Sex-specific microRNAome deregulation in the shielded bystander spleen of cranially exposed mice
10
©2008 LANDES BIOSCIENCE. DO NOT DISTRIBUTE. [Cell Cycle 7:11, 1658-1667; 1 June 2008]; ©2008 Landes Bioscience 1658 Cell Cycle 2008; Vol. 7 Issue 11 The bystander effect is a phenomenon that occurs when exposed cells signal distress to their naïve, unexposed neighbors. It is now accepted as a ubiquitous consequence of radiation exposure. It is well documented to occur in cultured cells, 3D tissue models, and in organs and organisms. Notwithstanding, the exact mechanisms of the bystander effect remain unclear. Recent studies hinted that bystander effects may, in part, be distinct in males and females, and possibly mediated via short non-coding RNAs, specifically, microRNAs. MicroRNAs are small, abundant and capable of regu- lating the expression of a wide variety of targets. Yet, their roles in bystander effects have not been analyzed in detail. The mechanisms behind sex differences observed in in vivo bystander effects also remain to be uncovered. We hypothesized that the radiation-induced expression of microRNAs in exposed and bystander tissue may be distinct in males and females. Using a well-establish bystander mouse model when the animal’s head is exposed, while the body is completely protected by a medical-grade shield, we have for the first time shown that radiation exposure triggers a significant and sex-specific deregula- tion of the microRNAome in the non-exposed bystander spleen. The altered miRNA levels were paralleled by sex-specific changes in the levels of the miRNA processing enzyme Dicer and compo- nents of the RNA-induced silencing complex (RISC). Sterilization of animals resulted in drastic microRNAome alterations and significantly affected radiation and bystander miRNA responses. Our data may provide a roadmap for further analysis of the role of microRNAome in genotoxic stress responses and may help us explain sex specificity of radiation-induced carcinogenesis. Introduction Ionizing radiation (IR) is an invaluable diagnostic and treat- ment tool. On other hand, it is a cytotoxic agent and a potential carcinogen. 1 Over the course of years, the central dogma in radiation biology stated that IR effects were restricted to directly exposed cells. However, it was shown that IR effects can also be seen in naïve cells that were in contact with directly irradiated cells, or in naïve cells that received certain irradiation ‘distress’ signals from directly exposed cells via growth medium. 2-11 Such an exposure communication process is termed a bystander effect, whereby radiation impact is localized not only in the area of exposure, but it can be extended into the tissue, 2,3 organ, 4-7 organism, 8-10 and even to other living organisms. 11 Recent studies report a number of alterations taking place in bystander cells and tissues, including induction of DNA strand breaks, 2,3,8,12,13 altered gene expression, 14 cell cycle arrest, apoptosis, 2,3,8,15,16 and a number of epigenetic events 3,9,10,17 resulting in genomic insta- bility. 2,3,18-21 Yet, the precise mechanisms of bystander effects and the nature of bystander signaling remain unclear. However, a newly emerged mechanism of epigenetic control mediated through the involvement of small regulatory RNAs 22,23 promises to bridge the gaps in our understanding of a bystander phenomenon. 9 There are a number of functional classifications of small regu- latory RNAs, and among them, microRNAs (miRNAs) are of a particular interest. MiRNAs are evolutionally conserved, small, single-stranded, non-protein-coding RNA molecules which are presently recognized as major regulators of gene expression. 22-24 MiRNAs are initially expressed as primary miRNAs (pri-miRNAs), which form a hairpin containing the mature miRNA sequence. This hairpin is released in the nucleus by a dsRNA-specific ribo- nuclease, Drosha, to form a precursor miRNA (pre-miRNA). 25-27 The pre-miRNAs are then exported to the cytoplasm, where Dicer, a member of the RNAse III superfamily, cleaves the pre-miRNA leading to the formation of a mature miRNA. 25-27 To regulate target mRNAs expression, mature miRNAs must associate with the RNA- induced silencing complex (RISC) proteins 27 such as Argonaute (Ago), 28 PACT (a protein activator of the interferon-induced protein *Correspondence to: Olga Kovalchuk; Department of Biological Sciences; University of Lethbridge; 4401 University Drive; Lethbridge, Alberta T1K3M4 Canada; Fax: 403.329.2242; Email: [email protected] Submitted: 02/13/08; Accepted: 03/23/08 Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/article/5981 Report Sex-specific microRNAome deregulation in the shielded bystander spleen of cranially exposed mice Igor Koturbash, 1,† Franz J. Zemp, 1,† Kristy Kutanzi, 1 Lidiya Luzhna, 1 Jonathan Loree, 1 Bryan Kolb 2 and Olga Kovalchuk 1, * 1 Department of Biological Sciences; University of Lethbridge; Alberta, Canada; 2 Department of Neuroscience; University of Lethbridge; Alberta, Canada † These authors contributed equally to this work. Abbreviations: RISC, RNA-induced silencing complex; IR, Ionizing radiation; pri-miRNAs, primary miRNAs; pre-miRNA, precursor miRNA; PACT, a protein activator of the interferon-induced protein kinase, PKR; qRT-PCR, quantitative real-time polymerase chain reac- tion; CAST, castration; OVX, ovariectomization; H&E, hematoxylin and eosin; TMAs, tissue microarrays Key words: bystander effect, microRNA, DICER, RISC, sex differences
Transcript of Sex-specific microRNAome deregulation in the shielded bystander spleen of cranially exposed mice

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
[Cell Cycle 7:11, 1658-1667; 1 June 2008]; ©2008 Landes Bioscience
1658 Cell Cycle 2008; Vol. 7 Issue 11
The bystander effect is a phenomenon that occurs when exposed cells signal distress to their naïve, unexposed neighbors. It is now accepted as a ubiquitous consequence of radiation exposure. It is well documented to occur in cultured cells, 3D tissue models, and in organs and organisms. Notwithstanding, the exact mechanisms of the bystander effect remain unclear. Recent studies hinted that bystander effects may, in part, be distinct in males and females, and possibly mediated via short non-coding RNAs, specifically, microRNAs. MicroRNAs are small, abundant and capable of regu-lating the expression of a wide variety of targets. Yet, their roles in bystander effects have not been analyzed in detail. The mechanisms behind sex differences observed in in vivo bystander effects also remain to be uncovered.
We hypothesized that the radiation-induced expression of microRNAs in exposed and bystander tissue may be distinct in males and females.
Using a well-establish bystander mouse model when the animal’s head is exposed, while the body is completely protected by a medical-grade shield, we have for the first time shown that radiation exposure triggers a significant and sex-specific deregula-tion of the microRNAome in the non-exposed bystander spleen. The altered miRNA levels were paralleled by sex-specific changes in the levels of the miRNA processing enzyme Dicer and compo-nents of the RNA-induced silencing complex (RISC). Sterilization of animals resulted in drastic microRNAome alterations and significantly affected radiation and bystander miRNA responses. Our data may provide a roadmap for further analysis of the role of microRNAome in genotoxic stress responses and may help us explain sex specificity of radiation-induced carcinogenesis.
Introduction
Ionizing radiation (IR) is an invaluable diagnostic and treat-ment tool. On other hand, it is a cytotoxic agent and a potential carcinogen.1 Over the course of years, the central dogma in radiation biology stated that IR effects were restricted to directly exposed cells. However, it was shown that IR effects can also be seen in naïve cells that were in contact with directly irradiated cells, or in naïve cells that received certain irradiation ‘distress’ signals from directly exposed cells via growth medium.2-11 Such an exposure communication process is termed a bystander effect, whereby radiation impact is localized not only in the area of exposure, but it can be extended into the tissue,2,3 organ,4-7 organism,8-10 and even to other living organisms.11 Recent studies report a number of alterations taking place in bystander cells and tissues, including induction of DNA strand breaks,2,3,8,12,13 altered gene expression,14 cell cycle arrest, apoptosis,2,3,8,15,16 and a number of epigenetic events3,9,10,17 resulting in genomic insta-bility.2,3,18-21 Yet, the precise mechanisms of bystander effects and the nature of bystander signaling remain unclear. However, a newly emerged mechanism of epigenetic control mediated through the involvement of small regulatory RNAs22,23 promises to bridge the gaps in our understanding of a bystander phenomenon.9
There are a number of functional classifications of small regu-latory RNAs, and among them, microRNAs (miRNAs) are of a particular interest. MiRNAs are evolutionally conserved, small, single-stranded, non-protein-coding RNA molecules which are presently recognized as major regulators of gene expression.22-24 MiRNAs are initially expressed as primary miRNAs (pri-miRNAs), which form a hairpin containing the mature miRNA sequence. This hairpin is released in the nucleus by a dsRNA-specific ribo-nuclease, Drosha, to form a precursor miRNA (pre-miRNA).25-27 The pre-miRNAs are then exported to the cytoplasm, where Dicer, a member of the RNAse III superfamily, cleaves the pre-miRNA leading to the formation of a mature miRNA.25-27 To regulate target mRNAs expression, mature miRNAs must associate with the RNA-induced silencing complex (RISC) proteins27 such as Argonaute (Ago),28 PACT (a protein activator of the interferon-induced protein
*Correspondence to: Olga Kovalchuk; Department of Biological Sciences; University of Lethbridge; 4401 University Drive; Lethbridge, Alberta T1K3M4 Canada; Fax: 403.329.2242; Email: [email protected]
Submitted: 02/13/08; Accepted: 03/23/08
Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/article/5981
Report
Sex-specific microRNAome deregulation in the shielded bystander spleen of cranially exposed miceIgor Koturbash,1,† Franz J. Zemp,1,† Kristy Kutanzi,1 Lidiya Luzhna,1 Jonathan Loree,1 Bryan Kolb2 and Olga Kovalchuk1,*
1Department of Biological Sciences; University of Lethbridge; Alberta, Canada; 2Department of Neuroscience; University of Lethbridge; Alberta, Canada
†These authors contributed equally to this work.
Abbreviations: RISC, RNA-induced silencing complex; IR, Ionizing radiation; pri-miRNAs, primary miRNAs; pre-miRNA, precursor miRNA; PACT, a protein activator of the interferon-induced protein kinase, PKR; qRT-PCR, quantitative real-time polymerase chain reac-tion; CAST, castration; OVX, ovariectomization; H&E, hematoxylin and eosin; TMAs, tissue microarrays
Key words: bystander effect, microRNA, DICER, RISC, sex differences

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
www.landesbioscience.com Cell Cycle 1659
kinase, PKR),29 fragile X mental retardation protein (FXR),30 and TudorSN,31 among others.32
Dicer was found to stably associate with the Ago2 protein,28 while the FxR protein interacts with Ago2 to modulate Dicer activity.30 PACT has been shown to interact with Dicer without facilitating its pre-miRNA cleavage activity,29 and Tudor-SN is important to the mechanism of RNA interference and gene regulation.31
When associated with RISCs, miRNAs bind to a specific section of target mRNAs in a sequence- specific manner, serving as translational suppressors and, thus, as regulators of protein production.26,33,34 By doing so, the RISC associated regulatory miRNAs impact cellular differentiation, proliferation, apoptosis, and even predisposition to cancer.35,36 Indeed, aberrant levels of miRNAs have been reported in a variety of human cancers.35-37 Furthermore, several studies pointed towards a role of the miRNA processing machinery in malignant transformation.38-41 However, the functions of miRNAs and the miRNA processing machinery in response to genotoxic stress42 and, in particular, the radiation-induced bystander effect, have yet to be studied in detail.
Our previous studies hinted at the involvement of miRNAs in bystander effects.9 The present study was designed to scrutinize the role of the microRNAome and microRNA machinery in the regu-lation of a cellular response to the direct (whole-body) or indirect (cranial) exposure to IR in the spleen tissue of male and female mice. Here, we show for the first time that radiation exposure results in a significant and sex-specific deregulation of microRNAome and miRNA processing machinery in the directly exposed and bystander spleen.
Results and Discussion
In this study, we utilized an in vivo murine model to identify and compare microRNAome changes in the spleen of whole-body and head-exposed animals. In the course of the study, experimental animals (males and females of comparable weight) were divided in several experimental cohorts. The ‘whole-body exposed’ cohort received 1 Gy of whole-body irradiation of X-rays. The ‘bystander’ cohort received 1 Gy of X-ray exposure to the skull while the rest of the animal body was protected by a medical grade ~3 mm thick lead shield (of the same type used for the human body protection in diag-nostic radiology). The identical shielding was used in the published studies on the in vivo bystander effect in rodent models.8-10 The protection of the shielded ‘bystander’ tissue was complete as verified by careful dosimetry. Control mice were sham treated. The animals were humanely sacrificed either 6 hours or 96 hours after exposure to analyze both the initial and persistent responses.
MicroRNAome changes in spleen of whole-body and head exposed male and female mice. Analysis of the murine spleen microRNAome revealed a number of intriguing patterns. Firstly, basal levels of miRNA expression in the male and female control spleen suggested a steady-state sex-specific miRNA expression pattern. We found eight microRNAs upregulated in the spleen tissue of intact male mice in comparison with females, and four upregu-lated in the intact female spleen in comparison with the males (Fig. 1). This control comparison was utilized to eliminate false assump-tions of sex-specific radiation- and bystander-induced miRNAs.
Next, we analyzed microRNAome changes upon X-ray expo-sure in the directly irradiated and bystander spleen. Since we had
previously noted that both direct irradiation and bystander effects in spleen were very pronounced 96 hours after exposure, we decided to use this time-point for the initial analysis of radiation-induced microRNAome changes.
We found that exposure to ionizing radiation resulted in gross and sex-specific microRNAome perturbations in the murine spleen tissue. In the whole-body exposure group, we found 12 significantly regulated miRNAs in males (four up and eight downregulated) and 20 significantly regulated in females (five up and 15 downregulated), when compared to their respective controls. Common to both males and females was the significant ~2.5-fold upregulation of miR-34a and a ~1.3-fold downregulation of miR-150. These expression levels were independently confirmed by qRT-PCR (data not shown).
The observed upregulation of miR-34a in the exposed spleen tissue of males and females constitutes an important finding. MiR-34a has recently been shown to be strongly regulated via the transcription factor p53.43,44 Thus, the significant upregulation of miR-34a is a logical response, considering that p53 has long been shown to be involved in the response to radiation.8,45,46 Further, miR-34a has been shown to influence the E2F family of transcription factors in human cells, leading to increases in apoptosis and G1 cell cycle arrest.8,47-
49 Consequently, miR-34a is assumed to be a tumor suppressor miRNA;43,44,47 therefore, its upregulation in the exposed spleen may be viewed as a positive protective effect. Importantly, miR-34a upregulation is well-correlated with our recently reported observation of the IR-induced upregulation of p53 in murine spleen.8
An interesting female-specific observation in the whole-body exposure effects on spleen involved three members of the ‘oncogenic’ miR-17 family that are known to be involved in the regulation of proliferation and apoptosis.36 MiRNAs 17-5p, 20a and 106a (Fig. 2) were all downregulated in the female spleen, while remaining unchanged in the male spleen. Such changes in radiation exposed tissues were, in part, expected as IR is well known to disturb the fine balance between proliferation and apoptosis in directly exposed tissue.2,3,8,19,20 However, the observed female-specific regulation of these important miRNAs is an important novel finding that could suggest a mechanism behind sex-specific radiation induced tumor formation.
Figure 1. Hierarchical clustering of differentially expressed miRNA genes in male and female spleen samples. Hierarchical clusters of significantly altered miRNAs (as determined by ANOVA) across the male and female mouse spleen. Red denotes high expression levels, whereas green depicts low expression levels. Each miRNA listed is significantly and differentially expressed (p < 0.05) between male and female mice.
©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
1660 Cell Cycle 2008; Vol. 7 Issue 11
The remaining regulated ‘sex-specific miRNAs’ in spleen tissue in response to whole-body irradiation could further be delineated, insofar as their regulation and analysis of their putative and/or known targets are concerned. However, this report was intended to focus on a bystander phenomenon, and therefore the direct exposure data will be analyzed in the context of future experiments.
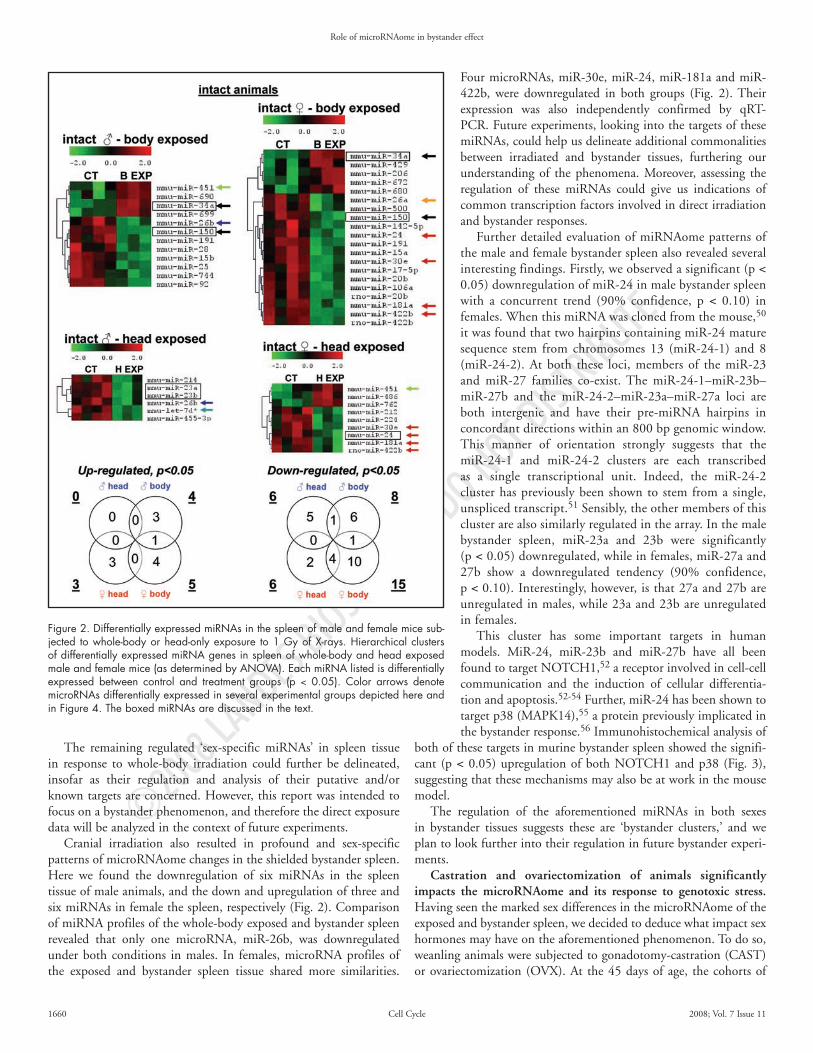
Cranial irradiation also resulted in profound and sex-specific patterns of microRNAome changes in the shielded bystander spleen. Here we found the downregulation of six miRNAs in the spleen tissue of male animals, and the down and upregulation of three and six miRNAs in female the spleen, respectively (Fig. 2). Comparison of miRNA profiles of the whole-body exposed and bystander spleen revealed that only one microRNA, miR-26b, was downregulated under both conditions in males. In females, microRNA profiles of the exposed and bystander spleen tissue shared more similarities.
Four microRNAs, miR-30e, miR-24, miR-181a and miR-422b, were downregulated in both groups (Fig. 2). Their expression was also independently confirmed by qRT-PCR. Future experiments, looking into the targets of these miRNAs, could help us delineate additional commonalities between irradiated and bystander tissues, furthering our understanding of the phenomena. Moreover, assessing the regulation of these miRNAs could give us indications of common transcription factors involved in direct irradiation and bystander responses.
Further detailed evaluation of miRNAome patterns of the male and female bystander spleen also revealed several interesting findings. Firstly, we observed a significant (p < 0.05) downregulation of miR-24 in male bystander spleen with a concurrent trend (90% confidence, p < 0.10) in females. When this miRNA was cloned from the mouse,50 it was found that two hairpins containing miR-24 mature sequence stem from chromosomes 13 (miR-24-1) and 8 (miR-24-2). At both these loci, members of the miR-23 and miR-27 families co-exist. The miR-24-1–miR-23b–miR-27b and the miR-24-2–miR-23a–miR-27a loci are both intergenic and have their pre-miRNA hairpins in concordant directions within an 800 bp genomic window. This manner of orientation strongly suggests that the miR-24-1 and miR-24-2 clusters are each transcribed as a single transcriptional unit. Indeed, the miR-24-2 cluster has previously been shown to stem from a single, unspliced transcript.51 Sensibly, the other members of this cluster are also similarly regulated in the array. In the male bystander spleen, miR-23a and 23b were significantly (p < 0.05) downregulated, while in females, miR-27a and 27b show a downregulated tendency (90% confidence, p < 0.10). Interestingly, however, is that 27a and 27b are unregulated in males, while 23a and 23b are unregulated in females.
This cluster has some important targets in human models. MiR-24, miR-23b and miR-27b have all been found to target NOTCH1,52 a receptor involved in cell-cell communication and the induction of cellular differentia-tion and apoptosis.52-54 Further, miR-24 has been shown to target p38 (MAPK14),55 a protein previously implicated in the bystander response.56 Immunohistochemical analysis of
both of these targets in murine bystander spleen showed the signifi-cant (p < 0.05) upregulation of both NOTCH1 and p38 (Fig. 3), suggesting that these mechanisms may also be at work in the mouse model.
The regulation of the aforementioned miRNAs in both sexes in bystander tissues suggests these are ‘bystander clusters,’ and we plan to look further into their regulation in future bystander experi-ments.
Castration and ovariectomization of animals significantly impacts the microRNAome and its response to genotoxic stress. Having seen the marked sex differences in the microRNAome of the exposed and bystander spleen, we decided to deduce what impact sex hormones may have on the aforementioned phenomenon. To do so, weanling animals were subjected to gonadotomy-castration (CAST) or ovariectomization (OVX). At the 45 days of age, the cohorts of
Figure 2. Differentially expressed miRNAs in the spleen of male and female mice sub-jected to whole-body or head-only exposure to 1 Gy of X-rays. Hierarchical clusters of differentially expressed miRNA genes in spleen of whole-body and head exposed male and female mice (as determined by ANOVA). Each miRNA listed is differentially expressed between control and treatment groups (p < 0.05). Color arrows denote microRNAs differentially expressed in several experimental groups depicted here and in Figure 4. The boxed miRNAs are discussed in the text.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
www.landesbioscience.com Cell Cycle 1661
CAST and OVX animals were exposed to radiation using the protocol described above.
Our study revealed that gonadotomy itself profoundly affected the steady-state murine spleen microRNAome. When comparing the CAST or OVX animals to their intact counterparts, we found 76 significantly regulated (26 up and 50 downregulated; p > 0.05) in males and 104 significantly regulated (35 up and 69 downregu-lated; p > 0.05) in females (Suppl. Fig. 1). This suggests that over one-quarter of female and one-fifth of male miRNAs tested in the entire array could potentially be regulated via the endocrine system. Furthermore, many miRNAs dropped to negligible expression levels as a result of gonadotomy (boxed-miRNAs, Suppl. Fig. 1), providing a plethora of excellent potential targets for the investigation of sex-regulated miRNAs.
Radiation exposure significantly influenced the microRNAome level of CAST and OVX animals. Whole-body irradiation led to the elevated levels of two and seven miRNAs and to the decreased levels of one and four miRNAs in CAST and OVX animals, respectively.
Relating the OVX and CAST whole-body exposed data to that of whole-body exposed intact animals, we can see that some of the radiation-induced regulation may in fact be due to the presence of sex-hormones. For example, the members of the miR-17 family, that were specifically downregulated in the intact female mice, were no longer responsive to radiation in the OVX mice, suggesting a role of estrogen in their regulation. Basal expression levels of these miRNAs dropped significantly in response to ovarectomization, further substantiating this observation (Fig. 4).
MiR-34a, which was upregulated in males and females in response to radiation, was still significantly regulated in CAST mice (p < 0.05) and while the trend still remained in OVX mice, suggesting this response has little or no relation to the presence of sex-hormones (Fig. 4).
In bystander spleen, radiation induced the upregulation of 11 and downregulation of three miRNAs in CAST males, and a complete lack of regulation in females (Fig. 4). This is an interesting result, as it suggests that estrogen may play a large role in altering the microR-NAome in the spleen of females. Moreover, the ‘bystander clusters’ previously discussed have lost their regulation in the sterilized animals. These miRNAs did have some sex-specificity in their expres-sion in intact bystander spleen tissue, lending more evidence that sex-hormones are attenuating the bystander response. Furthermore, bystander responsiveness of NOTCH1 and p38 was also lost in the CAST and OVX animals (data not shown).
Sex-specific changes in the levels of dicer and RISC proteins upon whole-body and cranial exposure in murine spleen. Production and function of miRNAs require a set of proteins collectively referred to as the microRNA processing machinery (miR machinery).24,26,32-
34 The altered balance of miR machinery components may result in altered miRNA levels, thus influencing the interaction between miRNAs and their cognate mRNAs.24,26,32-34
Having seen pronounced and sex-specific miRNA alterations in the exposed and bystander murine spleen, we decided to investigate
radiation and bystander-induced changes in the miR machinery proteins. In this study, we analyzed and compared the levels of the RNA processing enzyme Dicer and the RISC machinery compo-nents Ago2, FxR, TUDOR-SN and PACT in the male and female spleen upon whole-body and cranial irradiation.
IHC analysis revealed that whole-body and cranial radiation exposure led to a significant (p < 0.05) upregulation of Dicer expres-sion in the spleen of male and female mice 6 and 96 hours after exposure (Fig. 5). Interestingly, 96 hours after exposure upregulation was more pronounced in males than in females.
Dicer expression patterns in CAST and OVX animals differed from that of intact mice. In CAST animals, whole-body exposure let to a less pronounced Dicer induction, while cranial exposure led to a small but significant Dicer induction only 96 hours after irradia-tion. No Dicer changes were seen in CAST animals 6 hours after cranial exposure. Similarly, in the OVX spleen, small but statistically significant changes were seen only 6 hours after whole-body or head exposure.
Dicer is involved in the processing of pre-miRNAs in the cytoplasm, leading to the formation of mature miRNAs.57,58 Any alterations in Dicer levels can result in drastic changes in the microR-NAome. For example, Dicer knockdown cell lines had significant downregulation of a large variety of miRNAs.58 Further, Dicer expression levels have been implicated in the deregulation of the microRNAome of many cancers. It has recently been shown that
Figure 3. Elevated levels of p38 and NOTCH1 in the bystander spleen of the cranially exposed male and female mice. The levels of 38 and Notch1 positive cells relative to the control are shown, mean values ± SEM, n = 400, *p < 0.05, Student’s t-test, Dunnett’s test and Tukey-Kramer test.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
1662 Cell Cycle 2008; Vol. 7 Issue 11
Dicer levels were downregulated in lung carcinoma39 and upregu-lated in prostate adenocarcinoma38 and Burkitt lymphoma.59
Dicer upregulation in Burkitt lymphoma is especially intriguing. Radiation is known to induce lymphoma in both humans and mice,60,61 and spleen is an important radiation and lymphoma target organ.62 Therefore, the reported radiation-induced upregulation of Dicer in lymphoma may be in part correlated with our observation of Dicer upregulation in the exposed and bystander spleen. The impact of radiation-induced Dicer upregulation on the formation of radia-tion-induced lymphoma has to be further investigated.
Overall, our study is the first showing that Dicer levels may be changed in cells and tissues in response to genotoxic stress. Dicer, therefore, may play a crucial role in fine-tuning genotoxic stress responses mediated via microRNAs in both the directly exposed and bystander tissues. Besides being an important RNA interference regulator, Dicer is also known to effect the cell cycle G1 and DNA damage checkpoints.63 Furthermore, Dicer plays a substantial role in the maintenance of centromeric heterochromatin and chromosome
segregation,64-67 both of which are affected by direct irradiation68 and bystander effects (reviewed in refs. 18–21). Therefore, we may speculate that these processes may also be perturbed due to Dicer deregulation.
To regulate the expression of their target mRNAs, miRNAs need to be associated with proteins of the RISC complex. Amongst those, Ago2 is one of the most crucial.28,32 We noted that in the male spleen, whole-body radiation exposure resulted in a downregulation of Ago2 96 hours after irradiation, while cranial exposure led to upregulation of Ago2 6 hours after treatment (p < 0.05; Fig. 7). In females, whole-body irradiation induced a significant drop in Ago2 expression 6 and 96 hours after exposure, and cranial exposure alone caused a significant decrease in Ago2 levels only 96 hours after expo-sure (p < 0.05; Fig. 6).
Interestingly, sterilized animals exhibited different patterns of Ago2 expression upon irradiation. In response to whole-body and cranial irradiation, significant upregulation of Ago2 was found in the spleen. Upregulation was seen both 6 and 96 hours after treatment and was more pronounced in CAST than in OVX animals (Fig. 7).
Ago2 is very important for the final Dicer-mediated processing to mature miRNAs, and for further interaction with the other RISC members.69 Notably, studies have shown that Ago levels were significantly elevated in multiple tumor cells lines.70 These experi-ments lead us to hypothesize that upregulation of Ago2 may have a negative influence on radiation exposed and bystander tissue. Thus, future works should explore the dysregulation of Ago2 in radiation- and bystander-induced carcinogenesis in order to further understand these mechanisms.
FXR levels in the spleen were found to be elevated in cranially exposed intact males after 6 hours, whole-body exposed intact females after 96 hours, and in the whole-body exposed CAST animals after 6 hours of X-ray exposure (Fig. 6). Notably, in OVX animals, all treat-ments led to a significant upregulation of FXR. FXR interacts with mammalian Ago2 and is associated with Dicer activity.30
PACT is another vital member of the RISC complex, which inter-acts directly with Dicer and contributes to its stabilization.29 PACT levels were downregulated in the exposed male and female spleen 6 hours after whole-body exposure, while expression was elevated both 6 and 96 hours after cranial irradiation in the male spleen, and downregulated 96 hours in the female bystander spleen (p < 0.05). Interestingly, CAST and OVX animal groups exhibited different patterns of PACT induction upon irradiation. CAST animals showed upregulation of PACT only 6 hours after whole-body X-ray exposure, while OVX animals showed upregulation of PACT 96 hours following whole-body and cranial irradiation. Data on the cellular effects of PACT are relatively scarce. The upregulation of PACT was reported in bronchoalveolar carcinoma, and higher levels of PACT were consid-ered a poor prognostic factor.71 The role of PACT in genotoxic stress responses and carcinogenesis has yet to be established.
TUDOR-SN levels were significantly affected in the exposed male animals. Whole-body irradiation caused downregulated TUDOR-SN 6 and 96 hours after exposure, while cranial irradiation led to TUDOR-SN downregulation only at the 6 hour time point. In females, TUDOR-SN was significantly upregulated 96 hours following whole-body exposure.
Patterns of TUDOR-SN induction by radiation in the spayed mice differed significantly from those in the intact ones. In CAST
Figure 4. Differentially expressed miRNAs in spleen of castrated (CAST) and ovariectomized (OVX) mice subjected to whole-body and head exposure to 1 Gy of X-rays. Hierarchical clusters of differentially expressed miRNA genes in spleen of whole-body and head exposed CAST and OVX mice (as determined by ANOVA). Each miRNA listed is differentially expressed (p < 0.05) between control and treatment groups. Color arrows denote microR-NAs differentially expressed in several experimental groups depicted here and in Figure 2.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
www.landesbioscience.com Cell Cycle 1663
animals, whole-body exposure led to significantly increased levels of TUDOR-SN 6 and 96 hours after irradiation, and cranial exposure resulted in upregulation of TUDOR-SN 96 hours after treat-ment. In OVX animals, TUDOR-SN was upregulated after all treatments and all time-points. TUDOR-SN is an important RISC machinery component that harbors a recognizable nuclease domain and could that could contribute to the RNA degradation observed in RNAi.31 Its exact role in the RISC complexes has to be further delineated.
Overall, whole-body and cranial exposure induced patterns of miRNA machinery that were significantly different in males and females, and in CAST and OVX animals.
Intriguingly, we noted that upon whole body and cranial irradiation, Dicer levels in spleen of male and female mice exhibit somewhat different trends than levels of Ago2 and other members of the RISC complex (Fig. 6). We speculate that if Dicer expres-sion increases, this wouls result in the elevated levels of microRNAs, which might significantly alter a variety of cellular processes and alter cellular homeostasis. In an attempt to prevent this, from happening, cells may try to downregulate levels of the RISC complex members, especially Ago2, which guides the interaction between miRNAs and mRNAs. The balance between different members of a miRNA production pathway and its response to genotoxic insult is a completely novel area of research. The potential for discoveries in this area is vast, as miRNAs have a great deal of promise as regu-lators to cellular stress, and mechanisms behind their regulation are becoming increasingly important.
Both the deregulated microRNAome and the altered expression of Dicer and RISC components may directly or indirectly result in changes in the expression of various proteins that partake in crucial cellular pathways such as the maintenance of genome stability, DNA repair, cell cycle control, apoptosis, proliferation and differentiation. All these processes are affected by direct irradiation, as well as by bystander effects. The altered microRNAome and miR machinery may also be explored as potential radiation exposure and bystander effect biomarkers.
It is important to note that while passing through tissue, X-rays can be reflected, leading to the formation of a small ‘scatter’ dose of radiation in the protected tissue.8-10 Yet, exposure of animals to a scatter dose did not exert any changes in the exposed spleen (data not shown). We therefore concluded that changes observed in the shielded spleen tissue can be classified as true bystander effects. This conclusion is also supported by a series of our previous studies that also showed that the observed in vivo bystander effects are not initi-ated by the scatter-dose.8-10
Conclusions
In this study, we report for the first time the pronounced and sex-determined microRNAome deregulation that occurs in murine spleen tissue upon whole-body and cranial irradiation. The main findings of the present study are that: (i) male and female spleen
tissues are characterized by very distinct microRNA patterns; (ii) radiation exposure alters the microRNAome of male and female animals and induces sex-specific changes of microRNA levels in the exposed spleen tissue; (iii) cranial-only exposure also influences the spleen microRNAome and causes distinct changes in the male and female shielded bystander spleen; (iv) irradiation and bystander miRNA response may, in part, be regulated through altered levels of miRNA processing machinery. Importantly, these are the first data on the radiation- and bystander-induced microRNAome changes in animal tissue in vivo.
Furthermore, our study is the first to investigate the role of sex hormones in microRNAome regulation in vivo. Additionally, radia-tion and bystander responses of sterilized animals differ drastically from intact animal responses.
Our microRNAome datasets may provide a roadmap for further analysis of the role of the microRNAome in genotoxic stress responses. Specifically, our data provide an opportunity to integrate sex hormones and stress responses. While the radiation- and bystander-induced microRNAs identified in our study may further be explored as sex-specific biomarkers of direct and indirect radiation effects.
It is known that radiation-induced cancer occurs more frequently in males,72-74 yet the molecular grounds of this sex-specific bias are not well understood. The microRNA patterns observed in this study in the male and female spleen tissue may be used to further explore
Figure 5. Changes in the Dicer levels in the exposed and bystander spleen of male, female, castrated (CAST) and ovariectomized (OVX) mice. The levels of Dicer positive cells relative to the control are shown, mean values ± SEM, n = 400, *p < 0.05, Student’s t-test, Dunnett’s test and Tukey-Kramer test.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
1664 Cell Cycle 2008; Vol. 7 Issue 11
the molecular basis of sex differences in radiation-induced carcino-genesis.
Another important outcome of this study is a proof of radia-tion- and bystander-inducibility of Dicer and other RICS complex members. Dicer is a key RNAse involved in the final steps of microRNA maturation. It works in a tight collaboration with Ago2, PACT, FXR and TUDOR-SN to produce mature miRNAs. All of these proteins exhibit altered expression upon direct irradiation, as well as changes in the shielded bystander spleen. The altered levels of microRNA machinery compliment the observed microRNAome deregulation.
Cellular and organismal repercussions of the observed changes need to be further defined. Moreover, the mechanisms of sex differ-ences in the microRNAome and microRNA processing machinery levels have to be further explored. Altered Dicer and RISC levels, together with the altered microRNAome, may serve as novel and sex-specific biomarkers of exposure and bystander effects.
Materials and Methods
Animal model. In this study, we analyzed microRNAome changes in the spleen of male and female C57BL/6 mice following in vivo
cranial irradiation exposure. The 45 days old male mice (30 males and 30 female) were randomly assigned to different treatment groups. The whole-body exposed cohort (10 male and 10 female animals) received 1 Gy of whole-body exposure to X-rays (5 cGy/s, 90 kV, 5 mA). The second cohort (10 male and 10 female animals) received 1 Gy of X-rays exposure to a scull only, while the rest of animal body were protected by a ~3 mm thick lead shield, the same type as used for a human body protection in diagnostic radiology. The protection of shielded ‘bystander’ tissue was complete, as verified by careful dosimetry using a RAD-CHECK™ monitor (Nuclear Associates div. of Victoreen, Inc., FL, USA). The control mice (10 male and 10 female animals) were sham treated. The animals were humanely sacrificed either 6 hours or 96 hours (4 days) after exposure. These time points were chosen to analyze the initial (6 hours) as well as persistent (4 days) responses, if any. Spleen tissue was sampled upon sacrifice and processed for further molecular studies. The experiment was independently reproduced according to the same scheme using 18 male and 18 female animals. Both experiments yielded congruent results. Another cohort of 10 animals was exposed to an approximate scatter dose of ~0.01 Gy.8-10 To determine the bystander ventral/spleen/thigh skin dose resulting from photon scatter within a mouse
Figure 6. Changes in the Ago2, PACT, FXR and TUDOR-SN levels in the exposed and bystander spleen of male, female, castrated (CAST) and ovariectomized (OVX) mice. The levels of Ago2, PACT, FXR and TUDOR-SN positive cells relative to control are shown, mean values ± SEM, n = 400, *p < 0.05, #p < 0.10, Student’s t-test, Dunnett’s test and Tukey-Kramer test.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
www.landesbioscience.com Cell Cycle 1665
itself, a Monte Carlo simulation was performed.8-10 The skin was assumed to be 0.1 cm thick, and a 2 mm thick lead shield covered one half of the mouse. An absorbed dose was tallied within the skin region in several locations in both the unshielded dorsal regions and the shielded ventral/thigh regions. The absorbed dose to the ventral skin/spleen located right beneath the skin from a 90 kVp X-ray spectrum was determined to be approximately 0.014–0.017 Gy for a 1 Gy dose delivered to the dorsal skin. Additionally, the scatter dose upon cranial was directly measured, and was found to be in the range of ~0.01 Gy.
To dissect the effect of sex hormones on the bystander effects manifestations, a cohort of 30 male and 30 female animals were subjected to gonadotomy at the age of 18 days. At the age of 45 days, the castrated and ovariectomised (CAST and OVX) animals were subjected to IR treatment using the same scheme as the intact animals.
Handling and care of animals was in strict accordance with the recommendations of the Canadian Council for Animal Care and Use. The procedures were approved by the University of Lethbridge Animal Welfare Committee.
miRNA microarray expression analysis. Total RNA was extracted from mouse spleen tissues using TRIzol Reagent (Invitrogen, Burlington, Ontario) according to the manufacturer’s instruc-tions. Tissue from 3 animals per group was used for the analysis. The miRNA microarray analysis was performed by LC Sciences (Houston, TX). Ten micrograms of total RNA were size-fractionated (<200 nucleotides) by using a mirVana kit (Ambion, Austin, TX). Poly-A tails were added to the RNA sequences at the 3' ends using a poly(A) polymerase, and nucleotide tags were then ligated to the poly-A tails. The tagged RNAs were hybridized to the dual-channel microarray μParaFlo microfluidics chips (LC Sciences) containing 439 miRNA probes to rat and mouse miRNAs and then labeled with tag-specific dendrimer Cy3 and Cy5 fluorescent dyes. Dye switching was performed to eliminate the dye bias. The detection probes melting temperature was balanced by incorporating varying numbers of modified nucleotides with the increased binding affini-ties. Hybridization images were collected using a GenePix 4000B laser scanner (Molecular Devices, Sunnyvale, CA), and then digitized using the Array-Pro image analysis software (Media Cybernetics, Silver Spring, MD). The maximum signal level of background probes was 180. A miRNA detection signal threshold was defined as twice the maximum background signal. Normalization was performed with a cyclic LOWESS (locally weighted regression) method to remove system-related variations, as previously described.75,76 Data adjustments included data-filtering, log 2 transformation, and gene centering and normalization. The t-test analysis was conducted between the control, whole-body exposed and bystander sample groups. MicroRNAs with p-values <0.05 were selected for cluster analysis.
Quantitative real-time PCR (qRT-PCR) expression analysis. Quantitative real-time PCRs (qRT-PCRs) were performed by using SuperTaq Polymerase (Ambion) and a mirVana qRT-PCR miRNA Detection Kit (Ambion) following the manufacturer’s instructions. Reactions contained mirVana qRT-PCR Primer Sets specific for miR-34, miR-17-5p, miR-20, miR-26, miR-92, miR-106a and miR-150, human 5S rRNA served as a positive control. Quantitative real-time PCR was performed on a SmartCycler (Cepheid, Sunnyvale, CA).
End-point reaction products were also analyzed on a 3.5% high-resolution agarose gel stained with ethidium bromide.
Immunohistochemistry. Paraffin embedding and sectioning were conducted at Central Vet Labs, Edmonton, AB. The sections were stained with hematoxylin and eosin (H&E) for the histopatho-logical examination. Following the pathological examination, the tissues were assembled into tissue microarrays (TMAs) with 2 mm cores. TMAs offer great benefits for the fast, consistent and effi-cient analysis of large amounts of data. The TMAs were produced by Pantomics, Inc., (www.pantomics.com). Immunohistochemical staining was conducted using the antibodies against Dicer, FXR, PACT, TUDOR-SN, NOTCH 1 (Santa Cruz Biotechnology, Santa Cruz, CA), AGO 2 and p38 (Abcam, Cambridge, MA) in accordance with the manufacturer’s recommendations. In brief, upon the citrate buffer the epitope retrieval slides were rinsed and subjected to serum blocking to prevent the non-specific binding of immunoglobulin. Sections then were incubated with primary antibodies, rinsed and subjected to peroxidase blocking. Following the peroxidase blocking, the slides were incubated with the secondary biotinylated antibody, subjected to HRP-Streptavidin detection and counterstained with hematoxylin. Positive cells stained brown/purple, while negative cells stained blue. Positive cells were enumerated in, at least, 20 high power fields.
Statistical analysis. Statistical analysis (Student’s t-test, Bonferroni correction, Tukey-Kramer test and Dunnett’s tests) was performed using the MS Excel 2000 and JMP5 software packages.
Acknowledgements
We are grateful to Karen Dow-Cazal and Charlotte Holmes for animal care. We appreciate the help of Dr. Valentina Titova in proofreading this manuscript. This research was supported by ACB and CIHR operating grants to Olga Kovalchuk. Igor Koturbash is the recipient of the Dr. Cyril Kay Alberta Cancer Board (ACB) Graduate Scholarship. Kristy Kutanzi is the recipient of NSERC Canada Graduate Scholarship, Alberta Heritage Foundation for Medical Research (AHFMR) Graduate Scholarship, Keith and Hope Ferguson Memorial Scholarship and the Queen Elizabeth Graduate Scholarship. Lidiya Luzhna is a recipient of the Alberta Cancer Board Graduate Studentship. Jonathan Loree was a recipient of the NSERC Undergraduate Summer Research Award.
Note
Supplementary materials can be found at:www.landesbioscience.com/supplement/KoturbashCC7-11-Sup.pdf
References 1. Little JB. Radiation carcinogenesis. Carcinogenesis 2000; 21:397-404. 2. Belyakov OV, Mitchell SA, Parikh D, Randers Pehrson G, Marino SA, Amundson SA,
Geard CR, Brenner DJ. Biological effects in unirradiated human tissue induced by radiation damage up to 1 mm away. Proc Natl Acad Sci USA 2005; 102:14203-8.
3. Sedelnikova OA, Nakamura A, Kovalchuk O, Koturbash I, Mitchell SA, Marino SA, Brenner DJ, Bonner WM. DNA double-strand breaks form in bystander cells after microbeam irradiation of three-dimensional human tissue models. Cancer Res 2007; 67:4295-302.
4. Khan MA, Hill RP, Van Dyk J. Partial volume rat lung irradiation: an evaluation of early DNA damage. Int J Radiat Oncol Biol Phys 1998; 40:467-76.
5. Khan MA, Van Dyk J, Yeung IW, Hill RP. Partial volume rat lung irradiation; assessment of early DNA damage in different lung regions and effect of radical scavengers. Radiother Oncol 2003; 66:95-102.
6. Brooks AL, Retherford JC, McClellan RO. Effect of 239PuO2 particle number and size on the frequency and distribution of chromosome aberrations in the liver of the Chinese hamster. Radiat Res 1974; 59:693-709.
7. Brooks AL. Evidence for ‘bystander effects’ in vivo. Hum Exp Toxicol 2004; 23:67-70.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
1666 Cell Cycle 2008; Vol. 7 Issue 11
8. Koturbash I, Loree J, Kutanzi K, Koganow C, Pogribny I, Kovalchuk O. In Vivo Bystander Effect: Cranial X-Irradiation Leads to Elevated DNA Damage, Altered Cellular Proliferation and Apoptosis, and Increased p53 Levels in Shielded Spleen. Int J Radiat Oncol Biol Phys 2008; 70:554-62.
9. Koturbash I, Boyko A, Rodriguez Juarez R, McDonald RJ, Tryndyak VP, Kovalchuk I, Pogribny IP, Kovalchuk O. Role of epigenetic effectors in maintenance of the long-term persistent bystander effect in spleen in vivo. Carcinogenesis 2007; 28:1831-8.
10. Koturbash I, Rugo RE, Hendricks CA, Loree J, Thibault B, Kutanzi K, Pogribny I, Yanch JC, Engelward BP, Kovalchuk O. Irradiation induces DNA damage and modulates epigenetic effectors in distant bystander tissue in vivo. Oncogene 2006; 25:4267-75.
11. Mothersill C, Smith RW, Agnihotri N, Seymour CB. Characterization of a radiation-induced stress response communicated in vivo between zebrafish. Environ Sci Technol 2007; 41:3382-7.
12. Smilenov LB, Hall EJ, Bonner WM, Sedelnikova OA. A microbeam study of DNA double-strand breaks in bystander primary human fibroblasts. Radiat Prot Dosimetry 2006; 122:256-9.
13. Han W, Wu L, Chen S, Bao L, Zhang L, Jiang E, Zhao Y, Xu A, Hei TK, Yu Z. Constitutive nitric oxide acting as a possible intercellular signaling molecule in the initiation of radia-tion-induced DNA double strand breaks in non-irradiated bystander cells. Oncogene 2007; 26:2330-9.
14. Klokov D, Criswell T, Leskov KS, Araki S, Mayo L, Boothman DA. IR-inducible clus-terin gene expression: a protein with potential roles in ionizing radiation-induced adaptive responses, genomic instability and bystander effects. Mutat Res 2004; 568:97-110.
15. Belyakov OV, Folkard M, Mothersill C, Prise KM, Michael BD. Bystander-induced apop-tosis and premature differentiation in primary urothelial explants after charged particle microbeam irradiation. Radiat Prot Dosimetry 2002; 99:249-51.
16. Lyng FM, Maguire P, Kilmurray N, Mothersill C, Shao C, Folkard M, Prise KM. Apoptosis is initiated in human keratinocytes exposed to signalling factors from microbeam irradiated cells. Int J Radiat Biol 2006; 82:339-93.
17. Kaup S, Grandjean V, Mukherjee R, Kapoor A, Keyes E, Seymour CB, Mothersill CE, Schofield PN. Radiation-induced genomic instability is associated with DNA methylation changes in cultured human keratinocytes. Mutat Res 2006; 597:87-97.
18. Morgan WF, Sowa MB. Non-targeted bystander effects induced by ionizing radiation. Mutat Res 2007; 616:159-64.
19. Morgan WF. Non-targeted and delayed effects of exposure to ionizing radiation: I. Radiation-induced genomic instability and bystander effects in vitro. Radiat Res 2003; 159:567-80.
20. Morgan WF. Non-targeted and delayed effects of exposure to ionizing radiation: II. Radiation-induced genomic instability and bystander effects in vivo, clastogenic factors and transgenerational effects. Radiat Res 2003; 159:581-96.
21. Mothersill C, Seymour CB. Radiation-induced bystander effects—implications for cancer. Nat Rev Cancer 2004; 4:158-64.
22. Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regula-tion by microRNAs: are the answers in sight? Nat Rev Genet 2008; 9:102-14.
23. Niwa R, Slack FJ. The evolution of animal microRNA function. Curr Opin Genet Dev 2007; 17:145-50.
24. Nilsen TW. Mechanisms of microRNA-mediated gene regulation in animal cells. Trends Genet 2007; 23:243-9.
25. Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J, Lee J, Provost P, Radmark O, Kim S, Kim VN. The nuclear RNase III Drosha initiates miRNA processing. Nature 2003; 425:415-9.
26. Hutvagner G, Zamore PD. A miRNA in a multiple-turnover RNAi enzyme complex. Science 2002; 297:2056-60.
27. Rana TM. Illuminating the silence: understanding the structure and function of small RNAs. Nat Rev Mol Cell Biol 2007; 8:23-36.
28. Chendrimada TP, Gregory RI, Kumaraswamy E, Norman J, Cooch N, Nishikura K, Shiekhattar R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005; 436:740-4.
29. Lee Y, Hur I, Park SY, Kim YK, Suh MR, Kim VN. The role of PACT in the RNA silencing pathway. EMBO J 2006; 25:522-32.
30. Jin P, Zarnescu DC, Ceman S, Nakamoto M, Mowrey J, Jongens TA, Nelson DL, Moses K, Warren ST. Biochemical and genetic interaction between the fragile X mental retardation protein and the microRNA pathway. Nat Neurosci 2004; 7:113-7.
31. Scadden AD. The RISC subunit TudorsN binds to hyper-edited double-stranded RNA and promotes its cleavage. Nat Struct Mol Biol 2005; 12:489-96.
32. Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R. Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell 2005; 123:631-40.
33. Sevignani C, Calin GA, Siracusa LD, Croce CM. Mammalian microRNAs: a small world for fine-tuning gene expression. Mamm Genome 2006; 17:189-202.
34. Carthew RW. Gene regulation by microRNAs. Curr Opin Genet Devel 2006; 16:203-8. 35. Fabbri M, Ivan M, Cimmino A, Negrini M, Calin GA. Regulatory mechanisms of microR-
NAs involvement in cancer. Expert Opin Biol Ther 2007; 7:1009-19. 36. Esquela Kerscher A, Slack FJ. Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer
2006; 6:259-69. 37. Lu J, Getz G, Miska EA, Alvarez Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL,
Mak RH, Ferrando AA, Downing JR, Jacks T, Horvitz HR, Golub TR. MicroRNA expres-sion profiles classify human cancers. Nature 2005; 435:834-8.
38. Chiosea S, Jelezcova E, Chandran U, Acquafondata M, McHale T, Sobol RW, Dhir R. UpRegulation of Dicer, a Component of the MicroRNA Machinery, in Prostate Adenocarcinoma. Am J Pathol 2006; 169:1812-20.
39. Chiosea S, Jelezcova E, Chandran U, Luo J, Mantha G, Sobol RW, Dacic S. Overexpression of Dicer in precursor lesions of lung adenocarcinoma. Cancer Res 2007; 67:2345-50.
40. Karube Y, Tanaka H, Osada H, Tomida S, Tatematsu Y, Yanagisawa K, Yatabe Y, Takamizawa J, Miyoshi S, Mitsudomi T, Takahashi T. Reduced expression of Dicer associated with poor prognosis in lung cancer patients. Cancer Sci 2005; 96:111-5.
41. Cheng JC, Moore TB, Sakamoto KM. RNA interference and human disease. Mol Genet Metab 2003; 80:121-8.
42. Marsit CJ, Eddy K, Kelsey KT. MicroRNA responses to cellular stress. Cancer Res 2006; 66:10843-8.
43. He L, He X, Lim LP, de Stanchina E, Xuan Z, Liang Y, Xue W, Zender L, Magnus J, Ridzon D, Jackson AL, Linsley PS, Chen C, Lowe SW, Cleary MA, Hannon GJ. A microRNA compo-nent of the p53 tumour suppressor network. Nature 2007; 447:1130-4.
44. He L, He X, Lowe SW, Hannon GJ. microRNAs join the p53 network—another piece in the tumour-suppression puzzle. Nat Rev Cancer 2007; 7:819-22.
45. Blattner C, Sparks A, Lane D. Transcription factor E2F-1 is upregulated in response to DNA damage in a manner analogous to that of p53. Mol Cell Biol 1999; 19:3704-13.
46. Fujikawa K, Hasegawa Y, Matsuzawa S, Fukunaga A, Itoh T, Kondo S. Dose and dose-rate effects of X rays and fission neutrons on lymphocyte apoptosis in p53(+/+) and p53(-/-) mice. Radiat Res 2000; 41:113-27.
47. Welch C, Chen Y, Stallings RL. MicroRNA-34a functions as a potential tumor suppressor by inducing apoptosis in neuroblastoma cells. Oncogene 2007; 26:5017-22.
48. Tazawa H, Tsuchiya N, Izumiya M, Nakagama H. Tumor-suppressive miR-34a induces senescence-like growth arrest through modulation of the E2F pathway in human colon cancer cells. Proc Natl Acad Sci USA 2007; 104:15472-7.
49. Chang TC, Wentzel EA, Kent OA, Ramachandran K, Mullendore M, Lee KH, Feldmann G, Yamakuchi M, Ferlito M, Lowenstein CJ, Arking DE, Beer MA, Maitra A, Mendell JT. Transactivation of miR-34a by p53 broadly influences gene expression and promotes apop-tosis. Mol Cell 2007; 26:745-52.
50. Lagos-Quintana M, Rauhut R, Yalcin A, Meyer J, Lendeckel W, Tuschl T. Identification of tissue-specific microRNAs from mouse. Curr Biol 2002; 12:735-9.
51. Cai X, Hagedorn CH, Cullen BR. Human microRNAs are processed from capped, polyad-enylated transcripts that can also function as mRNAs. RNA 2004; 10:1957-66.
52. Fukuda Y, Kawasaki H, Taira K. Exploration of human miRNA target genes in neuronal differentiation. Nucleic Acids Symp Ser (Oxf ) 2005; 49:341-2.
53. Romer S, Saunders U, Jäck HM, Jehn BM. Notch1 enhances B-cell receptor-induced apoptosis in mature activated B cells without affecting cell cycle progression and surface IgM expression. Cell Death Differ 2003; 10:833-44.
54. Qi R, An H, Yu Y, Zhang M, Liu S, Xu H, Guo Z, Cheng T, Cao X. Notch1 signaling inhibits growth of human hepatocellular carcinoma through induction of cell cycle arrest and apoptosis. Cancer Res 2003; 63:8323-9.
55. Kiriakidou M, Nelson PT, Kouranov A, Fitziev P, Bouyioukos C, Mourelatos Z, Hatzigeorgiou A. A combined computational-experimental approach predicts human microRNA targets. Genes Dev 2004; 18:1165-78.
56. Lyng FM, Maguire P, McClean B, Seymour C, Mothersill C. The involvement of calcium and MAP kinase signaling pathways in the production of radiation-induced bystander effects. Radiat Res 2006; 165:400-9.
57. Tijsterman M, Plasterk RH. Dicers at RISC; the mechanism of RNAi. Cell 2004; 117:1-3. 58. Cummins JM, He Y, Leary RJ, Pagliarini R, Diaz LA Jr, Sjoblom T, Barad O, Bentwich Z,
Szafranska AE, Labourier E, Raymond CK, Roberts BS, Juhl H, Kinzler KW, Vogelstein B, Velculescu VE. The colorectal microRNAome. Proc Natl Acad Sci USA 2006; 103:3687-92.
59. Kaul D, Sikand K. Defective RNA-mediated c-myc gene silencing pathway in Burkitt’s lymphoma. Biochem Biophys Res Commun 2004; 313:552-4.
60. Dange P, Sarma H, Pandey BN, Mishra KP. Radiation-induced incidence of thymic lym-phoma in mice and its prevention by antioxidants. J Environ Pathol Toxicol Oncol 2007; 26:273-9.
61. Focosi D, Galimberti S, Petrini M. Acute myeloid leukemia and follicular lymphoma after very low dose radioiodine therapy for thyroid diseases. Haematologica 2007; 92:96-7.
62. Koturbash I, Pogribny I, Kovalchuk O. Stable loss of global DNA methylation in the radia-tion-target tissue—a possible mechanism contributing to radiation carcinogenesis? Biochem Biophys Res Commun 2005; 337:526-33.
63. Carmichael JB, Provost P, Ekwall K, Hobman TC. ago1 and dcr1, two core components of the RNA interference pathway, functionally diverge from rdp1 in regulating cell cycle events in Schizosaccharomyces pombe. Mol Biol Cell 2004; 15:1425-35.
64. Provost P, Silverstein RA, Dishart D, Walfridsson J, Djupedal I, Kniola B, Wright A, Samuelsson B, Radmark O, Ekwall K. Dicer is required for chromosome segregation and gene silencing in fission yeast cells. Proc Natl Acad Sci USA 2002; 99:16648-53.
65. Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI, Martienssen RA. Regulation of het-erochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science 2002; 297:1833-7.
66. Volpe T, Schramke V, Hamilton GL, White SA, Teng G, Martienssen RA, Allshire RC. RNA interference is required for normal centromere function in fission yeast. Chromosome Res 2003; 11:137-46.
67. Fukagawa T, Nogami M, Yoshikawa M, Ikeno M, Okazaki T, Takami Y, Nakayama T, Oshimura M. Dicer is essential for formation of the heterochromatin structure in vertebrate cells. Nat Cell Biol 2004; 6:784-91.
68. Rodemann HP, Blaese MA. Responses of normal cells to ionizing radiation. Semin Radiat Oncol 2007; 17:81-8.

©2008 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
Role of microRNAome in bystander effect
www.landesbioscience.com Cell Cycle 1667
69. Joshua-Tor L. The Argonautes. Cold Spring Harb Symp Quant Biol 2006; 71:67-72. 70. Jaronczyk K, Carmichael JB, Hobman TC. Exploring the functions of RNA interference path-
way proteins: some functions are more RISCy than others? Biochem J 2005; 387:561-71. 71. Roh MS, Kwak JY, Kim SJ, Lee HW, Kwon HC, Hwang TH, Choi PJ, Hong YS.
Expression of double-stranded RNA-activated protein kinase in small-size peripheral adeno-carcinoma of the lung. Pathol Int 2005; 55:688-93.
72. Yoshida K, Nemoto K, Nishimura M, Seki M. Exacerbating factors of radiation-induced myeloid leukemogenesis. Leuk Res 1993; 17:437-40.
73. Guizard AV, Boutou O, Pottier D, Troussard X, Pheby D, Launoy G, Slama R, Spira A. The incidence of childhood leukaemia around the La Hague nuclear waste reprocessing plant (France): a survey for the years 1978–1998. J Epidemiol Community Health 2001; 55:469-74.
74. Noshchenko AG, Moysich KB, Bondar A, Zamostyan PV, Drosdova VD, Michalek AM. Patterns of acute leukaemia occurrence among children in the Chernobyl region. Int J Epidemiol 2001; 30:125-9.
75. Bolstad BM, Irizarry RA, Astrand M, Speed TP. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003; 19:185-93.
76. Pogribny IP, Tryndyak VP, Boyko A, Rodriguez Juarez R, Beland FA, Kovalchuk O. Induction of microRNAome deregulation in rat liver by long-term tamoxifen exposure. Mutat Res 2007; 619:30-7.