
Size, shape and hue modulate attraction and landing responses of the braconid parasitoid Fopius...
10
Size, shape and hue modulate attraction and landing responses of the braconid parasitoid Fopius arisanus to fruit odour-baited visual targets Jeanneth Pe ´rez • Julio C. Rojas • Pablo Montoya • Pablo Liedo • Francisco J. Gonza ´lez • Alfredo Castillo Received: 25 June 2011 / Accepted: 9 October 2011 / Published online: 16 November 2011 Ó International Organization for Biological Control (IOBC) 2011 Abstract Female parasitoids are guided by multi- sensory information, including chemical and physical cues during host location. In the present study, we investigated the behavioural responses of naı ¨ve Fopius arisanus (Sonan) females to visual targets baited with guava odour. In non-choice wind tunnel tests, the attraction and landing responses of parasitoids to spheres painted with different colours, and targets of different shapes and sizes were evaluated. Females were more frequently attracted and landed more often on dark yellow targets than on targets with other colours. There was no correlation between the bright- ness of each colour and the attraction or landing responses. In contrast, both responses were correlated with relative reflectance (hue) of the coloured targets. A positive correlation was observed between attraction and hue, and a negative correlation between landing and hue. F. arisanus was attracted to and landed more often on spheres than on other shape models. The attraction response of this parasitoid was affected by the size of the targets, with spheres of 10 and 12 cm diameter being more attractive than spheres of 8, 6 and 4 cm diameter. The fact that F. arisanus females were able to discriminate among visual targets that differ in colour, shape and size stresses the importance of vision during host location by this species. Keywords Visual cues Spectral reflectance Attraction Landing Host location Braconidae Introduction The host selection by parasitoids is a sequential process that can be divided in host habitat location, host location, and host acceptance (Vinson 1976; Fatouros et al. 2008). At long-range, parasitoids forage between plants used by their host within their habitat, Handling Editor: Torsten Meiners J. Pe ´rez (&) J. C. Rojas P. Liedo A. Castillo El Colegio de la Frontera Sur (ECOSUR), Carretera Antiguo Aeropuerto km 2.5, 30700 Tapachula, Chiapas, Me ´xico e-mail: [email protected] J. C. Rojas e-mail: [email protected] P. Liedo e-mail: [email protected] A. Castillo e-mail: [email protected] P. Montoya Subdireccio ´n de Desarrollo de Me ´todos, Programa Moscas de la Fruta DGSV SAGARPA, Central Poniente 14, 30700 Tapachula, Chiapas, Me ´xico e-mail: [email protected] F. J. Gonza ´lez Coordinacio ´n para la Innovacio ´n y la Aplicacio ´n de la Ciencia y la Tecnologı ´a, Universidad Auto ´noma de San Luis Potosı ´, Sierra Leona 550, Lomas 2da. Seccio ´n, 78210 San Luis Potosı ´, SLP, Me ´xico e-mail: [email protected] 123 BioControl (2012) 57:405–414 DOI 10.1007/s10526-011-9416-0
Transcript of Size, shape and hue modulate attraction and landing responses of the braconid parasitoid Fopius...

Size, shape and hue modulate attraction and landingresponses of the braconid parasitoid Fopius arisanus to fruitodour-baited visual targets
Jeanneth Perez • Julio C. Rojas • Pablo Montoya •
Pablo Liedo • Francisco J. Gonzalez • Alfredo Castillo
Received: 25 June 2011 / Accepted: 9 October 2011 / Published online: 16 November 2011! International Organization for Biological Control (IOBC) 2011
Abstract Female parasitoids are guided by multi-
sensory information, including chemical and physicalcues during host location. In the present study, we
investigated the behavioural responses of naıveFopiusarisanus (Sonan) females to visual targets baited withguava odour. In non-choice wind tunnel tests, the
attraction and landing responses of parasitoids to
spheres painted with different colours, and targets of
different shapes and sizes were evaluated. Femaleswere more frequently attracted and landed more often
on dark yellow targets than on targets with other
colours. There was no correlation between the bright-ness of each colour and the attraction or landing
responses. In contrast, both responses were correlatedwith relative reflectance (hue) of the coloured targets.
A positive correlation was observed between attraction
and hue, and a negative correlation between landingand hue. F. arisanus was attracted to and landed more
often on spheres than on other shape models. The
attraction response of this parasitoid was affected bythe size of the targets, with spheres of 10 and 12 cm
diameter being more attractive than spheres of 8, 6 and
4 cm diameter. The fact that F. arisanus females wereable to discriminate among visual targets that differ in
colour, shape and size stresses the importance of vision
during host location by this species.
Keywords Visual cues ! Spectral reflectance !Attraction ! Landing ! Host location ! Braconidae
Introduction
The host selection by parasitoids is a sequentialprocess that can be divided in host habitat location,
host location, and host acceptance (Vinson 1976;
Fatouros et al. 2008). At long-range, parasitoids foragebetween plants used by their host within their habitat,
Handling Editor: Torsten Meiners
J. Perez (&) ! J. C. Rojas ! P. Liedo ! A. CastilloEl Colegio de la Frontera Sur (ECOSUR), CarreteraAntiguo Aeropuerto km 2.5, 30700 Tapachula, Chiapas,Mexicoe-mail: [email protected]
J. C. Rojase-mail: [email protected]
P. Liedoe-mail: [email protected]
A. Castilloe-mail: [email protected]
P. MontoyaSubdireccion de Desarrollo de Metodos, ProgramaMoscas de la Fruta DGSV SAGARPA, Central Poniente14, 30700 Tapachula, Chiapas, Mexicoe-mail: [email protected]
F. J. GonzalezCoordinacion para la Innovacion y la Aplicacion de laCiencia y la Tecnologıa, Universidad Autonoma de SanLuis Potosı, Sierra Leona 550, Lomas 2da. Seccion, 78210San Luis Potosı, SLP, Mexicoe-mail: [email protected]
123
BioControl (2012) 57:405–414
DOI 10.1007/s10526-011-9416-0

at medium-range they search for the hosts within hostplants, and finally at short-range females recognize
and accept their hosts (Volkl 2000). During the host
selection process, parasitoids are guided by multisen-sory information, including chemical, vibratory and
visual cues, which are used in an interactive manner.
Chemical cues seem to play an important role in thedifferent steps within the host selection process,
whereas visual cues are thought to be important only
after females have been stimulated by host odours(Vinson 1976).
The role of chemical cues during the host selection
process has been studied extensively in parasitoids,but the use of vision during this process is less
understood for this group of insects. Visual informa-
tion may consist of variables such as colour, shape,and size, among others. These parameters have
different behavioural relevance during host habitat
location at distance, and during host location after theparasitoid has landed on a plant. At long-range, colour
seems to be the most important visual parameter,
while at short-range, shape and size are crucial for hostrecognition and host acceptance (Wackers and Lewis
1999). Colour can be described in terms of three main
attributes: hue (dominant wavelength of reflectedlight), brightness (intensity of perceived reflected
light) and saturation (spectral purity of reflected light)
(Prokopy and Owens 1983). True colour vision ininsects has only been confirmed for a limit number of
orders, among them Hymenoptera (Chittka and Men-
zel 1992; Kelber et al. 2003). Most of hymenopteranspecies investigated so far have trichromatic vision
with UV (340 nm), blue (430 nm), and green
(535 nm) receptors (Peitsch et al. 1992). The capacityfor colour discrimination has been reported for several
species of parasitoids (Messing and Jang 1992;
Battaglia et al. 2000). For example, colour plays animportant role during host recognition and oviposition
by the parasitoid Aphidius ervi Haliday (Battaglia
et al. 2000). In addition to colour, other visualparameters may be used by parasitoids during host
location, including the size and shape (Vinson 1976).Host size and shape may be visual cues highly
distinguishable in almost any habitat because mor-
phological differences exist not only between plantsbut also between structures of the same plant (Wackers
and Lewis 1999). The use of these parameters during
host location has been documented for differentparasitoid species. For instance, Diachasmimorpha
longicaudata (Ashmead) females showed no prefer-ence for colour or shape, but displayed a clear
preference for large over smaller fruit-mimicking
models (Segura et al. 2007).Fopius arisanus (Sonan) is an egg-pupal endopar-
asitoid that attacks several species of tephritid fruit
flies, particularly those in the genus Bactrocera.Currently, F. arisanus is known to parasitize 21 fruit
fly species, and can develop in at least 18 species
(Quimio and Walter 2001; Rousse et al. 2005, 2006).The successful introductions of this parasitoid to
Hawaii and French Polynesia for the control of
Ceratitis capitata (Wiedemann) and Bactrocera dor-salis (Hendel), respectively, suggest that F. arisanushas potential for augmentative biological control of
fruit flies, alone or in combination with other controlmethods (Harris et al. 2000).
Host location behaviour of F. arisanus has been
described by Wang and Messing (2003). Chemicalcues play important roles during host location behav-
iour in this wasp (Liquido 1991; Altuzar et al. 2004;
Rousse et al. 2007a). However, there are contrastingreports regarding the role of visual preferences during
host location (Vargas et al. 1991; Cornelius et al. 1999;
Rousse et al. 2007b). With the aim of clarifying therole of visual preferences in host location by F.arisanus, this study was undertaken to investigate the
behavioural responses of female wasps to guavaodour-baited targets of different colours, sizes and
shapes in a laboratory wind tunnel.
Materials and methods
Insects
Parasitoids used in this study were obtained fromcolonies established at MOSCAFRUT mass-rearing
facility (SAGARPA-IICA) located in Metapa de
Domınguez, Chiapas, Mexico. This strain of parasit-oids was imported from Hawaii in 1998, where it was
reared on C. capitata eggs (Bautista et al. 1999).Parasitoids used for experiments had been reared from
Anastrepha ludens (Loew) eggs placed in pieces of
papaya at 22 ± 2"C, 60–80% RH, and 12:12 L:D hphotoperiod as previously described (Montoya et al.
2009). The eggs ofA. ludenswere obtained from adults
reared on artificial diet according to establishedprotocols (Domınguez et al. 2010). After emergence,
406 J. Perez et al.
123
parasitoids of both sexes were placed in wooden framecages 30 9 30 9 30 cm to allow mating. Adults were
given free access to honey and water dispensed on
cottonwool. One hour before the experiments, femaleswere placed individually in plastic containers (5 cm
high by 4 cm diameter) and allowed to acclimatize to
the wind tunnel room conditions. In all experiments,7–9 days old females were used. Parasitoids had no
prior contact with their hosts or host fruits.
Bioassays
The observations were performed in a wind tunnel(120 cm long 9 30 cm high 9 30 cm wide). The
wind tunnel was constructed using 1 cm thick clear
plexiglass. A fan was used to pull air, filtered byactivated charcoal, through the tunnel at a speed of
0.4 m s-1. Illumination was provided by four fluores-
cent tubes mounted 60 cm above the wind tunnelgiving a light intensity of 2380 lx. Green dots of
10 cm diameter made from cardboard were placed on
the tunnel floor to provide optomotor feedback for theparasitoids. Fruit odour consisted of five green guavas
(325.7 ± 13.2 g) (mean ± SD) without insect infes-
tation, placed inside a polyethylene bag that wassituated outside the wind tunnel. Filtered air
(1 l min-1) was passed through the bag and the odour
was conducted into the wind tunnel by means of tygontubing (0.4 cm internal diameter). The end of the tube
was connected to a hole (0.7 cm) in the centre of the
visual target from which odour emanated at a flow rateof 0.6 l min-1. The target stimulus was hung in the
centre of the wind tunnel, 16 cm from the upwind end.
The distance between platform release and the targetstimulus was about 1 m.
Each observation started by placing the plastic
container with one female on a 12 cm high platform atthe downwind end of the tunnel. The parasitoid was
released and observed for 5 min. Female behaviour
was recorded, specifically the zigzagging upwindflight towards the target stimulus and landing on the
target after such upwind flight. If the parasitoid did nottake off or did not show zigzagging upwind flight after
5 min, the test was stopped and the insect was
considered to be a non-responding individual. Here-after, the term attraction is used to indicate such
zigzagging upwind flights. Parasitoids were used
once. In all experiments, the responses of parasitoidsto the visual targets were evaluated in non-choice
tests. All bioassays were conducted between 08:00 hand 14:00 h at 24 ± 2"C, 65 ± 5% RH.
Response to models of different colours
The responses of F. arisanus females to styrofoamspheres of different colours baited with guava odour
were evaluated. Preliminary observations showed thatfemales were not attracted to spheres without the
odour. Spheres of 8 cm diameter were painted with the
following colours of acrylic paint (Comex Group,Mexico): white, black, brown, red, blue, light green,
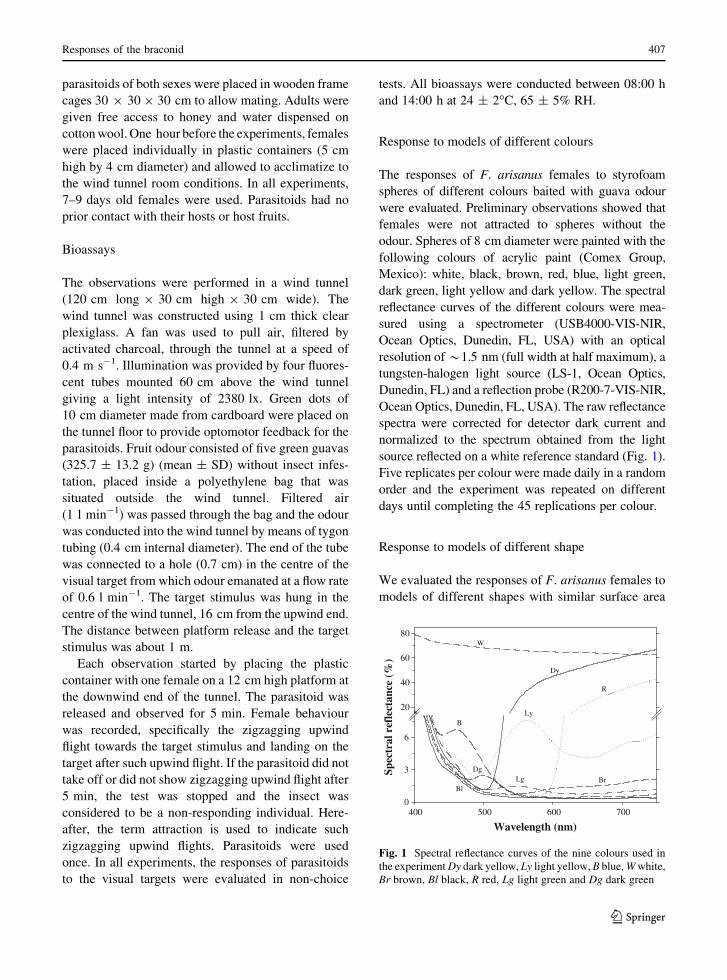
dark green, light yellow and dark yellow. The spectral
reflectance curves of the different colours were mea-sured using a spectrometer (USB4000-VIS-NIR,
Ocean Optics, Dunedin, FL, USA) with an optical
resolution of*1.5 nm (full width at half maximum), atungsten-halogen light source (LS-1, Ocean Optics,
Dunedin, FL) and a reflection probe (R200-7-VIS-NIR,
Ocean Optics, Dunedin, FL, USA). The raw reflectancespectra were corrected for detector dark current and
normalized to the spectrum obtained from the light
source reflected on a white reference standard (Fig. 1).Five replicates per colour were made daily in a random
order and the experiment was repeated on different
days until completing the 45 replications per colour.
Response to models of different shape
We evaluated the responses of F. arisanus females tomodels of different shapes with similar surface area
Wavelength (nm)400 500 600 700
Spec
tral
ref
lect
ance
(%)
0
3
6
20
40
60
80W
Dy
R
LyB
DgBrLg
Bl
Fig. 1 Spectral reflectance curves of the nine colours used inthe experimentDy dark yellow, Ly light yellow, B blue,Wwhite,Br brown, Bl black, R red, Lg light green and Dg dark green
Responses of the braconid 407
123

(313 cm2). All models were made using styrofoamand were painted dark yellow. This colour was chosen
because it was the most attractive to F. arisanusfemales in the previous experiment. The modelsevaluated had either three-dimensional (sphere, cube,
pyramid, and ellipse) or flat shapes (circle, square,
rectangle, and triangle). Five replicates per shape wereperformed daily in a random order and the experiment
was repeated on different days until completing 45
replicates per shape.
Response to models of different size
The responses of F. arisanus females to dark yellow
spheres of 14, 12, 10, 8, 6, and 4 cm diameter were
tested in this experiment. The size of spheres werechosen on the basis of previous studies with this
species and considering the range of size of fruits used
by F. arisanus hosts. Five replicates per size weremade daily in a random order and the experiment was
repeated on different days until completing 45 repli-
cates per size.
Statistical analysis
We used a log likelihood ratio contingency-table
analysis to test whether responses (attraction or
landing) differed among treatments. Fisher’s exacttest was used when counts were low. When significant
effects were found, odds ratios were used to make
comparisons between treatments.In the first experiment, numbers of attraction and
landing responses were correlated with the brightness
and hues of visual targets, using methods describedelsewhere (Katsoyannos and Kouloussis 2001).
According to this procedure whether the insect
response to coloured target depends only on bright-ness, a strong correlation between the frequency of
responses and total brightness was expected. If the
insect response depends on the specific wavelengths(hues), then the response should correlate with relative
reflectance values of the colours. Relative reflectancewas considered to be the amount of light reflected by a
colour within a given wavelength interval relative to
the total amount of light reflected by this colour withinthe range of the spectrum considered.
The total reflectance values of the colours used in this
study were obtained as percentages of the total amountof light reflected over a white reference sphere between
350 and 750 nm. Reflectance values were: white =100, dark yellow = 51, light yellow = 14.1, dark
green = 7.9, light green = 9.7, blue = 9, brown =
8.9, black = 7.1, and red = 24. Relative reflectancewere calculated for every colour as the amount of light
reflected by each successive 10 nm wide wavelength
interval, in relation to the total amount of light reflectedby that colour between 350 and 750 nm.
Results
Response to models of different colours
Colour influenced the attraction of F. arisanus females
towards the spheres baited with fruit odour (G = 21.1,df = 8, P = 0.007). Females were more attracted to
dark yellow models than to light yellow, blue, black,
red, light green, and dark green models (Fig. 2a).There were no differences in the numbers of females
that were attracted to dark yellow, white, and brown
targets. The landing behaviour of parasitoids on thetargets was also influenced by colour (G = 67.44,
df = 8, P\ 0.001). Females landed more often on the
dark yellow, light yellow, brown, black, red, and lightgreen models than on blue, white and dark green
targets (Fig. 2b).
There was no correlation between the total amountof reflected light of each colour and attraction
(r = 0.44) or landing (r = -0.399) responses (P[0.05). In contrast, both responses were correlated withrelative reflectance (hues) of the coloured targets. A
positive correlation was detected between attraction
and hue, and a negative correlation between landingand hue (Fig. 3). Females were more attracted to hues
reflecting maximally between 550 and 620 nm (dark
yellow), with a maximum peak at 600 nm (r = 0.795).Parasitoids landed less often on hues reflecting
maximally between 420 and 510 nm (blue, light
green), with a minimal peak at 490 nm (r = -0.943).
Response to models of different shape
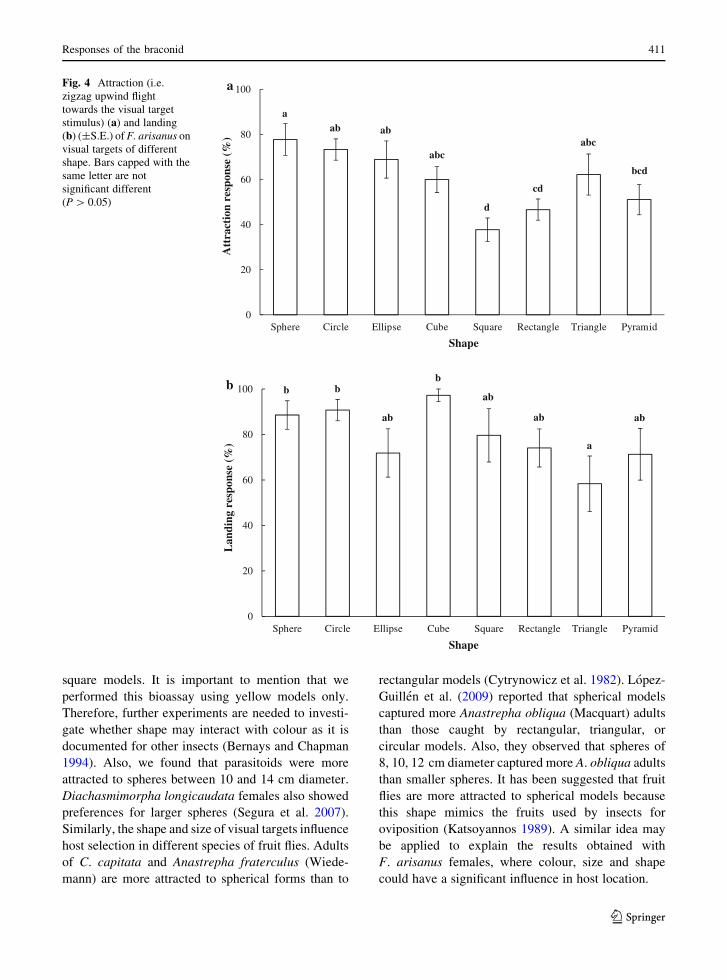
The shape of visual targets affected attraction
(G = 24.2, df = 7, P = 0.001) and landing (G =19.0, df = 7, P = 0.008) responses of F. arisanus.Parasitoids were more attracted to spheres than to
square, rectangle, or pyramid models. Attraction ofF. arisanus females to spheres was not significantly
408 J. Perez et al.
123

different from that of circle, ellipse, cube and triangle
models (Fig. 4a). Parasitoids landed more often on
sphere, circle, and cube models than on the trianglemodel. The landing response on ellipse, square,
rectangle and pyramid were intermediate and not
significantly different from that on sphere, circle andcube or triangle (Fig. 4b).
Response to models of different size
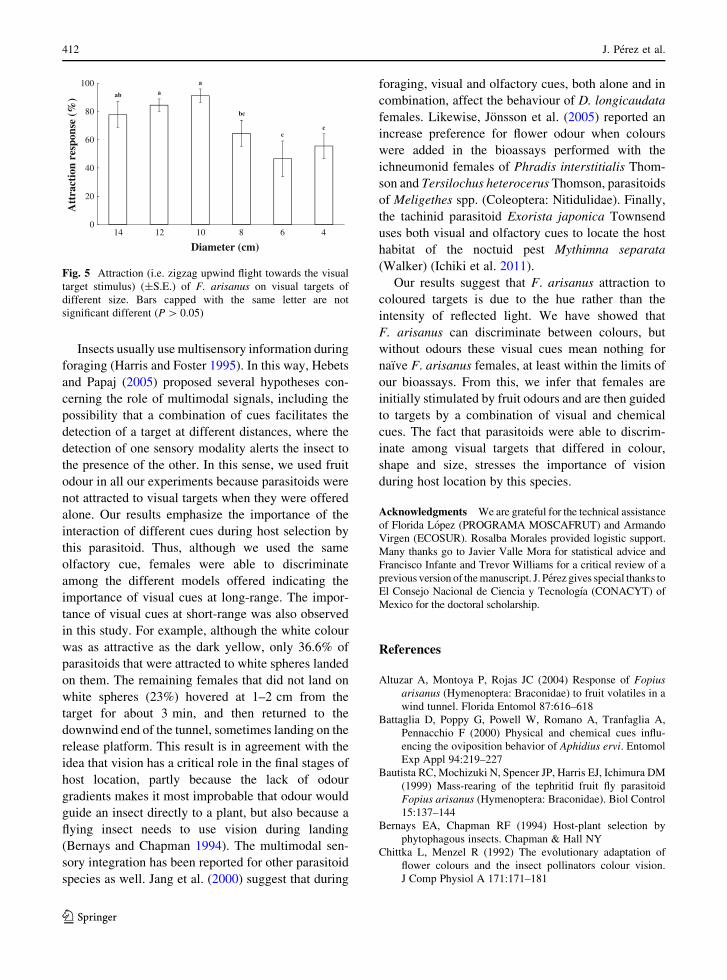
The size of the visual targets affected attraction of
F. arisanus females (G = 33.7, df = 5, P\ 0.001).In contrast, the landing response was not affected by
the size of the models tested (Fisher test, P = 0.76).
Parasitoids were more attracted to spheres of 10 and
12 cm diameter than to spheres of 4, 6 and 8 cmdiameter. The attraction of F. arisanus to spheres of
14 cm diameter was intermediate and not significantly
different from that to spheres of 10 and 12 cmdiameter (Fig. 5).
Discussion
We showed that some visual cues in combination withfruit odour might play a role in the host location
0
20
40
60
80
100
DarkYellow
LightYellow Green
DarkGreen
Att
ract
ion
resp
onse
(%)
Colour
bc
a
bc bcbc
bc
c
ab ab
a
0
20
40
60
80
100
DarkYellow
LightYellow
Blue White Brown Black Red Light
Blue White Brown Black Red LightGreen
DarkGreen
Lan
ding
res
pons
e (%
)
Colour
a
a a
aa
a
b
b
b
b
Fig. 2 Attraction (i.e.zigzag upwind flighttowards the visual targetstimulus) (a) and landing(b) (±S.E.) of F. arisanusfemales on visual targets ofdifferent colour. Barscapped with the same letterare not significant different(P[ 0.05)
Responses of the braconid 409
123

behaviour of naıve F. arisanus females. We found thatfemales of this parasitoid were attracted and landed
more often on yellow spheres compared with spheres
of other colours. Previously, Vargas et al. (1991)reported that yellow and orange traps captured more
F. arisanus than those caught by blue and black traps.
Rousse et al. (2007b) showed that naıve F. arisanusfemales landed less often on yellow and white spheres
than on the black and dark green spheres. These
authors concluded that F. arisanus females respondmainly to achromatic cues. However, the response that
we obtained towards the dark yellow and brown
spheres showed that they were equally attractive toF. arisanus, although they differ widely in the amount
of reflected light, suggesting that for this species the
hue is more important than brightness. An explanationfor differences between both studies may be due to the
experimental protocols. In our case, experiments were
performed using fruit odour in combination withvisual targets, and this may have affected the parasit-
oid responses. Thus, we consider that the response of
F. arisanus females to odour-baited visual targets maybe related to host foraging, while the responses of the
parasitoids reported by Rousse et al. (2007b) may only
imply the ability of females to detect and respond tovisual targets, without direct link to the host location
process. On other hand, our bioassays were performed
in a wind tunnel with controlled illumination, such thatthe amount of light received by the coloured visual
targets of the different treatments was constant
throughout the whole experiment. Rousse et al.
(2007b) performed experiments in field cages wherethe amount of light received by each visual target
likely varied depending on their position relative to the
sun and the target, and variation in solar and clima-tological condition, thus influencing the parasitoid’s
responses. Indeed, the received colour of an object
depends upon the interaction between the ambientlight and the reflectance colour of the object. Thus, an
object may have a different appearance in each
environment (Endler 1993). Another reason is that inour study the visual targets were not placed against a
particular background (black, white, gray). Consider-
ing that insects likely perceive foliage as gray, thusagainst this achromatic background, any different
wavelength other than that of foliage will produce at
least some colour contrast (Chittka et al. 1994).The preference for the colour yellow we found in
F. arisanus has been reported for other hymenopterans,
including egg parasitoids (Lobdell et al. 2005). In thecase of fruit fly parasitoids, Diachasmimorpha juglan-dis (Muesebeck) chose yellow walnut fruits over black
fruits despite that the latter are more likely to containhost larvae (Henneman 1998). In field cage tests,
D. longicaudata females discriminated between vari-
ous spheres painted with different colours with yellowas the most attractive colour (Messing and Jang 1992).
In the same way, several studies have shown that
yellow colour is highly attractive to different species offruit flies (Katsoyannos 1987; Vargas et al. 1991; Drew
et al. 2003; Pinero et al. 2006; Lopez-Guillen et al.
2009). For example, C. capitata females were mainlyattracted to yellow, followed by red, orange, black, and
green spheres, whereaswhite and blue sphereswere the
least attractive (Katsoyannos 1987).The shape and size of targets are other visual
parameters that can be used during host location by
parasitoids. The hue and brightness of visual targetscan be perceived before insect landing, whereas the
shape and size gain in importance at lesser distances.
Vinson (1976) pointed out that size and shape areusually secondary stimuli during foraging, and
become important during host acceptance. However,in some cases these stimuli seem to be crucial during
host location. For instance, Henneman et al. (2002)
suggested that at distance,D. juglandis females tend toignore the odour and use visual cues, because the
odour plume is turbulent and difficult to locate the host
fruit. We have found that F. arisanus females showedbetter responses to spherical than to rectangular or
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
350 390 430 480 520 560 600 650 690 740
Cor
rela
tion
coe
ffic
ient
(r)
Wavelength (nm)
Attraction Landing
P < 0.05
P < 0.05
Fig. 3 Correlation coefficient between the responses (attrac-tion and landing) of F. arisanus females to targets of ninedifferent colours, and the relative reflectance values of eachcolour at successive 10 nm wavelength, from 350 to 750 nm.Statistical differences are indicated above or below of the dash-lines
410 J. Perez et al.
123
square models. It is important to mention that we
performed this bioassay using yellow models only.Therefore, further experiments are needed to investi-
gate whether shape may interact with colour as it is
documented for other insects (Bernays and Chapman1994). Also, we found that parasitoids were more
attracted to spheres between 10 and 14 cm diameter.
Diachasmimorpha longicaudata females also showedpreferences for larger spheres (Segura et al. 2007).
Similarly, the shape and size of visual targets influence
host selection in different species of fruit flies. Adultsof C. capitata and Anastrepha fraterculus (Wiede-
mann) are more attracted to spherical forms than to
rectangular models (Cytrynowicz et al. 1982). Lopez-
Guillen et al. (2009) reported that spherical modelscaptured more Anastrepha obliqua (Macquart) adults
than those caught by rectangular, triangular, or
circular models. Also, they observed that spheres of8, 10, 12 cm diameter captured more A. obliqua adultsthan smaller spheres. It has been suggested that fruit
flies are more attracted to spherical models becausethis shape mimics the fruits used by insects for
oviposition (Katsoyannos 1989). A similar idea may
be applied to explain the results obtained withF. arisanus females, where colour, size and shape
could have a significant influence in host location.
0
20
40
60
80
100
Att
ract
ion
resp
onse
(%)
Shape
a
abab
abc
d
cd
abc
bcd
a
0
20
40
60
80
100
Sphere Circle Ellipse Cube Square Rectangle Triangle Pyramid
Sphere Circle Ellipse Cube Square Rectangle Triangle Pyramid
Lan
ding
res
pons
e (%
)
Shape
b
ab
a
ab
ab
ab
bbb
Fig. 4 Attraction (i.e.zigzag upwind flighttowards the visual targetstimulus) (a) and landing(b) (±S.E.) ofF. arisanus onvisual targets of differentshape. Bars capped with thesame letter are notsignificant different(P[ 0.05)
Responses of the braconid 411
123
Insects usually use multisensory information duringforaging (Harris and Foster 1995). In this way, Hebets
and Papaj (2005) proposed several hypotheses con-
cerning the role of multimodal signals, including thepossibility that a combination of cues facilitates the
detection of a target at different distances, where the
detection of one sensory modality alerts the insect tothe presence of the other. In this sense, we used fruit
odour in all our experiments because parasitoids were
not attracted to visual targets when they were offeredalone. Our results emphasize the importance of the
interaction of different cues during host selection by
this parasitoid. Thus, although we used the sameolfactory cue, females were able to discriminate
among the different models offered indicating the
importance of visual cues at long-range. The impor-tance of visual cues at short-range was also observed
in this study. For example, although the white colour
was as attractive as the dark yellow, only 36.6% ofparasitoids that were attracted to white spheres landed
on them. The remaining females that did not land on
white spheres (23%) hovered at 1–2 cm from thetarget for about 3 min, and then returned to the
downwind end of the tunnel, sometimes landing on the
release platform. This result is in agreement with theidea that vision has a critical role in the final stages of
host location, partly because the lack of odour
gradients makes it most improbable that odour wouldguide an insect directly to a plant, but also because a
flying insect needs to use vision during landing(Bernays and Chapman 1994). The multimodal sen-
sory integration has been reported for other parasitoid
species as well. Jang et al. (2000) suggest that during
foraging, visual and olfactory cues, both alone and incombination, affect the behaviour of D. longicaudatafemales. Likewise, Jonsson et al. (2005) reported an
increase preference for flower odour when colourswere added in the bioassays performed with the
ichneumonid females of Phradis interstitialis Thom-
son and Tersilochus heterocerus Thomson, parasitoidsof Meligethes spp. (Coleoptera: Nitidulidae). Finally,the tachinid parasitoid Exorista japonica Townsend
uses both visual and olfactory cues to locate the hosthabitat of the noctuid pest Mythimna separata(Walker) (Ichiki et al. 2011).
Our results suggest that F. arisanus attraction tocoloured targets is due to the hue rather than the
intensity of reflected light. We have showed that
F. arisanus can discriminate between colours, butwithout odours these visual cues mean nothing for
naıve F. arisanus females, at least within the limits of
our bioassays. From this, we infer that females areinitially stimulated by fruit odours and are then guided
to targets by a combination of visual and chemical
cues. The fact that parasitoids were able to discrim-inate among visual targets that differed in colour,
shape and size, stresses the importance of vision
during host location by this species.
Acknowledgments We are grateful for the technical assistanceof Florida Lopez (PROGRAMA MOSCAFRUT) and ArmandoVirgen (ECOSUR). Rosalba Morales provided logistic support.Many thanks go to Javier Valle Mora for statistical advice andFrancisco Infante and Trevor Williams for a critical review of aprevious version of themanuscript. J. Perez gives special thanks toEl Consejo Nacional de Ciencia y Tecnologıa (CONACYT) ofMexico for the doctoral scholarship.
References
Altuzar A, Montoya P, Rojas JC (2004) Response of Fopiusarisanus (Hymenoptera: Braconidae) to fruit volatiles in awind tunnel. Florida Entomol 87:616–618
Battaglia D, Poppy G, Powell W, Romano A, Tranfaglia A,Pennacchio F (2000) Physical and chemical cues influ-encing the oviposition behavior of Aphidius ervi. EntomolExp Appl 94:219–227
Bautista RC, Mochizuki N, Spencer JP, Harris EJ, Ichimura DM(1999) Mass-rearing of the tephritid fruit fly parasitoidFopius arisanus (Hymenoptera: Braconidae). Biol Control15:137–144
Bernays EA, Chapman RF (1994) Host-plant selection byphytophagous insects. Chapman & Hall NY
Chittka L, Menzel R (1992) The evolutionary adaptation offlower colours and the insect pollinators colour vision.J Comp Physiol A 171:171–181
0
20
40
60
80
100
14 12 10 8 6 4
Att
ract
ion
resp
onse
(%)
Diameter (cm)
bc
aab
a
cc
Fig. 5 Attraction (i.e. zigzag upwind flight towards the visualtarget stimulus) (±S.E.) of F. arisanus on visual targets ofdifferent size. Bars capped with the same letter are notsignificant different (P[ 0.05)
412 J. Perez et al.
123

Chittka L, Shmida A, Troje N, Menzel R (1994) Ultraviolet as acomponent of flower reflections, and the colour perceptionof Hymenoptera. Vision Res 34:1489–1508
Cornelius ML, Duan JJ, Messing RH (1999) Visual stimuli andthe response of female oriental fruit fly (Diptera: Tephri-tidae) to fruit-mimicking traps. J Econ Entomol 92:121–129
Cytrynowicz M, Morgante JS, de Souza HML (1982) Visualresponse of South American fruit flies, Anastrepha frat-erculus, and Mediterranean fruit flies, Ceratitis capitata, tocolored rectangles and spheres. Environ Entomol 11:1202–1210
Domınguez J, Artiaga-Lopez T, Solıs E, Hernandez E (2010)Metodos de colonizacion y crıa masiva. In: Montoya P,Toledo J, Hernandez E (eds) Moscas de la fruta: Funda-mentos y procedimientos para su manejo, Mexico,pp 259–276
Drew RAI, Prokopy RJ, Romig MC (2003) Attraction of fruitflies of the genus Bactrocera to colored mimics of hostfruit. Entomol Exp Appl 107:39–45
Endler JA (1993) The color of light in forests and its implica-tions. Ecol Monogr 63:1–27
Fatouros NE, Dicke M, Mumm R, Meiners T, Hilker M (2008)Foraging behavior of egg parasitoids exploiting chemicalinformation. Behav Ecol 19:677–685
Harris MO, Foster SP (1995) Behavior and integration. In:Carde RT, Bell W (eds) Chemical ecology of insects, 2ndedn. Chapman & Hall, NY, pp 3–46
Harris EJ, Bautista RC, Spencer JP (2000) Utilisation of the egg-larval parasitoid, Fopius (Biosteres) arisanus, for augmen-tative biological control of tephritid fruit flies. In: Tan KH(ed) Area-wide control of fruit flies and other insect pests.Penerbit Universiti Sains Malaysia, Penang, pp 725–732
Hebets EA, Papaj DR (2005) Complex signal function: devel-oping a framework of testable hypotheses. Behav EcolSociobiol 57:197–214
Henneman ML (1998) Maximization of host encounters byparasitoids foraging in the field: females can use a simplerule. Oecologia 116:467–474
HennemanML, Dyreson EG, Takabayashi J, Raguso RA (2002)Response to walnut olfactory and visual cues by the para-sitic wasp Diachasmimorpha juglandis. J Chem Ecol 28:2221–2244
Ichiki RT, Kainoh Y, Yamawaki Y, Nakamura S (2011) Theparasitoid fly Exorista japonica uses visual and olfactorycues to locate herbivore-infested plants. Entomol Exp Appl138:175–183
Jang EB, Messing RH, Klungness LM, Carvalho LA (2000)Flight tunnel response of Diachasmimorpha longicaudata(Ashmead) (Hymenoptera: Braconidae) to olfactory andvisual stimuli. J Insect Behav 13:525–538
Jonsson M, Lindkvist A, Anderson P (2005) Behaviouralresponses in three ichneumonid pollen beetle parasitoids tovolatiles emitted from different phenological stages ofoilseed rape. Entomol Exp Appl 115:363–369
Katsoyannos BI (1987) Effect of color properties of spheres ontheir attractiveness for Ceratitis capitata (Wiedemann)flies in the field. J Appl Entomol 104:79–85
Katsoyannos BI (1989) Response to shape, size and color. In:Robinson AS, Hooper G (eds) Fruit flies: their biology,
natural enemies and control, vol 3A. Elsevier, Amsterdam,pp 307–324
Katsoyannos BI, Kouloussis NA (2001) Captures of the olivefruit fly Bactrocera oleae on spheres of different colours.Entomol Exp Appl 100:165–172
Kelber A, Vorobyev M, Osorio D (2003) Animal colour vision-behavioural tests and physiological concepts. Biol Rev 78:81–118
Liquido NJ (1991) Effect of ripeness and location of papayafruits on the parasitization rates of oriental fruit fly andmelon fly (Diptera: Tephritidae) by Braconid (Hymenop-tera) parasitoids. Environ Entomol 20:1732–1736
Lobdell CE, Yong TH, Hoffmann MP (2005) Host color pref-erences and short range searching behavior of the eggparasitoid Trichogramma ostriniae. Entomol Exp Appl116:127–134
Lopez-Guillen G, Valle-Mora J, Llanderal-Cazares C, Rojas J(2009) Response of Anastrepha obliqua (Diptera: Tephri-tidae) to visual and chemical cues under seminatural con-ditions. J Econ Entomol 102:954–959
Messing RH, Jang EB (1992) Response of the fruit fly parasitoidDiachasmimorpha longicaudata (Hymenoptera: Braconi-dae) to host fruit stimuli. Environ Entomol 21:1189–1195
Montoya P, Suarez A, Lopez F, Cancino J (2009) Fopius aris-anus oviposition in four Anastrepha fruit fly species ofeconomic importance in Mexico. BioControl 54:437–444
Peitsch D, Fietz A, Hertel H, de Souza J, Ventura DF, Menzel R(1992) The spectral input systems of hymenopteran insectsand their receptor-based colour vision. J Comp Physiol A170:23–40
Pinero JC, Jacome I, Vargas R, Prokopy RJ (2006) Response offemale melon fly Bactrocera cucurbitae, to host-associatedvisual and olfactory stimuli. Entomol Exp Appl 121:261–269
Prokopy RJ, Owens ED (1983) Visual detection of plants byherbivorous insects. Annu Rev Entomol 28:337–364
Quimio GM, Walter GH (2001) Host preference and host suit-ability in an egg-pupal fruit fly parasitoid Fopius arisanus(Sonan) (Hym., Braconidae). J Appl Entomol 125:135–140
Rousse P, Harris EJ, Quilici S (2005) Fopius arisanus, an egg-pupal parasitoid of Tephritidae. Overv Biocontr News Info26:59–69
Rousse P, Gourdon F, Quilici S (2006) Host specificity of theegg pupal parasitoid Fopius arisanus (Hymenoptera: Bra-conidae) in La Reunion. Biol Control 37:284–290
Rousse P, Chiroleu F, Domerg C, Quilici S (2007a) NaıveFopius arisanus females respond mainly to achromaticcues. Biol Control 43:41–48
Rousse P, Chiroleu F, Veslot J, Quilici S (2007b) The host andmicrohabitat olfactory location by Fopius arisanus sug-gests a broad potential host range. Physiol Entomol 32:313–321
Segura DF, Viscarret MM, Carabajal-Paladino LZ, Ovruski SM,Cladera JL (2007) Role of visual information and learningin habitat selection by a generalist parasitoid foraging forconcealed hosts. Anim Behav 74:131–142
Vargas RI, Stark JD, Prokopy RJ, Green TA (1991) Response oforiental fruit fly (Diptera: Tephritidae) and associatedparasitoids (Hymenoptera: Braconidae) to different colorspheres. J Econ Entomol 84:1503–1507
Responses of the braconid 413
123

Vinson SB (1976) Host selection by insect parasitoids. AnnuRev Entomol 21:109–133
Volkl W (2000) Foraging behavior and sequential multisensoryorientation in the aphid parasitoid Pauesia picta (Hym.,Aphidiidae) at different spatial scales. J Appl Entomol124:307–314
Wackers FL, Lewis WJ (1999) A comparison of color, shape,and pattern-learning by the hymenopteran parasitoid Mi-croplitis croceipes. J Comp Physiol A 184:387–393
Wang XG, Messing RH (2003) Foraging behavior and patchtime allocation by Fopius arisanus (Hymenoptera: Bra-conidae), an egg-larval parasitoid of tephritid fruit flies.J Insect Behav 16:593–612
Author Biographies
Jeanneth Perez Received her MSc degree from El Colegio dela Frontera Sur, Mexico. At present she is a Ph. D candidate atthe same institution. Her research interests are chemicalecology, insect behaviour and biological control of tropicalinsects.
Julio C. Rojas Received his PhD from Oxford University in1998. He is a research entomologist at the Department ofTropical Entomology, El Colegio de la Frontera Sur, Mexico.His interests include the behaviour and the chemical ecology ofherbivorous and carnivorous insects.
Pablo Montoya Doctor in Sciences from the UniversidadNacional Autonoma de Mexico in 1999. He is a Sub-Director
and researcher in the Moscafrut Program SAGARPA-IICA ofMexico. He has experience in the management of fruit flies,particularly in biological control, detection of populations, andthe application of the Sterile Insect Technique.
Pablo Liedo Ph. D. in Entomology, University of California -Davis (1989). He is a research entomologist at the Departmentof Tropical Entomology, El Colegio de la Frontera Sur,Mexico. His area of interest is insect ecology and pest control,with particular emphasis on biodemography, population ecol-ogy, biological control and the Sterile Insect Technique.Research work has been focused on fruit flies.
Francisco J. Gonzalez Ph.D. in electrical engineering from theSchool of Optics and Photonics, University of Central Florida,Orlando, in 2003. He is currently a Professor at the UniversidadAutonoma de San Luis Potosı, Mexico. His research interestsare the application of physics, optics and engineering in non-invasive medical diagnosis.
Alfredo Castillo Doctor in Science from El Colegio dePosgraduados in 2005. He is an entomologist at the Departmentof Tropical Entomology, El Colegio de la Frontera Sur,Mexico. He conducts research on ecology and biology oftropical insects.
414 J. Perez et al.
123




















