
Sequence variation of ookinete surface proteins Pvs25 and Pvs28 of Plasmodium vivax isolates from...
10
Sequence variation of ookinete surface proteins Pvs25 and Pvs28 of Plasmodium vivax isolates from Southern Mexico and their association to local anophelines infectivity Lilia Gonza ´ lez-Cero ´n a, *, Alejandro Alvarado-Delgado b , Jesus Martı ´nez-Barnetche b , Mario H. Rodrı ´guez c , Marbella Ovilla-Mun ˜oz a , Fabia ´n Pe ´ rez a , Juan E. Hernandez-Avila d , Marco A. Sandoval a , Maria del Carmen Rodrı ´guez b , Cuauhte ´ moc Villarreal-Trevin ˜o a a Centro Regional de Investigacio ´n en Salud Pu ´blica, Instituto Nacional de Salud Pu ´blica, 4a Av. Norte and 19 calle Poniente, Tapachula, Chiapas, Mexico b Centro de Investigacio ´n en Enfermedades Infecciosas; Cuernavaca, Morelos, Mexico c Direccio ´n General, Cuernavaca, Morelos, Mexico d Direccio ´n de Informa ´tica: Instituto Nacional de Salud Pu ´blica, Av. Universidad 655, Cuernavaca, Morelos, Mexico 1. Introduction Plasmodium parasites are transmitted to their vertebrate host and mosquito vectors during the insect blood feeding. Mosquitoes obtain parasite sexual forms circulating in the blood of the vertebrate host, and after fecundation within the insect midgut, invasive forms (ookinetes) migrate to the external midgut surface, where sporozoites develop within oocysts. The sporozoites invade the mosquito salivary glands and are injected into new vertebrate hosts during the subsequent blood meals (Sinden, 1984). Several molecules that mediate the interactions of the different parasites stages with the mosquito organs have been identified (Vlachou et al., 2006); however, the molecular bases of the differences in infectivity of Plasmodia to different mosquito species remain unknown. The infectivity (measured by oocyst counts) of Southern Mexican Plasmodium vivax isolates to local Anopheles albimanus and Anopheles pseudopunctipennis was associated to the repeat central region sequence (CSPr) phenotypes (CSPrVk210 and Vk247) of their circumsporozoite proteins (Gonza ´ lez-Cero ´n Infection, Genetics and Evolution 10 (2010) 645–654 ARTICLE INFO Article history: Received 14 April 2009 Received in revised form 24 March 2010 Accepted 25 March 2010 Available online 2 April 2010 Keywords: Plasmodium vivax Pvs25 Pvs28 Ookinete surface molecules CSPr Polymorphism Haplotype Anopheles albimanus Anophelespseudopunctipennis ABSTRACT The polymorphism of Pvs25 and Pvs28 ookinete surface proteins, their association to circumsporozoite protein repeat (CSPr) genotypes (Vk210 and Vk247) and their infectivity to local Anopheles albimanus and Anopheles pseudopunctipennis were investigated in Plasmodium vivax-infected blood samples obtained from patients in Southern Mexico. The pvs25 and pvs28 complete genes were amplified, cloned and sequenced; and the CSPr genotype was determined by PCR amplification and hybridization. The amino acid Pvs25 and Pvs28 polymorphisms were mapped to their corresponding protein structure. Infected blood samples were simultaneously provided through artificial feeders to both mosquito species; the ratio of infected mosquitoes and oocyst numbers were recorded. The polymorphism of pvs25 and pvs28 was limited to few nucleotide positions, and produced three haplotypes: type A/A parasites presented Pvs25 and Pvs28 amino acid sequences identical to that of Sal I reference strain; parasites type B1 presented a mutation 130 Ile ! Thr in Pvs25, while type B2 presented 87 Gln ! Lys/130 Ile ! Thr in the same molecule. Both types B1 and B2 parasites presented changes in Pvs28 at 87 Asn ! Asp, 110 Tyr ! Asn and five GSGGE/D repeat sequences between the fourth EGF-like domain and the GPI. Most P. vivax parasites from the coastal plains and the overlapping region were Pvs25/28 A/A, CSPrVk210 and were infective only to An. albimanus (p 0.0001). Parasites originating in foothills were Pvs25/28 type B1/B or B2/B and CSPrVk210 or Vk247, and were more infective to An. pseudopunctipennis than to An. albimanus (p 0.001). These results and the analysis of Pvs25/28 from other parts of the world indicated that non-synonymous variations in these proteins occur in amino acid residues exposed on the surface of the proteins, and are likely to interact with midgut mosquito ligands. We hypothesize that these molecules have been shaped by co-evolutionary adaptations of parasites to their susceptible vectors. ß 2010 Elsevier B.V. All rights reserved. * Corresponding author at: ParasitologyDepartment, 4a Av. Norte y 19 calle Poniente, Tapachula, Chiapas 30700, Me ´ xico. Tel.: +52 962 6262219x112; fax: +52 962 6265782. E-mail address: [email protected] (L. Gonza ´ lez-Cero ´ n). Contents lists available at ScienceDirect Infection, Genetics and Evolution journal homepage: www.elsevier.com/locate/meegid 1567-1348/$ – see front matter ß 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.meegid.2010.03.014
Transcript of Sequence variation of ookinete surface proteins Pvs25 and Pvs28 of Plasmodium vivax isolates from...

Infection, Genetics and Evolution 10 (2010) 645–654
Sequence variation of ookinete surface proteins Pvs25 and Pvs28 ofPlasmodium vivax isolates from Southern Mexico and their associationto local anophelines infectivity
Lilia Gonzalez-Ceron a,*, Alejandro Alvarado-Delgado b, Jesus Martınez-Barnetche b,Mario H. Rodrıguez c, Marbella Ovilla-Munoz a, Fabian Perez a, Juan E. Hernandez-Avila d,Marco A. Sandoval a, Maria del Carmen Rodrıguez b, Cuauhtemoc Villarreal-Trevino a
a Centro Regional de Investigacion en Salud Publica, Instituto Nacional de Salud Publica, 4a Av. Norte and 19 calle Poniente, Tapachula, Chiapas, Mexicob Centro de Investigacion en Enfermedades Infecciosas; Cuernavaca, Morelos, Mexicoc Direccion General, Cuernavaca, Morelos, Mexicod Direccion de Informatica: Instituto Nacional de Salud Publica, Av. Universidad 655, Cuernavaca, Morelos, Mexico
A R T I C L E I N F O
Article history:
Received 14 April 2009
Received in revised form 24 March 2010
Accepted 25 March 2010
Available online 2 April 2010
Keywords:
Plasmodium vivax
Pvs25
Pvs28
Ookinete surface molecules
CSPr
Polymorphism
Haplotype
Anopheles albimanus
Anophelespseudopunctipennis
A B S T R A C T
The polymorphism of Pvs25 and Pvs28 ookinete surface proteins, their association to circumsporozoite
protein repeat (CSPr) genotypes (Vk210 and Vk247) and their infectivity to local Anopheles albimanus and
Anopheles pseudopunctipennis were investigated in Plasmodium vivax-infected blood samples obtained
from patients in Southern Mexico. The pvs25 and pvs28 complete genes were amplified, cloned and
sequenced; and the CSPr genotype was determined by PCR amplification and hybridization. The amino
acid Pvs25 and Pvs28 polymorphisms were mapped to their corresponding protein structure. Infected
blood samples were simultaneously provided through artificial feeders to both mosquito species; the
ratio of infected mosquitoes and oocyst numbers were recorded. The polymorphism of pvs25 and pvs28
was limited to few nucleotide positions, and produced three haplotypes: type A/A parasites presented
Pvs25 and Pvs28 amino acid sequences identical to that of Sal I reference strain; parasites type B1
presented a mutation 130 Ile! Thr in Pvs25, while type B2 presented 87 Gln! Lys/130 Ile! Thr in the
same molecule. Both types B1 and B2 parasites presented changes in Pvs28 at 87 Asn! Asp, 110
Tyr! Asn and five GSGGE/D repeat sequences between the fourth EGF-like domain and the GPI. Most P.
vivax parasites from the coastal plains and the overlapping region were Pvs25/28 A/A, CSPrVk210 and
were infective only to An. albimanus (p � 0.0001). Parasites originating in foothills were Pvs25/28 type
B1/B or B2/B and CSPrVk210 or Vk247, and were more infective to An. pseudopunctipennis than to An.
albimanus (p � 0.001). These results and the analysis of Pvs25/28 from other parts of the world indicated
that non-synonymous variations in these proteins occur in amino acid residues exposed on the surface of
the proteins, and are likely to interact with midgut mosquito ligands. We hypothesize that these
molecules have been shaped by co-evolutionary adaptations of parasites to their susceptible vectors.
� 2010 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsev ier .com/ locate /meegid
1. Introduction
Plasmodium parasites are transmitted to their vertebrate hostand mosquito vectors during the insect blood feeding. Mosquitoesobtain parasite sexual forms circulating in the blood of thevertebrate host, and after fecundation within the insect midgut,invasive forms (ookinetes) migrate to the external midgut surface,
* Corresponding author at: ParasitologyDepartment, 4a Av. Norte y 19 calle
Poniente, Tapachula, Chiapas 30700, Mexico. Tel.: +52 962 6262219x112;
fax: +52 962 6265782.
E-mail address: [email protected] (L. Gonzalez-Ceron).
1567-1348/$ – see front matter � 2010 Elsevier B.V. All rights reserved.
doi:10.1016/j.meegid.2010.03.014
where sporozoites develop within oocysts. The sporozoites invadethe mosquito salivary glands and are injected into new vertebratehosts during the subsequent blood meals (Sinden, 1984). Severalmolecules that mediate the interactions of the different parasitesstages with the mosquito organs have been identified (Vlachouet al., 2006); however, the molecular bases of the differences ininfectivity of Plasmodia to different mosquito species remainunknown.
The infectivity (measured by oocyst counts) of SouthernMexican Plasmodium vivax isolates to local Anopheles albimanus
and Anopheles pseudopunctipennis was associated to the repeatcentral region sequence (CSPr) phenotypes (CSPrVk210 andVk247) of their circumsporozoite proteins (Gonzalez-Ceron

L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654646
et al., 1999). The geographic distribution of these CSPr phenotypesfollowed that of the two vectors, An. albimanus on the coast,abundant during the late rainy season and An. pseudopunctipennis
in the foothills, abundant during the dry and the early rainyseasons (Rodriguez et al., 2000). However, as CSP is expressed afterthe successful implantation of the parasites on the externalmosquito midgut surface (Menard et al., 1997; Gonzalez-Ceronet al., 1998), other proteins expressed on invasive ookinetes,probably associated to the CSPr phenotypes, are most likelyresponsible for the differences in parasite infectivity of the twomosquito species.
Plasmodium P25 and P28 proteins are abundant on the ookinetesurface during parasite development in the mosquito blood mealbolus (Kaslow et al., 1988) and elicit transmission-blockingantibodies (Kaslow, 1996; Hisaeda et al., 2000; Arevalo-Herreraet al., 2005; Collins et al., 2006). Members of the P25/28 familyhave been detected in Plasmodium falciparum (Pfs25) (Kaslow et al.,1988); (Pfs28) (Duffy and Kaslow, 1997); Plasmodium reichenowi
(Prs25) (Lal et al., 1990), Plasmodium yoelii (Pys21/25) (Tsuboiet al., 1997a), Plasmodium berghei (Pbs21) (Paton et al., 1993);(Pbs25) (Tsuboi et al., 1997b), Plasmodium gallinaceum (Pgs25)(Kumar and Carter, 1985; Kaslow et al., 1989); (Pgs28) (Duffy et al.,1993), Plasmodium ovale (Pos28) (Tachibana et al., 2001) andPlasmodium vivax (Pvs25/28) (Tsuboi et al., 1998).
P25 proteins are composed of four Epithelial Growth Factor(EGF)-like domains, while P28 members are composed of threecomplete and one incomplete EGF-like domain (Kaslow et al.,1989; Tsuboi et al., 1998). Both proteins participate in theinteraction with midgut molecules (Tomas et al., 2001), and canactivate mosquito molecular immune responses (Danielli et al.,2005; Baton and Ranford-Cartwright, 2005). Compared to the P.
vivax Sal I reference strain, polymorphism of Pvs25 in Asianisolates is limited to few nucleotides and amino acid positions anda slightly larger polymorphism has been reported for Pvs28 (Tsuboiet al., 1998, 2004; Sattabongkot et al., 2003; Zakeri et al., 2009).Reported pvs25 gene sequences in parasites from Central and SouthAmerica were similar to that of the Salvador I strain (Arevalo-Herrera et al., 2005; Escalante et al., 2005), but to date, no data onthe Pvs28 sequence of parasites from the region has been reported.
In this study, the Pvs25/28 gene polymorphisms, theirassociation to CSPr phenotypes and infectivity to An. albimanus
and An. pseudopunctipennis of P. vivax isolates from SouthernMexico were investigated.
2. Materials and methods
2.1. P. vivax parasites
The project was approved by the Ethics Committee of theNational Institute of Public Health (INSP), Mexico. Infected bloodsamples were obtained from symptomatic patients diagnosed withP. vivax using Giemsa-stained blood smear examination, at theRegional Center for Public Health Research (CRISP-INSP) inSouthern Chiapas, Mexico between 2000 and 2005. After informedwritten consent was obtained, patients donated 5 ml of venousblood. In accordance with the Mexican Malaria Treatment Guide-lines (NOM-032-SSA2-2002) all patients received anti-malarialtreatment whether or not they agreed to give a blood sample(http://www.salud.gob.mx/unidades/cdi/nom/032ssa202.html;WHO, 1967). The sexual-stage parasite density per 200 leukocyteswas counted and expressed as the number of gametocytes permicroliter, assuming a leukocyte count of 7000/ml. The patient’splace of residence was recorded, and classified as originating fromthe coastal plains if they lived at �150 m above sea level (m.a.s.l.),and from the foothills if they lived from 250 m up to 1200 m.a.s.l.Tapachula City (150–250 m.a.s.l.) was considered within an
overlapping region. A total of 64 P. vivax-infected blood sampleswere collected; CSPr and pvs25 nucleotide sequences wereinvestigated in all samples, and pvs28 nucleotide sequence andinfectivity to anopheline mosquitoes was investigated andobtained in 20 of these samples. P. vivax DNA from infected bloodwas obtained using the QIAamp1 DNA Blood Mini kit, according tothe manufacturer’s instructions.
2.2. Circumsporozoite protein repeat (CSPr) genotyping
The method described by Kain et al. (1992) was used todetermine the CSPr phenotype in P. vivax-infected blood samples.Using a nested PCR, DNA from each sample was used as a templateto amplify the complete repeated central region of the CS protein.Specific oligonucleotides for the CSP gene: Pv5A (50-GAAAA-TAAGCTGAAACAACC-30) and Pv6A (50-TCCACAGGTTACACTGCAT-30) for the first amplification round, and primers Pv5A and Pv9 (50-GCCAACGGTAGCTCTAAC TTT-30) for the second amplification wereused. The PCR products were fixed to a Nylon membrane using aslot blot apparatus (Hoefer, Pharmacia Biotech, CA) and hybridizedwith fluorescein-labeled Vk210 and Vk247 specific probes,followed by detection with an anti-fluorescein antibody-HRPusing the ECLTM 30-oligolabeling and detection systems (Amer-shamInt, England). The reaction was registered by exposure ontolight sensitivity plates (Amershamplc, UK LTD) using chemilumi-nescence reagents (Amersham Int., England).
2.3. PCR amplification of pvs25 and pvs28 genes
To amplify pvs25 coding region, a mixture of 100 ng of DNA,2 mmol of each oligonucleotide: (PV25F: 50-GGA TCC CTG ACT TTCGTT TCA CA-30), and (A25R: 50-GAA TTC ACA TCA CAA GTC GGT A-30), 1� buffer (100 mM KCl, 20 mM Tris–HCl pH 7.5), 1.5 mmolMgCl2, 125 mmol of each deoxynucleotide triphosphate and 1.5 Uof expand polymerase (Expand High Fidelity PLUS PCR System,Roche, Penzberg) were used.
Reaction conditions for PCR were performed as follows: 94 8Cfor 5 min, followed by 35 cycles of 94 8C for 30 s, 50 8C for 30 s,72 8C for 1.20 min; held at 72 8C for 10 min. To incorporate adenineat the ends of the fragments, 2 U of Taq DNA polymerase(Invitrogen, CA) were added and the reaction incubated for15 min at 72 8C. From each sample, a 1007 bp fragment containingthe pvs25 ORF (657 bp, 219 amino acids) was amplified.
For pvs28 gene, the oligonucleotides flanking the coding regionof Salvador I strain (pvs28-F, 50-ACC CAA CTG AGC TCG ATT CC-30)and (pvs28-R, 50-CTG TGA GAC GCT TCT CCC CT-30) (accessionnumber PlasmoDB, Pv111180) were used. The PCR reaction used0.4 mmol of each oligonucleotide, 0.5 mmol dNTPs, 1� buffer(100 mmol KCl, 20 mmol Tris–HCl pH 7.5), 2 mmol MgSO4 for 2.5 Uof PlatinumR Taq DNA polymerase (PlatinumR Taq DNA polymeraseHigh Fidelity, Invitrogen, CA) and 100 ng of DNA.
The reaction conditions were performed as follows: 95 8C for5 min, followed by 35 cycles of 95 8C for 30 s, 60 8C for 30 s, 72 8C for60 s; held at 72 8C for 5 min. From each sample an 817 bp fragmentcontaining the pvs28 ORF (711 bp, 237 amino acids) was amplified.
The pvs25 gene PCR products were purified using the ConcertTM
Rapid PCR Purification System (Life Technologies Inc., MD)according to the manufacturer’s instructions. Sequencing of thesePCR products from 46 samples produced a good qualityelectropherogram and no cloning was required, while the other18 parasite samples were cloned before sequencing, using acloning kit (TOPO TA Cloning1, Invitrogen, CA) according to themanufacturer’s instructions. The plasmids were extracted using acommercial kit (Wizard1 Plus Minipreps DNA Purification System.Promega, USA). The 20 amplified pvs28 genes were purified andcloned as above.

L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654 647
2.4. Nucleotide sequencing and sequence analysis
The PCR products and plasmid clones were sequenced fromboth strands using PV25-F and A25-R primers for pvs25 and PV28-Fand PV28-R for pvs28 in an automatic sequencer ABI 3100 GeneticAnalyzer (Applied Biosystems, Ambion, Inc. USA). The quality ofthe electropherogram of each DNA sequence was analyzed withboth Phred-Phrap-Consed (Ewing et al., 1998) and by manualinspection. The obtained nucleotide sequences were edited andaligned, using the gene [GenBank; AAKM01002769.1] and thededuced amino acid sequences [GenBank; XP_001608460.1] forPvs25 and [GenBank; XP_001608461.1] for Pvs28, using the VectorNTI Suite software, InforMax, Inc., MD. The polymorphic regionshad Phred scores above 30 (Gordon et al., 1998).
2.5. Nucleotide sequence accession number
The complete gene sequences reported here have beensubmitted in the data base [GenBank; EU024409 to EU024472]for pvs25 and [GenBank; EU514769 to EU514785, EU514789,GU815237 and GU815238] for pvs28.
2.6. Structural analysis of Pvs25 and Pvs28 amino acid polymorphism
To gain further insight into the effect of amino acid variations inthe Pvs25 and Pvs28 structures and how these variations could affectthe interaction of Plasmodium parasites with mosquito midgutmolecules, the variants reported for P. vivax throughout the worldand those obtained in this study were mapped into a previouslyreported native Pvs25 crystal structure (Saxena et al., 2006). Thecrystallographic model of the native Pvs25 protein (PDB:1Z27) wasdownloaded from the Protein Data Bank (PDB; http://www.wwpdb.org/) and visualized in the DeepView/Swiss-PDBviewer (Guex and Peitsch, 1997). A Pvs25 model was generatedwith the mutant amino acid residues as input for homologymodeling using the First Approach mode at the Swiss-Model server(Schwede et al., 2003). The same approach was used to model Sal Itype Pvs28 and the corresponding mutant residues. In Pvs28modeling, the C-terminus (residues 173–236) including the repeatregion, was excluded due to the lack of homology with Pvs25.
2.7. Mosquitoes
An. albimanus white stripped strain and An. pseudopunctipennis
Tapachula colonies from the insectary at RCRPH-INSP were used(Villarreal et al., 1998).
2.8. Parasite infectivity to mosquitoes
The infectivity to An. albimanus and An. pseudopunctipennis ofparasites carrying distinct Pvs25/28-CSPr haplotypes was investi-gated in the 20 blood samples developing oocyst in any mosquitospecies. The plasma of fresh P. vivax infected blood samples wasreplaced (to a 40% hematocrit) by a same blood type non-immuneserum and each blood sample was offered simultaneously tobatches of An. albimanus and An. pseudopunctipennis females usingartificial feeders (Gonzalez-Ceron et al., 1999). Fed mosquitoeswere kept with 5% sugar solution in impregnated cotton pads at25–27 8C and 70–80% humidity. Six to seven days post-infection,midguts were inspected for oocysts and the number of infectedmosquitoes (based on the presence of at least one oocyst); theoocyst numbers per infected mosquito were recorded (Gonzalez-Ceron et al., 1999). Parasite isolates producing infection in eitherAn. albimanus or An. pseudopuntipennis were analyzed to identifythe CSPr genotype and to determine the pvs25 and pvs28
sequences.
2.9. Statistical analysis
A X2 test was used to compare the relation of each Pvs25phenotype with either CSPrVk210 or Vk247 phenotypes. Themedian and intercuartilar range of oocyst counts was calculatedper mosquito species and per Pvs25/28–CSPr haplotype. Onlythose haplotypes having at least two feeding experiments wereincluded in the analysis. A logistic model was used to compare theproportion of infected mosquitoes fed with P. vivax for each Pvs25/28–CSPr haplotype-infected blood. A negative binomial regres-sion model was used to compare oocyst counts in both mosquitospecies infected with different haplotypes. Both the logisticregression and the negative binomial regression models aregeneralized linear models (McCullag and Nelder, 1989), as theirname suggests, this type of models are generalizations of thelinear regression model that allows data such as the proportion ofinfection (logistic model) or the oocyst count per mosquito(negative binomial model), which do not have a normaldistribution, to be modeled. To adjust for the differences in thegametocyte counts in the blood used to feed the mosquitoes in thedifferent experiments, a variable which measures the number ofgametocytes in each blood batch was included in all statisticalmodels used to analyze our data. All statistical analyses werecarried out at the 95% significance level using Stata 9.2 (StataCorporation LP).
To determine linkage equilibrium for Pvs25, Pvs28 and CSPrvariant alleles of all isolates and within parasites isolates of thefoothill region we used the software LIAN V 3.5 at http://gump.auburn.edu/cgi-bin/lian/lian.cgi.pl (Haubold and Hudson,2000). Estimation of p value in case of Monte Carlo (PMC)simulation was performed with 100,000 re-samplings.
3. Results
3.1. Determination of P. vivax Pvs25/28-CSPr haplotypes and its
geographic origin
3.1.1. Pvs25 polymorphism
The pvs25 nucleotide sequences of 12 P. vivax isolates (18.75%)were identical to that of the Sal I strain sequence (type A). Genepolymorphism in the other 52 isolates was limited to threenucleotide positions: A! G at position 78, A! C at position 259,and T! C at position 389. The polymorphism observed at position78 was present in only one isolate and was synonymous, while theother two substitutions were non-synonymous. The transition atposition 389 changed Ile (codon ATC) to Thr (codon ACC) at aminoacid position 130 (EGF-like domain III) in 81.25% (52 out of 64) ofthe isolates, and 42.3% (22 of 52) of these parasites also showed thenucleotide substitution at position 259 changed Gln (codon CAG)to Lys (codon AAG) at amino acid position 87 (EGF-like domain II).These non-synonymous changes produced three distinctivephenotypes in the samples analyzed: 87 Gln/130 Ile (type A; asSal I strain), 87 Gln/130 Thr (type B1) and 87 Lys/130 Thr (type B2)(Table 1). It would be expected that random recombinationbetween parasite populations in the area would generate fourhaplotypes between codons 87 and 130. However, there was asignificant association between A, B1 and B2 alleles and theabsence of the 87 Lys/130 Ile, as assessed by Fisher’s Exact/Chisquared analysis (p � 0.005). The three Pvs25 alleles identifiedhere were in the 20-parasite isolates employed in mosquitofeeding experiments.
3.1.2. Pvs28 polymorphism
No synonymous nucleotide changes were detected. Out of the20 sequenced isolates, seven that were pvs25 Sal I type A also hadpvs28 nucleotide sequences identical to that reported for the Sal I
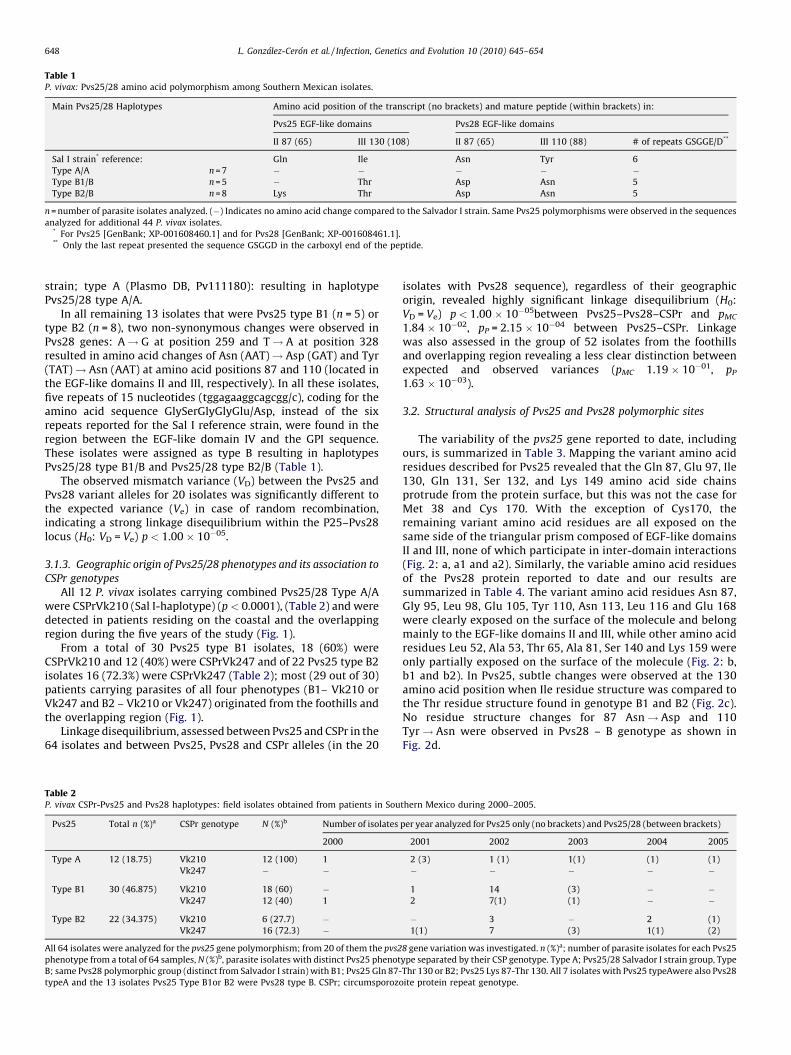
Table 1P. vivax: Pvs25/28 amino acid polymorphism among Southern Mexican isolates.
Main Pvs25/28 Haplotypes Amino acid position of the transcript (no brackets) and mature peptide (within brackets) in:
Pvs25 EGF-like domains Pvs28 EGF-like domains
II 87 (65) III 130 (108) II 87 (65) III 110 (88) # of repeats GSGGE/D**
Sal I strain* reference: Gln Ile Asn Tyr 6
Type A/A n = 7 � � � � �Type B1/B n = 5 � Thr Asp Asn 5
Type B2/B n = 8 Lys Thr Asp Asn 5
n = number of parasite isolates analyzed. (�) Indicates no amino acid change compared to the Salvador I strain. Same Pvs25 polymorphisms were observed in the sequences
analyzed for additional 44 P. vivax isolates.* For Pvs25 [GenBank; XP-001608460.1] and for Pvs28 [GenBank; XP-001608461.1].** Only the last repeat presented the sequence GSGGD in the carboxyl end of the peptide.
L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654648
strain; type A (Plasmo DB, Pv111180): resulting in haplotypePvs25/28 type A/A.
In all remaining 13 isolates that were Pvs25 type B1 (n = 5) ortype B2 (n = 8), two non-synonymous changes were observed inPvs28 genes: A! G at position 259 and T! A at position 328resulted in amino acid changes of Asn (AAT)! Asp (GAT) and Tyr(TAT)! Asn (AAT) at amino acid positions 87 and 110 (located inthe EGF-like domains II and III, respectively). In all these isolates,five repeats of 15 nucleotides (tggagaaggcagcgg/c), coding for theamino acid sequence GlySerGlyGlyGlu/Asp, instead of the sixrepeats reported for the Sal I reference strain, were found in theregion between the EGF-like domain IV and the GPI sequence.These isolates were assigned as type B resulting in haplotypesPvs25/28 type B1/B and Pvs25/28 type B2/B (Table 1).
The observed mismatch variance (VD) between the Pvs25 andPvs28 variant alleles for 20 isolates was significantly different tothe expected variance (Ve) in case of random recombination,indicating a strong linkage disequilibrium within the P25–Pvs28locus (H0: VD = Ve) p < 1.00 � 10�05.
3.1.3. Geographic origin of Pvs25/28 phenotypes and its association to
CSPr genotypes
All 12 P. vivax isolates carrying combined Pvs25/28 Type A/Awere CSPrVk210 (Sal I-haplotype) (p < 0.0001), (Table 2) and weredetected in patients residing on the coastal and the overlappingregion during the five years of the study (Fig. 1).
From a total of 30 Pvs25 type B1 isolates, 18 (60%) wereCSPrVk210 and 12 (40%) were CSPrVk247 and of 22 Pvs25 type B2isolates 16 (72.3%) were CSPrVk247 (Table 2); most (29 out of 30)patients carrying parasites of all four phenotypes (B1– Vk210 orVk247 and B2 – Vk210 or Vk247) originated from the foothills andthe overlapping region (Fig. 1).
Linkage disequilibrium, assessed between Pvs25 and CSPr in the64 isolates and between Pvs25, Pvs28 and CSPr alleles (in the 20
Table 2P. vivax CSPr-Pvs25 and Pvs28 haplotypes: field isolates obtained from patients in Sou
Pvs25 Total n (%)a CSPr genotype N (%)b Number of isolates
2000
Type A 12 (18.75) Vk210 12 (100) 1
Vk247 � �
Type B1 30 (46.875) Vk210 18 (60) �Vk247 12 (40) 1
Type B2 22 (34.375) Vk210 6 (27.7) �Vk247 16 (72.3) �
All 64 isolates were analyzed for the pvs25 gene polymorphism; from 20 of them the pvs2
phenotype from a total of 64 samples, N (%)b, parasite isolates with distinct Pvs25 phenot
B; same Pvs28 polymorphic group (distinct from Salvador I strain) with B1; Pvs25 Gln 87-
typeA and the 13 isolates Pvs25 Type B1or B2 were Pvs28 type B. CSPr; circumsporozo
isolates with Pvs28 sequence), regardless of their geographicorigin, revealed highly significant linkage disequilibrium (H0:VD = Ve) p < 1.00 � 10�05between Pvs25–Pvs28–CSPr and pMC
1.84 � 10�02, pP = 2.15 � 10�04 between Pvs25–CSPr. Linkagewas also assessed in the group of 52 isolates from the foothillsand overlapping region revealing a less clear distinction betweenexpected and observed variances (pMC 1.19 � 10�01, pP
1.63 � 10�03).
3.2. Structural analysis of Pvs25 and Pvs28 polymorphic sites
The variability of the pvs25 gene reported to date, includingours, is summarized in Table 3. Mapping the variant amino acidresidues described for Pvs25 revealed that the Gln 87, Glu 97, Ile130, Gln 131, Ser 132, and Lys 149 amino acid side chainsprotrude from the protein surface, but this was not the case forMet 38 and Cys 170. With the exception of Cys170, theremaining variant amino acid residues are all exposed on thesame side of the triangular prism composed of EGF-like domainsII and III, none of which participate in inter-domain interactions(Fig. 2: a, a1 and a2). Similarly, the variable amino acid residuesof the Pvs28 protein reported to date and our results aresummarized in Table 4. The variant amino acid residues Asn 87,Gly 95, Leu 98, Glu 105, Tyr 110, Asn 113, Leu 116 and Glu 168were clearly exposed on the surface of the molecule and belongmainly to the EGF-like domains II and III, while other amino acidresidues Leu 52, Ala 53, Thr 65, Ala 81, Ser 140 and Lys 159 wereonly partially exposed on the surface of the molecule (Fig. 2: b,b1 and b2). In Pvs25, subtle changes were observed at the 130amino acid position when Ile residue structure was compared tothe Thr residue structure found in genotype B1 and B2 (Fig. 2c).No residue structure changes for 87 Asn! Asp and 110Tyr! Asn were observed in Pvs28 – B genotype as shown inFig. 2d.
thern Mexico during 2000–2005.
per year analyzed for Pvs25 only (no brackets) and Pvs25/28 (between brackets)
2001 2002 2003 2004 2005
2 (3) 1 (1) 1(1) (1) (1)
� � � � �
1 14 (3) � �2 7(1) (1) � �
� 3 � 2 (1)
1(1) 7 (3) 1(1) (2)
8 gene variation was investigated. n (%)a; number of parasite isolates for each Pvs25
ype separated by their CSP genotype. Type A; Pvs25/28 Salvador I strain group, Type
Thr 130 or B2; Pvs25 Lys 87-Thr 130. All 7 isolates with Pvs25 typeAwere also Pvs28
ite protein repeat genotype.

Fig. 1. Spatial distribution of the P. vivax Pvs25/28–CSPr haplotypes detected in
Southern Mexico during 2000–2005: a) this map shows the study region as
indicated by the arrow. b) In here, the origin of all 64 patients carrying parasites
with Pvs25–CSPr haplotypes is shown as indicated with the symbols. Those
symbols with asterisk ( ) identify the 20 out of 64 isolates with complete Pvs25/28–
CSPr haplotype and mosquito infectivity results: parasites with Pvs25-type A were
Pvs28-type A resembling Sal I sequences, and parasites with Pvs25-B1 (QT) or
Pvs25-B2 (KT) all were Pvs28–B.
L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654 649
3.3. Association of the Pvs25/28–CSPr haplotypes to mosquito
infectivity
The results regarding infectivity to An. albimanus and An.
pseudopunctipennis by parasites carrying different Pvs25/28–CSPrhaplotypes are summarized in Table 5 and Supplementary Table 1.Parasite lots A/A-Vk210 (five feeding experiments) consistentlyproduced high proportions (0.74) of infected An. albimanus withhigh mean oocyst densities (138, IQR 5–25), while none of the An.
pseudopunctipennis fed with the same blood samples wereinfected. Similar results were obtained from two more feedingexperiments (80% infected An. albimanus and mean oocystdensities of 84 IQR 55–110 and 92.5 IQR 64.5–130, respectively),however no An. pseudopunctipennis were available to be includedin the experiments.
Parasite lots B1/B–Vk210 (3 feeding experiments), B1/B–Vk247(two feeding experiments), B2/B–Vk210 (only one feeding experi-ment), and B2/B–Vk247 (seven feeding experiments) consistentlyproduced higher proportions of infected An. pseudopuntipennis withhigh mean oocyst densities than in An. albimanus (Table 5). It wasconsidered relevant to show the infectivity results produced by theB2/B–Vk210 haplotype, even from a single experiment, but notadequate enough to include in the statistical analysis.
After adjusting for gametocyte counts, the logistic modelshowed that the proportion of infected An. albimanus fed withparasites A/A–Vk210 was 95.99 times higher than those fed withparasites B1/B–Vk210 (p < 0.0005; 95% CI: 22.08–417.31); 29.13
times higher than those fed with parasites B1/B–Vk247(p < 0.0005; 95% CI: 10.56–80.29) and 38.96 times higher thanthose fed with parasites B2/B–Vk247 (p < 0.0005; 95% CI: 19.30–78.63). Parasites with haplotype B2/B–Vk210 produced similarinfection pattern in a single feeding experiment.
Significant differences in oocyst counts were found between An.
albimanus fed with parasites A/A–Vk210 (3.77 times more oocystsper mosquito) and those fed with parasites B1/B–Vk247(p < 0.0005, 95% CI: 1.41–10-15). Oocysts counts in mosquitoesfed with parasites B/B1–Vk210 and B/B2–Vk247 were not differentto those fed with parasites A/A–Vk210 (p = 0.13 and 0.54,respectively).
The logistic model showed that An. pseudopunctipennis were 83times more likely to be infected with parasites B1/B–Vk210(p < 0.0001, 95% CI: 16.43–422.28), 30.7 times more with parasitesB1/B–Vk247 (p < 0.0001, 95% CI: 8.67–108.95) and 33.64 timesmore with parasites B2/B–Vk247 (p < 0.0001, 95% CI: 15.68–72.21), than An. albimanus. As not a single An. pseudopunctipennis
(n = 83) was infected when fed with parasites A/A–Vk210, it wasnot possible to analyze differences between mosquito speciesinfectivity using this model.
The model showed no differences in the likelihood of infectionof An. pseudopunctipennis fed with parasites B1/B–Vk210 and thosefed with B1/B–Vk247 and B2/B–Vk247 (p = 0.87 and 0.55,respectively) and between those fed with parasites B1/B–Vk247and B2/B–Vk247 (p = 0.60). There were also no differences in thelikelihood of infection of An. albimanus fed with parasites B1/B–Vk210 compared to those fed with parasites B1/B–Vk247 or B2/B–Vk247 (p = 0.08 and 0.13, respectively) or B1/B–Vk247 comparedto B2/B–Vk247 (p = 0.53). Adjusting for gametocyte counts, thenegative binomial model showed that the mean oocyst counts inAn. pseudopunctipennis fed with B1/B–Vk210 did not differ to thosein An. albimanus (p = 0.44). According to the model, the meanoocyst counts in An. pseudopunctipennis fed with B1/B–Vk247(8.89) and B2/B–Vk247: (2.52) was higher than those in An.
albimanus (p < 0.0001, 95% CI: 3.51–22.499 and p = 0.003, 95% CI:1.31–3.87, respectively).
4. Discussion
Previous studies in the region first identified two parasitepopulations (Vk210 and Vk247) according to their CSPr sequencesand documented that Vk210 parasites occurred only on the coasts,while Vk247 parasites were limited to the foothills (Gonzalez-Ceron et al., 1999; Rodriguez et al., 2000). Subsequent studies usingmicrosatellite analysis identified two related parasite geneticpopulations in the foothills (f1 and f2) and one (c1) along the coast(Joy et al., 2008), and a higher infectivity for their sympatric vectormosquito species was also documented (Gonzalez-Ceron et al.,1999; Rodriguez et al., 2000; Joy et al., 2008).
The results presented here confirmed that VK247 parasites aretransmitted only in the foothills; however, Vk210 parasitesdetected on these locations since 2001 represent a recentintroduction. Pvs25 and Pvs28 sequences in all parasites fromthe coast were similar to that of El Salvador I reference strain (Sal I),and were classified as haplotype A/A–Vk210. Based on mutationsin their Pvs25/28 proteins and their association to the two CSPr,parasites from the foothills were classified as four haplotypes (B1/B–Vk210, B1/B–Vk247, B2/B–Vk210 and B2/B–Vk247). The onegenetic population (A/A–Vk210), collected only in the coastalregion, definitely corresponds to c1 parasites while the four foothillhaplotypes could correspond to either of the other two geneticpopulations (f1 and f2) identified by Joy et al. (2008), althoughtheir association is pending further investigation.
Our studies on the development of P. vivax parasites in An.
albimanus and An. pseudopunctipennis revealed that both Vk210

Table 3Amino acid sequence variation of Pvs25 among P. vivax isolates from different geographic origin.
Geographic
origin (country)
Amino acid position of the transcript (no brackets) and mature peptide (between brackets) in each
EGF-like domain:
GenPept ID: R
I II III IV
38 (16) 87 (65) 97 (75) 130 (108) 131 (109) 132 (110) 149 (127) 170 (148)
Sal I: referencestrain Met Gln Glu Ile Gln Ser Lys Cys
Mexico/ Sm type A **
Honduras/ AAV33637.1 [3]
El Salvador/ � � � � � � � � AAV33638.1 [3]
Brazil ABG29074.1 *
Brazil/ AAV33632.1 [3]
Colombia � Lys � � � � � � AAV33633.1
Venezuela Thr Lys � � � � � � AAV33634.1 [3]
Nicaragua � � � � � � � Arg AAV33636.1 [3]
Mexico/ Sm type B1 **
Thailand/ BAC66002.1 [1]
North Korea/ � � � Thr � � � � AAV33640.1 [3]
Vietnam ABG29072.1 *
Mexico/ Sm type B2 **
Turkey/ ABG29073.1 *
Mauritania/ � Lys � Thr � � � � AAV33635.1 [3]
Iran ACJ54131.1 [4]
Thailand/ BAC66003.1 [1]
Bangladesh/ BAA94347.6 [2]
India/ BAA94346.1 [2]
Indonesia/ � � Gln Thr � � � � AAV33639.1 [3]
Iran ACJ54128.1 [4]
Thailand/ BAC66001.1 [1]
Bangladesh � � Thr Lys � � � BAA94348.1 [2]
Bangladesh � � Gln Thr Lys � � � BAA94350.1 [2]
Papua New Guinea � � � Thr Lys Arg Asn � AAV33641.1 [3]
All GenPept ID were obtained from the GenBank data base: Sal I strain (XP-001608460.1) was used as reference. Sm; Southern Mexico isolates. (�) Indicates no amino acid
change compared to the Salvador I strain. R; references: [1] Sattabongkot et al. (2003); [2] Tsuboi et al. (2004); [3] Escalante et al. (2005); [4] Zakeri et al. (2009).* Unpublished data.** Data from the present study.
L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654650
and Vk247 parasites are destroyed before midgut invasion in therefractory mosquito species (Gonzalez-Ceron et al., 2001, 2007),signaling the participation of ookinete proteins, responsible formosquito midgut interactions, as determinants for the differencesin parasite infectivity to the two mosquito species (Tomas et al.,2001). Coastal A/A-Vk210 parasites were more infective to An.
albimanus, but in the foothills, B1/B – and B2/B – parasites weremore infective to An. pseudopunctipennis, regardless of their Vk210or Vk247 phenotype. This indicates an irrelevance of CSPrphenotypes and an association between variant Pvs proteinsand the infectivity to the two vectors, although the participation ofother nearby genes that are crucial for the midgut invasive processcannot be ruled out. These studies together suggested a reciprocalselection between parasites and vectors restricted by thegeographic distribution of the two vectors.
The Pvs25 and Pvs28 variants found in Southern Mexico, havebeen documented in other parts of the world (Arevalo-Herreraet al., 2005; Tsuboi et al., 1998, 2004; Sattabongkot et al., 2003;Escalante et al., 2005; Zakeri et al., 2009). The Pvs25 phenotype Acorresponds to that of Sal I strain. Sal I was isolated from ElSalvador, Central America almost 40 years ago (Collins et al., 1972),and is also CSPrVk210 (McCutchan et al., 1985). This phenotype hasbeen documented in isolates from Nicaragua, El Salvador,Honduras and Colombia (Escalante et al., 2005) where An.
albimanus is widely distributed (Rodriguez, 2006). The Pvs25mutation Lys-87 found in a high proportion of samples from thefoothills and the overlapping region of Southern Mexico has beenreported in isolates from South America (Venezuela, Colombia andBrazil), Africa (Mauritania), Turkey, but not in isolates from CentralAmerica (Nicaragua, Honduras and El Salvador) (Escalante et al.,2005), and Asia (India, Bangladesh, Thailand and China) (Tsuboi
et al., 1998, 2004; Sattabongkot et al., 2003). This mutation hasbeen recently reported in P. vivax from Iran (Zakeri et al., 2009). Thelimited polymorphism of the Pvs25 gene found in the 64 P. vivax
isolates of Southern Mexico was comparable to that reported forIranian parasites (50 P. vivax isolates), a geographic region withlow malaria transmission (Zakeri et al., 2009).
Although Pvs25 amino acid variation 130-Thr has been reportedin P. vivax parasites from North Korea, Turkey, Mauritania, PapuaNew Guinea and Indonesia (Escalante et al., 2005), India (Tsuboiet al., 1998), Thailand (Sattabongkot et al., 2003), Bangladesh(Tsuboi et al., 2004) and Iran (Zakeri et al., 2009), this is the firstreport of such variation in the American Continent. Other aminoacid substitutions (97 Gln to Glu, 131 Gln to Lys, and 132 Ser toArg), identified in Asian parasites, were neither detected in ourMexican samples nor reported in other regions of the Americas sofar. The polymorphisms 97-Asp and 110-Asn (Pvs28 type B) foundin Mexican parasites, low infective to An. albimanus wereconserved, as well as in those isolates from Asia (Sattabongkotet al., 2003; Tsuboi et al., 2004; Zakeri et al., 2009); but numerousadditional polymorphisms were detected in isolates from Bangla-desh and Thailand (Sattabongkot et al., 2003; Tsuboi et al., 2004). P.vivax isolates from the old world were also low infective to An.albimanus (Li et al., 2001).
The p25–p28 family has been described in all malaria parasites;the two genes probably arose by gene duplication (Duffy andKaslow, 1997), and are separated only by 1.851 kb of contigCM000447 (http://plasmodb.org). The results presented hererevealed linkage disequilibrium between Pvs25 and Pvs28. Linkagebetween Pvs25–Pvs28 is not surprising due to their proximity inchromosome 6. Linkage disequilibrium also occurred betweenPvs25/28 haplotype A/A and CSPr 210, which is also coded on a

Fig. 2. Amino acid position variation of the Pvs25 and Pvs28 modeled structure surface protein. a) and b) Molecular variation on the Pvs25 and Pvs28 surface protein (Sal I
strain) model depicting non-synonymous changes (bright blue) from Tables 3 and 4 respectively. The surface (grey) was rendered semi-transparent to reveal EGF-like
domains I (light blue), II (green), III (red), and IV (orange). Three sides of the molecule were produced: a1 and a2 are rotations from a and b1 and b2 are rotations from b. c) The
truncated structure of the Pvs25 protein shows subtle changes induced by the residue 130 substitution: magnification of the molecular surface of 130 Ile is shown in blue and
the non-conserved amino acid variant 130 Thr is shown in red of Pvs25. d) The truncated structure of the Pvs28 protein shows subtle changes induced by the residue 110
substitution but not for residue 87: magnification of the molecular surface of 87 Asn and 110 Tyr are shown in blue and the amino acid variant 87 Asp and 110 Asn is shown in
red of Pvs28. The models were generated by homology with the Pvs25 model (PDB:1Z27; [33]).(For interpretation of the references to color in this figure legend, the reader is
referred to the web version of the article.)
L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654 651
different chromosome (chromosome 8). This apparent associationbetween Pvs and CSPr could be the result of a relative isolation ofmalaria transmission, caused by the geographic distribution of thevectors (An. pseudopunctipennis in foothills and An. albimanus in thecoastal region), their seasonal abundance (An. pseudopunctipennis
during the dry season and An. albimanus during the rainy season)(Rodriguez et al., 2000), combined with hypo-endemic malariafavoring parasite inbreeding. The recent introduction of Vk210parasites to the foothills, demonstrates that this phenotype cancombine with the Pvs25/28 haplotypes B1/B and B2/B.
Although P25–P28 protein functions may partially overlap, ithas been suggested that P25 and P28 may have varying functionsduring parasite development in the mosquito midgut (Tomaset al., 2001; Siden-Kiamos et al., 2000; Vlachou et al., 2001). P25–P28 molecules form homo- and heteromers in tile-like prisms onthe parasite surface, suggest that conserved surface and shapemay be important for the function of the molecules. The variabilityof P25/28 in P. vivax depends mostly on non-synonymousmutations, suggesting positive selection (adaptive) of the mutat-ed sites. In Pvs25 and Pvs28, the amino acid residues at positions87, 130 and 87, 110 respectively are clearly exposed on the surfaceof the molecule. Moreover, most polymorphisms describedworldwide are exposed on the same side of the triangularprism-shaped molecule (Saxena et al., 2006). The variations inPvs25 affect domain II (87 and 97 are in loops between b sheet5–b6 and b6–7, respectively) and III (130 and 131 in b10).The polymorphic positions do not affect residues participating inthe inter-phases between domains I, III and IV. None of thepolymorphic residues affect the eighteen-conserved residue (9
acids, 5 lysines and 4 glycines) whose lateral chains are exposed tothe solvent and participate in the interaction with othermonomer, and form triangular mosaics on the parasite surface(Saxena et al., 2006). The variability in Pvs28 is greater than thatdescribed in Pvs25 (the 20 isolates analyzed here and thosereported by Sattabongkot et al., 2003; Tsuboi et al., 2004;Escalante et al., 2005; Zakeri et al., 2009), and occur in at leastnine positions (52, 65, 87, 98, 105, 110, 113, 116 and 140), domainsII and III being the most affected. Similar to Pvs25, in Pvs28, noneof the variable residues participate in forming the inter-phasebetween domains I, III and IV and do not affect conserved residueswhose lateral chains are exposed to the solvent and mayparticipate in the formation of triangular mosaics.
Transmission blocking assays with monoclonal antibodies areinformative of the possible location of P25/28 amino acid residuesthat interact with their cognate ligand on vector mosquito midgutepithelium. A monoclonal antibody (mAb) that interacts with Q87and the E97 (both polymorphic residues) of domain II of Pvs25effectively blocks mosquito infection (Saxena et al., 2006).Likewise, the blocking capacity of another mAb (Stura et al.,1994; vanAmerongen et al., 1989) is due to their interaction with Bloop of domain III of Pfs25. These data suggest that the union of P25with its ligand on the mosquito midgut epithelium is mediated bydomains II and III, and supports the hypothesis that structuralvariations on these sites can affect interactions with its cognateligand.
The binding selectivity of ookinete surface proteins may reflectdifferent invasion strategies among different vector-parasitecombinations (Han et al., 2000). We have documented the
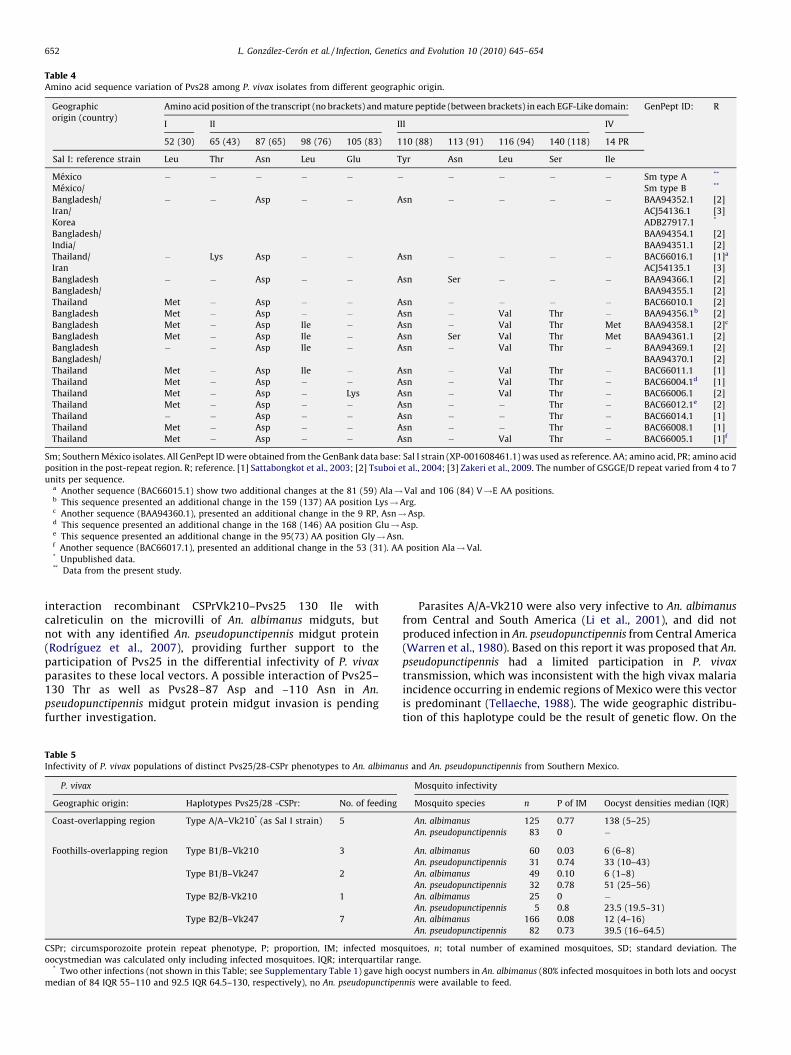
Table 4Amino acid sequence variation of Pvs28 among P. vivax isolates from different geographic origin.
Geographic
origin (country)
Amino acid position of the transcript (no brackets) and mature peptide (between brackets) in each EGF-Like domain: GenPept ID: R
I II III IV
52 (30) 65 (43) 87 (65) 98 (76) 105 (83) 110 (88) 113 (91) 116 (94) 140 (118) 14 PR
Sal I: reference strain Leu Thr Asn Leu Glu Tyr Asn Leu Ser Ile
Mexico � � � � � � � � � � Sm type A **
Mexico/ Sm type B **
Bangladesh/ � � Asp � � Asn � � � � BAA94352.1 [2]
Iran/ ACJ54136.1 [3]
Korea ADB27917.1 *
Bangladesh/ BAA94354.1 [2]
India/ BAA94351.1 [2]
Thailand/ � Lys Asp � � Asn � � � � BAC66016.1 [1]a
Iran ACJ54135.1 [3]
Bangladesh � � Asp � � Asn Ser � � � BAA94366.1 [2]
Bangladesh/ BAA94355.1 [2]
Thailand Met � Asp � � Asn � � � � BAC66010.1 [2]
Bangladesh Met � Asp � � Asn � Val Thr � BAA94356.1b [2]
Bangladesh Met � Asp Ile � Asn � Val Thr Met BAA94358.1 [2]c
Bangladesh Met � Asp Ile � Asn Ser Val Thr Met BAA94361.1 [2]
Bangladesh � � Asp Ile � Asn � Val Thr � BAA94369.1 [2]
Bangladesh/ BAA94370.1 [2]
Thailand Met � Asp Ile � Asn � Val Thr � BAC66011.1 [1]
Thailand Met � Asp � � Asn � Val Thr � BAC66004.1d [1]
Thailand Met � Asp � Lys Asn � Val Thr � BAC66006.1 [2]
Thailand Met � Asp � � Asn � � Thr � BAC66012.1e [2]
Thailand � � Asp � � Asn � � Thr � BAC66014.1 [1]
Thailand Met � Asp � � Asn � � Thr � BAC66008.1 [1]
Thailand Met � Asp � � Asn � Val Thr � BAC66005.1 [1]f
Sm; Southern Mexico isolates. All GenPept ID were obtained from the GenBank data base: Sal I strain (XP-001608461.1) was used as reference. AA; amino acid, PR; amino acid
position in the post-repeat region. R; reference. [1] Sattabongkot et al., 2003; [2] Tsuboi et al., 2004; [3] Zakeri et al., 2009. The number of GSGGE/D repeat varied from 4 to 7
units per sequence.a Another sequence (BAC66015.1) show two additional changes at the 81 (59) Ala!Val and 106 (84) V!E AA positions.b This sequence presented an additional change in the 159 (137) AA position Lys!Arg.c Another sequence (BAA94360.1), presented an additional change in the 9 RP, Asn!Asp.d This sequence presented an additional change in the 168 (146) AA position Glu!Asp.e This sequence presented an additional change in the 95(73) AA position Gly!Asn.f Another sequence (BAC66017.1), presented an additional change in the 53 (31). AA position Ala!Val.* Unpublished data.** Data from the present study.
L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654652
interaction recombinant CSPrVk210–Pvs25 130 Ile withcalreticulin on the microvilli of An. albimanus midguts, butnot with any identified An. pseudopunctipennis midgut protein(Rodrıguez et al., 2007), providing further support to theparticipation of Pvs25 in the differential infectivity of P. vivax
parasites to these local vectors. A possible interaction of Pvs25–130 Thr as well as Pvs28–87 Asp and –110 Asn in An.
pseudopunctipennis midgut protein midgut invasion is pendingfurther investigation.
Table 5Infectivity of P. vivax populations of distinct Pvs25/28-CSPr phenotypes to An. albiman
P. vivax
Geographic origin: Haplotypes Pvs25/28 -CSPr: No. of feeding
Coast-overlapping region Type A/A–Vk210* (as Sal I strain) 5
Foothills-overlapping region Type B1/B–Vk210 3
Type B1/B–Vk247 2
Type B2/B-Vk210 1
Type B2/B–Vk247 7
CSPr; circumsporozoite protein repeat phenotype, P; proportion, IM; infected mosq
oocystmedian was calculated only including infected mosquitoes. IQR; interquartilar r* Two other infections (not shown in this Table; see Supplementary Table 1) gave high
median of 84 IQR 55–110 and 92.5 IQR 64.5–130, respectively), no An. pseudopunctipe
Parasites A/A-Vk210 were also very infective to An. albimanus
from Central and South America (Li et al., 2001), and did notproduced infection in An. pseudopunctipennis from Central America(Warren et al., 1980). Based on this report it was proposed that An.
pseudopunctipennis had a limited participation in P. vivax
transmission, which was inconsistent with the high vivax malariaincidence occurring in endemic regions of Mexico were this vectoris predominant (Tellaeche, 1988). The wide geographic distribu-tion of this haplotype could be the result of genetic flow. On the
us and An. pseudopunctipennis from Southern Mexico.
Mosquito infectivity
Mosquito species n P of IM Oocyst densities median (IQR)
An. albimanus 125 0.77 138 (5–25)
An. pseudopunctipennis 83 0 �
An. albimanus 60 0.03 6 (6–8)
An. pseudopunctipennis 31 0.74 33 (10–43)
An. albimanus 49 0.10 6 (1–8)
An. pseudopunctipennis 32 0.78 51 (25–56)
An. albimanus 25 0 �An. pseudopunctipennis 5 0.8 23.5 (19.5–31)
An. albimanus 166 0.08 12 (4–16)
An. pseudopunctipennis 82 0.73 39.5 (16–64.5)
uitoes, n; total number of examined mosquitoes, SD; standard deviation. The
ange.
oocyst numbers in An. albimanus (80% infected mosquitoes in both lots and oocyst
nnis were available to feed.

L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654 653
other hand, haplotype Pvs25–130 Thr/Pvs28–87 Asp–110 Asnparasites also circulate in other distant geographic regions(Bangladesh and Iran) suggesting evolutive convergence drivenby a need to adapt to diverse binding mosquito midgut molecules(Morris, 2003; Shi and Yokoyama, 2003). As it occurs in SouthernMexico, the transmission of selective parasite strains by differentmosquito species suggests a selective pressure due to geographicisolation produced by the vector–parasite interaction and theecological conditions restricting mosquito breeding distribution(Li et al., 2001).
Until biochemical interactions between Pvs25/28 variants andmidgut mosquito molecules are demonstrated, it remains possiblethat these associations only reflect the prevalence of the variantswithin parasite populations. The possible interaction of Pvs25 130Thr with An. pseudopunctipennis midgut proteins, as well as theparticipation of Pvs28 variants in An. pseudopunctipennis and An.
albimanus midgut invasion is currently being studied; a differentialinteraction would provide evidence for the purported co-evolu-tion. From the epidemiological point of view, it is essential toinvestigate whether the association between Pvs25/28-CSPrhaplotypes with these two vectors found here, prevails in otherP. vivax affected regions of America.
Acknowledgements
This work was supported by funds of the INSP-Mexico (project476191) and CONACyT-Mexico (SALUD-2004-01-119). We thankthe insectary personnel at CIP/INSP–Tapachula for contributing tothe inbreeding of the mosquito species and to Frida Santillan, JoseNettel and Olga Palomeque for their technical assistance inmosquito blood feeding and oocyst count. We also thank D.Garboczi for providing useful suggestions for Pvs25 structurepresentation.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.meegid.2010.03.014.
References
Arevalo-Herrera, M., Solarte, Y., Yasnot, M.F., Castellanos, A., Rincon, A., Saul, A., Mu,J., Long, C., Miller, L., Herrera, S., 2005. Induction of transmission-blockingimmunity in Aotus monkeys by vaccination with a Plasmodium vivax clinicalgrade PVS25 recombinant protein. Am. J. Trop. Med. Hyg. 73, 32–37.
Baton, L.A., Ranford-Cartwright, L.C., 2005. Do malaria ookinete surface proteinsP25 and P28 mediate parasite entry into mosquito midgut epithelial cells.Mal. J. 4, 15.
Collins, W.E., Contacos, P.G., Krotoski, W.A., Howard, W.A., 1972. Transmission offour Central American strains of Plasmodium vivax from monkey to man. J.Parasitol. 58, 332–333.
Collins, W.E., Barnwell, J.W., Sullivan, J.S., Nace, D., Williams, T., Bounngaseng, A.,Roberts, J., Strobert, E., McClure, H., Saul, A., Long, C.A., 2006. Assessment oftransmission-blocking activity of candidate Pvs25 vaccine using gametocytesfrom chimpanzees. Am. J. Trop. Med. Hyg. 74, 215–221.
Danielli, A., Barillas-Mury, C., Kumar, S., Kafatos, F.C., Loukeris, T.G., 2005. Overexpression and altered nucleocytoplasmic distribution of Anopheles ovalbu-min-like SRPN10 serpins in Plasmodium-infected midgut cells. Cell. Microbiol. 7,181–190.
Duffy, P.E., Kaslow, D.C., 1997. A novel malaria protein, Pfs28, and Pfs25 aregenetically linked and synergistic as falciparum malaria transmission-blockingvaccines. Infect. Immun. 65, 1109–1113.
Duffy, P.E., Pimenta, P., Kaslow, D.C., 1993. Pgs28 belongs to a family of epidermalgrowth factor-like antigens that are targets of malaria transmission-blockingantibodies. J. Exp. Med. 177, 505–510.
Escalante, A.A., Cornejo, O.E., Freeland, D.E., Poe, A.C., Durrego, E., Collins, W.E., Lal,A.A., 2005. A monkey tales: the origin of Plasmodium vivax as a human malariaparasite. Proc. Natl. Acad. Sci. USA 102, 1980–1985.
Ewing, B., Hiller, L., Wendl, M., Green, P., 1998. Base-calling of automated sequencetraces using phred. I. Accuracy assessment. Genome Res. 8, 175–185.
Gonzalez-Ceron, L., Rodrıguez, M.H., Wirtz, R.A., Sina, B.J., Palomeque, O.L., Nettel,J.A., Tsutsumi, V., 1998. Plasmodium vivax: a monoclonal antibody recognizes a
circumsporozoite protein precursor on the sporozoite surface. Exp. Parasitol.90, 203–211.
Gonzalez-Ceron, L., Rodrıguez, M.H., Nettel, J.A., Villarreal, C., Kain, K., Hernandez,J.E., 1999. Differential susceptibilities of Anopheles albimanus and Anophelespseudopunctipennis to infection with coindigenous Plasmodium vivax variantsVK210 and VK247 in Southern Mexico. Infect. Immun. 67, 410–412.
Gonzalez-Ceron, L., Rodriguez, M.H., Santillan, F., Chavez, B., Nettel, J.A., Hernandez-Avila, J.E., Kain, K.C., 2001. Plasmodium vivax: ookinete destruction and oocystdevelopment arrest are responsible for Anopheles albimanus resistance tocircumsporozoite phenotyphe VK247 parasites. Exp. Parasitol. 98, 152–161.
Gonzalez-Ceron, L., Rodriguez, M.H., Chavez-Munguia, B., Santillan, F., Nettel, J.A.,Hernandez-Avila, J.E., 2007. Plasmodium vivax: impaired escape of Vk210phenotype ookinetes from the midgut blood bolus of Anopheles pseudopuncti-pennis. Exp. Parasitol. 115, 59–67.
Gordon, D., Abanjian, C., Green, P., 1998. Consed: a graphical tool for sequencefinishing. Gen. Res. 8, 195–202.
Guex, N., Peitsch, M.C., 1997. SWISS-MODEL and the Swiss-PdbViewer: an environ-ment for comparative protein modeling. Electrophoresis 18, 2714–2723.
Han, Y.S., Thompson, J., Kafatos, F.C., Barillas-Mury, C., 2000. Molecular interactionsbetween Anopheles stephensi midgut cells and Plasmodium berghei: the timebomb theory of ookinete invasion of mosquitoes. EMBO J. 19, 6030–6040.
Hisaeda, H., Stowers, A.W., Tsuboi, T., Collins, W.E., Sattabongkot, J.S., Suwanabun,N., Torii, M., Kaslow, D.C., 2000. Antibodies to malaria vaccine candidates Pvs25and Pvs28 completely block the ability of Plasmodium vivax to infect mosqui-toes. Infect. Immun. 68, 6618–6623.
Haubold, B., Hudson, R.R., 2000. LIAN 3. 0: detecting linkage disequilibrium inmultilocus data. Linkage analysis. Bioinformatics 16, 847–848.
Joy, D.A., Gonzalez-Ceron, L., Carlton, J.M., Gueye, A., Fay, M., McCutchan, T.F., Su,X.Z., 2008. Local adaptation and vector-mediated population structure inPlasmodium vivax malaria. Mol. Biol. Evol. 25, 1245–1252.
Kain, K.C., Wirtz, R.A., Fernandez, I., Franke, E.D., Rodrıguez, M.H., Lanar, D.E., 1992.Serologic and genetic characterization of Plasmodium vivax from whole bloodimpregnated filter paper discs. Am. J. Trop. Med. Hyg. 46, 473–479.
Kaslow, D.C., Quakyi, I.A., Syin, C., Raum, M., Keister, D.B., Coligan, J.E., McCutchan,T.F., Miller, L.H., 1988. A vaccine candidate from the sexual stage of humanmalaria that contains EGF-like domains. Nature 333, 74–76.
Kaslow, D.C., Syin, C., McCutchan, T.F., Miller, L.H., 1989. Comparison of the primarystructure of the 25 kDa ookinete surface antigen of Plasmodium falciparum andPlasmodium gallinaceum reveals six conserved regions. Mol. Biochem. Parasitol.33, 283–287.
Kaslow, D.C., 1996. Transmission-blocking vaccines. In: Hoffman, S.L. (Ed.), MalariaVaccines. 2nd ed. ASM Press, Washington DC, pp. 81–228.
Kumar, N., Carter, R., 1985. Biosynthesis of two stage-specific membrane proteinsduring transformation of Plasmodium gallinaceum zygotes into ookinetes. Mol.Biochem. Parasitol. 14, 127–139.
Lal, A.A., Goldman, I.F., Campbell, G.H., 1990. Primary structure of the 25-kilodaltonookinete antigen from Plasmodium reichenowi. Mol. Biochem. Parasitol. 43,143–145.
Li, J., Collins, W.E., Wirtz, R.A., Rathore, D., Lal, A., McCutchan, T.F., 2001. Geographicsubdivision of the range of the malaria parasite Plasmodium vivax. Emerg. Infect.Dis. 7, 35–42.
McCullag, P., Nelder, J.A., 1989. Generalized Linear Models, 2nd ed. Chapman andHall, New York.
McCutchan, T.F., Lal, A.A., de la Cruz, V.F., Miller, L.H., Maloy, W.L., Charoenvit, Yl,Beaudoin, R.L., Guerry, P., Wistar Jr., R., Hoffman, S.L., Hockmeyer, W.T., Collins,W.E., Wirth, D., 1985. Sequence of the immunodominant epitope for the surfaceprotein on sporozoites of Plasmodium vivax. Science 230, 1381–1383.
Menard, R., Sultan, A.A., Cortes, C., Altszuler, R., Van Dijk, M.R., Janse, C.J., Waters,A.P., Nussenzweig, R.S., Nussenzweig, V., 1997. Circumsporozoite protein isrequired for development of malaria sporozoites in mosquitoes. Nature 385,336–340.
Morris, S.C., 2003. Life’s Solution: Inevitable Humans in a Lonely Universe. Cam-bridge University Press, Cambridge.
Paton, M.G., Barker, G.C., Matsuoka, H., Ramesar, J., Janse, C.J., Waters, A.P., Sinden,R.E., 1993. Structure and expression of a post-transcriptionally regulated ma-laria gene encoding a surface protein from the sexual stages of Plasmodiumberghei. Mol. Biochem. Parasitol. 59, 263–275.
Rodriguez, M.H., Gonzalez-Ceron, L., Hernandez, J.E., Nettel, J.A., Villarreal, C.,Kain, K.C., Wirtz, R.A., 2000. Different prevalences of Plasmodium vivaxphenotypes VK210 and VK247 associated with the distribution of Anophelesalbimanusand Anopheles pseudopunctipennis in Mexico. Am. J. Trop. Med. Hyg.62, 122–127.
Rodriguez, M.H., 2006. Malaria and dengue vector biology and control in LatinAmerica. In: Knols, B.G.J., Louis, C., Bogers, R.J. (Eds.), Bridging Laboratory andField Research for Genetic Control of Disease Vectors. Series Wageningen U. R.Frontis Series, vol. 11. Springer, Dordrecht, TheNetherlands, pp. 129–141.
Rodrıguez, M.C., Martınez-Barnetche, J., Alvarado-Delgado, A., Batista, C., Argotte-Ramos, R.S., Hernandez-Martınez, S., Gonzalez-Ceron, L., Torres, J.A., Margos, G.,Rodrıguez, M.H., 2007. The surface protein Pvs25 of Plasmodium vivax ookinetesinteracts with calreticulin on the midgut apical surface of the malaria vectorAnopheles albimanus. Mol. Biochem. Parasitol. 153, 167–177.
Sattabongkot, J., Tsuboi, T., Hisaeda, H., Tachibana, M., Suwanabun, N., Rungruang,T., Cao, Y.M., Stowers, A.W., Sirichaisinthop, J., Coleman, R.E., Torii, M., 2003.Blocking of transmission to mosquitoes by antibody to Plasmodium vivaxmalaria vaccine candidates Pvs25 and Pvs28 despite antigenic polymorphismin field isolates. Am. J. Trop. Med. Hyg. 69, 536–541.

L. Gonzalez-Ceron et al. / Infection, Genetics and Evolution 10 (2010) 645–654654
Saxena, A.K., Singh, K., Su, H.P., Klein, M.M., Stowers, A.W., Saul, A.J., Long, C.A.,Garboczi, D.N., 2006. The essential mosquito-stage P25 and P28 proteins fromPlasmodium form tile-like triangular prisms. Nat. Struct. Mol. Biol. 13, 90–91.
Schwede, T., Kopp, J., Guex, N., Peitsch, M.C., 2003. SWISS-MODEL: an automatedprotein homology-modeling server. Nucleic Acids Res. 31, 3381–3385.
Shi, Y., Yokoyama, S., 2003. Molecular analysis of the evolutionary significance ofultraviolet vision in vertebrates. PNAS 100, 8308–8313.
Siden-Kiamos, I., Vlachou, D., Margos, G., Beetsma, A., Waters, A.P., Sinden, R.E.,Louis, C., 2000. Distinct roles for pbs21 and pbs25 in the in vitro ookinete tooocyst transformation of Plasmodium berghei. J. Cell. Sci. 113, 3419–3426.
Sinden, R.E., 1984. The biology of Plasmodium in the mosquito. Experientia 40,1330–1343 Review.
Stura, E.A., Sarrterthwait, A.C., Calvo, J.C., Stefancko, R.S., Langevels, J.P., Kaslow, D.C.,1994. Crystallization of an intact monoclonal antibody (4B7) against Plasmodi-um falciparum malaria with peptides from the Pfs25 protein antigen. ActaCrystallogr. D Biol. Crystallogr. 50, 556–562.
Tachibana, M., Tsuboi, T., Templeton, T.J., Kaneko, O., Torii, M., 2001. Presence ofthree distinct ookinete surface protein genes, Pos25 Pos28-1, and Pos28-2, inPlasmodium ovale. Mol. Biochem. Parasitol. 113, 341–344.
Tellaeche, A.M., 1988. La evolucion y la situacion actual del paludismo en Mexico:A cien anos del descubrimiento de Ross. In: Kumate, J., Martınez-Palomo, A.(Eds.), El paludismo en Mexico. El Colegio Nacional, Mexico city, pp. 209–218.
Tomas, A.M., Margos, G., Dimopoulos, G., Van Lin, L.H., De Koning-Ward, T.F., Sinha,R., Lupetti, P., Beetsma, A.L., Rodriguez, M.C., Karras, M., Hager, A., Mendoza, J.,Butcher, G.A., Kafatos, F., Janse, C.J., Waters, A.P., Sinden, R.E., 2001. P25 and P28proteins of the malaria ookinete surface have multiple and partially redundantfunctions. EMBO J. 15, 3975–3983.
Tsuboi, T., Kaslow, D.C., Cao, Y.M., Shiwaku, K., Torii, M., 1997a. Comparison ofPlasmodium yoelii ookinete surface antigens with human and avian malariaparasite homologues reveals two highly conserved regions. Mol. Biochem.Parasitol. 87, 107–111.
Tsuboi, T., Cao, Y.M., Kaslow, D.C., Shiwaku, K., Torii, M., 1997b. Primary structure ofa novel ookinete surface protein from Plasmodium berghei. Mol. Biochem.Parasitol. 85, 131–134.
Tsuboi, T., Kaslow, D.C., Margaret, G.M., Tachibana, M., Cao, Y.M., Torii, M., 1998.Sequence polymorphism in two novel Plasmodium vivax ookinete surfaceproteins Pvs25 and Pvs28, that are malaria transmission-bloking vaccine can-didates. Mol. Med. 4, 772–782.
Tsuboi, T., Kaneko, O., Cao, Y.M., Tachibana, M., Yoshihiro, Y., Nagao, T., Kanbara, H.,Torii, M., 2004. A rapid genotyping method for the vivax malaria transmission-blocking vaccine candidates Pvs25 and Pvs28. Parasitol. Int. 53, 211–216.
vanAmerongen, A., Sauerwein, R.W., Beckers, P.J., Meloen, R.H., Mewissen, J.H.,1989. Identification of a peptide sequence of the 25 kD surface protein ofPlasmodium falciparum recognized by transmission-blocking monoclonal anti-bodies: implications for synthetic vaccine development. Parasite Immunol. 11,425–428.
Villarreal, C., Arredondo-Jimenez, J.I., Rodriguez, M.H., Ulloa, A., 1998. Colonizationof Anopheles pseudopunctipennis from Mexico. J. Am. Mosq. Control Assoc. 14,369–372.
Vlachou, D., Lycett, G., Siden-Kiamos, I., Blass, C., Sinden, R.E., Louis, C., 2001.Anopheles gambiaelaminin interacts with the P25 surface protein of Plasmodiumbergheiookinetes. Mol. Biochem. Parasitol. 112, 229–237.
Vlachou, D., Schlegelmilch, T., Runn, E., Mendes, A., Kafatos, F.C., 2006. The devel-opmental migration of Plasmodium in mosquitoes. Curr. Opin. Genet. Dev. 16,384–391 Review.
Warren, M., Collins, W.E., Jeffery, G.M., Richardson, B.B., 1980. Anopheles pseudo-punctipennis: laboratory maintenance and malaria susceptibility of a strainfrom El Salvador. Am. J. Trop. Med. Hyg. 29, 503–506.
World Health Organization, 1967. Chemotherapy of malaria. World Health Organ.Tech. Rep. Ser. 375.
Zakeri, S., Razavi, S., Djadid, N.D., 2009. Genetic diversity of transmission blockingvaccine candidate (Pvs25 and Pvs28) antigen in Plasmodium vivax clinicalisolates from Iran. Acta Trop. 109, 176–180.




















