Structure of the DNA Repair Helicase Hel308 Reveals DNA Binding and Autoinhibitory Domains
Upload
independentCategory
view
1download
0
Genomics 69, 162–173 (2000)doi:10.1006/geno.2000.6281, available online at http://www.idealibrary.com on
SMARCAD1, a Novel Human Helicase Family-Defining MemberAssociated with Genetic Instability: Cloning, Expression, and
Mapping to 4q22–q23, a Band Rich in Breakpoints and DeletionMutants Involved in Several Human Diseases
Chaker N. Adra,* ,† ,1 Jose-Luiz Donato,* Rachel Badovinac,‡ Farzand Syed,* Reshma Kheraj,*Hongbo Cai,* Colin Moran,* Mitchell T. Kolker,* Helen Turner,† Stanislawa Weremowicz,§
Taro Shirakawa,¶,\ Cynthia C. Morton,§ Lowell E. Schnipper,* and Reed Drews*
*Department of Medicine and †Department of Pathology, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston,Massachusetts 02215; §Department of Pathology and Department of Obstetrics, Brigham and Women’s Hospital, Harvard Medical
School, Boston, Massachusetts 02215; ¶Experimental Medicine Unit, University of Wales, Swansea, Swansea, United Kingdom;\Department of Health Promotion and Human Behavior, Kyoto University Graduate School of Medicine, Kyoto, 606-8501, Japan;
and ‡Harvard School of Dental Medicine and Harvard School of Public Health, Boston, Massachusetts 02215
Received January 12, 2000; accepted June 6, 2000
Members of the DEAD/H box-containing helicase su-perfamily include proteins essential to genome replica-tion, repair, and expression. We report here the cloningand initial characterization of a novel human member ofthis protein family, designated hHel1 (human helicase1), now designated SMARCAD1 by HUGO. This DEAD/Hbox-containing molecule has seven highly conserved se-quence regions that allow us to place it in the SNF2family of the helicase superfamily. Uniquely, though,hHel1 contains two DEAD/H box motifs, a property notreported to be shared by any other SNF2 family mem-bers. This defines a new subfamily consisting of hHel1and its homologues. In addition to these DEAD/H box/ATP-binding motifs, hHel1 has a putative nuclear local-ization signal and several regions that may mediate pro-tein–protein interactions. Expression analysis indicatesthat hHel1 transcripts are ubiquitous, with particularlyhigh levels in endocrine tissue. We have mapped thegene for hHel1 to human chromosome 4q22–q23; thisregion is rich in breakpoints and deletion mutants ofgenes involved in several human diseases, notably softtissue leiomyosarcoma, hepatocellular carcinoma, andhematologic malignancies. Our observation that humanHel1 gene overexpression is present in an E1A-express-ing cell line with increased capacity for gene reactiva-tion events by genomic rearrangement suggests that hu-man Hel1 may play a role in genetic instabilitydevelopment. © 2000 Academic Press
Sequence data from this article have been deposited with theEMBL/GenBank Data Libraries under Accession No. AY008271.
1 To whom correspondence should be addressed at Department ofMedicine, Hematology/Oncology Division, Beth Israel DeaconessMedical Center, Harvard Medical School, 99 Brookline Avenue, Bos-ton, MA 02215. Telephone: (617) 667-3766. Fax: (617) 975-7905.
E-mail: [email protected].1620888-7543/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
INTRODUCTION
A vast array of proteins interact with DNA and RNA.While some of these proteins regulate transcriptionand translation, others are involved in the preserva-tion of the sequence integrity of genomic DNA. It isclear that many form complexes that contain function-ally specialized subunits. Complexity in the architec-ture of these protein machines apparently is highlyconserved throughout evolution. As our knowledge ofthe sequence of the human genome nears completion,there is an ever-growing need to understand the mech-anisms involved in the maintenance of the genome. Wetherefore have initiated an effort to clone and charac-terize novel DNA and RNA interacting proteins.
Members of the helicase superfamily are essential tomany aspects of nucleic acid metabolism, such as DNAreplication, transcription, and excision and repair ofmutated bases (Lohman and Bjornson, 1996; West,1996). Helicases comprise a superfamily of proteinsthat interact with DNA or RNA (Lohman and Bjorn-son, 1996). Proteins are classified as helicases basedupon specific biochemical activity (unwinding of DNAor RNA helices) or upon sequence homology. Manyhelicases contain a signature protein sequence motif,in which an ATPase B motif DEAD/H box (a conservedAsp-Glu-Ala-Asp/His motif) is found in tandem with anA-type ATP-binding site (AXXXXGKT) (Bork and Koo-nin, 1993; Luking et al., 1998). Thus helicases of thissuperfamily apparently use energy derived from NTPhydrolysis (Lohman and Bjornson, 1996). Despite theassignment of function to each motif, the functionaleffect of mutation of a given motif varies between he-licases. For example, in elf-4A, a mammalian RNA
helicase, mutation of the A type motif affected ATP
vBaDAl
tittdifnqutlolemo
dEb1htwslqnsf(
lh
ad
1i
mfi(
peole
163SMARCAD1, A MEMBER OF THE HUMAN HELICASE FAMILY
binding, while mutation of the DEAD/H region affectedATP hydrolysis (Pause et al., 1993). In at least one viralhelicase, the situation is different: helicase function isdisrupted by mutation of the DEAD/H box, leavingATPase activity intact (Heilek and Peterson, 1997).DEAD/H boxes are conserved within helicase super-family proteins from viruses, bacteria, and yeast tohigher eukaryotes.
By the nomenclature of Pazin (see Pazin and Ka-donaga, 1997) DEAD/H box-containing helicases canbe subdivided into several families based upon se-quence conservation. One such family is the SNF2-likeprotein family (subsequently referred to as the SNF2family). SNF2 helicases have seven distinct, highlyconserved motifs. The function of each of these sevenconserved motifs has not yet been defined. However, allmembers of this family have the ability to hydrolyzeNTPs, and many have been shown to function in tran-scriptional activation.
DEAD/H box-containing members of the helicase su-perfamily function in transcriptional regulation, re-combination, and DNA repair. It therefore is not sur-prising that many hereditary pathologic conditions areassociated with defects in their function (Ellis, 1997).Thus far, Cockayne syndrome B (Troelstra et al., 1992;an Gool et al., 1997), xeroderma pigmentosum group, and trichothiodystrophy (Weeda et al., 1997; Coin etl., 1998, 1999) have been ascribed to genes encodingEAD/H box-containing proteins. Moreover, the XNP/TRX gene, involved in mental retardation syndromes
inked to a-thalassemia, encodes a helicase (Villard etal., 1997). Bloom and Werner syndromes also arelinked to helicases (Ellis, 1997; Huang et al., 1998).Cell culture studies indicate that certain oncogenictransformations are accompanied by upregulation inthe levels of DEAD/H box proteins. This indicates ei-ther a dysregulation of genome maintenance pathwaysas part of overproliferation or an effort on the part oftransformed cells to contend with increased levels ofDNA damage (Godbout et al., 1998).
We report the cloning of a novel human member ofhe DEAD/H box-containing proteins, hHel1, now des-gnated SMARCAD1 by HUGO (SWI/SNF-related, ma-rix-associated, actin-dependent regulator of chroma-in, subfamily a, containing DEAD/H box 1). Weescribe its sequence features that are consistent withts inclusion in the SNF2 family of the helicase super-amily. Chromosomal mapping has positioned thisewly described human helicase at chromosome 4q22–23, a region linked to several human diseases. It isbiquitously expressed in human tissues, supportinghe essential role of this helicase in fundamental cel-ular function. Our observation that human Hel1 geneverexpression is present in an E1A-expressing celline with increased capacity for gene reactivationvents by genomic rearrangement suggests that hu-an Hel1 is associated with genetic instability devel-
pment.
MATERIALS AND METHODS
cDNA cloning, sequencing, and analysis. Duplicate filters of aphage cDNA library in the l gt11 vector (Clontech, Palo Alto, CA)
erived from human fetal brain RNAs were screened with mousetl-1 cDNA, a gift of Dr. Achim Gossler (Soininen et al., 1992), andy cDNA probes derived by PCR as described (Adra et al., 1994, 1996,997, 1998). Hybridization and washing were carried out underigh-stringency conditions (65°C, 63 standard saline citrate). Posi-ive clones were identified by autoradiography. Individual clonesere then isolated after two rounds of secondary screening under
imilar stringency conditions. The human cDNA inserts were ana-yzed by agarose electrophoresis and then sequenced by direct se-uencing and also by sequencing subclones using the BigDye Termi-ator Cycle Sequencing Ready Reaction Kit and ABI Prism 377 DNAequencer (PE Applied Biosystems, Foster City, CA). One potentialull-length cDNA clone was identified and designated human Hel1human helicase 1).
Expression analysis. Northern and Master blot filters of poly(A)1
RNA from normal human tissues were purchased from Clontech.The filters were hybridized with [a-32P]dCTP-labeled human Hel1cDNA at high stringency, under the conditions described by Adra etal. (1994, 1996, 1997, 1998).
Fluorescence in situ hybridization (FISH). Two micrograms ofthe cDNA containing the full-length human Hel1 insert was labeledwith biotin-11–dUTP using the BioNIck Labeling System (GibcoBRL, Gaithersburg, MD). DNA was coprecipitated with 60 mg ofCot-1 DNA (Gibco BRL) and resuspended in 13 TE at 200 mg/ml.
Hybridization of metaphase chromosomes from peripheral bloodymphocytes from two normal males was performed with the labeleduman Hel1 probe at a concentration of 20 mg/ml in Hybrisol VI as
previously described (Adra et al., 1996, 1998). Metaphase chromo-somes were stained with 4,6-diamidino-2-phenylindole dihydrochlo-ride (DAPI; Oncor, Gaithersburg, MD).
The map position of the human Hel1 probe was determined byvisual inspection of the fluorescent signal on the DAPI-stained meta-phase chromosomes. Twenty-eight metaphases were examined forprobe localization.
EN/NIH 2-20 E1A transfection and cDNA differential analysis.The derivation of the EN/NIH 2-20 cell line from NIH3T3 fibroblastswas described elsewhere (Drews et al., 1992, 1998). The EN/NIH2-20 cell line contains, by Southern analysis, two stable integrants ofa variant of the retrovirus vector pZipNeoSV(x)1 that was modifiedby (1) deleting enhancer elements from the long terminal repeats(LTRs) and (2) deleting the 39 splice acceptor site located between theBamHI site and neo. This retrovirus vector variant retains promoterelements in the 59 and 39 LTRs. Neo transcripts are absent byNorthern analysis.
The parental EN/NIH 2-20 cells were transfected (by electropora-tion) with the pMSG(E1AWT) construct expressing a gene insert forE1A and conferring mycophenolic acid resistance (MPAR) (Drews et
l., 1992). This was followed by selection in MPA-containing me-ium.Independent, randomly selected foci (200–500 cells) emerging
4–21 days into selection were harvested individually and expandednto the MPA medium to 10 3 106 to 30 3 106 cells for further
analysis. Before G418 selection, all expanded MPAR subclones wereaintained in nonselective medium for three passages to allow suf-cient cell growth recovery following transfer from selective mediumDrews et al., 1992, 1998).
We performed G418 selections [600 mg (total) of G418/ml, corre-sponding to 93 IC50, half-maximal inhibitory concentration, forG418] on a total of 3.0 3 106 cells plated at a density of 1.5 3 106 cells
er 150-mm tissue culture plate. We counted G418R foci (.50 cells)merging after 28 to 35 days to determine frequencies of G418R focibserved per cell placed into selection. Independent, randomly se-ected G418R foci (100–200 cells) were harvested individually andxpanded to 10 3 106 to 30 3 106 cells in G41 8 medium for further
analysis (Drews et al., 1992, 1998).

Rac1dipf
BwtEsfipWps
lps1b3esawth9pwmtn
D
hqThopGb
rnGsic
qnetehtwa
htbhSapFaia
h
gtspt
Cstuaaaaeicfiwt
tbmC
164 ADRA ET AL.
Quantitation of neo reactivation events and preparation of totalNA were performed as described in Drews et al. (1998). Differentialnalysis of EN/NIH 2-20 and E1A-expressing EN/NIH 2-20 sub-lones with varying capacities for neo reactivation (Drews et al.,998) was performed using RNAmap Kit A (GenHunter Corp.) asescribed in Liang and Pardee (1992). Band excision, PCR, subclon-ng, and sequence analysis were used to identify differentially ex-ressed RNA. Finally, confirmatory Northern analysis was per-ormed as described in Drews et al. (1998).
RESULTS
cDNA Cloning of an Etl-1-Related Human Protein
Soininen et al. (1992) reported the cloning of a novelmouse gene, Etl-1, with a number of interesting fea-tures. First, the deduced Etl-1 protein has considerablehomology to helicase proteins contained within theyeast SWI/SNF complex and to a Drosophila protein,
rahma, with putative helicase activity. Murine Etl-1as one of the first mammalian proteins of this ex-
ended family to be cloned. Second, in situ analysis oftl-1 shows a widespread tissue distribution; latertudies at the protein level (Schoor et al., 1993) con-rmed that Etl-1 is a nuclear protein and that Etl-1rotein levels increase in early embryo development.e hypothesized that the human homologue of Etl-1
lays an equally important role. Based on this, weought to clone the human counterpart of Etl-1.A cDNA fragment derived from the full-length pub-
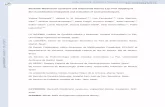
ished Etl-1 sequence was radiolabeled and used torobe a human fetal brain cDNA library. We isolated aingle cDNA, designated human Hel1 (human helicase), containing a 3081-bp open reading frame, flankedy 78 bp of 59 untranslated region (UTR) and a 1779-bp9 UTR, the latter with a polyadenine sequence at thextreme 39 end. The complete sequence of this cDNA ishown in Fig. 1A. We generated the predicted aminocid sequence encoded by the ORF and aligned thisith the published Etl-1 protein. Figure 1B confirms
hat the novel human Hel1 ORF is in fact the humanomologue of murine Etl-1, with 95.0% homology and2.0% identity, at the amino acid level. human Hel1 isredicted to encode a protein 1027 amino acids longith a calculated molecular mass of 117 kDa. No trans-embrane domains are predicted, although the pro-
ein has several extended hydrophilic stretches, a sig-ature for a globular structure.
omain Analysis of hHel1 Classifies It as a Helicaseby Homology
We carried out a Proscan analysis of hHel1, the noveluman Etl-1 homologue, to identify protein subse-uences that may reveal the function of the protein.he results of this analysis are summarized in Table 1.Hel1 has characteristics that indicate it is a memberf the helicase superfamily by homology. The mostrominent features are the presence of two “DEA/D/H” boxes, a conserved marker of DNA- and RNA-
inding proteins classified as helicases in organisms tanging from Escherichia coli to mouse (Bork and Koo-in, 1993; Lohman and Bjornson, 1996). Both DEA/D/H boxes are preceded by consensus NTP-binding
ites. Thus, hHel1 is likely to bind NTPs, which mayndicate an ATP-dependent helicase activity. This is aommon feature in DEA/GD/H box proteins.hHel1 contains a bipartite nuclear targeting se-
uence at positions 721–738. Additionally, a putativeuclear localization signal (KKRKK) (Iida et al., 1992)xists at positions 342–346. These findings suggesthat hHel1 functions in the nucleus. Finally, there isvidence that the hHel1 protein may be able to formomo- or hetero-complexes, with itself or other pro-eins, since the sequence contains motifs associatedith protein–protein interactions: the leucine zippernd helix-loop-helix motifs described in Table 1.As Etl-1 has been assigned to the SNF2 family of the
elicase superfamily (Schoor et al., 1999), we reasonedhat, by homology, hHel1, too, is an SNF2 family mem-er. The SNF2 family of proteins is defined by sevenighly conserved motifs, collectively considered theNF2 domain (Bork and Koonin, 1993). We performedn alignment analysis of the hHel1 protein againstreviously identified family members, presented inig. 2. hHel1 contains all seven conserved motifs thatre unique to the SNF2 family of DEAD/H box-contain-ng helicases (Table 1, Fig. 2). Thus, like Etl-1, hHel1 is
member of the SNF2 family.
Hel1 Shows a Ubiquitous Tissue Distribution
Helicase proteins are involved in many aspects ofene regulation. Depending on their specific function,hey can exhibit both tissue- and developmental stage-pecific expression patterns. We investigated the ex-ression pattern of hHel1 in a broad range of humanissues.
We used a human mRNA dot blot (RNA Master Blot,lontech) to analyze the expression of the hHel1 tran-cript in a wide range of mature and fetal humanissues. The data are shown in Fig. 3A. hHel1 shows abiquitous tissue distribution, as would be expected forprotein involved in a fundamental aspect of nucleic
cid metabolism. In mature tissues, expression levelsre highest in endocrine tissues, lung, colon, tracheand placenta. hHel1 is abundant in adult hematopoi-tic tissues (spleen, thymus, lymph nodes), but is lown mature peripheral leukocytes. In the brain, signifi-ant expression is localized to the cerebellum and therontal lobe, although all regions of the brain are pos-tive. hHel1 is abundant in all the fetal tissues tested,ith the strongest signals in kidney, liver, spleen, and
hymus.To ensure that we were specifically hybridizing a
ranscript of the correct size for hHel1in the Masterlot, we performed Northern analysis of multiple hu-an tissues (Fig. 3B, Multiple Tissue Northern Blot,lontech) using the same human Hel1 probe. Two
ranscripts, probably splice variants, in the range of

s
165SMARCAD1, A MEMBER OF THE HUMAN HELICASE FAMILY
FIG. 1. (A) Complete cDNA and deduced amino acid sequence of SMARCAD1 (human Hel1). Amino acid sequence of the longest ORF ishown, beginning at the presumed translational start site. TGA (stop) codon is followed by a 39 UTR containing an AATAAA polyadenylation
sequence (4898–4903 bp). (B) Alignment of murine Etl-1 and SMARCAD1 (hHel1) proteins. Clustal alignment of ORFs for murine Etl-1 andthe predicted protein sequence for the novel human cDNA, SMARCAD1 (hHel1), isolated in this study. Dark gray background indicates
identity; nonmatching residues are highlighted.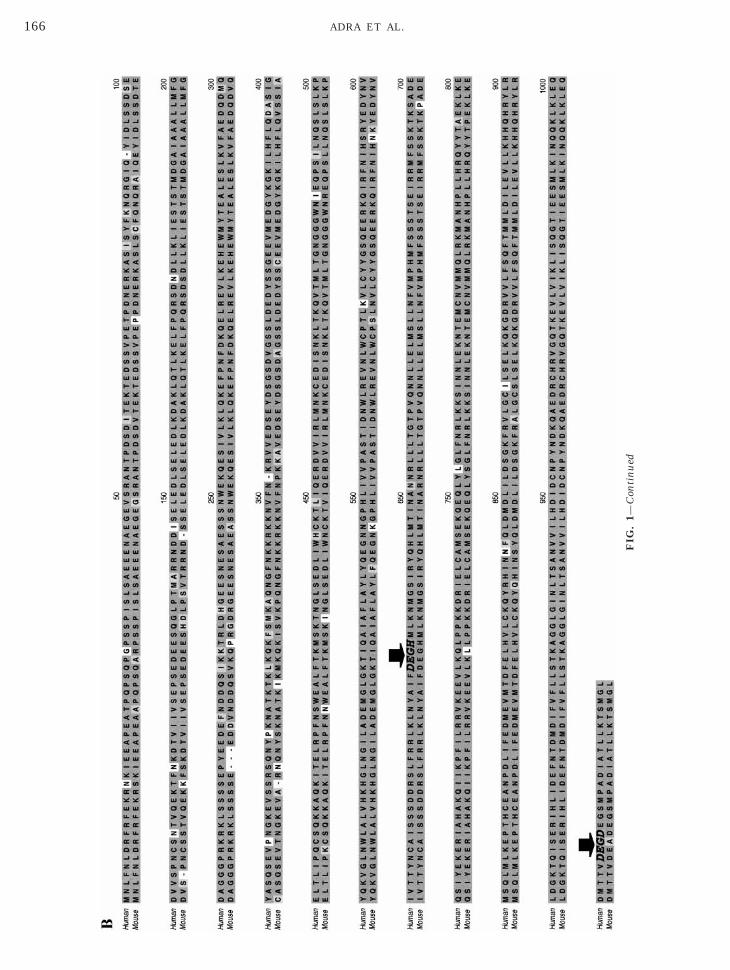
FIG
.1—
Con
tin
ued
166 ADRA ET AL.

ttR
167SMARCAD1, A MEMBER OF THE HUMAN HELICASE FAMILY
the predicted size for hHel1 mRNA (5 kb) were the onlyhybridizing species observed with our human Hel1probe.
It should be noted that there are discrepancies in thedetected levels of expression between the Master blotand the Northern blot. This is due to the fact that whilethe Master blot is standardized via normalization toallow semiquantitative evaluation, the Northern blot isnot. Thus, one should evaluate relative levels of tissue-specific expression from the Master blot and considerthe Northern blot a confirmation of Master blot valid-ity.
Human Hel1 Maps to Chromosome 4q22–q23
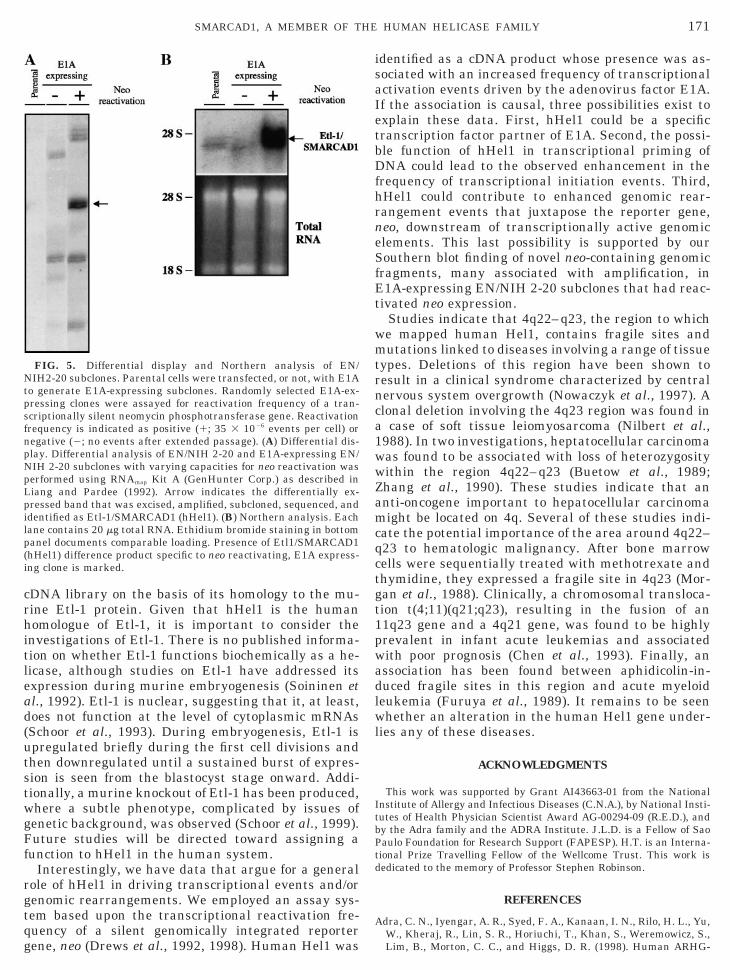
Helicase proteins are fundamental to DNA and RNAmetabolism. It therefore is not surprising that a num-ber of genetic disorders have been mapped to muta-tions or polymorphisms in helicase genes. For example,Cockayne syndrome B (CSB), a-thalassemia-associ-ated X-linked syndrome (ATRX), and xeroderma pig-mentosum group B disorders are all wholly or partlyattributable to a defect in a helicase protein (Ellis,1997). For this reason, we performed FISH analysis todetermine the chromosomal location of the humanHel1 gene and to ascertain its possible link to diseasesthrough chromosomal colocalization.
The human Hel1 gene was precisely assigned to
TABLE 1
Protein Sequence Features of SMARCADI (hHel1)and Implied Function
Postion(aa) Signature
Homology toconsensus (%) Implied function
87–94 RNP-1 89 Nucleic acidinteraction
500–788 SNF2 — —522–529 P-loop 79 ATP/GTP binding897–904 P-loop 79 ATP/GTP binding721–738 Bipartite nuclear
targetingsequence
100 Nucleartranslocation
629–632 DEA/GH/D 100 DEAD/H boxATP-dependenthelicasesignature
1006–1009 DEA/GH/D 100 DEAD/H boxATP-dependenthelicasesignature
553–576 Homeobox 77 Protein–proteindimerization
427–448 Leucine zipper 75 Protein–proteindimerization
845–866 Leucine zipper 75 Protein–proteindimerization
Note. Protein domain signatures with a high level of homology tohe consensus sequence are listed, with calculated percentage iden-ity to consensus. Prepared using Proscan and Prosite databases.NP, ribonucleoprotein; aa, amino acid.
4q22–q23 using FISH (Fig. 4). In all metaphase prep-
arations analyzed, the hybridization signal was foundto be present on the long arm of chromosome 4: in 25metaphase spreads, both copies of chromosome 4 werelabeled, and in 3 metaphase spreads, signal was de-tected on one chromosome 4.
Adenovirus E1A and Human Hel1 Cooperate for NeoAllelic Reactivation in Assays of GenomicInstability
We previously employed an assay system to identifyviral and other factors that can contribute to the de-velopment of genomic instability. This type of assayhas been used to identify cellular partners of the ade-novirus E1A transcription factor that can cooperate toenhance reactivation frequency of a stably integratedgenomic reporter gene (Drews et al., 1992, 1998). Invivo it is possible that these partners could contributeto the process of adenoviral oncogenesis. We examinedwhether human HelI could affect the frequency of E1A-driven reactivation of such a reporter, comprisinggenomically integrated transcriptionally silent neomy-cin phosphotransferase alleles.
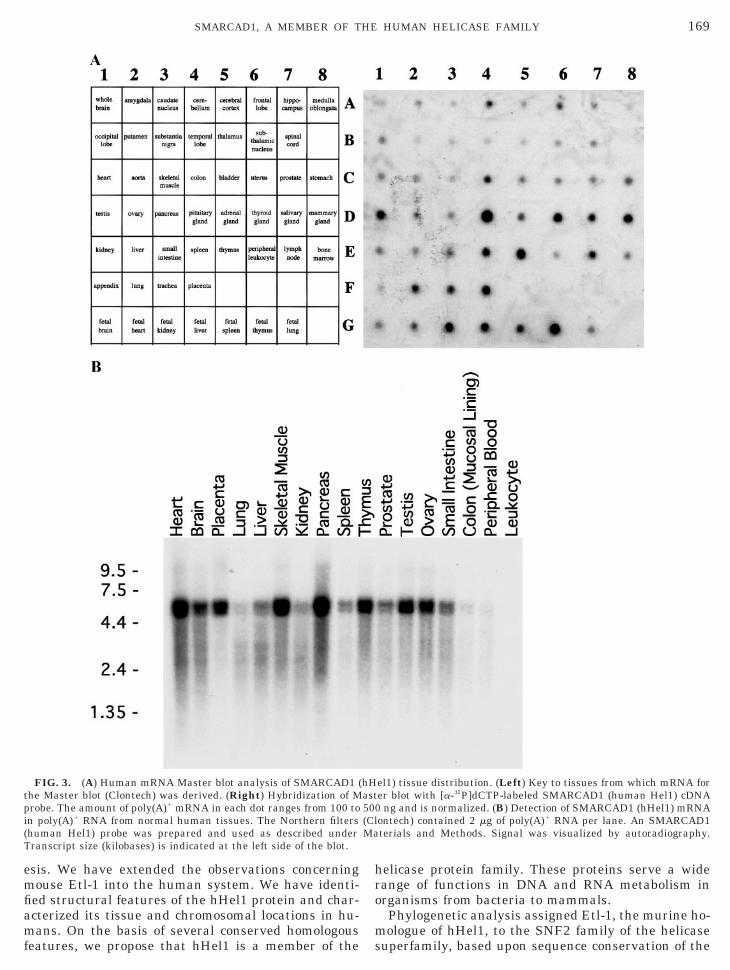
The EN/NIH 2-20 cell line is a derivative of NIH3T3fibroblasts containing two stable integrants of a tran-scriptionally silent neomycin phosphotransferase gene(neo). We transfected these cells with plasmid contain-ing wildtype adenovirus E1A (serotype 5) and ran-domly selected 21 E1A-expressing clones (E1A-EN/NIH 2-20) that displayed (n 5 9) or did not display (n 512) neo transcriptional reactivation. To identify reacti-vation-enhancing partners of E1A, we performed dif-ferential display on total RNA prepared from the fol-lowing sources: (1) parental EN/NIH 2-20, (2) E1A-EN/NIH 2-20 without neo reactivation (no reactivationobserved after extensive passage), and (3) E1A-EN/NIH 2-20 with neo reactivation (numerous reactivationevents; 35 3 1026/cell). Differential analysis revealedthe presence of a specific band in neo-reactivating E1A-expressing cells that was absent in E1A-expressing cellclones transcriptionally silent for neo reactivation (Fig.5A). Band excision, PCR, subcloning, and sequenceanalysis revealed that this difference product was infact Etl-1/hHel1 mRNA. Finally, Northern analysis ofthe parental EN-NIH 2-20- and E1A-expressing clonesconfirmed this finding (representative Northern blot;Fig. 5B). Thus the presence of Etl-1/hHel1 in neo-reac-tivating E1A-expressing cells suggests that it is asso-ciated with genomic rearrangements. If this associa-tion is causal, hHel1 could act alone or in cooperationwith E1A to produce these events (Drews et al., 1998).
DISCUSSION
The present report details the cloning and character-ization of human Hel1, a human gene encoding a pu-tative helicase by homology. human Hel1 is a homo-logue of the murine Etl-1 gene, identified on the basis
of its differential expression during mouse embryogen-
FIG
.2.
Par
tial
sequ
ence
alig
nm
ent
ofS
MA
RC
AD
1(h
Hel
1)an
dot
her
DE
A/G
D/H
box-
con
tain
ing
SN
F2
prot
ein
s.S
even
con
serv
edre
gion
sar
epr
esen
t,w
ith
the
nu
mbe
rof
sepa
rati
ng
resi
dues
indi
cate
dbe
twee
nth
ebl
ocke
dam
ino
acid
s.In
SM
AR
CA
D1
(hH
el1)
,th
eD
EA
/GD
/Hbo
xes
are
con
tain
edw
ith
indo
mai
nII
and
inth
eC
-ter
min
alre
gion
ofth
epr
otei
n.P
-loo
pN
TP
-bin
din
gsi
tes
(EM
GL
GK
T)
are
con
tain
edw
ith
inre
gion
Ian
dth
eC
-ter
min
us.
168 ADRA ET AL.
(T
169SMARCAD1, A MEMBER OF THE HUMAN HELICASE FAMILY
esis. We have extended the observations concerningmouse Etl-1 into the human system. We have identi-fied structural features of the hHel1 protein and char-acterized its tissue and chromosomal locations in hu-mans. On the basis of several conserved homologous
FIG. 3. (A) Human mRNA Master blot analysis of SMARCAD1 (the Master blot (Clontech) was derived. (Right) Hybridization of Mprobe. The amount of poly(A)1 mRNA in each dot ranges from 100 toin poly(A)1 RNA from normal human tissues. The Northern filtershuman Hel1) probe was prepared and used as described underranscript size (kilobases) is indicated at the left side of the blot.
features, we propose that hHel1 is a member of the
helicase protein family. These proteins serve a widerange of functions in DNA and RNA metabolism inorganisms from bacteria to mammals.
Phylogenetic analysis assigned Etl-1, the murine ho-mologue of hHel1, to the SNF2 family of the helicase
el1) tissue distribution. (Left) Key to tissues from which mRNA forer blot with [a-32P]dCTP-labeled SMARCAD1 (human Hel1) cDNA
ng and is normalized. (B) Detection of SMARCAD1 (hHel1) mRNAontech) contained 2 mg of poly(A)1 RNA per lane. An SMARCAD1terials and Methods. Signal was visualized by autoradiography.
hHast500(ClMa
superfamily, based upon sequence conservation of the

ittp1ha
c
170 ADRA ET AL.
seven protein motifs previously described (Eisen et al.,1995). SNF2 proteins have been identified in manyspecies. Those for which functional data are availablehave been found to serve in a variety of cellular pro-cesses, including transcriptional regulation, DNA re-pair, and maintenance of mitotic chromosomal stabil-ity (Eisen et al., 1995).
Despite the conservation within the seven motifs,the phylogenetic analysis revealed the low degree ofsequence similarity between Etl-1, and by extensionhHel1, and other SNF2 family members outside of thiscore region. Moreover (see below), hHel1/Etl-1 isunique in that it contains two DEAD/H box motifs, aproperty not shared by any other member of the SNF2family. We propose that hHel1, Etl-1, and their homo-logues form a novel helicase subfamily characterizedby the duplication of this important functional motif.
It is an important finding that hHel1 contains twoDEA/GD/H motifs. Each is preceded by an ATP-bind-ing site with almost precisely the same length of se-quence separation. On the basis of sequence informa-tion, we cannot differentiate between the two motifs interms of function. This duplication has been reportedfor only one other molecule, the yeast PRP44 geneproduct. PRP44 is involved in pre-mRNA splicing. Kimand Rossi (1999) report that overexpression of the first,but not the second, helicase/ATP domain containingdominant negative mutations gave rise to an accumu-lation in unspliced mRNAs. Moreover, in vitro, only thefirst ATPase domain actually was active. It is not yetpossible to assess the relative contributions of the two
FIG. 4. Chromosomal localization of SMARCAD1 (human Hel1)hromosome 4 showing mapped location of human Hel1 gene (arr
chromosomes counterstained with DAPI. The two chromosomes 4 arbiotin-labeled SMARCAD1 (human Hel1) probe on both chromosomesite. Hybridization was observed with a Zeiss Axioscop microscope,(Applied Imaging, Pittsburgh, PA).
ATPase/DEAD/H box domains in hHel1, as we do not
know the catalytic activity of the enzyme. However,hHel1 is the first human member of a potentially im-portant new subclass of human helicases that containduplicate ATPase/DEAD/H box domains.
In addition to the NTP-binding sites, hHel1 containsconsensus sequences of several types known to be in-volved in protein–protein interactions. The homeoboxand leucine zipper domains indicate that hHel1 mayform oligomers or may mediate interactions with otherproteins involved in nucleic acid metabolism. Effort isbeing directed at identifying protein partners of hHel1to begin defining its cellular function.
We do not offer specific analysis of whether hHel1 isa DNA or RNA directed helicase. This is an ambiguousquestion since, at least in vitro, some RNA helicasescan unwind DNA molecules. In extensive BLASTsearches using the whole hHel1 sequence (not limitedto the seven conserved domains), significant homolo-gies to hHel1 were seen almost exclusively with SNF2family members. Many members of this family exam-ined thus far have demonstrated the ability to functionin transcriptional activation. Focusing on more specificcomponents of the sequence, the presence of the RNP-1(Bandziulis et al., 1989) signature motif would seem tondicate a role for hHel1 in RNA metabolism, althoughhe consensus sequence contains considerable varia-ion. However, RNP-1 motifs in known RNA-bindingroteins often are repeated (Swanson and Dreyfuss,988a,b), and this does not appear to be the situation inHel1. This will be the subject of detailed analyses thatre beyond the scope of this report.
ne by fluorescence in situ hybridization. (Left) Idiogram of humanat position 4q22–q23. (Right) Photograph of human metaphase
dicated by numbers. Arrows point to the site of hybridization of thein band q22–q23; both sister chromatids show hybridization at thatphotographs were prepared using the CytoVision Imaging System
geow)e ins 4and
The hHel1 molecule was cloned from a human brain
ad(utstwgFf
rgtqg
isaIetbDfhrneSfEt
wmtrnca1wwZamcqctgt1pwadlwl
Lpil
171SMARCAD1, A MEMBER OF THE HUMAN HELICASE FAMILY
cDNA library on the basis of its homology to the mu-rine Etl-1 protein. Given that hHel1 is the humanhomologue of Etl-1, it is important to consider theinvestigations of Etl-1. There is no published informa-tion on whether Etl-1 functions biochemically as a he-licase, although studies on Etl-1 have addressed itsexpression during murine embryogenesis (Soininen et
l., 1992). Etl-1 is nuclear, suggesting that it, at least,oes not function at the level of cytoplasmic mRNAsSchoor et al., 1993). During embryogenesis, Etl-1 ispregulated briefly during the first cell divisions andhen downregulated until a sustained burst of expres-ion is seen from the blastocyst stage onward. Addi-ionally, a murine knockout of Etl-1 has been produced,here a subtle phenotype, complicated by issues ofenetic background, was observed (Schoor et al., 1999).uture studies will be directed toward assigning a
unction to hHel1 in the human system.Interestingly, we have data that argue for a general
ole of hHel1 in driving transcriptional events and/orenomic rearrangements. We employed an assay sys-em based upon the transcriptional reactivation fre-uency of a silent genomically integrated reporter
FIG. 5. Differential display and Northern analysis of EN/NIH2-20 subclones. Parental cells were transfected, or not, with E1Ato generate E1A-expressing subclones. Randomly selected E1A-ex-pressing clones were assayed for reactivation frequency of a tran-scriptionally silent neomycin phosphotransferase gene. Reactivationfrequency is indicated as positive (1; 35 3 1026 events per cell) ornegative (2; no events after extended passage). (A) Differential dis-play. Differential analysis of EN/NIH 2-20 and E1A-expressing EN/NIH 2-20 subclones with varying capacities for neo reactivation wasperformed using RNAmap Kit A (GenHunter Corp.) as described in
iang and Pardee (1992). Arrow indicates the differentially ex-ressed band that was excised, amplified, subcloned, sequenced, anddentified as Etl-1/SMARCAD1 (hHel1). (B) Northern analysis. Eachane contains 20 mg total RNA. Ethidium bromide staining in bottom
panel documents comparable loading. Presence of Etl1/SMARCAD1(hHel1) difference product specific to neo reactivating, E1A express-ing clone is marked.
ene, neo (Drews et al., 1992, 1998). Human Hel1 was
dentified as a cDNA product whose presence was as-ociated with an increased frequency of transcriptionalctivation events driven by the adenovirus factor E1A.f the association is causal, three possibilities exist toxplain these data. First, hHel1 could be a specificranscription factor partner of E1A. Second, the possi-le function of hHel1 in transcriptional priming ofNA could lead to the observed enhancement in the
requency of transcriptional initiation events. Third,Hel1 could contribute to enhanced genomic rear-angement events that juxtapose the reporter gene,eo, downstream of transcriptionally active genomiclements. This last possibility is supported by ourouthern blot finding of novel neo-containing genomic
ragments, many associated with amplification, in1A-expressing EN/NIH 2-20 subclones that had reac-
ivated neo expression.Studies indicate that 4q22–q23, the region to whiche mapped human Hel1, contains fragile sites andutations linked to diseases involving a range of tissue
ypes. Deletions of this region have been shown toesult in a clinical syndrome characterized by centralervous system overgrowth (Nowaczyk et al., 1997). Alonal deletion involving the 4q23 region was found in
case of soft tissue leiomyosarcoma (Nilbert et al.,988). In two investigations, heptatocellular carcinomaas found to be associated with loss of heterozygosityithin the region 4q22–q23 (Buetow et al., 1989;hang et al., 1990). These studies indicate that annti-oncogene important to hepatocellular carcinomaight be located on 4q. Several of these studies indi-
ate the potential importance of the area around 4q22–23 to hematologic malignancy. After bone marrowells were sequentially treated with methotrexate andhymidine, they expressed a fragile site in 4q23 (Mor-an et al., 1988). Clinically, a chromosomal transloca-ion t(4;11)(q21;q23), resulting in the fusion of an1q23 gene and a 4q21 gene, was found to be highlyrevalent in infant acute leukemias and associatedith poor prognosis (Chen et al., 1993). Finally, anssociation has been found between aphidicolin-in-uced fragile sites in this region and acute myeloideukemia (Furuya et al., 1989). It remains to be seenhether an alteration in the human Hel1 gene under-
ies any of these diseases.
ACKNOWLEDGMENTS
This work was supported by Grant AI43663-01 from the NationalInstitute of Allergy and Infectious Diseases (C.N.A.), by National Insti-tutes of Health Physician Scientist Award AG-00294-09 (R.E.D.), andby the Adra family and the ADRA Institute. J.L.D. is a Fellow of SaoPaulo Foundation for Research Support (FAPESP). H.T. is an Interna-tional Prize Travelling Fellow of the Wellcome Trust. This work isdedicated to the memory of Professor Stephen Robinson.
REFERENCES
Adra, C. N., Iyengar, A. R., Syed, F. A., Kanaan, I. N., Rilo, H. L., Yu,W., Kheraj, R., Lin, S. R., Horiuchi, T., Khan, S., Weremowicz, S.,
Lim, B., Morton, C. C., and Higgs, D. R. (1998). Human ARHG-
A
B
B
B
C
C
C
D
D
E
E
F
G
H
H
I
K
L
L
L
L
M
N
N
P
P
S
S
S
S
S
T
v
172 ADRA ET AL.
DIG, a GDP-dissociation inhibitor for Rho proteins: Genomicstructure, sequence, expression analysis, and mapping to chromo-some 16p13.3. Genomics 53: 104–109.
Adra, C. N., Lelias, J-M., Kobayashi, H., Kaghad, M., Morrison, P.,Rowley, J., and Lim, B. (1994). Cloning of the cDNA for a hema-topoietic cell-specific protein related to CD20 and the b subunit ofthe high-affinity IgE receptor: Evidence for a family of proteinswith four membrane-spanning regions. Proc. Natl. Acad. Sci. USA91: 10178–10182.
Adra, C. N., Manor, D., Ko, J. L., Zhu, S., Horiuchi, T., Van Aelst, L.,Cerione, R., and Lim, B. (1997). RhoGDIg: A GDP-dissociationinhibitor for Rho proteins with preferential expression in brainand pancreas. Proc. Natl. Acad. Sci. USA 94: 4279–4284.dra, C. N., Zhu, S., Ko, J. L., Guillemot, J. C., Cuervo, A. M.,Kobayashi, H., Horiuchi, T., Lelias, J. M., Rowley, J. D., and Lim,B. (1996). LAPTM5: A novel lysosomal-associated multispanningmembrane protein preferentially expressed in hematopoietic cells.Genomics 35: 328–337.andziulis, R. J., Swanson, M. S., and Dreyfuss, G. (1989). RNA-binding proteins as developmental regulators. Genes Dev. 3: 431–437.ork, P., and Koonin, E. V. (1993). An expanding family of helicaseswithin the ’DEAD/H’ superfamily. Nucleic Acids Res. 21: 751–752.uetow, K. H., Murray, J. C., Israel, J. L., London, W. T., Smith, M.,Kew, M., Blanquet, V., Brechot, C., Redeker, A., and Govindara-jah, S. (1989). Loss of heterozygosity suggests tumor suppressorgene responsible for primary hepatocellular carcinoma. Proc. Natl.Acad. Sci. USA 86: 8852–8856.hen, C. S., Hilden, J. M., Frestedt, J., Domer, P. H., Moore, R.,Korsmeyer, S. J., and Kersey, J. H. (1993). The chromosome 4q21gene (AF-4/FEL) is widely expressed in normal tissues and showsbreakpoint diversity in t(4;11)(q21;q23) acute leukemia. Blood 82:1080–1085.oin, F., Bergmann, E., Tremeau-Bravard, A., and Egly, J. M.(1999). Mutations in XPB and XPD helicases found in xerodermapigmentosum patients impair the transcription function of TFIIH.EMBO J. 18: 1357–1366.oin, F., Marinoni, J. C., Rodolfo, C., Fribourg, S., Pedrini, A. M.,and Egly, J. M. (1998). Mutations in the XPD helicase generesult in XP and TTD phenotypes, preventing interaction be-tween XPD and the p44 subunit of TFIIH. Nat. Genet. 20:184 –188. [See comments]rews, R., Chan, VT-W., and Schnipper, L. E. (1992). Oncogenesresult in genomic alterations that activate a transcriptionally si-lent, dominantly selectable reporter gene (neo) Mol. Cell. Biol. 12:198–206.rews, R., Kolker, M., Moran, C., Sachar, D., Chan, V., andSchnipper, L. (1998). Genetic analysis of adenovirus E1A: In-duction of genetic instability and altered cell morphologic andgrowth characteristics are segregatable functions. Mutat. Res.412: 9 –25.isen, J. A., Sweder, K. S., and Hanawalt, P. C. (1995). Evolution ofthe SNF2 family of proteins: Subfamilies with distinct sequencesand functions. Nucleic Acids Res. 23: 2715–2723.llis, N. A. (1997). DNA helicases in inherited human disorders.Curr. Opin. Genet. Dev. 7: 354–363.
uruya, T., Ochi, H., and Watanabe, S. (1989). Common fragile sitesin chromosomes of bone marrow cells and peripheral blood lym-phocytes from healthy persons and leukemia patients. CancerGenet. Cytogenet. 43: 131–138.odbout, R., Packer, M., and Bie, W. (1998). Overexpression of aDEAD box protein (DDX1) in neuroblastoma and retinoblastomacell lines. J. Biol. Chem. 273: 21161–21168.eilek, G. M., and Peterson, M. G. (1997). A point mutation abolishesthe helicase but not the nucleoside triphosphatase activity of hep-
atitis C virus NS3 protein. J. Virol. 71: 6264–6266.uang, S., Li, B., Gray, M. D., Oshima, J., Mian, I. S., andCampisi, J. (1998). The premature ageing syndrome protein,WRN, is a 39 3 59 exonuclease. Nat. Genet. 20: 114 –116. [Let-ter]
ida, K., Matsumoto, S., and Yahara, I. (1992). The KKRKK sequenceis involved in heat shock-induced nuclear translocation of the18-kDa actin-binding protein, cofilin. Cell Struct. Funct. 17: 39–46.im, D. H., and Rossi, J. J. (1999). The first ATPase domain of theyeast 246-kDa protein is required for in vivo unwinding of theU4/U6 duplex. RNA 5: 959–971.
iang, P., and Pardee A. B. (1992) Differential display of eukaryoticmessenger RNA by means of the polymerase chain reaction. Sci-ence 257: 967–971.
ohman, T. M., and Bjornson, K. P. (1996). Mechanisms of heli-case-catalyzed DNA unwinding. Annu. Rev. Biochem. 65: 169 –214.
ohman, T. M., Thorn, K., and Vale, R. D. (1998). Staying on track:Common features of DNA helicases and microtubule motors. Cell93: 9–12.
uking, A., Stahl, U., and Schmidt, U. (1998). The protein familyof RNA helicases. Crit. Rev. Biochem. Mol. Biol. 33: 259 –296.organ, R., Morgan, S. S., Hecht, B. K., and Hecht, F. (1988). Fragilesites at 4q23 and 7q11.23 unique to bone marrow cells. CancerGenet. Cytogenet. 31: 47–53.ilbert, M., Mandahl, N., Heim, S., Rydholm, A., Willen, H., Aker-man, M., and Mitelman, F. (1988). Chromosome abnormalities inleiomyosarcomas. Cancer Genet. Cytogenet. 34: 209–218. [Pub-lished erratum appears in Cancer Genet. Cytogenet. 1988, Nov;36(1):141]owaczyk, M. J., Teshima, I. E., Siegel-Bartelt, J., and Clarke, J. T.(1997). Deletion 4q21/4q22 syndrome: Two patients with de novo4q21.3q23 and 4q13.2q23 deletions. Am. J. Med. Genet. 69: 400–405.
ause, A., Methot, N., and Sonenberg, N. (1993). The HRIGRXXRregion of the DEAD box RNA helicase eukaryotic translation ini-tiation factor 4A is required for RNA binding and ATP hydrolysis.Mol. Cell. Biol. 13: 6789–6798.
azin, M. J., and Kadonaga, J. T. (1997). SWI2/SNF2 and relatedproteins: ATP-driven motors that disrupt protein–DNA interac-tions? Cell 88: 737–740.
choor, M., Schuster-Gossler, K., and Gossler, A. (1993). The Etl-1gene encodes a nuclear protein differentially expressed duringearly mouse development. Dev. Dyn. 197: 227–237.
choor, M., Schuster-Gossler, K., Roopenian, D., and Gossler, A.(1999). Skeletal dysplasias, growth retardation, reduced postnatalsurvival, and impaired fertility in mice lacking the SNF2/SWI2family member ETL1. Mech. Dev. 85: 73–83.
oininen, R., Schoor, M., Henseling, U., Tepe, C., Kisters-Woike,B., Rossant, J., and Gossler, A. (1992). The mouse Enhancertrap locus 1 (Etl-1): A novel mammalian gene related to Dro-sophila and yeast transcriptional regulator genes. Mech. Dev.39: 111–123.
wanson, M. S., and Dreyfuss, G. (1988a). Classification andpurification of proteins of heterogeneous nuclear ribonucleopro-tein particles by RNA-binding specificities. Mol. Cell. Biol. 8:2237–2241.
wanson, M. S., and Dreyfuss, G. (1988b). RNA binding specificity ofhnRNP proteins: A subset bind to the 39 end of introns. EMBO J.7: 3519–3529.
roelstra, C., van Gool, A., de Wit, J., Vermeulen, W., Bootsma, D.,and Hoeijmakers, J. H. (1992). ERCC6, a member of a subfamily ofputative helicases, is involved in Cockayne’s syndrome and pref-erential repair of active genes. Cell 71: 939–953.
an Gool, A. J., Citterio, E., Rademakers, S., van Os, R., Ver-meulen, W., Constantinou, A., Egly, J. M., Bootsma, D., and
Hoeijmakers, J. H. (1997). The Cockayne syndrome B protein,
V
W
W
Z
173SMARCAD1, A MEMBER OF THE HUMAN HELICASE FAMILY
involved in transcription-coupled DNA repair, resides in anRNA polymerase II-containing complex. EMBO J. 16: 5955–5965.illard, L., Lossi, A. M., Cardoso, C., Proud, V., Chiaroni, P., Col-leaux, L., Schwartz, C., and Fontes, M. (1997). Determination ofthe genomic structure of the XNP/ATRX gene encoding a potentialzinc finger helicase. Genomics 43: 149–155.eeda, G., Eveno, E., Donker, I., Vermeulen, W., Chevallier-Lag-
ente, O., Taieb, A., Stary, A., Hoeijmakers, J. H., Mezzina, M., andSarasin, A. (1997). A mutation in the XPB/ERCC3 DNA repairtranscription gene, associated with trichothiodystrophy. Am. J.Hum. Genet. 60: 320–329.est, S. C. (1996). DNA helicases: New breeds of translocatingmotors and molecular pumps. Cell 86: 177–180.
hang, W. D., Hirohashi, S., Tsuda, H., Shimosato, Y., Yokota, J.,Terada, M., and Sugimura, T. (1990). Frequent loss of heterozy-gosity on chromosomes 16 and 4 in human hepatocellular carci-
noma. Jpn. J. Cancer Res. 81: 108–111.Copyright © 2022 FDOKUMEN