Neurotoxicological effects of deltamethrin on the postnatal development of cerebellum of rat
14
J. Biosci., Vol. 22, Number 2, March 1997, pp 117–130. © Printed in India. Neurotoxicological effects of deltamethrin on the postnatal development of cerebellum of rat Ν PATRO*, S Κ MISHRA, Μ CHATTOPADHYAY and I K PATRO Neurobiology and Ageing Unit, School of Studies in Zoology, Jiwaji University, Gwalior 474011, India MS received 31 August 1996; revised 3 December 1996 Abstract. Deitamethrin (DLT) has been accepted to be 10,000 times less toxic to man than to insects. While toxicity of DLT in adult animals has been studied using biochemical and electrophysiological tools, reports on its developmental neurotoxicity are rather scanty. Wistar rat pups were exposed to DLT (0·7 mg/kg body wt/day, i.p., dissolved in propylene glycol) from postnatal day 9-13. Equal number of age matched pups were used as vehicle controls. The animals were weighed and perfused intracardially on postnatal days 12, 15, 21, and 30 and their brains dissected out. Cerebellum along with the brainstem was separated by a transverse section at the tectal level and processed for morphometric and toxicological studies. The micro- and inter-neurons in the cerebellum are known to differentiate and mature, both morphologically and biochemically, during the postnatal life of rats. Postnatal exposure to DLT has been observed to delay the cytogenesis and morphogenesis of these neurons. In addition to this, damage to the developing vasculature has also been recorded in the form of thrombus and haemorrhage. Focal degeneration and spongy appearance of the tissue in the vicinity of the damaged blood vessels have also been recorded. The study has opened up several questions on the safety of this substance to the pregnant mothers and infants in the habitats where this substance is in use for vector control. Keywords. Deitamethrin; cerebellum; developmental neurotoxicity 1. Introduction Pyrethroids are very active synthetic insecticides which act on the nervous system (Narahashi 1980; Hijzen and Slangen 1988; Aldridge 1990). The primary target site for pyrethroid action, in both insects and mammals, is the axonal sodium channel through a dissociable interaction with macromolecular components (Lund and Narahashi 1983; Aldridge 1990; Vijverberg and van den Bercken 1990; Narahashi 1992). Deitamethrin (DLT) is considered to be amongst the safest class of available insecti- cides with relatively low mammalian toxicity due to the presence of ester group in the molecule (Aldridge 1990; Vijverberg and van den Bercken 1990). Extensive literature available on the toxicokinetics of this insecticide reveals that, in mammals, the primary detoxification step is ester cleavage followed by hydroxylation (Casida et al 1983). The rate of detoxification is very high in mammals than in insects leading to high insect/mammalian toxicity ratio (Elliot 1976; Aldridge 1990). The main mechanism of action of DLT involves the inhibition of acetylcholine esterase (Coppage and Braidech 1976; Reddy et al 1991) resulting in a cholinergic syndrome (Moretto and Johnson 1987). Acute exposure to DLT results in decreased * Corresponding author 117
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Neurotoxicological effects of deltamethrin on the postnatal development of cerebellum of rat

J. Biosci., Vol. 22, Number 2, March 1997, pp 117–130. © Printed in India. Neurotoxicological effects of deltamethrin on the postnatal development of cerebellum of rat
Ν PATRO*, S Κ MISHRA, Μ CHATTOPADHYAY and I K PATRO Neurobiology and Ageing Unit, School of Studies in Zoology, Jiwaji University, Gwalior 474011, India MS received 31 August 1996; revised 3 December 1996 Abstract. Deitamethrin (DLT) has been accepted to be 10,000 times less toxic to man than to insects. While toxicity of DLT in adult animals has been studied using biochemical and electrophysiological tools, reports on its developmental neurotoxicity are rather scanty. Wistar rat pups were exposed to DLT (0·7 mg/kg body wt/day, i.p., dissolved in propylene glycol) from postnatal day 9-13. Equal number of age matched pups were used as vehicle controls. The animals were weighed and perfused intracardially on postnatal days 12, 15, 21, and 30 and their brains dissected out. Cerebellum along with the brainstem was separated by a transverse section at the tectal level and processed for morphometric and toxicological studies. The micro- and inter-neurons in the cerebellum are known to differentiate and mature, both morphologically and biochemically, during the postnatal life of rats. Postnatal exposure to DLT has been observed to delay the cytogenesis and morphogenesis of these neurons. In addition to this, damage to the developing vasculature has also been recorded in the form of thrombus and haemorrhage. Focal degeneration and spongy appearance of the tissue in the vicinity of the damaged blood vessels have also been recorded. The study has opened up several questions on the safety of this substance to the pregnant mothers and infants in the habitats where this substance is in use for vector control. Keywords. Deitamethrin; cerebellum; developmental neurotoxicity
1. Introduction Pyrethroids are very active synthetic insecticides which act on the nervous system (Narahashi 1980; Hijzen and Slangen 1988; Aldridge 1990). The primary target site for pyrethroid action, in both insects and mammals, is the axonal sodium channel through a dissociable interaction with macromolecular components (Lund and Narahashi 1983; Aldridge 1990; Vijverberg and van den Bercken 1990; Narahashi 1992). Deitamethrin (DLT) is considered to be amongst the safest class of available insecti-cides with relatively low mammalian toxicity due to the presence of ester group in the molecule (Aldridge 1990; Vijverberg and van den Bercken 1990). Extensive literature available on the toxicokinetics of this insecticide reveals that, in mammals, the primary detoxification step is ester cleavage followed by hydroxylation (Casida et al 1983). The rate of detoxification is very high in mammals than in insects leading to high insect/mammalian toxicity ratio (Elliot 1976; Aldridge 1990).
The main mechanism of action of DLT involves the inhibition of acetylcholine esterase (Coppage and Braidech 1976; Reddy et al 1991) resulting in a cholinergic syndrome (Moretto and Johnson 1987). Acute exposure to DLT results in decreased * Corresponding author 117

118 N Patro et al motor activity (Chanh et al 1984; Crofton and Reiter 1984; Gilbert et al 1990), decreased operant response rate (Glomot 1982; Peele and Crofton 1987; Bloom et al 1993) and impaired performance on an inclined plane (Rose and Deewar 1983). The Pyrethroids also have the potential of causing axonal damage in the peripheral nerves of rodents (Aldridge 1990). Most severe cases of pyrethroid poisoning reported in infants may be due to the insufficient hydrolysis of the pyrethrum esters (Hayes 1982).
Because of the lipophilic nature of these Pyrethroids, they have an easy as well as rapid access to the tissues, including the central nervous system (CNS), and thus even the very small doses are able to produce significant biological effects. The intravenous dosage required for intrinsic toxicity is very low. The in vivo biological effects of several Pyrethroids, including effects on CNS, are produced by a very small dose, e.g., an intraperitoneal administration of 5–8 mg DLT/kg body wt. has been recorded to be quite effective. Such an administration of DLT results in concentration of parental compound to the tune of 2·4-3·4, 0.12-0.45 and 0.23-0.38 nmol/g of blood, brain and spinal cord, respectively (Aldridge 1990).
Increasing use of pyrethroid insecticides in agricultural and public health, disinsec-tion of aircrafts and personal protection by using mosquito nets impregnated with DLT or permethrin and in various other ways have enhanced the chances of human exposure to this class of insecticides (Depeng et al 1994; Janakara et al 1995; MoyouSomo et al 1995). Developing mammalian brain is highly susceptible to environmental changes and toxicants, as compared to the adult brain, with permanent and devastating effects. Developmental neurotoxicology demonstrates and analyses the frank pathological changes caused by commonly used drugs, pesticides, occupa-tional hazards and other environmental agents otherwise claimed to be comparatively safe to adult and mature brain.
During normative cerebellar development of rat the basket cells are produced mostly on postnatal days 6 and 7, the stellate cells on postnatal days 8-11, and the granule cells beginning shortly after birth until day 21 when the external granular layer disappears (Altman 1969, 1972; Altman and Anderson 1972, 1973). Some of the experimental conditions like experimental undernutrition (Choudhary et al 1982; Gopinath 1984), X-ray irradiation (Altman and Anderson 1972, 1973), internal irradiation (Gopinath et al 1987), transient injury of the external granular layer (Woodward et al 1975), exposure to heavy metals like arsenic (Harding et al 1968) and lead (Pounds 1984) and chloroquine (Agarwal 1991), during this critical period of brain development have been recorded to variously affect the histogenesis and morphogen-esis of cerebellum.
The main aim of the present study was to investigate the effects of DLT on the morphological events occurring in the cerebellum of the rat brain. It has been observed that about 97% of the total cell number in the cerebellum of the rat are formed after birth (Balazas and Patel 1973). Although glial cell formation predominates postnatal development of brain in total, neurogenesis is also significant in cerebellum where most of the interneurons are formed during the period (Altman 1969). Considering the fact that the extent of damage depends on the developmental stage at which the infants are exposed, the rats of postnatal age 9-13 days, which is equivalent to the first year of the human postnatal life, were selected. During this period some of the very important events of cerebellar development occur e.g., granule cell proliferation and migration, formation of the cells of the molecular layer, Purkinje cell arborization, spine formation and synaptic formation (Epstein and Stanford 1977). DLT was selected amongst the

Developmental neurotoxicity of deltamethrin 119 Pyrethroids as its kinetics and pharmacologic, physiologic and behaviour effects have been studied extensively in adult animals (Ray 1980; Cremer and Seville 1982; Rickard and Brodie 1985; Crofton and Reiter 1984, 1987) and the developmental neurotoxicity remains to be evaluated. 2. Materials and methods Cerebellar growth in rats is active during the first two weeks of postnatal life (Altaian 1972; Gopinath et al 1987). We followed the views of Paxinos et al (1994) and considered the first 24 h following birth as P0. Wistar rats of postnatal day 9, born to healthy mothers were chosen from the breeding colony of the animal house of our Department. Average litter size for the study was chosen to be eight. Half of the pups of each litter were used as controls (group-I) and the other half was injected with DLT (group-II) from postnatal day 9 to 13 (P9-P13) as per the experimental plan given in table 1. The animals were weighed and dissected on postnatal days 12, 15, 21 and 30 (P12, P15, P21and P30).
On the respective days, the animals were anaesthetized with ether vapour and perfused intracardially with phosphate buffered saline (pH 7·2) followed by 10% buffered formalin (pH 7·2). Every care was taken to perfuse the control and the treated pups similarly to avoid any perfusion associated artefact. The brains were carefully dissected out and weighed. Cerebellum along with the brainstem were separated by a transverse section at the tectal level and post-fixed in buffered formalin (pH 7·2) for 48 h. The tissues were processed for paraffin embedding and serial sagittal sections were cut at 10 µm thickness by using a Weswox rotary microtome.
The sections were stained with hematoxylin and eosin (H and E) and cresyl violet stains to effectively assess the DLT toxicity. Cresyl violet stained sections were used to measure the thickness of the various cerebellar layers (restricted to vermis region only) by using on ocular micrometer scale at 30 randomly selected sites. The data obtained was analysed using student's t-test. 3. Results The body and brain weights were recorded to be very significantly low in the treated animals as compared to their respective controls. The control pups gained body weight
Table 1. Experimental plan
a Vehicle only (equal injectable volume of propylene glycol). b Deltamethrin dissolved in propylene glycol at a dosage of 0.7 mg/kg./body wt./day, i.p. c pups per age group. No mortality.

120 Ν Patro et al
Table 2. Effect of deltamethrin on average body weight and brain weight (g) during postnatal development of rat.
Values are expressed as mean Level of significance (Student’s t-test) aP< 0.00005; b P < 0.01; cp < 0.2; d p < 0.001.
Figure 1. Effect of deltamethrin on average (a) body weight and (b) brain weight of rats during postnatal development.
rapidly while the DLT treated pups grew only at a slower rate till the postnatal day 30. Both body and brain weight remained significantly low in the treated animals even after withdrawal of treatment (table 2; figure 1).
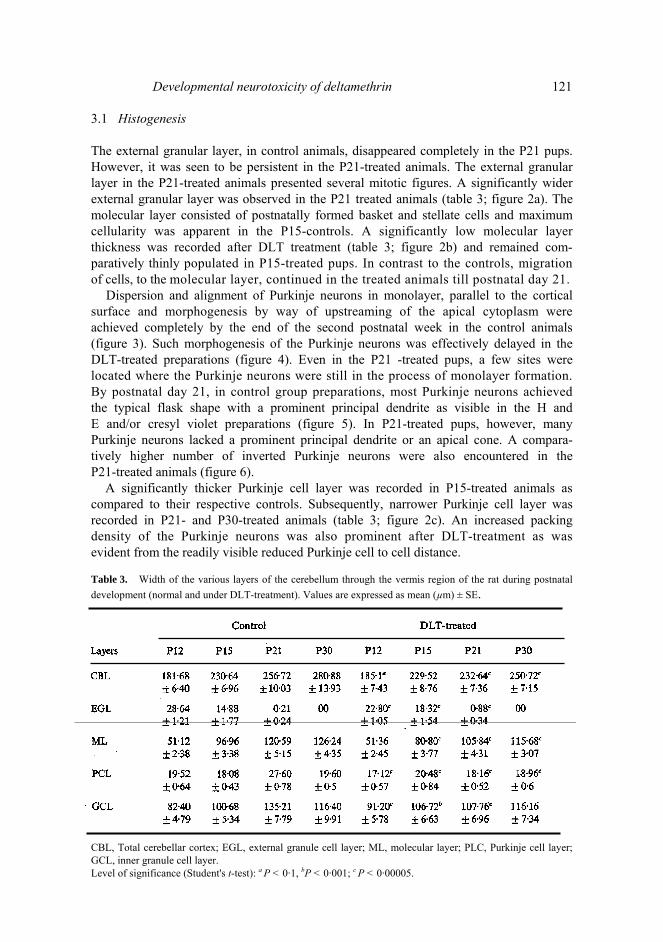
Developmental neurotoxicity of deltamethrin 121 3.1 Histogenesis The external granular layer, in control animals, disappeared completely in the P21 pups. However, it was seen to be persistent in the P21-treated animals. The external granular layer in the P21-treated animals presented several mitotic figures. A significantly wider external granular layer was observed in the P21 treated animals (table 3; figure 2a). The molecular layer consisted of postnatally formed basket and stellate cells and maximum cellularity was apparent in the P15-controls. A significantly low molecular layer thickness was recorded after DLT treatment (table 3; figure 2b) and remained com-paratively thinly populated in P15-treated pups. In contrast to the controls, migration of cells, to the molecular layer, continued in the treated animals till postnatal day 21.
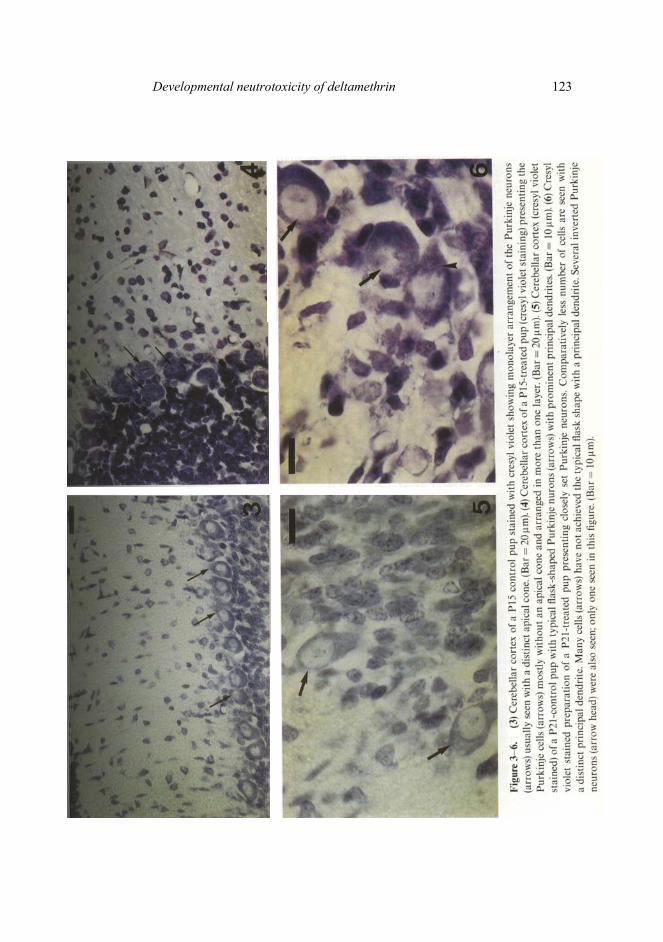
Dispersion and alignment of Purkinje neurons in monolayer, parallel to the cortical surface and morphogenesis by way of upstreaming of the apical cytoplasm were achieved completely by the end of the second postnatal week in the control animals (figure 3). Such morphogenesis of the Purkinje neurons was effectively delayed in the DLT-treated preparations (figure 4). Even in the P21 -treated pups, a few sites were located where the Purkinje neurons were still in the process of monolayer formation. By postnatal day 21, in control group preparations, most Purkinje neurons achieved the typical flask shape with a prominent principal dendrite as visible in the Η and Ε and/or cresyl violet preparations (figure 5). In P21-treated pups, however, many Purkinje neurons lacked a prominent principal dendrite or an apical cone. A compara-tively higher number of inverted Purkinje neurons were also encountered in the P21-treated animals (figure 6).
A significantly thicker Purkinje cell layer was recorded in P15-treated animals as compared to their respective controls. Subsequently, narrower Purkinje cell layer was recorded in P21- and P30-treated animals (table 3; figure 2c). An increased packing density of the Purkinje neurons was also prominent after DLT-treatment as was evident from the readily visible reduced Purkinje cell to cell distance. Table 3. Width of the various layers of the cerebellum through the vermis region of the rat during postnatal development (normal and under DLT-treatment). Values are expressed as mean (µm) ± SE.
CBL, Total cerebellar cortex; EGL, external granule cell layer; ML, molecular layer; PLC, Purkinje cell layer; GCL, inner granule cell layer. Level of significance (Student's t-test): a Ρ < 0·1, bP < 0·001; c Ρ < 0·00005.

122 Ν Patro et al
Figure 2. Width of the cerebellar layers in the vermis of control and deltamethrin treated rats during postnatal development, (a) External granule layer, (b) molecular layer, (c) Purkinje cell layer and (d) granule cell layer.
In the control group preparations the granule cell layer thickness constantly
increased during 12 to 21 day postpartum when the proliferation and migration of cells were prominent. Subsequently, by day 30 the granule cell layer thickness dropped significantly. In P21-treated group preparations the granule cell layer thickness remained very significantly low as compared to the controls. However, in P30 preparations the granule layer thickness was insignificantly different between the controls and treated (table 3; figure 2d). As has already been mentioned the prolifer-ation and migration of cells was delayed and continued even after day 21 in DLT-treated animals.
The total cerebellar cortex thickness also remained low in the DLT -treated animals as compared to the controls during the entire period of study (table 3). 3.2 DLT and the developing vasculature
The choroid plexus consisted of a mass of large thin walled and often fenestrated capillaries projecting into the ventricles and invested by modified ependymal cells. The
Developmental neutrotoxicity of deltamethrin 123

124 Ν Patro et al choroid epithelial cells were seen separated from the underlying capillaries and their supporting connective tissue by a basement membrane. This plexus remained unaffec-ted in the control animals (figure 7). In the DLT-treated animals starting from the postnatal day 12, major changes in the vasculature of the cerebellum were observed. The P15-treated animals had aggregates of blood cells in their capillaries which in the sections appeared to be clots and formed thrombus (figure 8). Such clots were recorded even in P21- and P30- treated pups. The vasculature in the subpial space and those extending into the perivascular space were dilated and filled with blood (hyperemia) (figure9). The tissue close to such dilated and/or thrombus capillaries, presented various degrees of infarct which appeared to have resulted from a sudden reduction in blood flow. At several sites, focal degeneration or even spongy appearance of the tissue has been recorded (figure 10). 4. Discussion Extensive period of development beginning from the embryonic organogenesis to the postnatal infantile period, the high susceptibility of the undifferentiated neural cells and the lack of self-renewing ability of neurons make the mammalian brain highly vulnerable to the environmental factors (Koestner and Norton 1991; Inouye 1995). During early postnatal development of the brain, the proper acquisition of position of the migrating neurons, through active migration, ultimately affects their morphology, synaptic connectivity and position. Any disturbance to the migrating neurons may cause congenital brain anomalies (Barth 1987; Hatten 1990).
During the normal development of the cerebellar cortex, the granule cell prolifer-ation and migration to the inner granular layer and deeper parts of the molecular layer are complete by the end of the postnatal day 20 and the external granular layer almost disappears in the 21 day postpartum animals (Clos et al 1977; Lewis et al 1975). Similar results have been recorded during the present investigation in the control group preparations. However, in the DLT-treated group animals, the external granular layer shows a slow migration of neurons as evident from the persistence of the layer along with several mitotic figures even at 21 day postpartum. This may probably be a compensatory proliferation beyond the normal growth period. Such results are in line with the reports available on the compensatory cell proliferation seen as a result of undernutrition (Gopinath 1984; Dobbing et al 1971; Barnes and Altman 1973). Continued existence of external granular layer and migration of granule cells have also been reported in thyroid deficiency (Legrand 1967) and treatment with corticosteroids (Cotterrell et al 1972). Such continued migration of granule cells till postnatal day 21, in DLT-exposed animals, adds further evidence to the fact that DLT delays and/or restricts differentiation of micro-neurons in the cerebellum.
The molecular layer widens up after first postnatal week with the onset of arboriz-ation of Purkinje cell dendrites (Altman 1969, 1972). In this study the animals exposed to DLT had low molecular layer thickness as compared to their respective controls. In the control animals the maximum cellularity of the molecular layer has been observed in the P15 pups, which is in accordance with the findings of Sima and Persson (1975), Pysh and Perkins (1975), Berry and Bradley (1976), Pysh et al (1979) and Gopinath et al (1987). However, the P15-treated animals had few differentiated basket cells, thereby, suggesting the possibility of delayed proliferation of pre-migratory cells in developing

Developmental neutrotoxicity of deltamethrin 125

126 Ν Patro et al brain consequent upon DLT-treatment. Many researchers have interpreted that a narrow molecular layer may result due to the reduced dendritic arborization of the Purkinje cells (McConnell and Berry 1978; Pysh et al 1979; Gopinath 1984). The delayed appearance of the apical cone in Purkinje neurons of DLT-treated animals, as prominently observed in the present investigation, may also result in the reduced Purkinje cell arborization.
By postnatal day 12, the Purkinje neurons have been observed to be clearly arranged in a monolayer in control animals which is in accordance with the findings of Sima and Persson (1975), Choudhary et al (1982) and Gopinath et al (1987). A delay in monolayer arrangement of Purkinje cells in P15-treated and even, at some sites in P21-treated animals suggest a general retardation of histogenesis and morphogenesis of the cerebellum. Such adverse effects have also been recorded consequent upon hy-pothyroidism (Clos et al 1973), low protein diet (West and Kemper 1976), undernutri-tion (Lewis et al 1975; Choudhary et al 1982), irradiation with high doses of X-rays (Altman and Anderson 1972, 1973; Berry and Bradely 1976), internal irradiation (Gopinath et al 1987) and chloroquine toxicity (Agarwal 1991).
Dobbing et al (1971), Dobbing and Smart (1974) and Gopinath (1984) observed that the brain weight improved considerably on withdrawal of the experimental undernutri-tion in comparison to body weight showing a brain sparing effect. Withdrawal of DLT exposure, however, did not present any brain sparing effect during the tenure of this study.
The capillaries in the sub-pial space presented various degrees of inflammation like vasodilatation, exudation and even immigration due to DLT administration. Several sites with thrombus and finally patchy haemorrhages were also observed. Ultrastruc-tural changes in the vascularization have been recorded by Banerjee et al (1988) due to internal administration of isotopes. The blood vessels are formed during the immediate postnatal period (Donahue and Pappas 1961) and exposure to any toxic substance at this stage may not only retard the process of proliferation of micro vessels but also result in progressive severity of changes with time. Severity of such changes recorded during 12-21 day postpartum are in accordance with the observations made by Dimitrievich et al (1984) and Banerjee et al (1988) on exposure to radiation. It has also been clearly observed that vascular damage is focal and neuronal tissue in the vicinity of such damage presents patchy degeneration. Koestner and Norton (1991) explained such changes and their resultant as: a reduction in blood supply to the nervous tissue could result in several types of alterations including generalized hypotension (causing hypoxia) and capillary damage (causing both hypoxia and ischemia). It is also believed that primary damage to capillaries is an acute response to high levels of cytotoxic agents such as heavy metals like arsenic (Harding et al 1968) and lead (Pounds 1984). Acute arsenic intoxication also cause petechial haemorrhages in many CNS areas similar to what has been observed in the present study.
It is evident that the DLT-treated pups had comparatively less populated and thinner granular layer, particularly at postnatal day 21, which can be explained on the basis of neural-darwinism. Ever since 1929, when Ramon Υ Cajal proposed that during neurogenesis perhaps a competitive struggle exists among the outgrowths and even among the nerve cells for space and nutrition, efforts are on to evaluate if neuronal development is darwinian (Purves et al 1996). Neural darwinists hold that the brain during early development has more neural elements (like neurons, neural branches, synapses and the groups of interconnected neurons) and loss of some neural elements has important implications in the development of the nervous system (Purves and

Developmental neutrotoxicity of deltamethrin 127 Lichtman 1985). It is known that excess number of neurons are generated in ontogeny which could be to the tune of two- or three-folds to the population present in the mature brain (Openheim 1981). While neural counting has raised technical problems, it is almost certain and rational to accept that the process of generation of excessive neural elements and subsequent pruning is a epigenetic strategy for matching the neuronal centers and their targets which Purves et al (1996) believe to have all the characteristics of competition. Electrophysiological studies also confirmed that inputs are significant-ly eliminated during postnatal development and the process is also thought to be competitive (Purves and Lichtman 1980). Thus it can safely be accepted that excessive cells in early development enables a suitable competition. In this study we have recorded an initial excessive thickening of the granule cell layer (particularly by day P21) and subsequent thinning could be due to pruning of granule cells which could not establish connections. In DLT-treated pups the formation of the cells of the inner granular layer is very significantly delayed and/or retarded, thereby, restricting a com-petition or even a suitable excess of connectivity to ensure that the required number of synapses are positively established. However, by day 30, the layer thickness nearly reaches that of the controls. It is thus evident that DLT interferes the initial develop-mental phenomenon substantially. Further studies with morphometric analysis of cellular density, which is in progress may add to the proposition.
Experimental studies with undernutrition have suggested significant adverse effects on the functional development of the brain as a result of retardation of morphological and biochemical maturation (Winick 1969; Balazas and Patel 1973; Dobbing and Smart 1974). Malnutrition in children, during early life, permanently impaired the intellectual performance (Hertzig et al 1972; Tizard 1974). DLT administration in postnatal rats have presented most changes similar to what has been observed in conditions of undernutrition, which raises a great concern. Postnatal exposure to DLT has been observed to generally delay the process of cytogenesis and/or morphogenesis of various types of micro-neurons and inter-neurons (including Purkinje neurons) during the tenure of treatment. The compound has also been observed to damage the developing vasculature in the cerebellum. Thus the present study opens up several questions on the safety of this substance to the pregnant mothers and new-born babies in the habitat where this substance is in common use for vector control. Acknowledgment The authors are grateful to the Department of Science and Technology, New Delhi for the financial support through the Young Scientist Project grant (Sanction No. SR/OY/Z-10) to NP. References Agarwal Μ 1991 Effects of chloroquine on the postnatal development of mouse cerebellar cortex, M. Phil
Dissertation, Jiwaji University, Gwalior Aldridge W Ν 1990 An assessment of the toxicological properties of Pyrethroids and their toxicity; CRC Rev.
Toxicol 21 89-104 Altman J 1969 Autoradiographic and histological studies of postnatal neurogenesis, II—A longitudinal
investigation of the kinetics, migration and transformation of cells incorporating tritiated thymidine in rats, with special reference to postnatal neurogenesis in some brain regions; J. Comp. Neurol. 128 431-474

128 N Patro et al Altman J 1972 Postnatal development of the cerebellar cortex in the rat: The external germinal layer and the
transition molecular layer; J. Comp. Neurol. 145 353-398 Altman J and Anderson W J 1972 Experimental reorganization of the cerebellar cortex, part I (Morphologi-
cal effects of elimination of all microneurons with prolonged X-irradiation started at birth); J. Comp. Neurol. 145 355-405
Altman J and Anderson W J 1973 Experimental reorganization of the cerebellar cortex, part II (Effects of elimination of most microneurons with X-irradiation started at 4 days); J. Comp. Neurol. 149 123-152
Balazas R and Patel A J 1973 Factors afecting the biochemical maturation of the brain. Effect of undernutrition during early life; Prog. Brain Res. 40 115-128
Banerjee R, Gopinath G and Gopinath Ρ G 1988 Vascular changes in the brain following internally administered radioisotope I131 in rats during postnatal period; Indian J. Med. Res. 87 484-493
Barnes D and Altman J 1973 Effects of different schedules of early undernutrition on the preweaning growth of the rat cerebellum; Exp. Neurol. 38 406-419
Barth Ρ G 1987 Disorders of neuronal migration; J. Neurol. Sci. 14 1-16 Berry Μ and Bradley Ρ 1976 The growth of the dendritic trees of Purkinje cells in irradiated agranular
cerebellar cortex; Brain Res. 116 361-387 Bloom A S, Staatz C G and Dieringer Τ 1993 Pyrethroid effects on operant responding and feeding;
Neurobehav. Toxicol. Teratol 5 321-324 Casida J E, Gammon D W, Glickman Α Η and Lawrence L J 1983 Mechanisms of selective action of
pyrethroid insecticides; Annu. Rev. Pharmacol Toxicol. 23 413-438 Chanh Ρ Η, Navarro-Delmasure C, Chanh ΑΡΗ , Cheav S L, Ziade F and Samaha F 1984 Pharmacological
effects of deltamethrin on the central nervous system; Arzneim.-Forsch 34 175-181 Choudhary C, Gopinath G and Roy S 1982 Effects of undernutrition on the maturation of Purkinje cells in the
rat; Indian. J. Med. Res. 75 559-566 Clos J, Favre C, Selme-Matrat and Legrand J 1977 Effects of undernutrition on cell formation in the rat brain
and specially on cellular composition of the cerebellum; Brain Res. 123 13-26 Clos J, Rebiere A and Legrand J 1973 Differential effects of hypothyroidism and undernutrition on the
development of glia in the rat cerebellum; Brain Res. 63 445-449 Coppage D L and Braidech Τ Ε 1976 River pollution by anticholinesterase agents; Water Res. 10 19-24 Cotterrell M, Balazs R and Johnson A L 1972 Effects of corticosteroids on the biochemical maturation of rat
brain: Postnatal cell formation; J. Neurochem. 19 2152-2167 Cremer J Ε and Seville Μ Ρ 1982 Comparative effects of two Pyrethroids, deltamethrin and cismethrin on
plasma catecholamines and on blood glucose and lactate; Toxicol. Appl. Pharmacol. 66 124-133 Crofton Κ Μ and Reiter L W 1984 Effects of two pyrethroid insecticides on motor activity and the acoustic
startle response in the rat; Toxicol. Appl. Pharmacol. 75 318-328 Crofton Κ Μ and Reiter L W 1987 Pyrethroid insecticides and the y-aminobutyric acid. A receptor complex:
Motor activity and the startle response in the rat; J. Pharmacol. Exp. Therap. 243 946-954 Depeng L, Deling L, Renguo Y, Peng L, Xueguang Η and Aimin L 1994 Alphamethrin impregnated bednets
for malaria and mosquito control in China; Trans. R. Soc. Trop. Med. Hyg. 88 625-628 Dimitrievich G S, Fischer-Dzoga Κ and Griem Μ L 1984 Radiosensitivity of vascular tissue. 1 Differential
radiosensitivity of capillaries. A quantitative in vivo study; Rad. Res. 99 511-535 Dobbing J and Smart J L 1974 Vulnerability of developing brain and behaviour; Br. Med. Bull. 30
164-168 Dobbing J, Hopewell J W and Lynch A 1971 Vulnerability of the developing brain. VII. Permanent deficit of
neurons in cerebral and cerebellar cortex following early mild undernutrition; Exp. Neurol. 32 439-447 Donahue S and Pappas G D 1961 The fine structure of capillaries in the cerebral cortex of the rat at various
stages of development; Am. J. Anat. 108 331-347 Elliot Μ 1976 Properties and application of Pyrethroids; Environ. Health Perspect. 14 3-13 Epstein Η Τ and Stanford Α Μ 1977 Nutr. Rep. Intl. 16 366 (Quoted from Gopinath 1984) Gilbert Μ Ε, Mack C and Crofton Κ Μ 1990 An examination of the proconvulsant actions of pyrethroid
insecticide using pentylentetrazol and amygdala kindling seizure models; Neurotoxicology 11 73-86 Glomot R 1982 Toxicity of Deltamentrin to higher vertebrates; Deltamethrin Monograph (Paris: Roussel-
Uclaaf) Gopinath G 1984 Experimental undernutrition and morphology of the brain; in INSA Status report series-I
(eds) Ρ Ν Tandon and G Gopinath, pp 9-37 Gopinath G, Banerjee R and Gopinath Ρ G1987 Effects of internal irradiation on the maturing Purkinje cells
in the rat—A Golgi study; J. Neurol. Sci. 78 93-109

Developmental neutrotoxicity ofdeltamethrin 129 Harding J D J, Lewis G and Done J Τ 1968 Experimental arsenilic acid poisoning in pigs; Vet. Rec. 83
560-564 Hatten Μ Ε 1990 Riding the glial monorail: A common mechanism for neuronal migration in different
regions of the developing mammalian brain; Trends Neurosci. 13 179-184 Hayes W J 1982 Pesticides derived from plants and other organisms; in: Pesticides studies in man (ed.)
W J Hayes (Baltomore: Williams and Winkins) pp 75-111 Hertzig Μ Ε, Birch Η G, Richardson S A and Tizard J 1972 Intellectual levels of school children severely
malnourished during the first two years of life; Pediatrics 49 814-824 Hijzen Τ Η and Slangen J L 1988 Effects of type I and type II Pyrethroids on the startle response in rats;
Toxicol. Lett. 40 141-152 Inouye Μ 1995 Radiation-induced apoptosis and developmental disturbance of the brain; Cong. Anom. 35
1-13 Janakara Β R, Jihullah W A, Shahi B, Dev V, Curtis C F and Sharma V Ρ 1995 Deltamethrin impregnated
bednets against Anopheles minimus transmitted malaria in Assam, India; J. Trop. Med. Hyg. 98 73-83 Koestner A and Norton S 1991 Nervous system: A Handbook of toxicologic pathology (London: Academic
Press) pp 625-674 Legrand J 1967 Analyses del'action morphogenetique des hormones thyroidiennes sur le cervelet du jeune
rat; Ach. Anat. Microsc Morphol. Exp. 56 205-244 Lewis Ρ D, Balazs R, Patel A J and Johnson A L 1975 The effect of undernutrition in early life on cell
generation in the rat brain; Brain Res. 83 235-247 Lund Α Ε and Narahashi Τ 1983 Kinetics of sodium channel modification as the basis for the variation in
nerve membrane, effects of the Pyrethroids and DDT analogs; Pestic. Biochem. Physiol. 20 203-216 McConnell Ρ and Berry Μ 1978 Effects of undernutrition on Purkinje cell dendritic growth in the rat; J.
Comp. Neurol. 177 159-172 Moretto A and Johnson Μ Κ 1987 Toxicology of organophosphates and carbamates; in Toxicology of
pesticides: Experimental, clinical and regulatory perspectives (eds) L G Costa, C L Galli and S D Murphy (Berlin: Springer-Verlag) pp 33-48
MoyouSomo R, Lehman L G, Awahmukalah S and Enyong Ρ A 1995 Deltamethrin impregnated bednets for the control of urban malaria in Kumba town, South-West province of Cameroon; J. Trop. Med. Hyg. 98 319-324
Narahashi Τ 1980 Site and type of action of Pyrethroids on nerve membrane; in Pyrethroid insecticides: Chemistry and action (eds) J Ε Casida and Μ Elliot (Paris: Table Ronde Roussel-Uclaaf, No. 37)
Narahashi Τ 1992 Nerve membrane Na+ channels as targets of insecticides; Trends Pharmacol. 13 236-241 Oppenheim R W 1981 Neuronal cell death and some related regressive phenomena during neurogenesis:
A historical review and a progress report; in Studies in developmental neurobiology: Essays in honour of Viktor Hamburger (ed.) W Μ Cowan (New York: Oxford University Press) pp 74-133
Paxinos G, Ashwell Κ W S and Tork I 1994 Atlas of the developing rat nervous system, 2 edition (London: Academic Press)
Peele D Β and Corfton Κ Μ 1987 Pyrethroid effect on schedule controlled behavior: Time and damage relationships; Neurotoxicol. Teratol. 9 387-394
Pounds J G 1984 Effects of lead intoxication on calcium homeostasis and calcium-mediated cell function: A review; Neurotoxicology 5 295-332
Purves D and Lichtman J W 1980 Elimination of synapses in the developing nervous system; Science 210 153-157
Purves D and Lichtman J W 1985 Principles of neural development (Sunderland, Massachusetts: Sinauer) Purves D, White L Ε and Riddle D R 1996 Is neural development darwinial?; Trends Neurosci. 19
460-464 Pysh J J and Perkins R Ε 1975 Undernutrition and Purkinje cell development; Neurosci. Abstr. 1 756 Pysh J J, Perkins R Ε and Singer Β L 1979 The effects of postnatal undernutrition on development of the
mouse Purkinje cell dendritic tree; Brain Res. 163 165-170 Ramon Υ and Cajal S 1929 Studies on vertebrate neurogenesis (Springfield, IL: Thomas) Ray D Ε 1980 An EEG investigation of decamethrin-induced choreoathetosis in the rat; Exp. Brain Res. 38
221-277 Reddy A T, Ayyanna Κ and Yellama Κ 1991 Sensitivity of brain Cholinesterase to cypermethrin toxicity in
freshwater teleost Telapia mossambica; Biochem. Int. 23 959-962 Rickard J and Brodie Μ Ε 1985 Correlation of blood and brain levels of the neurotoxic pyrethroid
deltamethrin with the onset of symptoms in rats; Pest. Biochem. Physiol. 23 143-156

130 N Patro et al Rose G Ρ and Deewar A J 1983 Intoxication of four synthetic Pyrethroids fails to show any correlation
between neuromuscular dysfunction and neurochemical abnormalities in rats; Arch. Toxicol. 53297-316 Sima A and Persson L 1975 The effects of pre- and post-natal undernutrition on the development of the rat
cerebellar cortex, I. Morphological observations; Neurobiology 5 23-43 Tizard J 1974 Early malnutrition, growth and mental development in man; Br. Med. Bull. 30 169-174 Vijverberg Η Ρ Μ and van den Bercken J 1990 Neurotoxicological effects and the mode of action of
pyrethroid insecticides; CRC Rev. Toxicol. 21 105-126 West C D and Kemper Τ L 1976 The effect of a low protein diet on the anatomical development of the rat
brain; Brain Res. 107 221-237 Winick Μ 1969 Malnutrition and brain development; J. Pediatr. 74 667-679 Woodward D J, Beckett D and Chanda A 1975 Purkinje cell dendritic alterations after transient develop-
mental injury of the external granular layer; Brain Res. 97 195-214
Corresponding editor: INDIRA NATH