
NK1, NMDA, 5HT1a, and 5HT2 Receptor Binding Sites in the Rat Lumbar Spinal Cord: Modulation...
14
NK1, NMDA, 5HT1a, and 5HT2 Receptor Binding Sites in the Rat Lumbar Spinal Cord: Modulation Following Sciatic Nerve Crush S. Croul,* A. Radzievsky,* A. Sverstiuk,* and M. Murray² *Departments of Pathology and Laboratory Medicine and ²Departments of Neurobiology and Anatomy, Allegheny University of the Health Sciences, Philadelphia, Pennsylvania 19102-1192 Received February 14, 1997; accepted June 16, 1998 Quantitative receptor binding autoradiography was used to study the NK1, NMDA, 5HT1a, and 5HT2 recep- tor binding densities in the adult rat lumbar spinal cord from 3 days to 20 weeks following a unilateral crush lesion of the sciatic nerve. NK1 binding density increased unilaterally in the superficial dorsal horn on the side of the sciatic crush to reach levels 60% above controls by 4 weeks following the lesion and returned to control values by 12 weeks. NMDA binding density increased bilaterally and equally in both the dorsal and ventral horns to reach 300% of control values at 2 weeks following the crush and returned to near control values by 20 weeks following the lesion. Serotonergic receptor binding did not change. The changes in NK1 receptor binding density on postsynaptic dorsal horn cells are consistent with a response to the decrease and recovery in the synthesis and transport of tachykinins by the dorsal root ganglion cells following peripheral nerve injury. The bilateral changes in NMDA receptor binding are more likely mediated by polysynap- tic pathways in the spinal cord that respond to the changes in metabolic events of the dorsal root ganglion cells evoked by axotomy and regeneration. r 1998 Academic Press Key Words: spinal cord; sciatic nerve; receptor; sub- stance P; NK; glutamate; NMDA; serotonin. INTRODUCTION Section of dorsal roots results in Wallerian degenera- tion of the central processes of dorsal root ganglion (DRG) cells and secondary changes within the spinal cord. These changes include modification of the synap- tic complement in the dorsal horn (31, 51) and perma- nent loss of the peptides and enzymes entirely supplied by the dorsal root ganglion cells such as fluoride- resistant acid phosphatase (FRAP) and calcitonin gene- related peptide (CGRP), (27, 47). An acute loss and chronic recovery of the dorsal horn content of other peptides associated with dorsal root afferents such as substance P (SP) has been observed (42, 47). In addi- tion, since dorsal roots do not regenerate unaided into the spinal cord, the synaptic reorganization in the dorsal horn following this lesion is attributable to sprouting of undamaged intraspinal and supraspinal projections. The partial restitution of SP content in the dorsal horn is therefore due to sprouting of intrinsic SP-containing axons (12). Injury to the peripheral processes of the DRG pro- duces a modest cell loss in the associated dorsal root ganglia with a consequent dorsal root degeneration that is much less pronounced than that following dorsal root section. It does, however, produce intraspinal modifications of dorsal horn synapses, enzymes, and peptides similar in many respects although less dra- matic than those found after dorsal root section (1). If regeneration of peripheral axons is prevented by sec- tion and ligation, the changes in the dorsal horn are more marked and more likely to be permanent than those following nerve crush or section and reapposition which allow peripheral regeneration to occur (1, 2, 10). Peripheral lesions deprive the DRG of target-derived growth factors such as nerve growth factor which may account both for synthetic changes in the DRG and the changes observed in the dorsal horn. Restoration of enzymes and peptides that occurs after regeneration is mediated by dorsal root ganglion cells (16, 18, 19). Receptor binding autoradiography has provided a measure of the effects of these lesions on the target neurons of the dorsal root projections. Thus, binding density for the dominant SP receptor subtype in the superficial dorsal horn (NK1) increases acutely on the experimental side of the spinal cord at 2 weeks follow- ing dorsal rhizotomy and then decreases to control levels by 6 weeks. These findings imply that spinal targets respond to partial denervation by increasing receptor density and that the intraspinal sprouting which restores the density of SP terminals also restores the receptor density to normal (9, 25, 32, 50). We have explored the possibility that sciatic nerve crush will also change the receptor properties of spinal targets even though the lesion produces little degenera- tion. We chose to measure the Neurokinin1 (NK1) and EXPERIMENTAL NEUROLOGY 154, 66–79 (1998) ARTICLE NO. EN986875 66 0014-4886/98 $25.00 Copyright r 1998 by Academic Press All rights of reproduction in any form reserved.
Transcript of NK1, NMDA, 5HT1a, and 5HT2 Receptor Binding Sites in the Rat Lumbar Spinal Cord: Modulation...

NK1, NMDA, 5HT1a, and 5HT2 Receptor Binding Sites in the RatLumbar Spinal Cord: Modulation Following Sciatic Nerve Crush
S. Croul,* A. Radzievsky,* A. Sverstiuk,* and M. Murray†*Departments of Pathology and Laboratory Medicine and †Departments of Neurobiology and Anatomy,
Allegheny University of the Health Sciences, Philadelphia, Pennsylvania 19102-1192
Received February 14, 1997; accepted June 16, 1998
Quantitative receptor binding autoradiography wasused to study the NK1, NMDA, 5HT1a, and 5HT2 recep-tor binding densities in the adult rat lumbar spinalcord from 3 days to 20 weeks following a unilateral crushlesion of the sciatic nerve. NK1 binding density increasedunilaterally in the superficial dorsal horn on the side ofthe sciatic crush to reach levels 60% above controls by 4weeks following the lesion and returned to control valuesby 12 weeks. NMDA binding density increased bilaterallyand equally in both the dorsal and ventral horns to reach300% of control values at 2 weeks following the crush andreturned to near control values by 20 weeks following thelesion. Serotonergic receptor binding did not change. Thechanges in NK1 receptor binding density on postsynapticdorsal horn cells are consistent with a response to thedecrease and recovery in the synthesis and transport oftachykinins by the dorsal root ganglion cells followingperipheral nerve injury. The bilateral changes in NMDAreceptor binding are more likely mediated by polysynap-ticpathwaysinthespinalcordthatrespondtothechangesin metabolic events of the dorsal root ganglion cellsevoked by axotomy and regeneration. r 1998 Academic Press
Key Words: spinal cord; sciatic nerve; receptor; sub-stance P; NK; glutamate; NMDA; serotonin.
INTRODUCTION
Section of dorsal roots results in Wallerian degenera-tion of the central processes of dorsal root ganglion(DRG) cells and secondary changes within the spinalcord. These changes include modification of the synap-tic complement in the dorsal horn (31, 51) and perma-nent loss of the peptides and enzymes entirely suppliedby the dorsal root ganglion cells such as fluoride-resistant acid phosphatase (FRAP) and calcitonin gene-related peptide (CGRP), (27, 47). An acute loss andchronic recovery of the dorsal horn content of otherpeptides associated with dorsal root afferents such assubstance P (SP) has been observed (42, 47). In addi-tion, since dorsal roots do not regenerate unaided into
the spinal cord, the synaptic reorganization in thedorsal horn following this lesion is attributable tosprouting of undamaged intraspinal and supraspinalprojections. The partial restitution of SP content in thedorsal horn is therefore due to sprouting of intrinsicSP-containing axons (12).
Injury to the peripheral processes of the DRG pro-duces a modest cell loss in the associated dorsal rootganglia with a consequent dorsal root degenerationthat is much less pronounced than that following dorsalroot section. It does, however, produce intraspinalmodifications of dorsal horn synapses, enzymes, andpeptides similar in many respects although less dra-matic than those found after dorsal root section (1). Ifregeneration of peripheral axons is prevented by sec-tion and ligation, the changes in the dorsal horn aremore marked and more likely to be permanent thanthose following nerve crush or section and reappositionwhich allow peripheral regeneration to occur (1, 2, 10).Peripheral lesions deprive the DRG of target-derivedgrowth factors such as nerve growth factor which mayaccount both for synthetic changes in the DRG and thechanges observed in the dorsal horn. Restoration ofenzymes and peptides that occurs after regeneration ismediated by dorsal root ganglion cells (16, 18, 19).
Receptor binding autoradiography has provided ameasure of the effects of these lesions on the targetneurons of the dorsal root projections. Thus, bindingdensity for the dominant SP receptor subtype in thesuperficial dorsal horn (NK1) increases acutely on theexperimental side of the spinal cord at 2 weeks follow-ing dorsal rhizotomy and then decreases to controllevels by 6 weeks. These findings imply that spinaltargets respond to partial denervation by increasingreceptor density and that the intraspinal sproutingwhich restores the density of SP terminals also restoresthe receptor density to normal (9, 25, 32, 50).
We have explored the possibility that sciatic nervecrush will also change the receptor properties of spinaltargets even though the lesion produces little degenera-tion. We chose to measure the Neurokinin1 (NK1) and
EXPERIMENTAL NEUROLOGY 154, 66–79 (1998)ARTICLE NO. EN986875
660014-4886/98 $25.00Copyright r 1998 by Academic PressAll rights of reproduction in any form reserved.

N-methyl D-aspartate (NMDA) receptors because theyrepresent targets of neurotransmitters supplied bydorsal root afferents. SP content in the dorsal horn ismodified following sciatic nerve crush (2, 19) and NK1receptor binding changes after deafferentation (9). Glu-tamate is colocalized in DRG neurons and dorsal rootterminals with SP (11, 28, 48), there is high NMDAreceptor binding density in the spinal dorsal horn(9, 30), and it has been implicated in synaptic stabiliza-tion and plasticity in other CNS systems (46). Seroto-nin 1a (5HT1a) receptors were also measured becausethey are associated with supraspinal rather than dor-sal root afferents and are present at high levels in thedorsal horn (24, 44).
Since sciatic nerve lesions also axotomize motorneurons, we examined receptors in the ventral horn aswell. In the case of NK and glutamate receptors, theanalysis in the ventral horn was performed for thesame subtypes as in the dorsal horn. However, for 5HTreceptors, we also analyzed the 5HT2 subtype becauseof its high receptor binding density in the ventral horn(24, 44).
MATERIALS AND METHODS
Experimental Surgeries
Adult female Sprague–Dawley rats weighing on aver-age 225 g were purchased from Ace Animals (Boyer-town, PA). They were housed in cages of two or three inan AAALAC accredited facility. Surgeries were per-formed using ketamine-xylazine-acepromazine anesthe-sia (50 mg, 5 mg, and 0.75 mg/kg i.p.) with aseptictechnique. The right sciatic nerve was exposed atmidthigh, the nerve was crushed with a number fivejewelers’ forceps for 5 s, released, and recrushed foranother 5 s. Postoperatively, the response of the rightpaw to pinch was tested weekly. Animals were sacri-ficed in groups of six at 3 days and 1, 2, 4, 12, and 20weeks postoperative. Six nonoperated rats were in-cluded as controls.
Tissue Processing
All animals were anesthetized with carbon dioxideprior to decapitation. Spinal cords were quickly re-moved, frozen in isopentane, and stored at 270°C.Segments of sciatic nerve 2–3 mm distal to the lesionwere immersion fixed in phosphate-buffered 4% parafor-maldehyde–1% glutaraldehyde, dehydrated throughascending alcohols, cleared in propylene oxide, andembedded in Epon. Semithin toluidine blue-stainedsections and selected thin sections for electron micros-copy from these blocks were used to confirm the com-pleteness of the crush lesion acutely and the regenera-tion of the nerve at the chronic survival times.
The midlumbar spinal cord was cut in the transverseplane into 20-µm-thick sections at 220°C and mountedon chrome alum, poly-L-lysine-coated slides. All slideswere subsequently desiccated for 12 to 16 h at 4°C andtransferred to 270°C for storage prior to autoradiography.
Receptor Binding Autoradiography
Reagents were purchased from Sigma Chemicals (St.Louis MO), except for radiochemicals. [125I]SP and[3H]8-OH-DPAT were purchased from Amersham LifeSciences (Arlington Heights, IL). [ 125I]DOI and[3H]MK801 were purchased from NEN Research Prod-ucts (Boston, MA).
For autoradiography, slides were removed from 270°Cstorage, thawed, and quickly dried with cool air. Follow-ing this, they were immersed in preincubation buffer.This was followed by immersion of half of the slides inincubation buffer with radioactive ligand to measuretotal binding. The other slides were incubated in buffercontaining radioactive ligand and a high concentrationof a blocking agent for the receptor binding site tomeasure nonspecific binding. Slides were then washed,dried quickly, and vacuum desiccated overnight atroom temperature. The next day, slides were apposed totritium-sensitive film (Hyperfilm-Amersham) in autora-diography cassettes which were exposed at 4°C. Twenty-micrometer-thick 125I or 3H standards (Amersham mi-croscales), depending on the ligand chosen, wereincluded in the cassettes. Exposed films were traydeveloped in D-19 (Kodak), fixed, and washed permanufacturer’s directions. Dried autoradiograms weremounted on glass slides. The exposed tissue sectionswere immersion fixed in 4% pararaformaldehyde, 0.1Mphosphate buffer, pH 7.4, and stained with Pearse’sChromoxane Cyanine R stain for myelin and a CresylViolet counter stain (34).
For the receptor subtypes investigated, our methodsfollowed previously published protocols (Table 1).
Analysis and Statistical Treatment
To determine if the crush lesion resulted in unilateralshrinkage of spinal gray matter, an analysis of selectedlaminar areas was performed with the MORPHON 5.0system (MORPHON Image Analysis Systems, BalaCynwyd, PA). Stained tissue sections were projected ona Sony Trinitron monitor at a magnification of 40X.Laminae I/II and IX, defined by the criteria of Molanderet al. (29), were outlined with the cursor to producecomputer-generated area measures. An average of foursections were measured from each animal.
For densitometric analysis of the autoradiograms, anaverage of six sections (four for total binding and twofor nonspecific binding) were analyzed per animal per
67NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH

ligand using the stained tissue section as a template.Both laminae I/II and IX in each autoradiogram wereoutlined with the cursor and optical densities weregenerated for those defined regions. Standardizationcurves were calculated from the optical densities of thefilm exposed to commercial 125I or 3H microscales usingthe manufacturers’ values of radioactive concentrationand decay. Regression analyses were performed on thestandard data using the method of least squares toyield a power curve (Y 5 aXb). Optical density valuesmeasured from the tissue sections were converted tonCi/mg tissue of ligand bound by comparison to thestandard curves and then converted to fmol/mg tissuefrom the ligand-specific activity. Nonspecific bindingwas subtracted from total binding in adjacent sectionsto yield values for specific binding density.
To compensate for interanimal variability in thestatistical analysis of laminar areas, the unlesionedside of each section was used as an internal control togive ratios of lesioned/unlesioned sides. Since ratiomeasures represent nonparametric data, these datawere analyzed using the Kruskal–Wallis one-way analy-sis of variance by ranks, setting the significance level at0.05.
For the analysis of the receptor binding density data,two methods were used. Absolute values of bindingdensities expressed as fmol/mg tissue for lesioned andunlesioned sides were initially analyzed using an Analy-sis of Variance (ANOVA) setting the significance levelat 0.05. Because of anatomical evidence that the DRGprojection to lumbar spinal cord is predominantly
unilateral, the data were also analyzed using theunlesioned side of each section as an internal control togive ratios of lesioned/unlesioned sides. This reducesthe effect of interanimal variability. The Kruskal–Wallis one-way analysis of variance by ranks wasemployed, setting the significance level at 0.05 as well.
RESULTS
In all operated animals, withdrawal of the right hindpaw to pinch was absent upon recovery from anesthesiaand did not return in any of the 3 day, 1 or 2 weeksurvivors. Most of the rats sacrificed at 4 weeks postop-erative exhibited withdrawal to pinch. Withdrawal topinch was present in all animals in the groups sacri-ficed at 12 and 20 weeks following crush. At 3 days aftercrush, 1-µm-thick sections of the sciatic nerve distal tothe crush site showed almost complete degeneration oflarge diameter fibers and electron microscopy showedmarked diminution in the number of small axons. By 20weeks postoperative, 1 µm and electron microscopicsections of the sciatic nerve showed both large andsmall diameter fibers.
Effect of Lesion on Laminar Areas (Fig. 1)
Lesioned/unlesioned ratios of cross-sectional areas oflaminae I/II and lamina IX show no significant differ-ences between groups (P # 0.05). Therefore, the crushlesion did not result in ipsilateral shrinkage of spinalgray matter in these laminae. Comparison of binding
TABLE 1
Receptor Binding Protocols
Receptor subtype
NK1 5HT1a 5HT2 NMDA
Reference 14 24 43 41Preincubation buffer 0.05 M Tris HCl, 0.02%
BSA, pH 7.4, 15 min RT170 mM Tris HCl, 4 mM
CaCl2, 0.01% ascorbicacid, 0.01% BSA, 1 µMfluoxetine, pH 7.6, 30min RT
0.05 M Tris HCl, 4 mMCaCl2, 0.1% ascorbicacid, 0.1% BSA, pH 7.4,30 min RT
None
Incubation buffer 0.05 M Tris HCl, 3 mMMnCl2, 4 mg/ml Baci-tracin, 4 mg/ml leu-peptin, 2 mg/ml chy-mostatin, pH 7.4, 30 minRT
Above buffer plus 10 µMpargyline, 60 min RT
Above buffer 60 min RT 0.03 M Hepes, 1 mMEDTA, 0.1 mM gluta-mate, 0.1 mM glycine, 75mM spermine, pH 7.45,2.5 h RT
Ligand [125I]BH-SP, 100 pM 8-OH-[3H]DPAT, 2 nM [125I]DOI, 0.2 nM [3H]MK801 25 nMNonspecific blocking agent SP, 1 µM 5HT, 1 µM Ketanserin, 1 µM Ketamine, 0.2 mMRinse 0.05 M Tris, pH 7.4, 15
s 3 5 changes, 3 dips 3 2changes DH2O, all 4°C
Incubation buffer 5min 3 3 changes, thendipped quickly DH2O 4°C
Incubation buffer 15min 3 2 changes, 4°C
Incubation buffer 20min 3 2 changes
Exposure time 4 to 7 days 8 weeks 66 h 8 weeks
68 CROUL ET AL.
densities between right and left sides of the samespinal cord or between different spinal cords weretherefore made without additional area corrections.
Receptor Binding Density
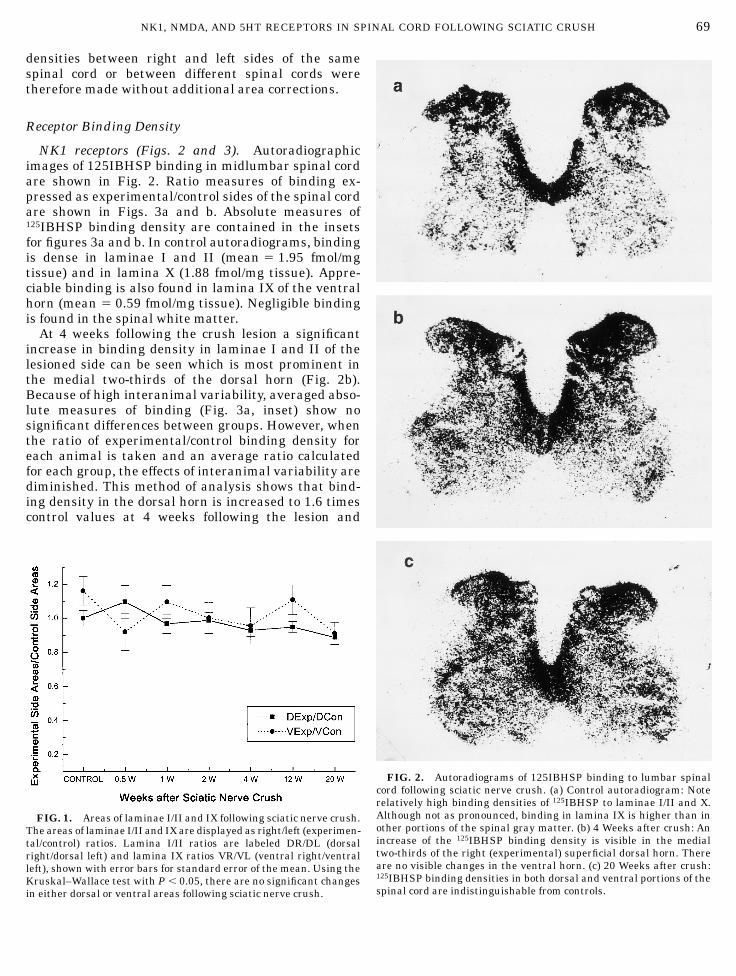
NK1 receptors (Figs. 2 and 3). Autoradiographicimages of 125IBHSP binding in midlumbar spinal cordare shown in Fig. 2. Ratio measures of binding ex-pressed as experimental/control sides of the spinal cordare shown in Figs. 3a and b. Absolute measures of125IBHSP binding density are contained in the insetsfor figures 3a and b. In control autoradiograms, bindingis dense in laminae I and II (mean 5 1.95 fmol/mgtissue) and in lamina X (1.88 fmol/mg tissue). Appre-ciable binding is also found in lamina IX of the ventralhorn (mean 5 0.59 fmol/mg tissue). Negligible bindingis found in the spinal white matter.
At 4 weeks following the crush lesion a significantincrease in binding density in laminae I and II of thelesioned side can be seen which is most prominent inthe medial two-thirds of the dorsal horn (Fig. 2b).Because of high interanimal variability, averaged abso-lute measures of binding (Fig. 3a, inset) show nosignificant differences between groups. However, whenthe ratio of experimental/control binding density foreach animal is taken and an average ratio calculatedfor each group, the effects of interanimal variability arediminished. This method of analysis shows that bind-ing density in the dorsal horn is increased to 1.6 timescontrol values at 4 weeks following the lesion and
FIG. 1. Areas of laminae I/II and IX following sciatic nerve crush.The areas of laminae I/II and IX are displayed as right/left (experimen-tal/control) ratios. Lamina I/II ratios are labeled DR/DL (dorsalright/dorsal left) and lamina IX ratios VR/VL (ventral right/ventralleft), shown with error bars for standard error of the mean. Using theKruskal–Wallace test with P , 0.05, there are no significant changesin either dorsal or ventral areas following sciatic nerve crush.
FIG. 2. Autoradiograms of 125IBHSP binding to lumbar spinalcord following sciatic nerve crush. (a) Control autoradiogram: Noterelatively high binding densities of 125IBHSP to laminae I/II and X.Although not as pronounced, binding in lamina IX is higher than inother portions of the spinal gray matter. (b) 4 Weeks after crush: Anincrease of the 125IBHSP binding density is visible in the medialtwo-thirds of the right (experimental) superficial dorsal horn. Thereare no visible changes in the ventral horn. (c) 20 Weeks after crush:125IBHSP binding densities in both dorsal and ventral portions of thespinal cord are indistinguishable from controls.
69NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH

returns to normal by 12 weeks (P , 0.05 by Kruskal–Wallace test) (Fig. 3a).
In lamina IX, no significant changes in NK1 bindingare found following the crush lesion (Fig. 3b).
NMDA (Figs. 4 and 5). Control autoradiograms of[3H]MK801 binding (Fig. 4a) show that binding in therat lumbar spinal cord is predominantly localized tolaminae I/II (mean 5 82 fmol [3H]MK801/mg tissue,approximately 40 times that for 125IBHSP). Consider-ably lower levels of binding are seen in lamina IX(mean 5 13 fmol [3H]MK801/mg tissue). Following sci-atic crush the autoradiograms demonstrate an increaseof binding density in both lesioned and unlesioneddorsal horns (Fig. 4b). The absolute values of binding(Fig. 5a) show a parallel threefold increase in bothdorsal horns in the first 2 weeks following crush. From2 to 20 weeks following the lesion, the binding densitiesfall again in parallel to 1.5 times control values. Thereare significant differences between control and experi-mental values bilaterally for all postoperative survivaltimes except 20 weeks. Analysis of the data as experi-
mental/control ratios (Fig. 5a, inset) shows no signifi-cant difference between the two sides from 3 days to 20weeks, indicating that the changes on the two sides aresymmetrical. In the ventral horn absolute values ofbinding show a similar bilateral and symmetric in-crease. Binding density reaches 3 times control valuesby 2 weeks postlesion and returns to 1.4 times controlvalues by 20 weeks postlesion (Fig. 5b). As in the dorsalhorn, there are significant differences between controland experimental values bilaterally for all postopera-tive survival times except 20 weeks. In the ventralhorn, experimental/control ratios again show no signifi-cant differences following the lesion which indicatessymmetrical changes ipsilateral and contralateral tothe nerve crush (Fig. 5b, inset).
5HT1A (Figs. 6a, 7a, and 7b). The highest levels of[3H]8-OH-DPAT receptor binding density are in lami-nae I, II, and III of the dorsal horn (mean value of 32fmol [3H]8-OH-DPAT/mg tissue, 16 times that of125IBHSP binding in the dorsal horn). In lamina IX,
FIG. 3. Quantitation of 125IBHSP BINDING density in lumbar spinal cord following sciatic nerve crush. (a) Superficial dorsal horn(laminae I/II). 125IBHSP BINDING density is shown for laminae I/II. The center bar graph shows the data as experimental/control or dorsalright/dorsal left (DR/DL) ratios. The inset graph displays the same data as the absolute values of binding density in fmol/mg. Vertical barsdelineate standard errors of the mean. Although no significant changes in binding density are found through the analysis of absolute values,the ratio measures show a significant increase in binding density at 4 weeks following the lesion (*P , 0.05; Kruskal–Wallace test). (b) Ventralhorn (lamina IX). The center graph again shows binding density as a right/left or experimental/control ratio. The inset graph displays thesame data as absolute levels of binding (fmol/mg). Neither analysis yields changes of significance in lamina IX.
70 CROUL ET AL.

FIG. 3—Continued
FIG. 4. Autoradiograms of 3HMK801 binding to lumbar spinalcord following sciatic nerve crush. (a) Control: Binding density inspinal gray matter is greatest in laminae I/II. Lamina IX is difficult todistinguish from the remainder of the spinal gray. (b) 2 weeksfollowing crush: Binding density in laminae I/II is greater thancontrols on both control and experimental sides. Again, the bindingdensity in lamina IX is difficult to distinguish from the remainder ofthe spinal gray. (c) 20 weeks following crush: Binding density in theentire spinal gray matter is indistinguishable from controls.
71NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH

mean values of binding are 4 fmol [3H]8-OH-DPAT/mgtissue. There are no significant changes in either dorsalor ventral horns at any time following crush.
5HT2 (Figs. 6b, 7c, and 7d). The highest levels ofreceptor binding are concentrated in lamina IX. Meanvalues are 0.17 fmol [125I]DOI/mg tissue in laminae I/IIand 0.5 fmol [125I]DOI/mg tissue in lamina IX, whichare less than 1/10 that for 125IBHSP. There are no
significant changes in [125I]DOI binding density follow-ing the lesion.
DISCUSSION
The Dorsal Horn
NK1 receptor binding. After lumbosacral deafferen-tation, 125IBHSP receptor binding densities in the
FIG. 5. Quantitation of 3HMK801 BINDING density in lumbar spinal cord following sciatic nerve crush. (a) Superficial dorsal horn(lamina I/II). 3HMK801 binding density is shown for laminae I/II. The center bar graph shows the data as absolute values of binding density infmol/mg. Note that scale differs from that for 125IBHSP in Fig. 3. The inset graph displays the same data as experimental/control or dorsalright/dorsal left (DR/DL) ratios. Vertical bars delineate standard errors of the mean. The analysis of absolute values in this case showsignificant increases in binding density beginning at 3 days, peaking at approximately 3 times controls at 2 weeks following the lesion on bothexperimental and control sides (*P , 0.05; ANOVA). At times after 2 weeks, binding densities decrease to 20 weeks, at which time they are stillsignificantly different from controls. Because of the bilaterality of the process, ratios of experimental/control sides show no change. (b) Ventralhorn (lamina IX). The center graph again shows binding density as absolute levels of binding (fmol/mg). The scale is the same as that in (a) toallow comparison of absolute levels. The inset graph displays the same data as right/left or experimental/control ratios. Again, absolute valuesreveal a smooth, bilateral change peaking at 3 times control values at 2 weeks following crush at decreasing to 20 weeks. Note that all absolutelevels of binding in lamina IX are much lower (approximately 20%) of those in laminae I/II. Because of the bilateral response, the dataexpressed as experimental/control ratios are again without significant change.
72 CROUL ET AL.

FIG. 5—Continued
FIG. 6. Autoradiograms of binding to lumbar spinal cord. (a) 3H8OHDPAT binding in control spinal cord. The highest binding density isclearly in the dorsal horn, extending into lamina III. High binding is also seen in the medial aspect of the dorsal horn extending into lamina X.Lamina IX also shows binding density above that seen in the rest of the spinal gray matter. (b) 125IDOI binding in control spinal cord. Bindingdensity above the rest of the spinal gray is found in the medial dorsal horn extending into lamina X. The highest binding density is seen inlamina IX with a particular preference for the medial motor nuclei.
73NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH

superficial dorsal horn increase acutely reaching 250%at 2 weeks and return to normal by 6 weeks followingthe lesion (9, 25, 32, 50). The acute changes followingdeafferentation have been attributed to denervationsupersensitivity for SP, and the return to normal valuesat chronic time points to the proliferation of synapticterminals (central sprouting) that restores SP innerva-tion in the dorsal horn to near normal levels (9, 25, 32,49–51).
125IBHSP receptor binding density in the medialtwo-thirds of the dorsal horn increases 60% during thefirst 4 weeks following sciatic nerve crush and returnsto normal values by 12 weeks following the lesion. Webelieve that these receptor binding changes are due tothe decrease and recovery in tachykinin synthesis andtransport to the dorsal horn rather than denervation-induced sprouting. While there is some afferent remod-eling in the dorsal horn and some loss of dorsal rootganglion cells following the crush lesion, synthetic and
transport functions in surviving dorsal root ganglioncells are greatly modified after any peripheral lesion(5, 15). Thus, electron microscopy of the dorsal hornacutely following crush shows darkening, shrinkage,and vesicular change of primary afferent terminals butdoes not reveal more substantial degenerative changes(17). Dorsal root ganglion cell loss is between 10 and20% (38). A 30% decrease in SP content and a 20 to 40%decrease in synthetic rate of SP in the DRG occuracutely following crush (5, 33) and SP immunoreactiv-ity in the superficial dorsal horn is decreased by 60%between 11 and 15 days following a crush lesion (2, 19).
The return of many of these measures to normal atlater time points after a peripheral lesion is primarilydue to restoration of the synthetic activity by DRGneurons which occurs in parallel with regeneration ofthe peripheral axon and probably results from access totarget-derived growth factors (15). SP content in theDRG returns to normal values by 35 to 45 days
FIG. 7. Quantitation of 3H8OHDPAT and 125IDOI binding density in lumbar spinal cord following sciatic nerve crush. (a and b)3H8OHDPAT binding densities in dorsal (laminae I/II) and ventral (lamina IX) spinal cord. In both figures, the central graph shows levels ofbinding expressed as right/left (control/experimental) ratios. Inset graphs show the same data as absolute values in fmol/mg. Scales for thesetwo graphs are identical to allow comparisons of dorsal and ventral values. They differ from scales in Figs. 3 and 5. None of these analysesreveals a significant change following sciatic crush. (c and d) 125IDOI binding densities in dorsal (laminae I/II) and ventral (lamina IX) spinalcord. In both figures, the central graph shows levels of binding expressed as right/left (control/experimental) ratios. Again, dorsal and ventralscale are identical but differ from those in Figs. 3, 5, and 7a. Inset graphs show the same data as absolute values in fmol/mg. None of theseanalyses reveals a significant change following sciatic crush.
74 CROUL ET AL.

following crush and synthetic rate becomes normal by64 days (5, 33). SP content of the superficial dorsal hornalso returns to normal (2, 19).
Our results indicate that tachykinin receptors in thedorsal horn are mediated in response to loss andreplacement of SP containing synaptic contacts and arealso changed, but to a lesser extent, by modifications inthe synthesis and transport of tachykinins by survivingDRG cells.
NMDA receptor binding. Using [3H]MK801 as theligand for NMDA receptors, we have demonstrated abilateral and symmetrical threefold increase in bindingdensity in the superficial dorsal horns at 2 weeksfollowing sciatic crush which returns to near normalvalues by 20 weeks following the lesion. A retrospectiveanalysis of [3H]MK801 binding densities in the dorsalhorn following deafferentation (9) shows a similarbilateral, symmetrical increase in binding with maxi-mum values reached at 1 week following surgery (datanot shown). The response of NMDA receptors is thusquantitatively much greater than that of NK1 receptors(i.e., 3 fold versus 1.5 fold). Because of the largeresponse, this effect is significant with absolute mea-sures. In that the response is bilateral and symmetri-cal, it is not revealed by side to side relative measures.
Glutamate and SP are colocalized in dorsal root affer-ents (11, 28, 48), nevertheless these differences inlaterality and magnitude of response further suggestthat the changes in NK1 and NMDA receptor bindingon target cells may reflect different mechanisms.
At lumbar levels, dorsal root projections are predomi-nantly ipsilateral, although Light and Perl (20, 21)have demonstrated that a few primary afferents crossto contralateral Lamina III in every level of the lumbarspinal cord. Thus, while a unilateral sciatic nervelesion may have some direct effect on the contralateraldorsal horn through these connections, it is unlikelythat loss of these direct projections account for thesymmetrical bilateral effects seen in these experi-ments. Since there are few monosynaptic connectionsto explain this effect, the bilateral effects are mostlikely mediated by polysynaptic pathways. In recentyears there have been a number of reports of bilateralresponses to unilateral peripheral nerve lesions. Thus,partial sciatic nerve ligation increases the receptivefield size of neurons in the dorsal horn bilaterally (3).Microdialysis probes in the dorsal horn show bilateralrelease of SP following unilateral sciatic stimulation (7)and bilateral release of aspartate and glutamate follow-ing unilateral intradermal capsaicin injection (40).
FIG. 7—Continued
75NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH

FIG. 7—Continued
FIG. 7—Continued

Deoxyglucose autoradiography of the spinal cord re-veals a bilateral increase in dorsal horn glucose utiliza-tion 2 min following formalin injection into one paw pad(36). Unilateral formalin injection into the paw padacutely results in a bilateral increase in NADPH diapho-rase staining of both dorsal root ganglia and superficialdorsal horn (45). Experimental monoarthritis causes abilateral increase in SP and CGRP (23) and in opiodreceptor binding density (4) in the dorsal horns. Particu-larly germane to our findings with NMDA receptorbinding is the report of a bilateral increase in GluR1gene expression in the dorsal horn following monoarthri-tis (35). In contrast to our findings, these studies haveshown that the contralateral effects, while significant,are of less magnitude than those ipsilateral to thelesion.
5HT receptors. The absence of a difference in [3H]8-OH-DPAT binding in the dorsal horn following crush isconsistent with our previous studies (9) which failed todemonstrate a significant change in the deafferenteddorsal horn. We have focused on [3H]8-OH-DPAT recep-tor binding density as a measure of the 5HT1a receptorbased on the predominance of [3H]8-OH-DPAT bindingwithin the spinal cord (24). Analysis of [125I]DOI bind-ing densities in the dorsal horn as an index of theresponse of 5HT2 receptors (44) also fails to show asignificant change. It remains possible that the re-sponse of 5HT receptors following either peripheralnerve lesion or deafferentation might be reflected at thelevel of signal transduction which would not be re-vealed by the technique of receptor binding autoradiog-raphy since receptor number and affinity would not bechanged (22).
The Ventral Horn
Injury to ventral roots results in chromatolysis ofmotoneurons with acute synaptic detachment and longlasting changes in dendritic morphology (6, 8, 26). Nochanges were found in binding densities for NK1 or5HT receptors, but the changes in the NMDA receptorin lamina IX were quite similar to those in the superfi-cial dorsal horn, with a bilateral and symmetricalincrease peaking at 2 weeks and returning to nearcontrol values by 20 weeks. As in the case of the dorsalhorn, there are reports of bilateral changes in motornuclei following a unilateral lesion. Following a unilat-eral hypoglossal nerve lesion, bilateral increases insuccinoxiadase activity (13), nucleolar and total cellRNA (39), and glial activation (37) have been shown. Asin the dorsal horn, monoarthritis results in a change inGluR1 gene expression in both ventral horns (35). Weassume that polysynaptic pathways may play a part ingenerating the bilateral ventral horn response as well.
In summary, sciatic nerve crush is associated withbiphasic changes in NK1 and NMDA receptor binding
densities in rat spinal cord. The changes in the NK1receptor occur ipsilateral to the lesion and are re-stricted to the dorsal horn. The NMDA receptor changesare bilateral, symmetrical, and occur in both dorsal andventral horns. We propose that the changes in the NK1receptor are due to changes in synthesis and transportof tachykinins by dorsal root ganglion cells. The bilat-eral changes in the NMDA receptor may reflect changesin synthesis and transport of glutamate and are prob-ably mediated by polysynaptic pathways.
REFERENCES
1. Aldskogius, H., J. Arvidsson, and G. Grant. 1985. The reactionof primary sensory neurons to peripheral nerve injury withparticular emphasis on transganglionic changes. Brain Res.Rev. 10: 27–46.
2. Barbut, D., J. M. Polak, and P. D. Wall. 1981. Substance P inspinal cord dorsal horn decreases following peripheral nerveinjury. Brain Res. 205: 289–298.
3. Behbehani, M. M., and O. Dollberg-Stolik. 1994. Partial sciaticnerve ligation results in an enlargement of the receptive fieldand enhancement of the response of dorsal horn neurons tonoxious stimulation by an adenosine agonist. Pain 58: 421–428.
4. Besse, D., J. Weil-Fugazza, M. C. Lombard, S. H. Butler, andJ. M. Besson. 1992. Monoarthritis induces complex changes inmu-, delta- and kappa-opioid binding sites in the superficialdorsal horn of the rat spinal cord. Eur. J. Pharmacol. 223:123–131.
5. Bisby, M. A., and P. Keen. 1986. Regeneration of primaryafferent neurons containing substance P-like immunoreactivity.Brain Res. 365: 85–95.
6. Bowe, C. M., C. H. Yu, and S. G. Waxman. 1988. Morphologicalchanges in spinal motor neurons giving rise to long-termregenerated sciatic nerve axons. Brain Res. 463: 69–77.
7. Brodin, E., B. Linderoth, B. Gazelius, and U. Ungerstedt. 1987.In vivo release of substance P in cat dorsal horn studied withmicrodialysis. Neurosci. Lett. 76: 357–362.
8. Chen, D. H. 1978. Qualitative and quantitative study of synap-tic displacement in chromatolyzed spinal motoneurons of thecat. J. Comp. Neurol. 177: 635–664.
9. Croul, S., A. Sverstiuk, A. Radzievsky, and M. Murray. 1995.Modulation of neurotransmitter receptors following unilateralL1-S2 deafferentation: NK1, NK3, NMDA, and 5HT1a receptorbinding autoradiography. J. Comp. Neurol. 361: 633–644.
10. Csillik, B., and E. Knyihar-Csillik. 1981. Regenerative synapto-neogenesis in the Mammalian spinal cord: Dynamics of synapto-chemical restoration in the Rolando Substance after transgangli-onic degenerative atrophy. J. Neural Trans. 52: 303–317.
11. De Biasi, S., and A. Rustioni. 1988. Glutamate and substance Pcoexist in primary afferent terminals in the superficial laminaeof spinal cord. Proc. Natl. Acad. Sci. USA 85: 7820–7824.
12. Goldberger, M. E., M. Murray, and A. Tessler. 1993. Sproutingand regeneration in the spinal cord. Their roles in recovery offunction after spinal injury. In Neuroregeneration (A. Gorio,Ed.), pp. 241–263. Raven Press, New York.
13. Hamberger, A., and J. Sjostrand. 1966. Respiratory enzymeactivities in neurons and glial cells of the hypoglossal nucleusduring nerve regeneration. Acta Physiol. Scand. 67: 76–88.
14. Helke, C. J., C. G. Charlton, and R. G. Wiley. 1986. Studies onthe cellular localization of spinal cord substance P receptors.Neuroscience 19: 523–533.
77NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH

15. Henken, D. B., W. P. Battisti, M. F. Chesselet, M. Murray, and A.Tessler. 1990. Expression of beta-preprotachykinin mRNA andtachykinins in rat dorsal root ganglion cells following peripheralor central axotomy. Neuroscience 39: 733–742.
16. Himes, B. T., and A. Tessler. 1989. Death of some dorsal rootganglion neurons and plasticity of others following sciatic nervesection in adult and neonatal rats. J. Comp. Neurol. 284:215–230.
17. Knyihar-Csillik, E., P. Rakic, and B. Csillik. 1985. Fine struc-ture of growth cones in the upper dorsal horn of the adultprimate spinal cord in the course of reactive synapto-neogen-esis. Cell Tissue Res. 239: 633–641.
18. Knyihar-Csillik, E., and A. Torok. 1989. Cytochemical restora-tion in the upper dorsal horn after transganglionic degenerativeatrophy: Temporospatial and fine structural correlates. Neuro-science 33: 75–91.
19. Knyihar-Csillik, E., A. Torok, and B. Csillik. 1990. Primaryafferent origin of substance P-containing axons in the superfi-cial dorsal horn of the rat spinal cord: Depletion, regenerationand replenishment of presumed nociceptive central terminals.J. Comp. Neurol. 297: 594–612.
20. Light, A. R., and E. R. Perl. 1979. Spinal termination offunctionally identified primary afferent neurons with slowlyconducting myelinated fibers. J. Comp. Neurol. 186: 133–150.
21. Light, A. R., and E. R. Perl. 1979. Reexamination of the dorsalroot projection to the spinal dorsal horn including observationson the differential termination of coarse and fine fibers. J.Comp. Neurol. 186: 117–131.
22. Lund, A., N. Mjellem-Jolly, and K. Hole. 1992. Desipramine,administered chronically, influences 5-Hydroxytryptamine 1areceptors, as measured by behavioral tests and receptor bindingin rats. Neuropharmacology 31: 25–32.
23. Mapp, P. I., G. Terenghi, D. A. Walsh, S. T. Chen, S. C. Cruwys,N. Garrett, B. L. Kidd, J. M. Polak, and D. R. Blake. 1993.Monoarthritis in the rat knee induces bilateral and time-dependent changes in substance P and calcitonin gene-relatedpeptide immunoreactivity in the spinal cord. Neuroscience 57:1091–1096.
24. Marlier, L., J. R. Teilhac, C. Cerruti, and A. Privat. 1991.Autoradiographic mapping of 5-HT1, 5-HT1A, 5-HT1B and5-HT2 receptors in the rat spinal cord. Brain Res. 550: 15–23.
25. Massari, V. J., C. W. Shults, C. H. Park, Y. Tizabi, T. W. Moody,B. M. Chronwall, M. Culver, and T. N. Chase. 1985. Deafferenta-tion causes a loss of presynaptic bombesin receptors and super-sensitivity of substance P receptors in the dorsal horn of the catspinal cord. Brain Res. 343: 268–274.
26. Matthews, M. R., and G. Raisman. 1972. A light and electronmicroscopic study of the cellular response to axonal injury in thesuperior cervical ganglion of the rat. Proc. R. Soc. London Ser. B:Biol. Sci. 181: 43–79.
27. McNeill, D. L., S. M. Carlton, and C. E. Hulsebosch. 1991.Intraspinal sprouting of calcitonin gene-related peptide contain-ing primary afferents after deafferentation in the rat. Exp.Neurol. 114: 321–329.
28. Merighi, A., J. M. Polak, and D. T. Theodosis. 1991. Ultrastruc-tural visualization of glutamate and aspartate immunoreactivi-ties in the rat dorsal horn, with special reference to theco-localization of glutamate, substance P and calcitonin-generelated peptide. Neuroscience 40: 67–80.
29. Molander, C., Q. Xu, and G. Grant. 1984. The cytoarchitectonicorganization of the spinal cord in the rat. I. The lower thoracicand lumbosacral cord. J. Comp. Neurol. 280: 133–141.
30. Monaghan, D. T., and C. W. Cotman. 1985. Distribution of
N-methyl-D-aspartate-sensitive L-[3H]glutamate-binding sitesin rat brain. J. Neurosci. 5: 2909–2919.
31. Murray, M., and M. E. Goldberger. 1986. Replacement ofsynaptic terminals in lamina II and Clarke’s nucleus afterunilateral lumbosacral dorsal rhizotomy in adult cats. J. Neuro-sci. 6: 3205–3217.
32. Nakata, Y., Y. Kusaka, and T. Segawa. 1979. Supersensitivity tosubstance P after dorsal root section. Life Sci. 24: 1651–1654.
33. Nielsch, U., M. A. Bisby, and P. Keen. 1987. Effect of cutting orcrushing the rat sciatic nerve on synthesis of substance P byisolated L5 dorsal root ganglia. Neuropeptides 10: 137–145.
34. Page, K. 1965. A stain for myelin using solochrome cyanin. J.Med. Lab. Technol. 22: 224.
35. Pellegrini-Giampietro, D. E., S. Fan, B. Ault, B. E. Miller, andR. S. Zukin. 1994. Glutamate receptor gene expression in spinalcord of arthritic rats. J. Neurosci. 14: 1576–1583.
36. Porro, C. A., M. Cavazzuti, A. Galetti, L. Sassatelli, and G. C.Barbieri. 1991. Functional activity mapping of the rat spinalcord during formalin-induced noxious stimulation. Neurosci-ence 41: 655–665.
37. Reisert, I., G. Wildemann, D. Grab, and C. Pilgrim. 1984. Theglial reaction in the course of axon regeneration: A stereologicalstudy of the rat hypoglossal nucleus. J. Comp. Neurol. 229:121–128.
38. Rich, K. M., H. K. Yip, P. A. Osborne, R. E. Schmidt, and E. M.Johnson, Jr. 1984. Role of nerve growth factor in the adultdorsal root ganglia neuron and its response to injury. J. Comp.Neurol. 230: 110–118.
39. Rossignol, S., and L. Gauthier. 1980. An analysis of mechanismscontrolling the reversal of crossed spinal reflexes. Brain Res.182: 31–45.
40. Sorkin, L. S., and D. J. Mcadoo. 1993. Amino acids and serotoninare released into the lumbar spinal cord of the anesthetized catfollowing intradermal capsaicin injections. Brain Res. 607:89–98.
41. Subramaniam, S., and P. McGonigle. 1991. Quantitative autora-diographic characterization of the binding of (1)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5, 10-imine ([3H]MK-801)in rat brain: Regional effects of polyamines. J. Pharmacol. Exp.Ther. 256: 811–819.
42. Tessler, A., E. Glazer, R. Artymyshyn, M. Murray, and M. E.Goldberger. 1980. Recovery of substance P in the cat spinal cordafter unilateral lumbosacral deafferentation. Brain Res. 191:459–470.
43. Thor, K. B., A. Blitz-Siebert, and C. J. Helke. 1992. Autoradio-graphic localization of 5HT1 binding sites in the medullaoblongata of the rat. Synapse 10: 185–205.
44. Thor, K. B., S. Nickolaus, and C. J. Helke. 1993. Autoradio-graphic localization of 5-hydroxytryptamine1A, 5-hydrox-ytryptamine1B and 5-hydroxytryptamine1C/2 binding sites inthe rat spinal cord. Neuroscience 55: 235–252.
45. Traub, R. J., A. Solodkin, and G. F. Gebhart. 1994. NADPH-diaphorase histochemistry provides evidence for a bilateral,somatotopically inappropriate response to unilateral hindpawinflammation in the rat. Brain Res. 647: 113–123.
46. Ulas, J., D. T. Monaghan, and C. W. Cotman. 1990. Plasticresponse of hippocampal excitatory amino acid receptors todeafferentation and reinnervation. Neuroscience 34: 9–17.
47. Wang, S. D., M. E. Goldberger, and M. Murray. 1991. Normaldevelopment and the effects of early rhizotomy on spinalsystems in the rat. Brain Res. Dev. Brain 57–69.
48. Weinberg, R. J., F. Conti, S. L. Van Eyck, P. Petrusz, and A.
78 CROUL ET AL.

Rustioni. 1987. Glutamate immunoreactivity in superficial lami-nae of rat dorsal horn and spinal trigeminal nucleus. Excit.Amino Acid Trans. 173–176.
49. Wright, D. M., and M. H. Roberts. 1978. Supersensitivity to asubstance P analogue following dorsal root section. Life Sci. 22:19–24.
50. Yashpal, K., T. V. Dam, and R. Quirion. 1991. Effects of dorsal
rhizotomy on neurokinin receptor sub-types in the rat spinalcord: A quantitative autoradiographic study. Brain Res. 552:240–247.
51. Zhang, B., M. E. Goldberger, and M. Murray. 1993. Proliferationof SP- and 5HT-containing terminals in lamina II of rat spinalcord following dorsal rhizotomy: Quantitative EM-immunocyto-chemical studies. Exp. Neurol. 123: 51–63.
79NK1, NMDA, AND 5HT RECEPTORS IN SPINAL CORD FOLLOWING SCIATIC CRUSH






![Differential Occupancy of Somatodendritic and Postsynaptic 5HT1A Receptors by Pindolol: A Dose-Occupancy Study with [11C]WAY 100635 and Positron Emission Tomography in Humans](https://static.fdokumen.com/doc/165x107/631c4aedd5372c006e045dcf/differential-occupancy-of-somatodendritic-and-postsynaptic-5ht1a-receptors-by-pindolol-1675040353.jpg)












