Neurotoxicological effects of deltamethrin on the postnatal development of cerebellum of rat

B
O
Pcm
Va
b
c
ARAA
KDNAOLAN
1
tpetcpthheiz
2h
ARTICLE IN PRESSG ModelIOMAG-116; No. of Pages 8
Biomedicine & Aging Pathology xxx (2013) xxx–xxx
Available online at
ScienceDirectwww.sciencedirect.com
riginal article
yrethroid deltamethrin-induced developmental neurodegenerativeerebral injury and ameliorating effect of dietary glycoside naringin inale wistar rats
inayagam Magendira Mania, Sivaji Ashab, Abdul Majeeth Mohamed Sadiqc,∗
Department of Biochemistry, Islamiah College (Autonomous), Vaniyambadi 635751, Tamil Nadu, IndiaDepartment of Biochemistry, DKM College for Women’s (Autonomous), Vellore 632001, Tamil Nadu, IndiaDepartment of Biochemistry, Adhiparasakthi College of Arts and Science (Autonomous), Kalavai 632506, Tamil Nadu, India
a r t i c l e i n f o
rticle history:eceived 17 October 2013ccepted 14 November 2013vailable online xxx
eywords:eltamethrinaringinntioxidant enzymesxidative stressipid peroxidationcetyl choline esteraseeurotoxicity
a b s t r a c t
Deltamethrin (DEL) is a type II � - cyano group containing synthetic pyrethroid insecticide that is usedextensively for controlling flies, mosquitoes, insects, pests worldwide. DEL exposure leads to pathophysi-ology of a broad spectrum of cerebrovascular & neurodegenerative disorders like Parkinson’s, Lou Gehrig’sdisease, Alzheimer’s disease, developmental deficits, birth defects, and learning disabilities. In these stud-ies, we have demonstrated that the deltamethrin-induced neurotoxicity and ameliorating effect of dietaryflavonoid naringin by its antioxidant and neuro-protective ability in male wistar rat. Adult male wistarrats were divided into four different groups. Group I vehicle treated control group; group II receiveddeltamethrin dissolved in corn oil 12.8 mg/kg BW orally (1/10 LD 50) for three weeks (21 days) to induceneurotoxicity; group III received naringin (100 mg/kg BW for 21 days) orally. Group IV naringin alonetreated. DEL-induced neurotoxicity was evidenced by increased activities of creatine phosphokinase,lactate dehydrogenase, TBARS in DEL administered rat brain tissue homogenate, and decrease activity ofacetylcholinesterase (AChE) activities, however it was reversed by naringin treatment. DEL administered
rats showed reduction in the levels of enzymic (superoxide dismutase, catalase, glutathione peroxidase,glutathione reductase and glutathione-S-transferase) and non-enzymic antioxidants (glutathione, vita-min C and vitamin E) levels. However, normalized antioxidant and non-enzymic antioxidant defenseswere reported in the naringin treated rats. These findings highlight the efficacy of naringin as a neuro-protectant in DEL-induced neurotoxicity which is also supported by native gel electrophoresis, agarosepatho
gel electrophoresis, histo. Introduction
Pestidices are worldwide used synthetic chemicals, designedo kill pests such as insects, weeds, microbes, but they also haveotential harmful toxic effects on human health. Humans arexposed to pesticides via inhalation, dermal contact and inges-ion due to occupational exposure, in the environment throughontaminated food and suicidal exposure [1]. Pesticides are sus-ected to have harmful neurological, cardiotoxic, hepatotoxic, renaloxic, immunological, reproductive and cancerous health effects toumans [1]. Agricultural pesticides have become a common house-old item in rural areas of the developing world. Due to their
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
asy availability, pesticides have also become commonly used forntentional self-poisoning [2]. According to World Health Organi-ation (WHO) estimates published in 2010 [3], around 5 million
∗ Corresponding author.E-mail address: [email protected] (A.M.M. Sadiq).
210-5220/$ – see front matter © 2013 Published by Elsevier Masson SAS.ttp://dx.doi.org/10.1016/j.biomag.2013.11.001
logical studies of rat brain tissue.© 2013 Published by Elsevier Masson SAS.
poisoning cases with 3, 50,000 deaths occur annually. Pesticide self-poisoning accounts for one third of the world suicides [4]. In India,last September 2013, it was reported that there were over a quar-ter of a million farmers’ suicides between 1995 and 2013, the worstever recorded wave of suicides in human history. In 2009, the stateof Tamil Nadu in India witnessed a total of 14,424 suicides [5,6]. InIndia, July 2013, 56 school children’s were died after eating lunchcontaminated with “very toxic” levels of an agricultural pesticidenamely monocrotophos.
The exposure of humans to pyrethroid pesticide may thereforebe extensive [7], which increases the health risk. Pyrethroids aresynthetic insecticides derived from the naturally occurring botan-ical insecticide mixture, pyrethrum. Deltamethrin ((S) � – cyano- 3- phenoxybenzyl - (1R) – cis – 3 - (2, 2 - dibromovinyl) -2, 2-dimethylcyclopropane carboxylate), one of the most potent
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
pyrethroid insecticides with �- cyano group, produces type IIneurological syndrome “choreoathetosis with salivation” or “CS”syndrome, which is characterized by salivation and convulsionschoreoathetosis [8]. Furthermore, epidemiological studies have

ING ModelB
2 ne & A
soedhatcti
bawaiddav[aagpaiapopmtn
2
2
9elAu
2
hmn(cian
2
3wia
ARTICLEIOMAG-116; No. of Pages 8
V. Magendira Mani et al. / Biomedici
hown that the use of deltamethrin pesticide leads increase the riskf neurodegenerative disorders like Parkinson’s, Lou Gehrig’s Dis-ase (ALS), Alzheimer’s Disease, ADHD, autism, and developmentaleficits, birth defects, learning disabilities [9]. Several mechanismsave been suggested for the neural toxicity of deltamethrin. Incute exposure, deltamethrin blocks nerve impulse by modifyinghe kinetics of sodium channels, GABA receptors, nicotinic acetyl-holine receptors and excitatory glutamate receptors [8]. Hence,he present study to evaluate the deltamethrin induced neurotox-city and the ameliorating effect of naringin.
Naringin (4′, 5, 7-trihydroxyflavanone-7-rhamnoglucoside)elongs to a family of C6-C3-C6 polyphenol compounds whichre widely distributed in foods of plant origin [10]. Naringin isidely distributed in citrus and occurs in grapefruits, mandarins
nd other variety of citrus fruits [11]. It gives bittering principaln citrus juices. Flavonoids occur ubiquitously in the plant king-om and are common components of the human diet [12]. Theiverse effects of naringin is to their structural similarity to ATPnd hence to their ability to compete with ATP for binding toarious enzymatic sites and easy to move in blood brain barriers13]. Naringin possess significant anti-hepatotoxic, anti-allergic,nti-inflammatory, anti-osteoporotic, anti-cancer, hypolipidemicctivities metal chelating [14], antioxidant by up-regulating theene expression of superoxide dismutase, catalase and glutathioneeroxidase, free radical scavenging properties, antiradical agents,ntimicrobial [15,16] and anti-lipoperoxidation activities; anti-nflammatory, anti-apoptotic, anti-atherogenic, anti-ulcer activity,nti-hyperlipidemic [17]. Previously studies shown that naringinrevents the radiation induced tumor development. [15,18]. More-ver, naringin demonstrated inhibition of breast cancer cellroliferation and delay of mammary tumorigenesis [19], yet noutagenic activity. Therefore, the present study was undertaken
o investigate the effect of naringin against deltamethrin-inducedeurotoxicity in male wistar rats.
. Martials and methods
.1. Chemicals
Deltamethrin commercial formulation Decis, deltamethrin7.3% EC [denotes 97.3% of technical grade deltamethrin (w/w) inmulsifiable concentrate] was procured from Bayer crop scienceimited, Mumbai, India. Naringin were purchased from Sigma-ldrich Co. (St. Louis, MO, USA). All other chemicals and reagentssed in this study were of analytical grade.
.2. Animal model
Wistar male albino rats weighing between 200 g–240 g wereoused in animal cages with food and water ad libitum. Six ani-als were housed per cage, and maintained on 12/12 hr/day and
ight cycle. The animals were fed with commercial pellet dietHindustan lever Ltd., Bangalore, India). The experiments wereonducted according to ethical norms approved by the Min-stry of Social Justices and Empowerment, Government of Indiand Institutional Animal Ethics Committee Guidelines (Approvalo. IAEC/APCAS/01/2013/01).
.3. Standardization of induction of neurotoxicity in rat model
Deltamethrin [(S) – alpha - cyano – 3 – phenoxybenzyl (1R, 3R) –
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
- (2, 2-dibromovinyl) - 2, 2- Dimethylcyclo - propanecarboxylate]as dissolved in corn oil, and it was used for inducing neurotoxicity
n wistar rat, for a period of three weeks. For the induction, malelbino wistar rats (n = 6) was orally administered with deltamethrin
PRESSging Pathology xxx (2013) xxx–xxx
at a dosage (12.8 mg/kg b.wt. daily for three weeks). The admin-istration for deltamethrin induction used was based on its LD 50value and was administered orally to promote neurotoxicity. Theoral LD50 value of deltamethrin in male rats ranged 128 mg/kg B.W.Here, 1/10 LD 50 value of 12.8 mg/kg BW was chosen for this studies.
2.4. Experimental design
Experimental animals were divided into four groups of six ratseach as follows:
• group I – control rats (vehicle treated);• group II – rats induced with deltamethrin dissolved in corn oil
12.8 mg/kg BW orally (1/10 LD 50) for three weeks (21 days);• group III – rats administered with deltamethrin dissolved in
corn oil 12.8 mg/kg BW orally and simultaneously administered100 mg/kg BW of naringin dissolved in water orally for 21 days;
• group IV – drug alone rats (100 mg/kg BW of naringin alone dis-solved in water orally for 21 days).
After the experimental period, the animals were fastedovernight, anaesthetized with sodium-pentothal and blood col-lected from jugular vein for serum isolation and sacrificed bycervical decapitation. The brain tissue was excised immediately anda portion of the tissue was homogenized in 0.1 M Tris buffer, pH 7.4and used for various biochemical assays.
2.5. Histological examination
A portion of the brain tissue was fixed in 10% neutral bufferedformalin and embedded in paraffin wax for histological evaluation.Sections with thickness 5 �m were stained with hamatoxylin andeosin (H & E), examined under high power light microscope.
2.6. Biochemical assays
The collected serum was used to estimate antioxidant enzymessuch as superoxide dismutase SOD [20], catalase CAT [21], glu-tathione peroxidase GPx [22], glutathione reductase GR [23], andnon-enzymic antioxidants reduced glutathione GSH [24], vitamin C[25], vitamin E [26]. The brain tissue homogenate was used to mea-sure lactate dehydrogenase (LDH), was measured by the method of[27], cretinine kinase was measured by the method of [28]. Lipidperoxidation level was determined by measuring thio barbituricacid reactive substances (TBARS) according to the method of [29].The activity of AChE in the homogenate was assayed by the methodof [30].
2.7. Activity staining of SOD and CAT by native gel electrophoresis
The levels of protein were estimated by [31], were separated bynative gel electrophoresis in the presence of Tris (50 mM), glycine(300 mM) and EDTA (1.8 mM) at constant current (50 mA) for4–5 hrs. After electrophoresis, the 12% native gel was incubated instaining solution containing riboflavin (0.028 mM), nitroblue tetra-zolium (0.25 mM), EDTA (1 mM) and TEMED (28 mM), in 50 mMphosphate buffer. After 30 mins incubation in dark, the stainingsolution was removed and replaced with 50 mM phosphate buffer.The gel was then exposed to light for a few minutes and areasof SOD activity appeared as white bands in a blue background
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
[32]. Similarly, the 8% native gel of CAT after electrophoresis wassoaked in 50 mM phosphate buffer (pH 7.8) containing 0.01 MH2O2 for 20 mins at room temperature in dark with gentle rock-ing. The gel was then placed in 2% ferric chloride and 2% potassium
ARTICLE IN PRESSG ModelBIOMAG-116; No. of Pages 8
V. Magendira Mani et al. / Biomedicine & Aging Pathology xxx (2013) xxx–xxx 3
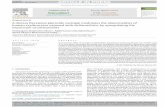
Fig. 1. Effect of DEL and naringin on the level of lipid peroxides in brain tissuesof control and experimental group rats. Results were expressed as mean ± SD, forsix animals in each group. Statistical significant variations are compared as fol-lv
fb
2
efi(m1im
2
eSafs
3
3
hgeclg
3
Grtaibo
Table 1Effect of DEL and naringin on the activities of SOD, CAT, GPx, GST and GR in theserum of control and experimental rats.
Particulars Control DEL-induced DEL-induced+naringin
Naringin alone
SOD 2.72 ± 0.57 1.40 ± 0.28* 2.09 ± 0.42* 2.57 ± 0.59NS
CAT 64.22 ± 5.99 42.63 ± 4.48* 57.05 ± 6.01* 62.88 ± 9.60NS
GPx 6.84 ± 0.85 3.65 ± 0.43* 5.34 ± 0.71* 6.65 ± 0.91NS
GST 2.98 ± 0.62 1.13 ± 0.33* 2.33 ± 0.44* 2.62 ± 0.97NS
GR 1.22 ± 0.03 0.74 ± 0.07* 1.01 ± 0.16* 1.16 ± 0.05NS
Units: SOD - 50% inhibition of epinephrine auto-oxidation/min/mg protein; CAT-�mole H2O2 consumed/min/mg protein; GPx - �gm of GSH consumed/min/mg pro-tein; GST - �moles of 1–chloro - 2- 4 - dinitrobenzene formed/min/mg protein andGR - �gm GSSH utilized/min/mg protein. Results were expressed as mean ± SD,
Fig. 5 shows the effect of DEL and naringin on tissue antioxidantsstatus of SOD and CAT activity staining by native gel electrophoresis
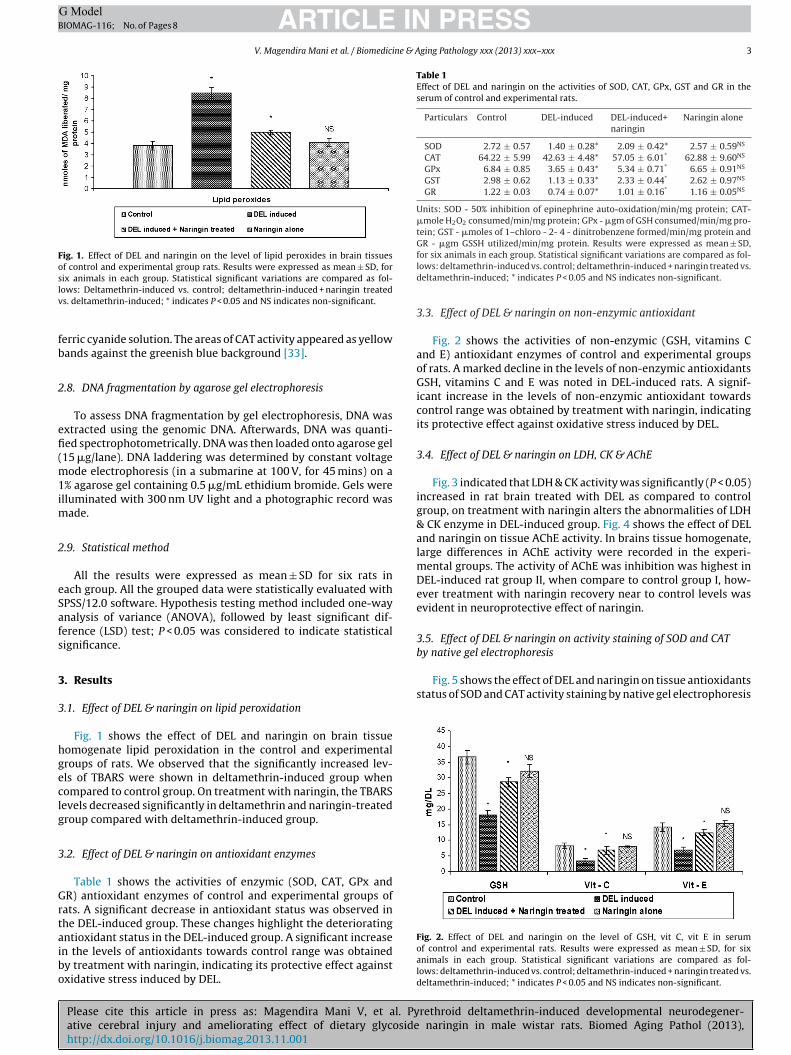
Fig. 2. Effect of DEL and naringin on the level of GSH, vit C, vit E in serum
ows: Deltamethrin-induced vs. control; deltamethrin-induced + naringin treateds. deltamethrin-induced; * indicates P < 0.05 and NS indicates non-significant.
erric cyanide solution. The areas of CAT activity appeared as yellowands against the greenish blue background [33].
.8. DNA fragmentation by agarose gel electrophoresis
To assess DNA fragmentation by gel electrophoresis, DNA wasxtracted using the genomic DNA. Afterwards, DNA was quanti-ed spectrophotometrically. DNA was then loaded onto agarose gel15 �g/lane). DNA laddering was determined by constant voltage
ode electrophoresis (in a submarine at 100 V, for 45 mins) on a% agarose gel containing 0.5 �g/mL ethidium bromide. Gels were
lluminated with 300 nm UV light and a photographic record wasade.
.9. Statistical method
All the results were expressed as mean ± SD for six rats inach group. All the grouped data were statistically evaluated withPSS/12.0 software. Hypothesis testing method included one-waynalysis of variance (ANOVA), followed by least significant dif-erence (LSD) test; P < 0.05 was considered to indicate statisticalignificance.
. Results
.1. Effect of DEL & naringin on lipid peroxidation
Fig. 1 shows the effect of DEL and naringin on brain tissueomogenate lipid peroxidation in the control and experimentalroups of rats. We observed that the significantly increased lev-ls of TBARS were shown in deltamethrin-induced group whenompared to control group. On treatment with naringin, the TBARSevels decreased significantly in deltamethrin and naringin-treatedroup compared with deltamethrin-induced group.
.2. Effect of DEL & naringin on antioxidant enzymes
Table 1 shows the activities of enzymic (SOD, CAT, GPx andR) antioxidant enzymes of control and experimental groups of
ats. A significant decrease in antioxidant status was observed inhe DEL-induced group. These changes highlight the deterioratingntioxidant status in the DEL-induced group. A significant increase
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
n the levels of antioxidants towards control range was obtainedy treatment with naringin, indicating its protective effect againstxidative stress induced by DEL.
for six animals in each group. Statistical significant variations are compared as fol-lows: deltamethrin-induced vs. control; deltamethrin-induced + naringin treated vs.deltamethrin-induced; * indicates P < 0.05 and NS indicates non-significant.
3.3. Effect of DEL & naringin on non-enzymic antioxidant
Fig. 2 shows the activities of non-enzymic (GSH, vitamins Cand E) antioxidant enzymes of control and experimental groupsof rats. A marked decline in the levels of non-enzymic antioxidantsGSH, vitamins C and E was noted in DEL-induced rats. A signif-icant increase in the levels of non-enzymic antioxidant towardscontrol range was obtained by treatment with naringin, indicatingits protective effect against oxidative stress induced by DEL.
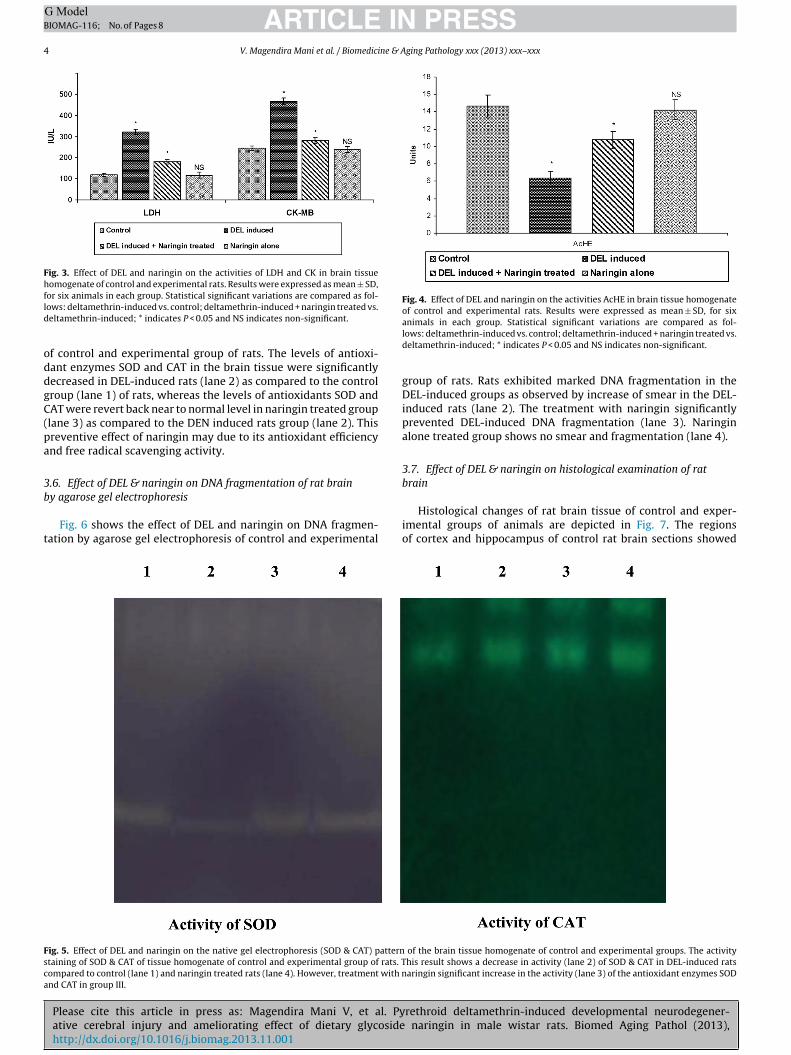
3.4. Effect of DEL & naringin on LDH, CK & AChE
Fig. 3 indicated that LDH & CK activity was significantly (P < 0.05)increased in rat brain treated with DEL as compared to controlgroup, on treatment with naringin alters the abnormalities of LDH& CK enzyme in DEL-induced group. Fig. 4 shows the effect of DELand naringin on tissue AChE activity. In brains tissue homogenate,large differences in AChE activity were recorded in the experi-mental groups. The activity of AChE was inhibition was highest inDEL-induced rat group II, when compare to control group I, how-ever treatment with naringin recovery near to control levels wasevident in neuroprotective effect of naringin.
3.5. Effect of DEL & naringin on activity staining of SOD and CATby native gel electrophoresis
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
of control and experimental rats. Results were expressed as mean ± SD, for sixanimals in each group. Statistical significant variations are compared as fol-lows: deltamethrin-induced vs. control; deltamethrin-induced + naringin treated vs.deltamethrin-induced; * indicates P < 0.05 and NS indicates non-significant.
ARTICLE IN PRESSG ModelBIOMAG-116; No. of Pages 8
4 V. Magendira Mani et al. / Biomedicine & Aging Pathology xxx (2013) xxx–xxx
Fig. 3. Effect of DEL and naringin on the activities of LDH and CK in brain tissuehomogenate of control and experimental rats. Results were expressed as mean ± SD,fld
oddgC(pa
3b
t
Fig. 4. Effect of DEL and naringin on the activities AcHE in brain tissue homogenateof control and experimental rats. Results were expressed as mean ± SD, for six
Fsca
or six animals in each group. Statistical significant variations are compared as fol-ows: deltamethrin-induced vs. control; deltamethrin-induced + naringin treated vs.eltamethrin-induced; * indicates P < 0.05 and NS indicates non-significant.
f control and experimental group of rats. The levels of antioxi-ant enzymes SOD and CAT in the brain tissue were significantlyecreased in DEL-induced rats (lane 2) as compared to the controlroup (lane 1) of rats, whereas the levels of antioxidants SOD andAT were revert back near to normal level in naringin treated grouplane 3) as compared to the DEN induced rats group (lane 2). Thisreventive effect of naringin may due to its antioxidant efficiencynd free radical scavenging activity.
.6. Effect of DEL & naringin on DNA fragmentation of rat brain
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
y agarose gel electrophoresis
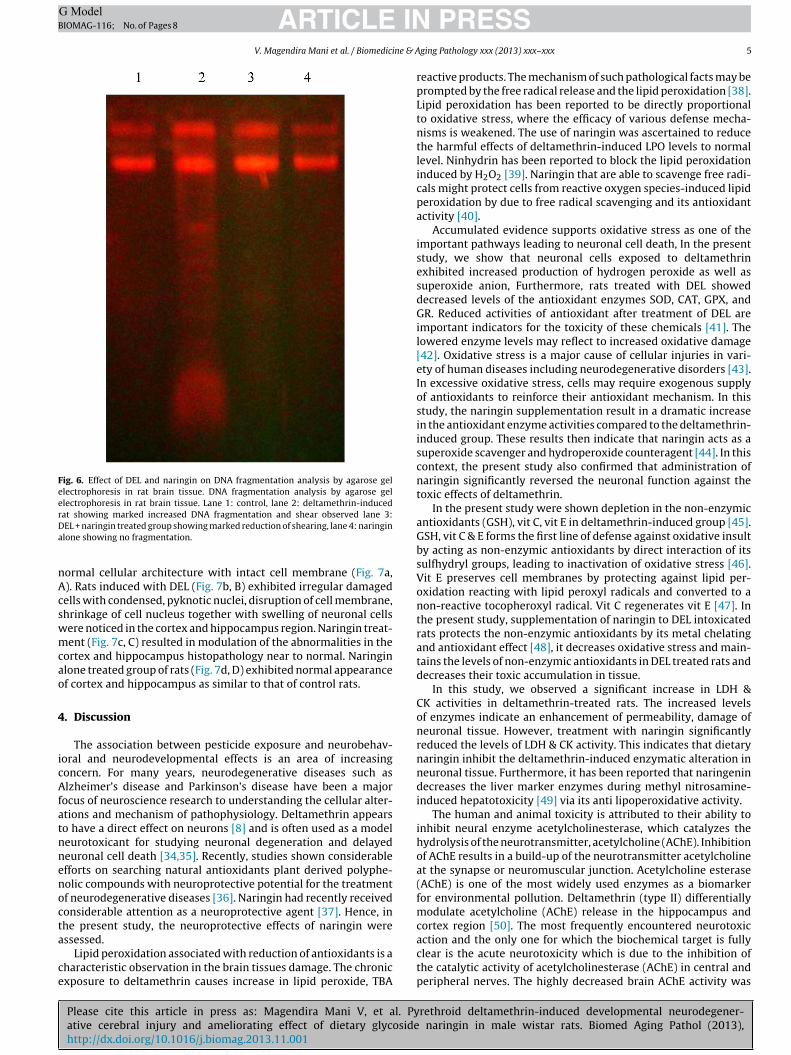
Fig. 6 shows the effect of DEL and naringin on DNA fragmen-ation by agarose gel electrophoresis of control and experimental
ig. 5. Effect of DEL and naringin on the native gel electrophoresis (SOD & CAT) patterntaining of SOD & CAT of tissue homogenate of control and experimental group of rats. Tompared to control (lane 1) and naringin treated rats (lane 4). However, treatment with nnd CAT in group III.
animals in each group. Statistical significant variations are compared as fol-lows: deltamethrin-induced vs. control; deltamethrin-induced + naringin treated vs.deltamethrin-induced; * indicates P < 0.05 and NS indicates non-significant.
group of rats. Rats exhibited marked DNA fragmentation in theDEL-induced groups as observed by increase of smear in the DEL-induced rats (lane 2). The treatment with naringin significantlyprevented DEL-induced DNA fragmentation (lane 3). Naringinalone treated group shows no smear and fragmentation (lane 4).
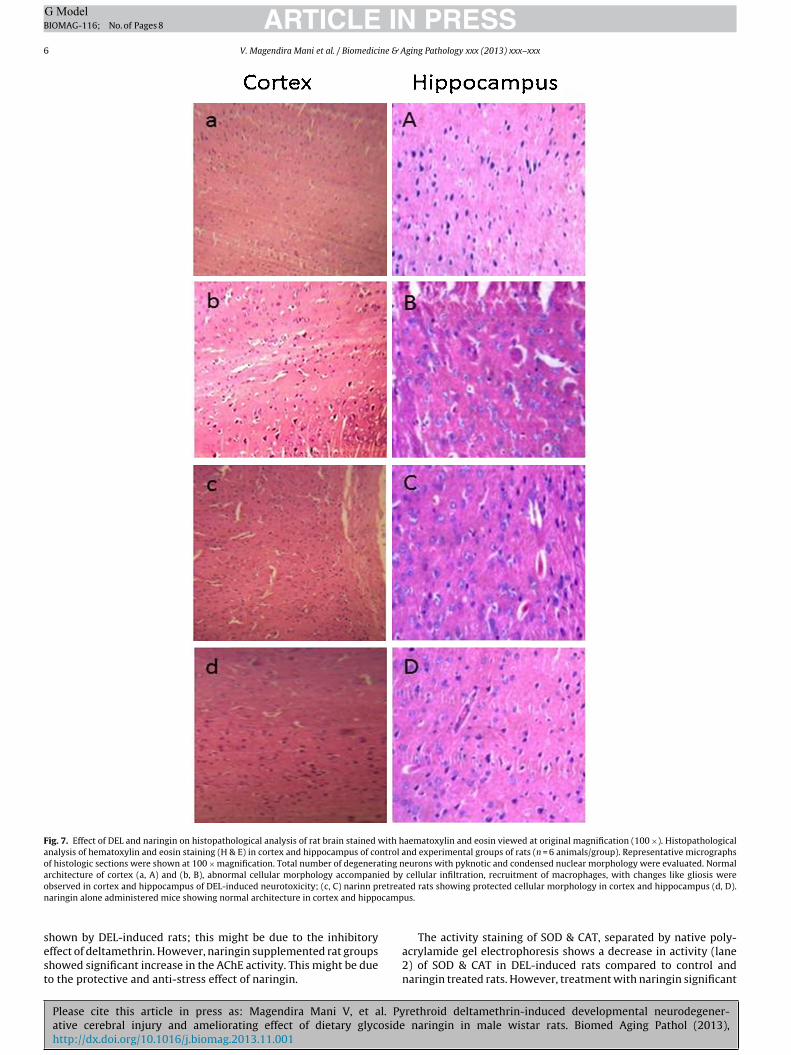
3.7. Effect of DEL & naringin on histological examination of ratbrain
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
Histological changes of rat brain tissue of control and exper-imental groups of animals are depicted in Fig. 7. The regionsof cortex and hippocampus of control rat brain sections showed
of the brain tissue homogenate of control and experimental groups. The activityhis result shows a decrease in activity (lane 2) of SOD & CAT in DEL-induced ratsaringin significant increase in the activity (lane 3) of the antioxidant enzymes SOD
ARTICLE ING ModelBIOMAG-116; No. of Pages 8
V. Magendira Mani et al. / Biomedicine & A
Fig. 6. Effect of DEL and naringin on DNA fragmentation analysis by agarose gelelectrophoresis in rat brain tissue. DNA fragmentation analysis by agarose gelelectrophoresis in rat brain tissue. Lane 1: control, lane 2: deltamethrin-inducedrDa
nAcswmcao
4
icAfatnnenocta
ce
at showing marked increased DNA fragmentation and shear observed lane 3:EL + naringin treated group showing marked reduction of shearing, lane 4: naringinlone showing no fragmentation.
ormal cellular architecture with intact cell membrane (Fig. 7a,). Rats induced with DEL (Fig. 7b, B) exhibited irregular damagedells with condensed, pyknotic nuclei, disruption of cell membrane,hrinkage of cell nucleus together with swelling of neuronal cellsere noticed in the cortex and hippocampus region. Naringin treat-ent (Fig. 7c, C) resulted in modulation of the abnormalities in the
ortex and hippocampus histopathology near to normal. Naringinlone treated group of rats (Fig. 7d, D) exhibited normal appearancef cortex and hippocampus as similar to that of control rats.
. Discussion
The association between pesticide exposure and neurobehav-oral and neurodevelopmental effects is an area of increasingoncern. For many years, neurodegenerative diseases such aslzheimer’s disease and Parkinson’s disease have been a major
ocus of neuroscience research to understanding the cellular alter-tions and mechanism of pathophysiology. Deltamethrin appearso have a direct effect on neurons [8] and is often used as a modeleurotoxicant for studying neuronal degeneration and delayedeuronal cell death [34,35]. Recently, studies shown considerablefforts on searching natural antioxidants plant derived polyphe-olic compounds with neuroprotective potential for the treatmentf neurodegenerative diseases [36]. Naringin had recently receivedonsiderable attention as a neuroprotective agent [37]. Hence, inhe present study, the neuroprotective effects of naringin were
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
ssessed.Lipid peroxidation associated with reduction of antioxidants is a
haracteristic observation in the brain tissues damage. The chronicxposure to deltamethrin causes increase in lipid peroxide, TBA
PRESSging Pathology xxx (2013) xxx–xxx 5
reactive products. The mechanism of such pathological facts may beprompted by the free radical release and the lipid peroxidation [38].Lipid peroxidation has been reported to be directly proportionalto oxidative stress, where the efficacy of various defense mecha-nisms is weakened. The use of naringin was ascertained to reducethe harmful effects of deltamethrin-induced LPO levels to normallevel. Ninhydrin has been reported to block the lipid peroxidationinduced by H2O2 [39]. Naringin that are able to scavenge free radi-cals might protect cells from reactive oxygen species-induced lipidperoxidation by due to free radical scavenging and its antioxidantactivity [40].
Accumulated evidence supports oxidative stress as one of theimportant pathways leading to neuronal cell death, In the presentstudy, we show that neuronal cells exposed to deltamethrinexhibited increased production of hydrogen peroxide as well assuperoxide anion, Furthermore, rats treated with DEL showeddecreased levels of the antioxidant enzymes SOD, CAT, GPX, andGR. Reduced activities of antioxidant after treatment of DEL areimportant indicators for the toxicity of these chemicals [41]. Thelowered enzyme levels may reflect to increased oxidative damage[42]. Oxidative stress is a major cause of cellular injuries in vari-ety of human diseases including neurodegenerative disorders [43].In excessive oxidative stress, cells may require exogenous supplyof antioxidants to reinforce their antioxidant mechanism. In thisstudy, the naringin supplementation result in a dramatic increasein the antioxidant enzyme activities compared to the deltamethrin-induced group. These results then indicate that naringin acts as asuperoxide scavenger and hydroperoxide counteragent [44]. In thiscontext, the present study also confirmed that administration ofnaringin significantly reversed the neuronal function against thetoxic effects of deltamethrin.
In the present study were shown depletion in the non-enzymicantioxidants (GSH), vit C, vit E in deltamethrin-induced group [45].GSH, vit C & E forms the first line of defense against oxidative insultby acting as non-enzymic antioxidants by direct interaction of itssulfhydryl groups, leading to inactivation of oxidative stress [46].Vit E preserves cell membranes by protecting against lipid per-oxidation reacting with lipid peroxyl radicals and converted to anon-reactive tocopheroxyl radical. Vit C regenerates vit E [47]. Inthe present study, supplementation of naringin to DEL intoxicatedrats protects the non-enzymic antioxidants by its metal chelatingand antioxidant effect [48], it decreases oxidative stress and main-tains the levels of non-enzymic antioxidants in DEL treated rats anddecreases their toxic accumulation in tissue.
In this study, we observed a significant increase in LDH &CK activities in deltamethrin-treated rats. The increased levelsof enzymes indicate an enhancement of permeability, damage ofneuronal tissue. However, treatment with naringin significantlyreduced the levels of LDH & CK activity. This indicates that dietarynaringin inhibit the deltamethrin-induced enzymatic alteration inneuronal tissue. Furthermore, it has been reported that naringenindecreases the liver marker enzymes during methyl nitrosamine-induced hepatotoxicity [49] via its anti lipoperoxidative activity.
The human and animal toxicity is attributed to their ability toinhibit neural enzyme acetylcholinesterase, which catalyzes thehydrolysis of the neurotransmitter, acetylcholine (AChE). Inhibitionof AChE results in a build-up of the neurotransmitter acetylcholineat the synapse or neuromuscular junction. Acetylcholine esterase(AChE) is one of the most widely used enzymes as a biomarkerfor environmental pollution. Deltamethrin (type II) differentiallymodulate acetylcholine (AChE) release in the hippocampus andcortex region [50]. The most frequently encountered neurotoxic
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
action and the only one for which the biochemical target is fullyclear is the acute neurotoxicity which is due to the inhibition ofthe catalytic activity of acetylcholinesterase (AChE) in central andperipheral nerves. The highly decreased brain AChE activity was
ARTICLE IN PRESSG ModelBIOMAG-116; No. of Pages 8
6 V. Magendira Mani et al. / Biomedicine & Aging Pathology xxx (2013) xxx–xxx
Fig. 7. Effect of DEL and naringin on histopathological analysis of rat brain stained with haematoxylin and eosin viewed at original magnification (100 ×). Histopathologicalanalysis of hematoxylin and eosin staining (H & E) in cortex and hippocampus of control and experimental groups of rats (n = 6 animals/group). Representative micrographsof histologic sections were shown at 100 × magnification. Total number of degenerating neurons with pyknotic and condensed nuclear morphology were evaluated. Normalarchitecture of cortex (a, A) and (b, B), abnormal cellular morphology accompanied by cellular infiltration, recruitment of macrophages, with changes like gliosis wereo retrean camp
sest
bserved in cortex and hippocampus of DEL-induced neurotoxicity; (c, C) narinn paringin alone administered mice showing normal architecture in cortex and hippo
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
hown by DEL-induced rats; this might be due to the inhibitoryffect of deltamethrin. However, naringin supplemented rat groupshowed significant increase in the AChE activity. This might be dueo the protective and anti-stress effect of naringin.
ted rats showing protected cellular morphology in cortex and hippocampus (d, D).us.
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
The activity staining of SOD & CAT, separated by native poly-acrylamide gel electrophoresis shows a decrease in activity (lane2) of SOD & CAT in DEL-induced rats compared to control andnaringin treated rats. However, treatment with naringin significant

ING ModelB
ne & A
i(foombwtfDp
tpncbNi
5
weaaeOdtmio
D
c
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLEIOMAG-116; No. of Pages 8
V. Magendira Mani et al. / Biomedici
ncreases in the activity of the antioxidant enzymes SOD and CATlane 3). It might flavonoids naringin inhibit lipid peroxidation andree radical formation by acting as free radical scavengers of super-xide anions and hydroxyl radicals [41]. Electrophoretic analysisf DNA extracted from whole brain tissue (Fig. 6) of rats exhibitedarked DNA fragmentation in the DEL-induced groups as observed
y increase of smear in the DEL-induced rats (lane 2). The treatmentith naringin significantly prevented DEL-induced DNA fragmen-
ation (lane 3). Naringin alone treated group shows no smear andragmentation (lane 4). The result indicates that naringin preventsNA integrity that was challenged due to DEL induction therebyrotecting the brain tissues against toxic substances.
The histological features of DEL-induced rat as observed inhis study exhibited irregular damaged, cells with condensed andyknotic nuclei, disruption of cell membrane, shrinkage of cellucleus together with swelling of neuronal cells were noticed in theortex and hippocampus region. These histological changes coulde attributed to increased oxidative stress elicited by deltamethrin.aringin ameliorated the histopathological alterations in DEL-
nduced rats due to its neuroprotective effects.
. Conclusion
In conclusion, the oral administration of deltamethrin to ratsas found to significantly increase the TBARS, LDH, CK lev-
ls, but significantly decreased the enzymic and non-enzymicntioxidants enzyme activities, AChE, the treatment with naringinfter deltamethrin administration prevented abnormalities innzyme levels when compared to deltamethrin-induced group.ur results demonstrate that treatment with naringin, shortly aftereltamethrin treatment leads to a significant revival of the oxida-ive status, which confirms the protective effect of naringin. Further
olecular studies should be focused to find apoptotic pathwaynvolved in deltamethrin neurotoxicity and neuroprotective effectf naringin.
isclosure of interest
The authors declare that they have no conflicts of interest con-erning this article.
eferences
[1] Gilden RC, Huffling K, Sattler B. Pesticides and health risks. JOGNN2010;39:103–10.
[2] Eddleston M, Buckley NA, Checketts H, Senarathna L, Mohamed F, Sheriff MH,et al. Speed of initial atropinisation in significant organophosphorus pesticidepoisoning a systematic comparison of recommended regimens. J Toxicol ClinToxicol 2004;42(6):865–75.
[3] World Health Organization. Public health impact of pesticides used in agricul-ture. Geneva: WHO; 2010.
[4] Karalliedde L, Eddleston M, Murray V. The global picture of organophosphateinsecticide poisoning. Organophosphates Health 2001:431–71.
[5] Vijayakumar L, Shujaath Ali Z, Kesavan K, Umamaheswari C, Devaraj P.Intervention for suicide attempters: a randomized controlled study. Indian JPsychiatry 2011;53(3):244.
[6] Gunnell D, Eddleston M, Phillips MR, Konradsen F. The global distribu-tion of fatal pesticide self-poisoning: systematic review. BMC Public Health2007;7:357.
[7] Morgan MK, Sheldon LS, Croghan CW, Jones PA, Chuang JC. An observationalstudy of 127 perschool children at their homes and daycare centers in Ohio:environmental pathways to cis- and trans-permethrin exposure. Environ Res2007;104(2):266–74.
[8] Soderlund DM, Clark JM, Sheet LP, Mullin LS, Piccirillo VJ, Sargent D, et al.Mechanisms of pyrethroid neurotoxicity: implications for cumulative riskassessment. Toxicology 2002;171:3–59.
[9] Grandjean P, Landrigan PJ. Developmental neurotoxicity of industrial chemi-
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
cals. Lancet 2006;16:2167–78.10] Jagetia GC, Reddy TK. Modulation of radiation induced alteration in the antiox-
idant status of mice by naringin. Life Sci 2005;77:780–94.11] Singh SV, Jain RK, Gupta AK, Dhatt AS. Debittering of citrus juices- a-review. J
Food Sci Technol 2003;40(3):247–53.
[
PRESSging Pathology xxx (2013) xxx–xxx 7
12] Nijveldt RJ, van Nood E, van Hoorn DEC, Boelens PG, van Norren K, van LeeuwenPAM. Flavonoids: a review of probable mechanisms of action and potentialapplications. Clin Nutr 2001;74:418–25.
13] Wang Y, Chang CF, Chou J, Chen H, Deng X, Harvey BK, et al. Dietary supplemen-tation with blueberries, spinach, or spirulina reduces ischemic brain damage.Exp Neurol 2005;193:75–84.
14] Bok SH, Shin YW, Bae KH, Jeong TS, Kwon YK, Park YB, et al. Effects of naringinand lovastatin on plasma and hepatic lipids in high-fat and highcholesterol fedrats. Nutr Res 2000;20:1007–15.
15] Jagetia GC, Reddy TK. The grapefruit flavanone naringin protects against theradiation-induced genomic instability in the mice bone marrow: a micro-nucleus study. Mutat Res 2002;519:37–48.
16] Kim DH, Jung EA, Song IS, Han JA, Kim TH, Han MJ. Intestinal bacterialmetabolism of flavonoids and its relation to some biological activities. ArchPharmacol Res 1998;21:17–23.
17] Ribeiro IA, Rocha J, Sepodes B, Mota-Filipe H, Ribeiro MH. Effect of naringinenzymatic hydrolysis towards naringenin on the anti-inflammatory activity ofboth compounds. J Molecular Catalysis B: Enzymatic 2008;52-53:13–8.
18] Tripoli E, Guardia M, Giammanco S, Majo D, Giammanco M. Citrus flavonoids:molecular structure, biological activity and nutritional properties: a review.Food Chem 2007;104:466–79.
19] So FV, Guthrie N, Chambers AF, Moussa M, Carroll KK. Inhibition of humanbreast cancer cell proliferation and delay of mammary tumorigenesis byflavonoids and citrus juices. Nutr Cancer 1996;26:167–81.
20] Misra HP, Fridovich I. The role of superoxide anion in the auto oxidationof epinephrine and a simple assay for superoxide dismutase. J Biol Chem1972;247:3170–5.
21] Takahara S, Hamilton BH, Neell JV, Kobra TY, Ogura Y, Nishimura ET. Hypocata-lasemia: a new genetic carrier state. J Clin Invest 1960;29:10–619.
22] Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, Hoekstra WG.Selenium: biochemical role as a component of glutathione peroxidase. Science1973;79:588–90.
23] Staal GE, Visser J, Veeger C. Purification and properties of glutathione reductaseof human erythrocytes. Biochim Biophys Acta 1969;185:39–48.
24] Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys 1959;82:70–7.
25] Omaye ST, Tumball JD, Sauberlich HE. Selected methods for the determina-tion of ascorbic acid in animal cells, tissues and fluids. Meth Enzymol 1979;2:1–11.
26] Desai I. Vitamin E analysis methods for animal tissues. Meth Enzymol1984;105:138–47.
27] King J. The dehydrogenases or oxidoreductases–lactate dehydrogenase. In: VanD, editor. Practical clinical enzymology. London: Nostrand; 1965. p. 83–93.
28] Hall N, Deluca M. Electrophoretic separation and quantitation of creatine kinaseisozymes. Anal Biochem 1976;76:561–7.
29] Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxidation in animal tissues bythiobarbituric acid reaction. Anal Biochem 1979;195:351–8.
30] Ellman GL, Courtney KD, Andres JV, Featherstone RM. A new and rapid col-orimetric determination of acetylcholinesterase activity. Biochem Pharmacol1961;7:88–95.
31] Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. Protein measurement withFolin–Phenol reagent. J Biol Chem 1951;193:265–75.
32] Beauchamp CO, Fridovich I. Superoxide dismutase: improved assays and anassay applicable to acrylamide gels. Anal Biochem 1971;44:276–87.
33] Sun Y, Elwell JH, Oberley LW. A simultaneous visualization of the antioxidantenzymes glutathione peroxidase and catalase on polyacrylamide gels. FreeRadic Res Commun 1988;5:67–75.
34] Aziz MH, Agrawal AK, Adhami VM, Shukla Y, Seth PK. Neurodevelopmentalconsequences of gestational exposure (GD14-GD20) to low dose deltamethrinin rats. Neurosci Lett 2001;300:161–5.
35] Ray DE, Fry JR. A reassessment of the neurotoxicity of pyrethroid insecticides.Pharmacol Ther 2006;111:174–93.
36] Kelsey NA, Wilkins HM, Linseman DA. Nutraceutical antioxidants as novel neu-roprotective agents. Molecules 2010;15:7792–814.
37] Kim HJ, Song JY, Park HJ, Park HK, Yun DH, Chung JH. Naringin protects againstrotenone-induced apoptosis in human neuroblastoma SH-SY5Y cells. Korean JPhysiol Pharmacol 2009;13:281–5.
38] Dinu D, Marinescu D, Munteanu MC, Staicu AC, Costache M, Dinischiotu A.Modulatory effects of deltamethrin on antioxidant defense mechanisms andlipidperoxidation in Carassius auratus gibelio liver and intestine. Arch EnvironContam Toxicol 2010;58:757–64.
39] Kanno S, Shouji A, Asou K, Ishikawa M. Effects of naringin on hydrogenperoxide-induced cytotoxicity and apoptosis in p388 cells. J Pharmacol Sci2003;92:166–70.
40] Kanno SI, Shouji A, Asou K, Ishikawa M. Effects of naringin on hydrogenperoxide-induced cytotoxicity and apoptosis in P388 cells. J Pharmacol Sci2003;92:166–70.
41] Jayakumar T, Sakthivel M, Thomas PA, Geraldine P. Pleurotus ostreatus, an oys-ter mushroom, decreases the oxidative stress induced by carbon tetrachloridein rat kidneys, heart and brain. Chem Biol Interact 2008;176:108–20.
42] Srinivasan M, Rukkumani R, Ram Sudheer A, Menon VP. Ferulic acid, a natural
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
protector against carbon tetrachloride-induced toxicity. Fundam Clin Pharma-col 2005;19:491–6.
43] Keating DJ. Mitochondrial dysfunction, oxidative stress, regulation of exo-cytosis and their relevance to neurodegenerative diseases. J Neurochem2008;104:298–305.

ING ModelB
8 ne & A
[
[
[
[
[
ARTICLEIOMAG-116; No. of Pages 8
V. Magendira Mani et al. / Biomedici
44] Jeon SM, Bok SH, Jang MK, Kim YH, Nam KT, Jeong TS, et al. Comparison ofantioxidants effect of naringin and probucol in cholesterol-fed rabbits. ClinChim Acta 2002;317:181–90.
45] Tos-Luty S, Haratym-Maj A, Latuszynska J, Obuchowska-Przebirowska D,Tokarska-Rodak M. Oral toxicity of deltamethrin and fenvalerate in Swiss mice.
Please cite this article in press as: Magendira Mani V, et al. Pyative cerebral injury and ameliorating effect of dietary glycosidehttp://dx.doi.org/10.1016/j.biomag.2013.11.001
Ann Agric Environ Med 2001;8:245–54.46] Rahman SH, Ibrahim K, Larvin M, Kingsnorth A, McMahon MJ. Asso-
ciation of antioxidant enzyme gene polymorphisms and glutathionestatus with severe acute pancreatitis. Gastroenterology 2004;126:1312–22.
[
[
PRESSging Pathology xxx (2013) xxx–xxx
47] Clarke MW, Burnett JR, Croft KD. Vitamin E in human health and disease. CritRev Clin Lab Sci 2008;45:417–50.
48] Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, Telser J. Free radicalsand antioxidants in normal physiological functions and human disease. Int JBiochem Cell Biol 2007;39:44–84.
rethroid deltamethrin-induced developmental neurodegener- naringin in male wistar rats. Biomed Aging Pathol (2013),
49] Lee MH, Soon S, Moon JO. The flavonoid naringen ininhibits dimethylnitrosamine induced liver damage in rats. Biol Pharm Bull 2004;27:72–6.
50] Hossain MM, Suzuki T, Sato I, Takewaki T, Suzuki K, Kobayashi H. The modula-tory eVect of pyrethroids on acetylcholine release in the hippocampus of freelymoving rats. Neurotoxicology 2004;25:825–33.
Copyright © 2022 FDOKUMEN