
Analisi semiotica del testo videoludico: l'enunciazione cinetica

25
SINTESI
CONTESTO - Diversi funghi (muffe, lieviti) possono essere presenti in vari ambienti di lavoro ed esserecausa di un’ampia gamma di disturbi respiratori tra cui allergie ed irritazioni. In effetti, numerose proteinedi origine fungina sono state identificate come importanti allergeni. Pertanto, in determinati ambienti dilavoro risulta essenziale rilevare con metodi rapidi ed accurati la presenza di ceppi fungini per monitorarei livelli di esposizione biologica e per disporre di eventuali misure di tutela della salute pubblica. Nelpresente studio è stato utilizzato l’approccio molecolare e filogenetico per evitare che gli isolati funginirisultassero caratterizzati dal solo impiego di metodi basati sull’isolamento in coltura su piastra e suosservazioni di tratti morfologici e fisiologici. Inoltre si è investigato, a livello di sequenza del DNA e deicorrispondenti amminoacidi, su alcuni dei principali allergeni provenienti sia da lieviti (appartenenti agliAscomiceti ed ai Basidiomiceti) che da muffe (Ascomiceti).OBIETTIVI - Lo scopo principale del presente lavoro è consistito nell’ottenere nuove sequenze di DNA,ricavare filogenesi molecolari ed indagare le relazioni evolutive dei funghi isolati da bioaerosol. Inoltre, unaltro importante obiettivo è stato l’analizzare le sequenze degli aminoacidi dedotti delle serina-preoteasialcaline/vacuolari di funghi potenzialmente allergenici e costruire modelli molecolari al fine di localizzareamminoacidi importanti per la struttura e la funzione di questi enzimi.METODI - Le analisi molecolari sono state basate sulla tecnica dell’amplificazione genica mediantePolymerase Chain Reaction (PCR) e sul successivo sequenziamento del DNA di geni nucleari emitocondriali di funghi isolati da bioaerosol di ambienti interni ed esterni. Inoltre, il sequenziamento delDNA e l’analisi degli amminoacidi dedotti sono stati utilizzati per studiare determinati geni funginicodificanti proteine potenzialmente coinvolte in reazioni allergiche. In particolare, le sequenze delleserina-proteasi alcaline/vacuolari sono state elaborate per ricavare un modello tridimensionale edinferenze sulla filogenesi molecolare di queste proteine fungine con potenziale allergico.
SEQUENZIAMENTO DEL DNA ED ANALISIFILOGENETICA SU GENI CODIFICANTI ALLERGENIDI MUFFE E LIEVITI ISOLATI DA BIOAEROSOLDomenico Davolos, Biancamaria Pietrangeli
Dipartimento Installazioni di Produzione e Insediamenti Antropici (DIPIA), Istituto Superiore per la Prevenzione e laSicurezza del Lavoro (ISPESL)
(Parole chiave: serina-proteasi alcalina/vacuolare, bioaerosol, sequenziamento di DNA, introni GT-AG,allergeni fungini )
Vol. 3, n. 3, 25-37
BOW PO/base indexing:EUOSHA - OSH: Fungi [28681C], Allergens [33801D], Toxicity testing [27001D]CIS: Moulds [Fapo], Fungi [Fap], Yeast [Fapasy], Molecular structure [Czym], Allergens [Fyhg], Electrical aerosolanalysis [Qepie], DNA [Frmra]
EUOSHA - OSH: Funghi [28681C], Allergeni [33801D], Analisi di tossicità [27001D]CIS: Muffe [Fapo], Funghi [Fap], Lieviti [Fapasy], Struttura molecolare [Czym], Allergeni [Fyhg], Analisi con aerosolelettrico [Qepie], DNA [Frmra]

RISULTATI - I dati molecolari ottenuti mediante sequenziamento del DNA sono risultati di fondamentaleimportanza per la rapida ed accurata identificazione a vari livelli tassonomici dei funghi isolati dabioaerosol e potenzialmente allergenici. Inoltre, è stata determinata la struttura esone-introne ed è stataricavata una filogenesi molecolare, basata sulle sequenze degli aminoacidi dedotti, delle serina-preoteasialcaline/vacuolari di Penicillium chrysogenum e di altri funghi. Infine, è stato costruito un modellotridimensionale della serina-proteasi al fine di localizzare epitopi riconosciuti da anticorpi IgE ed altriimportanti residui tra i quali amminoacidi conservati che risultano essenziali per la funzione e la strutturaenzimatica.I nostri studi basati sul sequenziamento del DNA di determinati geni fungini integrati con le informazioniricavabili da genomi di muffe e lieviti, di recente completamente sequenziati o in fase di ultimazione,daranno un contributo notevole per incrementare le conoscenze sugli aero-allergeni fungini e sullecaratteristiche generali dell’esposizione biologica a funghi allergenici.
INTRODUZIONE
Studi recenti hanno evidenziato la presenza in vari ambienti di lavoro di diversi taxa fungini, tra cui muffee lieviti, che possono essere causa di problemi per la salute pubblica, quali ad esempio reazioniallergiche. Ai fini del monitoraggio dei livelli di esposizione biologica [1] e dell’eventuali misure a tuteladella salute pubblica, in un determinato ambiente (interno o esterno) è di fondamentale importanzarilevare in modo rapido ed accurato la presenza di ceppi fungini così come è estremamente utilecondurre ricerche a livello molecolare sugli allergeni di specie fungine isolate da bioaerosol. I funghi generalmente vengono caratterizzati su basi morfologiche e fisiologiche ma l’identificazionepuò richiedere tempi lunghi e può rivelarsi problematica per i risultati che spesso presentano variabilitàsignificative. Per ovviare a tali difficoltà e per indagare sulle relazioni evolutive [2,3], qui sono statiutilizzati metodi molecolari basati sulla tecnica dell’amplificazione mediante la reazione a catena dellaDNA polimerasi (PCR, Polymerase Chain Reaction) e sul sequenziamento del DNA. Come marcatorimolecolari sono stati utilizzati geni nucleari tra cui il 18S rDNA, gli spaziatori interni trascritti (ITS1,ITS2), il 5.8S rDNA, il 28S rDNA, il gene codificante la beta-tubulina, il gene del fattore di allungamento1-a (EF1-a) della traduzione e geni mitocondriali (ad esempio cytb codificante la citocromo b) che sonorisultati utili al fine della identificazione a vari livelli tassonomici di funghi isolati da bioaereosol [4-9]. Inoltre, poiché le spore fungine aerodisperse possono essere importanti agenti eziologici di disturbidel tratto respiratorio di natura allergica, sono state condotte delle ricerche su determinati geni funginiche codificano per proteine che potenzialmente possono causare reazioni allergiche. In questo studiovengono presentati i risultati preliminari riguardanti i geni che codificano per le serina-proteasi alcalineresponsabili di reazioni allergiche provenienti da diversi taxa appartenenti al genere Penicillium (muffepotenzialmente allergeniche generalmente presenti in ambienti interni). Infine, vengono presentatidati preliminari relativi alle indagini molecolari condotte su lieviti basidiomiceti aerodispersi raccolti indiversi ambienti interni ed esterni, nell’ambito dell’attività di ricerca del piano triennale 2005-2007dell’ISPESL-DIPIA, con finalità lo studio della variabilità genetica di alcuni dei più importanti allergenifungini.
26
Prev
enzi
one
Ogg
i - lu
glio
- se
ttem
bre
2007

27
Seq
uenz
iam
ento
del
DN
Aed
ana
lisi f
iloge
netic
a su
gen
i cod
ifica
nti a
llerg
eni d
i muf
fe e
liev
iti is
olat
i da
bioa
eros
ol
1. MATERIALI E METODI
1.1 Campionamento e caratterizzazione morfologica dei ceppi fungini
Volumi di aria sono stati raccolti da ambienti esterni (discariche) ed interni (allevamenti di suini; polveri sucarte) generalmente mediante un campionatore Surface Air System (SAS; International PBI) utilizzandodiversi valori di flusso (litri/min). Singole colonie di funghi sono state successivamente isolate conmetodi di coltura su piastre Sabouraud agar. La morfologia degli isolati fungini è stata esaminata tramitemicroscopia a contrasto di fase e microscopia a scansione laser confocale (nel secondo casoaggiungendo ai campioni una soluzione Nano-Orange specifica per proteine; Molecular Probes Inc.).
1.2 Amplificazione e sequenziamento del DNA
Colture pure di 2/3 giorni da piastre agar Sabouraud sono state lisate con un metodo semplice e rapido(Davolos, in preparazione) in modo tale da ottenere DNA genomico amplificabile mediante PCR. Lereazioni di PCR per i geni nucleari dell’RNA ribosomiale (18S-, 28S- e 5.8S- rDNA) e per la regione deglispaziatori interni trascritti (ITS1, ITS2) del DNA ribosomiale sono state eseguite utilizzando i primersuniversali NS1, NS4, NS5, NS8, ITS1, ITS4 [10] e NL-4 [11]. Una regione del gene codificante la beta-tubulina è stata amplificata utilizzando il primer benA1 [12] ed il nuovo primer reverse benA3 (5’-GGAAGGGAACGATGTTGAC-3’). Una regione del gene del fattore EF1-a di allungamento della traduzioneè stata amplificata attraverso l’impiego di nuovi primers: EF1-f (5’-ATGGGTAAGGACGACAAGAC-3’), EF1-r(5’-GTGCCGATACCACCGATCTT-3’) modificati rispetto a quelli riportati da Rehner [13]. Nuovi primers forwarde reverse per i geni che codificano le serina-proteasi alcaline, Pen-f (5’-GCTAGAAATGTTCATGCTCG-3’) ePen-r (5’-TCCAGATAGGGTCTTTGAG-3’), Pen2-f (5’-GGTCTCTCTCGCATCTCT-3’) e Pen2-r (5’-AGTTGGTGAAGGAAGCAC-3’) sono stati basati su regioni conservate di geni fungini delle serina-proteasi alcaline disponibili in GenBank (National Center for Biotechnology Information, NCBI). Unaregione del gene mitocondriale cytb per la citocromo b è stata amplificata mediante i due nuovi primersCytb-f2 (5´-CGTTTGTGATTCACCAC-3’) e Cytb-r2 (5’-GTTTGCATAGGGTTAGC-3’). L’amplificazione delDNA mediante PCR è stata eseguita utilizzando Taq DNA polimerasi (Polymed) con aliquote di ciascunlisato cellulare come DNA genomico. Le condizioni per l’amplificazione genica mediante PCR sono statele seguenti: 2 min a 92-95°C, seguiti da 30-35 cicli, ognuno dei quali costituito dalle seguenti fasi: 95°Cper 20-60 sec, 50°-58°C per 40-50 sec e 72°C per 1-3 min, ed una fase finale di 7-10 min a 72°C. Iframmenti di DNA amplificati sono stati separati tramite elettroforesi su gel d’agarosio esuccessivamente evidenziati con specifici coloranti (SYBR Green). Nel caso in cui i prodotti PCRpresentavano bande non specifiche si è proceduto ad ottimizzare le condizioni di PCR. I prodotti dellaPCR sono stati purificati e successivamente sequenziati (chimica del BigDye terminator v3.1) inentrambe le direzioni utilizzando gli stessi primers di PCR. I prodotti del cycle sequencing sono statianalizzati mediante il sequenziatore di DNA ABI PRISM 3100 (Applied Biosystems).
1.3 Analisi delle sequenze
I nucleotidi ed i residui amminoacidici dedotti sono stati confrontati con dati disponibili in Genbankutilizzando il server BLAST del database NCBI. Gli allineamenti delle sequenze sono stati eseguitiutilizzando il software Clustal X 1.8. Le analisi filogenetiche sono state effettuate utilizzando sia il metodoNeighbor-Joining (NJ) [14] con il modello di Kimura 2P ed includendo sia le transizioni che le transversioniche con il metodo di Massima Parsimonia (MP) [14]. In tutte le analisi filogenetiche, sono state inserite

sequenze di geni nucleari o mitocondriali di funghi estratte da GenBank e sono state calcolate 1.000repliche di bootstrap. Inoltre, l’analisi filogenetica ha riguardato le sequenze amminoacidiche delleserina-proteasi e di proteine omologhe usando come confronto dati provenienti prevalentemente dagenomi sia di Ascomiceti che di Basidiomiceti completamente sequenziati o in fase di completamentodisponibili presso banche dati pubbliche. Le distanze evolutive derivate dagli allineamenti dei residuiamminoacidici dedotti sono state calcolate utilizzando la correzione di Poisson [14]. Le analisifilogenetiche con i metodi NJ e MP sono state eseguite utilizzando il programma MEGA versione 3.1[14]. Inoltre, gli allineamenti delle sequenze amminoacidiche sono stati sottoposti ad analisi dellaMassima Verosimiglianza (ML, Maximum Likelihood) con il modello JTT utilizzando il programmaPHYML versione 2.4.4 [15] con valori di default e 500 repliche di bootstrap.
2. RISULTATI E DISCUSSIONE
2.1 Analisi filogenetica degli isolati fungini
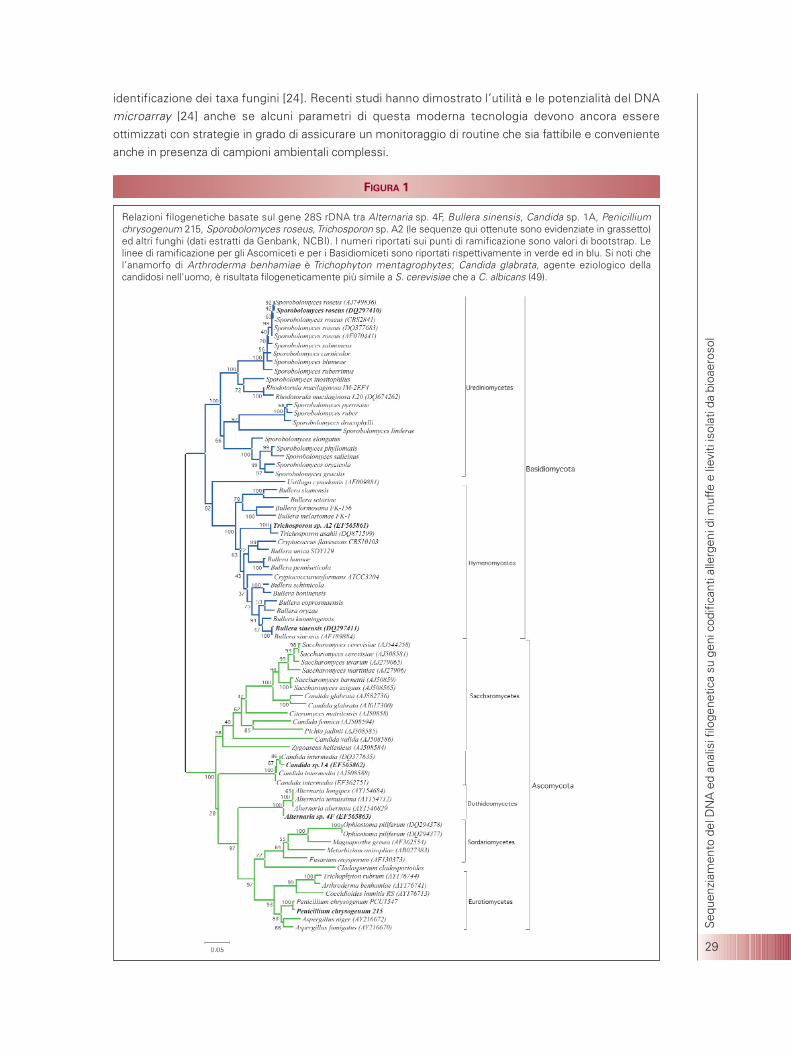
Recenti studi riguardanti contaminazioni di origine microbiologica in ambienti di lavoro sia interni cheesterni hanno evidenziato la necessità di individuare in modo efficace la presenza di funghi aerodispersi[16]. Una conoscenza approfondita della diversità fungina difatti consente una migliore comprensionedel ruolo dei funghi ad esempio in processi allergici [17]. Nel presente studio gli isolati fungini sono stati principalmente identificati attraverso l’analisi filogeneticabasata su sequenze di DNA [1,9,18]. Al fine di dedurre le relazioni evolutive sono stati utilizzati diversimarcatori molecolari, tra i quali il 18S rDNA, gli ITS1 e ITS2, il 5.8S rDNA ed il 28S rDNA; alcunesequenze sono state già depositate in GenBank con i seguenti numeri di accesso: DQ297408;DQ297409; DQ297410; DQ297411; EF565861-EF565863; EF611042-3). Gli alberi evolutivi dedotti da taligeni (una filogenesi basata sul gene 28S rDNA è riportata in Figura 1) hanno evidenziato congruenzanella delineazione dei vari gruppi filogenetici [19,20]. Le muffe appartenenti principalmente ai generiAlternaria (Dothideomycetes), Aspergillus (Eurotiomycetes), Cladosporium (Dothideomycetes incertaesedis) e Penicillium (Eurotiomycetes) (Figura 1) sono state isolate dai campioni di bioaerosol provenientida diversi ambienti (Alternaria e Cladosporium sono risultati prevalentemente diffusi in ambienti esterni).Inoltre, lieviti appartenenti sia agli Ascomiceti che ai Basidiomiceti sono stati individuati in varie fasi diprocessi lavorativi; i lieviti ascomiceti che sono stati isolati più frequentemente appartenevano al genereCandida (C. intermedia e Candida sp.; Saccharomycetes ) (Figura 1) mentre i lieviti basidiomicetiappartenevano ai generi Bullera (B. sinensis; Hymenomycetes), Sporobolomyces (S. roseus;Urediniomycetes) eTrichosporon (Hymenomycetes) (Figura 1). L’analisi morfologica eseguita sui taxa fungini del presente studio ha confermato le descrizioni originali, adesempio per le cellule gemmanti di forma sferica o semisferica (3-4 µm) di B. sinensis e per le cellulegemmanti ellissoidali (3-4 x 5-6 µm) di S. roseus. In particolare, i lieviti basidiomiceti analizzati sono risultatidisposti in clusters (Figura 1) che correlano con specifici caratteri morfologici [21,22]: le specie di Bullera eTrichosporon al contrario delle specie di Sporobolomyces e Rhodotorula (Figura 1) contengono xilosio nellepareti cellulari. Inoltre, i nostri risultati sul gene della citocromo b mitocondriale (una sequenza è statadepositata in GenBank con numero di accesso: EF570138), integrati con dati relativi a sequenze omologhedisponibili presso banche dati pubbliche, hanno confermato che queste regioni mitocondriali contengonoinformazioni filogenetiche sufficienti per dedurre relazioni evolutive all’interno di singoli gruppi [21,22]. Taliindagini molecolari sono di fondamentale importanza per lo studio di specie morfologiche distribuite suun’ampia area che possono comprendere taxa filogenetici geograficamente circoscritti, come riportatorecentemente per Aspergillus fumigatus e per Penicillium chrysogenum [2, 23].Inoltre, sequenze geniche come quelle qui ottenute sono basilari per sviluppare moderni strumentibasati sul DNA, come ad esempio il DNA microarray, per una rapida, accurata e simultanea
28
Prev
enzi
one
Ogg
i - lu
glio
- se
ttem
bre
2007
29
Seq
uenz
iam
ento
del
DN
Aed
ana
lisi f
iloge
netic
a su
gen
i cod
ifica
nti a
llerg
eni d
i muf
fe e
liev
iti is
olat
i da
bioa
eros
ol
identificazione dei taxa fungini [24]. Recenti studi hanno dimostrato l’utilità e le potenzialità del DNAmicroarray [24] anche se alcuni parametri di questa moderna tecnologia devono ancora essereottimizzati con strategie in grado di assicurare un monitoraggio di routine che sia fattibile e convenienteanche in presenza di campioni ambientali complessi.
FIGURA 1
Relazioni filogenetiche basate sul gene 28S rDNA tra Alternaria sp. 4F, Bullera sinensis, Candida sp. 1A, Penicilliumchrysogenum 215, Sporobolomyces roseus, Trichosporon sp. A2 (le sequenze qui ottenute sono evidenziate in grassetto)ed altri funghi (dati estratti da Genbank, NCBI). I numeri riportati sui punti di ramificazione sono valori di bootstrap. Lelinee di ramificazione per gli Ascomiceti e per i Basidiomiceti sono riportati rispettivamente in verde ed in blu. Si noti chel’anamorfo di Arthroderma benhamiae è Trichophyton mentagrophytes; Candida glabrata, agente eziologico dellacandidosi nell'uomo, è risultata filogeneticamente più simile a S. cerevisiae che a C. albicans (49).

2.2 Analisi dei geni fungini che codificano proteine allergiche
Nonostante diverse indagini hanno generalmente permesso di associare disturbi respiratori alla qualitàdell’aria presente in determinati ambienti di lavoro, in realtà le informazioni esistenti riguardantil’esposizioni biologiche associate a funghi allergenici nell’ambito di specifici ambienti di lavoro interni edesterni sono limitate. In particolare, gli studi molecolari sui geni che codificano per le proteine fungineallergeniche sono generalmente scarsi e, di conseguenza, sono limitate le analisi filogenetiche eseguite sutali allergeni, soprattutto se si considerano taxa fungini conspecifici o di livelli tassonomici bassi [25, 26]. Muffe appartenenti ai generi Aspergillus e Penicillium sono state identificate come importanti agentieziologici di allergie [27-32]. Le nostre indagini a livello del DNA sono state condotte principalmente sudiversi campioni appartenenti a P. chrysogenum (conosciuto anche come P. notatum) [23] poiché,nonostante si tratti di una muffa riconosciuta come importante agente eziologico di allergie, gli studicondotti sulla variabilità genetica dei suoi principali allergeni sono limitati. Sebbene P. chrysogenum siaun ascomiceto industrialmente importante (e.g. come produttore di penicillina), gli studi genetici suimportanti vie metaboliche di tale muffa sono tuttora scarsi. Difatti, anche se il genoma di P.chrysogenum è di dimensioni relativamente ridotte (34-38 Mb distribuite su 4 cromosomi) [33], ilsequenziamento dell’intero genoma di tale fungo filamentoso o di altri taxa appartenenti al generePenicillium ad oggi non è stato realizzato. Inoltre, qui sono stati analizzati lieviti basidiomiceti appartenenti ai generi Bullera, Rhodotorula eSporobolomyces (Figura 1) considerando che tali lieviti, anche se generalmente non risultano patogeni,possono rivelarsi importanti allergeni respiratori. Difatti, la potenziale allergenicità di molte speciefungine risulta poco investigata [34]. In particolare, per numerosi funghi, il ruolo di diversi enzimi, tra iquali omologhi della subtilisina, come allergeni o come agenti responsabili di reazioni infiammatorie oallergiche dell’apparato respiratorio umano non è stato ancora definito [35]. Recentemente, Shen et al. [36] hanno dimostrato che due importanti allergeni di P. chrysogenum sono leserina-proteasi vacuolari (gruppo 18, ad esempio Pen ch 18) e le serina-proteasi alcaline (gruppo 13, adesempio Pen ch 13 e Pen n 13). In tale contesto, i nostri studi molecolari si sono indirizzati particolarmentesui geni che codificano per le serina-proteasi alcaline/vacuolari: la sequenza della serina-proteasi alcalinadal ceppo 215 di P. chrysogenum (Figura 1), campionato da polvere su carte, è stata depositata in GenBank(con numero di accesso EF579962) ed è stata inclusa nell’analisi filogenetica riportata nella Figura 2.Basandoci sulla struttura esone-introne, qui determinata attraverso il confronto di sequenze omologhe(introni spliceosomali che presentano delle tipiche sequenze consenso GT e AG alle estremità 5’ e 3’ edappartengono agli introni di fase 0; Figura 3), è stata eseguita un’analisi filogenetica sulle sequenze diamminoacidi delle serina-proteasi alcaline e delle serina-proteasi vacuolari di P. chrysogenum e di altrifunghi. L’analisi filogenetica condotta utilizzando i metodi NJ, MP e ML ha evidenziato la stessa tipologiaevolutiva (Figura 2), in generale confermando precedenti inferenze sull’evoluzione delle serina-proteasi,comprese le relazioni di paralogia e di ortologia [26,37-9]. Considerando che i due phyla degli Ascomiceti edei Basidiomiceti costituiscono due gruppi monofiletici discendenti da unica linea evolutiva (Dikarya; Figura1), è interessante notare che a stadi basali della linea evolutiva degli Ascomiceti si è verificata unasostanziale diversificazione delle serina-proteasi e che più recentemente si sono verificati, in modoindipendente, diversi eventi di duplicazione genica in specifici gruppi di ascomiceti patogeni (Figura 2),come riportato per i generi Metarhizium [37] e Trichophyton [26]. Apparentemente, nella storia evolutivadei Basidiomiceti non si è verificata una equivalente divergenza di geni che codificano per le proteasisecrete (Figura 2). Tuttavia, per chiarire se il livello di divergenza della sequenza di aminoacidi osservato trale serina-proteasi vacuolari dei lieviti basidiomiceti inclusi nel presente studio, quali ad esempioRhodotorula mucilaginosa (nota anche come R. rubra) [40] e di alcuni lieviti ascomiceti (Figura 2), è dovutoa recenti eventi di trasferimento genico orizzontale o se è conseguenza di evoluzione convergente, sononecessari campionamenti ed analisi filogenetiche estesi ad un numero maggiore di taxa. 30
Prev
enzi
one
Ogg
i - lu
glio
- se
ttem
bre
2007

31
Seq
uenz
iam
ento
del
DN
Aed
ana
lisi f
iloge
netic
a su
gen
i cod
ifica
nti a
llerg
eni d
i muf
fe e
liev
iti is
olat
i da
bioa
eros
ol
FIGURA 2
Analisi filogenetica basata sulle sequenze amminoacidiche delle serina-proteasi alcaline di Penicillium chrysogenum 215(in grassetto) e da altri funghi (dati estratti da GenBank, NCBI). Nell’analisi sono state incluse le sequenze di geni checodificano per altre serina-proteasi fungine, comprese le serina-proteasi vacuolari di P. chrysogenum e le sequenze diproteasi alcaline da A. fumigatus (ALP1, ALP2); l'ortologia di SUB2 e delle proteasi alcaline secrete di Aspergillus è stataconfermata dalle analisi sulla struttura esone-introne (26); Metarhizium Pr1H non conteneva sequenze tipiche disecrezione fungina (37). Le ramificazioni filogenetiche per gli Ascomiceti e per i Basidiomiceti sono rispettivamente inverde ed in blu. Si noti: Gibberella zeae PH-1 (anamorfo: Fusarium graminearum), Hypocrea lixii (anamorfo: Trichodermaharzianum) e Ashbya gossypii ATCC 10895 (Eremothecium gossypii ATCC 10895).

Difatti, a tutt’oggi i dati molecolari disponibili sui geni codificanti allergeni per i lieviti basidiomicetirisultano ancora limitati. Una ricerca di geni omologhi delle serina-proteasi alcaline (Ec 3.4.21.-) e delleserina-proteasi vacuolari (EC 3.4.24) nei dati di sequenza attualmente disponibili per le 10 Mbp delgenoma del lievito basidiomiceto S. roseus (S. roseus v1.0; http://genome.jgi-psf.org/Sporo1/Sporo1.home.html) ha evidenziato un’omologia di sequenza con regioni geniche che codificano per proteasidella famiglia delle subtilasi: gli aminoacidi dedotti da una di queste regioni geniche (e_gw1.11.77.1,Sporo1:15367) sono stati inclusi nell’analisi filogenetica riportata in Figura 2. L’esame dei dati disponibilidel genoma di circa 20 Mb del lievito basidiomiceto (e patogeno umano) Cryptococcus neoformans(Hymenomycetes) (Figura 1) [41] ed il completamento di progetti sul sequenziamento di genomi fungini- e.g. attualmente si stanno sequenziando i genomi (tra le 25 e le 90 Mbp) di diversi Urediniomiceti -potranno accrescere le conoscenze sugli allergeni da funghi e su altri fattori di virulenza [26, 41-5]. Tuttavia, ai fini di una migliore comprensione delle relazioni evolutive delle serina-proteasi alcaline frataxa conspecifici, abbiamo rivolto la nostra attenzione su vari campioni di P. chrysogenum raccolti inluoghi diversi. Sulla base di questi risultati, stiamo completando i nostri studi volti a delineare gruppi
32
Prev
enzi
one
Ogg
i - lu
glio
- se
ttem
bre
2007
FIGURA 3
Allineamento delle sequenze degli amminoacidi (i relativi simboli sono colorati nel modo seguente: arancione: GPST;rosso: HKR; blu: FWY; verde: ILMV) dedotti da una regione della serina-proteasi alcalina di Penicillium chrysogenum 215(sottolineato) ed altri funghi. I residui amminoacidici (GHADFGGR) inseriti nel riquadro sono un epitopo di P.chrysogenum. In alto: le barre aperte rappresentano gli introni mentre quelle solide gli esoni; le frecce indicano leposizioni di due (D e H) dei tre residui catalitici (vedi testo). Gli amminoacidi completamente conservati sono segnalati daun asterisco mentre il gap di allineamento è indicato con un trattino.

33
Seq
uenz
iam
ento
del
DN
Aed
ana
lisi f
iloge
netic
a su
gen
i cod
ifica
nti a
llerg
eni d
i muf
fe e
liev
iti is
olat
i da
bioa
eros
ol
filogenetici ed a correlare i genotipi con le località di raccolta. È interessante notare che dalle nostreindagini preliminari per la maggior parte degli isolati fungini emerge, nonostante i livelli alti di variazionenucleotidica, un alto grado di somiglianza tra gli aminoacidi delle serina-proteasi alcaline (Figura 3), laquale suggerisce una conservazione di funzione (Figura 3). Come previsto, inoltre, tali sequenzecontengono aminoacidi conservati e caratteristici del sito catalitico delle serina-proteasi [38,46]costituito dalla triade di aminoacidi (Figura 3) composta da istidina (H), acido aspartico (D) e serina (S).Mediante il programma Cn3D versione 4.1 è stato possibile costruire un modello di strutturatridimensionale della serina-proteasi utilizzando i nostri dati ed altre sequenze delle subtilasi estratte daGenbank (NCBI) (Figura 4). Tali studi di modellistica molecolare si sono dimostrati utili al fine di localizzaregli epitopi di legame alle IgE ed altri importanti residui, tra i quali aminoacidi fondamentali che si sonoconservati nel corso dell’evoluzione [32]. Difatti diversi epitopi di legame alle IgE risultano già mappatinella struttura delle serina-proteasi tramite modellistica molecolare [44]. Ad esempio, il principaleepitopo (GHADFGGR) (Figura 3) di Pen ch13 di P. chrysogenum [43] forma una struttura ad ansa e risultasituato sulla superficie proteica mentre, come emerge dall’analisi delle Figure 2 e 3, la proteina omologadi P. chrysogenum PG 222 [47] in quella regione mostra tre sostituzioni amminoacidiche.Infine, una parte delle nostre ricerche è stata incentrata su determinati geni fungini di lievitibasidiomiceti che codificano per proteine potenzialmente coinvolte in reazioni allergiche che sono tuttorascarsamente conosciute. L’analisi delle sequenze dei genomi completi dei funghi da poco resi pubblici[48,49] risulta essenziale per individuare allergeni non ancora identificati, come recentemente riportatoper proteasi secrete e proteine di superficie del conidio [50,51]. Tutti questi studi molecolari sicuramenteincrementano le conoscenze in materia di esposizione biologica e dei livelli di rischio per malattierespiratorie di origine lavorativa causate da specie fungine aerodisperse potenzialmente allergeniche.
FIGURA 4
A sinistra: coltura di Penicillium chrysogenum di 2/3 giorni su piastra Sabouraud agar. A destra: modellisticatridimensionale della serina-proteasi (vedi testo). La molecola (appartenente alle subtilisine della famiglia S8) è costituitada un dominio alfa/beta contenente eliche alfa e foglietti beta. I radicali (o gruppi R) di ciascun amminoacido sono riportaticome filamenti gialli; l'alfa elica è rappresentata come solenoide verde ed il foglietto beta come freccia marrone-ocra.

3. CONCLUSIONI
Nel presente lavoro è stata evidenziata l’utilità sia del sequenziamento del DNA che degli studifilogenetici per una rapida ed accurata identificazione dei funghi potenzialmente allergenici. Inoltre, lericerche a livello molecolare su allergeni da funghi hanno fornito informazioni che possono risultareessenziali in riferimento al monitoraggio dei livelli di esposizione biologica ed agli interventi diprevenzione per le allergie da funghi. Infine i nostri studi integrati con le informazioni ricavabili dalsequenziamento di genomi fungini risultano di fondamentale importanza per lo sviluppo di metodimolecolari basati sul DNA, ad esempio DNA microarray, che possono essere utilizzati al fine dimonitorare nei funghi in modo simultaneo l’espressione di geni che codificano per allergeni come peraltri fattori di virulenza e di patogenicità.
RINGRAZIAMENTI
Desideriamo ringraziare la Prof.ssa O. Maggi (Università di Roma “La Sapienza”, Dipartimento di BiologiaVegetale) in particolare per i campioni di Penicillium chrysogenum e la Dott.ssa M. Manganelli (ISPESL-DIPIA) per l’assistenza fornita durante l’analisi morfologica al microscopio confocale.
RIFERIMENTI BIBLIOGRAFICI
1. Stetzenbach LD, Buttner MP, Cruz P. Detection and enumeration of airborne biocontaminants. CurrOpin Biotechnol 2004;15:170-4
2. Pringle A, Baker DM, Platt JL, Wares JP, Latge JP, Taylor JW. Cryptic speciation in the cosmopolitanand clonal human pathogenic fungus Aspergillus fumigatus. Evolution Int J Org Evolution2005;59:1886-99
3. Rakeman JL, Bui U, Lafe K, Chen YC, Honeycutt RJ, Cookson BT. Multilocus DNA sequencecomparisons rapidly identify pathogenic molds. J Clin Microbiol 2005;43:3324-33
4. Bai FY, Zhao JH, Takashima M, Jia JH, Boekhout T, Nakase T. Reclassification of the Sporobolomycesroseus and Sporidiobolus pararoseus complexes, with the description of Sporobolomyces phaffii sp.nov. Int J Syst Evol Microbiol 2002;52:2309-14
5. Fell JW, Boekhout T, Fonseca A, Scorzetti G, Statzell-Tallman A. Biodiversity and systematics ofbasidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. IntJ Syst Evol Microbiol 2000;50:1351-71
6. Kurtzman CP, Robnett CJ. Identification and phylogeny of ascomycetous yeasts from analysis ofnuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek1998;73:331-71
7. Wu Z, Wang X, Blomquist G. Evaluation of PCR primers and PCR conditions for specific detection ofcommon airborne fungi. J Environ Monit 2002;4:377-82
8. Wu Z, Tsumura Y, Blomquist G, Wang X. 18S rRNA gene variation among common airborne Fungi,and development of specific oligonucleotide probes for the detection of fungal isolates. ApplEnvironm Microb 2003;69:5389-97
9. Davolos D, Persiani AM, Pietrangeli B, Maggi O. Metarhizium sp., an Antarctic mitosporic fungus(Ascomycota) with a distinct evolutionary lineage based on sequences of the ribosomal DNA andbeta tubulin coding genes. In: BAGECO 9 - 9th Symposium on Bacterial Genetics and Ecology.Wernigerode Germany, 23-27 June 2007. 2007. p.01-2
34
Prev
enzi
one
Ogg
i - lu
glio
- se
ttem
bre
2007

35
Seq
uenz
iam
ento
del
DN
Aed
ana
lisi f
iloge
netic
a su
gen
i cod
ifica
nti a
llerg
eni d
i muf
fe e
liev
iti is
olat
i da
bioa
eros
ol
10. White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNAgenes for phylogenetics. In: Innis MA, Gelfand H, Sninsky JS, White TJ, (eds.) PCR protocols: aguide to methods and applications. New York: Academic Press; 1990. p.315-22
11. O’Donnell K. Fusarium and its near relatives. In: Reynolds DR, Taylor JW, eds. The fungal holomorph:mitotic, meiotic and pleomorphic speciation in fungal systematics. Wallingford, UK: CABInternational; 1993. p.225-33
12. Geiser DM, Frisvad JC, Taylor JW. Evolutionary relationships in Aspergillus section Fumigati inferredfrom partial b-tubulin and hydrophobin DNA sequences. Mycologia 1998;90:831-45
13. Rehner S. Primers for Elongation Factor 1-· (EF1-·) [online] 2001. URL: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf
14. Kumar S, Tamura K, Nei M. MEGA3: Integrated software for Molecular Evolutionary GeneticsAnalysis and sequence alignment. Brief Bioinformatics 2004;5:150-63
15. Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies bymaximum likelihood. Syst Biol 2003;52:696-704
16. Ahmad S, Al-Mahmeed M, Khan ZU. Characterization of Trichosporon species isolated from clinicalspecimens in Kuwait. J Med Microbiol 2005;54:639-46
17. Hageskal G, Knutsen AK, Gaustad P, de Hoog GS, Skaar I. Diversity and significance of mold speciesin Norwegian drinking water. Appl Environ Microbiol 2006;72:7586-93
18. Davolos D, Pietrangeli B. Molecular analysis on the Fungi isolated from bioaerosols. In: AnnualConference of the Association for General and Applied Microbiology 2006 (VAAM). Jena Germany,19-22 March 2006. 2006. p.182
19. Bai FY, Takashima M, Zhao JH, Jia JH, Nakase T. Bullera anomala sp. nov. and Bullerapseudovariabilis sp. nov., two new ballistoconidium-forming yeast species from Yunnan, China.Antonie Van Leeuwenhoek 2003;83:257-63
20. Hamamoto M, Nakase T. Phylogenetic analysis of the ballistoconidium-forming yeast genusSporobolomyces based on 18S rDNA sequences. Int J Syst Evol Microbiol 2000;50:1373-80
21. Biswas SK, Yokoyama K, Nishimura K, Miyaji M. Molecular phylogenetics of the genus Rhodotorulaand related basidiomycetous yeasts inferred from the mitochondrial cytochrome b gene. Int J SystEvol Microbiol 2001;51:1191-9
22. Biswas SK, Wang L, Yokoyama K, Nishimura K. Molecular phylogenetics of the genus Trichosporoninferred from mitochondrial cytochrome b gene sequences. J Clin Microbiol 2005;43:5171-8
23. Scott J, Untereiner WA, Wong B, Straus NA, Malloch D. Genotypic variation in Penicilliumchrysogenum from indoor environments. Mycologia 2004;96:1095-105
24. Leinberger DM, Schumacher U, Autenrieth IB, Bachmann TT. Development of a DNA microarray fordetection and identification of fungal pathogens involved in invasive mycoses. J Clin Microbiol2005;43:4943-53
25. Hong SG, Cramer RA, Lawrence CB, Pryor BM. Alt a 1 allergen homologs from Alternaria andrelated taxa: analysis of phylogenetic content and secondary structure. Fungal Genet Biol2005;42:119-29
26. Jousson O, Lechenne B, Bontems O, Mignon B, Reichard U, Barblan J, Quadroni M, Monod M.Secreted subtilisin gene family in Trichophyton rubrum. Gene 2004;339:79-88
27. Chow LP, Su NY, Yu CJ, Chiang BL, Shen HD. Identification and expression of Pen c 2, a novelallergen from Penicillium citrinum. Biochem J 1999;341:51-9
28. Chow LP, Chiou SH, Hsiao MC, Yu CJ, Chiang BL. Characterization of Pen n 13, a major allergen fromthe mold Penicillium notatum. Biochem Biophys Res Commun 2000;269:14-20

29. Gupta R, Sharma V, Sridhara S, Singh BP, Arora N. Identification of serine protease as a majorallergen of Curvularia lunata. Allergy 2004;59:421-7
30. Su, NY, Yu CJ, Shen HD, Pan FM, Chow LP. Pen c 1, a novel enzymic allergen protein fromPenicillium citrinum. Purification, characterization, cloning and expression. Eur J Biochem1999;261:115-23
31. Yu CJ, Chiou SH, Lai WY, Chiang BL, Chow LP. Characterization of a novel allergen, a majorIgEbinding protein from Aspergillus flavus, as an alkaline serine protease. Biochem Biophys ResCommun 1999;261:669-75
32. Yu CJ, Chen YM, Su SN, Forouhar F, Lee SH, Chow LP. Molecular and immunological characterizationand IgE epitope mapping of Pen n 18, a major allergen of Penicillium notatum. Biochem J2002;363:707-15
33. Xu Z, van den Berg MA, Scheuring C, Covaleda L, Lu H, Santos FA, Uhm T, Lee MK, Wu C, Liu S,Zhang HB. Genome physical mapping from large-insert clones by fingerprint analysis with capillaryelectrophoresis: a robust physical map of Penicillium chrysogenum. Nucleic Acids Res2005;33(5):e50
34. Westwood GS, Huang SW, Keyhani NO. Allergens of the entomopathogenic fungus Beauveriabassiana. Clin Mol Allergy 2005;3:1
35. Spreer A, Rüchel R, Reichard U. Characterization of an extracellular subtilisin protease of Rhizopusmicrosporus and evidence for its expression during invasive rhinoorbital mycosis. Med Mycol2006;44:723-31
36. Shen HD, Chou H, Tam MF, Chang CY, Lai HY, Wang SR. Molecular and immunologicalcharacterization of Pen ch 18, the vacuolar serine protease major allergen of Penicilliumchrysogenum. Allergy 2003;58:993-1002
37. Bagga S, Hu G, Screen SE, St Leger RJ. Reconstructing the diversification of subtilisins in thepathogenic fungus Metarhizium anisopliae. Gene 2004;324:159-69
38. Wang C, Typas MA, Butt TM. Phylogenetic and exon-intron structure analysis of fungal subtilisins:support for a mixed model of intron evolution. J Mol Evol 2005;60:238-46
39. Hu G, Leger RJ. A phylogenomic approach to reconstructing the diversification of serine proteasesin fungi. J Evol Biol 2004;17:1204-14
40. Chou H, Tam MF, Lee SS, Tai HY, Chang CY, Chou CT, Shen HD. A vacuolar serine protease (Rho m 2)is a major allergen of Rhodotorula mucilaginosa and belongs to a class of highly conservedpanfungal allergens. Int Arch Allergy Immunol 2005;138:134-41
41. Loftus BJ, Fung E, Roncaglia P, Rowley D, Amedeo P, Bruno D, et al. The genome of thebasidiomycetous yeast and human pathogen Cryptococcus neoformans. Science 2005;307:1321-24
42. Bowyer P, Denning DW. Genomic analysis of allergen genes in Aspergillus spp: the relevance ofgenomics to everyday research. Med Mycol 2007;45:17-26
43. Lai HY, Tam MF, Chou H, Lee SS, Tai HY, Shen HD. Molecular and structural analysis ofimmunoglobulin E-binding epitopes of Pen ch 13, an alkaline serine protease major allergen fromPenicillium chrysogenum. Clin Exp Allergy 2004;34:1926-33
44. Lee LH, Tam MF, Chou H, Tai HY, Shen HD. Lys, Pro and Trp are critical core amino acid residuesrecognized by FUM20, a monoclonal antibody against serine protease pan-fungal allergens. Int ArchAllergy Immunol 2007;143:194-200
45. Kaur R, Domergue R, Zupancic ML, Cormack BP. A yeast by any other name: Candida glabrata andits interaction with the host. Curr Opin Microbiol 2005;8:378-84
36
Prev
enzi
one
Ogg
i - lu
glio
- se
ttem
bre
2007

37
Seq
uenz
iam
ento
del
DN
Aed
ana
lisi f
iloge
netic
a su
gen
i cod
ifica
nti a
llerg
eni d
i muf
fe e
liev
iti is
olat
i da
bioa
eros
ol
46. Chou H, Lai HY, Tam MF, Chou MY, Wang SR, Han SH, Shen HD. cDNA cloning, biological andimmunological characterization of the alkaline serine protease major allergen from Penicilliumchrysogenum. Int Arch Allergy Immunol 2002;127:15-26
47. Benito MJ, Connerton IF, Cordoba JJ. Genetic characterization and expression of the novel fungalprotease, EPg222 active in dry-cured meat products. Appl Microbiol Biotechnol 2006;73:356-65
48. Galagan, JE, Calvo SE, Cuomo C, Ma LJ, Wortman JR, Batzoglou S. Sequencing of Aspergillusnidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 2005;438:1105-15
49. Dujon B, Sherman D, Fischer G, Durrens P, Casaregola S, Lafontaine I, et al. Genome evolution inyeasts. Nature 2004;430:35-44
50. Asif AR, Oellerich M, Amstrong VW, Riemenschneider B, Monod M, Reichard U. Proteome ofconidial surface associated proteins of Aspergillus fumigatus reflecting potential vaccine candidatesand allergens J Proteome Res 2006;5:954-62
51. Nierman WC, Pain A, Anderson MJ, Wortman JR, Kim HS, Arroyo J, et al. Genomic sequence of thepathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005;38:1151-6
Copyright © 2022 FDOKUMEN




















