In situ forming, resorbable graft copolymer hydrogels providing controlled drug release
Upload
independentCategory
view
4download
0
Skin-Derived Stem Cells Transplanted into ResorbableGuides Provide Functional Nerve RegenerationAfter Sciatic Nerve ResectionC. MARCHESI,1 M. PLUDERI,2 F. COLLEONI,1,2 M. BELICCHI,1 M. MEREGALLI,1 A. FARINI,1 D. PAROLINI,1
L. DRAGHI,3 M. E. FRUGUGLIETTI,1 M. GAVINA,1 L. PORRETTI,4 A. CATTANEO,4 M. BATTISTELLI,1
A. PRELLE,1 M. MOGGIO,1 S. BORSA,2 L. BELLO,2 D. SPAGNOLI,2 S. M. GAINI,2 M. C. TANZI,3
N. BRESOLIN,1 N. GRIMOLDI,2 AND Y. TORRENTE1*1Fondazione IRCCS Ospedale Maggiore Policlinico-Mangiagalli e Regina Elena of Milan, Stem Cell Laboratory,Department of Neurological Sciences, Centro Dino Ferrari, University of Milan, Italy2Unit of Neurosurgery, Fondazione IRCCS Ospedale Maggiore Policlinico-Mangiagalli e Regina Elena of Milan, Italy3BioMatLab. Bioengineered Department, Politecnico di Milano, Milano, Italy4Centro Trasfusionale e di Immunologia dei Trapianti, Fondazione IRCCS Ospedale MaggiorePoliclinico-Mangiagalli e Regina Elena of Milan, Italy
KEY WORDSperipheral nerve; regeneration; skin-derived stem cells; arti-ficial graft
ABSTRACTThe regeneration in the peripheral nervous system is oftenincomplete and the treatment of severe lesions with nervetissue loss is primarily aimed at recreating nerve continuity.Guide tubes of various types, filled with Schwann cells, stemcells, or nerve growth factors are attractive as an alternativetherapy to nerve grafts. In this study, we evaluated whetherskin-derived stem cells (SDSCs) can improve peripheralnerve regeneration after transplantation into nerve guides.We compared peripheral nerve regeneration in adult ratswith sciatic nerve gaps of 16 mm after autologous transplan-tation of GFP-labeled SDSCs into two different types ofguides: a synthetic guide, obtained by dip coating with aL-lactide and trimethylene carbonate (PLA-TMC) copolymerand a collagen-based guide. The sciatic function index andthe recovery rates of the compound muscle action potentialwere significantly higher in the animals that receivedSDSCs transplantation, in particular, into the collagenguide, compared to the control guides filled only with PBS.For these guides the morphological and immunohistochemi-cal analysis demonstrated an increased number of myelin-ated axons expressing S100 and Neurofilament 70, suggest-ing the presence of regenerating nerve fibers along the gap.GFP positive cells were found around regenerating nervefibers and few of them were positive for the expression ofglial markers as S-100 and glial fibrillary acidic protein. RT-PCR analysis confirmed the expression of S100 and myelinbasic protein in the animals treated with the collagen guidefilled with SDSCs. These data support the hypothesis thatSDSCs could represent a tool for future cell therapy applica-tions in peripheral nerve regeneration. VVC 2006 Wiley-Liss, Inc.
INTRODUCTION
Despite the capacity for axonal regeneration in the pe-ripheral nervous system (PNS), nerve transection is char-acterized by poor functional recovery (Fawcett and Keynes,1990; Stoll and Muller, 1999). Several approaches have
been attempted to overcome this deficiency. Although au-tologous nerve grafts are the first choice strategy in recon-struction of peripheral nerve defects, they have severaldisadvantages caused by the limited availability of donortissue, loss of donor nerve function, and unsatisfactory re-covery for long gaps. An alternative to autograft is repre-sented by the use of nerve guidance channel, i.e. tubularstructures implanted to support the axonal regrowth to-ward the distal stump (Archibald et al., 1991; Belkaset al., 2004; Longo et al., 1984; Lundborg et al., 1982; Wil-liams et al., 1983). Advantages of tubulization repair are:prevention of fibrous entrapment of the injured nerve andreduced problems related to the correct alignment andsuture of nerve fascicles. Over recent years many materialsfor nerve guides, either biological or synthetic, have beeninvestigated with regard to biodegradation, immune bio-compatibility, and revascularization. Along with others,collagen and polylactide filaments have been reported par-ticularly useful for nerve regeneration (Evans et al., 1999,2000; Ngo et al., 2003; Widmer et al., 1998). In fact thesematerials are biodegradable and biocompatible and theirfibrillar structure permits the adhesion of residentSchwann cells allowing for their survival and proliferation(Ahmed et al., 2005; Archibald et al., 1991; Li et al., 1992).Because artificial tubular devices do not contain the cellu-lar elements of nerve stumps the regeneration tends tofail when tubes are used to bridge extended nerve defects(Archibald et al., 1991). To enhance the axonal regenera-tion in experimental models of peripheral nerve injurybioengineered conduits filled with neurotrophic factors or
Grant sponsors: The Association Francaise Contre les Myopathies, The ItalianMinistry of Health, The Centro Dino Ferrari, Department of Neurological Science,University of Milan, Italy.
*Correspondence to: Dr. Torrente Yvan, Department of Neurological Science, Uni-versity of Milan, Stem Cell Laboratory, Fondazione IRCCS Ospedale Maggiore Poli-clinico-Mangiagalli e Regina Elena of Milan, via Francesco Sforza 35, 20122 Milan,Italy. E-mail: [email protected]
Received 1 June 2006; Revised 13 November 2006; Accepted 16 November 2006
DOI 10.1002/glia.20470
Published online 3 January 2007 in Wiley InterScience (www.interscience.wiley.com).
GLIA 55:425–438 (2007)
VVC 2006 Wiley-Liss, Inc.

cells, such as Schwann cells, were proposed (Ansellinet al., 1997; Evans et al., 2002; Levi et al., 1994, 1997;Mosahebi et al., 2001). Moreover, neural stem cells (NSCs)were also reported to have contributed to the axonalregeneration through their differentiation into Schwanncells when injected into a conduit spanning a 15-mm gapin the rat sciatic nerve (Heine et al., 2004; Murakamiet al., 2003). However, the use of Schwann cells or NSCshas several ethical and clinical limitations including thereduced proliferative capacity of Schwann cells and noreadily accessible source of NSCs. Recently, stem cellsderived from non-neural tissue, such as mesenchymalstem cells (MSCs), have attracted interest because,although adult, their differentiation potential is remark-able (Pittenger et al., 1999; Prockop, 1997) and they areeasily accessible through aspiration of bone marrow. Sev-eral groups demonstrated the ability of MSCs to transdif-ferentiate in vitro into myelinating ‘‘Schwann cell-like’’cells which are capable of improving myelin formation andnerve regeneration in vivo after their transplantation intodifferent models of peripheral nerve injury (Dezawa et al.,2001; Hou et al., 2006; Keilhoff et al., 2006; Mimura et al.,2004; Tohill et al., 2004). However, there is controversyabout the capacity of the undifferentiated MSCs to pro-mote nerve regeneration in rat animal models with pe-ripheral nerve injury (Cuevas et al., 2002, 2004; Tohillet al., 2004). To extend these data we focused our attentionon an accessible source of multipotent stem cell populationderived from the dermis, called skin-derived stem cells(SDSCs). These cells were reported to be able to generateboth endothelial and neural derivatives in vitro and invivo (Belicchi et al., 2004; Fernandes et al., 2004; Tomaet al., 2001). This ability could be of great benefit in pro-viding nerve regeneration by an intrinsic microenviron-ment which enhances the migration of endogenous cellu-lar elements into the guides and promotes neoangiogen-esis. However, little is known about the effects of guidematerials on the behavior of these stem cells. Hence, thesize of the tube wall pores and their stability over timeseem to be important factors in determining stem cellcapacity to promote or inhibit regeneration. In this studywe evaluated, by means of functional and histologicalmethods, the axonal regeneration after resection of the ratsciatic nerve leaving a 16-mm gap repaired with biore-sorbable and durable tubes with different degrees of per-meability filled with SDSCs.
MATERIALS AND METHODSCell Culture
Cells were isolated from 2-day-old newborn Wistar rats.Skin tissues were dissected carefully and subsequentlyminced into smaller pieces using sharp razor blades. Speci-men were washed three times with essential balanced saltsolution and enzymatically dissociated by adding 0.2% col-lagenase type XI and 0.1% trypsin for 1 h at 37�C. Thecell extract was filtered with a 70 lm nylon mesh (DAKOglostrup, Denmark) and subsequently plated on non-coated flasks to offer the advantage to isolate different
populations of skin-derived cells. Preplate 1 (pp1) repre-sented a population of skin-derived cells that adhered inthe first 24 h after isolation and highly enriched of fibro-blasts. Preplate 2 (pp2), which represented a population ofskin-derived cells that did not adhere, was used as the tar-get population. The growth media was composed of DMEM/F-12 (1:1), 20% FBS, including HEPES buffer (5 mM),glucose (0.6%), sodium bicarbonate (3 mM), and glutamine(2 mM). A defined hormone and salt mixture composed ofinsulin (25 lg/mL), transferrin (100 lg/mL), progesterone(20 nM), putrescine (60 lM), and sodium selenite (30 nM)was also used. To the above medium, epidermal growthfactor (EGF) (20 ng/mL) and fibroblast growth factor(bFGF) (10 ng/mL) (murine recombinant; Chiron Corpora-tion, Emeryville, CA) were added as mitogens.
Self-Maintenance and In Vitro Differentiationof SDSCs
To test whether the skin-derived progenitors exhibitself-maintenance, single cells generated from the super-natant of the pp2 were plated into 96-well plates. Forplating single cells, the clusters of floating cells (spheres)were collected in vitro with a pipette, mechanically disso-ciated, and serially diluted to yield �1–2 cells per 10 lLaliquot. A 10 lL aliquot was added to each well of a 96-well plate containing 200 lL of EGF 1 bFGF-containingmedium. All wells that contained one viable cell 24 hlater were scored 8 days later for the presence of spheres.Under these conditions, secondary single spheres weretransferred to 500 lL eppendorf tubes containing 200 lLof medium, triturated 20–40 times, and replated as singlecells into a new 96-well plate. The plates were scored8 days later for the number of tertiary spheres derivedfrom a single secondary sphere. In experiments of differ-entiation, primary or secondary single isolated sphereswere plated on glass coverslips in individual wells of 24-well (1.0 mL/well) (Nunc, USA) culture dishes in DMEM/F-12 medium with the hormone and salt mixture andEGF 1 bFGF. Medium was not changed for the rest ofthe experiment and coverslips were processed 21–25 dayslater for indirect immunocytochemistry.
In Vitro Immunocytochemistry and FlowCytometry Characterization
To verify the expression of stem cells markers, cells in thesupernatant of pp2 were resuspended in PBS and incubatedwith anti-CD34PE (Pharmingen), anti-CDW90 (Thy-1)-fluo-rescein-isotiocyanate (FITC, Pharmingen), anti-CD45-FITC(Becton Dickinson Immunocytometry Systems, MountainView, CA). Isotype-matched mouse immunoglobulin servedas controls. After each incubation performed at 4�C for20 min, cells were washed in PBS containing 1% heat inac-tivated FCS and 0.1% sodiumazide. Finally, cells were ana-lyzed using a FACS Calibur flow cytometer and Cell Questsoftware (Becton Dickinson). Each analysis included atleast 5,000–10,000 events. A light-scatter gate was set up
426 MARCHESI ET AL.
GLIA DOI 10.1002/glia

to eliminate cell debris from analysis. By using isotype-controls for PE and FITC, gates for phenotypic analysis ofCD341 cells were set so that the lower left panel containedat least 98% of the total cells analyzed. pp2 Cells were alsotested with mouse monoclonal antibodies against the neu-rofilament-M protein (1:100; Chemicon), neuronal specificenolase (NSE) (1:2000; Polysciences), glial fibrillary acidicprotein (GFAP) (1:50) from Boehringer Mannheim (India-napolis, IN); b-tubulin type III (TuJ1; Sigma; 1:100) and tomicrotubule-associated protein (MAP-2ab isoform) (1:200)from Sigma; nestin (1:500; Pharmingen) and galactocereb-roside (Gal-C) from Promega (South San Fracisco, CA);anti p75NFGR polyclonal (1:500; Promega). For quantita-tive analysis, after immunostaining, coverslips were coun-terstained with DAPI (Sigma, 1:200). Indirect immuno-staining of pp2 cells was also done using monoclonal anti-bodies against the CD31 (1:100; Santa Cruz), VE-Cad(1:100; Santa Cruz), CD34 (1:50; Becton Dickinson, Moun-tain View, CA), fibronectin (1:150; Sigma), cytokeratin(1:150; Dako), S100, and vimentin (1:200; Santa Cruz).Indirect immunocytochemistry was carried out with spheresattached to glass coverslips, either immediately after plat-ing (for nestin) or after 21 days in vitro. Coverslips testedfor neuronal markers were fixed in 4% paraformaldehyde(in PBS, pH 7.2) for 30 min, followed by three washes(10 min each) in PBS, pH 7.2. Non-neuronal antibodieswere tested into coverslips fixed in ethanol 70% in PBS for1 min and permeabilized for 5 min with 0.5% Triton 100-Xin PBS. After washing with PBS, cells were incubatedwith appropriate secondary FITC/TRITC-conjugated IgGantibodies (1:100) for 1 h at room temperature and exam-ined by a fluorescence microscope (LEICA DMIR2) eq-uipped with LEICA Qfluoro software.
ELISA Assay
Supernatant of the rat SDSCs after 3 days of cultureand the tissues into the guides 7 and 90 days after thetransplantation were collected and tested in duplicate forneurotrophin dosage. Neurotrophin 4/5 (NT4/5) and brain-derived neurotrophic factor (BDNF) were measured usingcommercially available kits (Chemicon) according to themanufacturer instructions. Standard dilution curves andconcentrations within the samples were measured with aspectrophotometer (Celbio, Milan, Italy) at 450 nm. Signifi-cance of differences in the values was determined usingANOVA.
Nerve Guides
PLA-TMC guide
For the preparation of porous PLA-TMC nerve guides, a10% solution of PLA-TMC (Resomer� LT 706, BoehringerIngelheim) in chloroform was filled with a fine powder ofglucose, accurately sieved to about 30 lm particles. Theglucose, in amount equal to that of the copolymer, wasthoroughly suspended on a magnetic stirring plate. The
suspension was then used to dip coat 16G syringe needlesin a home-made dipper apparatus, consisting of a car-riage sliding in a Teflon lubricated vertical guide anddriven by a stepping motor connected to a pulley. Rota-tion speed and pulley spooling were dimensioned to ac-hieve very slow linear speed of extraction (�15 mm/min),to reduce the thickness variation along the guide. Dip-ping process was performed in a closed chamber to bothreduce solvent evaporation speed and control humidity.After drying, the conduits were gently removed from theneedles and cut to length by removing both their ends, toobtain guides with uniform thickness. Water extractionfor 48 h at 40�C was sufficient to free the samples fromsugar particles. Nerve guides prepared were sputter-coated with gold–palladium and observed by scanningelectron microscopy (SEM) with an accelerating voltageof 10 kV. Diameter reduction under compressive loadswas analyzed using as load the maximum in vivo loadestimated by Belkas et al. 2005 on 12-mm long PHEMA-MMA nerve guides implanted to bridge an 8 mm gap onrat sciatic nerve. Chemical structure of the guide isshown in the scheme below:
Collagen guide
Collagen nerve guides were fabricated (NeuraGen, Inte-gra Lifesciences, Plainsboro, NJ) from highly purifiedType 1 collagen (<0.2% hexosamine and <0.1% trichloro-acetic acid-insoluble residues, with four or fewer tyrosineper molecule), derived from bovine deep flexor tendon.The construction and physical properties of this guidesused have been described earlier (Archibald et al., 1995;Li et al., 1992).
Grafting Procedure
We used male Wistar rats weighing 150–200 g, aged2 months. Animals were kept in our animal facilities understandard housing conditions (22�C 6 2�C, 65% humidity,artificial lights from 06.00 to 20.00 h). A standard dry dietand water were available ad libitum. All experimental pro-tocols were performed in accordance with the Italianguidelines for laboratory animals which conform with theEuropean Union guidelines. All procedures were per-formed using aseptic techniques and with the aid of anoperating microscope. Anesthesia was induced with an in-traperitoneal injection of Cloralium Hydrates (2 mL at6.25%). The animals were kept in prone position and a lin-ear skin incision on the right posterolateral surface of thethigh was performed. The right sciatic nerve was exposed
427TRANSPLANTATION OF SKIN-DERIVED STEM CELLS
GLIA DOI 10.1002/glia

deep to the biceps femoris muscle, and it was microsurgi-cally transected. A segment of the nerve was excised andthe appropriate 20-mm long tube guide was put at thelevel of the resulting gap. To leave a 16 mm gap, the proxi-mal and the distal nerve stumps were inserted into thelumen of the tube (inner diameter of 1 mm) at a distance of2 mm from each end, and kept in place without suturesbut only with fibrin glue to reduce nervous tissue damageand scar. At the end of the procedure, muscle incision wasreapproximated with 3-0 Polysorb sutures, and skin withinterrupted 3-0 silk sutures. Animals were randomly di-vided into four groups according to the guides used forrepair: PLA-TMC guide (Resomer�, Boehringer Ingelheim)filled with SDSCs (n 5 30) or PBS (n 5 20); Type 1 col-lagen guide (NeuraGen, Integra Neurosciences) filled withSDSCs (n 5 30) or PBS (n 5 20). Rat SDSCs were isolatedat second pp2 passage and transduced with the vesicularstomatitis virus glycoprotein pseudotyped human lenti-viral vector expressing the enhanced green fluorescentprotein (eGFP). The vector was produced by cotransfectionof NIH 3T3 cells with three plasmids. The packaging plas-mid (pCMVDR8.74) provides gag, pol, tat, and rev helpervector proteins driven by the human cytomegalovirus pro-moter. The envelope plasmid (pMD.G) encodes the heterol-ogous VSV/G. The reporter eGFP gene expression is drivenby the internal human phosphoglycerate kinase promoterand the post-transcriptional cis-acting regulatory elementof the woodchuck hepatitis virus is inserted in the senseorientation to increase the transgene expression (Follenziet al., 2000). The central polypurine tract is associatedwith the central termination site to enhance nuclear trans-location (Zennou et al., 1999; Zufferey et al., 1999). Lenti-viral vector preparations were purified and concentratedfrom the conditioned media of transfected cells by ultra-centrifugation. Lentiviral vector stocks were titered onHT1080 cells and generally exceeded 5 3 109 infectiousunits (IU)/mL. About 1 3 106 cells were suspended in15 lL of PBS and transplanted into the lumen of the tubeswith an Hamilton syringe in treated animals. Control ani-mals received 15 lL of PBS injected into the tubes in thesame manner.
Functional Assessment
Walking track analysis
The degree of recovery was monitored by the assessmentof walking patterns in the hind limb to obtain a sciaticfunction index (SFI), according to the method of De Medi-naceli et al. 1982. Paw prints were recorded by paintingrats’ hind paws with black ink and having them walkalong a narrow corridor lined with white paper. To obtainmeasurable and reproducible prints all animals werewalked multiple times before surgery and cell implanta-tion. Paw prints were measured for both operated and non-operated sides. Three different parameters of the rodent’spaw print were measured to determine the SFI with thefollowing formula 2 38.3 (PLF) 1 109.5 (TSF)1 13.3 (ITF)2 8.8 [PLF 5 print length function 5 (experimental PL 2
normal PL)/normal PL, TSF 5 toe spread function 5 (ex-perimental TS 2 normal TS)/normal TS (1st–5th toe), ITF5 intermedian toe spread function 5 (experimental IT 2normal IT)/normal IT (2nd–4th toe)]. A value of 2100represents total impairment. The evaluation was donebefore surgery, 7 days after injury, then once a week until30 days and subsequently every 15 days until the end ofthe study and was expressed as a percentage of SFI fol-lowing a previously reported method.
Electrophysiological study: Measurementof CMAPS
Nerve conduction studies were performed 90 days afterthe operation in all the animals before sacrifice. After theinduction of anesthesia the right sciatic nerve (trans-planted side) was re-exposed and directly stimulated withsingle electrical pulses (200 ms duration and supramaxi-mal intensity) delivered by monopolar needles placed atthe sciatic notch at the proximal stump, 2 mm before itsinsertion into the nerve guide. The compound muscleaction potentials (CMAP) were recorded both in the con-trols and SDSCs-transplanted animals by a recording nee-dle electrode placed on the gastrocnemius muscle throughpercutaneous puncture in the distal third of the paw, at adistance of about 3 cm from the stimulation electrode.
The CMAP were amplified (Viking 4 System, Nicolet,Madison, WI) and recorded on an ink-jet writer via a digitalmemory store. The peak to peak amplitude of the CMAPwas measured, and the latency of the response was mea-sured to the first deflection from the baseline. Currentwas gradually increased until a response was recorded,but in absence of recordable CMAP for an 8 mA stimulus,the presence of muscle reinnervation was excluded. Nerveconduction velocity recordings were carried out in vivo byplacing the recording electrodes in the plantar muscles ofthe foot/gastrocnemius muscle and stimulating the sciaticnerve proximally and distally to the nerve guide using amonopolar stimulating electrode under general anesthe-sia. The CMAP obtained by stimulating the proximal anddistal sites of the nerve guides were then used to calculatethe latency time, i.e., from stimulus to the onset of thefirst negative deflection. The distance between the stimu-lation sites from the proximal and distal ends was meas-ured. The unoperated right side was also evaluated ascontrol.
Morphological Analysis
For quantitative studies, specimens were fixed in 4%glutaraldehyde in phosphate buffer for 24 h. After dissec-tion and postfixation in 2% osmium tetroxide for 1 h thespecimens were dehydrated and embedded in Spurr resin.Morphometric analysis was performed on 0.5 lm semithinsections stained with toluidine blue, using a Leica lightmicroscope computer assisted. For study of the myelinatedfibers, the endoneural space was observed under an oil-immersion lens (1,0003). The number of axons and theirdiameter were calculated using the public domain NIH
428 MARCHESI ET AL.
GLIA DOI 10.1002/glia

Image program (developed at the National Institute ofHealt, Bethesda, MD).
Tissue Immunohistochemical Analysis
Immunofluorescent stainings were done at differenttime points: 7, 40, or 90 days after surgery. Artificialgrafts were removed, frozen in liquid nitrogen-cooled iso-pentane, stored at 280�C until sectioning. Both longitu-dinal and transverse section of 10 lm thickness were cuton a cryostat.
For immunostaining of Schwann cell markers and ba-sal lamina, sections were fixed in 4% paraformaldehydefor 4 min, then incubated with anti-S-100 (Chemicon),antiglial fibrillary acidic protein (GFAP-cy3 coniugated,SIGMA Saint Louis, Missouri) anti-laminin (Chemicon)antibodies for 90 min at room temperature. For myelindetection was used an anti-myelin basic protein (MBP,Chemicon) antibody with overnight incubation at 4�C af-ter fixation with methanol for 3 min. For endothelial im-munostaining sections were fixed in ethanol 80% in PBSfor 5 min and incubated with anti-alpha smooth actin(SMA, SIGMA) and anti-CD31 (Santa Cruz Biotechnol-ogy, Santa Cruz, CA) for 90 min at room temperature. Wealso evaluated the expression of nestin (marker of neuralprecursors). Sections were then washed with PBS andincubated with secondary antibodies (Alexa Fluor 488 or594-conjugated anti-mouse IgG or anti-rabbit IgG, 1:100,Molecular Probes). Cell nuclei were stained for 5 min atroom temperature with DAPI. Grafted cells were detectedby immunofluorescence assays for GFP positive cells withpolyclonal anti-GFP 594 (1:50 Molecular Probes). To ob-tain an estimate of S100, CD31, and SMA positive cellsten random fields were selected, photographed, and foreach field the total number of cells positive for glial or en-dothelial markers was determined.
Reverse Transcription-PolymeraseChain Reaction
Total RNA was isolated from rat sciatic nerve frozensections (about 30–40 frozen sections/sample) using Trizol
Reagent as described by the manufacturer’s protocol(Invitrogen, Life Technologies). Aliquots of total RNA (2lg) were used to analyze rat regeneration markers bymeans of RT-PCR technique and the primers used aresummarized in Table 1. First strand cDNA was preparedby using SuperScript First Strand Synthesis System IIIfor RT-PCR (Invitrogen, Life Technologies) with oligo(dT)12–18 priming. Polymerase chain reaction (PCR) wasperformed in a reaction mixture containing 1.5 mMMgCl2, 200 lM dNTPs, 2.5 unit of Platinum Taq DNAPolymerase (Invitrogen, Life Technologies),10 pmol of 50
and 30 primers. PCR was performed under the followingconditions: 94�C, 5 min; then 35 cycles at 92�C, 1 min;62�C, 1 min; 72�C, 1 min; and the last cycle 72�C, 5 min.
Statistical Analysis
The experimental data are expressed as the mean 6SD. Statistical analysis was performed by using a two-tailed Student t-test. One-way ANOVA was used only toevaluate differences in neuotrophins expression.
RESULTSEvaluation of Rat SDSC Cultures
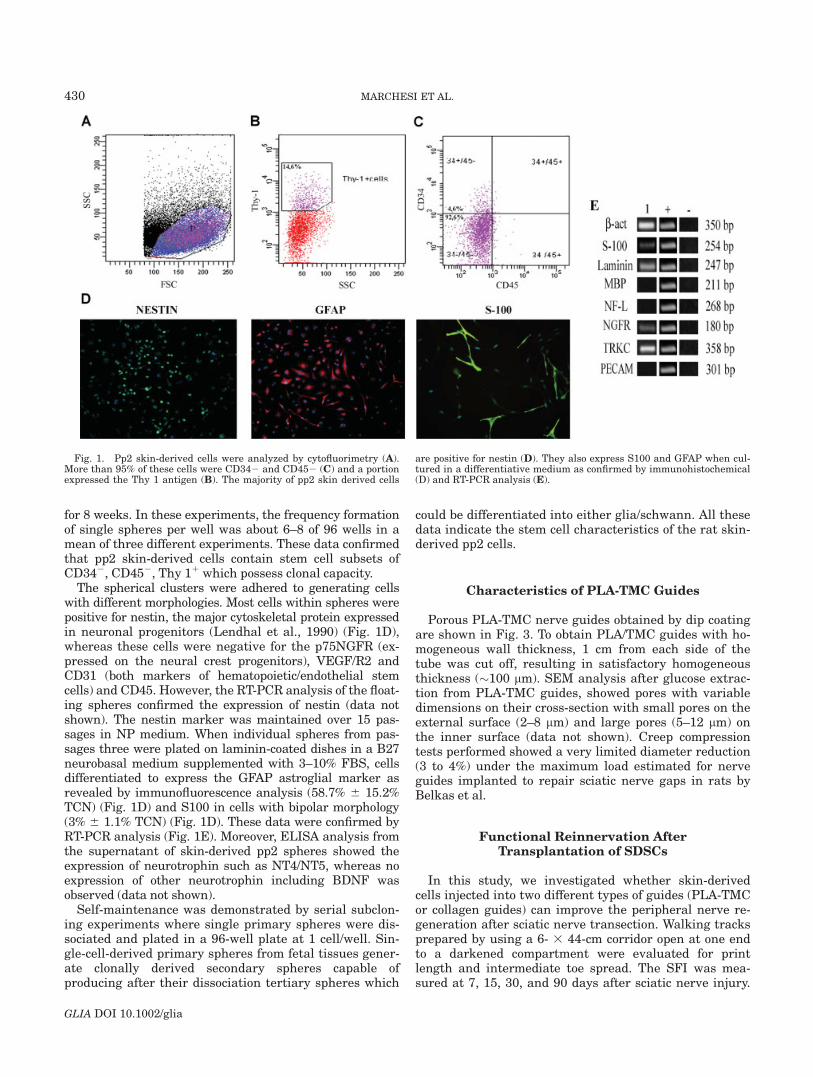
After passage two (pp2) from the total skin-derivedcells, the average of CD34 positive cells represented lessthen 5% as determined by flow cytometry. A portion ofpp2 skin-derived cells coexpressed Thy-1 antigen (5%)(Fig. 1A–C). Moreover, more than 95% of the selected pp2skin-derived cells have lineage negative phenotype (CD332,CD382, CD452). These findings suggest that the pp2population from the skin did not contain differentiatedblood cells.
The pp2 skin-derived cells isolated from adult rat skinwere plated in a growth medium containing EGF and bFGF.After 7 days in vitro, floating clusters of cells with morpho-logical resemblance to spheres were seen as previously de-scribed (Toma et al., 2001). To test the clonality of single pp2derived cells, the spheres were dissociated to single cells,plated into wells of a 96-well plate (1 cell/well) and cultured
TABLE 1. Primers for Markers of Nerve Regeneration
Gene SymbolPCR
fragments Primers sequence
Calcium-binding proteinm-RNA
S-100 254 bp 50-CAA GGG GAA GGA GAT GCC C-3050-CGC CTG AAA TTT GCA CCC ACA C-30
Myelin basic protein MBP 211 bp 50-CGA CGC GCA TCT TGT TAA TCC-3050-CTG TGC TGT CTG GAT GAA GCC-30
Light molecular-weightneurofilament
NF-L 268 bp 50-CTG CTA AGA AGA AAG ATT GAG CC-3050-CTG AAC TCA TAA GCA TGG ACC-30
Laminin chain 247 bp 50-GAC CGC TTT GTT CTG TAC ATG-3050-GGG CCA CTG TGT CCC-30
Nerve growth factorreceptor
NGFR 180 bp 50-GCA ATT CCC AGA CAC CTG G-3050-GAG TTC ATA GTC ACC AGC TGG-30
b–actin b–act 350 bp 50-TGG CAC CAC ACC TTC TAC AAT GAG-3050-CCG TGG TGG TGA AGC TGT AGC C-30
Neurotrophic turosinekinase receptor, type 3
TRKC 358 bp 50-CCT CCT GAA AGC ATC ATG TAC C-3050-CAG GGA GAG AGA GGG CAA C-30
Platelet/endothelial cellsadhesion molecule
PECAM 301 bp 50-CTC AAG CCG GCA ATG GC-3050-CAA AAT GGA GGC ATC TCC C-30
429TRANSPLANTATION OF SKIN-DERIVED STEM CELLS
GLIA DOI 10.1002/glia
for 8 weeks. In these experiments, the frequency formationof single spheres per well was about 6–8 of 96 wells in amean of three different experiments. These data confirmedthat pp2 skin-derived cells contain stem cell subsets ofCD342, CD452, Thy 11 which possess clonal capacity.
The spherical clusters were adhered to generating cellswith different morphologies. Most cells within spheres werepositive for nestin, the major cytoskeletal protein expressedin neuronal progenitors (Lendhal et al., 1990) (Fig. 1D),whereas these cells were negative for the p75NGFR (ex-pressed on the neural crest progenitors), VEGF/R2 andCD31 (both markers of hematopoietic/endothelial stemcells) and CD45. However, the RT-PCR analysis of the float-ing spheres confirmed the expression of nestin (data notshown). The nestin marker was maintained over 15 pas-sages in NP medium. When individual spheres from pas-sages three were plated on laminin-coated dishes in a B27neurobasal medium supplemented with 3–10% FBS, cellsdifferentiated to express the GFAP astroglial marker asrevealed by immunofluorescence analysis (58.7% 6 15.2%TCN) (Fig. 1D) and S100 in cells with bipolar morphology(3% 6 1.1% TCN) (Fig. 1D). These data were confirmed byRT-PCR analysis (Fig. 1E). Moreover, ELISA analysis fromthe supernatant of skin-derived pp2 spheres showed theexpression of neurotrophin such as NT4/NT5, whereas noexpression of other neurotrophin including BDNF wasobserved (data not shown).
Self-maintenance was demonstrated by serial subclon-ing experiments where single primary spheres were dis-sociated and plated in a 96-well plate at 1 cell/well. Sin-gle-cell-derived primary spheres from fetal tissues gener-ate clonally derived secondary spheres capable ofproducing after their dissociation tertiary spheres which
could be differentiated into either glia/schwann. All thesedata indicate the stem cell characteristics of the rat skin-derived pp2 cells.
Characteristics of PLA-TMC Guides
Porous PLA-TMC nerve guides obtained by dip coatingare shown in Fig. 3. To obtain PLA/TMC guides with ho-mogeneous wall thickness, 1 cm from each side of thetube was cut off, resulting in satisfactory homogeneousthickness (�100 lm). SEM analysis after glucose extrac-tion from PLA-TMC guides, showed pores with variabledimensions on their cross-section with small pores on theexternal surface (2–8 lm) and large pores (5–12 lm) onthe inner surface (data not shown). Creep compressiontests performed showed a very limited diameter reduction(3 to 4%) under the maximum load estimated for nerveguides implanted to repair sciatic nerve gaps in rats byBelkas et al.
Functional Reinnervation AfterTransplantation of SDSCs
In this study, we investigated whether skin-derivedcells injected into two different types of guides (PLA-TMCor collagen guides) can improve the peripheral nerve re-generation after sciatic nerve transection. Walking tracksprepared by using a 6- 3 44-cm corridor open at one endto a darkened compartment were evaluated for printlength and intermediate toe spread. The SFI was mea-sured at 7, 15, 30, and 90 days after sciatic nerve injury.
Fig. 1. Pp2 skin-derived cells were analyzed by cytofluorimetry (A).More than 95% of these cells were CD342 and CD452 (C) and a portionexpressed the Thy 1 antigen (B). The majority of pp2 skin derived cells
are positive for nestin (D). They also express S100 and GFAP when cul-tured in a differentiative medium as confirmed by immunohistochemical(D) and RT-PCR analysis (E).
430 MARCHESI ET AL.
GLIA DOI 10.1002/glia
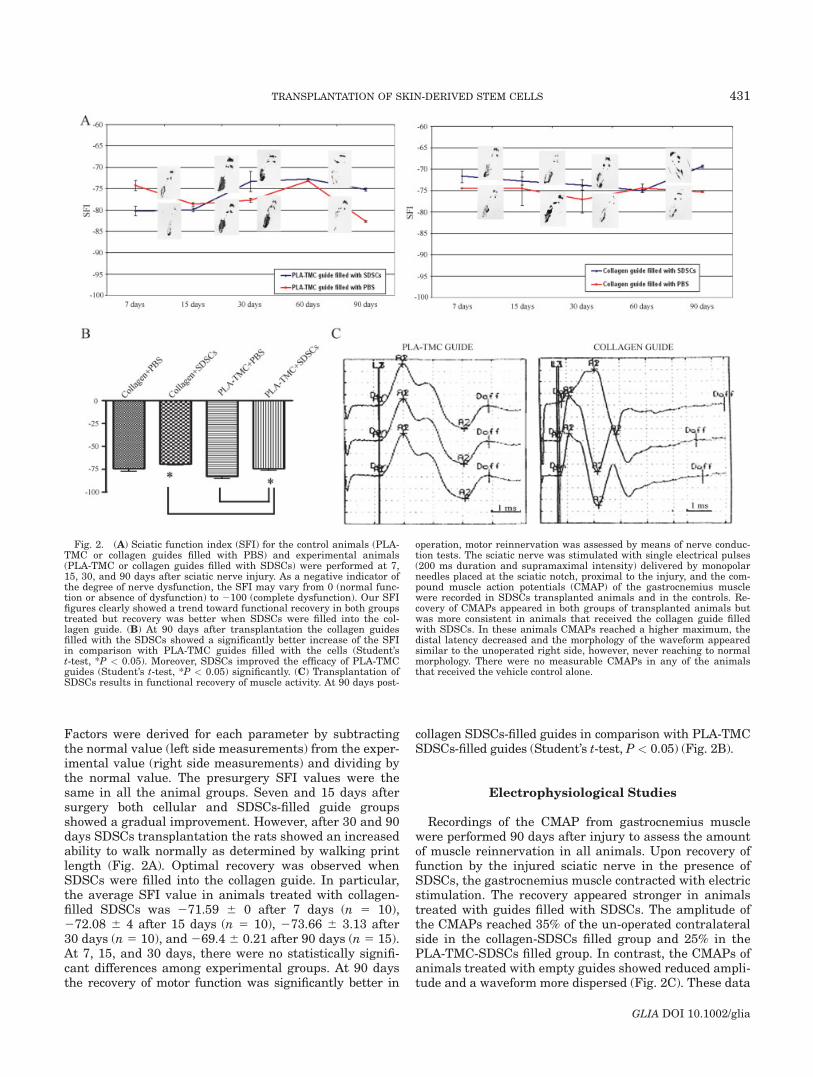
Factors were derived for each parameter by subtractingthe normal value (left side measurements) from the exper-imental value (right side measurements) and dividing bythe normal value. The presurgery SFI values were thesame in all the animal groups. Seven and 15 days aftersurgery both cellular and SDSCs-filled guide groupsshowed a gradual improvement. However, after 30 and 90days SDSCs transplantation the rats showed an increasedability to walk normally as determined by walking printlength (Fig. 2A). Optimal recovery was observed whenSDSCs were filled into the collagen guide. In particular,the average SFI value in animals treated with collagen-filled SDSCs was 271.59 6 0 after 7 days (n 5 10),272.08 6 4 after 15 days (n 5 10), 273.66 6 3.13 after30 days (n 5 10), and 269.4 6 0.21 after 90 days (n 5 15).At 7, 15, and 30 days, there were no statistically signifi-cant differences among experimental groups. At 90 daysthe recovery of motor function was significantly better in
collagen SDSCs-filled guides in comparison with PLA-TMCSDSCs-filled guides (Student’s t-test, P < 0.05) (Fig. 2B).
Electrophysiological Studies
Recordings of the CMAP from gastrocnemius musclewere performed 90 days after injury to assess the amountof muscle reinnervation in all animals. Upon recovery offunction by the injured sciatic nerve in the presence ofSDSCs, the gastrocnemius muscle contracted with electricstimulation. The recovery appeared stronger in animalstreated with guides filled with SDSCs. The amplitude ofthe CMAPs reached 35% of the un-operated contralateralside in the collagen-SDSCs filled group and 25% in thePLA-TMC-SDSCs filled group. In contrast, the CMAPs ofanimals treated with empty guides showed reduced ampli-tude and a waveform more dispersed (Fig. 2C). These data
Fig. 2. (A) Sciatic function index (SFI) for the control animals (PLA-TMC or collagen guides filled with PBS) and experimental animals(PLA-TMC or collagen guides filled with SDSCs) were performed at 7,15, 30, and 90 days after sciatic nerve injury. As a negative indicator ofthe degree of nerve dysfunction, the SFI may vary from 0 (normal func-tion or absence of dysfunction) to 2100 (complete dysfunction). Our SFIfigures clearly showed a trend toward functional recovery in both groupstreated but recovery was better when SDSCs were filled into the col-lagen guide. (B) At 90 days after transplantation the collagen guidesfilled with the SDSCs showed a significantly better increase of the SFIin comparison with PLA-TMC guides filled with the cells (Student’st-test, *P < 0.05). Moreover, SDSCs improved the efficacy of PLA-TMCguides (Student’s t-test, *P < 0.05) significantly. (C) Transplantation ofSDSCs results in functional recovery of muscle activity. At 90 days post-
operation, motor reinnervation was assessed by means of nerve conduc-tion tests. The sciatic nerve was stimulated with single electrical pulses(200 ms duration and supramaximal intensity) delivered by monopolarneedles placed at the sciatic notch, proximal to the injury, and the com-pound muscle action potentials (CMAP) of the gastrocnemius musclewere recorded in SDSCs transplanted animals and in the controls. Re-covery of CMAPs appeared in both groups of transplanted animals butwas more consistent in animals that received the collagen guide filledwith SDSCs. In these animals CMAPs reached a higher maximum, thedistal latency decreased and the morphology of the waveform appearedsimilar to the unoperated right side, however, never reaching to normalmorphology. There were no measurable CMAPs in any of the animalsthat received the vehicle control alone.
431TRANSPLANTATION OF SKIN-DERIVED STEM CELLS
GLIA DOI 10.1002/glia

are consistent with the formation of newly regeneratedthinly myelinated axons and suggest that SDSCs trans-plantation stimulated recovery of the sciatic nerve, enablinggastrocnemius muscle contraction. Three months after, theconduction velocity was much slower in all the experimen-tal groups than the normal control sides (22.66 6 4.89 m/s)and no significant differences were observed among theSDSCs-filled guides. In fact, the mean value observed inthe SDSCs filled collagen and PLA-TMC guides groupswas 16.516 2.19 and 14.716 2.91, respectively.
Gross Findings
At 9 and 12 weeks after surgical procedure, the nervetube was still present in every examined animal. At the
moment of the sacrifice, neither prosthesis displacementsnor septic collections were found in the surgical field. Theproximal and distal segments of the sciatic nerve appearedtightly connected to the nerve guide (Figs. 3E,F). Thefibrin glue injected during the previous surgical procedurewas partially reabsorbed. A thin fibrous reaction envelopingthe channel guide was evident. The scar tissue was slightlythicker in animals implanted with the polymeric PLA-TMCtube than in collagen tube groups. The fibrous tissue waseasily separable from the nerve guide through microsurgi-cal blunt dissection. After opening the nerve tube, a whitishparenchymatous tissue macroscopically connected to bothproximal and distal nerve stumps was found. In animalsimplanted with SDSCs, we noticed a lesser amount of fi-brous proliferation around the nerve stumps and a more
Fig. 3. Synthetic nerve guides wereproduced by copolymerization of L-lactideand trimethylenecarbonate. Scanningelectron microscopy images of cross sec-tions of PLA-TMC nerve guides (scalebar 5 100 lm) (A). The wall of the nerveguide is semi-permeable to allow for nu-trient exchange (scale bar 5 10 lm) (B).In this study we made a long distancegap in the sciatic nerve of adult maleWistar rats and transplanted GFP1SDSCs. The GFP lentiviral constructmap is shown in C. After injection, theGFP1 cells can be seen within the midportion of the guides clustered near theguide wall (D). Right sciatic nerve wasexposed before transection and thenrepaired with a permeable PLA-TMCguide (E) or type 1 collagen guide (F).Most GFP1 SDSCs conserved an undif-ferentiated morphology and did not cola-bel with S-100 glial marker (G).
432 MARCHESI ET AL.
GLIA DOI 10.1002/glia

stable and firm connection between distal and proximalnerve segments and the parenchymatous tissue inside theguide lumen. No tumorigenesis was found at the implanta-tion site or along the sciatic nerve segments.
Sciatic Nerve Regeneration Induced byTransplantation of SDSCs
After 90 days of sciatic nerve injury the H&E-stainednerve sections showed a complete connexion betweenproximal and distal stumps in empty and SDSCs filledPLA-TMC and collagen guides. Nevertheless, the tissueappeared more stable and more consistently connected inanimals treated with SDSCs. All the groups underwentstandard morphometric analysis of the total number ofmyelinated fibers and fiber diameter to gain informationabout the stage of axonal regeneration. Remyelination,axonal degeneration, as well as regeneration, were pres-ent in all specimens without statistically significant dif-ferences. Thickening and fibrosis of both endoneuriumand perinerium associated with an increase in endoneur-
ial collagen was present in all specimens (Fig. 4). In bothPLA-TMC and collagen guides morphometric analysis ofsemithin sections showed a severe loss of myelinatedfibers in both SDSCs-transplanted (Figs. 4A,D,G,I) andnot transplanted (Figs. 4B,E,H,M) animals. The decreaseaffected all types of fibers, but predominantly the largestones, at both 6 and 12 weeks. Compared to the emptyguide controls, the mean number of myelinated axonswas significantly higher in the SDSCs-filled collagenguides (3835 6 539 vs. 2779 6 312; Student’s t-test P <0.05) and in the SDSCs-filled PLA-TMC guides (3361 6301 vs. 2361 6 146; Student’s t-test P < 0.05) (Fig. 4).These data suggest that SDSCs confer a beneficial effecton axonal regeneration with no significant differencesbetween collagen and PLA-TMC filled guides (Fig. 4). Toextend these observations, the average axon diameters ofall experimental groups was calculated and compared tothe controlateral normal nerves (6.7 lm). In particular,the mean fiber diameter was 3.3 6 0.81 lm and 2.8 60.48 lm, respectively in SDSCs-filled collagen and PLA-TMC guides.
Fig. 4. Morphological analysis of axonal regeneration in adult ratsciatic nerve. Toluidine blue stain, 3400. At 6 and 12 weeks, SDSCstransplanted animals show numerous regenerated axons (A, D, G, L)with both collagen and PLA-TMC guides; in nontransplanted animalsthere are few regenerated axons and a lower density of myelinated fibers(B, E, H, M). The controlateral normal sciatic nerve is shown in C, F, I,and N. The average and the standard deviation were calculated in three
parts of the same nerve (proximal, central, and distal). The average andthe standard deviation of controlateral nerves were calculated in onepart of the nerve. The bar-graph (chart) shows an increased density ofmyelinated fibers in nerves of SDCSs-transplanted animals compared tonerves of no SDSCs-transplanted animals, except for PLA-TMC guidetreated rats at 12 weeks.
433TRANSPLANTATION OF SKIN-DERIVED STEM CELLS
GLIA DOI 10.1002/glia

Characterization of Transplanted GFP1 SDSCs
Immunohistochemical analysis of the conduits contain-ing SDSCs identified transplanted cells due to their expres-sion of GFP (Figs. 3D,G). At the proximal end of both col-lagen and PLA-TMC guides, GFP1 SDSCs were found tobe adherent to intraluminal nerve fibers (Fig. 3D). To verifythe involvement of SDSCs with axonal regeneration wequantified the fate of transplanted GFP positive cells bycolabeling for glial markers (GFAP and S-100). After7 days of nerve injury we observed several clusters of liv-ing cells within the nerve guides (data not shown). In bothPLA-TMC and collagen guides transplanted cells weredetected at various sites along the tube, in particular, inthe distal and mid portion, demonstrating migration fromthe grafting site. The total number of GFP-labeled cells at2 months after transplantation was about 25–38% oftotal injected SDSCs. Most of the cells conserved an un-differentiated morphology and did not colabel with glialmarkers (Fig. 3G). Only a small percentage colabeledwith S-100 (4.5%) or GFAP (6.1%) suggesting glial dif-ferentiation (data not shown). However, in PLA-TMCconduits we found a small number of transplanted cellscoexpressing GFP and S100 at the distal ends while ahigher percentage of the GFP-positive cells colabeledwith GFAP were found inside the distal ends of collagenguides. In both guides the GFP-expressing Schwanncells formed many small myelin sheaths which sur-rounded GFP negative host neurofilament positiveaxons. All these data suggested an effect of local cellularand humoral factors on SDSCs differentiation.
S-100 and GFAP Expressionin Regenerating Nerves
The regenerating tissue at the proximal stump showedseveral endogenous GFP negative Schwann cells expres-sing GFAP and S100 in all animals. Ninety days aftertransplantation a progressive increasing expression ofneurofilament, S100, and GFAP indicating a nerve fiberregrowth into the gap was observed only in animals thatreceived guides filled with SDSCs (Fig. 5A). The meannumber of regenerated S-100 positive axons in the col-lagen SDSCs-transplanted group was significantly higherthan in PLA-TMC SDSCs-transplanted animals (1125.5 6345 vs. 708.5 6 109; Student’s t-test, P < 0.05) (Fig. 5C).However, the number of CD31 or SMA positive vesselswas similar in all groups of animals (Fig. 5C). In the trea-ted animals the increased expression of S 100 and MBPwas also confirmed by RT-PCR analysis (Fig. 5D).
Neurotrophin’s Expressionin Regenerating Nerves
To evaluate whether the better regeneration observedin the transplanted animals was related to the release ofneurotrophins from SDSCs, we performed ELISA analysisfor BDNF and NT4/5 in the transplanted tissues 7 and
90 days after transaction. In transacted sciatic nerves re-ceiving the PLA-TMC guides we found no significantdifferences in the expression of BDNF or NT4/5 in com-parison with collagen guides after 7 or 90 days of transac-tion (Table 2).
DISCUSSIONDamage and Regeneration of Peripheral Nerves
After injury to a peripheral nerve, axonal elongationmay occur by spontaneous regenerative capacity (Fu andGordon, 1997) but functional recovery is often subopti-mal. Bioengineered grafts are a promising method fornerve repair. A variety of conduits have been producedfor bridging nerve gaps where both synthetic and naturalmaterials have been used. Literature shows that materi-als with the highest regenerative activity are collagenand other synthetic biodegradable polymers (poly-lacticacid) and copolymers of lactic acid and ‹-caprolactone(Yannas and Hill, 2004). The efficacy of nerve guides canbe improved by means of growth factors, such as nervegrowth factor (NGF), and/or biological cues, such asSchwann cells or bone-marrow stem cells that can beintroduced along the graft to promote and guide regen-eration of peripheral axons in rats (Bryan et al., 2000;Guenard et al., 1992; Hadlock et al., 2000; Paıno et al.,1994; Rodriguez et al., 2000). In this regard, we tried tocombine the use of bioengineered grafts and cell-therapyusing SDSCs to enhance the nerve regeneration of adultrats after their sciatic transaction. As the excellent spon-taneous regeneration potential of rodents for short dis-tances is renowned, larger nerve defects were investi-gated on a 20 mm (Ijkema-Paassen et al., 2004; Madisonet al., 1988; Stang et al., 2005; Yoshii and Oka, 2001) and30 mm gap (Yoshii et al., 2003). More recently new ratmodels have been used to study regeneration across a40 mm gap (Bertelli et al., 2004; Sinis et al., 2006). Themaximum nerve gap distance that can be successfullybridged may depend on the nature of the nerve guide andthe experimental animal. Silicone and collagen tubesseem to be able to promote recovery between longer nervegaps in the rat sciatic nerve (Ansellin et al., 1997; Madi-son et al., 1988; Yoshii and Oka, 2001; Yoshii et al., 2003).Here, we bridged a gap length of 16 mm in the right scia-tic nerve of adult rats with two different types of resorb-able nerve guides filled with SDSCs: a biodegradable poly-meric tube (PLA-TMC) and a Type I collagen guide.
Regeneration of Peripheral Nerves Using SDSCs
Macroscopic observation, done 9 or 12 weeks after in-jury, showed that both the conduits were integrated intothe host tissue with minimal signs of fibrosis and onlymoderate foreign body reaction. Proximal and distalnerve ends maintained their adherence to the conduitthrough the use of fibrin glue. Repair with fibrin glue isan alternative to conventional suture repair and recentlydifferent experiments in rat models have shown that it
434 MARCHESI ET AL.
GLIA DOI 10.1002/glia

Fig. 5. SDSCs effect on nerve regeneration. Ninety days after SDSCstransplantation immunofluorescence analysis showed an increasedexpression of laminin, neurofilament and S100 in treated animals com-pared to the controls. Also the staining for endothelial markers such asCD 31 and a-SMA, revealed a higher number of vessels in SDSCs trans-planted animals (A, B). Quantification of expression of S 100, CD31 anda-SMA is shown in C. In particular statistical analysis showed that theexpression of S100 was significantly higher in collagen guides filled with
SDSCs: compared to collagen guide control group (*P < 0.001) and toPLA-TMC guide, filled with SDSCs (*P < 0.05). The S100 expressionwas also significantly higher in the animals that received the collagenguide filled with PBS in comparison with the PLA-TMC control group(Student’s t-test, *P < 0.05). RT-PCR analysis confirmed these data andalso showed an higher expression of myelin basic protein (MBP) andTRK-C (receptor of neurotrophin 3) in animals that received Type 1 col-lagen guide filled with SDSCs (D and E).
435TRANSPLANTATION OF SKIN-DERIVED STEM CELLS
GLIA DOI 10.1002/glia

seems to provide better conditions for regeneration thansuture after sciatic nerve transaction (Martins et al.,2005; Ornelas et al., 2006a,b). Electrophysiological andmorphological outcome measures, such as the immuno-fluorescence analysis, showed that nerve regeneration af-ter sciatic transection can be accomplished by transplant-ing SDSCs. In fact the amount of fibers within the tubeswas clearly influenced by the presence of these cells. Thevery high number of regenerating axons obtained by mor-phological counts may be related to the well-known phe-nomenon of axonal sprouting from the proximal stump ofthe lesioned peripheral nerve (Angelov et al., 1993, 1996).The regeneration success rate was higher in the conduitsfilled with SDSCs, in comparison to the controls and inparticular in the Type I collagen guide. Thus, the SDSCsmay stimulate the host axons to extend and to fill thetransection gap in both types of guides. In fact regenera-tion fails through long gaps most likely because the re-generative capabilities of the nerve stumps have beenexceeded and fibroblasts and Schwann cells are not ableto migrate through the gap and provide a permissiveenvironment for axonal elongation. After 7 days of im-plantation we observed several clusters of GFP positivecells within the nerve guides as demonstrated by the his-tological stains. One explanation for the low number oftransplanted GFP positive cells after long observations isthe failure of survival of GFP positive hSDSCs caused bylocal inflammation and cytokines secretion or tissue cul-ture processing. Otherwise, these results can be attribu-ted to the differentiation of SDSCs into Schwann cellswhich formed a myelin sheath surrounding axons withdissemination of the GFP protein along the joined nerve.This hypothesis seems to be confirmed by the presence ofGFP positive myelin sheaths around unlabeled host axonsof the marginal area of the guides. We observed that onlya small percentage of SDSCs appear to differentiate di-rectly into supportive cells such as S100 positive Schwanncells. Hence their number was too small to conclude thatthe observed regeneration was provoked solely by thismechanism.
Neurotrophic Factors and Angiogenesis MaySustain the Axonal Regeneration
It is thought that the microenvironment of a tissue hasan important role to control stem cell behavior due to a
combination of locally secreted factors, cell–cell interac-tions mediated by integral membrane proteins and theextracellular matrix. We observed increased numbers ofaxons in the guides after the transplantation of SDSCs. Areasonable hypothesis to explain the effect of SDSCs isthat, when placed in an injury environment, they expresstrophic factors which may contribute to neurogenesis asconfirmed by the evidence that in vitro these cells are ca-pable of expressing neurotrophic factors such as NT3 andNT4. Neurotrophins are implicated in the endogenousregulation of the repair process. Application of variousneurotrophins and growth factors has been demonstratedto encourage axonal outgrowth across the space betweenthe disconnected proximal and distal parts of the axoto-mized peripheral nerve. The therapeutic potential of neu-rotrophic factors for the treatment of peripheral nerveinjury has already been investigated in several studies(Makwana and Raivich, 2005). Because we observed anincreased regeneration in the transplanted animals, com-pared to the controls, but had no significant evidence of amassive differentiation of SDSCs into Schwann cells, wetested the expression of NT4/5 and BDNF to evaluate ifSDSCs may have regenerative capacity able to produceneurotrophic factors or influence their release. Sevendays after transaction we observed the release of bothBDNF and NT4/NT5 in the PLA-TMC and collagenguides filled with cells and 90 days after the transactionwe observed lower levels of BDNF and NT4/5 in bothtype cell filled guides. However, no significant differenceswere observed in any of the conditions or timings sug-gesting that neurotrophin release can not be the only ex-planation for the improved nerve regeneration observed incollagen guides filled with SDSCs. Immunofluorescenceevaluation of endothelial markers into the conduits filledwith the SDSCs showed traversing blood vessels whichallowed nutritive support to advancing axons and pro-vided delivery of induction factors. There is a likelihoodthat transplanted SDSCs may also promote the angiogen-esis of the host tissue, facilitating the formation of a tissuethat could fill the gap at the site of injury and an appropri-ate substrate for axonal regeneration across it.
Type I Collagen Guide Filled with SDSCs ExertPositive Effects on Axonal Regeneration
In this study we evaluated the effects of our SDSCs af-ter transplantation into commercial type 1 collagen con-duits, or house-made PLA-TMC guides, used to fill a gapof 16 mm in the rat sciatic nerve. The use of trimethylene-carbonate-co-‹-caprolactone (TMC/CL) guide as scaffoldsfor regenerating axons was described in a previous study(Sinis et al., 2005). Although the poly(L-lactide-co-tri-methylene carbonate) (PLA/TMC) polymer is quite differ-ent from the TMC/CL, these two types of guides are com-parable for their high biocompatibility without the releaseof toxic degradation products. Moreover, the microporousstructure of these tubes allows a nutritive exchangebetween the external and internal microenvironmentswhile preventing scar-forming cells from penetrating. A
TABLE 2. Expression of NT4/5 and BDNF in Treated Animals7 and 90 Days After Nerve Transaction
Treatment NT4/5 BDNF
7 days after transectionCollagen guide 1 SDSCs 631.3 6 217.5 30.8 6 8Collagen guide 1 PBS 571.3 6 75 33.9 6 4.5PLA-TMC guide 1 SDSCs 652.6 6 61.3 70 6 11PLA-TMC guide 1 PBS 551.3 6 0 46.4 6 0.8
90 days after transectionCollagen guide 1 SDSCs 582.8 6 95 5.4 6 6.5Collagen guide 1 PBS 601.2 6 90 14 6 2.5PLA-TMC guide 1 SDSCs 541.2 6 25 4.3 6 5.1PLA-TMC guide 1 PBS 400.4 6 55.8 8 6 9.2
No statistically significant differences were observed using one-way ANOVA.
436 MARCHESI ET AL.
GLIA DOI 10.1002/glia

better outcome in terms of functional recovery and nerveregeneration was observed in the Type I collagen guidesin bioengineered conduits and several parameters caninfluence the possibility of regeneration: structural condi-tions (e.g. inner diameter), biocompatibility, porousness de-gradation, and mechanical strength. The differences betweenthe collagen and PLA-TMC guides in the above parame-ters can markedly alter their ability to promote the axon’sregeneration. However, in our in vitro tests these differ-ences did not affect the survival and differentiation capa-city of SDSCs (data not shown). Furthermore, pore size,tube diameter, and wall thickness of the nerve guideswere monitored through SEM. The internal diameter ofthe PLA/TMC and collagen guides was similar (�1 mm)and their wall thickness measured �100 lm. Finally, nosignificant difference in nerve regeneration was foundwithin the group of animals treated with the PLA/TMCguides, suggesting a homogenous in vivo effect. Althoughthe guarantees offered by an industrial product certainlyhave influence, the results obtained in our in vivo experi-ments seems more related to the biological properties ofthe guides than to their structural parameters. It is likelythe collagen guide can permit an enhanced adhesion ofthe injected cells to its walls and, in consequence, easiersurvival and higher contribution to regeneration.
CONCLUSIONS
The results of this study show that SDSCs, isolated andexpanded from small pieces of rat skin and transplantedwithin a bioresorbable and permeable Type 1 collagenguide or a PLC guide, enhance peripheral nerve regenera-tion across a gap of 16 mm in the adult rat, a length thatnormally limits regeneration. The SDSCs may provide apotential autologous repair strategy for supplying nutri-tional and mechanical support in the PNS. Further eva-luation is needed to analyze the fates of grafted cells andclarify the mechanism of nerve regeneration.
ACKNOWLEDGMENTS
We thank S. Gatti, M. Fava, F. Pisati, M. Furlanetto,and D. Ripolone for technical assistance.
REFERENCES
Ahmed MR, Vairamuthu S, Shafiuzama M, Basha SH, Jayakumar R.2005. Microwave irradiated collagen tubes as a better matrix for pe-ripheral nerve regeneration. Brain Res 1046:55–67.
Angelov DN, Gunkel A, Stennert E, Neiss WF. 1993. Recovery of originalnerve supply after hypoglossal-facial anastomosis causes permanentmotor hyperinnervation of the whisker-pad muscles in the rat. J CompNeurol 338:214–224.
Angelov DN, Neiss WF, Streppel M, Andermahr J, Mader K, Stennert E.1996. Nimodipine accelerates axonal sprouting after surgical repair ofrat facial nerve. J Neurosci 16:1041–1048.
Ansellin AD, Fink T, Davey DF. 1997. Peripheral nerve regenerationthrough nerve guides seeded with adult Schwann cells. NeuropatholAppl Neurobiol 23:387–398.
Archibald SJ, Krarup C, Shefner J, Li ST, Madison RD. 1991. A col-lagen-based nerve guide conduit for peripheral nerve repair: An elec-trophysiological study of nerve regeneration in rodents and nonhuman primates. J Comp Neurol 306:685–696.
Archibald SJ, Shefner J, Krarup C, Madison RD. 1995. Monkey mediannerve repaired by nerve graft or collagen nerve guide tube. J Neurosci15:4109–4123.
Belicchi M, Pisati F, Lopa R, Poretti L, Fortunato F, Sironi M,Scalamogna M, Parati EA, Bresolin N, Torrente Y. 2004. Human skin-derived stem cells migrate throughout forebrain and differentiate intoastrocytes after injection into adult mouse brain. J Neurosci Res 77:475–486.
Belkas JS, Munro CA, Shoichet MS, Johnston M, Midha R. 2005. Long-term in vivo biomechanical properties and biocompatibility of poly(2-hydroxyethyl methacrylate-co-methyl methacrylate) nerve conduits.Biomaterials 26:1741–1749.
Belkas JS, Shoichet MS, Midha R. 2004. Peripheral nerve regenerationthrough guidance tubes. Neurol Res 26:151–160.
Bertelli JA, Dos Santos AR, Taleb M, Calixto JB, Mira JC, Ghizoni MF.2004. Long interpositional nerve graft consistently induces incompletemotor and sensory recovery in the rat. An experimental model to testnerve repair. J Neurosci Methods 15:75–80.
Bryan D, Holway A, Wang KK, Silva AE, Trantolo DJ, Wise D, Summer-hayes I. 2000. Influence of glial growth factor and Schwann cells in abioresorable guidance channel on peripheral nerve regeneration. Tis-sue Eng 6:129–138.
Cuevas P, Carceller F, Dujovny M, Garcia-Gomez I, Cuevas B, Gonzalez-Corrochano R, Diaz-Gonzalez D, Reimers D. 2002. Peripheral nerveregeneration by bone marrow stromal cells. Neurol Res 24:634–638.
Cuevas P, Carceller F, Garcia-Gomez I, Yan M, Dujovny M. 2004. Bonemarrow stromal cell implantation for peripheral nerve repair. NeurolRes 26:230–232.
De Medinaceli L, Freed WJ, Wayatt RJ. 1982. An index of the functionalcondition of rat sciatic nerve based on measurements made from walk-ing tracks. Exp Neurol 77:634–643.
Dezawa M, Takahashi I, Esaki M, Takano M, Sawada H. 2001. Sciaticnerve regeneration in rats induced by transplantation of in vitro dif-ferentiated bone-marrow stromal cells. Eur J Neurosci 14:1771–1776.
Evans GR, Brandt K, Katz S, Chavuin P, Otto L, Bogle M, Wang B, Mes-zlenyi RK, Lu L, Mikos AG, Patrick CW Jr. 2002. Bioactive poly(L-lac-tic acid) conduits seeded with Schwann cells for peripheral nerveregeneration. Biomaterials 23:841–848.
Evans GR, Brandt K, Niederbichler AD, Chauvin P, Hermann S, BogleM, Otta L, Wang B, Patrick CW Jr. 2000. Clinical long-term in vivoevaluation of poly(L-lactic acid) porous conduits for peripheral nerveregeneration. J Biomater Sci 11:787–792.
Evans GR, Brandt K, Widmer M, Lu L, Meszlenyi RK, Gupta PK,Mikos AG, Hodges J, Williams J, Gurlek A, Nabawi A, Lohman R,Patrick CW Jr. 1999. In vivo evaluation of poly (L-lactic acid) porousconduits for peripheral nerve regeneration. Biomaterials 20:1109–1115.
Fawcett JW, Keynes RJ. 1990. Peripheral nerve regeneration. Annu RevNeurosci 13:43–60.
Fernandes KJL, McKenzie IA, Mill P, Smith KM, Akhavan M, Barnebe-Heider F, Biernaskie J, Junek A, Kobayashi NR, Toma JG, Kaplan DR,Labosky PA, Rafuse V, Hui CC, Miller FD. 2004. A dermal niche formultipotent adult skin-derived precursor cells. Nat Cell Biol 6:1082–1093.
Follenzi A, Ailles LE, Bakovic S, Geuna M, Naldini L. 2000. Gene trans-fer by lentiviral vectors is limited by nuclear translocation and rescuedby HIV-1 pol sequences. Nat Genet 25:217–222.
Fu SY, Gordon T. 1997. The cellular and molecular basis of peripheralnerve regeneration. Mol Neurobiol 14:67–116.
Guenard V, Kleitman N, Morrissey TK, Bunge RP, Aebischer P. 1992.Syngeneic Schwann cells derived from adult nerves seeded in semi-permeable guidance channels enhance peripheral nerve regeneration.J Neurosci 12:3310–3320.
Hadlock T, Sundback C, Hunter D, Cheney M, Vacanti JP. 2000. A poly-mer foam conduit seeded with Schwann cells promotes guided periph-eral nerve regeneration. Tissue Eng 6:119–127.
Heine W, Conant K, Griffin JW, Hoke A. 2004. Transplanted neural stemcells promote axonal regeneration through chronically denervated pe-ripheral nerves. Exp Neurol 189:231–240.
Hou SY, Zhang HY, Quan DP, Liu XL, Zhu JK. 2006. Tissue-engineeredperipheral nerve grafting by differentiated bone marrow stromal cells.Neuroscience 140:101–110.
Ijkema-Paassen J, Jansen K, Gramsbergen A, Meek MF. 2004. Transec-tion of peripheral nerves, bridging strategies and effect evaluation.Biomaterials 25:1583–1592.
Keilhoff G, Goihl A, Langnase K, Fansa H, Wolf G. 2006. Transdifferen-tiation of mesenchymal stem cells into Schwann cell-like myelinatingcells. Eur J Cell Biol 85:11–24.
Lendahl U, Zimmerman LB, McKay RDG. 1990. CNS stem cells expressa new class of intermediate filament protein. Cell 60:585–595.
Levi AD, Guenard V, Aebischer P, Bunge RP. 1994. The functional char-acteristics of Schwann cells cultured from human peripheral nerve
437TRANSPLANTATION OF SKIN-DERIVED STEM CELLS
GLIA DOI 10.1002/glia

after transplantation into a gap within the rat sciatic nerve. J Neurosci14:1309–1319.
Levi AD, Sonntag VK, Dickman C, Mather J, Li RH, Cordoba SC,Bichard B, Berens M. 1997. The role of cultured Schwann cell graftsin the repair of gaps within the peripheral nervous system of primates.Exp Neurol 143:25–36.
Li ST, Archibald SJ, Krarup C, Madison R. 1992. Peripheral nerve repairwith collagen conduits. Clin Mater 9:95–200.
Longo FJ, Hayman EG, Davis GE, Ruoslahti E, Engvall E, Manthorpe M,Varon S. 1984. Neurite promoting factors and extracellular matrix com-ponents accumulating in vivo within nerve regenerating chambers.Brain Res 309:107–117.
Lundborg G, Longo FM, Varon S. 1982. Nerve regeneration model andtrophic factors in vivo. Brain Res 232:57–61.
Madison RD, Da Silva CF, Dikkes P. 1988. Entubulation repair with pro-tein additives increases the maximum nerve gap distance successfullybridged with tubular prostheses. Brain Res 447:325–334.
Makwana M, Raivich G. 2005. Molecular mechanisms in successful pe-ripheral regeneration. FEBS J 272:2628–2638.
Martins RS, Siqueira MG, Da Silva CF, Plese JP. 2005. Overall assess-ment of regeneration in peripheral nerve lesion repair using fibringlue, suture, or a combination of the 2 techniques in a rat model.Which is the ideal choice? Surg Neurol 64 (Suppl 1):S10–S16.
Mimura T, Dezawa M, Kanno H, Sawada H, Yamamoto I. 2004. Periph-eral nerve regeneration by transplantation of bone marrow stromalcell-derived Schwann cells in adult rats. J Neurosurg 101:806–812.
Mosahebi A, Woodward B, Wiberg M, Martin R, Terenghi G. 2001. Retro-viral labeling of Schwann cells: In vitro characterization and in vivotransplantation to improve peripheral nerve regeneration. Glia 34:8–17.
Murakami T, Fujimoto Y, Yasunaga Y, Ishida O, Tanaka N, Ikuta Y,Ochi M. 2003. Transplanted neuronal progenitor cells in a peripheralnerve gap promote nerve repair. Brain Res 974:17–24.
Ngo TT, Waggoner PJ, Romero AA, Nelson KD, Eberhart RC, Smith GM.2003. Poly(L-lactide) microfilaments enhance peripheral nerve regen-eration across extended nerve lesions. J Neurosci Res 72:227–238.
Ornelas L, Padilla L, Di Silvio M, Schalch P, Esperante S, Infante PL,Bustamante JC, Avalos P, Varela D, Lopez M. 2006a. Fibrin glue: An al-ternative technique for nerve coaptation. I. Wave amplitude, conductionvelocity, and plantar length factors. J Reconstr Microsurg 22:119–122.
Ornelas L, Padilla L, Di Silvio M, Schalch P, Esperante S, Infante PL,Bustamante JC, Avalos P, Varela D, Lopez M. 2006b. Fibrin glue: Analternative technique for nerve coaptation. II. Nerve regeneration andhistomorphometric assessment. J Reconstr Microsurg 22:123–128.
Paıno CL, Fernandez-Valle C, Bates ML, Bunge MB. 1994. Regrowth ofaxons in lesioned adult rat spinal cord: Promotion by implants of cul-tured Schwann cells. J Neurocytol 23:433–452.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD,Moorman MA, Simonetti DW, Craig S, Marshak DR. 1999. Multiline-
age potential of adult human mesenchymal stem cells. Science 284:143–147.
Prockop DJ. 1997. Marrow stromal cells as stem cells for nonhemato-poietic tissues. Science 276:71–74.
Rodriguez FJ, Verdu E, Ceballos D, Navarro X. 2000. Nerve guides seededwith autologous Schwann cells improve nerve regeneration. Exp Neurol161:571–584.
Sinis N, Schaller H-E, Becker ST, Lanaras T, Schulte-Eversum C, M€uller H-W,Vonthein R, R€osner H, Haerle M. 2006. Cross-chest median nervetransfer: A new model for the evaluation of nerve regeneration across a40 mm gap in the rat. J Neurosci Methods 156:166–172.
Sinis N, Schaller HE, Schulte-Eversum C, Schlosshauer B, Doser M,Dietz K, Rosner H, Muller HW, Haerle M. 2005. Nerve regenerationacross a 2-cm gap in the rat median nerve using a resorbable nerveconduit filled with Schwann cells. J Neurosurg 103:1067–1076.
Stang F, Fansa H, Wolf G, Reppin M, Keilhoff G. 2005. Structural pa-rameters of collagen nerve grafts influence peripheral nerve regenera-tion. Biomaterials 26:3083–3091.
Stoll G, Muller HW. 1999. Nerve injury, axonal degeneration and neuralregeneration: Basic insights. Brain Pathol 9:313–325.
Tohill M, Mantovani C, Mikael W, Terenghi G. 2004. Rat bone marrowmesenchymal stem cells express glial markers and stimulat nerveregeneration. Neurosci Lett 362:200–203.
Toma JG, Akhavan M, Fernandes KJL, Barnab�e-Heider F, Sadikot A,Kaplan DR, Miller FD. 2001. Isolation of multipotent adult stem cellsfrom the dermis of mammalian skin. Nat Cell Biol 3:778–784.
Widmer MS, Gupta PK, Lu L, Meszlenyi R, Evans GRD, Brandt K, SavelT, Gurlek A, Patrick CW Jr, Mikos AG. 1998. Manufacture of porousbiodegradable polymer conduits by an extrusion process for guided tis-sue regeneration. Biomaterials 19:1945–1955.
Williams LR, Longo FM, Powell HC, Lundborg G, Varon S. 1983. Spatial-temporal progress of peripheral nerve regeneration within a siliconechamber: Parameters for a bioassay. J Comp Neurol 218:160–170.
Yannas IV, Hill BJ. 2004. Selection of biomaterials for peripheral nerveregeneration using data from the nerve chamber model. Biomaterials25:1593–1600.
Yoshii S, Oka M. 2001. Peripheral nerve regeneration along collagen fila-ments. Brain Res 888:158–162.
Yoshii S, Oka M, Shima M, Taniguchi A, Akagi M. 2003. Bridging a30-mm nerve defect using collagen filaments. J Biomed Mater Res A67:467–474.
Zennou V, Serguera C, Sarkis C, Colin p, Perret E, Mallet J, CharneauP. 1999. The HIV-1 DNA flap stimulates HIV vector mediated celltransduction in the brain. Nat Biotechnol 19:446–450.
Zufferey R, Donello JE, Trono D, Hope TJ. 1999. Woodchuck hepatitis vi-rus posttranscriptional regulatory element enhances expression oftransgenes delivered by retroviral vectors. J Virol 73:2886–2892.
438 MARCHESI ET AL.
GLIA DOI 10.1002/glia
Copyright © 2022 FDOKUMEN