
Metazoan parasite communities of three endemic cichlid fish ...

Experimental Parasitology 103 (2003) 127–135
www.elsevier.com/locate/yexpr
Plasmodium falciparum: limited genetic diversity of MSP-2in isolates circulating in Brazilian endemic areas
S. Sallenave-Sales,a,* M.F. Ferreira-da-Cruz,a C.P. Faria,a C. Cerruti Jr.,b
C.T. Daniel-Ribeiro,a and M.G. Zalisc,d
a WHO Collaborating Center for Research and Training in the Immunology of Parasitic Diseases, Departamento de Imunologia,
Instituto Oswaldo Cruz/Fiocruz, Avenida Brasil 4365, Manguinhos, CEP 21045-900, Rio de Janeiro, Brazilb Departamento de Medicina Social, Universidade Federal do Esp�ıırito Santo, Rio de Janeiro, Brazil
c Instituto de Biof�ıısica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazild Departamento de Medicina Preventiva, Programa de Doenc�as Infecto-Parasit�aarias, Hospital Universit�aario Clementino Fraga Filho,
UFRJ, CEP 21949-000, Rio de Janeiro, Brazil
Received 13 August 2002; received in revised form 24 April 2003; accepted 5 May 2003
Abstract
The genetic polymorphism of the surface merozoite protein 2 (MSP-2) was evaluated in Plasmodium falciparum isolates from
individuals with uncomplicated malaria living in a Brazilian endemic area of Peixoto de Azevedo. The frequency of MSP-2 alleles
and the survival of genetically different populations clones in 104 isolates were verified by Southern blot and SSCP-PCR. Single and
mixed infections were observed in similar frequencies and the rate of detection of FC27 and 3D7 allelic families was equivalent.
Eight alleles were identified and among them, the sequence polymorphism was mainly attributed to variations in the repetitive
region. Interestingly, in three alleles nucleotide polymorphism was identical to that detected in a previous study, conducted in 1992,
in a near Brazilian endemic area. This finding demonstrated the genetic similarity between two isolate groups, besides the certain
temporal stability in the allelic patterns. The implications of these data for studies on the genetic diversity are also discussed.
� 2003 Elsevier Science (USA). All rights reserved.
Index Descriptors and Abbreviations: MSP-2; Plasmodium falciparum; allelic diversity; SSCP-PCR; MSP-2, merozoite surface protein 2; PCR,
polymerase chain reaction; SSCP, single strand conformational polymorphism; SDS, sodium dodecyl sulfate; EDTA, ethylenediaminetetraacetic acid
1. Introduction
The great genetic variability of Plasmodium falcipa-
rum contributes to generate a number of biologicallyand antigenically different parasite populations (Anders
et al., 1993; Babiker et al., 1997; Druilhe et al., 1998).
This genetic diversity can occur at any endemic level and
appears to be related to pathology as well as acquired
immunity, since multiple allelic forms could differ in
their ability to escape recognition by the host�s immuneresponse (Engelbrecht et al., 1995; Gupta et al., 1994;
Muller et al., 2001; Zevering et al., 1994). Presumably,this polymorphism would also restrict the effectiveness
of subunit vaccines against P. falciparum infection if
* Corresponding author. Fax: +55-21-280-1589.
E-mail address: [email protected] (S. Sallenave-Sales).
0014-4894/03/$ - see front matter � 2003 Elsevier Science (USA). All rights
doi:10.1016/S0014-4894(03)00091-2
variable proteins are included (Genton et al., 2002).
Additionally, the nature and extent of antigenic diversity
could also give new information on the dynamics of
host–parasite relationships and on parasite evasionmechanisms.
Results from epidemiological investigations using
PCR-based assays and allele specific hybridization
studying P. falciparum polymorphic antigens have given
valuable data on parasite population structure in the
human host. These typing schemes estimate the extent
of antigenic polymorphism by the frequency evaluation
of different circulating allelic variants and by the ge-notype complexity of individual infections (Babiker et
al., 1999; Daubersies et al., 1994; Mercereau-Puijalon,
1996; Ntoumi et al., 1995; Sallenave-Sales et al., 2000).
In this context, particular attention has been given to
antigens associated with the surface of merozoite stage,
reserved.

128 S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135
such as the 35–56-kDa merozoite surface protein-2(MSP-2) of P. falciparum. The location of this protein,
and its possible role in the invasion process, as well as
the ability of anti-MSP-2 antibodies to inhibit parasite
growth in vitro and in vivo, motivated its use as a
potential malaria vaccine candidate (Genton et al.,
2000; Lawrence et al., 2000). The MSP-2 gene contains
blocks with tandem repeated units that can vary in size
and nucleotide sequence in P. falciparum isolates(Smythe et al., 1988, 1990). The MSP-2 alleles described
generally fall into two allelic families, FC27 and 3D7,
which differ considerably by the dimorphic structure of
the variable central region. The FC27 family shows
varying numbers of structurally conserved R1 (96 bp)
and R2 (36 bp) repetitive regions, in contrast to those
observed in the 3D7 family, where the R1 region is less
conserved being highly variable in length, copy number,and sequence.
Field studies on natural populations of P. falciparum
carried out mainly in hyper- and holoendemic regions
have demonstrated that MSP-2 gene is highly poly-
morphic, showing considerable size and sequence vari-
ation which can reflect in complexes infections with a
mixture of allelic variants belonging to the same and/or
different allelic family (Babiker et al., 1999; Felger et al.,1999; Kyes et al., 1997; Paul et al., 1995). Sequence
variation may also result from the recombination events
between allelic variants of FC27 and 3D7 families within
the mosquito vector (Irion et al., 1997; Keer et al., 1994;
Marshall et al., 1991; Snewin et al., 1991). This extensive
polymorphism raises the important question about the
impact of MSP-2 diversity on the immune response, as
well as the ability of different allelic variants to inducedifferent clinical manifestations (Al-Yaman et al., 1997;
Engelbrecht et al., 1995). In fact, specific effects of a
malaria vaccine containing 3D7 allele of MSP-2 antigen
on parasites with this particular MSP-2 genotype were
demonstrated, besides the vaccination with 3D7 may
have contributed to the increase the rate of FC27-as-
sociated morbidity in vaccinated children (Genton et al.,
2002).MSP-2 alleles and their frequencies have been com-
monly determined by PCR-RFLP and/or Southern blot
hybridization using specific probes and rely on size dif-
ferences between repeat regions. However, such meth-
ods underestimate the true level of diversity and further
most of them cannot identify hybrid recombinant MSP-
2 alleles. The association of restriction analysis methods
to nucleotide sequencing provides a more accurate de-termination of the genetical microheterogeneity of MSP-
2. Considering these aspects together with the fact that
few data are available about the complexity and be-
havior of P. falciparum populations in Brazilian endemic
areas (Creasey et al., 1990; Hoffmann et al., 2001;
Sallenave-Sales et al., 2000), we decided to study the
allelic diversity of MSP-2 gene in P. falciparum Brazilian
isolates from residents of Peixoto de Azevedo village(MT-Brazil).
2. Materials and methods
2.1. Study site
Studies have been carried out between 1995 and 1996in Peixoto de Azevedo village, which is localized in the
Mato Grosso State in the southern part of the Brazilian
Amazon Forest. The population consists mainly of mi-
grants individuals exposed during all the year to the risk
of malaria infection (Duarte and Fontes, 1998). At time
collection, the village had the highest malaria transmis-
sion rates in Brazil with an annual parasitic index (po-
sitive slides/1000 inhabitants) of 284.8 and 263.1 in 1995and 1996, respectively. The number of malaria cases in-
duced by P. falciparum and Plasmodium vivax was very
similar, corresponding to 47 and 52% in 1995 and to 43
and 56% in 1996, respectively. In 1995, the rate of bites/
person/hour by Anopheles darlingi vector was 10. Some
P. falciparum isolates collected in 1992 in a similar study
carried out in Porto Velho, another Brazilian endemic
area, were used in a comparative analysis.
2.2. Isolates
After signed consent venous blood samples were ta-
ken from 104 symptomatic individuals from the mu-
nicipality of Peixoto de Azevedo (MT-Brazil). The
P. falciparum parasite density was determined by mi-
croscopic examination of Giemsa stained thick bloodsmears. The fresh blood samples were centrifuged
(10min at 350g) to remove the plasma and the leuko-
cytes were depleted by repeated washes with an equal
volume of 0.15M phosphate-buffered saline. The red
blood cell pellet was stored in 50% (v/v) glycerolyte so-
lution (0.9% NaCl/4.2% sorbitol/20% glycerol) in liquid
nitrogen (N2). Samples from Porto Velho were similarly
collected and stored.
2.3. PCR amplification and electrophoresis
One milliliter of red blood cells was lysed by 2–3
washing cycles with distilled water. Free parasites were
resuspended in 5 volumes of TEN buffer (10mM Tris–
HCl, pH 8.0, 1mM EDTA, pH 8.0, 0.15M NaCl, 0.5%
Triton X-100, and 0.5% SDS) and 5mg/ml proteinase Kand incubated at 37 �C for 2–3 h. The DNA was phenol-
extracted, precipitated, and amplified using P. falciparum
oligonucleotides specific for the central polymorphic re-
gion of MSP-2 antigen, as described elsewhere (Conta-
min et al., 1995). Two microliters of DNA was amplified
in a 100 ll reaction volume containing 10 nmol of each
dNTP, 100 pmol of each primer, 2.5U TaqI DNA

Fig. 1. MSP-2 gene polymorphism by PCR in P. falciparum isolates
from Peixoto de Azevedo. I—520 bp; II—500 bp; III—600bp; and IV—
620 bp.
S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135 129
Polymerase (Perkin–Elmer), and 5 ll of 10� buffer(Perkin–Elmer, 2.0mM MgCl2). The PCRs were carried
out using a Hybaid automated heating block for 35 cy-
cles (2 s at 94 �C, 1min at 55 �C, and 2min at 72 �C). Tenmicroliters of PCR was loaded onto a 2.5% Nusieve
GTG agarose gel (FCM Bioproduct, USA) in 1� TAE
buffer (0.04M Tris–acetate, 1mM EDTA) in the pres-
ence of ethidium bromide (0.5 lg/ml).
2.4. Hybridization conditions
Following gel analysis, amplified products from
MSP-2 were transferred onto a Nylon membrane (Hy-
bond-N, Amersham), as recommended by suppliers, and
hybridized with specific radioactive probes representing
each one of the two allelic families of the MSP-2 gene:
FC27 and 3D7. The probes were obtained by amplifi-cation of P. falciparum DNA from reference isolates and
clones (Contamin et al., 1995). These probes were la-
beled by Nick Translation (Boehringer–Mannheim,
France) and the unincorporated nucleotides were re-
moved by spin dialysis (Quickspin, Amersham). Hy-
bridizations were performed overnight at 65 �C in the
presence of 6� SSC (0.9M NaCl, 0.09M sodium cit-
rate), 0.1% SDS, 2.5% skimmed milk, and 100 lg/ml ofsheared salmon sperm DNA (Sigma). The membranes
were successively washed at 65 �C in 6� SSC, 2� SSC,
0.5� SSC, and 0.1� SSC. After each wash, the mem-
branes were exposed to an X-OMAT film (Kodak) with
an intensifying screen for 6–48 h at )70 �C.
2.5. Single-strand conformational polymorphism
To assess sequence microheterogeneity within frag-
ments defined as the same genotype by PCR and allelic
typing, each amplified fragment was analyzed by SSCP.
The PCR sample was previously concentrated with 0.3M
of sodium acetate, resuspended in 30 ll of distilled water,and loaded onto a 2% Nusieve GTG agarose gel (FCM
Bioproduct, USA) in 1� TAE buffer (0.04M Tris–ace-
tate, 1mM EDTA). The amplified fragments were indi-vidually gel-purified using Wizard DNA preps as
recommended by suppliers (Promega) and resuspended
in 40 ll of distilled water. PCR fragments were digested
to completion with RsaI (Promega), then extracted with
phenol–chloroform, and precipitated with ethanol. The
samples were denatured prior to analysis by adding 1 llof 0.5MNaOH/10mM EDTA and then electrophoresed
for 3 h at 10V/cm using 0.5� Tris–borate EDTA (TBE)buffer (pH 8.4). SSCP patterns were visualized by silver
staining as described elsewhere (Gonc�alves et al., 1990).
2.6. DNA sequencing
To verify sequence modifications in the alleles of
MSP-2, the allelic types from Peixoto de Azevedo were
sequenced. In addition, four fragments from PortoVelho (Rondoonia State) that were previously typed
(Sallenave-Sales et al., 2000) were chosen for a com-
parative analysis: two FC27 alleles—PV54 (500 bp) and
PV56 (520 bp) and two 3D7 alleles—PV25 (620 bp) and
PV61 (560 bp). Uncloned amplified fragments were se-
quenced using the di-deoxy chain termination procedure
with Taq polymerase and fluorescently labeled (Dye
terminator kit, Perkin–Elmer) with the primers 50-GAGTAT AAG GAG AAG TAT GG-30 (forward) and 50-CCT GTA CCT TTA TTC TCT GG-30 (reverse). Se-quencing products were separated on an ABI Model
373A automated DNA Sequencer and the sequences
were analyzed using University of Wisconsin Genetics
Computer Group software package. The resulting MSP-
2 allele sequences have been submitted to EMBL/Gen-
Bank Data libraries under Accession Nos. AY137758 toAY137765.
2.7. Statistical analysis
The data were analyzed using the program GrapPad
Instat, version 2.05a. The v2 test was applied for the
analysis of allelic frequencies.
3. Results
3.1. Allelic polymorphism of MSP-2
One hundred and seventeen PCR fragments corre-
sponding to 104 P. falciparum isolates from Peixoto de
Azevedo were obtained from the central polymorphicregion of MSP-2 (Fig. 1). Thirty-four (24%) fragments
that were not visible by ethidium bromide staining were
detected after probing, performing a total of 154 frag-
ments identified in this area. All of them hybridized
against only one of the two probes (either FC27 or
3D7) at low stringency conditions (0.1� SSC) (data not
shown). Ninety (58%) out of 154 fragments were
identified as FC27 family—type I (520 bp), type II(500 bp), and type III (600 bp), whereas the 3D7 family

Fig. 2. Single strand conformational polymorphism analysis of MSP-2
PCR fragments. MSP-2 PCR fragments were digested with RsaI and
analyzed by SSCP in a silver staining 10% polyacrylamide gel: Ia and
Ib—520 bp patterns; II—500 bp pattern; III—600bp pattern; and IVa–
IVd—620bp patterns. The PhiX174 HaeIII (left) and 100 bp (right)
were used as DNA size markers.
130 S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135
fragments—type IV (620 bp)—represented about 42%
(64/154). Single (one fragment) and mixed (two frag-
ments) MSP-2 infections were observed in similar fre-
quencies, representing 53 and 47% of the 104 isolates,
respectively. The presence of fragments of both allelic
families in mixed infections was 96%.
A degree of DNA microheterogeneity that was un-
detectable by PCR alone was verified by SSCP analysisof the 117 fragments detected in electrophoresis gel (Fig.
2). Alteration sequences were observed only in I (FC27/
520 bp) and IV (3D7/620 bp) types, whereas no mobility
shift was observed for fragments II (FC27/500 bp) and
Table 1
Infection patterns of the 104 P. falciparum isolates analyzed by SSCP-PCR
Complexity infection Fragment (bp) Hybridi
Single infections1
S1 520 FC27
S2 520 FC27
S3 500 FC27
S4 600 FC27
S5 620 3D7
S6 620 3D7
Mixed infections2
M7 520/500 FC27/F
M8 520/600 FC27/F
M9 520/620 FC27/3D
M10 520/620 FC27/3D
M11 520/620 FC27/3D
M12 520/620 FC27/3D
M13 500/620 FC27/3D
1 Isolates with one fragment.2 Isolates with two fragments; bp—base pair
III (FC27/600 bp). Two SSCP patterns were observedfor type I, designated alleles Ia and Ib, and four were
found for type IV—IVa, IVb, IVc, and IVd alleles. The
Ib (59/117) and IVa (28/117) alleles were the most fre-
quent types detected among the 104 isolates. In 4% of
SSCP pattern obtained, we detected additional bands of
low intensity most probably corresponding to the minor
clonal populations that were not discriminated by PCR
technique (data not shown).Considering the size and number of fragments found
by the isolate characterized, the allelic typing, and the
SSCP pattern, we observed nine different MSP-2 infec-
tion profiles in Peixoto de Azevedo (Table 1).
3.2. Sequencing of MSP-2 fragments
The FC27 alleles showed repetitive sequences thatwere related but not identical to each other. The repeat
regions of all compared alleles of this family consist of
one (Ia, Ib, and II alleles) or three (III allele) copies of
96 bp (R1) and two (II allele), three (Ia and Ib alleles) or
none (III allele) copies of 32 bp (R2), thus displaying a
more conserved organization than the 3D7 allelic family
(Fig. 3). Point mutations were spread by the regions�sequences, although some of them were shared by allalleles, such as that which resulted in a non-synonymous
substitution of the six amino acids (SSGNAP) in the E3
region (Fig. 4). Few nucleotide substitutions, spread in
the E1 (family specific region 1), R1, and R2 regions,
account for the difference between the three FC27 alleles
(Ia, Ib, and II), all leading to non-synonymous changes
(Fig. 4). More diversity was observed when referring to
allele III (600 bp) that presented three identical copies of96 bp repeats in R1 region contrasting with the lack of
32 bp repeats in the R2 region. The deletion of two
codons and some nucleotide substitutions were also
and allelic typing
zation SSCP pattern No. of isolates, N (%)
Ia 13 (12)
Ib 50 (48)
II 5 (5)
III 2 (2)
IVa 20 (19)
IVd 1 (1)
C27 Ia/II 1 (1)
C27 Ib/III 1 (1)
7 Ia/IVa 1 (1)
7 Ib/IVa 6 (6)
7 Ib/IVb 1 (1)
7 Ib/IVc 2 (2)
7 II/IVa 1 (1)

Fig. 5. Schematic representation of the predicted 3D7 MSP-2 protein
sequences from Peixoto de Azevedo isolates. The central variable re-
gion of each allelic type is compared to the 3D7 reference strain
(Smythe et al., 1990), which is represented in the upper position. Un-
filled blocks of 3D7 protein sequence indicate conserved regions. The
black blocks represent family-specific regions (E1–3) and the two
blocks of tandem repeats (R1 and R2) are indicated. The numbers
represent amino acid positions corresponding to published sequences
of 3D7 strain. The arrows show the location of the primers used for
PCR (2.1 and 2.2) and sequencing (A and B).
Fig. 3. Schematic representation of the predicted FC27 MSP-2 protein
sequences from Peixoto de Azevedo isolates. The central variable re-
gion of each allele is compared to the FC27 reference strain (Smythe et
al., 1988), which is represented in the upper position. Unfilled blocks of
FC27 protein sequence indicate conserved regions. The dotted blocks
represent family-specific regions (E1–3) and the two blocks of tandem
repeats (R1 and R2) are indicated. The numbers represent amino acid
positions corresponding to published sequence of FC27 strain. The
arrows show the location of the primers used for PCR (2.1 and 2.2)
and sequencing (A and B).
S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135 131
detected, but only two of them were situated in positions
not shared by at least two FC27 alleles (Fig. 4). In
contrast, the variability observed in repetitive sequences
of FC27, E2 (family specific region 2) and E3 (family
specific region 3) regions was extremely conserved in all
alleles.
The sequence modifications of 3D7 antigenic family
were mainly situated in the R1 and E2 regions, i.e., thetwo most polymorphic segments of the molecule in our
study (Fig. 5). However, as described to FC27 alleles,
several nucleotide substitutions occurred in shared
Fig. 4. Alignment of the predicted amino acid sequence of MSP-2 of the FC2
region of allelic types is compared to the reference strain FC27 sequence (
represent no homology residues and have been inserted to optimize the alignm
The family-specific regions (E1–3) and the two tandem repeats (R1 and R2)
points of allelic sequences, demonstrating a non-
randomic pattern of distribution. As expected, the first
repetitive region (R1) showed to be extremely poly-morphic, beginning in all alleles with a short variable
segment, followed by multiple copies of 4–10 amino
acids that differed in number, sequence, and length from
one allele to other (Figs. 5 and 6). The richness of gly-
cine (G), serine (S), and alanine (A) residues, besides
the biased codon usage with an exclusive use of NNT
7 allelic family from Peixoto de Azevedo isolates. The central variable
Smythe et al., 1988). Residues identical are indicated by dots. Gaps
ent of the sequence. The first residue of each unit repeat is underlined.
are indicated.
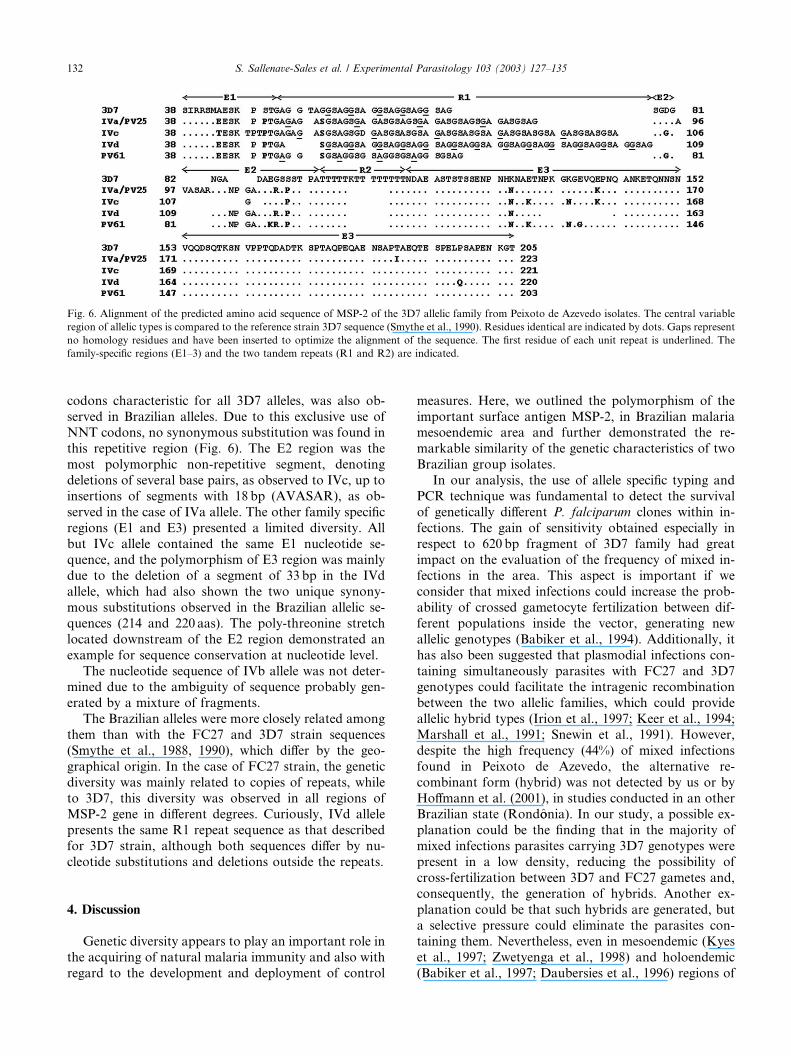
Fig. 6. Alignment of the predicted amino acid sequence of MSP-2 of the 3D7 allelic family from Peixoto de Azevedo isolates. The central variable
region of allelic types is compared to the reference strain 3D7 sequence (Smythe et al., 1990). Residues identical are indicated by dots. Gaps represent
no homology residues and have been inserted to optimize the alignment of the sequence. The first residue of each unit repeat is underlined. The
family-specific regions (E1–3) and the two tandem repeats (R1 and R2) are indicated.
132 S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135
codons characteristic for all 3D7 alleles, was also ob-
served in Brazilian alleles. Due to this exclusive use ofNNT codons, no synonymous substitution was found in
this repetitive region (Fig. 6). The E2 region was the
most polymorphic non-repetitive segment, denoting
deletions of several base pairs, as observed to IVc, up to
insertions of segments with 18 bp (AVASAR), as ob-
served in the case of IVa allele. The other family specific
regions (E1 and E3) presented a limited diversity. All
but IVc allele contained the same E1 nucleotide se-quence, and the polymorphism of E3 region was mainly
due to the deletion of a segment of 33 bp in the IVd
allele, which had also shown the two unique synony-
mous substitutions observed in the Brazilian allelic se-
quences (214 and 220 aas). The poly-threonine stretch
located downstream of the E2 region demonstrated an
example for sequence conservation at nucleotide level.
The nucleotide sequence of IVb allele was not deter-mined due to the ambiguity of sequence probably gen-
erated by a mixture of fragments.
The Brazilian alleles were more closely related among
them than with the FC27 and 3D7 strain sequences
(Smythe et al., 1988, 1990), which differ by the geo-
graphical origin. In the case of FC27 strain, the genetic
diversity was mainly related to copies of repeats, while
to 3D7, this diversity was observed in all regions ofMSP-2 gene in different degrees. Curiously, IVd allele
presents the same R1 repeat sequence as that described
for 3D7 strain, although both sequences differ by nu-
cleotide substitutions and deletions outside the repeats.
4. Discussion
Genetic diversity appears to play an important role in
the acquiring of natural malaria immunity and also with
regard to the development and deployment of control
measures. Here, we outlined the polymorphism of the
important surface antigen MSP-2, in Brazilian malariamesoendemic area and further demonstrated the re-
markable similarity of the genetic characteristics of two
Brazilian group isolates.
In our analysis, the use of allele specific typing and
PCR technique was fundamental to detect the survival
of genetically different P. falciparum clones within in-
fections. The gain of sensitivity obtained especially in
respect to 620 bp fragment of 3D7 family had greatimpact on the evaluation of the frequency of mixed in-
fections in the area. This aspect is important if we
consider that mixed infections could increase the prob-
ability of crossed gametocyte fertilization between dif-
ferent populations inside the vector, generating new
allelic genotypes (Babiker et al., 1994). Additionally, it
has also been suggested that plasmodial infections con-
taining simultaneously parasites with FC27 and 3D7genotypes could facilitate the intragenic recombination
between the two allelic families, which could provide
allelic hybrid types (Irion et al., 1997; Keer et al., 1994;
Marshall et al., 1991; Snewin et al., 1991). However,
despite the high frequency (44%) of mixed infections
found in Peixoto de Azevedo, the alternative re-
combinant form (hybrid) was not detected by us or by
Hoffmann et al. (2001), in studies conducted in an otherBrazilian state (Rondoonia). In our study, a possible ex-
planation could be the finding that in the majority of
mixed infections parasites carrying 3D7 genotypes were
present in a low density, reducing the possibility of
cross-fertilization between 3D7 and FC27 gametes and,
consequently, the generation of hybrids. Another ex-
planation could be that such hybrids are generated, but
a selective pressure could eliminate the parasites con-taining them. Nevertheless, even in mesoendemic (Kyes
et al., 1997; Zwetyenga et al., 1998) and holoendemic
(Babiker et al., 1997; Daubersies et al., 1996) regions of

S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135 133
Africa, where multiclonal infections are prevalent andpresumably the rate sexual reproduction is higher, this
event is rare. Thus, independently of the endemicity of
the area, how frequently new genotypes are introduced
into a population and how they are maintained seem to
be crucial points to better understand the generation of
hybrid types.
The microheterogeneity sequences suggested by the
different degrees of probe homology in the allelic typing(data not shown) were confirmed by the SSCP technique
and sequencing analysis. The two groups of allelic
families presented a sequence polymorphism very simi-
lar to that already described where the inter-allelic MSP-
2 diversity was mainly related to repetitive regions
(Felger et al., 1997; Marshall et al., 1994). One of the
consequences of these variations was the size polymor-
phism, which was less evident among 3D7 alleles, be-cause the number and size alterations of the repeats were
compensated by deletions and insertions in other points
of sequence.
A picture of strong conservation in repeat structure
and nucleotide sequence was observed for FC27 alleles.
It is possible that mechanisms of amplification and/or
unequal recombination during the meiosis could con-
tribute to generate the FC27 allelic variation. These twoprocesses could explain, for example, the duplication
and the deletion of repeats units in R1 and R2 regions of
the III allele, respectively (Fig. 3). In this sense, the II
allele could have evolved from Ia by deleting one repeat
unit or Ia could have derived from II by repeat ampli-
fication. Despite the variability of the two repetitive
units among the isolates, some underlying patterns can
be clearly observed suggesting an ancestral repeat unit.The complete deletion of R2 region (32 bp) seems to be
frequent, as this deletion has also been observed in
isolates from other endemic areas (Felger et al., 1994;
Marshall et al., 1994; Ranford-Cartwright et al., 1996).
This finding led to the conclusion that this repetitive
region is not important to the parasite, since the para-
sites without this region are able to survive. In contrast,
the complete deletion of R1 region (96 bp unit) has notbeen described until now. Moreover, this region con-
tains the epitope STNS that is considered the reactivity
point for blocking monoclonal antibodies, suggesting its
importance in the invasion process (Epping et al., 1988).
The most striking feature of 3D7 nucleotide repeat
sequence was the exclusive presence of NNT codons,
which clearly causes a strong bias in codon usage and
can contribute considerably to increase antigenic diver-sity. It has been suggested by Felger et al. (1997) that
NNT codon exclusivity results from homogenization
mechanisms at nucleotide sequence level, such as un-
equal crossing over and biased gene conversion. In this
case, these proposed events acting only at the DNA level
would not necessarily reflect selection acting on the
amino acid sequence that would not maintain the
homogeneity between repeats. On the other hand, themajor sources of interallelic diversity in 3D7 allelic
family have been proposed to be replication slippage,
causing either duplication or deletion of a hexamer
GGT GCT or multiples of it, and mitotic or meiotic
unequal exchange, which can also increase or reduce the
copy number of repeats (Felger et al., 1997). In this
light, the presence of identical repeats in IVd, PV25, and
PV61 alleles could be explained by amplification mech-anisms, whereas the imperfect nucleotide repeats as seen
in IVa and IVc alleles may be caused by DNA poly-
merase slippage.
The majority of point mutations, observed in both
Brazilian allelic groups, conducted to non-synonymous
substitutions that could contribute to increase the anti-
genic diversity. Several of such mutations occurred in
specific sites of sequences demonstrating a non-randomdistribution pattern that might indicate an immunolog-
ical significance if clustered in putative epitopes involved
in the parasite evasion mechanism. The position-specific
feature of some mutations could suggest the existence of
typical patterns, although these patterns are not exclu-
sive of Brazilian endemic areas (Felger et al., 1994, 1997;
Marshall et al., 1994).
Our data suggest that the P. falciparum parasitepopulations of MSP-2 circulating in two Brazilian
communities are genetically homogeneous and tempo-
rally stable, since isolates from Peixoto de Azevedo and
Porto Velho presented MSP-2 alleles with identical or
similar sequence of central variable regions. In addition,
similar to the data reported by Hoffmann et al. (2000),
there was no remarkable evidence of differences in allelic
family frequencies and infection complexity between thetwo areas, in spite of the fact that P. falciparum isolates
had been collected in two different periods and in geo-
graphically distinct areas. Considering the extensive
polymorphism of MSP-2 gene, the finding of identical
sequences or different combinations of the motifs in
isolates from different geographical areas might be un-
expected. It is possible that all MSP-2 alleles circulating
in wild parasite populations were derived from a com-mon ancestor (Doba~nno et al., 1997) or alternatively the
extensive homoplasy results from unifying selection of
MSP-2 alleles in separate parasite lineages (McCutchan
et al., 1992).
The simplicity of the allelic profiles from isolates of
Peixoto de Azevedo, when compared to those observed
in highly endemic areas such as Senegal (Daubersies
et al., 1996), Tanzania (Babiker et al., 1999; Felger et al.,1999), and Papua New Guinea (Felger et al., 1994), was
consistent with the endemicity of the Brazilian endemic
areas. Thus, diversity is expected to be correlated with
transmission intensity, since in holoendemic areas the
higher complexity of human infections could be a result
of a major exposition to different parasite genotypes
carried by the mosquito vector. On the other hand, the

134 S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135
fact that the samples were obtained from symptomaticindividuals should also be considered an alternative or
additional explanation for the limited diversity ob-
served, since minor quantitative and qualitative varia-
tions in the parasite antigen alleles harbored by
symptomatic individuals had been reported (Daubersies
et al., 1994; Sallenave-Sales et al., 2000). In fact, the
higher frequency of a multiplex pattern observed among
asymptomatic infections is claimed to be related topremunition state (Babiker et al., 1997; Daubersies et al.,
1996).
Our molecular epidemiological studies clearly showed
an important, although restricted, genetic diversity of
MSP-2 in two Brazilian endemic areas. Considering that
the genetic diversity would not necessarily reflect selec-
tion acting at protein level, works are in progress to
associate these data with MSP-2 antibody reactivity inorder to evaluate the impact of this polymorphism on
the immune response to MSP-2 antigens. Moreover,
this approach could also open the possibility of investi-
gating the clinical impact of a particular MSP-2 geno-
type, a crucial question in understanding P. falciparum
pathogenesis.
Acknowledgments
We thank Dr. Odile Mercereau Puijalon for the
provision of the MSP-2 probes and all the patients of
Peixoto de Azevedo who volunteered to provide us with
blood samples. We also thank the help of Drs. Adeilton
Alves Brand~aao and Alberto D��AAvila, Department of
Biochemistry and Molecular Biology, and Dr. Rosa,Department of Genetics of Institute Oswaldo Cruz. This
work was supported by CNPq (Brazilian Research
Council) and Fundac�~aao Oswaldo Cruz (Fiocruz).
References
Al-Yaman, F., Genton, B., Reeder, J.C., Mokela, D., Anders, R.F.,
Alpers, M.P., 1997. Humoral response to defined Plasmodium
falciparum antigens in cerebral and uncomplicated malaria and
their relationship to parasite genotype. American Journal of
Tropical Medicine Hygiene 56, 430–435.
Anders, R.F., Mc Coll, D.J., Coppel, R.L., 1993. Molecular variation
in Plasmodium falciparum: polymorphic antigens of asexual erytr-
ocytic stages. Acta Tropica 53, 239–253.
Babiker, H.A., Ranford-Cartwright, L.C., Currie, D., Charlwood,
J.D., Billingsley, P., Teuscher, T., Walliker, D., 1994. Random
mating in a natural population of the malaria parasite Plasmodium
falciparum. Parasitology 109, 413–421.
Babiker, H.A., Lines, J., Hill, W.G., Walliker, D., 1997. Population
structure of Plasmodium falciparum in villages with different
malaria endemicity in east Africa. American Journal of Tropical
Medicine Hygiene 56, 141–147.
Babiker, H.A., Ranford-Cartwright, L.C., Walliker, D., 1999. Epide-
miology of multiple Plasmodium falciparum infections. 3. Genetic
structure and dynamics of Plasmodium falciparum infections in the
Kilombero region of Tanzania. Transactions of the Royal Society
of Tropical Medicine and Hygiene 93, S1/11–S1/14.
Contamin, H., Fandeur, T., Bonnefoy, S., Skouri, F., Ntoumi, F.,
Mercereau-Puijalon, O., 1995. PCR typing of field isolates of
Plasmodium falciparum. Journal of Clinical Microbiology 33,
944–951.
Creasey, A., Fenton, B., Walker, A., Thaithong, S., Oliveira, S.,
Mutanbu, S., Walliker, D., 1990. Genetic diversity of Plasmodium
falciparum shows geographical variation. American Journal of
Tropical Medicine and Hygiene 42, 403–413.
Daubersies, P., Sallenave-Sales, S., Trape, J.F., Raharimalala, L.,
Rogier, C., Contamin, H., Fandeur, T., Daniel-Ribeiro, C.T.,
Mercereau-Puijalon, O., Druilhe, P., 1994. PCR characterization of
isolates from various endemic areas: diversity and turn over of
Plasmodium falciparum populations are correlated with transmis-
sion. Mem�oorias do Instituto Oswaldo Cruz 89, 9–12.
Daubersies, P., Sallenave-Sales, S., Magne, S., Trape, J.F., Contamin,
H., Fandeur, T., Rogier, C., Mercereau-Puijalon, O., Druilhe, P.,
1996. Rapid turnover of Plasmodium falciparum populations in
asymptomatic individuals living in a high transmission area.
American Journal of Tropical Medicine and Hygiene 54, 18–26.
Doba~nno, C., Khan, A., Robinson, J.V., Taylor, R.R., McBride, J.S.,
1997. Identical alleles of Plasmodium falciparum merozoite surface
protein 2 in distant geographic areas and times. Parasitology
International 46, 137–142.
Druilhe, P., Daubersies, P., Patarapotikul, J., Gentil, C., Chene, L.,
Chongsuphajaisiddhi, T., Mellouk, S., Langsley, G., 1998. A
primary malarial infection is composed of a very wide range of
genetically diverse but related parasites. Journal of Clinical
Investigation 101, 2008–2016.
Duarte, E.C., Fontes, C.J.F., 1998. Associac�~aao entre o �ıındiceparasit�aario anual de mal�aaria e produc�~aao garimpeira de ouro em
Mato Grosso—1989–1986. Revista da Sociedade Brasileira de
Medicina Tropical 31, I.
Engelbrecht, F., Felger, I., Genton, B., Alpers, M., Beck, H., 1995.
Plasmodium falciparum: Malaria morbidity is associated with
specific merozoite surface antigen 2 genotypes. Experimental
Parasitology 81, 90–96.
Epping, R.J., Goldstone, S.D., Ingram, L.T., Ucproft, J.A., Ramasay,
R., Cooper, J.A., Bushell, G.R., Geysen, H.M., 1988. An epitope
recognised by inhibitory monoclonal antibodies that react with a 51
kilodalton merozoite surface antigen in Plasmodium falciparum.
Molecular and Biochemical Parasitology 28, 1–10.
Felger, I., Tavul, L., Kabintik, S., Marshall, V., Genton, B., Alpers,
M., Beck, H.P., 1994. Plasmodium falciparum extensive polymor-
phism in merozoite surface antigen 2 alleles in an area with endemic
malaria in Papua New Guinea. Experimental Parasitology 79,
106–116.
Felger, I., Marshal, V.M., Reeder, J.C., Hunt, J.A., Mgone, C.S.,
Beck, H., 1997. Sequence diversity and molecular evolution of the
merozoite surface antigen 2 Plasmodium falciparum. Journal of
Molecular Evolution 45, 154–160.
Felger, I., Irion, A., Steiger, S., Beck, H.P., 1999. Epidemiology of
multiple Plasmodium falciparum infections. 2. Genotypes of mer-
ozoite surface protein 2 of Plasmodium falciparum in Tanzania.
Transactions of the Royal Society of Tropical Medicine and
Hygiene 93, S1/3–S1/9.
Genton, B., Al-Yaman, F., Anders, R., Saul, A., Brown, G., Pye, D.,
Irving, D.O., Briggs, W.R.S., Mai, A., Ginny, M., Adiguma, T.,
Rare, L., Giddy, A., Reber-Liske, R., Sturchler, D., Alpers, M.P.,
2000. Safety and immunogenicity of a three-component blood-
stage malaria vaccine in adults living in an endemic area of Papua
New Guinea. Vaccine 18, 2504–2511.
Genton, B., Betuela, I., Felger, I., Al-Yaman, F., Anders, R.F., Saul,
A., Rare, L., Baisor, M., Lorry, K., Brown, G.V., Pye, D., Irving,
D.O., Smith, T.A., Beck, H.P., Alpers, M.P., 2002. A recombinant
blood-stage malaria vaccine reduces Plasmodium falciparum density

S. Sallenave-Sales et al. / Experimental Parasitology 103 (2003) 127–135 135
and exerts selective pressure on parasite populations in a phase
1-2b trial in Papua New Guinea. Journal of Infectious Diseases
185, 820–827.
Gonc�alves, A.M., Nehme, N.S., Morel, C., 1990. An improved silver
staining procedure for schizodeme analysis in polyacrylamide
gradient gels. M�eemorias do Instituto Oswaldo Cruz 85, 101–106.
Gupta, S., Trenholme, K., Anderson, R.M., Day, K.P., 1994.
Antigenic diversity and the transmission dynamics of Plasmodium
falciparum. Science 263, 961–963.
Hoffmann, E.H., da Silveira, L.A., Tonhosolo, R., Pereira, F.J.,
Ribeiro, W.L., Tonon, A.P., Kawamoto, F., Ferreira, U.M., 2001.
Geographical patterns of allelic diversity in the Plasmodium
falciparum malaria-vaccine candidate, merozoite surface protein-
2. Annals of Tropical Medicine and Parasitology 95, 117–132.
Irion, A., Beck, H., Felger, I., 1997. New repeat unit and hot spot of
recombination in FC27-type alleles of the gene coding for
Plasmodium falciparum merozoite surface protein 2. Molecular
and Biochemical Parasitology 90, 367–370.
Keer, P.J., Ranford-Cartwright, L.C., Walliker, D., 1994. Proof of
intragenic recombination in Plasmodium falciparum. Molecular and
Biochemical Parasitology 29, 75–147.
Kyes, S., Harding, R., Black, G., Graig, A., Peshu, N., Newblod, C.,
Marsh, K., 1997. Limited spatial clustering of individual
Plasmodium falciparum alleles in field isolates from coastal
Kenya. American Journal of Tropical Medicine and Hygiene
57, 205–215.
Lawrence, G., Cheng, Q., Reed, C., Taylor, D., Stowers, A., Cloonan,
N., Rzepczyk, C., Smillie, A., Anderson, K., Pombo, D., Allworth,
A., Eisen, D., Anders, R., Saul, A., 2000. Effect of vaccination with
3 recombinant asexual-stage malaria antigens on initial growth
rates of Plasmodium falciparum in non-immune volunteers. Vaccine
18, 1925–1931.
Marshall, V.M., Coppel, R.L., Martin, R.K., Oduola, A.M.J.,
Anders, R.F., Kemp, D.J., 1991. A Plasmodium falciparum
MSA-2 gene apparently generated by intragenic recombination
between the two allelic families. Molecular and Biochemical
Parasitology 63, 13–21.
Marshall, V.M., Anthony, R.L., Bangs, M.J., Purnomo, T., Anders,
R.F., Coppel, R.L., 1994. Allelic variants of the Plasmodium
falciparum merozoite surface antigen 2 (MSA-2) in a geographi-
cally restricted area of Irian Jaya. Molecular and Biochemical
Parasitology 63, 13–21.
McCutchan, T.F., Lal, A.A., Do Ros�aario, V.E., Waters, A.P., 1992.
Two types of sequence polymorphism in the circumsporozoite gene
of Plasmodium falciparum. Molecular and Biochemical Parasitol-
ogy 50, 37–46.
Mercereau-Puijalon, O., 1996. Revisiting host/parasite interactions:
molecular analysis of parasites collected during longitudinal and
cross-sectional surveys in humans. Parasite Immunology 18, 173–
180.
Muller, D.A., Charlwood, J.D., Felger, I., Ferreira, C., do Ros�aario, V.,
Smith, T., 2001. Prospective risk of morbidity in relation to
multiplicity of infection with Plasmodium falciparum in Sao Tome.
Acta Tropica 69, 996–1001.
Ntoumi, F., Contamin, H., Rogier, C., Bonnefoy, S., Trape, J.F.,
Mercereau-Puijalon, O., 1995. Age-dependent carriage of multiple
Plasmodium falciparum merozoite surface antigen-2 alleles in
asymptomatic malaria infections. American Journal of Tropical
Medicine and Hygiene 52, 81–88.
Paul, R.E.L., Packer, M.J., Walmsley, M., Lagog, M., Ranford-
Cartwright, L.C., Paru, R., Day, K.P., 1995. Mating patterns in
malaria parasite populations of Papua New Guinea. Science 269,
1709–1711.
Ranford-Cartwright, L.C., Taylor, R.R., Asgari-Jirhandeh, N., Smith,
D.B., Roberts, P.E., Robinson, V.J., Babiker, H.A., Riley, E.M.,
Walliker, D., McBride, J.S., 1996. Differential antibody recognition
of FC27–like Plasmodium falciparum merozoite surface protein
MSP-2 antigens which lack 12 amino acid. Parasite Immunology
18, 411–420.
Sallenave-Sales, S., Daubersies, P., Mercereau-Puijalon, O., Rahimal-
ala, L., Contamin, H., Druilhe, P., Daniel-Ribeiro, C.T., Ferreira-
da-Cruz, 2000. Plasmodium falciparum: a comparative analysis of
the genetic diversity in malaria-mesoendemic areas of Brazil and
Madagascar. Parasitology Research 86, 692–698.
Smythe, J.A., Coppel, R.L., Brown, G.V., Ramasamy, R., Kemp, D.J.,
Anders, R.F., 1988. Identification of two integral membrane
proteins of Plasmodium falciparum. Proceedings National Academy
Science of the USA 85, 5195–5199.
Smythe, J.A., Peterson, M.G., Coppel, R.L., Saul, A.J., Kemp, D.J.,
Anders, R.F., 1990. Structural diversity in the 45-kilodalton
merozoite surface antigen of Plasmodium falciparum. Molecular
and Biochemical Parasitology 39, 227–234.
Snewin, V.A., Herrera, M., Sanchez, G., Scherf, A., Langsley, G.,
Herrera, S., 1991. Polymorphism of the alleles of the merozoite
surface antigens MSA1 and MSA2 in Plasmodium falciparum wild
isolates from Colombia. Molecular and Biomedical Parasitology
49, 265–276.
Zevering, Y., Khamboonruang, C., Good, M.F., 1994. Effect of
polymorphism of sporozoite antigens on T-cell activation. Re-
search Immunology 145, 469–476.
Zwetyenga, J., Rogier, C., Tall, A., Fontenille, D., Snounou, G.,
Trape, J.-F., Mercereau-Puijalon, O., 1998. No influence of age on
infection complexity and allelic distribution in Plasmodium falci-
parum infections in Ndiop, a Senegalese village with seasonal,
mesoendemic malaria. American Journal of Tropical Medicine and
Hygiene 59, 726–735.
Copyright © 2022 FDOKUMEN




















