
Short-term climate changes in the southern Aegean Sea over the last 48,000 years
Upload
independentCategory
view
2download
0
Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332
wwwelseviercomlocatepalaeo
Short-term climate changes in the southern Aegean Sea
over the last 48000 years
Maria Geragaa Stella Tsaila-Monopolisb Chrysanthi Ioakimc
George Papatheodoroua George FerentinosaTaLaboratory of Marine Geology and Physical Oceanography Department of Geology University of Patras Patra Greece
bLaboratory of Paleontology Department of Geology University of Patras Patra GreececInstitute of Geology and Mineral Exploration (IGME) 70 Messogion Str Athens Greece
Received 4 June 2003 received in revised form 29 November 2004 accepted 28 January 2005
Abstract
High-resolution palaeoenvironmental changes corresponding to a mean time interval of 450 years covering the last 48000
years were examined in a core from the Cretan Basin in the southern Aegean Sea The intensity and duration of the climatic and
oceanographic events were determined by examining the compositional changes in the planktonic foraminifera and pollen
assemblages along with the y18O signal of Globigerinoides ruber A reconstruction of sea-surface temperatures was attempted
using the Modern Analogue Technique (MAT) In total 10 stadials and 6 interstadials occurred over the last 48000 years These
fluctuations in climatic conditions coincide with fluctuations documented in the western and central Mediterranean and seem to
be associated with DansgaardndashOeschger events Some of these climatic fluctuations are correlated with changes in the
vegetation in the surrounding land
Between 48 and 10 cal kyr BP the most pronounced stadials occurred at 41 cal kyr BP (C69-ST10) and at 13 cal kyr BP
(C69-ST4) These events are characterized by (i) high positive y18O values of Globigerinoides ruber (ii) drops in SST and (iii)
increases in aridity These events may be correlated with the Heinrich H4 event and the Younger Dryas event respectively Two
other stadials at 23 cal kyr BP (C69-ST6) and at 16 cal kyr BP (C69-ST5) which are characterized by increases in the abundance
of the cold plaktonic foraminifera species and increases in aridity may be correlated with the H2 and H1 events respectively
The dominant planktonic foraminiferal species during the stadials witch are correlated with the Heinrich events were
Turborotalita quinqueloba and Globorotalia scitula The most pronounced interstadials occurred between 395 and 385 cal kyr
BP (C69-IST6) and between 25 and 24 cal kyr BP (C69-IST3) and are characterized by depletion in y18O values increases in
SST and increases in humidity The former event coincides with the formation of the sapropelitic layer S2
In the Holocene the most pronounced stadial occurred between 8 and 65 cal kyr BP (C69-ST2) during the interruption of
S1 and is characterized by a reduction in SST and an increase in aridity The most pronounced interstadials of Holocene
occurred during the formation of S1a and S1b between 9 and 8 cal kyr BP (C69-IST1) and between 65 and 55 cal kyr BP
0031-0182$ - s
doi101016jpa
T Correspondi
E-mail addr
ee front matter D 2005 Elsevier BV All rights reserved
laeo200501010
ng author Tel +30 2610997646 fax +30 2610996275
ess gferenupatrasgr (G Ferentinos)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332312
(C69-IST2) respectively These events are characterized by depletion in y18O values increased SST and an increase in humidity
as is indicated by the expansion of temperate evergreen and Mediterranean taxa in the pollen record
D 2005 Elsevier BV All rights reserved
Keywords Late Quaternary Eastern Mediterranean Climatic variability Planktonic foraminifera Pollen Sapropel
1 Introduction
The documentation of climatic variability at millen-
nium scale in the northern Atlantic during the Late
Quaternary referred to as DansgaardndashOeschger cli-
matic cycles (Dansgaard et al 1993 Bond and Lotti
1995 Bond et al 1997) recently led scientists to
investigate the presence of similar events in the
Mediterranean Sea For this reason high-resolution
palaeoclimatic studies have recently been carried out
on marine sediments of the western (Cacho et al 1999
2000 2002) and easternMediterranean Sea (Rohling et
al 1997 De Rijk et al 1999 Geraga et al 2000) and
on terrestrial sediments in the Middle-East (Bar-
Matthews et al 1999 Gvirtzman and Wieder 2001)
The purpose of the present study is to further
extend our knowledge of the climatic variability over
the last 48000 years in the Cretan Basin in the
southern Aegean Sea Furthermore within the frame-
work of this work the microfauna assemblages are
compared with those of the central and the western
Mediterranean Sea (Hayes et al 1999 Sbaffi et al
2001 Perez-Folgado et al 2003) and the formation of
sapropels S1 and S2 is discussed
2 Material and methods
The present paper is based on the sedimentological
micropalaeontological and palynological examination
of a 2 m long core C69 collected from the Cretan
Basin in the southern Aegean Sea (Fig 1) The core
was collected at a water depth of 632 m at Longitude
36832769E and Latitude 24812782NNinety-four samples were used for foram analysis
and 76 samples for pollen analysis The mean
sampling interval for the faunal analyses was 2 cm
Each sample covered approximately a 1 cm thick
interval in the core
For the foraminiferal studies the samples were
disintegrated by hydrogen peroxide and were then
sieved through a 150 Am mesh The dry and weighed
samples were split into separate aliquots
The organic carbon content (Corg) was determined
by the K2Cr2O7ndashFe(NH4)26(H2O) titration method
according to Gaudette et al (1974)
For the palynological analyses the samples were
treated with HCl and HF and ultrasonic sieving (10
Am) Samples were stored and mounted in glycerin In
the diagrams of the total palynomorphs the dino-
flagellate cysts observed in the samples were
excluded The pollen count was between 210 and
930 grains per sample For the construction of the
pollen diagrams selected taxa from the pollen
percentage data were plotted against age together
with the total amount of Arboreal and Non Arboreal
Pollen (AP and NAP) concentration The percentages
of the selected data were based on the total of the
pollen assemblage
A total number of 52 samples along the core were
analyzed for the determination of the oxygen isotopes
on the tests of the planktonic foraminifera Globiger-
inoides ruber (N125 Am) The analyses were carried
out at the SOCFAC Laboratory of Southampton
University and at the Laboratory of Geology and
Geophysics in Edinburgh University
Sea-surface palaeotemperatures in C69 were esti-
mated using the Modern Analogue Technique (MAT
Hutson 1980 Overpeck et al 1985) The MAT
method is based on a database of modern samples
from the north Atlantic and the Mediterranean Sea and
by selecting those whose faunal composition is most
similar to a given fossil sample the sea-surface
palaeotemperature is thus determined The database
used in the present study for the MAT method
comprises the CLIMAP database and the database
of Kallel et al (1997) The former database gives the
sea-surface temperature (SST) for summer (SSTs) and
winter (SSTw) The latter database is based on
samples selected mainly from the Mediterranean Sea
and gives SST on a seasonal basis For the estimation
of the SSTcool (SSTc) in the present study the SSTw
Fig 1 Location map of the study area (CB) in the Aegean Sea showing (a) the present day coastline (thick line) and bathymetry (b) the Last
Glacial Maximum coastline (thin line) according to Van Andel and Lianos (1984) (c) the major rivers and torrents and (d) the location of the
studied core (C69) MB Myrtoon Basin NAT North Aegean Trough SkB Skopelos Basin NSB North Skyros Basin SSB South Skyros
Basin NIB North Ikaria Basin SIB South Ikaria Basin and CB Cretan Basin
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 313
by CLIMAP database and the SST of February by
Kallel were combined For the SSTwarm (SSTw) the
SSTs by CLIMAP and the SST of August by Kallel
were combined
Seven Accelerator Mass Spectrometry (AMS)
datings on specimens of Globigerinoides ruber were
carried out at the laboratories of Beta Analytic (USA)
(Table 1)
3 Data presentation
31 Sediments
Six lithological units have been identified at core
C69 (Fig 2a) on the basis of colour grain size
organic carbon content and sedimentary structures
The first unit A (0ndash23 cm) consists of yellowish
brown structureless mud with a small proportion of
sand Volcanic ash has been observed at about 15 cm
below surface but does not form a discrete tephra
layer This horizon most probably corresponds to the
most recent ash-layer in the Aegean Sea Z2 Santorini
which has a calibrated age of 3570 yr BP (Ramsey et
al 2004)
The second unit B (23ndash44 cm) represents a
sapropel sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) with the Corg content ranging
between 05 and 21 (Figs 2a and 3b) Based on
the AMS dating (Table 1) this sapropel sequence
corresponds to the S1 sapropel which was deposited
in the Aegean Sea and in the eastern Mediterranean
sometime between 6 and 12 14C kyr BP (Cita et al
1977 Rossignol-Strick et al 1982 Fontugne et al
Table 1
AMS dates and dated control points used in core C69
Depth (cm) AMS laboratory
code or source
Uncorrected
AMS 14C
(yr BP)
Calibrated
(yr BP)
150 Z2 Santorini
ash layer
3570a
280 Beta-166995 5810F40 5964ndash6269b
400 Beta-142621 8750F70 8919ndash9458b
600 Beta-190861 14470F70 16164ndash17173b
64 Boundary BZ67 18100c
84 Boundary BZ78a 24200c
930 Beta-168207 29120F270 Turbidited
990 Beta-166996 37900F1200 Turbidited
111 Boundary BZ89 32100c
1520 Beta-133928 36070F300 39855e
1890 Beta-166997 44300F2000 47126e
a Ramsey et al (2004)b Converted into calendar age using the program bcalib 44Q
(Stuiver and Reimer 1993) DR=88F54 (Reimer 2000)c From the study by Sbaffi et al (2001) and Capotondi et al
(1999)d These dates were not used in the construction of the age model
curve in Fig 2e Converted into calendar age with the polynomial provided by
Bard et al (2004)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332314
1994 Aksu et al 1995) Based on the fluctuations of
Corg content (Fig 3) the sapropel sequence can be
divided into three zones a lower and an upper zone
which are characterized by a maximum Corg content
of 21 and 16 respectively and a middle zone
separating the two others with a minimum Corg
content of 11 The lower (40ndash35 cm) and the upper
(30ndash26 cm) zones of high Corg content will hereforth
be referred to as S1a and S1b respectively Previous
studies on the formation of S1 have also described an
interruption in the deposition of the sapropel sedi-
ments (Rossignol-Strick et al 1982 Rohling et al
1997 Geraga et al 2000)
The third unit C (44ndash87 cm) consists of brownish
structureless mud with rare shell fragments The
fourth unit D (87ndash132 cm) is similar to unit C but
is grayish in colour Two layers of structureless silt
appear between 93 and 105 cm The lower boundary
of this interval at 105 cm is sharp and the upper one at
93 cm appears gradational The boundary between the
two layers is indistinct and occurs at approximately 97
cm The two layers correspond to two different
turbidites according to their sedimentological charac-
teristics The lower turbidite layer between 105 and 97
cm consists of material dated at 37900 14C kyr BP
(Table 1) and the upper between 97 and 93 cm
consists of material dated at 29120 14C kyr BP (Table
1) suggesting that the supplying source of each
turbidite was situated at two different locations
The fifth unit E (132ndash153 cm) represents a
sapropelitic sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) Within this sequence the max-
imum Corg content (05ndash154) occurs between 153
and 143 cm This sapropelic layer consists of olive
coloured structureless mud During the interval that
follows between 143 and 132 cm the Corg content
declines from 05 to 03 Based on the AMS
dating (Table 1) the age of this sapropelitic sequence
is 355ndash335 14C kyr BP and seems to correspond to
the S2 sapropel The rare appearance of sapropel S2
often called the bghost sapropelQ in the sediments of
the eastern Mediterranean has led to approximate
estimations of its age (Ryan 1972 Vergnaud-Grazzini
et al 1977)
The sixth unit F (153ndash195 cm) consists of
brownish mud
The sediments of units C D and E contain sand
and silt sized volcanic ash almost everywhere but no
discrete ash layers The highest concentration of the
ash was obtained between 97 and 125 cm
32 Age assessment
The chronological framework of the present study
is based on the five AMS dates (Table 1) The
turbidite layers have not been taken into consideration
in the construction of the age model curve for the
estimation of the sedimentation rate
The first three dates (b24 kyr BP) were converted
into calendar years using the program bcalib 44Q(Stuiver and Reimer 1993 Reimer 2000) while the
other two were converted using the polynomials
provided by Bard et al (2004) (Table 1)
In addition four dated control points were used for
the age assessment (Table 1) The upper dating point
is based on the age (357 cal kyr BP) (Ramsey et al
2004) of the ash layer at 15 cm which is considered to
be the Z2 Santorini ash layer The other three are
based on the biostratigraphic study by Sbaffi et al
(2001) That study has revealed 9 biozones over the
last 34 kyr based on changes in the planktonic
foraminifera and coccolithophores assemblages in the
Fig 2 (a) Lithology of core C69 Ages displayed to the left of the core are AMS 14Cnc dates (b) Age model curve for the estimation of the mean
sedimentation rate The diagram is based on five calibrated AMS 14C ages (arrows) and four dating points (in cal yr BP) based on (i) the
Santorini Z2 ash layer (Ramsey et al 2004) (square) and (ii) the estimated age for upper and lower boundaries of biozone 7 and upper boundary
of biozone 9 by Sbaffi et al (2001) (circles) The turbidite layers have not been taken into consideration in the construction of the age model
curve for the estimation of the sedimentation rate Thus the older dating points (at 47126 cal kyr BP 39855 cal kyr BP and 32100 cal kyr BP)
appear in shallower depths in the age model curve than in the core
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 315
central Mediterranean dated in calendar years BP
These biozones are comparable with the ecozones
proposed by Capotondi et al (1999) and are detect-
able throughout the central Mediterranean and as far
as the southern Adriatic Sea (Giunta et al 2003
Principato et al 2003) In our study the lower and the
upper boundaries (in calendar years BP) of the
biozone 7 and the upper boundary of biozone 9 were
used The former which corresponds to the ecozone 9
includes the Last Glacial Maximum interval and is
well identified in our records by the planktonic
assemblages and the isotopic signal The isotopic
Fig 3 Downcore variation of (a) y18O values (x) and (b) Corg content () versus age in cal kyr BP
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332316
record for this interval is also consistent with the
isotopic record in the study of Siani et al (2001) in the
southern Adriatic Sea suggesting a realistic age
control Biozone 9 represents a warm interval
corresponding to interstadial 5 (Cacho et al 1999)
In our data biozone 9 is well identified by the
planktonic assemblages and the depletions of the
isotope values
Based on the above age model curve the mean
sedimentation rate in core C69 is 38 cmkyr (Fig 2)
The mean time resolution of planktonic and MAT
records is 450 years while that of oxygen isotopes
and pollen is 850 yr and 550 yr respectively
33 Oxygen isotopes
The downcore variation of the y18O values in core
C69 seen in Fig 3 reveals that in the isotopic stage
ST3 (48ndash24 cal kyr BP) the y18O data exhibits
relatively lower values between 40 and 38 cal kyr BP
(~00x) and around 34 cal kyr BP (06x) The
former corresponds to the deposition of the S2
sapropel and the latter corresponds to the Interstadial
5 (Ist5) which has been also identified in cores from
the central and western Mediterranean (Cacho et al
1999 Sbaffi et al 2001 Perez-Folgado et al 2003)
The small decrease in y18O values (19x) around 24
cal kyr BP may correspond to the isotopic event 31
(Martinson et al 1987) High y18O values occur at
time intervals from 415 to 405 cal kyr BP (18ndash
19x) 355ndash345 cal kyr BP (15ndash13x) and 29ndash27
cal kyr BP (27x) The first and the third time
intervals are around the ages of the Heinrich 4 (39 cal
kyr BP) and Heinrich 3 (30 cal kyr BP) events
respectively (Bond et al 1992)
In the isotopic stage ST2 (24ndash11 cal kyr BP) the
strongest enrichment of y18O values (28ndash25x) occur
between 215 and 195 cal kyr BP suggesting the Last
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332312
(C69-IST2) respectively These events are characterized by depletion in y18O values increased SST and an increase in humidity
as is indicated by the expansion of temperate evergreen and Mediterranean taxa in the pollen record
D 2005 Elsevier BV All rights reserved
Keywords Late Quaternary Eastern Mediterranean Climatic variability Planktonic foraminifera Pollen Sapropel
1 Introduction
The documentation of climatic variability at millen-
nium scale in the northern Atlantic during the Late
Quaternary referred to as DansgaardndashOeschger cli-
matic cycles (Dansgaard et al 1993 Bond and Lotti
1995 Bond et al 1997) recently led scientists to
investigate the presence of similar events in the
Mediterranean Sea For this reason high-resolution
palaeoclimatic studies have recently been carried out
on marine sediments of the western (Cacho et al 1999
2000 2002) and easternMediterranean Sea (Rohling et
al 1997 De Rijk et al 1999 Geraga et al 2000) and
on terrestrial sediments in the Middle-East (Bar-
Matthews et al 1999 Gvirtzman and Wieder 2001)
The purpose of the present study is to further
extend our knowledge of the climatic variability over
the last 48000 years in the Cretan Basin in the
southern Aegean Sea Furthermore within the frame-
work of this work the microfauna assemblages are
compared with those of the central and the western
Mediterranean Sea (Hayes et al 1999 Sbaffi et al
2001 Perez-Folgado et al 2003) and the formation of
sapropels S1 and S2 is discussed
2 Material and methods
The present paper is based on the sedimentological
micropalaeontological and palynological examination
of a 2 m long core C69 collected from the Cretan
Basin in the southern Aegean Sea (Fig 1) The core
was collected at a water depth of 632 m at Longitude
36832769E and Latitude 24812782NNinety-four samples were used for foram analysis
and 76 samples for pollen analysis The mean
sampling interval for the faunal analyses was 2 cm
Each sample covered approximately a 1 cm thick
interval in the core
For the foraminiferal studies the samples were
disintegrated by hydrogen peroxide and were then
sieved through a 150 Am mesh The dry and weighed
samples were split into separate aliquots
The organic carbon content (Corg) was determined
by the K2Cr2O7ndashFe(NH4)26(H2O) titration method
according to Gaudette et al (1974)
For the palynological analyses the samples were
treated with HCl and HF and ultrasonic sieving (10
Am) Samples were stored and mounted in glycerin In
the diagrams of the total palynomorphs the dino-
flagellate cysts observed in the samples were
excluded The pollen count was between 210 and
930 grains per sample For the construction of the
pollen diagrams selected taxa from the pollen
percentage data were plotted against age together
with the total amount of Arboreal and Non Arboreal
Pollen (AP and NAP) concentration The percentages
of the selected data were based on the total of the
pollen assemblage
A total number of 52 samples along the core were
analyzed for the determination of the oxygen isotopes
on the tests of the planktonic foraminifera Globiger-
inoides ruber (N125 Am) The analyses were carried
out at the SOCFAC Laboratory of Southampton
University and at the Laboratory of Geology and
Geophysics in Edinburgh University
Sea-surface palaeotemperatures in C69 were esti-
mated using the Modern Analogue Technique (MAT
Hutson 1980 Overpeck et al 1985) The MAT
method is based on a database of modern samples
from the north Atlantic and the Mediterranean Sea and
by selecting those whose faunal composition is most
similar to a given fossil sample the sea-surface
palaeotemperature is thus determined The database
used in the present study for the MAT method
comprises the CLIMAP database and the database
of Kallel et al (1997) The former database gives the
sea-surface temperature (SST) for summer (SSTs) and
winter (SSTw) The latter database is based on
samples selected mainly from the Mediterranean Sea
and gives SST on a seasonal basis For the estimation
of the SSTcool (SSTc) in the present study the SSTw
Fig 1 Location map of the study area (CB) in the Aegean Sea showing (a) the present day coastline (thick line) and bathymetry (b) the Last
Glacial Maximum coastline (thin line) according to Van Andel and Lianos (1984) (c) the major rivers and torrents and (d) the location of the
studied core (C69) MB Myrtoon Basin NAT North Aegean Trough SkB Skopelos Basin NSB North Skyros Basin SSB South Skyros
Basin NIB North Ikaria Basin SIB South Ikaria Basin and CB Cretan Basin
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 313
by CLIMAP database and the SST of February by
Kallel were combined For the SSTwarm (SSTw) the
SSTs by CLIMAP and the SST of August by Kallel
were combined
Seven Accelerator Mass Spectrometry (AMS)
datings on specimens of Globigerinoides ruber were
carried out at the laboratories of Beta Analytic (USA)
(Table 1)
3 Data presentation
31 Sediments
Six lithological units have been identified at core
C69 (Fig 2a) on the basis of colour grain size
organic carbon content and sedimentary structures
The first unit A (0ndash23 cm) consists of yellowish
brown structureless mud with a small proportion of
sand Volcanic ash has been observed at about 15 cm
below surface but does not form a discrete tephra
layer This horizon most probably corresponds to the
most recent ash-layer in the Aegean Sea Z2 Santorini
which has a calibrated age of 3570 yr BP (Ramsey et
al 2004)
The second unit B (23ndash44 cm) represents a
sapropel sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) with the Corg content ranging
between 05 and 21 (Figs 2a and 3b) Based on
the AMS dating (Table 1) this sapropel sequence
corresponds to the S1 sapropel which was deposited
in the Aegean Sea and in the eastern Mediterranean
sometime between 6 and 12 14C kyr BP (Cita et al
1977 Rossignol-Strick et al 1982 Fontugne et al
Table 1
AMS dates and dated control points used in core C69
Depth (cm) AMS laboratory
code or source
Uncorrected
AMS 14C
(yr BP)
Calibrated
(yr BP)
150 Z2 Santorini
ash layer
3570a
280 Beta-166995 5810F40 5964ndash6269b
400 Beta-142621 8750F70 8919ndash9458b
600 Beta-190861 14470F70 16164ndash17173b
64 Boundary BZ67 18100c
84 Boundary BZ78a 24200c
930 Beta-168207 29120F270 Turbidited
990 Beta-166996 37900F1200 Turbidited
111 Boundary BZ89 32100c
1520 Beta-133928 36070F300 39855e
1890 Beta-166997 44300F2000 47126e
a Ramsey et al (2004)b Converted into calendar age using the program bcalib 44Q
(Stuiver and Reimer 1993) DR=88F54 (Reimer 2000)c From the study by Sbaffi et al (2001) and Capotondi et al
(1999)d These dates were not used in the construction of the age model
curve in Fig 2e Converted into calendar age with the polynomial provided by
Bard et al (2004)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332314
1994 Aksu et al 1995) Based on the fluctuations of
Corg content (Fig 3) the sapropel sequence can be
divided into three zones a lower and an upper zone
which are characterized by a maximum Corg content
of 21 and 16 respectively and a middle zone
separating the two others with a minimum Corg
content of 11 The lower (40ndash35 cm) and the upper
(30ndash26 cm) zones of high Corg content will hereforth
be referred to as S1a and S1b respectively Previous
studies on the formation of S1 have also described an
interruption in the deposition of the sapropel sedi-
ments (Rossignol-Strick et al 1982 Rohling et al
1997 Geraga et al 2000)
The third unit C (44ndash87 cm) consists of brownish
structureless mud with rare shell fragments The
fourth unit D (87ndash132 cm) is similar to unit C but
is grayish in colour Two layers of structureless silt
appear between 93 and 105 cm The lower boundary
of this interval at 105 cm is sharp and the upper one at
93 cm appears gradational The boundary between the
two layers is indistinct and occurs at approximately 97
cm The two layers correspond to two different
turbidites according to their sedimentological charac-
teristics The lower turbidite layer between 105 and 97
cm consists of material dated at 37900 14C kyr BP
(Table 1) and the upper between 97 and 93 cm
consists of material dated at 29120 14C kyr BP (Table
1) suggesting that the supplying source of each
turbidite was situated at two different locations
The fifth unit E (132ndash153 cm) represents a
sapropelitic sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) Within this sequence the max-
imum Corg content (05ndash154) occurs between 153
and 143 cm This sapropelic layer consists of olive
coloured structureless mud During the interval that
follows between 143 and 132 cm the Corg content
declines from 05 to 03 Based on the AMS
dating (Table 1) the age of this sapropelitic sequence
is 355ndash335 14C kyr BP and seems to correspond to
the S2 sapropel The rare appearance of sapropel S2
often called the bghost sapropelQ in the sediments of
the eastern Mediterranean has led to approximate
estimations of its age (Ryan 1972 Vergnaud-Grazzini
et al 1977)
The sixth unit F (153ndash195 cm) consists of
brownish mud
The sediments of units C D and E contain sand
and silt sized volcanic ash almost everywhere but no
discrete ash layers The highest concentration of the
ash was obtained between 97 and 125 cm
32 Age assessment
The chronological framework of the present study
is based on the five AMS dates (Table 1) The
turbidite layers have not been taken into consideration
in the construction of the age model curve for the
estimation of the sedimentation rate
The first three dates (b24 kyr BP) were converted
into calendar years using the program bcalib 44Q(Stuiver and Reimer 1993 Reimer 2000) while the
other two were converted using the polynomials
provided by Bard et al (2004) (Table 1)
In addition four dated control points were used for
the age assessment (Table 1) The upper dating point
is based on the age (357 cal kyr BP) (Ramsey et al
2004) of the ash layer at 15 cm which is considered to
be the Z2 Santorini ash layer The other three are
based on the biostratigraphic study by Sbaffi et al
(2001) That study has revealed 9 biozones over the
last 34 kyr based on changes in the planktonic
foraminifera and coccolithophores assemblages in the
Fig 2 (a) Lithology of core C69 Ages displayed to the left of the core are AMS 14Cnc dates (b) Age model curve for the estimation of the mean
sedimentation rate The diagram is based on five calibrated AMS 14C ages (arrows) and four dating points (in cal yr BP) based on (i) the
Santorini Z2 ash layer (Ramsey et al 2004) (square) and (ii) the estimated age for upper and lower boundaries of biozone 7 and upper boundary
of biozone 9 by Sbaffi et al (2001) (circles) The turbidite layers have not been taken into consideration in the construction of the age model
curve for the estimation of the sedimentation rate Thus the older dating points (at 47126 cal kyr BP 39855 cal kyr BP and 32100 cal kyr BP)
appear in shallower depths in the age model curve than in the core
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 315
central Mediterranean dated in calendar years BP
These biozones are comparable with the ecozones
proposed by Capotondi et al (1999) and are detect-
able throughout the central Mediterranean and as far
as the southern Adriatic Sea (Giunta et al 2003
Principato et al 2003) In our study the lower and the
upper boundaries (in calendar years BP) of the
biozone 7 and the upper boundary of biozone 9 were
used The former which corresponds to the ecozone 9
includes the Last Glacial Maximum interval and is
well identified in our records by the planktonic
assemblages and the isotopic signal The isotopic
Fig 3 Downcore variation of (a) y18O values (x) and (b) Corg content () versus age in cal kyr BP
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332316
record for this interval is also consistent with the
isotopic record in the study of Siani et al (2001) in the
southern Adriatic Sea suggesting a realistic age
control Biozone 9 represents a warm interval
corresponding to interstadial 5 (Cacho et al 1999)
In our data biozone 9 is well identified by the
planktonic assemblages and the depletions of the
isotope values
Based on the above age model curve the mean
sedimentation rate in core C69 is 38 cmkyr (Fig 2)
The mean time resolution of planktonic and MAT
records is 450 years while that of oxygen isotopes
and pollen is 850 yr and 550 yr respectively
33 Oxygen isotopes
The downcore variation of the y18O values in core
C69 seen in Fig 3 reveals that in the isotopic stage
ST3 (48ndash24 cal kyr BP) the y18O data exhibits
relatively lower values between 40 and 38 cal kyr BP
(~00x) and around 34 cal kyr BP (06x) The
former corresponds to the deposition of the S2
sapropel and the latter corresponds to the Interstadial
5 (Ist5) which has been also identified in cores from
the central and western Mediterranean (Cacho et al
1999 Sbaffi et al 2001 Perez-Folgado et al 2003)
The small decrease in y18O values (19x) around 24
cal kyr BP may correspond to the isotopic event 31
(Martinson et al 1987) High y18O values occur at
time intervals from 415 to 405 cal kyr BP (18ndash
19x) 355ndash345 cal kyr BP (15ndash13x) and 29ndash27
cal kyr BP (27x) The first and the third time
intervals are around the ages of the Heinrich 4 (39 cal
kyr BP) and Heinrich 3 (30 cal kyr BP) events
respectively (Bond et al 1992)
In the isotopic stage ST2 (24ndash11 cal kyr BP) the
strongest enrichment of y18O values (28ndash25x) occur
between 215 and 195 cal kyr BP suggesting the Last
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

Fig 1 Location map of the study area (CB) in the Aegean Sea showing (a) the present day coastline (thick line) and bathymetry (b) the Last
Glacial Maximum coastline (thin line) according to Van Andel and Lianos (1984) (c) the major rivers and torrents and (d) the location of the
studied core (C69) MB Myrtoon Basin NAT North Aegean Trough SkB Skopelos Basin NSB North Skyros Basin SSB South Skyros
Basin NIB North Ikaria Basin SIB South Ikaria Basin and CB Cretan Basin
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 313
by CLIMAP database and the SST of February by
Kallel were combined For the SSTwarm (SSTw) the
SSTs by CLIMAP and the SST of August by Kallel
were combined
Seven Accelerator Mass Spectrometry (AMS)
datings on specimens of Globigerinoides ruber were
carried out at the laboratories of Beta Analytic (USA)
(Table 1)
3 Data presentation
31 Sediments
Six lithological units have been identified at core
C69 (Fig 2a) on the basis of colour grain size
organic carbon content and sedimentary structures
The first unit A (0ndash23 cm) consists of yellowish
brown structureless mud with a small proportion of
sand Volcanic ash has been observed at about 15 cm
below surface but does not form a discrete tephra
layer This horizon most probably corresponds to the
most recent ash-layer in the Aegean Sea Z2 Santorini
which has a calibrated age of 3570 yr BP (Ramsey et
al 2004)
The second unit B (23ndash44 cm) represents a
sapropel sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) with the Corg content ranging
between 05 and 21 (Figs 2a and 3b) Based on
the AMS dating (Table 1) this sapropel sequence
corresponds to the S1 sapropel which was deposited
in the Aegean Sea and in the eastern Mediterranean
sometime between 6 and 12 14C kyr BP (Cita et al
1977 Rossignol-Strick et al 1982 Fontugne et al
Table 1
AMS dates and dated control points used in core C69
Depth (cm) AMS laboratory
code or source
Uncorrected
AMS 14C
(yr BP)
Calibrated
(yr BP)
150 Z2 Santorini
ash layer
3570a
280 Beta-166995 5810F40 5964ndash6269b
400 Beta-142621 8750F70 8919ndash9458b
600 Beta-190861 14470F70 16164ndash17173b
64 Boundary BZ67 18100c
84 Boundary BZ78a 24200c
930 Beta-168207 29120F270 Turbidited
990 Beta-166996 37900F1200 Turbidited
111 Boundary BZ89 32100c
1520 Beta-133928 36070F300 39855e
1890 Beta-166997 44300F2000 47126e
a Ramsey et al (2004)b Converted into calendar age using the program bcalib 44Q
(Stuiver and Reimer 1993) DR=88F54 (Reimer 2000)c From the study by Sbaffi et al (2001) and Capotondi et al
(1999)d These dates were not used in the construction of the age model
curve in Fig 2e Converted into calendar age with the polynomial provided by
Bard et al (2004)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332314
1994 Aksu et al 1995) Based on the fluctuations of
Corg content (Fig 3) the sapropel sequence can be
divided into three zones a lower and an upper zone
which are characterized by a maximum Corg content
of 21 and 16 respectively and a middle zone
separating the two others with a minimum Corg
content of 11 The lower (40ndash35 cm) and the upper
(30ndash26 cm) zones of high Corg content will hereforth
be referred to as S1a and S1b respectively Previous
studies on the formation of S1 have also described an
interruption in the deposition of the sapropel sedi-
ments (Rossignol-Strick et al 1982 Rohling et al
1997 Geraga et al 2000)
The third unit C (44ndash87 cm) consists of brownish
structureless mud with rare shell fragments The
fourth unit D (87ndash132 cm) is similar to unit C but
is grayish in colour Two layers of structureless silt
appear between 93 and 105 cm The lower boundary
of this interval at 105 cm is sharp and the upper one at
93 cm appears gradational The boundary between the
two layers is indistinct and occurs at approximately 97
cm The two layers correspond to two different
turbidites according to their sedimentological charac-
teristics The lower turbidite layer between 105 and 97
cm consists of material dated at 37900 14C kyr BP
(Table 1) and the upper between 97 and 93 cm
consists of material dated at 29120 14C kyr BP (Table
1) suggesting that the supplying source of each
turbidite was situated at two different locations
The fifth unit E (132ndash153 cm) represents a
sapropelitic sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) Within this sequence the max-
imum Corg content (05ndash154) occurs between 153
and 143 cm This sapropelic layer consists of olive
coloured structureless mud During the interval that
follows between 143 and 132 cm the Corg content
declines from 05 to 03 Based on the AMS
dating (Table 1) the age of this sapropelitic sequence
is 355ndash335 14C kyr BP and seems to correspond to
the S2 sapropel The rare appearance of sapropel S2
often called the bghost sapropelQ in the sediments of
the eastern Mediterranean has led to approximate
estimations of its age (Ryan 1972 Vergnaud-Grazzini
et al 1977)
The sixth unit F (153ndash195 cm) consists of
brownish mud
The sediments of units C D and E contain sand
and silt sized volcanic ash almost everywhere but no
discrete ash layers The highest concentration of the
ash was obtained between 97 and 125 cm
32 Age assessment
The chronological framework of the present study
is based on the five AMS dates (Table 1) The
turbidite layers have not been taken into consideration
in the construction of the age model curve for the
estimation of the sedimentation rate
The first three dates (b24 kyr BP) were converted
into calendar years using the program bcalib 44Q(Stuiver and Reimer 1993 Reimer 2000) while the
other two were converted using the polynomials
provided by Bard et al (2004) (Table 1)
In addition four dated control points were used for
the age assessment (Table 1) The upper dating point
is based on the age (357 cal kyr BP) (Ramsey et al
2004) of the ash layer at 15 cm which is considered to
be the Z2 Santorini ash layer The other three are
based on the biostratigraphic study by Sbaffi et al
(2001) That study has revealed 9 biozones over the
last 34 kyr based on changes in the planktonic
foraminifera and coccolithophores assemblages in the
Fig 2 (a) Lithology of core C69 Ages displayed to the left of the core are AMS 14Cnc dates (b) Age model curve for the estimation of the mean
sedimentation rate The diagram is based on five calibrated AMS 14C ages (arrows) and four dating points (in cal yr BP) based on (i) the
Santorini Z2 ash layer (Ramsey et al 2004) (square) and (ii) the estimated age for upper and lower boundaries of biozone 7 and upper boundary
of biozone 9 by Sbaffi et al (2001) (circles) The turbidite layers have not been taken into consideration in the construction of the age model
curve for the estimation of the sedimentation rate Thus the older dating points (at 47126 cal kyr BP 39855 cal kyr BP and 32100 cal kyr BP)
appear in shallower depths in the age model curve than in the core
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 315
central Mediterranean dated in calendar years BP
These biozones are comparable with the ecozones
proposed by Capotondi et al (1999) and are detect-
able throughout the central Mediterranean and as far
as the southern Adriatic Sea (Giunta et al 2003
Principato et al 2003) In our study the lower and the
upper boundaries (in calendar years BP) of the
biozone 7 and the upper boundary of biozone 9 were
used The former which corresponds to the ecozone 9
includes the Last Glacial Maximum interval and is
well identified in our records by the planktonic
assemblages and the isotopic signal The isotopic
Fig 3 Downcore variation of (a) y18O values (x) and (b) Corg content () versus age in cal kyr BP
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332316
record for this interval is also consistent with the
isotopic record in the study of Siani et al (2001) in the
southern Adriatic Sea suggesting a realistic age
control Biozone 9 represents a warm interval
corresponding to interstadial 5 (Cacho et al 1999)
In our data biozone 9 is well identified by the
planktonic assemblages and the depletions of the
isotope values
Based on the above age model curve the mean
sedimentation rate in core C69 is 38 cmkyr (Fig 2)
The mean time resolution of planktonic and MAT
records is 450 years while that of oxygen isotopes
and pollen is 850 yr and 550 yr respectively
33 Oxygen isotopes
The downcore variation of the y18O values in core
C69 seen in Fig 3 reveals that in the isotopic stage
ST3 (48ndash24 cal kyr BP) the y18O data exhibits
relatively lower values between 40 and 38 cal kyr BP
(~00x) and around 34 cal kyr BP (06x) The
former corresponds to the deposition of the S2
sapropel and the latter corresponds to the Interstadial
5 (Ist5) which has been also identified in cores from
the central and western Mediterranean (Cacho et al
1999 Sbaffi et al 2001 Perez-Folgado et al 2003)
The small decrease in y18O values (19x) around 24
cal kyr BP may correspond to the isotopic event 31
(Martinson et al 1987) High y18O values occur at
time intervals from 415 to 405 cal kyr BP (18ndash
19x) 355ndash345 cal kyr BP (15ndash13x) and 29ndash27
cal kyr BP (27x) The first and the third time
intervals are around the ages of the Heinrich 4 (39 cal
kyr BP) and Heinrich 3 (30 cal kyr BP) events
respectively (Bond et al 1992)
In the isotopic stage ST2 (24ndash11 cal kyr BP) the
strongest enrichment of y18O values (28ndash25x) occur
between 215 and 195 cal kyr BP suggesting the Last
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

Table 1
AMS dates and dated control points used in core C69
Depth (cm) AMS laboratory
code or source
Uncorrected
AMS 14C
(yr BP)
Calibrated
(yr BP)
150 Z2 Santorini
ash layer
3570a
280 Beta-166995 5810F40 5964ndash6269b
400 Beta-142621 8750F70 8919ndash9458b
600 Beta-190861 14470F70 16164ndash17173b
64 Boundary BZ67 18100c
84 Boundary BZ78a 24200c
930 Beta-168207 29120F270 Turbidited
990 Beta-166996 37900F1200 Turbidited
111 Boundary BZ89 32100c
1520 Beta-133928 36070F300 39855e
1890 Beta-166997 44300F2000 47126e
a Ramsey et al (2004)b Converted into calendar age using the program bcalib 44Q
(Stuiver and Reimer 1993) DR=88F54 (Reimer 2000)c From the study by Sbaffi et al (2001) and Capotondi et al
(1999)d These dates were not used in the construction of the age model
curve in Fig 2e Converted into calendar age with the polynomial provided by
Bard et al (2004)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332314
1994 Aksu et al 1995) Based on the fluctuations of
Corg content (Fig 3) the sapropel sequence can be
divided into three zones a lower and an upper zone
which are characterized by a maximum Corg content
of 21 and 16 respectively and a middle zone
separating the two others with a minimum Corg
content of 11 The lower (40ndash35 cm) and the upper
(30ndash26 cm) zones of high Corg content will hereforth
be referred to as S1a and S1b respectively Previous
studies on the formation of S1 have also described an
interruption in the deposition of the sapropel sedi-
ments (Rossignol-Strick et al 1982 Rohling et al
1997 Geraga et al 2000)
The third unit C (44ndash87 cm) consists of brownish
structureless mud with rare shell fragments The
fourth unit D (87ndash132 cm) is similar to unit C but
is grayish in colour Two layers of structureless silt
appear between 93 and 105 cm The lower boundary
of this interval at 105 cm is sharp and the upper one at
93 cm appears gradational The boundary between the
two layers is indistinct and occurs at approximately 97
cm The two layers correspond to two different
turbidites according to their sedimentological charac-
teristics The lower turbidite layer between 105 and 97
cm consists of material dated at 37900 14C kyr BP
(Table 1) and the upper between 97 and 93 cm
consists of material dated at 29120 14C kyr BP (Table
1) suggesting that the supplying source of each
turbidite was situated at two different locations
The fifth unit E (132ndash153 cm) represents a
sapropelitic sequence (Kidd et al 1978 Anastasakis
and Stanley 1984) Within this sequence the max-
imum Corg content (05ndash154) occurs between 153
and 143 cm This sapropelic layer consists of olive
coloured structureless mud During the interval that
follows between 143 and 132 cm the Corg content
declines from 05 to 03 Based on the AMS
dating (Table 1) the age of this sapropelitic sequence
is 355ndash335 14C kyr BP and seems to correspond to
the S2 sapropel The rare appearance of sapropel S2
often called the bghost sapropelQ in the sediments of
the eastern Mediterranean has led to approximate
estimations of its age (Ryan 1972 Vergnaud-Grazzini
et al 1977)
The sixth unit F (153ndash195 cm) consists of
brownish mud
The sediments of units C D and E contain sand
and silt sized volcanic ash almost everywhere but no
discrete ash layers The highest concentration of the
ash was obtained between 97 and 125 cm
32 Age assessment
The chronological framework of the present study
is based on the five AMS dates (Table 1) The
turbidite layers have not been taken into consideration
in the construction of the age model curve for the
estimation of the sedimentation rate
The first three dates (b24 kyr BP) were converted
into calendar years using the program bcalib 44Q(Stuiver and Reimer 1993 Reimer 2000) while the
other two were converted using the polynomials
provided by Bard et al (2004) (Table 1)
In addition four dated control points were used for
the age assessment (Table 1) The upper dating point
is based on the age (357 cal kyr BP) (Ramsey et al
2004) of the ash layer at 15 cm which is considered to
be the Z2 Santorini ash layer The other three are
based on the biostratigraphic study by Sbaffi et al
(2001) That study has revealed 9 biozones over the
last 34 kyr based on changes in the planktonic
foraminifera and coccolithophores assemblages in the
Fig 2 (a) Lithology of core C69 Ages displayed to the left of the core are AMS 14Cnc dates (b) Age model curve for the estimation of the mean
sedimentation rate The diagram is based on five calibrated AMS 14C ages (arrows) and four dating points (in cal yr BP) based on (i) the
Santorini Z2 ash layer (Ramsey et al 2004) (square) and (ii) the estimated age for upper and lower boundaries of biozone 7 and upper boundary
of biozone 9 by Sbaffi et al (2001) (circles) The turbidite layers have not been taken into consideration in the construction of the age model
curve for the estimation of the sedimentation rate Thus the older dating points (at 47126 cal kyr BP 39855 cal kyr BP and 32100 cal kyr BP)
appear in shallower depths in the age model curve than in the core
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 315
central Mediterranean dated in calendar years BP
These biozones are comparable with the ecozones
proposed by Capotondi et al (1999) and are detect-
able throughout the central Mediterranean and as far
as the southern Adriatic Sea (Giunta et al 2003
Principato et al 2003) In our study the lower and the
upper boundaries (in calendar years BP) of the
biozone 7 and the upper boundary of biozone 9 were
used The former which corresponds to the ecozone 9
includes the Last Glacial Maximum interval and is
well identified in our records by the planktonic
assemblages and the isotopic signal The isotopic
Fig 3 Downcore variation of (a) y18O values (x) and (b) Corg content () versus age in cal kyr BP
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332316
record for this interval is also consistent with the
isotopic record in the study of Siani et al (2001) in the
southern Adriatic Sea suggesting a realistic age
control Biozone 9 represents a warm interval
corresponding to interstadial 5 (Cacho et al 1999)
In our data biozone 9 is well identified by the
planktonic assemblages and the depletions of the
isotope values
Based on the above age model curve the mean
sedimentation rate in core C69 is 38 cmkyr (Fig 2)
The mean time resolution of planktonic and MAT
records is 450 years while that of oxygen isotopes
and pollen is 850 yr and 550 yr respectively
33 Oxygen isotopes
The downcore variation of the y18O values in core
C69 seen in Fig 3 reveals that in the isotopic stage
ST3 (48ndash24 cal kyr BP) the y18O data exhibits
relatively lower values between 40 and 38 cal kyr BP
(~00x) and around 34 cal kyr BP (06x) The
former corresponds to the deposition of the S2
sapropel and the latter corresponds to the Interstadial
5 (Ist5) which has been also identified in cores from
the central and western Mediterranean (Cacho et al
1999 Sbaffi et al 2001 Perez-Folgado et al 2003)
The small decrease in y18O values (19x) around 24
cal kyr BP may correspond to the isotopic event 31
(Martinson et al 1987) High y18O values occur at
time intervals from 415 to 405 cal kyr BP (18ndash
19x) 355ndash345 cal kyr BP (15ndash13x) and 29ndash27
cal kyr BP (27x) The first and the third time
intervals are around the ages of the Heinrich 4 (39 cal
kyr BP) and Heinrich 3 (30 cal kyr BP) events
respectively (Bond et al 1992)
In the isotopic stage ST2 (24ndash11 cal kyr BP) the
strongest enrichment of y18O values (28ndash25x) occur
between 215 and 195 cal kyr BP suggesting the Last
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254
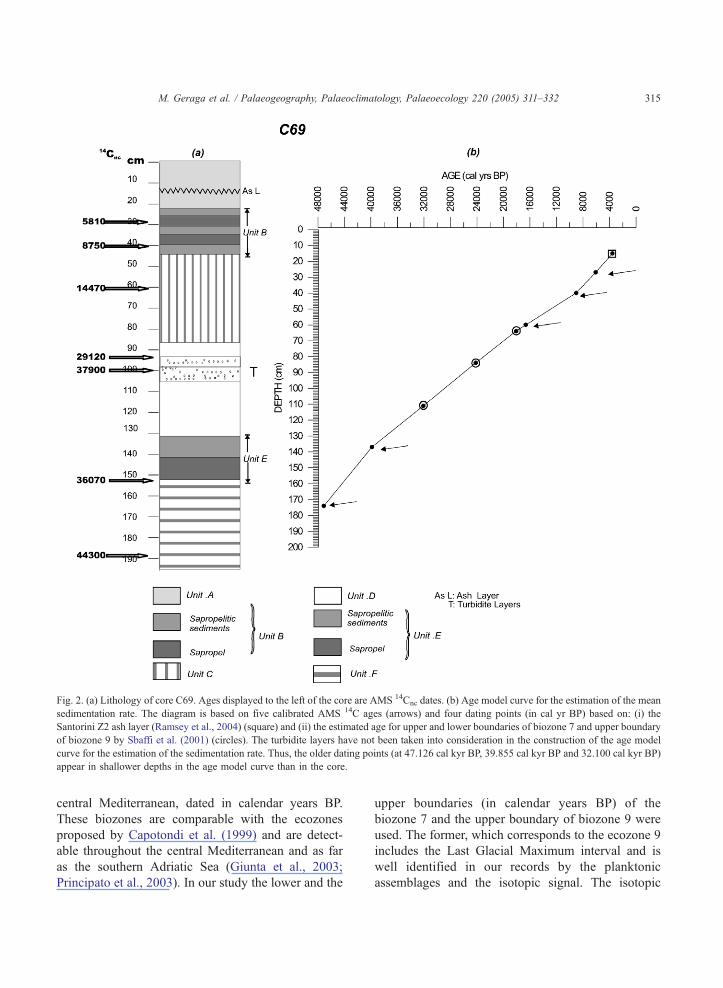
Fig 2 (a) Lithology of core C69 Ages displayed to the left of the core are AMS 14Cnc dates (b) Age model curve for the estimation of the mean
sedimentation rate The diagram is based on five calibrated AMS 14C ages (arrows) and four dating points (in cal yr BP) based on (i) the
Santorini Z2 ash layer (Ramsey et al 2004) (square) and (ii) the estimated age for upper and lower boundaries of biozone 7 and upper boundary
of biozone 9 by Sbaffi et al (2001) (circles) The turbidite layers have not been taken into consideration in the construction of the age model
curve for the estimation of the sedimentation rate Thus the older dating points (at 47126 cal kyr BP 39855 cal kyr BP and 32100 cal kyr BP)
appear in shallower depths in the age model curve than in the core
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 315
central Mediterranean dated in calendar years BP
These biozones are comparable with the ecozones
proposed by Capotondi et al (1999) and are detect-
able throughout the central Mediterranean and as far
as the southern Adriatic Sea (Giunta et al 2003
Principato et al 2003) In our study the lower and the
upper boundaries (in calendar years BP) of the
biozone 7 and the upper boundary of biozone 9 were
used The former which corresponds to the ecozone 9
includes the Last Glacial Maximum interval and is
well identified in our records by the planktonic
assemblages and the isotopic signal The isotopic
Fig 3 Downcore variation of (a) y18O values (x) and (b) Corg content () versus age in cal kyr BP
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332316
record for this interval is also consistent with the
isotopic record in the study of Siani et al (2001) in the
southern Adriatic Sea suggesting a realistic age
control Biozone 9 represents a warm interval
corresponding to interstadial 5 (Cacho et al 1999)
In our data biozone 9 is well identified by the
planktonic assemblages and the depletions of the
isotope values
Based on the above age model curve the mean
sedimentation rate in core C69 is 38 cmkyr (Fig 2)
The mean time resolution of planktonic and MAT
records is 450 years while that of oxygen isotopes
and pollen is 850 yr and 550 yr respectively
33 Oxygen isotopes
The downcore variation of the y18O values in core
C69 seen in Fig 3 reveals that in the isotopic stage
ST3 (48ndash24 cal kyr BP) the y18O data exhibits
relatively lower values between 40 and 38 cal kyr BP
(~00x) and around 34 cal kyr BP (06x) The
former corresponds to the deposition of the S2
sapropel and the latter corresponds to the Interstadial
5 (Ist5) which has been also identified in cores from
the central and western Mediterranean (Cacho et al
1999 Sbaffi et al 2001 Perez-Folgado et al 2003)
The small decrease in y18O values (19x) around 24
cal kyr BP may correspond to the isotopic event 31
(Martinson et al 1987) High y18O values occur at
time intervals from 415 to 405 cal kyr BP (18ndash
19x) 355ndash345 cal kyr BP (15ndash13x) and 29ndash27
cal kyr BP (27x) The first and the third time
intervals are around the ages of the Heinrich 4 (39 cal
kyr BP) and Heinrich 3 (30 cal kyr BP) events
respectively (Bond et al 1992)
In the isotopic stage ST2 (24ndash11 cal kyr BP) the
strongest enrichment of y18O values (28ndash25x) occur
between 215 and 195 cal kyr BP suggesting the Last
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254
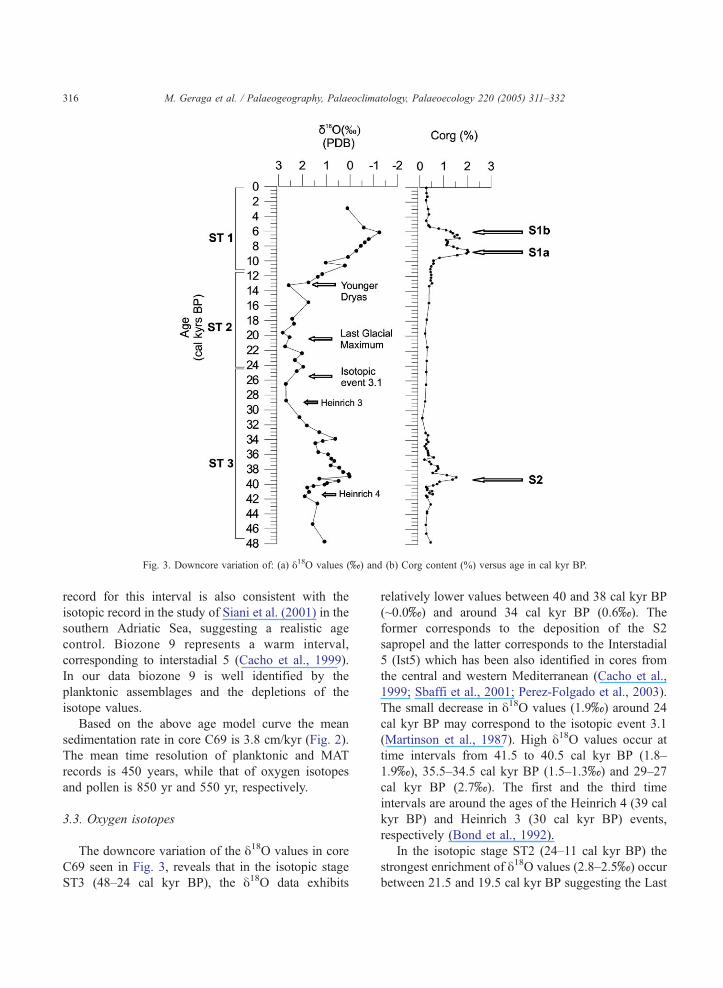
Fig 3 Downcore variation of (a) y18O values (x) and (b) Corg content () versus age in cal kyr BP
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332316
record for this interval is also consistent with the
isotopic record in the study of Siani et al (2001) in the
southern Adriatic Sea suggesting a realistic age
control Biozone 9 represents a warm interval
corresponding to interstadial 5 (Cacho et al 1999)
In our data biozone 9 is well identified by the
planktonic assemblages and the depletions of the
isotope values
Based on the above age model curve the mean
sedimentation rate in core C69 is 38 cmkyr (Fig 2)
The mean time resolution of planktonic and MAT
records is 450 years while that of oxygen isotopes
and pollen is 850 yr and 550 yr respectively
33 Oxygen isotopes
The downcore variation of the y18O values in core
C69 seen in Fig 3 reveals that in the isotopic stage
ST3 (48ndash24 cal kyr BP) the y18O data exhibits
relatively lower values between 40 and 38 cal kyr BP
(~00x) and around 34 cal kyr BP (06x) The
former corresponds to the deposition of the S2
sapropel and the latter corresponds to the Interstadial
5 (Ist5) which has been also identified in cores from
the central and western Mediterranean (Cacho et al
1999 Sbaffi et al 2001 Perez-Folgado et al 2003)
The small decrease in y18O values (19x) around 24
cal kyr BP may correspond to the isotopic event 31
(Martinson et al 1987) High y18O values occur at
time intervals from 415 to 405 cal kyr BP (18ndash
19x) 355ndash345 cal kyr BP (15ndash13x) and 29ndash27
cal kyr BP (27x) The first and the third time
intervals are around the ages of the Heinrich 4 (39 cal
kyr BP) and Heinrich 3 (30 cal kyr BP) events
respectively (Bond et al 1992)
In the isotopic stage ST2 (24ndash11 cal kyr BP) the
strongest enrichment of y18O values (28ndash25x) occur
between 215 and 195 cal kyr BP suggesting the Last
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 317
Glacial Maximum High positive values ranging from
3x to 48x have been observed for the same interval
in the eastern Mediterranean (Vergnaud-Grazzini et
al 1977 1986 Thunell and Williams 1989)
Towards the end of the ST2 the gradual depletion of
the y18O values from 24x to 12x indicates the
onset of deglaciation during the Late Glacial period
(Vergnaud-Grazzini et al 1986 Thunell and Wil-
liams 1989) The rapid and temporal enrichment of
y18O at 13 cal kyr BP (26x) may correspond to the
Younger Dryas event which has also been identified
in cores from the central Mediterranean (Vergnaud-
Grazzini et al 1977 1986)
During the Holocene (ST1) the most depleted
values of y18O occur during the sapropel sequence S1
(12x) High negative values of y18O ranging
between 00x and 20x are documented in all
palaeoceanographic records of the eastern Mediterra-
nean indicating the presence of warm climate
accompanied by the establishment of a low salinity
surface layer during that interval (Cita et al 1977
Williams et al 1978 Aksu et al 1995) The
depletion of y18O between the full glacial and the
Holocene conditions is estimated at around 43x in
the eastern Mediterranean Sea (Thunell and Williams
1989) Before the deposition of S1 a short and rapid
shift to higher value of y18O (10x) occurs at 10 cal
kyr BP and probably corresponds to the first Holocene
stadial (Bond et al 1997) Just after the deposition of
S1 the y18O values appear to increase to 0x
34 Planktonic foraminifera biostratigraphy
Planktonic foraminifera are abundant throughout
the core and the downcore variations of their
abundance are shown in Figs 4 and 5
341 Globigerinoides ruber (white and pink
varieties)
This species which is considered as indicative of
warm and oligotrophic surface waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) is present
throughout the core covering between 2 and 41
of the total planktonic fauna (Fig 4A)
The Globigerinoides ruber abundance between 48
and 42 cal kyr BP ranges between 21 and 28 Its
abundance between 42 and 14 cal kyr BP is low less
than 15 Towards the end of the Glacial period
between 14 and 10 cal kyr BP G ruber increases
gradually from 10 to 20 A sharp decrease of G
ruber at ~12 cal kyr BP (8) (Fig 4A) is just after the
enrichment of y18O at 13 cal kyr BP and may
therefore be attributed to the Younger Dryas event
A similar decrease in the abundance of G ruber
between 135 and 125 cal kyr BP has been recorded
in core C40 in the vicinity of the study area in the
Myrtoon Basin (Geraga et al 2000)
During the Holocene the presence of Globiger-
inoides ruber is dominant (N20) and appears in
maximum percentages between 95 and 8 cal kyr BP
(35ndash42) between 65 and 6 cal kyr BP (26ndash36)
and between 4 and 25 cal kyr BP (27ndash33) The
former two intervals almost coincide with the for-
mation of the two sapropelic layers S1a and S1b
respectively Increase of G ruber during the deposi-
tion of S1 have been observed throughout all of the
eastern Mediterranean (Casford et al 2002 Princi-
pato et al 2003) Temporal decreases in the abun-
dance of G ruber occur at 10 cal kyr BP 8ndash65 cal
kyr BP and 55 cal kyr BP The first coincides with
enrichment of values of y18O and is related to the first
Holocene stadial The other two coincide with the
interruption and the end of S1 respectively During
the Holocene a decrease in the abundance of G ruber
have also been observed in core C40 in the Myrtoon
Basin at 95 cal kyr BP 8ndash75 cal kyr BP and 7 cal
kyr BP
The high abundance of Globigerinoides ruber
during the late Glacial and Holocene periods has
been observed all throughout the Mediterranean Sea
(Buckley et al 1982 Hayes et al 1999)
342 Minor warm planktonic indicators
Other planktonic species recognized in the core
and indicative of warm surface waters are Globiger-
inoides sacculifer Globigerinoides tenellus Globi-
gerinoides trilobus Globigerinoides obliquus
Orbulina universa and Globigerinella aequilateralis
Their presence in the planktonic assemblages are
generally low (b10) and discontinuous (Fig 5) G
sacculifer is present only in the last 10 kyr and
exhibits the highest percentages (~8) between 7 and
6 cal kyr BP which almost coincides with the
formation of the sapropelic layer S1b This also
occurred in core C40 selected from the Myrtoon
Basin (Geraga et al 2000) An increase in abundance
Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

Fig 4 Downcore variation of selected planktonic species versus age in calibrated kyr BP On the right side of each diagram are shown the intervals where the species abundance peaks
in cores from the western Mediterranean Sea according to Perez-Folgado et al 2003 (Ro to Ra2 high abundance of Globigerinoides ruber alba R1a to R1c high abundance of
Globigerinoides ruber pink Q1 to Q6 high abundance of Turborotalita quinqueloba B1 to B4 high abundance of Globigerina bulloides I0 to I5 high abundance of Globorotalia
inflata P1 to P5 high abundance of Neogloboquadrina pachyderma (d) Pm3 decrease of N pachyderma (d) SPDF Small Foraminifera Dominance Zone recorded in cores from
the western and the central Mediterranean) (Pujol and Vergnaud Grazzini 1995 Perez-Folgado et al 2003) BZ The boundaries of the planktonic biozones in the central
Mediterranean (Sbaffi et al 2001)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
318
Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

Fig 5 Downcore variation of selected planktonic species against age in calibrated kyr BP (left axis) and radiocarbon kyr BP (right axis) BZ
The boundaries of the planktonic biozones in the central Mediterranean (Sbaffi et al 2001) Stippled bands show the zones in the sapropel
sequence with TOC contentN13
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 319
of G sacculifer at the top-most of S1 has been
observed in cores from the eastern Mediterranean and
has been related to the occurrence of a surface
pycnocline during that period (Principato et al
2003)
Globigerinoides trilobus is absent in the planktonic
assemblages between 47 and 18 cal kyr BP except for
two intervals between 48 and 44 cal kyr BP and
between 38 and 36 cal kyr BP where its percentage is
always b5 This species is present during the last 18
kyr but in very low percentages and shows a
maximum (4) between 75 and 55 cal kyr BP
around the formation of the S1b layer
Orbulina universa occurs only between 48 and 44
cal kyr BP between 40 and 34 cal kyr BP between 28
and 24 cal kyr BP and during the last 11 kyr Its
presence is generally low and exhibits the highest
percentages between 10 and 6 cal kyr BP (5ndash10)
Globigerinella aequilateralis is present in the core
only during the last 11 kyr but its percentages are
always lower than 5
The increases in all the aforementioned species
during the Holocene coincide with the decrease in the
y18O values suggesting a general increase in the sea
surface temperature The appearance of the above-
mentioned warm species between 16 and 11 14C kyr
BP and the following increase in abundance have
been observed in cores across the Mediterranean
(Buckley and Johnson 1988 Jorissen et al 1993
Hayes et al 1999)
343 Globigerina bulloides
This species is indicative of sub-polar water masses
highly dependent on enhanced food levels (upwelling
strong seasonal mixing or freshwater inputs (Lourens
et al 1994) Its presence is significant throughout the
core by a mean percentage of 11 (Fig 4C)
Globigerina bulloides occurs in highest abundance
between 48 and 43 cal kyr BP (10ndash23) between 35
and 32 cal kyr BP (10ndash21) between 26 and 18 cal
kyr BP (6ndash15) between 16 and 13 (5) and
between 9 and 55 cal kyr BP (7ndash30) during the
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332320
formation of the sapropel sequence of S1 During the
interruption of S1 G bulloides appears slightly
decreased
An increase in the abundance of Globigerina
bulloides during the formation of sapropel S1 has
also been observed in most of the cores in the
eastern Mediterranean (Aksu et al 1995 Rohling
et al 1997 Geraga et al 2000) This increase has
been attributed to lower oxygen content within the
photic zone since this species can survive at the
end of phyto- and zooplankton blooms when
oxygen had been rapidly consumed (Principato et
al 2003)
344 Globigerinita glutinata
This cosmopolitan species has a mean participa-
tion in the planktonic assemblages of about 9 (Fig
5) The concentration is high during the Glacial
period between 43 and 40 cal kyr BP (6ndash18)
Globigerinita glutinata appears to increase gradually
from 9 to 37 between 28 and 95 cal kyr BP
with two declines at 19 cal kyr BP (3) and 16 cal
kyr BP (9) The maximum percentage of G
glutinata (23ndash37) occurs between 125 and 95
cal kyr BP during the transition from the Last
Glacial period to Holocene G glutinata is almost
absent during the formation of the two sapropelic
layers S1a and S1b This trend has also been
observed in the sediments of cores throughout the
Aegean Sea (Casford et al 2002)
345 Globorotalia truncatulinoides
The abundance of this species in the Mediterranean
seems to be controlled mainly by the winter con-
vection and vertical mixing (Pujol and Vergnaud
Grazzini 1995) In C69 it is mainly absent except
between 40 and 34 cal kyr BP and between 18 and 9
cal kyr BP when it never exceeds 4 in the planktonic
assemblages (Fig 5)
346 Globorotalia inflata
This species in the Mediterranean is associated
with cool and deep mixed waters (Thunell 1978
Pujol and Vergnaud Grazzini 1995) In C69 (Fig 4D)
it participates with a low mean percentage (3) in the
planktonic fauna Globorotalia inflata appears to
increase slightly between 43 and 40 cal kyr BP (2ndash
8) between 36 and 33 cal kyr BP (5ndash12) between
26 and 23 cal kyr BP (2ndash5) and between 155 and
12 cal kyr BP (2ndash3)
During the Holocene it appears in low percen-
tages except for the interruption of S1 at 8 cal kyr
BP and immediately after the end of S1 at 5 cal kyr
BP when its percentages are 1ndash4 and 5ndash13
respectively The rapid and temporal increase of
Globorotalia inflata after the end of the sapropel S1
is documented in almost all the cores from the
eastern Mediterranean and is attributed to deep water
ventilation (De Rijk et al 1999 Geraga et al 2000
Casford et al 2002)
347 Globorotalia scitula
This species which is considered as indicative of
cool-subpolar waters (Thunell 1978 Lourens et al
1994) is absent in the last 10 kyr throughout the
Holocene and appears only during the Glacial period
(Fig 4F) Its participation in the planktonic assemb-
lages never exceeds 8 in the total fauna The
highest abundances of Globorotalia scitula are
recorded between 42 and 38 cal kyr BP (4ndash8) and
between 255 and 22 cal kyr BP (1ndash5)
348 Turborotalita quinqueloba
This species is considered as indicative of cool
waters (Thunell 1978 Lourens et al 1994) but is
also tolerant to fairly low salinity andor enhanced
fertility in surficial waters (Rohling et al 1997) In
C69 (Fig 4B) Turborotalita quinqueloba constitutes
a significant species and its mean percentage is 18
in the total fauna T quinqueloba from 46 to 42 cal
kyr BP and from 38 to 34 cal kyr BP appears in
relatively low percentages (5ndash12) A high abun-
dance of T quinqueloba occurs between 42 and 40
cal kyr BP (22ndash35) and from 32 to 26 cal kyr BP
(30ndash65) and coincides with an enrichment of y18Oin the same intervals (Fig 2) Between 26 and 10 cal
kyr BP T quinqueloba exhibits a gradual decline
from about 65 to 10 However during this
interval abrupt increases in T quinqueloba occur at
23 cal kyr BP (47) 195 cal kyr BP (35) 175ndash
165 cal kyr BP (25) and 13 cal kyr BP (18)
During these intervals the y18O data exhibits high
positive values During the last 11 kyr the abundance
of T quinqueloba ranges between 5 and 10
except for a spike (15) at 65 cal kyr BP during the
interruption of S1
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 321
349 Neogloboquadrina dutertrei Neogloboqua-
drina pachyderma
Neogloboquadrinid species are indicative of cool
waters and also of the eutrophicated waters associated
with the formation of a Deep Chlorophyll Maximum
(DCM) layer (Fairbanks and Wiebe 1980) Among
these species Neogloboquadrina pachyderma (s)
thrives in colder waters (Pujol and Vergnaud Grazzini
1995) In C69 N pachyderma (s) is almost absent as
only rare specimens occur during the glacial period
The presence of Neogloboquadrina dutertrei (Fig 5)
ranges between 0 and 21 that of N pachyderma
(d) (Fig 4E) between 0 and 37 and these two
species present similar trends in their downcore
variations The highest percentages of both species
though with abrupt fluctuations occur between 40 and
32 cal kyr BP and between 22 and 12 cal kyr BP
Spikes of Neogloboquadrina pachyderma occur at
40ndash33 cal kyr BP (20ndash37) (which includes the
deposition of S2) 29 cal kyr BP (14) 20ndash19 cal kyr
BP (23ndash25) and 17ndash13 cal kyr BP (28ndash22)
During the Holocene N pachyderma appears in
low percentages (b4) or is absent Slight increases
occur around 8 cal kyr BP during the interruption of
S1 (2ndash5) and around 6ndash5 cal kyr BP immediately
after the end of S1 (6ndash9)
Similar trends in the downcore variation of Neo-
globoquadrina pachyderma (d) for the Late Glacial
period and Holocene have been observed all through-
out the eastern Mediterranean (Casford et al 2002
Principato et al 2003)
35 Pollen
Pollen and spores are well preserved between 42
and 35 cal kyr BP and for the last 25 kyr and the
downcore variations of selected taxa presented in Fig
6 correspond to these intervals
The pollen spectrum ranges from semi-desert and
steppe to deciduous and coniferous forests In general
the pollen stratigraphy of the C69 core shows
fluctuations from open vegetation communities to
forest in response to glacialndashinterglacial climatic
conditions The forested periods are characterized by
the significant presence of Quercus ilexcoccifera
type followed by Corylus Carpinus Ulmus and
sometimes Fagus The steppic environment is char-
acterized by the significant presence of Gramineae
Chenopodiaceae Compositae and Artemisia The
main change in the pollen records occurs at 11 cal
kyr BP where the Mediterranean taxa appear and
afterwards remain constant suggesting the ameliora-
tion of the climate during the Holocene
For the intervals 42 to 35 cal kyr BP and 25 to 10
cal kyr BP coniferous and cool-temperate deciduous
trees were continuously present accounting for 25ndash
55 and 25ndash70 of the total palynomorphs assemb-
lages respectively (Figs 6 and 7E) The almost
continuous presence of cool-temperate trees in these
intervals is evident in all pollen records in marine ant
terrestrial sediments of Greece and the surrounding
seas and indicates relatively mild wet climatic
conditions (Tzedakis et al 2002)
Variations in APNAP values also occur in the
abovementioned intervals (Fig 7E) Between 42 and
35 cal kyr BP the NAP values exhibit three peaks at
41ndash40 cal kyr BP 375 cal kyr BP and 35 cal kyr BP
(Fig 7E) During these peaks Gramineae and Com-
positae appear together with Chenopodiaceae Arte-
misia and Polygonaceae (Figs 6 and 7EF)
suggesting the prevalence of low temperature and
low annual precipitation (Tarasov et al 1998 Peyron
et al 1998) However the co-existence of the boreal
taxa Alnus Betula and the cool-temperate elements
Fagus Corylus and Cedrus (Fig 6) indicates
sufficient precipitation and temperatures not below
that of a tundra in the uplands (Walter 1974
Rossignol-Strick et al 1992) The first of these
intervals coincides with the strong enrichment of
y18O values which is related to the period of H4
event Increased y18O values also occur at the time of
the third interval
The increase of AP values at 40 cal kyr BP and
between 37 and 36 cal kyr BP which account for 70
of the total palynomorphs with the concurrent decline
of cool steppic and semi-desertic taxa and the increase
in the warm- and cool-temperate taxa (Fig 6) suggests
an increase in humidity The peak at 40 cal kyr BP
occurs just before the deposition of S2 (39ndash385 cal
kyr BP) and coincides with a depletion of y18O values
Between 25 and 11 cal kyr BP Gramineae
Chenopodiaceae Compositae and Artemisia domi-
nate indicating the presence of a steppic environment
(Fig 6) The former exhibits constant percentage
abundance while the latter three exhibit variable
percentage abundance
Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

Fig 6 Pollen diagrams of selected taxa The percentage occurrence values of each taxa are plotted against a calibrated radiocarbon time scale Cool-temperate summergreen taxa
include Carpinus Corylus Fagus Tilia and Ulmus Warm-temperate summergreen taxa include Platanus Ostrya and Fraxinus Mediterranean taxa include Olea Pistacia and
Phyllirea Other woody taxa include Acer Juglans and Salix Other herbaceous taxa include Ranunculus Rosaceae and Rumex Stippled bands show the zones in the sapropel
sequence with TOC content N13
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
322
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254
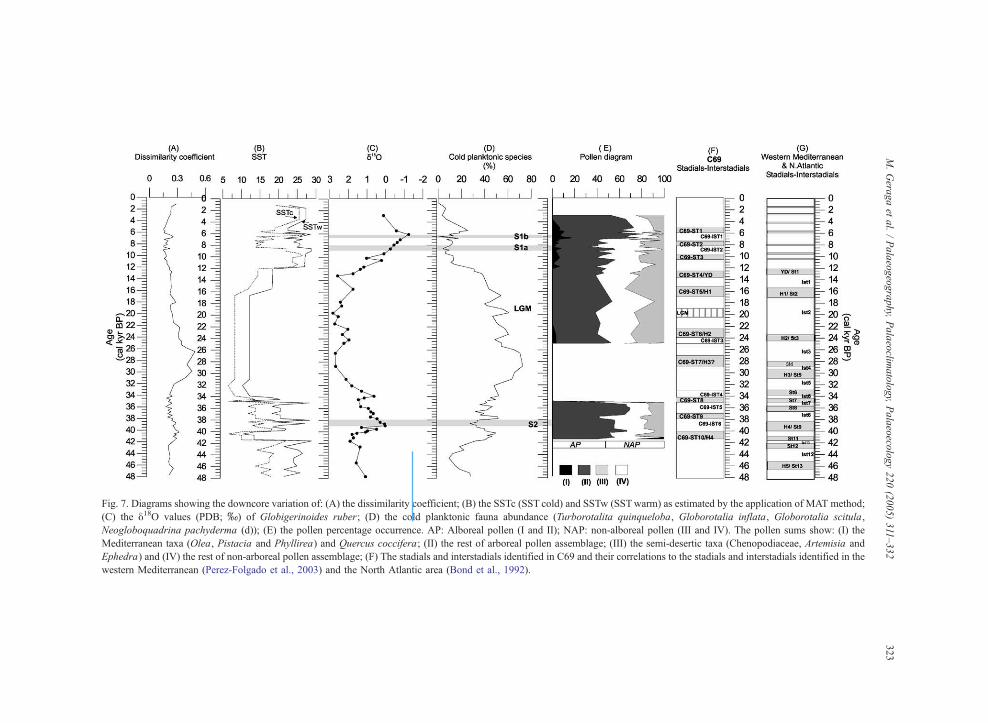
Fig 7 Diagrams showing the downcore variation of (A) the dissimilarity coefficient (B) the SSTc (SST cold) and SSTw (SSTwarm) as estimated by the application of MAT method
(C) the y18O values (PDB x) of Globigerinoides ruber (D) the cold planktonic fauna abundance (Turborotalita quinqueloba Globorotalia inflata Globorotalia scitula
Neogloboquadrina pachyderma (d)) (E) the pollen percentage occurrence AP Alboreal pollen (I and II) NAP non-alboreal pollen (III and IV) The pollen sums show (I) the
Mediterranean taxa (Olea Pistacia and Phyllirea) and Quercus coccifera (II) the rest of arboreal pollen assemblage (III) the semi-desertic taxa (Chenopodiaceae Artemisia and
Ephedra) and (IV) the rest of non-arboreal pollen assemblage (F) The stadials and interstadials identified in C69 and their correlations to the stadials and interstadials identified in the
western Mediterranean (Perez-Folgado et al 2003) and the North Atlantic area (Bond et al 1992)
MGera
gaet
alPalaeogeographyPalaeoclim
atologyPalaeoeco
logy220(2005)311ndash332
323
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332324
Between 25 and 24 cal kyr BP the presence of
Mediterranean taxa accompanied by temperate decid-
uous taxa and Abies Cedrus Quercus coccifera and
Pinus suggests a brief climatic amelioration with
increased moisture at least in summer Soon after at
24 cal kyr BP the decline of these taxa and the
increase of Artemisia and Ephedra suggest a climatic
deterioration The above change in pollen assemb-
lages has also been recorded in Ioannina Lake (NW
Greece Galanidou et al 2000) where an amelioration
in the climate at around 25 kyr followed by a short
period of climatic deterioration around 24ndash22 kyr
(H2 event) was observed Furthermore similar
changes for the same time interval have been recorded
in central Greece (Bottema 1974 Tarasov et al
1998 Digerfeldt et al 2000 Karkanas 2001)
At 16 cal kyr BP Gramineae exhibit a slight
decline while Artemisia and Chenopodiaceae
increase suggesting a brief expansion of the semi-
desert This climatic deterioration coincides with the
age of H1 event (~16 cal kyr BP) (Bond et al 1992)
At 13 cal kyr BP the high abundance of Chenopo-
diaceae coincides with the strong enrichment of y18Ovalues (Fig 3) and suggests an increase in aridity
during the Younger Dryas event Expansions of
Chenopodiaceae at the time of the Younger Dryas
event have been documented in all the pollen records
in the eastern Mediterranean region (Rossignol-Strick
1995)
At around 11 cal kyr BP the appearance of Quercus
coccifera and other Mediterranean taxa (Pistacia
Olea and Phyllirea) and soon after at 10 cal kyr BP
the appearance of the temperate deciduous forests
mark the onset of the Holocene The above taxa along
with Quercus Corylus Ulmus Tilia Fagus indicate
a significant available moisture increase with no
drought in summer in the Mediterranean region
Similar trends have been observed by Bottema
(1974) and Rossignol-Strick (1995) in almost all the
pollen records of the eastern Mediterranean region
The populations of the deciduous and Mediterra-
nean taxa decline between 8 and 7 cal kyr BP during
the sapropel interruption between S1a and S1b at
around 8 cal kyr BP whereas an increase of Artemisia
accompanied by the presence of Hedera indicate an
increase in aridity This climatic change is in phase
with the 82 cal kyr BP global event first observed in
Greenland ice cores (Alley et al 1997) Here the
climatic optimum began in the early Holocene at 105
cal kyr BP and ended at 3 cal kyr BP This climatic
optimum resulted in the great expansion of the
temperate deciduous and Mediterranean forests
36 Application of MAT
The downcore variation of SSTc and SSTw based
on the application of the MAT is presented in Fig 7B
According to Overpeck et al (1985) and Kallel et
al (1997) estimations of SSTs by MAT are consid-
ered reliable when the dissimilarity coefficients
(bdistanceQ) is lower than 025 If it is higher than
030 it indicates that the fossil samples in this
interval have no close modern analogue samples
The dissimilarity coefficient (Euclidean) in our data
(Fig 7A) is higher than 025 between 17 and 32 cal
kyr BP indicating no reliable SSTs estimations for
this interval High dissimilarity coefficient around
this time interval has also been recorded in previous
studies in the Mediterranean Sea (Sbaffi et al 2001)
In the present data the downcore variation of the
dissimilarity coefficient seems to be linked to
variations in the abundance of Turborotalita quin-
queloba The correlation coefficient of T quinque-
loba abundance and dissimilarity coefficient is 083
along the core and increases to 093 in the interval
105ndash30 cal kyr BP
At present the SST in the southern Aegean ranges
in winter between 149 and 181 8C and in summer
between 212 and 254 8C (Levitus and Boyer 1994)
The SSTs estimated for the core-top studied sample
(selected at a depth of 5 cm below the surface) are for
winter (SSTc) 193 8C and for summer (SSTw) 228
8C This shows that core-top SSTw is within the
present day range but core-top SSTc is higher by ~15
8C in relation to the present SSTc
During the Holocene the SST is continuously high
with short intervals of rapid fluctuations The mean
SSTc is 195 8C and the mean SSTw is 249 8C Themean difference in temperature between winter and
summer (seasonality) appears slightly decreased (54
8C) in relation to present day (Levitus and Boyer
1994)
During the Holocene from 8 to 7 cal kyr BP and
from 55 to 5 cal kyr BP the SST decreases The mean
annual SST at these intervals compared to the mean
annual SST value for the Holocene (222 8C) is about
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 325
25 8C lower SST appears increased at 95ndash8 cal kyr
BP and 65ndash55 cal kyr BP The mean annual SST
during these intervals compared to the mean annual
SST value for the Holocene (222 8C) is about 4 8Chigher A minor decrease of SSTc of about 1 8C is
observed at 10 cal kyr BP
Between 17 and 105 cal kyr BP the estimated
SSTc and SSTw are lower compared to the Holocene
The mean SSTc is 117 8C and the mean SSTw is 152
8C The mean difference of SSTc between the
abovementioned interval and the Holocene is 78 8Cand that of SSTw is slightly higher (97 8C) Thisreveals a weaker seasonality (358C) between 17 and
10 cal kyr BP than during the Holocene (54 8C)Although absolute values differ Perez-Folgado et al
(2003) have also recorded a decrease in the seasonal
gradient (due to high drops of SSTw) in the Glacial
Period (5 8C) in relation to the Holocene (10 8C) inthe western Mediterranean Sea
Between 47 and 32 cal kyr BP the SST appears
significantly higher compared to the previous interval
(ie 17 to 135 cal kyr BP) The mean SSTc is 135 8Cand the mean SSTw is 194 8C The seasonal gradientis more pronounced in the early part of the interval
between 47 and 42 cal kyr BP (76 8C) than in the latepart (55 8C)
Between 47 and 32 cal kyr BP the SST fluctuates
rapidly for short intervals Reduction of SST occurs
between 41 and 40 cal kyr BP and at 35 cal kyr BP
The mean annual SST drops during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C An increase in SST occurs at around
42 cal kyr BP 39ndash38 cal kyr BP and 35ndash34 cal kyr
BP The mean annual SST rises during these intervals
compared to the mean annual SST for this period
(165 8C) is 1 8C
4 Discussion
41 Stadial and interstadials
The present study of the downcore variation of (i)
SST (ii) the abundance of cold planktonic foramin-
ifera (iii) the y18O values and (iv) the pollen record
reveals a series of short-term palaeoclimatic changes
during the last 48 kyr (Fig 7) In total 10 stadials and
6 interstadials have been identified hereafter referred
to as C69-ST (1ndash10) and C69-IST (1ndash6) respectively
(Fig 7F) One stadial (C69-ST10) and four intersta-
dials (C69-IST1 C69-IST2 C69-IST3 and C69-IST6)
are evident in all four diagrams Two stadials (C69-
ST4 and C69-ST2) are evident in three diagrams Four
stadials (C69-ST5 C69-ST6 C69-ST7 and C69-ST8)
are evident in two diagrams Three stadials (C69-ST1
C69-ST3 and C69-ST9) and two interstadials (C69-
IST4 and C69-IST5) are evident in one diagram The
events that are evident in the three or four downcore
variation diagrams are considered as most pronounced
events The events that are evident in one or two
downcore variation diagrams are considered as least
pronounced events
All these climatic events are comparable with
similar events recognized in the North Atlantic
(Dansgaard et al 1993 Bond et al 1997) and the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) (Fig 7G) The difference in age
of the events occurrence between the present study
and in the abovementioned studies is in the range of a
few hundreds to one millennium of years and may be
attributed to (i) the lack of direct dating of the events
in the present data (ii) microscale changes in the
sedimentation rate in core C69 between the dating
points or (iii) the calibration method of the 14C age
Between 48 and 11 cal yr BP seven stadials (C69-
ST10-4) have been observed (Fig 7F) The C69-ST10
(415ndash40 cal kyr BP) shows a drop in SST and an
increase in y18O values and in semi-desertic vegeta-
tion (Fig 7) This event can be correlated with events
of St11 (~41 cal kyr BP) or St9H4 (40ndash385 cal kyr
BP) (Fig 7G) (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) However the
intensity and the duration of C69-ST10 event encour-
age us to suggest that this event is most probably
correlative with the H4 The C69-ST9 (38ndash37 cal kyr
BP) is characterized only by an increase in aridity
since the y18O values although exhibiting an increas-
ing trend are still low and the SST remains
moderately high This event may be correlated with
the St8 (365ndash355 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
The C69-ST8 (35ndash345 cal kyr BP) is characterized by
a rapid decline in SST and enrichment in y18O values
(Fig 7) This event occurs at times equivalent to St7
(~345 cal kyr BP) (Dansgaard et al 1993 Cacho et
al 1999 Perez-Folgado et al 2003)
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332326
An increase in cold fauna accompanied by enrich-
ment in y18O values characterize the C69-ST7 (29ndash27
cal kyr BP) (Fig 7) This event occurs at the same
time with St4 (29ndash28 cal kyr BP) and appears 2000 yr
later than H3 (31ndash295 cal kyr BP) (Dansgaard et al
1993 Cacho et al 1999 Perez-Folgado et al 2003)
However its intensity as identified by the very low
y18O values and the extremely high abundance of cold
species encourage us to suggest that may corresponds
to H3
The C69-ST6 (24ndash22 cal kyr BP) shows an
increase in cold planktonic fauna and aridity (Fig
7) The y18O values although exhibiting a decreasing
trend are still high (Fig 7C) This event can be
correlated to the H2 (24 cal kyr BP) identified in the
North Atlantic and western Mediterranean Sea (Dans-
gaard et al 1993 Cacho et al 1999 Perez-Folgado
et al 2003) Between 205 and 19 cal kyr BP the
increase of cold fauna accompanied with an enrich-
ment of y18O values and an increase in aridity suggest
the prevalence of Last Glacial Maximum event The
C69-ST5 (165ndash155 cal kyr BP) is characterized by
an increase in cold planktonic fauna at 155 cal kyr BP
and just after and at 165 cal kyr BP an increase in
aridity (Fig 7) The y18O values although exhibiting
a decreasing trend are still high (Fig 7C) This event
can be correlated to the H1 event (16 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-ST4
(135ndash125 cal kyr BP) shows low SST increased
y18O values and an increase in semi-desertic vegeta-
tion (Fig 7) This event corresponds to Younger
Dryas event which has also been observed in the
western Mediterranean Sea (Cacho et al 1999 Perez-
Folgado et al 2003) Climatic deterioration during
the Younger Dryas with a decrease in temperature
and an increase in aridity has been documented all
over the eastern Mediterranean in the marine and
terrestrial records (Rossignol-Strick 1995)
The intervals between the abovementioned stadials
are characterized by milder climatic conditions
However within four of them the signal is enhanced
(C69-IST3ndash6) The C69-IST6 (395ndash385 cal kyr BP)
shows a strong depletion in y18O values increased
SST a decline in semi-desertic taxa and an increase of
temperate trees This event coincides with the
formation of S2 If we assume that the C69-ST10
correlates with the H4 event then the C69-IST6 may
be correlated with the Ist8 (~38ndash37 cal kyr BP)
identified in the North Atlantic and western Medi-
terranean Sea (Dansgaard et al 1993 Cacho et al
1999 Perez-Folgado et al 2003) The C69-IST5 (37ndash
35 cal kyr BP) shows an increase of temperate taxa in
the pollen record The SST is moderate and increases
rapidly at the end of the event but the y18O values are
high The C69-IST4 (34ndash33 cal kyr BP) shows only a
depletion in the y18O values The appearance of
Mediterranean and temperate tree taxa characterizes
the C69-IST3 (25ndash24 cal kyr BP) During this event
the y18O values exhibit a decreasing trend and the
abundance of cold fauna also decreases This event
can be correlated with the Ist3 (Dansgaard et al 1993
Cacho et al 1999 Perez-Folgado et al 2003)
The microfauna associations identified in the
present data during the Heinrich events differ from
those recorded in the western Mediterranean at the
same intervals (Fig 4) (Perez-Folgado et al 2003)
The planktonic assemblages during the H1ndashH4 in the
western Mediterranean display sharp increases of
Neogloboquadrina pachyderma (s) Turborotalita
quinqueloba Globorotalia scitula and Globigerina
bulloides while Globorotalia inflata Globigerinoides
ruber and N pachyderma (d) tend to decline or
disappear (Perez-Folgado et al 2003) In the Cretan
Basin T quinqueloba and G scitula dominate the
microfauna during the H1ndashH4 but without forming
peaks as strong as those present in the western
Mediterranean data The participation of N pachy-
derma (d) and G inflata in the microfauna assemb-
lages is high though appear to decline during the
Heinrich events G bulloides does not exhibit any
specific trend and N pachyderma (s) is almost absent
The presence of Turborotalita quinqueloba is in
general higher between 48 and 11 kyr BP in the
southern Aegean Sea than in the central Mediterra-
nean (Capotondi et al 1999 Sbaffi et al 2001) It
seems that this cool species replaces other cold
species such as Globorotalia scitula and Neoglobo-
quadrina pachyderma (s) The abundance of G
scitula in the planktonic assemblages appears to
decrease eastwards in the Mediterranean Sea during
the early-Late Glacial period while N pachyderma (s)
is almost absent in the eastern Mediterranean (Hayes
et al 1999) Today N pachyderma (s) thrives in
seawater where SST is below 7 8C (Reynolds and
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 327
Thunell 1986) This may indicate that the cooling
during the stadials was not as intense in the eastern
Mediterranean as in the western Mediterranean The
MAT SST seems to support this hypothesis as it
shows that during H4 the SSTc was 9 8C while in thesame interval in the western Mediterranean it was 7
8C (Perez-Folgado et al 2003)
Increases in aridity attributed to Heinrich events
have been recorded in Greece in speleothems at cave
sites (Theopetra Cave Central Greece Karkanas
2001) and in terrestrial pollen records in Greece
(Galanidou et al 2000 Tzedakis et al 2002) Studies
in lake Xinias (central Greece) have shown lake-level
lowering at around 41ndash40 cal kyr BP and 32ndash30 cal
kyr BP (Digerfeldt et al 2000) The former interval
coincides with the C69-ST10H4 while the latter
appear about 2 kyr earlier than C69-ST7H3 In
addition oscillations of short duration between open
and forest communities during the Glacial Period have
been recognized in terrestrial pollen records from
Greece (Wijmstra 1969 Bottema 1974 Van Andel
and Tzedakis 1996 Tzedakis et al 2002) suggesting
climatic changes of short duration on land Stable
isotopic profiles of speleothems in Soreq cave in
Israel suggest the presence of Younger Dryas (132ndash
114 cal kyr BP) H1 (165 cal kyr BP) LGM (21ndash19
cal kyr BP) H2 (25 cal kyr BP) and H5 (46 cal kyr
BP) while H3 and H4 are not reflected in the record
(Bar-Matthews et al 1999) In the same study a
warm interval at 36 cal kyr BP probably correlates
with the C69-IST6 At the Lake Lisan (Israel) water
level drops have been documented at the times of H1ndash
H5 where the signal of H4 is very strong (Bartov et
al 2003)
The cooling of the sea surface temperature during
the Heinrich events in the southern Aegean Sea could
be a direct result of climatic deterioration in the
eastern Mediterranean region or an indirect effect of
the cold Atlantic surficial water flow The increase in
aridity shown in the pollen records during the stadials
suggests synchronicity between terrestrial and marine
data in the south Aegean region in response to short-
term climatic changes (Figs 6 and 7EF) Similar
suggestions have been made for marine pollen records
from the western Mediterranean region (Sanchez Goni
et al 2002) where during stadials the pollen zones are
characterized by the dominance of steppic plants
(Artemisia Chenopodiaceae and Ephedra) and during
the interstadials by the dominance of Quercus and the
reduction of the steppes
Climatic variability is also recorded during the
Holocene where three stadials (C69-ST1ndash3) have
occurred The C69-ST3 (10 cal kyr BP) is charac-
terized by a brief increase in y18O values and a brief
reduction in the abundance of the warm species
Globigerinoides ruber (Fig 4) A brief cooling event
around the same age (103 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) Although the SST is rather high
in this interval it shows a slight drop during winter
The C69-ST2 (8ndash65 cal kyr BP) is the most
prominent Holocene stadial and is characterized by a
rapid increase of semi-desertic vegetation and a
decline in SST During this event the y18O values
remain low The C69-ST2 seems to correspond to the
most widespread Holocene cold event of Europe
which occurred at around 82 cal kyr BP (Alley et al
1997) This event has been also identified in the UkV37
SST records in the western (Cacho et al 1999) and
the central Mediterranean Sea (Sbaffi et al 2001)
The C69-ST1 (55 cal kyr BP) is characterized by a
brief drop in SST During this interval the y18O values
show an increasing trend A brief cooling event
around the same age (54 cal kyr BP) has been
identified in the UkV37 SST records in the western
(Cacho et al 1999) and the central Mediterranean Sea
(Sbaffi et al 2001) The C69-ST2 and C69-ST1
events coincide with the interruption and the end of
the sapropel S1 respectively
All the abovementioned Holocene events in the
Cretan Basin are related to Holocene stadials docu-
mented in cores from high latitudes (Bond et al
1997) The C69-ST1 and C69-ST2 are the most
widespread Holocene events in the marine sediments
of the eastern Mediterranean Sea correlated with the
interruption and the final end of S1 (Rohling et al
1997 De Rijk et al 1999)
Two interstadials have been observed in the
Holocene C69-IST1 (65ndash6 cal kyr BP) and C69-
IST2 (9ndash8 cal kyr BP) These events which are
characterized by increased SSTs strong depletions in
y18O values and an expansion of temperate evergreen
and Mediterranean taxa coincide with the deposition
of the sapropelic layers S1a and S1b An increase in
humidity around this time has been documented in all
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332328
marine and terrestrial pollen records in the eastern
Mediterranean region (Rossignol-Strick 1995)
The abovementioned stadials and interstadials
coincide well with the climatic cycles of dry and
wet events observed in the coastal belt of Israel in
combination with the level fluctuations of the Dead
Sea and Lake Lisan (Gvirtzman and Wieder 2001)
The dry events E5 (65ndash50 cal kyr BP) E7 (75ndash7 cal
kyr BP) and E9 (105ndash10 cal kyr BP) of the
abovementioned study coincide rather well with the
C69-ST1ndash3 in the present study and the wet events E6
(7ndash65 cal kyr BP) and E8 (10ndash75 cal kyr BP)
coincide rather well with the C69-IST1ndash2 in the
present data The three Holocene stadials and the two
interstadials which coincide with the formation of S1a
and S1b have also been recorded in the sediments of
C40 from the Myrtoon Basin based on the fluctua-
tions of the planktonic and pollen assemblages
(Geraga et al 2000)
42 Sapropel S1 and sapropelic S2
The sapropel S1 (S1a and S1b) and the sapropelic
S2 have shown a few common characteristics The
formation of both sapropels coincides with intersta-
dials S1a and S1b with the interstadials in Holocene
(C69-IST1 and C69-IST2 respectively) and S2 with
(C69-IST6) which probably corresponds to Ist8
(Dansgaard et al 1993 Cacho et al 2002) In both
sapropels the high depletions in y18O values (Fig 3)
do not seem to be accounted for only the increases in
the SSTs but seem also to be associated with the
prevalence of a low salinity surface layer (Vergnaud-
Grazzini et al 1977 Rossignol-Strick et al 1982)
During the formation of S1 the major sources of the
low salinity surface layer are considered to be the
southern Mediterranean catchment and the fresh water
discharges from the Black Sea to the Aegean Sea
(Aksu et al 1999 Sperling et al 2003) The ages of
S1 and S2 almost coincide with peak values of the
moonsonal index indicating increased precipitation
although the moonsonal index is greater during S1
(42) than during S2 (19 at 33 14C kyr BP) (Rossignol-
Strick 1983 Rossignol-Strick and Paterne 1999)
The high level of Lake Xinias in Greece (Digerfeldt et
al 2000) and the increase in temperate deciduous
trees in the pollen records of the present study during
the formation of S1 and S2 (Fig 6) indicate an
increase in humidity over the region Furthermore
regarding the formation of S2 an increase in fresh
water discharges from the Black Sea to the Aegean
Sea could have taken place due to the retreat of the
Scandinavian ice sheet during the Alesund period
(385ndash325 14C kyr) (Baumann et al 1995)
Differences between S1 (S1a and S1b) and S2
occur in the microfauna assemblages (Figs 4 and 5)
During the formation of S1 the planktonic foramini-
feral assemblages dominated by the Globigerinoides
ruber together with other warm-water species indicate
the presence of warm surficial waters During S2 the
G ruber hardly increased while cool species such as
Globorotalia scitula are present indicating that the
SST was lower than during S1 The planktonic
assemblage is dominated by Neogloboquadrina
which indicates the development of a DCM (Deep
Chlorophyll Maximum) layer (Fairbanks and Wiebe
1980) Furthermore the presence of infauna benthic
foraminifera in core C69 (unpublished data) during S2
indicates the development of dysoxic bottom waters
conditions whereas the absence of benthic foramin-
ifera in core C69 during the S1 indicates the
prevalence of anoxic bottom waters
The abovementioned suggests that during S2 the
prevalence of lower salinity surficial waters at the
Levantine region where the LIW (Levantine Inter-
mediate Water) is forming reduced the density
gradient between the Surficial Atlantic Water and
Intermediate Water masses and caused (i) a decline in
the water mass vertical circulation and (ii) a shoaling
of the pycnocline in the euphotic zone (Rohling and
Gieskes 1989) The latter resulted in the development
of the DCM layer and caused an increase in primary
productivity as is suggested by the increase of
Neogloboquadrina The water mass circulation
although reduced never ceased as is indicated by
the presence of benthic fauna and the existence of
oxygen in the bottom waters Increases in organic
fluxes and reductions in oxygen supply increased the
preservation of the organic material on the sea floor
resulting in the formation of the sapropel S2 The
above mechanism has already been proposed for the
formation of the sapropel layers during the Glacial
Period (Rohling and Gieskes 1989)
The formation of S1 is a combination of changed
water mass circulation and increased productivity At
that time the establishment of warm and low salinity
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 329
surficial waters due to increased rainfall in the
surrounding area and the consequently freshwater
run-off resulted in an enhanced stratification of the
water column (Vergnaud-Grazzini et al 1977 Rossi-
gnol-Strick et al 1982 Rohling and Gieskes 1989
Aksu et al 1999) The increased productivity during
the formation of S1 is related to the increased nutrient
discharge of the Nile (Rossignol-Strick et al 1982
Aksu et al 1995) although in varying intensity
(Meier et al 2004) Furthermore early studies based
on calcareous nanofossils and chemical records
suggest the presence of a DCM layer although in a
varying duration (Principato et al 2003 Meier et al
2004) Brief deep water ventilation events are
associated with the interruption in the formation of
S1 (Casford et al 2002) In the present study the
planktonic assemblages suggest an enhanced stratifi-
cation of the water column during the deposition of
S1a and S1b and a brief water ventilation during the
interruption of S1 A shoaling of the pycnocline can
be assumed during S1b due to the increase in the
abundance of the Globigerinoides sacculifer (Princi-
pato et al 2003) while the presence of Globorotalia
inflata just after the end of S1 mark the return of the
deep water ventilation
5 Conclusions
High resolution micropaleontological and pollen
analyses in combination with y18O data in the
sediments of core C69 from the southern Aegean
Sea showed that the study area is characterized by
climatic variability at millennium scale over the last
48 kyr The climate variations seem to correspond to
cycles of stadialndashinderstadials attributed to Dans-
gaardndashOeschger events although some of them differ
slight in age
Four of the recognized stadials in the Glacial period
seem to correspond to the Younger Dryas and the
Heinrich events H1 H2 and H4 One stadial recog-
nized in the Holocene seems to correspond to the most
widespread stadial in Holocene at 8 cal kyr BP In most
of the stadials the isotopes planktonic and pollen data
show synchronicity suggesting an additional opera-
tion of atmospheric processes The climatic signal
based on planktonic assemblages appears attenuated in
comparison to the western Mediterranean during the
stadials which are correlated with the Heinrich events
in the present study During these events the planktonic
fauna is dominated by Turborotalita quinqueloba and
Globorotalia scitula
The sapropel S1 appears in two layers (S1a and
S1b) Their formations coincide with the prevalence
of interstadials The interruption and the end of S1
coincide with stadials A sapropelitic sequence
corresponding to S2 occurs between 395 and 385
cal kyr BP and coincides with the prevalence of an
interstadial
Acknowledgments
We wish to thank the anonymous reviewers and
Prof MB Cita for their suggested improvements to
the manuscript This study was funded by the Hellenic
Scholarship Foundation (IKY)
References
Aksu AE Yasar D Mudie PJ 1995 Paleoclimatic and
paleoceanographic conditions leading to development of sapro-
pel layer S1 in the Aegean Sea Palaeogeography Palae-
oclimatology Palaeoecology 116 71ndash101
Aksu AE Hiscott RN Yasar D 1999 Oscillating Quaternary
water levels of the Marmara Sea and vigorous outflow into the
Aegean Sea from the Marmara SeandashBlack Sea drainage corridor
Marine Geology 153 275ndash302
Alley RB Mayewski PA Sowers T Stuiver M Taylor KC
Clark PU 1997 Holocene climatic instability a prominent
widespread event 8200 yr ago Geology 25 483ndash486
Anastasakis GC Stanley DJ 1984 Sapropels and organic-rich
variants in the Mediterranean sequence development and
classification In Stow DAV Piper DJW (Eds) Fine
Grained Sediments Deep-Water Processes and Facies Geo-
logical Society Special Publication vol 15 pp 497ndash510
Bard E Rostek F Menot-Combes G 2004 Radiocarbon
calibration 20000 14C yr BP by means of planktonic
foraminifera of the Iberian Margin Quaternary Research 61
204ndash214
Bar-Matthews M Ayalon A Kaufman A Wasserbourg GJ
1999 The eastern Mediterranean palaeoclimate as a reflection of
regional events Soreq Cave Israel Earth Planet Science Letters
166 85ndash95
Bartov Y Goldstein SL Stein M Enzel Y 2003 Catastrophic
arid episodes in the Eastern Mediterranean linked with the North
Atlantic Heinrich events Geology 31 439ndash442
Baumann K-H Lachschewitz KS Mangerud J Spielhagen
RF Wolf-Welling TCW Henrich R Kassens H 1995
Reflection of Scandinavian Ice Sheet Fluctuations in Norwegian
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332330
Sea sediments during the past 150000 years Quaternary
Research 43 185ndash197
Bond GC Lotti R 1995 Iceberg discharges into the North
Atlantic on millennial time scales during the last glaciation
Science 267 1005ndash1010
Bond G Heinrich H Broecker W Labeyrie L McManus J
Andrews J Huon S Jantschik R Clasen S Simet C
Tedesco K Klas M Bonani G Ivy S 1992 Evidence for
massive discharges of icebergs into the North Atlantic Ocean
during the last glacial period Nature 360 245ndash249
Bond G Showers W Cheseby M Lotti R Almasi P de
Menocal P Priore P Cullen H Hadjas I Bonani G 1997
A pervasive millennial scale cycle in North Atlantic Holocene
and glacial climates Science 278 1257ndash1266
Bottema S 1974 Late Quaternary vegetation History of North-
western Greece PhD thesis Rijksuniversiteit te Groningen
Buckley HA Johnson LR 1988 Late Pleistocene to Recent
sediment deposition in the central and Western Mediterranean
Deep-Sea Research 35 (5) 749ndash766
Buckley HA Johnson LR Shackleton NJ Blow RA 1982
Late glacial to recent cores from the eastern Mediterranean
Deep-Sea Research 29 739ndash766
Cacho I Grimalt JO Pelejero C Canals M Sierro FJ
Flores JA Shackleton N 1999 DansgaardndashOeschger and
Heinrich event imprints in Alboran Sea paleotemperatures
Paleoceanography 14 698ndash705
Cacho I Grimalt JO Sierro FJ Shackleton N Canals M
2000 Evidence for enhanced Mediterranean thermohaline
circulation during rapid climatic coolings Earth and Planetary
Science Letters 183 417ndash429
Cacho I Grimalt JO Canals M 2002 Response of the Western
Mediterranean Sea to rapid climatic variability during the last
50000 years a molecular biomarker approach Journal of
Marine Systems 33ndash34 253ndash272
Capotondi L Borsetti AM Morigi C 1999 Foraminiferal
ecozones a high resolution proxy for the Late Quaternary
biochronology in the central Mediterranean Sea Marine
Geology 153 253ndash274
Casford JSL Rohling EJ Abu-Zied R Cooke S Fontanier
C Leng M Lykousis V 2002 Circulation changes and
nutrient concentrations in the late Quaternary Aegean Sea a
nonsteady state concept for sapropel formation Paleoceanog-
raphy 17 (2) 1024ndash1034
Cita MB Vergnaud-Grazzini C Robert C Chamley H
Ciaranfi N drsquoOnofrio S 1977 Paleoclimatic record of a
long deep sea core from the eastern Mediterranean Quaternary
Research 8 205ndash235
Dansgaard W Johnsen SJ Clausen HB Dahl-Jensen D
Gundestrup NS Hammer CU Hvidberg CS Steffensen
JP Sveinbjornsdottir AE Jouzel J Bond G 1993
Evidence of general instability of past climate from a 250 kyr
ice core record Nature 364 218ndash220
De Rijk S Hayes A Rohling EJ 1999 Eastern Mediterranean
sapropel S1 interruption an expression of the onset of climatic
deterioration around 7 ka BP Marine Geology 153 337ndash343
Digerfeldt G Olsson S Sandgren P 2000 Reconstruction of
lake-level changes in lake Xinias central Greece during the last
40000 years Palaeogeography Palaeoclimatology Palaeoecol-
ogy 158 65ndash82
Fairbanks RG Wiebe PH 1980 Foraminifera and Chlorophyll
Maximum vertical distribution seasonal succession and
paleoceanographic significance Science 209 1524ndash1526
Fontugne MR Arnold M Labeyrie L Paterne M Calvert
SE Duplessy JC 1994 Paleoenvironment sapropel chro-
nology and Nile River discharge during the last 20000 yr as
indicated by deep-sediment records in the Eastern Mediterra-
nean In Bar-Yosef O Kra RS (Eds) Late Quaternary
Chronology and Paleoclimates of the Eastern Mediterranean
Radiocarbon Arizona Board of Regents University of Arizona
pp 75ndash88
Galanidou N Tzedakis PC Lawson IT Frogley MR 2000
A revised chronological and paleoenvironmental framework
for the Kastritsa rockshelter northwest Greece Antiquity 74
349ndash355
Gaudette H Flight W Toner L Folger D 1974 An
inexpensive titration method for the determination of organic
carbon in recent sediments Journal Sedimentary Petrology 44
249ndash253
Geraga M Tsaila-Monopoli St Ioakim Ch Papatheodorou G
Ferentinos G 2000 An evaluation of paleoenvironmental
changes during the last 18000 yrs BP in the Myrtoon Basin
SW Aegean Sea Palaeogeography Palaeoclimatology Palae-
oecology 156 1ndash17
Giunta S Negri A Morigi C Capotondi L Combourieu-
Nebout N Emeis KC Sangiorgi F Vigliotti L 2003
Coccolithophorid ecostratigraphy and multi-proxy paleoceano-
graphic reconstruction in the Southern Adriatic Sea during the
last deglacial time (Core AD91-17) Palaeogeography Palae-
oclimatology Palaeoecology 190 39ndash59
Gvirtzman G Wieder M 2001 Climate of the last 53000 years
in the eastern Mediterranean based on soil-sequence stratig-
raphy in the coastal plain of Israel Quaternary Science Reviews
20 1827ndash1849
Hayes A Rohling EJ De Rijk S Kroon D Zachariasse WJ
1999 Mediterranean planktic foraminiferal faunas during the
last glacial cycle Marine Geology 153 239ndash252
Hutson WH 1980 The Agulhas Current during the Late
Pleistocene analysis of modern fauna analogs Science 207
64ndash66
Jorissen FJ Asioli A Borsetti AM Capotondi L de Visser
JP Hilgen FJ Rohling EJ van der Borg K Vergnaud
Grazzini C Zachariasse W 1993 Late Quaternary central
Mediterranean biochronology Marine Micropaleontology 21
169ndash189
Kallel N Paterne M Duplessy JC Vergnaud-Grazzini C
Pujol C Labeyrie L Arnold M Fontugne M Pierre C
1997 Enhanced rainfall in the Mediterranean region during the
last sapropel event Oceanological Acta 20 697ndash712
Karkanas P 2001 Site formation processes in Theopetra Cave a
record of climatic change during the Late Pleistocene and Early
Holocene in Thessaly Greece Geoarchaeology 16 373ndash399
Kidd RB Cita MB Ryan WBF 1978 Stratigraphy of eastern
Mediterranean sapropel sequences recovered during DSDP LEG
42A and their paleoenvironmental significance In Hsu KJ
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332 331
Mondrader L et al (Eds) Initial Reports of the Deep Sea
Drilling Project US Government Printing Office Washington
DC pp 421ndash443
Levitus S Boyer T 1994 World ocean atlas 1994 volume 4
temperature NOAA Atlas NESDIS vol 4 US Department of
Commerce Washington DC
Lourens LJ Hilgen FJ Gudjonsson L Zachariasse WJ 1994
Late Pliocene to Early Pleistocene astronomically forced sea
surface productivity and temperature variations in the Medi-
terranean In Lourens LJ (Ed) Astronomical Forcing of
Mediterranean Climate During the Last 53 Million Years
Universiteit Utrecht pp 37ndash58
Martinson DG Pisias NG Hays JD Imbrie J Moore Jr
TC Shackleton NJ 1987 Age dating and the orbital theory
of the Ice Ages development of a high-resolution 0 to 300000-
year chronostratigraphy Quaternary Research 27 1ndash29
Meier KJS Zonneveld KAF Kasten S Willems H 2004
Different nutrient sources forcing increased productivity during
eastern Mediterranean S1 sapropel formation as reflected by
calcareous dinoflagellate cysts Paleoceanography 19 1ndash12
(PA 1012)
Overpeck JT Webb III T Prentice IC 1985 Quantitative
interpretation of fossil pollen spectra dissimilarity coefficients
and the method of modern analogs for pollen data Quaternary
Research 23 87ndash108
Perez-Folgado M Sierro FJ Flores JA Cacho I Grimalt
JO Zahn R Shackleton N 2003 Western Mediterranean
planktonic foraminifera events and millenial climatic variability
during the last 70 kyr Marine Micropaleontology 48 49ndash70
Peyron O Guiot J Cheddadi R Tarasov P Reille M de
Beaulieu JL Bottema S Andrieu V 1998 Climatic
reconstruction in Europe for 18000 years BP from pollen data
Quaternary Research 49 183ndash196
Principato MS Giunta S Corselli C Negri A 2003 Late
PleistoceneHolocene planktic assemblages in three box-cores
from the Mediterranean Ridge area (WndashSW of Crete) paleo-
ecological and paleoceanographic reconstruction of sapropel S1
interval Palaeogeography Palaeoclimatology Palaeoecology
190 61ndash77
Pujol C Vergnaud Grazzini C 1995 Distribution of live planktic
forminifers as related to regional hydrography and productive
systems of the Mediterranean Sea Marine Micropaleontology
25 187ndash217
Ramsey CB Manning SW Galimberti M 2004 Dating the
volcanic eruption at Thera Radiocarbon 46 (1) 325ndash344
Reimer P 2000 Marine reservoir correction database in http
radiocarbonpaqubacukmarine
Reynolds LA Thunell RC 1986 Seasonal production and
morphologic variation of Neogloboquadrina pachyderma
(Ehrenberg) in the northeast Pacific Micropaleontology 32
1ndash18
Rohling EJ Gieskes WWC 1989 Late Quaternary changes in
Mediterranean intermediate water density and formation
Paleoceanography 4 531ndash545
Rohling EJ Jorissen FJ De Stigter HC 1997 200 year
interruption of Holocene sapropel formation in the Adriatic Sea
Journal of Micropalaeontology 16 97ndash108
Rossignol-Strick M 1983 African monsoon an immediate
response to orbital insolation Nature 303 46ndash49
Rossignol-Strick M 1995 Seandashland correlation of pollen records
in the Eastern Mediterranean for the GlacialndashInterglacial
transition biostratigraphy versus radiometric time-scale Qua-
ternary Science Reviews 14 893ndash915
Rossignol-Strick M Paterne M 1999 Synthetic pollen record of
the eastern Mediterranean sapropels of the last 1 Ma
implications for the timescale and formation of the sapropels
Marine Geology 153 221ndash237
Rossignol-Strick M Nesteroff WD Olive P Vergnaud-Graz-
zini C 1982 After the deluge Mediterranean stagnation and
sapropel formation Nature 295 105ndash110
Rossignol-Strick M Planchais N Paterne M Duzer D 1992
Vegetation dynamics and climate during deglaciation in the
south Adriatic basin from a marine record Quaternary Science
Reviews 11 415ndash425
Ryan WBF 1972 Stratigraphy of late Quaternary sediments in the
Eastern Mediterranean In Stanley DJ (Ed) Mediterranean
Sea A Natural Sedimentation Laboratory Dowden Hutchinson
and Ross Inc Stroudsburg Pennsylvania pp 149ndash169
Sanchez Goni MF Cacho I Turon JL Guiot J Sierro FJ
Peypouquet JP Grimalt JO Shackelton NJ 2002 Syn-
chroneity between marine and terrestrial responses to millennial
scale climatic variability during the last glacial period in the
Mediterranean region Climate Dynamics 19 95ndash105
Sbaffi L Wezel FC Kallel N Paterne M Cacho I Ziveri P
Shackleton N 2001 Response of the pelagic environment to
palaeoclimatic changes in the central Mediterranean Sea during
the Late Quaternary Marine Geology 178 39ndash62
Siani G Paterne M Michel E Sulpizio R Sbrana A Arnold
M Haddad G 2001 Mediterranean Sea surface radiocarbon
reservoir age changes since the Last Glacial Maximum Science
294 1917ndash1920
Sperling M Schmiedl G Hemleben Ch Emeis KC Erlen-
keuser H Grootes PM 2003 Black Sea impact on the
formation of eastern Mediterranean sapropel S1 Evidence from
the Marmara Sea Palaeogeography Palaeoclimatology Palae-
oecology 190 9ndash21
Stuiver M Reimer PJ 1993 Extended 14C database and revised
CALIB radiocarbon program Radiocarbon 35 215ndash230
Tarasov PE Cheddadi R Guiot J Bottema S Peyron O
Belmonte J Ruiz-Sanchez V Saadi F Brewer S 1998 A
method to determine warm and cool steppe biomes from pollen
data application to the Mediterranean and Kazakhstan regions
Journal of Quaternary Science 13 (4) 335ndash344
Thunell RC 1978 Distribution of recent planktonic foraminifera
in surface sediments of the Mediterranean Sea Marine Micro-
paleontology 3 147ndash173
Thunell RC Williams DF 1989 GlacialndashHolocene salinity
changes in the Mediterranean Sea hydrographic and deposi-
tional effects Nature 338 493ndash496
Tzedakis PC Lawson IT Frogley MR Hewitt G Preece R
2002 Buffered tree population changes in a Quaternary
refugium evolutionary implications Science 297 2044ndash2047
Van Andel T Lianos N 1984 High resolution seismic reflection
profiles for the reconstruction of post-glacial transgressive
M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254

M Geraga et al Palaeogeography Palaeoclimatology Palaeoecology 220 (2005) 311ndash332332
shorelines An example from Greece Quaternary Research 22
31ndash45
Van Andel TH Tzedakis P 1996 Palaeolithic landscapes of
Europe and environs 150000ndash25000 years ago an overview
Quaternary Science Review 15 481ndash500
Vergnaud-Grazzini C Ryan WBF Cita MB 1977 Stable
isotopic fractionation climate change and episodic stagnation in
the eastern Mediterranean during the Late Quaternary Marine
Micropaleontology 2 353ndash370
Vergnaud-Grazzini C Devaux M Znaidi J 1986 Stable isotope
banomaliesQ in Mediterranean Pleistocene records Marine
Micropaleontology 10 35ndash69
Walter W 1974 Die Vegetation Ost-Europas Nord-und Zentrala-
siens Gustav Fischer Verlag Stuttgart 452 pp
Wijmstra TA 1969 Palynology of the first 30 meters of a 120 m
deep section in Northern Greece Acta Botanica Neerlandica 18
511ndash527
Williams DF Thunell RC Kennett JP 1978 Periodic fresh-
water flooding and stagnation of the Eastern Mediterranean Sea
during the Late Quaternary Science 201 252ndash254
Copyright © 2022 FDOKUMEN




















