
dietary flavanone glycoside naringin modulates the abnormalities ofhuman erythrocytes exposed with...
6
Please cite this article in press as: Magendira Mani V, et al. A dietary flavanone glycoside naringin modulates the abnormali- ties of human erythrocytes exposed with deltamethrin, by upregulating the expression of antioxidants. Biomed Prev Nutr (2013), http://dx.doi.org/10.1016/j.bionut.2013.10.003 ARTICLE IN PRESS G Model BIONUT-191; No. of Pages 6 Biomedicine & Preventive Nutrition xxx (2013) xxx–xxx Available online at ScienceDirect www.sciencedirect.com Original article A dietary flavanone glycoside naringin modulates the abnormalities of human erythrocytes exposed with deltamethrin, by upregulating the expression of antioxidants Vinayagam Magendira Mani a , Adikesavan Gokulakrishnan a , Abdul Majeet Mohammed Sadiq b,∗ a Department of Biochemistry, Islamiah College (Autonomous), Vaniyambadi, 635 752 Tamil Nadu, India b Department of Biochemistry, Adiparasakthi College of Arts and Science, Kalavai, Tamil Nadu, India a r t i c l e i n f o Article history: Received 21 September 2013 Accepted 16 October 2013 Keywords: Deltamethrin Naringin Antioxidant enzymes Oxidative stress Lipid peroxidation Hemolysis a b s t r a c t The protective effect of naringin on deltamethrin poisoning in human erythrocyte was studied using an in vitro model. Hemolysis, percentage met-hemoglobin, lipid peroxidation, glutathione, antioxidant enzymes and erythrocyte ghost protein pattern were assessed to investigate the effect of naringin. Eryth- rocytes at a hematocrit of 10% were incubated with 500 ppm of deltamethrin and/or 0.1 M naringin under physiological conditions of temperature and pH for 2 h. Deltamethrin significantly increased the percent- age of hemolysis and met-hemoglobin in human erythrocytes as compared to the control erythrocytes and naringin significantly (P < 0.05) inhibited the percentage of hemolysis and met-hemoglobin. The levels of lipid peroxides and conjugated diene increased whereas the level of glutathione decreased signifi- cantly (P < 0.05) by deltamethrin-incubated erythrocytes. Naringin significantly inhibited the formation of lipid peroxides and conjugated diene while increased the glutathione level in erythrocytes incubated with deltamethrin. The activity of antioxidant enzymes and non-enzymic antioxidants were decreased in erythrocytes incubated with deltamethrin whereas naringin improved the activities of these antioxidant and non-enzymic antioxidants. SDS–PAGE of erythrocyte ghost protein pattern showed an alteration in the protein bands by deltamethrin poisoning but naringin significantly inhibited the alteration in protein profile. The present study divulges that naringin can reduce the abnormalities of deltamethrin poisoning by ameliorating oxidative stress. This finding raises the possibility that naringin may provide protection from pesticide poisoning. © 2013 Published by Elsevier Masson SAS. 1. Introduction Agricultural pesticide poisoning is a major public health prob- lem in the developing world. Suicide and deliberate self-harm using pesticides is a major but under-recognized public health problem in the developing world. Each year 250,000–370,000 thousand peo- ple die from deliberate ingestion of pesticides [1]. Recently, more than 56 school students died after eating pesticide, poisoned food in Patna and Bhopal, India, in the month of July 2013. Pyrethroids are structural derivatives of naturally occurring pyrethrins, which are present in pyrethrum, an extract from the flowers, Chrysanthemum cinerarifolium. Two distinct classes of pyrethroids have been identified type I and II, based upon ∗ Corresponding author. Department of Biochemistry, Adiparasakthi College of Arts and Science, Kalavai, Tamil Nadu, India. Tel.: +91 4173 242 644; Mobile: +91 9443 449 881. E-mail addresses: [email protected] (V. Magendira Mani), [email protected] (A.M. Mohammed Sadiq). their chemical structure and clinical manifestations of acute exposure [2]. Pyrethroids are a class of synthetic insecticides involved in different neurological disorders affecting humans, especially infants/fetus, the elderly, and those with chronic neu- rological or immune conditions [3]. Studies have documented that chronic exposures can cause chronic health conditions or symptoms similar to such conditions as Parkinson’s, Lou Gehrig’s disease (ALS), Alzheimer’s disease, ADHD/autism, and develop- mental deficits/birth defects/learning disabilities [4]. Pyrethroids cross the blood-brain barrier and exert their effect on dopami- nergic system, contributing to the burden of oxidative stress. Deltamethrin (DLM), [(S)––cyano–3-phenoxybenzyl-(1R, 3R)–3- (2,2-dibromovinyl)-2,2-dimethylcyclo-propane–1-carboxylate], is one of the most neurotoxic pyrethroids. It acts by delaying closure of sodium channels, resulting in a tail current that is characterized by a slow influx of sodium during the end of neuronal depolariza- tion [5]. As the pharmacological action of the pyrethroids including deltamethrin, it has been shown that they modify the gating kinet- ics of axonal Na + channels involved in the inward flow of Na + ions, producing the action potential in cells that are normally closed at 2210-5239/$ – see front matter © 2013 Published by Elsevier Masson SAS. http://dx.doi.org/10.1016/j.bionut.2013.10.003
Transcript of dietary flavanone glycoside naringin modulates the abnormalities ofhuman erythrocytes exposed with...

B
O
Ahe
VAa
b
ARA
KDNAOLH
1
lpipti
pto
AM
m
2h
ARTICLE IN PRESSG ModelIONUT-191; No. of Pages 6
Biomedicine & Preventive Nutrition xxx (2013) xxx–xxx
Available online at
ScienceDirectwww.sciencedirect.com
riginal article
dietary flavanone glycoside naringin modulates the abnormalities ofuman erythrocytes exposed with deltamethrin, by upregulating thexpression of antioxidants
inayagam Magendira Mania, Adikesavan Gokulakrishnana,bdul Majeet Mohammed Sadiqb,∗
Department of Biochemistry, Islamiah College (Autonomous), Vaniyambadi, 635 752 Tamil Nadu, IndiaDepartment of Biochemistry, Adiparasakthi College of Arts and Science, Kalavai, Tamil Nadu, India
a r t i c l e i n f o
rticle history:eceived 21 September 2013ccepted 16 October 2013
eywords:eltamethrinaringinntioxidant enzymesxidative stressipid peroxidationemolysis
a b s t r a c t
The protective effect of naringin on deltamethrin poisoning in human erythrocyte was studied usingan in vitro model. Hemolysis, percentage met-hemoglobin, lipid peroxidation, glutathione, antioxidantenzymes and erythrocyte ghost protein pattern were assessed to investigate the effect of naringin. Eryth-rocytes at a hematocrit of 10% were incubated with 500 ppm of deltamethrin and/or 0.1 M naringin underphysiological conditions of temperature and pH for 2 h. Deltamethrin significantly increased the percent-age of hemolysis and met-hemoglobin in human erythrocytes as compared to the control erythrocytes andnaringin significantly (P < 0.05) inhibited the percentage of hemolysis and met-hemoglobin. The levelsof lipid peroxides and conjugated diene increased whereas the level of glutathione decreased signifi-cantly (P < 0.05) by deltamethrin-incubated erythrocytes. Naringin significantly inhibited the formationof lipid peroxides and conjugated diene while increased the glutathione level in erythrocytes incubatedwith deltamethrin. The activity of antioxidant enzymes and non-enzymic antioxidants were decreased in
erythrocytes incubated with deltamethrin whereas naringin improved the activities of these antioxidantand non-enzymic antioxidants. SDS–PAGE of erythrocyte ghost protein pattern showed an alteration inthe protein bands by deltamethrin poisoning but naringin significantly inhibited the alteration in proteinprofile. The present study divulges that naringin can reduce the abnormalities of deltamethrin poisoningby ameliorating oxidative stress. This finding raises the possibility that naringin may provide protection.
from pesticide poisoning. Introduction
Agricultural pesticide poisoning is a major public health prob-em in the developing world. Suicide and deliberate self-harm usingesticides is a major but under-recognized public health problem
n the developing world. Each year 250,000–370,000 thousand peo-le die from deliberate ingestion of pesticides [1]. Recently, morehan 56 school students died after eating pesticide, poisoned foodn Patna and Bhopal, India, in the month of July 2013.
Pyrethroids are structural derivatives of naturally occurring
Please cite this article in press as: Magendira Mani V, et al. A dities of human erythrocytes exposed with deltamethrin, by upregulahttp://dx.doi.org/10.1016/j.bionut.2013.10.003
yrethrins, which are present in pyrethrum, an extract fromhe flowers, Chrysanthemum cinerarifolium. Two distinct classesf pyrethroids have been identified type I and II, based upon
∗ Corresponding author. Department of Biochemistry, Adiparasakthi College ofrts and Science, Kalavai, Tamil Nadu, India. Tel.: +91 4173 242 644;obile: +91 9443 449 881.
E-mail addresses: [email protected] (V. Magendira Mani),[email protected] (A.M. Mohammed Sadiq).
210-5239/$ – see front matter © 2013 Published by Elsevier Masson SAS.ttp://dx.doi.org/10.1016/j.bionut.2013.10.003
© 2013 Published by Elsevier Masson SAS.
their chemical structure and clinical manifestations of acuteexposure [2]. Pyrethroids are a class of synthetic insecticidesinvolved in different neurological disorders affecting humans,especially infants/fetus, the elderly, and those with chronic neu-rological or immune conditions [3]. Studies have documentedthat chronic exposures can cause chronic health conditions orsymptoms similar to such conditions as Parkinson’s, Lou Gehrig’sdisease (ALS), Alzheimer’s disease, ADHD/autism, and develop-mental deficits/birth defects/learning disabilities [4]. Pyrethroidscross the blood-brain barrier and exert their effect on dopami-nergic system, contributing to the burden of oxidative stress.Deltamethrin (DLM), [(S)–�–cyano–3-phenoxybenzyl-(1R, 3R)–3-(2,2-dibromovinyl)-2,2-dimethylcyclo-propane–1-carboxylate], isone of the most neurotoxic pyrethroids. It acts by delaying closureof sodium channels, resulting in a tail current that is characterizedby a slow influx of sodium during the end of neuronal depolariza-
etary flavanone glycoside naringin modulates the abnormali-ting the expression of antioxidants. Biomed Prev Nutr (2013),
tion [5]. As the pharmacological action of the pyrethroids includingdeltamethrin, it has been shown that they modify the gating kinet-ics of axonal Na+ channels involved in the inward flow of Na+ ions,producing the action potential in cells that are normally closed at

ING ModelB
2 & Pre
tdac
ciTibflna[gruamadeaapmnta
2
2
Sp(dfnp
2
ae[obrwri
etnp
2
b
ARTICLEIONUT-191; No. of Pages 6
V. Magendira Mani et al. / Biomedicine
he resting potential [6]. Most recently, it has been reported thateltamethrin stimulates the glutamate receptor, or blocks the �-minobutyric acid (GABAA) receptor [7], presumably leading tohronic seizures [8].
Flavonoids are a group of polyphenolic compounds diverse inhemical structure and characteristics. They are widely distributedn foods of plant origin, such as vegetables, fruits, tea and wine.he diverse effects of flavonoids may relate to their structural sim-larity to ATP and hence, to their ability to compete with ATP forinding to various enzymatic sites [9]. Citrus fruits contain variousavonoids, and among these naturally occurring citrus flavonoids,aringenin, and naringin have been pharmacologically evaluateds a potential anticancer agent [10] and a hypolipidemic agent11]. Naringin (4′,5,7-trihydroxy flavanone-7-rhamnoglucoside), alycoside, is the predominant flavanone found in grapefruit andelated citrus species e.g., Citrus paradise, Citrus sinensis, Citrusnshiu, and Artemisia selengensis, roots of Cudrania cochinchinensisnd fruits of Pon cirus [12]. Naringin have a broad spectrum of phar-acological and therapeutic properties, including metal chelating,
ntioxidant by upregulating the gene expression of superoxideismutase, catalase and glutathione peroxidase, free radical scav-nging properties [13], antiradical agents, antimicrobial [14] andnti-lipoperoxidation activities, anti-inflammatory, anti-apoptotic,nti-atherogenic, anti-ulcer and metal chelating activity, antihy-erlipidemic [15]. It has been reported to offer protection againstutagenesis, inhibition of tumor development [9,13], Moreover,
aringin demonstrated inhibition of breast cancer cell prolifera-ion and delay of mammary tumorigenesis [16], yet, no mutagenicctivity.
. Materials and methods
.1. Chemicals
All fine chemicals including naringin, were purchased fromigma–Aldrich Chemical Co., USA. Deltamethrin is a syntheticyrethroid insecticide (C22H19Br2NO3). CAS chemical name�–cyano–3–phenoxy benzyl (1R,3R)–3-(2,2-dibromovinyl)-2,2-imethyl cyclopropanecarboxylate). Deltamethrin commercialormulation [Decis, deltamethrin 97.3% EC denotes 97.3% of tech-ical grade deltamethrin (w/w) in emulsifiable concentrate] wasrocured from Bayer crop science limited, Mumbai, India.
.2. Erythrocyte incubation with deltamethrin and naringin
The experiments were conducted according to the ethical normspproved by Ministry of Social Justices and Empowerment, Gov-rnment of India. The erythrocytes were isolated by the method17]. Venous blood was collected from healthy individuals after anvernight fast using EDTA as anticoagulant. After two hours, thelood sample were centrifuged at 4 ◦C for 10 min at 1000 × g toemove plasma and buffy coat and the isolated erythrocytes wereashed 4 to 5 times with 0.154 mol/L NaCl and finally packed eryth-
ocytes were obtained. These packed erythrocytes were used forncubation with 500 ppm of deltamethrin and 0.1 M naringin.
Isolated erythrocytes at a hematocrit of 10% (10 volumes ofrythrocytes to 90 volume of phosphate buffer saline, pH 7.4) werereated for 1 h with a dose of 500 ppm of deltamethrin and/oraringin at 37 ◦C. The erythrocytes incubated in buffer withoutesticide was used as control.
Please cite this article in press as: Magendira Mani V, et al. A dities of human erythrocytes exposed with deltamethrin, by upregulahttp://dx.doi.org/10.1016/j.bionut.2013.10.003
.3. Segregation of experimental groups
Erythrocytes incubated with deltamethrin and naringin as givenelow and the protocol was repeated for six times:
PRESSventive Nutrition xxx (2013) xxx–xxx
• group I: erythrocyte alone• group II: erythrocyte incubated with 500 ppm of deltamethrin• group III: erythrocyte incubated with 500 ppm of deltamethrin
and 0.1 M naringin• group IV: erythrocyte incubated with 0.1 M naringin alone
2.4. Determination of hemolysis and % met-hemoglobin
The ratio of hemolysis was calculated from the equation asdescribed by [18]. H (%) = (Apb – A0)/(Awater – A0) × 100% where H(%) is the hemolysis of erythrocytes incubated with deltamethrin,Apb is a absorbance of sample, Awater is the absorbance after com-plete hemolysis and A0 is the absorbance of sample with 155 mMof NaCl. The concentration of hemoglobin was measured by mea-suring the absorption spectra of hemoglobin in the wavelengthrange of 440–700 nm using Schimadzu UV–visible spectropho-tometer. The percentage of met-Hb in the total Hb content wascalculated from absorbance at 630 and 700 nm both for hemoglobinof control erythrocytes and hemoglobin of erythrocytes treatedwith deltamethrin and naringin as described by [18]. Hemoglobintreated with potassium ferricyanide (100% met) was used as a pos-itive control.
2.5. Assay of MDA and membrane bound enzymes
Erythrocyte MDA levels were measured using a thiobarbi-turicacid (TBA) method [19]. After the reaction with TBA, thereaction product was measured spectrophotometrically at 535 mm.The MDA level was expressed as nmol/g Hb. Erythrocyte mem-brane antioxidant enzymes, such as superoxide dismutase (SOD)[20], catalase (CAT) [21], glutathione peroxidase (GPx) [22], glu-tathione reductase (GR) [23], and non-enzymic antioxidants, suchas reduced glutathione (GSH) [24], vitamin C [25], vitamin E [26]were assayed. The activity of Na+/K +-ATPase was determinedby the method of [27]. Ca2+-ATPase was measured accordingto [28]. Mg2+-ATPase was assayed according to the method of[29]. Inorganic phosphorus was estimated by the method of[30].
2.6. Determination of protein pattern in erythrocyte membrane
Erythrocyte membranes were isolated by the method of[31]. After the incubation period, an aliquot of erythrocyteswere subjected to hemolysis by adding hypotonic Tris–buffer,0.015 M, pH 7.2. After 4–6 h, the erythrocyte ghosts were sedi-ment by high-speed centrifugation at 12,000 rpm, for 40–45 min,at 4–6 ◦C. The erythrocyte membrane pellets were suspendedin 0.02 M Tris–buffer, pH 7.2 and subjected to Sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE) accord-ing to the method of [32]. Gels were stained with silver stain asdescribed by [33].
2.7. Statistical analysis
All data were analyzed with SPSS/12 student software.Hypothesis testing methods included one way analysis of vari-ance (ANOVA) followed by LSD. The values are expressedas mean ± SD and the results were considered significantlydifferent if P < 0.05. Statistically significant variations are com-
etary flavanone glycoside naringin modulates the abnormali-ting the expression of antioxidants. Biomed Prev Nutr (2013),
pared as follows: erythrocyte + DEL vs normal erythrocyte,erythrocyte + DEL vs erythrocyte + DEL + naringin and erythro-cyte + naringin vs normal erythrocyte.* indicates P < 0.05 and NS
indicates non-significant.

ARTICLE IN PRESSG ModelBIONUT-191; No. of Pages 6
V. Magendira Mani et al. / Biomedicine & Preventive Nutrition xxx (2013) xxx–xxx 3
*
*NS
*
*
NS
-1
0
1
2
3
4
5
6
7
Erythrocyte + DEL+Erythrocyte + DELErythrocyteNaringin
Erythrocyte + Naringin
% o
f Hem
olys
is &
met
-Hb
Hemolysis % met-Hb
Fig. 1. Effect of deltamethrin and naringin on the percentage of hemolysis and % met-Hb in control and experimental group of erythrocytes. Results are expressed asm fferena yte + Da
3
3
aTcdmei
3
irnagog
Fmaa
ean ± SD for 6 different sets of experiments. Values are considered significantly dis follows: erythrocyte + DEL vs normal erythrocyte, erythrocyte + DEL vs erythrocnd NS indicates non-significant.
. Results
.1. Effect of DEL and naringin on hemolysis and met-hemoglobin
The percentage of hemolysis and met-hemoglobin of the controlnd experimental groups of erythrocytes were presented in Fig. 1.he percentage of hemolysis significantly increased in erythro-ytes incubated with DEL as compared to control erythrocytes. DELecreased oxyhemoglobin concentration and increased the level ofet-Hb in DEL-treated erythrocytes as compared to control. How-
ver, naringin prevent the hemolysis and met-Hb in deltamethrinncubated erythrocytes (group III).
.2. Effect of DEL and Nringin on lipid peroxidation
In this present study, we observed that the significantlyncreased level of MDA were shown in deltamethrin-treated eryth-ocytes when compared to controls (Fig. 2). On treatment witharingin, the MDA levels decreased significantly in deltamethrin
Please cite this article in press as: Magendira Mani V, et al. A dities of human erythrocytes exposed with deltamethrin, by upregulahttp://dx.doi.org/10.1016/j.bionut.2013.10.003
nd naringin-treated group compared with deltamethrin-treatedroup. It can be said that naringin decreased the deleterious effectsf deltamethrin in lipid peroxidation. Also, naringin alone treatedroup does not cause any significant alteration.
ig. 2. Effect of deltamethrin and naringin on the levels of lipid peroxides, conjugated dean ± SD for 6 different sets of experiments. Values are considered significantly differen
s follows: erythrocyte + DEL vs normal erythrocyte, erythrocyte + DEL vs erythrocyte + Dnd NS indicates non-significant.
t at P < 0.05 with post-hoc LSD test. Statistically significant variations are comparedEL + naringin and erythrocyte + naringin vs normal erythrocyte.* indicates P < 0.05
3.3. Effect of DEL and naringin on antioxidant enzymes
Table 1 shows the levels of antioxidant enzymes of the controland experimental groups of erythrocytes. The antioxidant enzymesSOD, CAT, GR and GPx activities significantly (P < 0.05) decreased inall experimental groups as compared to the control erythrocytes.However, on treatment with naringin, the antioxidant enzymeactivity significantly (P < 0.05) increased in group III and IV as com-pared to group II.
3.4. Effect of DEL and naringin on non-enzymic antioxidants
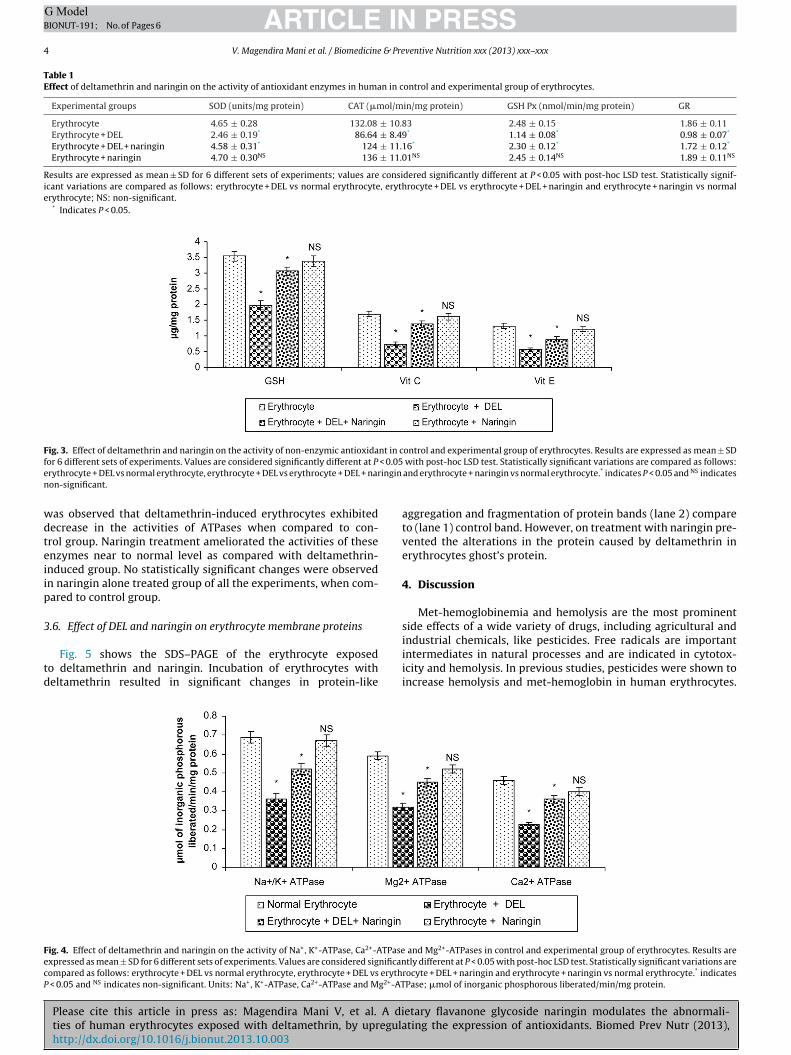
Fig. 3 shows that GSH, vitamin C and vitamin E levels were signif-icantly decreased (P ≤ 0.05) in the DLM-treated groups compared tothe control group. The greatest reduction was occurred in the DLM-treated group. Moreover, there was a significant increase (P ≤ 0.05)in non-enzymic antioxidants found in the naringin-treated groupscompared to the DEL induced group.
etary flavanone glycoside naringin modulates the abnormali-ting the expression of antioxidants. Biomed Prev Nutr (2013),
3.5. Effect of DEL and naringin on ATPases
Fig. 4 shows the activities of ATPases (Na+/K+-ATPase, Mg2+-ATPase and Ca2+-ATPase) in control and experimental groups. It
ienes in control and experimental group of erythrocytes. Results are expressed ast at P < 0.05 with post-hoc LSD test. Statistically significant variations are comparedEL + naringin and erythrocyte + naringin vs normal erythrocyte.* indicates P < 0.05
ARTICLE IN PRESSG ModelBIONUT-191; No. of Pages 6
4 V. Magendira Mani et al. / Biomedicine & Preventive Nutrition xxx (2013) xxx–xxx
Table 1Effect of deltamethrin and naringin on the activity of antioxidant enzymes in human in control and experimental group of erythrocytes.
Experimental groups SOD (units/mg protein) CAT (�mol/min/mg protein) GSH Px (nmol/min/mg protein) GR
Erythrocyte 4.65 ± 0.28 132.08 ± 10.83 2.48 ± 0.15 1.86 ± 0.11Erythrocyte + DEL 2.46 ± 0.19* 86.64 ± 8.49* 1.14 ± 0.08* 0.98 ± 0.07*
Erythrocyte + DEL + naringin 4.58 ± 0.31* 124 ± 11.16* 2.30 ± 0.12* 1.72 ± 0.12*
Erythrocyte + naringin 4.70 ± 0.30NS 136 ± 11.01NS 2.45 ± 0.14NS 1.89 ± 0.11NS
Results are expressed as mean ± SD for 6 different sets of experiments; values are considered significantly different at P < 0.05 with post-hoc LSD test. Statistically signif-icant variations are compared as follows: erythrocyte + DEL vs normal erythrocyte, erythrocyte + DEL vs erythrocyte + DEL + naringin and erythrocyte + naringin vs normalerythrocyte; NS: non-significant.
* Indicates P < 0.05.
Fig. 3. Effect of deltamethrin and naringin on the activity of non-enzymic antioxidant in control and experimental group of erythrocytes. Results are expressed as mean ± SDf < 0.05e inginn
wdteiip
3
td
FecP
or 6 different sets of experiments. Values are considered significantly different at Prythrocyte + DEL vs normal erythrocyte, erythrocyte + DEL vs erythrocyte + DEL + naron-significant.
as observed that deltamethrin-induced erythrocytes exhibitedecrease in the activities of ATPases when compared to con-rol group. Naringin treatment ameliorated the activities of thesenzymes near to normal level as compared with deltamethrin-nduced group. No statistically significant changes were observedn naringin alone treated group of all the experiments, when com-ared to control group.
.6. Effect of DEL and naringin on erythrocyte membrane proteins
Please cite this article in press as: Magendira Mani V, et al. A dities of human erythrocytes exposed with deltamethrin, by upregulahttp://dx.doi.org/10.1016/j.bionut.2013.10.003
Fig. 5 shows the SDS–PAGE of the erythrocyte exposedo deltamethrin and naringin. Incubation of erythrocytes witheltamethrin resulted in significant changes in protein-like
ig. 4. Effect of deltamethrin and naringin on the activity of Na+, K+-ATPase, Ca2+-ATPasexpressed as mean ± SD for 6 different sets of experiments. Values are considered significanompared as follows: erythrocyte + DEL vs normal erythrocyte, erythrocyte + DEL vs eryth
< 0.05 and NS indicates non-significant. Units: Na+, K+-ATPase, Ca2+-ATPase and Mg2+-AT
with post-hoc LSD test. Statistically significant variations are compared as follows: and erythrocyte + naringin vs normal erythrocyte.* indicates P < 0.05 and NS indicates
aggregation and fragmentation of protein bands (lane 2) compareto (lane 1) control band. However, on treatment with naringin pre-vented the alterations in the protein caused by deltamethrin inerythrocytes ghost’s protein.
4. Discussion
Met-hemoglobinemia and hemolysis are the most prominentside effects of a wide variety of drugs, including agricultural and
etary flavanone glycoside naringin modulates the abnormali-ting the expression of antioxidants. Biomed Prev Nutr (2013),
industrial chemicals, like pesticides. Free radicals are importantintermediates in natural processes and are indicated in cytotox-icity and hemolysis. In previous studies, pesticides were shown toincrease hemolysis and met-hemoglobin in human erythrocytes.
and Mg2+-ATPases in control and experimental group of erythrocytes. Results aretly different at P < 0.05 with post-hoc LSD test. Statistically significant variations are
rocyte + DEL + naringin and erythrocyte + naringin vs normal erythrocyte.* indicatesPase; �mol of inorganic phosphorous liberated/min/mg protein.

ARTICLE ING ModelBIONUT-191; No. of Pages 6
V. Magendira Mani et al. / Biomedicine & Pre
Fig. 5. Effect of deltamethrin and naringin on the SDS–PAGE protein patternof the erythrocytes of control and experimental groups. Protein fragmentationanalysis by SDS–PAGE on control and experimental group of human erythro-c3
Pttsdqlmdiht
ahoodsoip
wiocoi[sarGt
aMrdb
[tation with blueberries, spinach, or spirulina reduces ischemic brain damage.
ytes. Lane 1–control erythrocytes; lane 2–deltamethrin-induced erythrocytes; lane–deltamethrin + naringin-treated; lane 4–naringin alone treated.
esticide like 2,4-dichlorophenoxyacetic acid have been reportedo decrease oxygen affinity of haemoprotein and at the sameime increases the level of methaemoglobin [18]. In this presenttudy, deltamethrin cause increase in hemolysis in erythrocytesue to deltamethrin react with oxyhemoglobin with the conse-uent reduction of molecular oxygen to active oxygen species
eading to hemotoxic damage [34]. The dose- and time-dependentet-hemoglobin and hemolysis responses suggest that these
eltamethrin as potential active pesticide may mediate hemotox-city. However, naringin prevent the factors responsible for theemolysis and reduced the hemolysis and methhemoglobin in DEL-reated erythrocytes.
Lipid peroxidation associated with reduction of antioxidants is characteristic observation in the erythrocytes and malondialde-yde is used as an indicator of tissue damage [35]. LPO is a markerf oxidative damage, which plays an important role in the toxicityf many xenobiotics, including pesticides. They found significantecrease in MDA concentration in naringin-treated erythrocyteshortly after treating with deltamethrin. The inhibition of lipid per-xidation by naringin is mainly due to free radical scavenging andts antioxidant activity. Naringin has been reported to inhibit lipideroxidation in iron-induced, brain, kidney and liver damage [36].
The formation of free radicals is prevented by antioxidants,hich block some of destructive reactions in the cell. Reduced activ-
ties of antioxidant enzymes (SOD, CAT, GR and GPx) after treatmentf deltamethrin are important indicators for the toxicity of thesehemicals [37]. The lowered enzyme levels may reflect to increasedxidative damage [38]. The natural antioxidants may be helpfuln preventing or reducing the harmful effects of ROS on pesticide39]. Naringin has metal chelating, antioxidant, and free radicalcavenging properties [16]. It has been reported to offer protectiongainst mutagenesis and lipid peroxidation [13]. Naringin has beeneported to inhibit radiation-induced depletion in the SOD, CAT, GR,SHpx, GSH, vitamin C and E [13]. It has also been reported to inhibit
he streptozotocin-induced depletion in SOD CAT and GSHpx [40].The effects of deltamethrin were evaluated at cellular level,
nd primary effects were determined to be on sodium channels.onoamine oxidase and Na–K ATPase activities are reported to be
Please cite this article in press as: Magendira Mani V, et al. A dities of human erythrocytes exposed with deltamethrin, by upregulahttp://dx.doi.org/10.1016/j.bionut.2013.10.003
educed in deltamethrin intoxication [41]. The neurotoxic effect ofeltamethrin is related to its binding to the sodium channels of therain and interaction with the GABA receptor ionophore complex
[
PRESSventive Nutrition xxx (2013) xxx–xxx 5
[42]. Pyrethroids owe their insecticidal activity to their ability todisrupt electrical signalling in the nervous system by prolongingthe opening of voltage-gated sodium channels and deltamethrinshown significant heterogeneity in the sodium channel openingsthat produce the nature of the poisoning syndromes in insects,amphibia, and mammals [43]. Voltage-gated sodium channels arealso considered to be the primary targets for the central neuro-toxic effects of pyrethroids in mammals [44]. The decrease in theactivities of these ATPases could be due to the enhanced lipid perox-idation by free radicals in deltamethrin-induced erythrocytes andthe effective protection against decrease in ATPases activity is evi-dent among naringin-treated erythrocytes due to the decline ingeneration of free radical and lipid peroxidation.
Previous reports were shown that the aggregation in the proteinbands was documented in erythrocyte ghosts incubated with pes-ticide [45]. In this study, protein erythrocytes were incubated withdeltamethrin that cause fragmentation and aggregation of proteinbands, this is the evidence of deltamethrin powerful toxicants. Theprevention of alteration in protein bands by naringin indicates theprotective role offered by naringin on deltamethrin-induced eryth-rocytes.
5. Conclusion
In conclusion, in this present work, it has shown thatdeltamethrin cause dramatic changes in biochemical, enzymaticand structural changes in erythrocytes, the present data confirmedthat naringin exerts protective action on deltamethrin-inducederythrocytes by modulating the abnormalities by upregulating theexpression of antioxidants. Naringin can therefore potentially beused as a pharmaceutical protective agent. Further investigationsare in progress to demonstrate the mechanism of deltamethrin inneuronal damage and the neuroprotective mechanism of naringin.
Disclosure of interest
The authors declare that they have no conflicts of interest con-cerning this article.
References
[1] Gunnell D, Fernando R, Hewagama M, Priyangika WD, Konradsen F. Theimpact of pesticide regulations on suicide in Sri Lanka. Int J Epidemiol2007;36:1235–42.
[2] Dawson A, Buckley N. Integrating approaches to paraquat poisoning. CeylonMed 2007;52:45–7.
[3] Parrón T, Requena M, Hernández AF, Alarcón R. Association between environ-mental exposure to pesticides and neurodegenerative diseases. Toxicol ApplPharmacol 2011;256:379–85.
[4] Kanavouras K, Tzatzarakis MN, Mastorodemos V, Plaitakis A, Tsatsakis AM.A case report of motor neuron disease in a patient showing significant levelof DDTs, HCHs and organophosphate metabolites in hair as well as levels ofhexane and toluene in blood. Toxicol Appl Pharmacol 2011;256:399–404.
[5] Tuzmen N, Candan N, Kaya E, Demiryas N. Biochemical effects of chlorpyri-fos and deltamethrin on altered antioxidative defense mechanisms and lipidperoxidation in rat liver. Cell Biochem Funct 2011;26:119–24.
[6] Klaassen CD. Ca2+-permeable AMPA receptors and selective vulnerability ofmotor neurons. Basic Sci Poisons 2001:784–7.
[7] Narahashi T. Neuroreceptors and ion channels as the basis for drug action: past,present, and future. J Pharmacol Exp Therap 2000;294:1–26.
[8] Hossain MM, Suzuki T, Unno T, Komori S, Kobayashi H. Differential presynapticactions of pyrethroid insecticides on glutamatergic and GABAergic neurons inthe hippocampus. Toxicology 2008;243:155–63.
[9] Tripoli E, Guardia M, Giammanco S, Majo D, Giammanco M. Citrus flavonoids:molecular structure, biological activity and nutritional properties: a review.Food Chem 2007;104:466–79.
10] Wang Y, Chang CF, Chou J, Chen HL, Deng X, Harvey BK, et al. Dietary supplemen-
etary flavanone glycoside naringin modulates the abnormali-ting the expression of antioxidants. Biomed Prev Nutr (2013),
Exp Neurol 2005;193:75–84.11] Bok SH, Shin YW, Bae KH, Jeong TS, Kwon YK, Park YB, et al. Effects of naringin
and lovastatin on plasma and hepatic lipids in high-fat and high cholesterol fedrats. Nutr Res 2000;20:1007–15.

ING ModelB
6 & Pre
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLEIONUT-191; No. of Pages 6
V. Magendira Mani et al. / Biomedicine
12] Rajadurai M, Stanely MP. Preventive effect of naringin on lipid peroxides andantioxidants in isoproterenol-induced cardio toxicity in Wistar rats: biochem-ical and histopathological evidences. Toxicology 2006;228:259–68.
13] Jagetia GC, Reddy TK. Modulation of radiation-induced alteration in the antiox-idant status of mice by naringin. Life Sci 2005;77:780–94.
14] Kim DH, Jung EA, Song IS, Han JA, Kim TH, Han MJ. Intestinal bacterialmetabolism of flavonoids and its relation to some biological activities. ArchPharmacol Res 1998;21:17–23.
15] Ribeiro IA, Rocha J, Sepodes B, Mota-Filipe H, Ribeiro MH. Effect of naringinenzymatic hydrolysis towards naringenin on the anti-inflammatory activity ofboth compounds. J Mol Catal B: Enzym 2008;52:13–8.
16] Nijveldt RJ, van Nood E, van Hoorn DEC, Boelens PG, van Norren K, van LeeuwenPAM, et al. a review of probable mechanisms of action and potential applica-tions. Clin Nutr 2001;74:418–25.
17] Rizvi SI, Zaid MA. Impairment of sodium pump and Na/H exchanger in eryth-rocytes from non-insulin dependent diabetes mellitus patients: effect of teacatechins. Clin Chem Acta 2005;354:59–67.
18] Pieniazek D, Bukowska B, Duda W. Comparison of the effect of Roundup Ultra360 SL pesticide and its active component glyphosate on human erythrocytes.Pest Biochem Physiol 2004;79:58–63.
19] Bartosz G. Prooxidative effects of TEMPO on human erythrocytes. PWNWarszawa 2004:358–9.
20] Misra HP, Fridovich I. The role of superoxide anion in the auto oxidationof epinephrine and a simple assay for superoxide dismutase. J Biol Chem1972;247:3170–5.
21] Takahara S, Hamilton BH, Neell JV, Kobra TY, Ogura Y, Nishimura ET, et al. Anew genetic carrier state. J Clin Invest 1960;29:10–619.
22] Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, Hoekstra WG,et al. Biochemical role as a component of glutathione peroxidase. Science1973;79:588–90.
23] Staal G, Visser E, Veeger JC. Purification and properties of glutathione reductaseof human erythrocytes. Biochim Biophys Acta 1969;185:39–48.
24] Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys 1959;82:70–7.25] Omaye ST, Tumball JD, Sauberlich HE. Selected methods for the determina-
tion of ascorbic acid in animal cells, tissues and fluids. Meth Enzymol 1979;62:1–11.
26] Desai I. Vitamin E analysis methods for animal tissues. Meth Enzymol1984;105:138–47.
Please cite this article in press as: Magendira Mani V, et al. A dities of human erythrocytes exposed with deltamethrin, by upregulahttp://dx.doi.org/10.1016/j.bionut.2013.10.003
27] Bonting SL. Sodium-potassium activated adenosine triphosphatase cationtransport. In: Bitter EE, editor. Membrane Ion Transport, 1. Inter Science Chich-ester; 1970. p. 257–63.
28] Hjerten S, Pan H. Purification and characterisation of low affinity Ca2+ ATPaseform erythrocyte membranes. Biochim Biophys Acta 1983;728:281–8.
[
PRESSventive Nutrition xxx (2013) xxx–xxx
29] Ohnishi T, Suzuki T, Suzuki Y, Ozawa K. A comparative study of plasma mem-brane Mg2+-ATPase activities in normal, regenerating and malignant cells.Biochem Biophys Acta 1982;684:67–74.
30] Fiske CH, Subbarow Y. Colorimetric determination of phosphorus. J Biol Chem1925;66:375–400.
31] Rajasekaran NS, Devaraj NS, Devaraj H. Modulation of rat erythrocyte antioxi-dant defense system by buthionine sulfoximine and its reversal by glutathionemonoester therapy. Biochem Biophys Acta 2004;1688:121–9.
32] Laemmli UK. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature 1970;227:680–5.
33] Tsai M, Frasch C. Sensitive silver staining for detecting lipopolysaccharides inpolyacrylamide gels. Anal Biochem 1982;119:115–9.
34] Prchal J, Gregg X. Red cell enzymopathies. Hematology: basic principles andpractice. 3rd ed. New York: Churchill Livingstone; 2000. p. 561–76.
35] Ohkawa H, Ohishi N, Tagi K. Assay for lipid peroxides in animal tissues bythiobarbituric acid reaction. Anal Chem 1979;95:351–8.
36] Wang TB, Ng F, Liu ZT. Antioxidative activity of natural products from plants.Life Sci 2001;66:709–23.
37] Jayakumar T, Sakthivel M, Thomas PA, Geraldine P. Pleurotus ostreatus, an oystermushroom, decreases the oxidative stress induced by carbon tetrachloride inrat kidneys, heart and brain. Chem Biol Interact 2008;176:108–20.
38] Srinivasan M, Rukkumani R, Ram Sudheer A, Menon VP. Ferulic acid, a natu-ral protector against carbon tetrachloride-induced toxicity. Fundamental ClinPharmacol 2005;19:491–6.
39] Yousef MI, Vitamin E. modulates reproductive toxicity of pyrethroid lambda-cyhalothrin in male rabbits. Food Chem Toxicol 2010;48:1152–9.
40] Ali MM, Abd El Kader MA. The influence of naringin on the oxidative stateof rats with streptozotocin-induced acute hyperglycaemia. Z Naturforsch2004;59:726–33.
41] Calore EE, Cavaliere MJ, Puga FR, Calore NMP, Rosa AR, Weg R, et al. Histologicperipheral nerve changes in rats induced by deltamethrin. Ecotoxicol EnvironSaf 2000;47:82–6.
42] Lindblade KA, Dotson E, Hawley WA, Bayoh N, Williamson J, Mount D, et al.Evaluation of long-lasting insecticidal nets after 2 years of household use. TropMed Int Health 2005;10:1141–50.
43] Ray DE, Fry JR. A reassessment of the neurotoxicity of pyrethroid insecticides.Pharmacol Ther 2006;111:174–93.
44] Soderlund DM, Clark JM, Sheet LP, Mullin LS, Piccirillo VJ, Sargent D, et al.
etary flavanone glycoside naringin modulates the abnormali-ting the expression of antioxidants. Biomed Prev Nutr (2013),
Mechanisms of pyrethroid neurotoxicity: implications for cumulative riskassessment. Toxicology 2002;171(1):3–59.
45] Prasanthi K, Muralidhara, Rajini PS. Morphological and biochemical pertur-bations in rat erythrocytes following in vitro exposure to Fenvalerate and itsmetabolite. Toxicol In Vitro 2005;19(4):449–56.
















