, de-spite high androgen concentrations (7), but growth can bereactivated via unknown mechanisms in prostate diseaseleading to enlarged prostates and urinary retention. Tissuerecombination studies indicate that epithelial cell prolif-eration depends on stromal, rather than epithelial, andro-gen action (30). Interestingly, epithelial cell proliferationwas not significantly altered in our KO VP, suggesting thatSM AR does not regulate epithelial cell proliferation undernormal hormonal conditions. Estrogens have also beenshown to stimulate prostate cell proliferation with pros-tates normally expressing both AR and ER (17, 18, 31);therefore, we tested whether exogenous estrogens couldelicit the same response in the absence of SM AR. Micewere treated with exogenous T�E2, which resulted in nor-mal serum T but hugely elevated serum E2 concentrationscompared with untreated control or KO mice (24). Thisregime increased prostate weight in both controls and KO,demonstrating that lack of SM AR does not prevent E2-driven prostate growth. However, the response was con-siderably enhanced in KO (213% increase in VP size) com-pared with controls (25% increase), such that KO VPbecame significantly larger than T�E2-treated control
VP. This is opposite to the SV responseto E2 in KO, because E2 increased con-trol, but not KO, SV size (24). TheT�E2-induced increase in VP size wasassociated with pronounced urinary re-tention in all treated KO males. Thissuggests that SM AR may normallylimit adult prostate proliferation andgrowth and that, in its absence, pros-tate homeostasis and cell numbers arepoorly controlled if the hormonal en-vironment is altered. Interestingly,SM AR expression is often lost in menwith aggressive metastatic PCa (32)whereas circulating estrogen concen-trations are either stable or slightly in-creased in aging men whereas T concen-trations fall, leading to an androgen-estrogen imbalance similar to, althoughnot as pronounced as, that induced byour T�E2 treatments. Our presentfindings suggest that this hormonal im-balance and loss of SM AR expressioncould permit uncontrolled prostate cellproliferation and so facilitate the devel-opment and/or progression of prostatedisease in aging men. This is not sur-prising given that stromal AR is sug-gested to regulate epithelial cell prolif-eration and apoptosis (7, 30). Loss of
SM AR may thus be one of the ways in which the epithe-lium is released from its normal tight control by the stromaandsoallowsPCa toproceed.This is inagreementwith thehypothesis that SM-epithelial interactions are perturbedin PCa, resulting in adverse consequences for both com-partments and disease progression (33). PTM-ARKOmice provide a new tool with which to investigate this andits mechanisms.
Despite the lack of increased epithelial cell proliferationin untreated adult PTM-ARKO prostates, apparent stro-mal and epithelial overcrowding or hyperplasia was ob-served at d 35 and increased with age. This suggests thatthe epithelial hyperplasia in KO VP reflects similar cellnumbers in a smaller space rather than increased prolif-eration per se. This results in an increased proportion ofepithelial and stromal cells and increased infolding, over-crowding, and desquamation of the epithelium in adultKO prostates, suggesting that SM AR is important formaintaining normal adult prostate structure and homeo-stasis. Furthermore, we also demonstrated that SM AR isimportant in maintaining adult prostate cell identitybecause expression of several cell identity markers was
FIG. 5. Gross VP morphology and histology of adult PTM-ARKO mice exposed to exogenousT and E2. A, Quantification of VP weight in adult control and KO mice exposed to T�E2 for13 wk. Values are means � SEM (n � 6 mice). *, P � 0.05, compared with control littermates.B, Hematoxylin and eosin staining of T�E2-treated adult KO and control VP. Histologicalabnormalities are obvious in T�E2 KO VP, with epithelial cell hyperplasia (arrowhead)resulting in epithelial cell desquamation into the lumens (L). Stromal cell hyperplasia is alsoobvious in T�E2-treated control and KO VP (arrow). Note the dense eosinophilic secretions inthe T�E2-treated KO lumens (*) and the squamous metaplasia in both the T�E2-treatedcontrol and KO VP (F). Con, Control.
3548 Welsh et al. Smooth Muscle AR in Prostate Disease Endocrinology, September 2011, 152(9):3541–3551
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

reduced in KO prostates in adulthood, but not at d 35.This highlights a progressive prostatic phenotype inthese mice, suggesting that absence of SM AR expres-sion, directly or indirectly, leads to less differentiatedprostates. This is similar to observations in cancer-as-sociated reactive stroma in which SM-marker expres-sion is reduced, associated with a poorer patient prog-nosis (34), and is consistent with studies showing thatstromal androgen action determines and maintains thefate of the epithelium (7, 30).
Given these observations, it is not surprising that epi-thelial cell identity was also altered in PTM-ARKO pros-tates. For example, E-cadherin expression was lost fromsome acini in adult KO prostates. E-cadherin normallymediates cell adhesion in prostatic luminal epithelium andmaintains epithelial differentiation and homeostasis (35).Aberrant or decreased expression of E-cadherin is associ-ated with epithelial-mesenchymal transition (36) andlarger more aggressive prostate tumors (37). The loss ofE-cadherin from KO epithelial cells may explain the des-quamation of epithelial cells and could be a direct conse-quence of the loss of SM AR or could be indirect due to theloss of SM identity. Epithelial stem cell markers Nkx3.1and p63 were also both reduced in adult KO VP. Nkx3.1is androgen dependent (38), regulates prostate epithelialcell differentiation, and marks a luminal stem cell that iscastrate resistant (13), whereas p63 is expressed in basalstem cells. Our results suggest that SM AR might regulateexpression of stem cell markers and/or the stem cell niche,possibly by altering the production of growth factors (39).Pten expression is also reduced in these mice, and studiesin mice and humans have associated loss of Pten (40),Nkx3.1 (41), or p63 (42) with initiation of PCa; however,PCa per se was not identified in KO mice. Further inves-tigations are required to understand the role of SM AR,and the associated changes, on PCa development andprogression.
Abnormal infiltration of immune cells was seen in thestroma and glands of KO VP at both d 35 and in adult-hood, but it is unclear whether this is a primary effect ofthe absence of SM AR or is secondary to the histologicalabnormalities observed in these mice. Some of the histo-logical and gene expression changes discussed above maybe induced indirectly by the inflammation seen in thismodel. For example, fibrosis was observed in the stromalcompartment at d 100 in KO VP, which is a commonconsequence of inflammation and/or aberrant changes ingrowth factors (26). Inflammatory mediators secreted bystromal cells attract immune cells and induce proliferationof both the stroma and the epithelium (43); therefore, fur-ther investigation is required to identify how SM AR sig-naling drives prostate fibrosis and inflammation and their
relationship to prostate homeostasis, gene expression, anddisease.
Based on the histological and differentiation abnormal-ities discussed above, it is not surprising that VP lumenvolume and epithelial cell-secretory function was im-paired in adult KO. For example, expression of secretorygenes Spink 3, prostate ß defensin 1, and Cox-1 in adultKO VP was reduced, whereas no change was seen in themRNA expression of probasin or SBP. This suggests thatprostate epithelial secretory function is not globally im-paired in the KO mice but that secretion of specific prod-ucts is affected. This may be an indirect effect of the epi-thelial cell abnormalities discussed above, such as loss ofepithelial cell identity, or the inflammation observed inthese mice because it is believed that epithelial, rather thanstromal, AR mediates epithelial secretory function (44).
In conclusion, we have demonstrated an essential rolein vivo for AR signaling via the SM cells in mediatingnormal androgen-driven stromal-epithelial interactions inthe mouse adult prostate, determining cell identity andfunction and limiting hormone-dependent epithelial cellproliferation. These results differ from those recently re-ported by Yu et al. (45) who ablated AR from the SM ofthe AP and demonstrated reduced epithelial cell prolifer-ation and infolding and little effect on differentiation.These differences in effects are most likely due to the useof different Cre and ARflox models; for example, theSM22-Cre line used by Yu et al. appears to target only theAP whereas we show that the MHC-Cre targets all pros-tate lobes. The PTM-ARKO mouse model therefore pro-vides a unique tool for identifying the underlying molec-ular mechanisms of SM androgen action in the prostatewithout preventing prostate formation. Further investiga-tions with these mice could provide new insight into therole for SM AR in human adult-onset prostate diseases.
Acknowledgments
We thank Karel De Gendt and Guido Verhoeven (Leuven Cath-olic University, Leuven, Belgium) for providing the ARflox miceand Michael Kotlikoff (Cornell University, Ithaca, New York)for the smMHC-Cre mice. We thank Axel Thomson (Universityof Edinburgh, Edinburgh, UK) for advice with the hormonetreatments and Mark Fisken (University of Edinburgh, Edin-burgh, UK) and the members of the imaging facility and assaylaboratory for technical assistance.
Address all correspondence and requests for reprints to: Dr.Michelle Welsh, University of Edinburgh, MRC HRSU, QMRI,47 Little France Crescent, Edinburgh, United Kingdom, EH164TJ. E-mail: [email protected].
This work was supported by the United Kingdom MedicalResearch Council.
Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3549
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

Present address for M.W.: The Beatson Institute for CancerResearch, Garscube Estate, Switchback Road, Glasgow G611BD, Scotland, United Kingdom.
Disclosure Summary: M.W., L.M., A.M., D.B., P.T.K.S.,R.M.S., and L.B.S. have nothing to declare.
References
1. George FW, and Wilson, J 1994 Gonads and ducts in mammals. In:Knobil E, Neill JD, eds. The physiology of reproduction. 2nd ed.New York: Raven Press; 3–27
2. Mooradian AD, Morley JE, Korenman SG 1987 Biological actionsof androgens. Endocr Rev 8:1–28
3. Wilson JD, George FW, Griffin JE 1981 The hormonal control ofsexual development. Science 211:1278–1284
4. Franks LM, Riddle PN, Carbonell AW, Gey GO 1970 A compar-ative study of the ultrastructure and lack of growth capacity of adulthuman prostate epithelium mechanically separated from its stroma.J Pathol 100:113–119
5. Cunha GR, Donjacour A 1987 Stromal-epithelial interactions innormal and abnormal prostatic development. Prog Clin Biol Res239:251–272
6. Chung LW, Cunha GR 1983 Stromal-epithelial interactions: II. Reg-ulation of prostatic growth by embryonic urogenital sinus mesen-chyme. Prostate 4:503–511
7. Cunha GR, Hayward SW, Wang YZ 2002 Role of stroma in carci-nogenesis of the prostate. Differentiation 70:473–485
8. Condon MS, Bosland MC 1999 The role of stromal cells in prostatecancer development and progression. In Vivo 13:61–65
9. McNeal JE 1988 Normal anatomy of the prostate and changes inbenign prostatic hypertrophy and carcinoma. Semin Ultrasound CTMR 9:329–334
10. Richardson GD, Robson CN, Lang SH, Neal DE, Maitland NJ,Collins AT 2004 CD133, a novel marker for human prostatic epi-thelial stem cells. J Cell Sci 117:3539–3545
11. Kurita T, Medina RT, Mills AA, Cunha GR 2004 Role of p63 andbasal cells in the prostate. Development 131:4955–4964
12. Lawson DA, Xin L, Lukacs RU, Cheng D, Witte ON 2007 Isolationand functional characterization of murine prostate stem cells. ProcNatl Acad Sci USA 104:181–186
13. Wang X, Kruithof-de Julio M, Economides KD, Walker D, Yu H,Halili MV, Hu YP, Price SM, Abate-Shen C, Shen MM 2009 Aluminal epithelial stem cell that is a cell of origin for prostate cancer.Nature 461:495–500
14. Pritchard C, Mecham B, Dumpit R, Coleman I, Bhattacharjee M,Chen Q, Sikes RA, Nelson PS 2009 Conserved gene expression pro-grams integrate mammalian prostate development and tumorigen-esis. Cancer Res 69:1739–1747
15. Majdic G, Millar MR, Saunders PT 1995 Immunolocalisation ofandrogen receptor to interstitial cells in fetal rat testes and to mes-enchymal and epithelial cells of associated ducts. J Endocrinol 147:285–293
16. Quigley CA, De Bellis A, Marschke KB, el-Awady MK, Wilson EM,French FS 1995 Androgen receptor defects: historical, clinical, andmolecular perspectives. Endocr Rev 16:271–321
17. Couse JF, Lindzey J, Grandien K, Gustafsson JA, Korach KS 1997Tissue distribution and quantitative analysis of estrogen receptor-�(ER�) and estrogen receptor-� (ER�) messenger ribonucleic acid inthe wild-type and ER�-knockout mouse. Endocrinology 138:4613–4621
18. Bianco JJ, Handelsman DJ, Pedersen JS, Risbridger GP 2002 Directresponse of the murine prostate gland and seminal vesicles to estra-diol. Endocrinology 143:4922–4933
19. Simanainen U, Allan CM, Lim P, McPherson S, Jimenez M, ZajacJD, Davey RA, Handelsman DJ 2007 Disruption of prostate epi-
thelial androgen receptor impedes prostate lobe-specific growth andfunction. Endocrinology 148:2264–2272
20. Wu CT, Altuwaijri S, Ricke WA, Huang SP, Yeh S, Zhang C, NiuY, Tsai MY, Chang C 2007 Increased prostate cell proliferation andloss of cell differentiation in mice lacking prostate epithelial andro-gen receptor. Proc Natl Acad Sci USA 104:12679–12684
21. Niu Y, Altuwaijri S, Yeh S, Lai KP, Yu S, Chuang KH, Huang SP,Lardy H, Chang C 2008 Targeting the stromal androgen receptor inprimary prostate tumors at earlier stages. Proc Natl Acad Sci USA105:12188–12193
22. Cunha GR, Alarid ET, Turner T, Donjacour AA, Boutin EL, FosterBA 1992 Normal and abnormal development of the male urogenitaltract. Role of androgens, mesenchymal-epithelial interactions, andgrowth factors. J Androl 13:465–475
23. Welsh M, Saunders PT, Atanassova N, Sharpe RM, Smith LB 2009Androgen action via testicular peritubular myoid cells is essential formale fertility. FASEB J 23:4218–4230
24. Welsh M, Moffat L, Jack L, McNeilly A, Brownstein D, SaundersPT, Sharpe RM, Smith LB 2010 Deletion of androgen receptor in thesmooth muscle of the seminal vesicles impairs secretory function andalters its responsiveness to exogenous testosterone and estradiol.Endocrinology 151:3374–3385
25. Xin HB, Deng KY, Rishniw M, Ji G, Kotlikoff MI 2002 Smoothmuscle expression of Cre recombinase and eGFP in transgenic mice.Physiol Genomics 10:211–215
26. Shappell SB, Thomas GV, Roberts RL, Herbert R, Ittmann MM,Rubin MA, Humphrey PA, Sundberg JP, Rozengurt N, Barrios R,Ward JM, Cardiff RD 2004 Prostate pathology of genetically engi-neered mice: definitions and classification. The consensus reportfrom the Bar Harbor meeting of the Mouse Models of Human Can-cer Consortium Prostate Pathology Committee. Cancer Res 64:2270–2305
27. Ricke WA, Ishii K, Ricke EA, Simko J, Wang Y, Hayward SW,Cunha GR 2006 Steroid hormones stimulate human prostate cancerprogression and metastasis. Int J Cancer 118:2123–2131
28. Hooley RP, Paterson M, Brown P, Kerr K, Saunders PT 2009 Intra-testicular injection of adenoviral constructs results in Sertoli cell-specific gene expression and disruption of the seminiferous epithe-lium. Reproduction 137:361–370
29. Welsh M, Saunders PT, Fisken M, Scott HM, Hutchison GR, SmithLB, Sharpe RM 2008 Identification in rats of a programming win-dow for reproductive tract masculinization, disruption of whichleads to hypospadias and cryptorchidism. J Clin Invest 118:1479–1490
30. Cunha GR, Ricke W, Thomson A, Marker PC, Risbridger G, Hay-ward SW, Wang YZ, Donjacour AA, Kurita T 2004 Hormonal,cellular, and molecular regulation of normal and neoplastic pros-tatic development. J Steroid Biochem Mol Biol 92:221–236
31. Risbridger G, Wang H, Young P, Kurita T, Wang YZ, Lubahn D,Gustafsson JA, Cunha G, Wong YZ 2001 Evidence that epithelialand mesenchymal estrogen receptor-� mediates effects of estrogenon prostatic epithelium. Dev Biol 229:432–442
32. Wikstrom P, Marusic J, Stattin P, Bergh A 2009 Low stroma an-drogen receptor level in normal and tumor prostate tissue is relatedto poor outcome in prostate cancer patients. Prostate 69:799–809
33. Cunha GR, Hayward SW, Dahiya R, Foster BA 1996 Smooth mus-cle-epithelial interactions in normal and neoplastic prostatic devel-opment. Acta Anat (Basel) 155:63–72
34. Ayala G, Tuxhorn JA, Wheeler TM, Frolov A, Scardino PT, OhoriM, Wheeler M, Spitler J, Rowley DR 2003 Reactive stroma as apredictor of biochemical-free recurrence in prostate cancer. ClinCancer Res 9:4792–4801
35. Imhof BA, Vollmers HP, Goodman SL, Birchmeier W 1983 Cell-cellinteraction and polarity of epithelial cells: specific perturbation us-ing a monoclonal antibody. Cell 35:667–675
36. Gravdal K, Halvorsen OJ, Haukaas SA, Akslen LA 2007 A switchfrom E-cadherin to N-cadherin expression indicates epithelial tomesenchymal transition and is of strong and independent impor-
3550 Welsh et al. Smooth Muscle AR in Prostate Disease Endocrinology, September 2011, 152(9):3541–3551
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.

tance for the progress of prostate cancer. Clin Cancer Res 13:7003–7011
37. Rubin MA, Mucci NR, Figurski J, Fecko A, Pienta KJ, Day ML 2001E-cadherin expression in prostate cancer: a broad survey using high-density tissue microarray technology. Hum Pathol 32:690–697
38. Lei Q, Jiao J, Xin L, Chang CJ, Wang S, Gao J, Gleave ME, WitteON, Liu X, Wu H 2006 NKX3.1 stabilizes p53, inhibits AKT ac-tivation, and blocks prostate cancer initiation caused by PTEN loss.Cancer Cell 9:367–378
39. Berry PA, Maitland NJ, Collins AT 2008 Androgen receptor sig-nalling in prostate: effects of stromal factors on normal and cancerstem cells. Mol Cell Endocrinol 288:30–37
40. Wang S, Gao J, Lei Q, Rozengurt N, Pritchard C, Jiao J, Thomas GV,Li G, Roy-Burman P, Nelson PS, Liu X, Wu H 2003 Prostate-specificdeletion of the murine Pten tumor suppressor gene leads to meta-static prostate cancer. Cancer cell 4:209–221
41. Shen MM, Abate-Shen C 2003 Roles of the Nkx3.1 homeobox gene
in prostate organogenesis and carcinogenesis. Dev Dyn 228:767–778
42. Signoretti S, Waltregny D, Dilks J, Isaac B, Lin D, Garraway L, YangA, Montironi R, McKeon F, Loda M 2000 p63 is a prostate basal cellmarker and is required for prostate development. Am J Pathol 157:1769–1775
43. Begley LA, Kasina S, MacDonald J, Macoska JA 2008 The inflam-matory microenvironment of the aging prostate facilitates cellularproliferation and hypertrophy. Cytokine 43:194–199
44. Donjacour AA, Cunha GR 1993 Assessment of prostatic proteinsecretion in tissue recombinants made of urogenital sinus mesen-chyme and urothelium from normal or androgen-insensitive mice.Endocrinology 132:2342–2350
45. Yu S, Zhang C, Lin CC, Niu Y, Lai KP, Chang HC, Yeh SD, ChangC, Yeh S 2011 Altered prostate epithelial development and IGF-1signal in mice lacking the androgen receptor in stromal smooth mus-cle cells. Prostate 71:517–524
Become a member of The Endocrine Society and receive discounts on color and page charges
www.endo-society.org/join
Endocrinology, September 2011, 152(9):3541–3551 endo.endojournals.org 3551
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 23 January 2015. at 02:22 For personal use only. No other uses without permission. . All rights reserved.




















