
Simultaneous characterization of progenitor cell compartments in adult human liver
10
Simultaneous Characterization of Progenitor Cell Compartments in Adult Human Liver Laura Porretti, 1 * Alessandra Cattaneo, 1 Federico Colombo, 1 Raffaella Lopa, 1 Giorgio Rossi, 2 Vincenzo Mazzaferro, 3 Carlo Battiston, 3 Gianluca Svegliati-Baroni, 4 Francesco Bertolini, 5 Paolo Rebulla, 1 Daniele Prati 1,6 Abstract The human liver is a complex tissue consisting of epithelial, endothelial, hematopoietic, and mesenchymal elements that probably derive from multiple lineage-committed pro- genitors, but no comprehensive study aimed at identifying and characterizing intrahe- patic precursors has yet been published. Cell suspensions for this study were obtained by enzymatic digestion of liver specimens taken from 20 patients with chronic liver dis- ease and 13 multiorgan donors. Stem and progenitor cells were first isolated, amplified, and characterized ex vivo according to previously validated methods, and then opti- mized flow cytometry was used to assess their relative frequencies and characterize their immunophenotypes in the clinical specimens. Stem and progenitor cells committed to hematopoietic, endothelial, epithelial, and mesenchymal lineages were clearly identifia- ble in livers from both healthy and diseased subjects. Within the mononuclear liver cell compartment, epithelial progenitors [epithelial cell adhesion molecule (EpCAM) 1 / CD49f 1 /CD29 1 /CD45 2 ] accounted for 2.7–3.5% whereas hematopoietic (CD34 1 / CD45 1 ), endothelial [vascular endothelial growth factor-2 (KDR) 1 /CD146 1 /CD45 2 ], and mesenchymal [CD73 1 /CD105 1 /CD90 (Thy-1) 1 /CD45 2 ] stem cells and progeni- tors accounted for smaller fractions (0.02–0.6%). The patients’ livers had higher per- centages of hematopoietic and endothelial precursors than those of the donors. In con- clusion, we identified and characterized precursors committed to four different lineages in adult human liver. We also optimized a flow cytometry approach that will be useful in exploring the contribution of these cells to the pathogenesis of liver disease. ' 2009 International Society for Advancement of Cytometry Key terms stem and progenitor cells; human liver; flow cytometry ADULT human liver is a complex tissue consisting of epithelial, endothelial, hemato- poietic, and mesenchymal elements. This supports the view that the intrahepatic stem and progenitor cell compartment probably consists of a heterogeneous pool with various molecular and functional markers. Studies characterizing hepatic stem cells and progenitors have so far mainly concentrated on the epithelial compartment, and generally used human fetal or rodent liver (1,2); only a few have identified and characterized the hematopoietic (3,4), endothelial (5), and mesenchymal lineages (6), although it is well known that their progenies are deeply involved in the pathoge- nesis of liver disease (7). Despite recent growth of interest in liver stem and progenitor cell research and great expectations concerning the potential of these cells in the field of regenerative medicine (8), we are unaware of comprehensive studies of the entire stem/progenitor cell pool. This could be due to the rare occurrence of these cells in liver tissue, and the possible overlap in the immunological profiles of the cells committed to different lineages (9–11), thus making their identification difficult. 1 Center of Transfusion Medicine, Cellular Therapy and Cryobiology, IRCCS Fondazione Ospedale Maggiore Policlinico, Mangiagalli e Regina Elena, Milan, Italy 2 Liver Transplant Unit, IRCCS Fondazione Ospedale Maggiore Policlinico, Mangiagalli e Regina Elena, Milan, Italy 3 Gastroenterology Surgery Department, National Cancer Institute, Milan, Italy 4 Clinic of Gastroenterology, Universit a Politecnica delle Marche, Ancona, Italy 5 Hemato-oncology Department, European Oncology Institute, Milan, Italy 6 Department of Transfusion Medicine and Hematology, Ospedale A. Manzoni, Lecco, Italy Received 20 August 2009; Revision Received 2 November 2009; Accepted 3 November 2009 Additional Supporting Information may be found in the online version of this article. Grant sponsor: COPEV (Comitato per la Prevenzione dell’Epatite Virale), Milan; Grant sponsor: Associazione Italiana Studio Fegato (AISF). *Correspondence to: Laura Porretti, DSc, Center of Transfusion Medicine, Cellular Therapy and Cryobiology, IRCCS Fondazione Ospedale Maggiore Policlinico, Mangiagalli e Regina Elena, Via F. Sforza 35, 20122 Milan, Italy Email: [email protected] Published online 3 December 2009 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/cyto.a.20834 © 2009 International Society for Advancement of Cytometry Original Article Cytometry Part A 77A: 3140, 2010
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Simultaneous characterization of progenitor cell compartments in adult human liver

Simultaneous Characterization of Progenitor Cell
Compartments in Adult Human Liver
Laura Porretti,1 * Alessandra Cattaneo,1 Federico Colombo,1 Raffaella Lopa,1 Giorgio Rossi,2
Vincenzo Mazzaferro,3 Carlo Battiston,3 Gianluca Svegliati-Baroni,4 Francesco Bertolini,5
Paolo Rebulla,1 Daniele Prati1,6
� AbstractThe human liver is a complex tissue consisting of epithelial, endothelial, hematopoietic,and mesenchymal elements that probably derive from multiple lineage-committed pro-genitors, but no comprehensive study aimed at identifying and characterizing intrahe-patic precursors has yet been published. Cell suspensions for this study were obtainedby enzymatic digestion of liver specimens taken from 20 patients with chronic liver dis-ease and 13 multiorgan donors. Stem and progenitor cells were first isolated, amplified,and characterized ex vivo according to previously validated methods, and then opti-mized flow cytometry was used to assess their relative frequencies and characterize theirimmunophenotypes in the clinical specimens. Stem and progenitor cells committed tohematopoietic, endothelial, epithelial, and mesenchymal lineages were clearly identifia-ble in livers from both healthy and diseased subjects. Within the mononuclear liver cellcompartment, epithelial progenitors [epithelial cell adhesion molecule (EpCAM)1/CD49f1/CD291/CD452] accounted for 2.7–3.5% whereas hematopoietic (CD341/CD451), endothelial [vascular endothelial growth factor-2 (KDR)1/CD1461/CD452],and mesenchymal [CD731/CD1051/CD90 (Thy-1)1/CD45 2] stem cells and progeni-tors accounted for smaller fractions (0.02–0.6%). The patients’ livers had higher per-centages of hematopoietic and endothelial precursors than those of the donors. In con-clusion, we identified and characterized precursors committed to four different lineagesin adult human liver. We also optimized a flow cytometry approach that will be usefulin exploring the contribution of these cells to the pathogenesis of liver disease. ' 2009
International Society for Advancement of Cytometry
� Key termsstem and progenitor cells; human liver; flow cytometry
ADULT human liver is a complex tissue consisting of epithelial, endothelial, hemato-
poietic, and mesenchymal elements. This supports the view that the intrahepatic
stem and progenitor cell compartment probably consists of a heterogeneous pool
with various molecular and functional markers. Studies characterizing hepatic stem
cells and progenitors have so far mainly concentrated on the epithelial compartment,
and generally used human fetal or rodent liver (1,2); only a few have identified and
characterized the hematopoietic (3,4), endothelial (5), and mesenchymal lineages
(6), although it is well known that their progenies are deeply involved in the pathoge-
nesis of liver disease (7).
Despite recent growth of interest in liver stem and progenitor cell research and
great expectations concerning the potential of these cells in the field of regenerative
medicine (8), we are unaware of comprehensive studies of the entire stem/progenitor
cell pool. This could be due to the rare occurrence of these cells in liver tissue, and
the possible overlap in the immunological profiles of the cells committed to different
lineages (9–11), thus making their identification difficult.
1Center of Transfusion Medicine, CellularTherapy and Cryobiology, IRCCSFondazione Ospedale Maggiore Policlinico,Mangiagalli e Regina Elena, Milan, Italy
2Liver Transplant Unit, IRCCS FondazioneOspedale Maggiore Policlinico,Mangiagalli e Regina Elena, Milan, Italy
3Gastroenterology Surgery Department,National Cancer Institute, Milan, Italy
4Clinic of Gastroenterology, Universit�aPolitecnica delle Marche, Ancona, Italy
5Hemato-oncology Department, EuropeanOncology Institute, Milan, Italy
6Department of Transfusion Medicineand Hematology, Ospedale A. Manzoni,Lecco, Italy
Received 20 August 2009; RevisionReceived 2 November 2009; Accepted 3November 2009
Additional Supporting Information may befound in the online version of this article.
Grant sponsor: COPEV (Comitato per laPrevenzione dell’Epatite Virale), Milan;Grant sponsor: Associazione ItalianaStudio Fegato (AISF).
*Correspondence to: Laura Porretti, DSc,Center of Transfusion Medicine, CellularTherapy and Cryobiology, IRCCSFondazione Ospedale MaggiorePoliclinico, Mangiagalli e Regina Elena,Via F. Sforza 35, 20122 Milan, Italy
Email: [email protected]
Published online 3 December 2009 in WileyInterScience (www.interscience.wiley.com)
DOI: 10.1002/cyto.a.20834
© 2009 International Society forAdvancement of Cytometry
Original Article
Cytometry Part A � 77A: 31�40, 2010

However, new-generation FCM techniques allowing the
immunological detection of larger numbers of cells have sig-
nificantly improved cell characterization by making it possible
to analyze up to six to eight markers on the same cell. As a
result, FCM has become the gold standard to characterize clin-
ical grade cell preparations in several fields of regenerative
medicine. In our own laboratories, it has allowed the identifi-
cation of rare stem cells and progenitors in peripheral blood
and parenchymal tissues (12–14), thus prompting us to refine
FCM protocols for liver stem cell analysis with a view to future
clinical applications.
The aim of this study was to implement a FCM approach
to investigate the heterogeneous pool of intrahepatic progeni-
tor cells suitable for both healthy and diseased livers. To this
aim, intrahepatic progenitor cells committed to four different
lineages were first isolated, amplified, and characterized ex
vivo using previously validated culture conditions and anti-
body staining techniques (15–18). This enabled us to select an
optimal panel of monoclonal antibodies that was then used to
determine the relative frequencies of the four lineages and fur-
ther characterize them in clinical specimens.
MATERIALS AND METHODS
Tissue Specimens
We analyzed liver samples taken from 13 organ donors
(seven males/six females; median age 56 years, range 33–82) at
the time of organ harvest, and from 20 patients with chronic
liver disease (11 males/9 females; median age 66 years, range
48–82) who had undergone liver resection because of isolated
hepatocellular carcinoma. The liver disease was cryptogenic in
three cases, and related to Hepatitis C virus (HCV) in eleven,
Hepatitis B virus (HBV) in three and alcohol abuse in three.
The study was approved by the local Ethics Committee,
and informed consent was obtained from the patients and the
donors’ relatives.
Cell Isolation
The cells were isolated within 2–8 hours of sample collec-
tion. After tissue digestion (5), the cell suspension was resus-
pended in HBSS (Euroclone, Pero, Italy), and centrifuged at
50 g for 1 min to remove mature hepatocytes. The mononu-
clear liver cells (MNLCs) were pelleted by centrifuging the su-
pernatant at 650g for 5 min. Cell counts and viability were
assessed by means of trypan blue staining.
In Vitro Cell Cultures
The samples from three organ donors were used for spe-
cific culture assays of the four liver stem/progenitor cell com-
partments.
Liver hematopoietic stem cells (L-HSCs). CD341 or
CD1331 cells were immunoselected by means of magnetic cell
separation (Miltenyi Biotech, Florence, Italy), and their purity
was evaluated by FCM. Next, 200 CD341 or CD1331 cells
were added to 2 ml of culture medium (Methocult, StemCells
Technologies, Milan, Italy) in Petri dishes, and then incubated
in humidified air with 5% CO2 at 378C. After 14 days, colonies
were identified using an inverted microscope: Burst Forming
Unit-Erythroid (BFU-E) as closely packed red/orange cells,
Colony Forming Unit-Granulocyte/Monocyte (CFU-GM) as
flat, nonhemoglobinized colonies consisting of translucent
cells, and mixed colonies containing elements of both (4).
Liver endothelial (L-EnPCs) and epithelial (L-EpPCs) pro-
genitor cells. MNLCs were plated at 378C onto 6-well tissue
plates precoated with human collagen (StemCells Technolo-
gies) for L-EnPCs or uncoated for L-EpPCs, and cultured as
described by Yoder et al. (16) and Duret et al. (17), respec-
tively. After 24 hours of culture, nonadherent cells were dis-
carded.
L-EnPCs were cultured with complete endothelial growth
medium 2 (EGM-2, Lonza, Treviglio, Italy); colonies of endo-
thelial cells appeared between 5 and 22 days of culture and
were identified as well-circumscribed monolayer cells.
L-EpPCs were cultured with minimal essential medium
supplemented with 10% FCS (Invitrogen, San Giuliano Mila-
nese, Italy), 20 ng/ml hepatocyte growth factor (Peprotech,
Segrate, Italy), 10 ng/ml epidermal growth factor (Peprotech),
25 mM glucose, 1 lM hydrocortisone, 10 lg/ml insulin, 50
lg/ml albumin-linoleic acid, 0.1 lM selenium acid, 0.75 mg/
ml zinc sulfate, 10 mM nicotinamide (all from Sigma-Aldrich,
Milan, Italy), streptomycin and penicillin (Invitrogen) (L-
EpPC medium). Media were changed three times a week. L-
EnPCs and L-EpPCs were counted at each passage and, at the
first and fifth passages, characterized for the expression of var-
ious markers (1,2,16,17,19–21).
At fifth passage L-EpPCs were cultured in L-EpPC me-
dium without serum (17) for 3 weeks, and then the activity of
cytochrome P450 enzymes was evaluated (15). For this pur-
pose, cells were washed with PBS and then left overnight at
378C at 5% CO2 with 0.02 mM 5-(and 6)-chloromethylfluor-
escein diethyl ether (Invitrogen). This substrate is converted
by cytochrome P450 in a fluorescent product detectable by flu-
orescence microscopy.
Liver mesenchymal stem cells (L-MSCs). Cell suspensions
were plated at a density of 5 3 106/cm2 on plastic flasks. After
24 hours, nonadherent cells were removed and MSC Growth
Medium BulletKit (Lonza) was added and replaced twice a
week (18). After 3 weeks, the confluent cells with a fibroblastic
morphology were split; at the first and fifth passages the cells
were evaluated by FCM and fluorescence microscopy and cul-
tured at the fifth passage under different conditions in order
to evaluate their osteogenic and adipogenic (18), endothelial
(16), and hepatic (17) differentiation capacity. In particular,
cells were then cultured for 3 weeks with the following specific
media: MSC Osteogenic or Adipogenic Differentiation Bullet-
Kit (Lonza), EGM-2 medium and L-EpPC medium, respec-
tively. Differentiation was assessed by Alizarin red (Sigma),
Red-O-oil (Sigma), CD31/CD144/vascular endothelial growth
factor receptor-2 (KDR) expression, and cytochrome P450
activity, respectively.
ORIGINAL ARTICLE
32 Progenitor Cell Characterization in Human Liver

FCM Characterization of Cultured Cells
A total of 1 3 105 liver cells/tube were stained at 48C for
20 min with 10 ll of 7-AAD (Becton Dickinson Biosciences,
BD, San Jose, CA) in order to distinguish dead cells, and with
appropriate mAbs (Supporting Information, Table 1S) conju-
gated with different fluorochromes. The samples were
acquired and analyzed on a FACSCanto I flow cytometer
(standard configuration) with FACSDiva 5.0.3 software (BD).
Quality control included regular check-up with 7-Color Setup
Beads (BD). To avoid channel overlapping, fluorescence com-
pensation was initially assessed creating an automated six
color compensation matrix specific for MNLCs. However, flu-
orescence compensation was further checked and adjusted in
each sample during data analysis.
Immunofluorescence Microscopy of Cultured Cells
Fixed plated cells (4% paraformaldehyde or 70% ethanol)
were permeabilized using HEPES-Triton X-100 buffer and
then immunostained with anti-cytokeratin (CK)-18, anti-a1-
antitrypsin (Sigma), anti-CK19 (Novocastra, Newcastle, UK),
anti-albumin (Dako-Cytomation, Milan, Italy), anti-CD31,
anti-CD146, anti-CD144, and anti-b-catenin (BD) mAbs. The
cell nuclei were stained with DAPI (Sigma) and evaluated with
a fluorescence microscope (DM IRE2, Leica Microsystems,
Wetzlar, Germany).
FCM Characterization of Dissociated Tissues
Samples from liver tissues were used to characterize the
hematopoietic, endothelial, epithelial and mesenchymal com-
partments by means of FCM. Specific analyses were made
with reference to the antigen profiles previously determined
on cultured cells.
Four tubes per sample were set up with at least 106 cells/
tube: Tube 1: Thy-1/ATP Binding Cassette sub-family G mem-
ber 2 (ABCG2)/7-AAD/CD34/CD133-2/CD45 (L-HSCs).
Tube 2: CD146/KDR/7-AAD/CD34/CD133-2/CD45 (L-
EnPCs). Tube 3: CD49f/CD29/7-AAD/CD56/CD133-1/CD45
(L-EpPCs). Tube 4: CD105/CD73/7-AAD/ CD34/ Thy-1/
CD45 (L-MSCs).
Appropriate isotype controls for all mAbs were used to
measure the non-specific binding on MNLCs. The four cell
compartments were further evaluated for the expression of
other antigens (Supporting Information, Table 1S) in order to
characterize their immunophenotype in more detail.
Statistical Analysis
The data are expressed as median and range. The groups
were compared using the Mann-Whitney test; P values of
\0.05 were considered statistically significant.
RESULTS
In Vitro Cell Cultures
L-HSCs. CD341/CD451, but not CD1331/CD451 cells, pro-
duced CFUs, a few myeloid and many erythroid colonies, after
14 days of culture (Fig. 1, panel A).
L-EnPCs. Upon the fifth passage in culture (doubling time,
50–70 hours), the flattened morphology and KDR1/CD1461/
CD311/CD1441/CD452 phenotype of the cells were similar
to those of circulating endothelial colony-forming cells
(ECFCs) (16) (Fig. 1, panels B–D). Moreover, more than 95%
of the cultured L-EnPCs expressed CD34 (Fig. 1, panel D) but
not CD133-2 (data not shown). This feature distinguishes cir-
culating from bone marrow immature endothelial progenitors,
as reported by Mobius-Winkler (22).
L-EpPCs. A confluent monolayer of cells with typical cuboid
morphology was obtained after 3 weeks of culture (Fig. 2,
panel A). The cells were replated five times (doubling time 160
hours), and evaluated for marker expression. Between the first
and fifth passages, they showed uniform CK19 and CK18
staining, membrane-localized b-catenin, and weak albumin
reactivity; they were not a-1-antitrypsin positive after the first
passage and only weakly positive after the fifth passage (Fig. 2,
Table 1. Commitment of intrahepatic stem cells towards different lineages
DONORS (n 510) PATIENTS (n 520) P
Median (range) Median (range)
Hematopoietic stem cells
CD341/CD451 0.05 (0.01–0.2) 0.2 (0.03–1) 0.01
CD133-21/CD451 0.08 (0.01–0.33) 0.08 (0.01–1) NS
Endothelial progenitors
KDR1/CD1461/CD451 0.025 (0–0.22) 0.11 (0–0.17) NS
KDR1/CD1461/CD452 0.02 (0–0.13) 0.13 (0.01–0.77) 0.02
Epithelial stem cells
CD291/CD49f1/CD452 3.5 (0.7–13.6) 2.7 (0.1–18.4) NS
Mesenchymal stem cells
CD731/CD1051/CD452 0.6 (0.4–2.5) 0.6 (0.1–3.7) NS
Median percentages (range) of freshly isolated mononuclear liver cell fraction.
The groups were compared using the Mann-Whitney test.
P values of\0.05 were considered significant.
ORIGINAL ARTICLE
Cytometry Part A � 77A: 31�40, 2010 33

Figure 1. Liver hematopoietic and endothelial progenitor cells. (A) L-HSCs: Erythroid, myeloid, and mixed colonies of CD341/CD451 cells
after 14 days culture in methylcellulose. (B) L-EnPCs: Representative cell morphology of endothelial progenitors at the first (left) and fifth
culture passage (right) (original magnification 320). (C) CD31 (left) and CD144 (right) immunofluorescence staining of cultured L-EnPCs at
the fifth culture passage (original magnification 320). (D) FCM analysis of cultured endothelial progenitors at fifth passage: KDR and
CD146 (upper panel), CD45, CD144, CD31, and CD34 antigen expression (middle and lower panels). In all histograms grey line represents
isotype control. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
ORIGINAL ARTICLE
34 Progenitor Cell Characterization in Human Liver

Figure 2. Liver epithelial progenitors and mesenchymal stem cells. (A) Cell morphology (310) of L-EpPCs at the first (left panel) and fifth
culture passage (middle panel), and L-MSCs morphology at the fifth culture passage (right panel, 320). (B) L-EpPCs immunofluorescence
staining at the first (upper panels) and fifth passage (lower panels): albumin (green)/CK19 (red) (left panels), CK18 (green)/a1-antitrypsin (red)
(middle panels), b catenin (green) (right panels), and DAPI (blue); magnification 320 (for negative control and single staining see Supporting
Information, Fig. 4S). (C) FCM analysis of L-EpPCs: CD29 and CD49f antigen expression after one (upper left panel) and five culture passages
(upper right panel); CD166 overlay expression on first (blue) and fifth (red) passage cells (lower panel). (D) Cytochrome P450 expression after
hepatic differentiation of fifth passage L-EpPCs (left panel), negative control (right panel); original magnification 320. (E) L-MSCs at fifth cul-
ture passage: CD105 and CD73 expression (upper-left panel); Thy-1 (upper-right panel) and CD34 (lower panel) expression on CD1051/CD731
cells. (F) Differentiation capability: osteogenic differentiation (Alizarin red staining, original magnification 320, left panel) and adipogenic dif-
ferentiation (Red-O-oil staining, right panel, original magnification 320) of fifth passage L-MSCs. In all histograms grey line represents isotype
control. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
ORIGINAL ARTICLE
Cytometry Part A � 77A: 31�40, 2010 35

panel B). The CD291/CD49f1/CD452 phenotype was main-
tained by more than 90% of the cells at the fifth passage (Fig.
2, panel C). Moreover, freshly isolated L-EpPCs expressed
epithelial cell adhesion molecule (EpCAM) and CD133-1,
which have recently been identified as markers of hepatic pro-
genitor cells (20,21,23), but this expression progressively
decreased in culture, whereas the expression of neural cell ad-
hesion molecule (NCAM) and CD166 antigens increased dur-
ing the five culture passages (Fig. 2, panel C). The cultured
cells gradually became larger and flatter and a number of
binuclear cells appeared. After hepatic maturation, P450 cyto-
chrome enzymes were expressed in 70% of the cells (Fig. 2,
panel D).
L-MSCs. During culture, the cells acquired their characteristic
spindle shape and showed the CD1051/CD731/Thy-11/
CD452/CD342 antigen profile (Fig. 2, panels A and E). The
cells harvested at the fifth passage had differentiated to osteo-
cytes and adipocytes (Fig. 2, panel F). No mature endothelial
and epithelial markers, including P450 cytochrome activity,
were observed after lineage specific commitments.
Clinical Specimens
The liver specimens weighed between 0.5 g and 388 g
(median 2.64 g). A median of 5.8 3 106 liver cells were iso-
lated per gram of tissue (0.7–55 3 106), and median cell via-
bility was 86% (70–95%). There was no significant difference
in recovery or viability rates between donors and patients. The
analyses of all staining tubes were performed on viable cells
(not stained with 7-AAD) with a scatter profile typical of
stem/progenitor cells: i.e. intermediate/high FSC and low/in-
termediate SSC (10,12,14). Viable MNLCs were then divided
into CD451 and CD452 cells.
Flow cytometry L-HSC analysis. CD341/CD451 cells (Fig. 3,
panel A) were significantly increased in the patients MNLCs
(0.08%) as compared with the donors (0.05%), whereas
CD133-21/CD451 cells accounted for about 0.08%, without
any differences between donors and patients (Table 1). As
shown in Figure 3, panel A, the CD341/CD451 cells were dis-
tinct from the CD133-21/CD451 cells: very few CD451 cells
(1–2%) coexpressed CD34 and CD133-2 antigens. Both sub-
populations expressed the same levels of Thy-1 (17%, 9–53%)
and ABCG2 (4%, 1–35%) (Supporting Information, Fig. 1S).
Flow cytometry L-EnPC analysis. KDR1/CD1461/CD452
cells were the output of cultured endothelial progenitors or
ECFCs (16); another cell population expressing CD45 antigen
(KDR1/CD1461/CD451) and usually indicated as colony
forming unit-endothelial cells (CFU-ECs), was also identified
(16). The analysis therefore included a first gate for KDR1
cells, followed by a quadrant analysis for the expression of
CD146 and CD45 antigens (Fig. 3, panel B). The ECFC subset
was significantly increased in the patients as compared to the
donors (0.13% vs. 0.02%), but there were no between-group
differences in the CFU-EC population (Table 1). CD34 and
CD133-2 antigens were expressed by 30–40% of the ECFCs
and 1–5% of the CFU-ECs (Supporting Information, Fig. 2S).
Flow cytometry L-EpPC analysis. Epithelial progenitor cells,
which were defined by the expression of the two integrins
CD49f and CD29 in the absence of CD45, accounted for 2.7-
3.5% of the MNLCs (Fig. 3, panel C) without differences
between patients and donors (Table 1). About 15–20% of the
CD49f1/CD291/CD452 cells were positive for NCAM anti-
gen, and about 50–70% for CD133-1 (Fig. 3, panel C). All of
the L-EpPCs were EpCAM positive, whereas there was no
CD34 antigen reactivity (Supporting Information, Fig. 3S).
Flow cytometry L-MSC analysis. MSCs, identified as
CD1051/CD731/Thy-11/CD452/CD342 cells (Fig. 3, panel
D), accounted for 1–10% of the CD452 MNLCs.
Table 2 summarizes the immunophenotypical character-
istics of the intrahepatic stem cells and progenitors committed
to different lineages. We found that a combination of five
mAbs (EpCAM/KDR/CD34/Thy-1/CD45) could identify the
four lineages in a single tube reaction on the basis of their dif-
ferent mAb reactivities. Figure 4 represents 3D plots showing
the four lineage-committed stem/progenitor cells.
DISCUSSION
Necroinflammation, fibrogenesis, neoangiogenesis, and
accelerated hepatocellular turnover are common features of
almost all forms of chronic liver disease, and are respectively
sustained by hematopoietic, mesenchymal, endothelial, and
epithelial elements. Although all of these elements derive from
lineage-specific progenitors, there is no comprehensive study
identifying and characterizing the heterogeneous hepatic pro-
genitor cell pool.
This study confirmed that our FCM analysis, based on an
optimized mAb panel, is useful in identifying rare intrahepatic
progenitors, committed to four lineages, both in healthy and
diseased livers. These progenitors are capable of expansion
and differentiation to their lineage commitment.
In order to ensure an adequate number of observations,
at least four million cells were evaluated for each liver speci-
men and characterized on the basis of more than one marker
in order to improve the specificity of the detection (9,24).
We first identified the hematopoietic and endothelial
lineages which, given their common mesodermal origin, share
some antigens. As they differently express CD45, KDR and the
endothelial/mesenchymal marker CD146 (16), these antigens
were used to distinguish them. Our data indicate that the HSC
compartment, firstly evidenced by Crosbie et al. (4), accounts
for 0.05–0.2% of intrahepatic mononuclear cells, 10 times
more than in peripheral blood. Interestingly, we observed that
CD341/CD451 cells were approximately four times more fre-
quent in the patients with chronic liver disease than in the
donors, probably because of increased cell trafficking due to
chronic inflammation.
Unlike bone marrow and peripheral blood, in which
60–70% of CD34bright cells express CD133 antigen (25), less
than 3% of the intrahepatic CD341 cells coexpressed CD133
ORIGINAL ARTICLE
36 Progenitor Cell Characterization in Human Liver
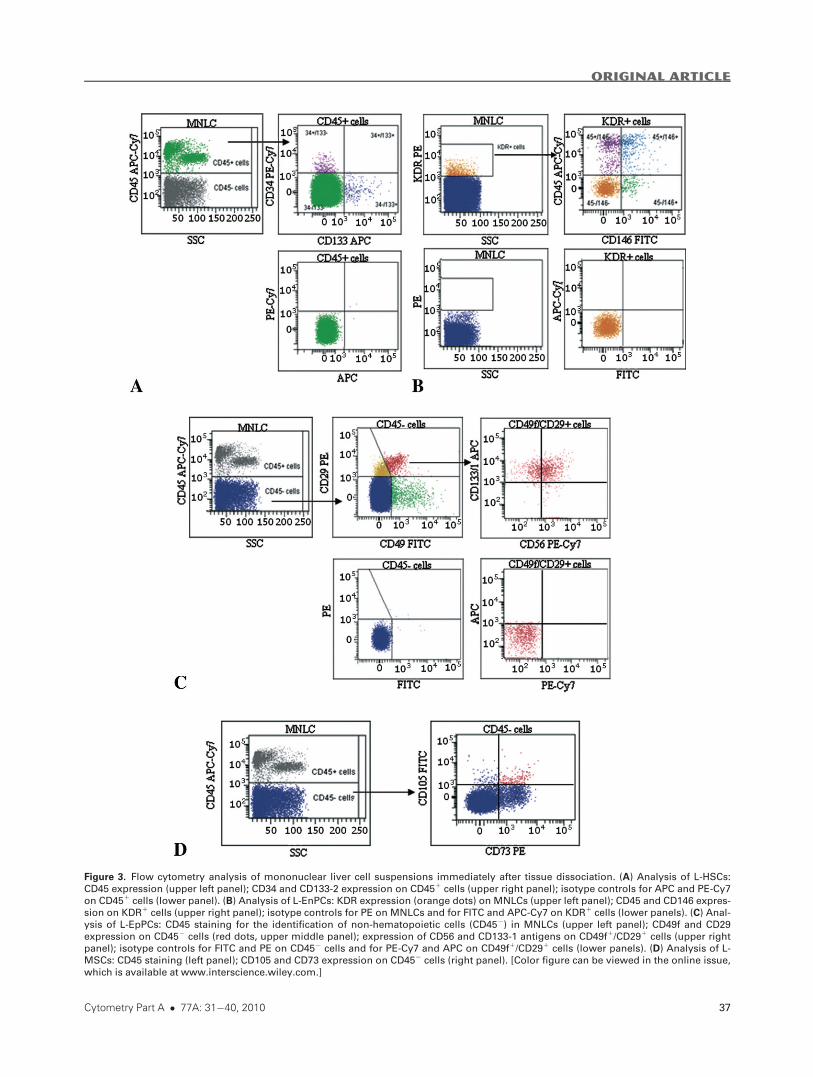
Figure 3. Flow cytometry analysis of mononuclear liver cell suspensions immediately after tissue dissociation. (A) Analysis of L-HSCs:
CD45 expression (upper left panel); CD34 and CD133-2 expression on CD451 cells (upper right panel); isotype controls for APC and PE-Cy7
on CD451 cells (lower panel). (B) Analysis of L-EnPCs: KDR expression (orange dots) on MNLCs (upper left panel); CD45 and CD146 expres-
sion on KDR1 cells (upper right panel); isotype controls for PE on MNLCs and for FITC and APC-Cy7 on KDR1 cells (lower panels). (C) Anal-
ysis of L-EpPCs: CD45 staining for the identification of non-hematopoietic cells (CD452) in MNLCs (upper left panel); CD49f and CD29
expression on CD452 cells (red dots, upper middle panel); expression of CD56 and CD133-1 antigens on CD49f1/CD291 cells (upper right
panel); isotype controls for FITC and PE on CD452 cells and for PE-Cy7 and APC on CD49f1/CD291 cells (lower panels). (D) Analysis of L-
MSCs: CD45 staining (left panel); CD105 and CD73 expression on CD452 cells (right panel). [Color figure can be viewed in the online issue,
which is available at www.interscience.wiley.com.]
ORIGINAL ARTICLE
Cytometry Part A � 77A: 31�40, 2010 37

antigen. However, a cell population bearing a CD1331/
CD451/CD342 phenotype and without hematopoietic poten-
tial in culture was also clearly identifiable. The hematopoietic
colonies were mainly erythroid, which fits well with the evi-
dence that the majority of BFU-E and CFU-mixed are pro-
duced by CD341/CD1332 stem cells (25). A small population
of ‘‘early’’ stem cells (about 4%) coexpressing ABCG2, which
is the molecular determinant of the side population (SP) phe-
notype (26), could be identified in almost all liver specimens,
suggesting the presence of cells with potent blood-repopulat-
ing potential (27) and capable of differentiating into hepato-
cyte-like cells in vitro (28).
Table 2. Patterns of mAb reactivities (mean percentages, n 5 3) on freshly isolated intrahepatic stem cells committed to different
lineages
COMMITMENT OF STEM CELLS
AND PROGENITORS CD34 CD45 CD133 ABCG2 THY-1 KDR CD146 CD31 CD105 CD73 CD29 CD49F EPCAM CD56 CD166
Hematopoietic 33 100 56 4 17 \1 0 0 0 NT NT NT 0 NT NT
Endothelial 35 0 70 NT 8 100 93 100 98 NT NT NT 0 NT NT
Epithelial 0 0 63 \1 \1 0 20 0 0 100 100 99 99 28 45
Mesenchymal 0 0 0 0 100 \1 83 0 87 88 100 0 \1 5 12
NT, not tested.
Figure 4. Three-dimensional plot of liver stem and progenitor cells analyzed in a single tube combination of EpCAM/KDR/7-AAD/CD34/
Thy-1 (CD90)/CD45. FACSDiva files, adequately compensated as reported in the text, were exported as FCS 2.0 in Paint-a-gate software.
The following gating strategy was then applied: L-HSCs (viable CD451/CD341 cells, yellow dots), ECFCs (viable CD452/KDR1 cells, black
dots), CFU-ECs (viable CD451/KDR1 cells, blue dots), L-EpPCs (viable CD452/EpCAM1 cells, green dots), L-MSCs (viable CD452/Thy-11
cells, violet dots). [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
ORIGINAL ARTICLE
38 Progenitor Cell Characterization in Human Liver

Since no studies on liver endothelial progenitors have
been published yet, we took advantage of the recent work by
Yoder et al. (16) and used the same culture conditions to iso-
late and maintain L-EnPCs. The morphology and phenotype
of the CD1461/KDR1/CD311/CD1441/CD452 cell popula-
tion were similar to those of circulating ECFCs, the only sub-
population capable of forming vessels in vivo (16). They were
more frequent in patients than in donors, which is in line with
the current view that hepatic inflammation and fibrogenesis
are frequently associated with angiogenesis (29).
The liver specimens also contained another cell popula-
tion coexpressing endothelial and hematopoietic markers
(KDR1/CD1461/CD451), which represented approximately
50% of the total KDR1 population. This immunophenotype
corresponded to CFU-EC, which are currently considered to
be descendents of hematopoietic stem cells that may facilitate
neoangiogenesis but cannot form perfused vessels in vivo (16).
We first cultivated epithelial progenitor cells under selec-
tive conditions (17) and obtained clusters with a typical mor-
phology that lasted for more than two months. After evaluat-
ing a large panel of markers because no single antigen had
been definitely validated (30), we observed that all of the cul-
tured cells expressed the a6 and b1 integrins, fundamental in
ensuring cell adhesion and clonality of hepatic progenitor cells
(1,2). EpCAM and CD133 antigens were also expressed on
freshly isolated L-EpPCs, but their reactivity progressively
decreased as the cells matured, as also suggested by the gradual
increase in CD166 and a1-antitrypsin positivity during
culture. In line with this, upon the fifth culture passage, the
cells were frequently binucleated and after maturation they
expressed P450 cytochrome, which are characteristic of mature
hepatocytes (31). The typical mesenchymal marker Thy-1, which
was not initially expressed by fresh L-EpPCs, progressively
appeared during culture, thus suggesting epithelial to mesenchy-
mal transition; however, the cells maintained the membrane
expression of b-catenin typical of epithelial cells (32).
In the clinical specimens, L-EpPCs accounted for the lar-
gest cluster of committed lineage within the MNLCs. Almost
all of these cells coexpressed EpCAM and a variable propor-
tion of CD166, thus suggesting the presence of cells at differ-
ent stages of maturation. In line with the results of recent stu-
dies (25,33), CD34 was not expressed at membrane level
whereas NCAM was detected in some cells.
MSCs were originally identified in bone marrow, but it is
currently agreed that they exist in all organs and tissues (34);
however, adult human liver MSCs have so far been poorly
described. Using the classic isolation protocol, we cultured
mesenchymal elements from liver specimens with the typical
hallmarks of MSCs, i.e. the expression of CD73, CD105, and
Thy-1, the absence of hematopoietic markers and osteogenic
and adipogenic differentiation potential (34). Mature liver
mesenchymal cells (mainly stellate cells) provide the basis for
the progression of fibrosis (35). Thus, a better characterization
of L-MSCs seems particularly important to improve our
knowledge of the pathogenesis of chronic liver disease.
In conclusion, healthy and diseased adult human livers
contain not only tissue-specific, but also mesenchymal, endo-
thelial, and hematopoietic progenitors, which can be distin-
guished by immunophenotyping and FCM analysis. A method
of single-tube staining with five-colour monoclonal antibo-
dies, that we defined in this study, can be used for future
investigations aimed at exploring the role and reciprocal inter-
actions of different subfractions of stem/progenitor cells in the
pathogenesis of liver disease.
ACKNOWLEDGMENTS
The authors are grateful to the Nord Italia Transplant
program (NITp) for providing liver tissue from organ donors
unsuitable for transplantation.
LITERATURE CITED
1. Suzuki A, Zheng Y-W, Kondo R, Kusakabe M, Takada Y, Fukao K, Nakauchi H, Tani-guchi H. Flow-cytometric separation and enrichment of hepatic progenitor cells inthe developing mouse liver. Hepatology 2000;32:1230–1239.
2. Suzuki A, Zheng Y-W, Kaneko S, Onodera M, Fukao K, Nakauchi H, Taniguchi H.Clonal identification and characterization of self-renewing pluripotent stem cells inthe developing liver. J Cell Biol 2002;156:173–184.
3. Taniguchi H, Toyoshima T, Fukao K, Nakauchi H. Presence of hematopoietic stemcells in the adult liver. Nat Med 1996;2:198–203.
4. Crosbie OM, Reynolds M, McEntee G, Traynor O, Hegarty JE, O’Farrelly C. In vitroevidence for the presence of hematopoietic stem cells in the adult human liver. Hepa-tology 1999;29:1193–1198.
5. Aicher A, Rentsch M, Sasaki K, Ellwart JW, Fandrich F, Siebert R, Cooke JP, Dimme-ler S, Heeschen C. Nonbone marrow-derived circulating progenitor cells contributeto postnatal neovascularization following tissue ischemia. Circ Res 2007;100:581–589.
6. Dan YY, Riehle KJ, Lazaro C, Teoh N, Haque J, Campbell JS, Fausto N. Isolation ofmultipotent progenitor cells from human fetal liver capable of differentiating intoliver and mesenchymal lineages. Proc Natl Acad Sci USA 2006;103:9912–9917.
7. Friedman SL. Hepatic stellate cells: Protean, multifunctional, and enigmatic cells ofthe liver. Physiol Rev 2008;88:125–172.
8. Forbes SJ. Stem cell therapy for chronic liver disease: Choosing the right tools for thejob. Gut 2008;57:153–155.
9. Donnenberg AD, Donnenberg VS. Rare-event analysis in flow cytometry. Clin LabMed 2007;27:627–652.
10. Crosby HA, Kelly DA, Strain AJ. Human hepatic stem-like cells isolated using c-kit orCD34 can differentiate into biliary epithelium. Gastroenterology 2001;120:534–544.
11. Adams V, Challen GA, Zuba-Surma E, Ulrich H, Vereb G, Tarnok A, Where newapproaches can stem from: Focus on stem cell identification, Cytometry Part A 2009;75A:1–3.
12. Torrente Y, Belicchi M, Sampaolesi M, Pisati F, Meregalli M, D’Antona G, TonlorenziR, Porretti L, Gavina M, Mamchaoui K, Pellegrino MA, Furling D, Mouly V, Butler-Browne GS, Bottinelli R, Cossu G, Bresolin N. Human circulating AC133(1) stemcells restore dystrophin expression and ameliorate function in dystrophic skeletalmuscle. J Clin Invest 2004;114:182–195.
13. Mancuso P, Antoniotti P, Quarna J, Calleri A, Rabascio C, Tacchetti C, Braidotti P,Wu HK, Zurita AJ, Saronni L, Cheng JB, Shalinsky DR, Heymach JV, Bertolini F. Vali-dation of a standardized method for enumerating circulating endothelial cells andprogenitors: Flow cytometry and molecular and ultrastructural analyses. Clin CancerRes 2009;15:267–273.
14. Marchesi C, Belicchi M, Meregalli M, Farini A, Cattaneo A, Parolini D, Gavina M,Porretti L, D’Angelo MG, Bresolin N, Cossu G, Torrente Y. Correlation of circulatingCD1331 progenitor subclasses with a mild phenotype in Duchenne muscular dystro-phy patients. PLoS ONE 2008;3:e2218.
15. Herrera MB, Bruno S, Buttiglieri S, Tetta C, Gatti S, Deregibus MC, Bussolati B,Camussi G. Isolation and characterization of a stem cell population from adulthuman liver. Stem Cells 2006;24:2840–2850.
16. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, Krasich R, Temm CJ,Prchal JT, Ingram DA. Redefining endothelial progenitor cells via clonal analysis andhematopoietic stem/progenitor cell principals. Blood 2007;109:1801–1809.
17. Duret C, Gerbal-Chaloin S, Ramos J, Fabre JM, Jacquet E, Navarro F, Blanc P,Sa-Cunha A, Maurel P, Daujat-Chavanieu M. Isolation, characterization, and differ-entiation to hepatocyte-like cells of nonparenchymal epithelial cells from adulthuman liver. Stem Cells 2007;25:1179–1190.
18. Chamberlain G, Fox J, Ashton B, Middleton J. Concise review: Mesenchymal stemcells: Their phenotype, differentiation capacity, immunological features and potentialfor homing. Stem Cells 2007;25:2739–2749.
19. Delorme B, Basire A, Gentile C, Sabatier F, Monsonis F, Desouches C, Blot-ChabaudM, Uzan G, Sampol J, Dignat-George F. Presence of endothelial progenitor cells, dis-tinct from mature endothelial cells, within human CD1461 blood cells. ThrombHaemost 2005;94:1270–1279.
20. Yovchev MI, Grozdanov PN, Zhou H, Racherla H, Guha C, Dabeva MD. Identifica-tion of adult hepatic progenitor cells capable of repopulating injured rat liver. Hepa-tology 2008;47:636–647.
ORIGINAL ARTICLE
Cytometry Part A � 77A: 31�40, 2010 39

21. Schmelzer E, Zhang L, Bruce A, Wauthier E, Ludlow J, Yao H, Moss N, Melhem A,McClelland R, Turner W, Kulik M, Sherwood S, Tallheden T, Cheng N, Furth ME,Reid LM. Human hepatic stem cells from fetal and postnatal donors. J Exp Med2007;204:1973–1987.
22. Mobius-Winkler S, Hollriegel R, Schuler G, Adams V, Endothelial progenitor cells:Implications for cardiovascular disease, Cytometry Part A 2009;75A:25–37.
23. Suzuki A, Sekiya S, Onishi M, Oshima N, Kiyonari H, Nakauchi H, Taniguchi H.Flow cytometric isolation and clonal identification of self-renewing bipotent hepaticprogenitor cells in adult mouse liver. Hepatology 2008;48:1–15.
24. Hughes OR, Stewart R, Dimmick I, Jones EA, A critical appraisal of factors affectingthe accuracy of results obtained when using flow cytometry in stem cell investigation:Where do you put your gates? Cytometry Part A 2009;75A:803–810.
25. Yin AH, Miraglia S, Zanjani ED, Almeida-Porada G, Ogawa M, Leary AG, Olweus J,Kearney J, Buck DW. AC133, a novel marker for human hematopoietic stem and pro-genitor cells. Blood 1997;90:5002–5012.
26. Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J, Morris JJ, Lagutina I,Grosveld GC, Osawa M, Nakauchi H, Sorrentino BP. The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant ofthe side-population phenotype. Nat Med 2001;7:1028–1034.
27. Kotton DN, Fabian AJ, Mulligan RC. A novel stem cell population in adult liver withpotent hematopoietic reconstitution activity. Blood 2005;106:1574–1580.
28. Hussain SZ, Strom SC, Kirby MR, Burns S, Langemeijer S, Ueda T, Hsieh M, TisdaleJF. Side population cells derived from adult human liver generate hepatocyte-like cellsin vitro. Dig Dis Sci 2005;50:1755–1763.
29. Medina J, Arroyo AG, Sanchez-Madrid F, Moreno-Otero R. Angiogenesis in chronicinflammatory liver disease. Hepatology 2004;39:1185–1195.
30. Dan YY, Yeoh GC. Liver stem cells: A scientific and clinical perspective. J Gastroen-terol Hepatol 2008;23:687–698.
31. Guidotti JE, Bregerie O, Robert A, Debey P, Brechot C, Desdouets C. Liver cell poly-ploidization: A pivotal role for binuclear hepatocytes. J Biol Chem 2003;278:19095–19101.
32. Eger A, Stockinger A, Schaffhauser B, Beuh H, Foisner R. Epithelial mesenchymaltransition by c-Fos estrogen receptor activation involves nuclear translocation ofbeta-catenin and upregulation of beta-catenin/lymphoid enhancer binding factor-1transcriptional activity. J Cell Biol 2000;148:173–188.
33. Schmelzer E, Wauthier E, Reid LM. The Phenotypes of pluripotent human hepaticprogenitors. Stem Cells 2006;24:1852–1858.
34. da Silva Meirelles L, Chagastelles PC, Beyer Nardi N. Mesenchymal stem cells residein virtually all post-natal organs and tissues. J Cell Sci 2006;119:2204–2213.
35. Svegliati-Baroni G, De Minicis S, Marzioni M. Hepatic fibrogenesis in response tochronic liver injury: Novel insights on the role of cell-to-cell interaction and transi-tion. Liver Int 2008;28:1052–1064.
ORIGINAL ARTICLE
40 Progenitor Cell Characterization in Human Liver




















