Ultrastructure of erythrocytes from Aotus trivirgatus and Saimiri sciureus monkeys infected by...
7
Parasitol Res (1991) 77:395 401 004432559100057W Parasitnlngy Research Springer-Verlag1991 Ultrastructure of erythrocytes from Aotus trivirgatus and SMmiri sciureus monkeys infected by Plasmodium vivax H, Norbert Lanners* Delta Regional Primate Research Center, 18703 Three Rivers Road, Covington, LA 70433, USA and Department of Tropical Medicine, School of Public Health and Tropical Medicine, Tulane University Medical Center, 1501 Canal Street, New Orleans, LA 70112, USA Accepted December 20, 1990 Abstract. Erythrocytes from Aotus and Saimiri monkeys parasitized by Plasmodium vivax show dramatic changes starting during the early stages of parasite development. Invaginations of the erythrocyte membrane, caveolae, are found during all parasite development stages. Up to six vesicles can be fused with one caveola, forming a caveola-vesicle complex. As the parasite grows, large accumulations of these vesicles can be seen within the erythrocyte cytoplasm. In addition to these caveolae- vesicle complexes, knob-like structures appear on the erythrocyte surface that are similar to those seen on the host-cell surface of P. falciparum-infected red cells. Ex- tensive membrane-bound clefts spread throughout the erythrocytic cytoplasm, sometimes forming stacks or large whorls. The density of the red cell cytoplasm begins to decrease at an early stage of parasite development. All of these changes may be responsible for an increased fragility of the P. vivax-infected red cell from Aotus or Saimiri monkeys. Moreover, the large amount of para- site material that is released during rupture of the red cell may account for the high fever paroxysms that are characteristic of P. vivax malaria infection. The malarial parasite Plasmodium vivax is one of the major scourges of mankind, causing high morbidities of benign tertian malaria in the tropics and subtropics. Although it is not as deadly as P. faleiparum, P. vivax nevertheless causes a most debilitating disease character- ized by intermittent high fever paroxysms (Boyd 1949). An understanding of the biology and pathogenicity of this parasite is of great interest, particularly in view of future drug and vaccine developments. Notes on the ultrastructure of P. vivax and related species have previously been reported (Aikawa 1977, 1988; Aikawa etal. 1975, 1977; Mackenstedt etal. 1989). The present report attempts to summarize the * Offprint requests to: Delta Regional Primate Research Center, 18703 Three Rivers Road, Covington, LA 70433, USA pathology caused by the parasite on and within the host cell. As a new finding, knobs appearing on the infected erythrocyte surface are described. Materials and methods Blood from two splenectomized Aotus lemurinus griseimembra was used. These animals had previously been infected with Plasmodium falciparum during an active immunization experiment (Trager et al. 1983). They were infected with blood from another Aotus monkey by inoculation of approximately 2 x 10 6 parasites into the saphen- ous vein. The parasite strain that was used originated from CDC (South Vietnam Palo Alto strain) and was passed through five animals before the monkeys in the present study were inoculated. The infection rate in these Aotus monkeys seldom rose above 1%. Blood from three splenectomized Saimiri sciureus monkeys of the Bolivian phenotype was inoculated into the saphenous vein of ex- perimental animals along with cryopreserved material obtained from the American Type Culture Collection (Rockville, Md., Ches- son strain, ATCC 30060). The history of simian adaptation of this material is not known. All blood was collected by venipuncture into preservative-free heparin after monkeys had been restrained by the injection of 10 mg/kg ketamine. Parasitemia was monitored by frequent exami- nation of Giemsa-stained thin blood films. Blood cells were addi- tionally stained with brilliant cresyl blue (Retic-Set, Curtin Mathe- son Scientific, Inc.) for identification of reticulocytes. Under gentle stirring, infected blood was slowly added to a fixative consisting of 2% (v/v) glutaraldehyde, 4% (w/v) sucrose, and 20 mM CaClz, in 0.1 M Na-cacodylate-HC1 buffer at pH 7.3, on ice. In some experiments the cacodylate buffer was replaced by 50 mM PIPES buffer. The cells were postfixed in 1% (w/v) OsO4, stained en bloc with uranyl acetate, dehydrated in ethanol, and embedded in Epon 812 via propylene oxide. Stained sections were examined with a Philips 300 or a JEOL 1200EXII electron microscope. Tracer studies were done using microperoxidase (Kar- novsky and Rice 1969) and kationized ferritin (Miles-Yeda; see Kawakami and Hirano 1986). Results Ultrastructure of the parasite The ultrastructure of the parasite Plasmodium vivax is only slightly different from the reported fine structure
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Ultrastructure of erythrocytes from Aotus trivirgatus and Saimiri sciureus monkeys infected by...

Parasitol Res (1991) 77:395 401 004432559100057W Parasitnlngy
Research �9 Springer-Verlag 1991
Ultrastructure of erythrocytes from Aotus trivirgatus and SMmiri sciureus monkeys infected by Plasmodium vivax H, Norbert Lanners*
Delta Regional Primate Research Center, 18703 Three Rivers Road, Covington, LA 70433, USA and Department of Tropical Medicine, School of Public Health and Tropical Medicine, Tulane University Medical Center, 1501 Canal Street, New Orleans, LA 70112, USA
Accepted December 20, 1990
Abstract. Erythrocytes f rom Aotus and Saimiri monkeys parasitized by Plasmodium vivax show dramatic changes starting during the early stages of parasite development. Invaginations of the erythrocyte membrane , caveolae, are found during all parasite development stages. Up to six vesicles can be fused with one caveola, forming a caveola-vesicle complex. As the parasite grows, large accumulations of these vesicles can be seen within the erythrocyte cytoplasm. In addition to these caveolae- vesicle complexes, knob-like structures appear on the erythrocyte surface that are similar to those seen on the host-cell surface of P. falciparum-infected red cells. Ex- tensive membrane-bound clefts spread throughout the erythrocytic cytoplasm, sometimes forming stacks or large whorls. The density of the red cell cytoplasm begins to decrease at an early stage of parasite development. All of these changes may be responsible for an increased fragility of the P. vivax-infected red cell f rom Aotus or Saimiri monkeys. Moreover, the large amount of para- site material that is released during rupture of the red cell may account for the high fever paroxysms that are characteristic of P. vivax malaria infection.
The malarial parasite Plasmodium vivax is one of the major scourges of mankind, causing high morbidities of benign tertian malar ia in the tropics and subtropics. Although it is not as deadly as P. faleiparum, P. vivax nevertheless causes a most debilitating disease character- ized by intermittent high fever paroxysms (Boyd 1949). An understanding of the biology and pathogenicity of this parasite is of great interest, particularly in view of future drug and vaccine developments.
Notes on the ultrastructure of P. vivax and related species have previously been reported (Aikawa 1977, 1988; Aikawa eta l . 1975, 1977; Mackenstedt e ta l . 1989). The present report a t tempts to summarize the
* Offprint requests to: Delta Regional Primate Research Center, 18703 Three Rivers Road, Covington, LA 70433, USA
pathology caused by the parasite on and within the host cell. As a new finding, knobs appearing on the infected erythrocyte surface are described.
Materials and methods
Blood from two splenectomized Aotus lemurinus griseimembra was used. These animals had previously been infected with Plasmodium falciparum during an active immunization experiment (Trager et al. 1983). They were infected with blood from another Aotus monkey by inoculation of approximately 2 x 10 6 parasites into the saphen- ous vein. The parasite strain that was used originated from CDC (South Vietnam Palo Alto strain) and was passed through five animals before the monkeys in the present study were inoculated. The infection rate in these Aotus monkeys seldom rose above 1%. Blood from three splenectomized Saimiri sciureus monkeys of the Bolivian phenotype was inoculated into the saphenous vein of ex- perimental animals along with cryopreserved material obtained from the American Type Culture Collection (Rockville, Md., Ches- son strain, ATCC 30060). The history of simian adaptation of this material is not known.
All blood was collected by venipuncture into preservative-free heparin after monkeys had been restrained by the injection of 10 mg/kg ketamine. Parasitemia was monitored by frequent exami- nation of Giemsa-stained thin blood films. Blood cells were addi- tionally stained with brilliant cresyl blue (Retic-Set, Curtin Mathe- son Scientific, Inc.) for identification of reticulocytes.
Under gentle stirring, infected blood was slowly added to a fixative consisting of 2% (v/v) glutaraldehyde, 4% (w/v) sucrose, and 20 mM CaClz, in 0.1 M Na-cacodylate-HC1 buffer at pH 7.3, on ice. In some experiments the cacodylate buffer was replaced by 50 mM PIPES buffer. The cells were postfixed in 1% (w/v) OsO4, stained en bloc with uranyl acetate, dehydrated in ethanol, and embedded in Epon 812 via propylene oxide. Stained sections were examined with a Philips 300 or a JEOL 1200EXII electron microscope. Tracer studies were done using microperoxidase (Kar- novsky and Rice 1969) and kationized ferritin (Miles-Yeda; see Kawakami and Hirano 1986).
Results
Ultrastructure o f the parasite
The ultrastructure of the parasite Plasmodium vivax is only slightly different f rom the reported fine structure

396 H.N. Lanners : Erythrocytes infected by Plasmodium vivax

H.N. Lanners: Erythrocytes infected by P&smodium vh)ax 397
of P. falciparum. The earliest stage, the ring stage (Fig. 1), is cup-shaped and therefore appears in cross section as a ring, with its nucleus bending around a cen- tral erythrocytic plasma portion. Within the cytoplasm, organelles found at this stage include cytostomes; small phagocytic vacuoles; pigment granules; and simple, acr- istate mitochondria.
At the trophozoite stage (Fig. 3), the parasitophorous vacuole has expanded greatly and sections show several profiles filled with a flocculent material. The nucleus is elongated such that several sections are visible in a micrograph. Except for small, electron-dense granules, no distinct structures can be discerned within the nuc- leoplasm. Small, stacked Golgi vesicles are present (Fig. 3, insert), and both rough and smooth endoplasmic reticulum (ER) are abundant (Fig. 3), displaying pro- nounced connections to the nuclear envelope. Pigment is not concentrated in a large vacuole, as in P. falcipar- urn, but is distributed in small (0.1 gm) granules throughout the cytoplasm. Even at high magnifications, the micrographs did not reveal any structural basis for the trophozoite motility that is so prominent in P. vivax. The merozoites of the schizont stage show the same ul- trastructure as those of P. falciparurn, exhibiting pellicu- lar membranes, rhoptries, and micronemes (see Figs. 10, 11).
Erythrocyte modifications
All stages of erythrocytes from our experimental animals could be invaded by P. vivax merozoites. No bluish- staining cells (as seen in Giemsa-stained thin blood smears) or erythrocytes with reticulum (after staining with brilliant cresyl blue for reticulocytes) were preferen- tially invaded as reported elsewhere (Kitchen 1938). Consequently, we found parasites in mature erythrocytes as well as in reticulocytes. During a P. vivax infection,
Fig. 1. Ring stage of Plasmodium vivax. The parasite bends around an extension of the erythrocytic cytoplasm (asterisk). The micro- graph shows caveolae with associated vesicles (arrowhead) in addi- tion to cytoplasmic clefts (C). One cleft is sectioned tangentially. The parasite contains small food vacuoles (arrows) and a small pigment granule (Pi). Other visible organelles include the nucleus (N) and ER studded with ribosomes. • 16500
Fig. 2. Small portion of a parasite (P) within a nucleated erythro- cyte. Note the condensed host-cell nucleus (N). The erythrocytic cytoplasm contains numerous polyribosomes and rhopheocytotic vesicles (R). x 15500
Fig. 3. Late trophozoite stage of P. vivax. The cell shows abundant rough endoplasmic reticulum, two pigment granules (P), and mito- chondria (M). Two nuclear profiles (N) are also visible. In addition to extensive membrane whorls within the erythrocytic cytoplasm, several branches of the parasitophorous vacuole (PIO are sec- tioned. The erythrocyte membrane shows several caveolae-vesicle complexes, x 23 500. Insert: Stack of Golgi vesicles within the cyto- plasm of a parasite (arrowhead). x 23 500
Aotus monkey 104 experienced a vitamin E-responsive anemia with a high incidence of normoblasts. The blood from this animal showed parasites even within these nuc- leated erythrocytes (Fig. 2).
As early as at the ring stage, the infected erythrocytes exhibit dramatic modifications (Fig. 1). Some changes involve the erythrocyte surface, which shows invagina- tions of the cell membrane. These invaginations, or ca- veolae, have been related to Schfiffner's dots (Aikawa et al. 1975; Sterling et al. 1975). These structures are an important diagnostic tool for P. vivax-type malaria in- fections and can be seen by light microscopy, as a promi- nent stippling on the erythrocyte surface. The caveolae are of slightly varying size, generally measuring 70 nm in depth and 100 nm in width, and they are present at all stages during the parasite's life in its host cell. Usually caveolae are associated with 50- to 60-nm-wide vesicles with an electron-dense content. In cross sections, up to six vesicles could surround one invagination (Fig. 5). Electron-dense tracers such as kationized ferritin or mi- croperoxidase, which were applied in the course of the present study, did enter the caveolae but could not be found within the associated vesicles even after an incuba- tion time of up to 3 h. As the parasite matures, increas- ingly more electron-dense vesicles are found within the erythrocyfic cytoplasm, until large accumulations of this material are present (Fig. 6).
A second type of erythrocyte membrane change con- cerns knob-like protrusions associated with cup-shaped electron-dense material underneath the erythrocyte cell membrane (Figs. 7-11). This material is directly attached to the erythrocyte membrane and often masks the unit membrane structure at the site of the knob. In P. vivax these knobs are 50 150 nm wide and occur in much smaller numbers than those in P.falciparurn. These alter- ations persist during the late schizont stage (Fig. 11).
A third change in the erythrocyte membrane involves delaminations of the membrane, particularly of the outer leaflet (about 50 x 100 nm; Figs. 4, 6). These observed changes may be fixation artifacts, but the mere observa- tion that they occur more frequently in P. vivax-infected cells than in those infected with P. falciparurn indicates that the erythrocyte membrane's reaction to fixation stress in P. vivax infections is different from that in P. falciparum infections.
In addition to these modifications of the erythrocyte surface, changes also occur within the cytoplasm. Apart from the above-mentioned accumulation of vesicles, membrane-bound clefts appear in the erythrocytic cyto- plasm. Similar clefts occur in P. falciparurn at the tro- phozoite stage and are referred to as Maurer's clefts (Trager et al. 1966). In P. vivax, they appear as early as at the ring stage (Fig. t), and large accumulations of these membranes can be found in later stages of the parasite (Fig. 7). In extreme cases, the erythrocyte cyto- plasm seems to be completely filled with membranes and vesicles, whereas the parasite maintains its normal ap- pearance. This condition is more prominent in erythro- cytes from Sairniri monkeys than in those from Aotus. Membrane whirls often show membrane fusions that are
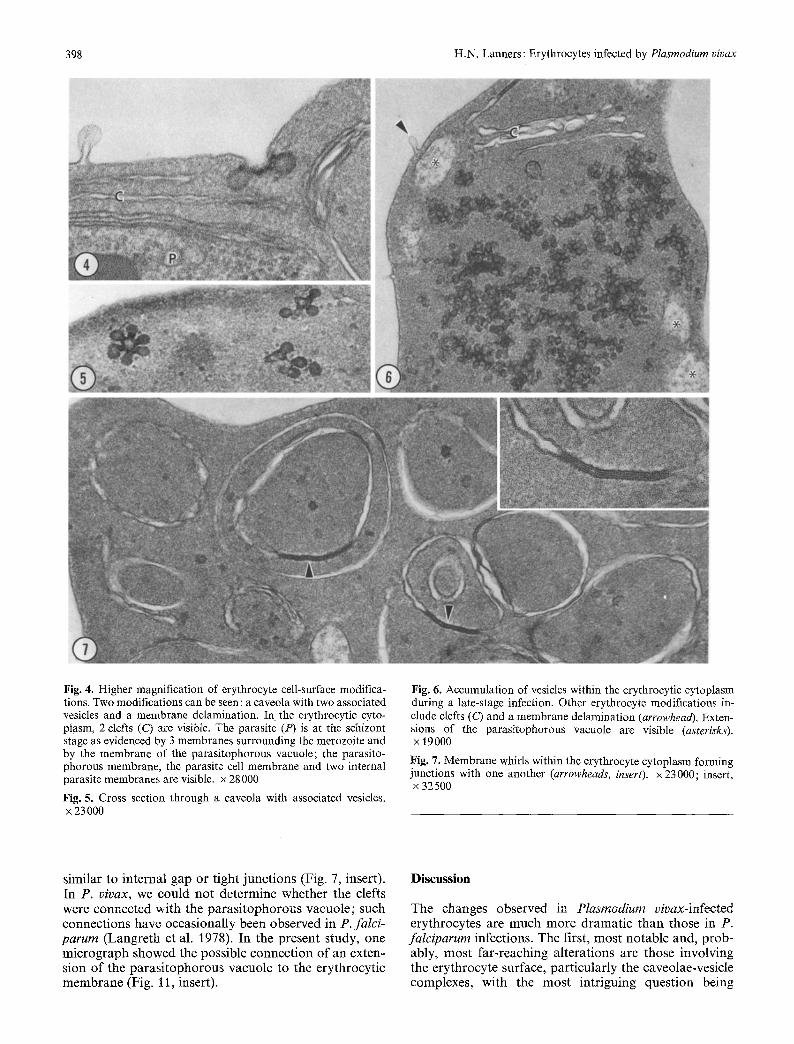
398 H.N. Lanners : Erythrocytes infected by Plasmod&rn vivax
Fig. 4. Higher magnification of erythrocyte cell-surface modifica- tions. Two modifications can be seen : a caveola with two associated vesicles and a membrane delamination. In the erythrocytic cyto- plasm, 2 clefts (C) are visible. The parasite (P) is at the schizont stage as evidenced by 3 membranes surrounding the merozoite and by the membrane of the parasitophorous vacuole; the parasito- phorous membrane, the parasite cell membrane and two internal parasite membranes are visible, x 28000
Fig. 5. Cross section through a caveola with associated vesicles. x 23 000
Fig. 6. Accumulation of vesicles within the erythrocytic cytoplasm during a late-stage infection. Other erythrocyte modifications in- clude clefts (C) and a membrane delamination (arrowhead). Exten- sions of the parasitophorous vacuole are visible (asterisks). x 19000
Fig. 7. Membrane whirls within the erythrocyte cytoplasm forming junctions with one another (arrowheads, insert), x 23000; insert, x 32 500
similar to internal gap or tight junct ions (Fig. 7, insert). In P. vivax, we could no t determine whether the clefts were connected with the pa ras i tophorous vacuole ; such connect ions have occasional ly been observed in P. falci- parurn (Langre th et al. 1978), In the present study, one mic rograph showed the possible connect ion o f an exten- sion o f the pa ras i tophorous vacuole to the erythrocyt ic membrane (Fig. 11, insert).
Discussion
The changes observed in Plasrnodium vivax-infected erythrocytes are much more dramat ic than those in P. faIciparurn infections. The first, mos t notable and, prob- ably, mos t far-reaching alterations are those involving the erythrocyte surface, part icularly the caveolae-vesicle complexes, with the mos t intriguing quest ion being

H.N. Lanners: Erythrocytes infected by Plasmodium vivax 399
Fig. 8. Knob of a Plasmodium vivax-infected erythrocyte next to a caveola, x 23000
Fig. 9. Knob of an eryhtrocyte infected with a trophozoite of P. vivax, x 24000
Fig. 10. Merozoites o fP . vivax within their host cell's parasitophor- ous vacuole. The merozoites are surrounded by 3 membranes and
contain rhoptries (Rh). The arrowhead points to a knob-like struc- ture and the arrow indicates a caveola. • 27000
Fig. 11. Late-stage parasite segmenter showing merozoites within an extensively branched parasi tophorous vacuole (PV). The arrow- head points to a knob on the erythrocyte surface, Rh, Rhoptries. x23000. Insert: The arrowhead points to a possible opening of
the parasi tophorous vacuole to the outside, x 23 000

400 H.N. Lanners: Erythrocytes infected by Plasmodium vivax
whether the vesicles are of endocytic or exocytic nature. Although we could not repeat the results of Aikawa et al. (1975) using electron-dense tracers, we do support the suggestion of these authors that the vesicles are endo- cytic. The accumulation of vesicles within the erythro- cyte cytoplasm and the absence of any vesicles connected with the parasitophorous vacuole supports this notion. Matsumoto et al. (1988) have described the presence of P. vivax antigens within the vesicle complexes and sug- gested that they transport malarial antigen from the par- asite to the host-cell surface. The function of these vesi- cles remains to be elucidated in future research, most likely when a continuous culture system becomes avail- able for P. vivax.
Similar, albeit more uniform, knobs were first de- scribed by Rudzinska and Trager (1968) for P. coatneyi and have subsequently been found in a number of malar- ial species. In contrast to P. coatney and P. falciparum, in which the knobs serve to mediate sequestration of the late-stage parasite within the postcapillary venules of inner organs, the function of the knobs observed in P. vivax, which is a nonsequestering parasite, is not known. This may be because the knobs in P. vivax are nonfunctioning for sequestration or because there are too few knobs to mediate effective sequestration. In this respect, P. vivax appears to be similar to other nonse- questering parasites such as P. simiovale, which also ex- presses both knobs and caveolae. The structure of the knobs with the underlying electron-dense material is dif- ferent from that reported for P. falciparum; in that their size is much more variable and the electron-dense mate- rial seems to be associated to a greater degree with the erythrocyte membrane. Whereas in P. falciparum the electron-dense material is separated by an electron-lu- cent gap, in P. vivax it is directly associated with and, sometimes, integrated into the membrane. The extent and molecular organization of this association must be elucidated in future research.
One change in the P. vivax-infected erythrocyte cyto- plasm involves the large amount of intraerythrocytic membranes. We did not observe any connection of these membranes to the erythrocytic cell membrane or to the membrane of the parasitophorous vacuole. These mem- branes may be homologous to the Maurer's clefts de- scribed in P. falciparum (Trager et al. 1966). Their func- tion in P. vivax is not known, but it has been shown that Maurer's clefts in P. falciparum are involved in the transport of parasite-derived material to the erythrocyte surface (Aikawa et al. 1986). Using immunogold stain- ing, Barnwell et al. (1990) showed that in P. vivax, some parasite-derived material is located within the erythro- cytic clefts.
All of these changes illustrate the alterations under- gone by a P. vivax-infected red blood cell. One indication of the physiological consequences of these alterations is our observation that during handling of blood infected with P. vivax, the erythrocyte membranes became ex- tremely fragile. Even brief handling and agitation of the blood resulted in breakage of the red blood cells and liberation of the parasite.
The cause of fever in malaria is little understood and
various factors, including the release of malaria pigment and other cell debris from ruptured erythrocytes, have been implicated in this process (Bruce-Chwatt 1985). One enigma in P. vivax malaria is the finding that this disease is connected with highly debilitating febrile par- oxysms, although the accompanying parasitemia is rela- tively low. The present report suggests a hypothesis to account for this phenomenon. With the rupture of a P. vivax-infected erythrocyte, a large amount of foreign (parasite-derived) material is released systemically. This material may then act as exogenous pyrogen, stimulating various types of reactive phagocytic cells (e.g., granulo- cytes, monocytes, macrophages, Kupffer's cells) to sec- rete endogenous pyrogen, which in turn would act on the thermoregulatory areas in the CNS (particularly the hypothalamic areas; Kluger 1979). The severe fever par- oxysms in a P. vivax malaria attack could thus be caused by the large amount of foreign material released. A com- parison of published micrographs of P. falciparum (e.g., Langreth et al. 1978) with those obtained in the present study reveals that P. vivax induces a much greater extent of alteration in its host cell than does P. falciparum. As a consequence, the amount of parasite-derived mate- rial is much higher in a P. vivax infection, with its low parasitemia, than in a P. falciparum infection, despite the higher parasitemia observed in the latter.
The question as to the ultrastructure of P. vivax para- sites in human red blood cells remains largely unans- wered. Judging from the case in P. falciparum, for which data on both monkey and human erythrocytes are avail- able, one would assume that the ultrastructure of P. vivax-infected human red blood cells would mainly be the same as that of infected monkey cells. Published electron micrographs (Trager et al. 1966; Djaldetti et al. 1984; Mackenstedt et al. 1989) confirm this. However, knobs have never been reported in human infections of P. vivax. It remains to be seen whether they are a peculiar expression of the parasite in a nonnatural splen- ectomized host.
Acknowledgements. The skillful help of M. Dodd during all phases of animal work and electron microscopy fixation is greatly appre- ciated and acknowledged, as is S. Nastasi's expert help in the prepa- ration of this manuscript. This work was supported by NIH grants RR 00164 (Division of Research Resources) and AI-26189 (Nation- al Institute of Allergy and Infectious Diseases).
References
Aikawa M (1977) Variations in structure and function during the life cycle of malarial parasites. Bull WHO 55 : 139-156
Aikawa M (1988) Morphological changes in erythrocytes induced by malarial parasites. Biol Cell 64:173-181
Aikawa M, Miller LH, Rabbege J (1975) Caveola-vesicle complexes in the plasmalemma of erythrocytes infected by Plasmodium vivax and P. cynomolgi. Unique structures related to Schiiffner's dots. Am J Pathol 79:285-292
Aikawa M, Hsieh CL, Miller LH (1977) Ultrastructural changes of the erythrocyte membrane in ovale-type malarial parasites. J Parasitol 63 : 152-154
Aikawa M, Uni S, Andruitis AT, Howard RJ (1986) Membrane associated electron dense material of the asexual stages of Plas- modium falciparum: evidence for movement from the intracellu-

H.N. Lanners: Erythrocytes infected by Plasmodium vivax 401
lar parasite to the erythrocyte membrane. Am J Trop Med Hyg 34 : 3~36
Barnwell JW, Ingravallo P, Galinski MR, Matsumoto Y, Aikawa M (1990) Plasmodium vivax: malarial proteins associated with the membrane-bound caveola-vesicle complexes and cytoplas- mid cleft structures of infected erythrocytes. Exp Parasitol 70: 85-99
Boyd MF (1949) Malariology (2 vols). W.B. Saunders, Philadel- phia
Bruce-Chwatt LJ (1985) Essential malariology, 2nd edn. William Heinemann Medical Books, London
Djatdetti M, Mittelman M, Malinsky H, Hart J (1984) Ultrastruc- tural alterations of human red blood cells infected by Plasmo- dium vivax. Acta Haematol 72:121-127
Karnovsky M, Rice DF (1969) Exogenous cytochrome c as an ultrastructural tracer. J Histochem Cytochem 17:751-753
Kawakami H, Hirano H (1986) Rearrangement of the open-canali- cular system of the human blood platelet after incorporation of surface-bound ligands. Cell Tissue Res 245:465-471
Kitchen SF (1938) The infection of reticulocytes by Plasrnodium vivax. Am J Trop Med Hyg 18:342359
Kluger MJ (1979) Fever, its biology, evolution and function. Prin- ceton University Press, Princeton
Langreth SG, Jensen JB, Reese RT, Trager W (i 978) Fine structure of human malaria in vitro. J Protozool 25 : 443-452
Mackenstedt U, Brockelman CR, Mehlhorn H, Raether W (1989) Comparative morphology of human and animal malaria para- sites: I. Host-parasite interface. Parasitol Res 75: 528-535
Matsumoto Y, Aikawa M, Barnwell JW (1988) Immunoelectron microscopic localization of vivax malaria antigens to the clefts and caveola-vesicle complexes of infected erythrocytes. Am J Trop Med Hyg 39:317-322
Rudzinska MA, Trager W (1968) The fine structure of trophozoites and gametocytes in Plasmodium coatneyi. J Protozool 15:73- 88
Sterling CR, Seed TM, Aikawa M, Rabbege J (1975) Erythrocyte membrane alterations induced by Plasmodium simium infection in Saimiri sciureus: relation to Sch/iffner's dots. J Parasitol 61 : 177-254
Trager W, Rudzkinsa MA, Bradbury PC (1966) The fine structure of Plasmodium falciparum and its host erythrocytes in natural malarial infections in man. Bull WHO 35:883-885
Trager W, Lanners HN, Stanley HA, Langreth SG (1983) Immuni- zation of owl monkeys to Plasrnodiurn falciparum with mero- zoites from cultures of a knobless clone. Parasite Immunol 5 : 225-236