
Polymorphisms in cg2 and pfcrt genes and resistance to chloroquine and other antimalarials in vitro...
7
TRANSACTIONS OF THE ROYAL SOCIETYOF TROPICALMEDICINE AND HYGIENE (2003) 97, 318-324 Polymorphisms in co2 and pfcrt genes and resistance to chloroquine and other antimalarials in vitro in Plasmodium falciparum isolates from Colombia Iveth J. Gonzfilez 1, Rub6n E. Varela 1, Claribel Murillo 1, Beatriz E. Ferro 1, Jenny Salas I, Luis E. Giraldo ~*, Mariano G. Zalis 2 and Nancy G. Saravia 1 ZCentro Internacional de Entrenamiento e Investigaciones NlJdieas, Cali, Colombia; 2 Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil Abstract Polymorphisms in Plasmodium falciparum cg2 and pfcrt genes and their association with chloroquine resistance in vitro in Colombian parasites were evaluated in this study. Association of chloroquine resistance with resistance to other antimalarial drugs in vitro was also examined. Polymerase chain reactions (PCR) for tc and o2 cg2 regions and nested PCR and digestion with ApoI enzyme for K-76T pfcrt point mutation defined corresponding polymorphisms in 83 samples collected between 1995 and 1999. The isotopic microtest was used to evaluate sensitivity in vitro in a subgroup of 18 isolates. The predominant cg2 pattern observed was 13~/14c0 repeats (46/83 [55.4%]) and all samples presented the K-76T mutant allele. Seventy-eight percent of samples were resistant to chloroquine in vitro, 35.3% to amodiaquine, 16.7% to mefloquine, and 5.6% to quinine. Significant correlations (P< 0.05) were observed between the ICs0s of chloroquine and arteether, and among ICs0s of arteether, mefloquine, and quinine. These results suggest the development of multiple and cross-resistance of Colombian P. falciparum isolates to second- and third-line antimalarials and new alternative drugs. Keywords: malaria, Plasmodiumfalciparum, drug resistance, chloroquine, cg2, pfcrt, Colombia Introduction Resistance of Plasmodium falciparum to chloroquine (CO.) has been one of the principal contributing factors to increased morbidity and mortality due to malaria worldwide (Barat & Bloland, 1997; Marsh, 1998). Dif- ferent approaches have been developed to determine the extent of CQ resistance and to establish the bio- logical mechanisms whereby the parasite has been able to evade the action of the drug. The evaluation of CQ efficacy in the treatment of uncomplicated P. falciparum malaria has allowed the determination of thresholds of treatment failure that are critical to the adjustment of antimalarial drug policies (WHO, 1990). However, the risk of loss of patients during long follow-up periods and the logistical demands of clinical response studies in endemic areas have motivated the development of laboratory strategies for the identification of CQ resis- tance (WHO, 1994). The latter include drug sensitivity tests in vitro and the evaluation of molecular markers associated with drug resistance (Bickii et al., 1998; Warhurst, 2001). Tests of antimalarial drug sensitivity in vitro provide information on the frequency of the resistant phenotype among the populations of parasites being transmitted and, at the same time, the possible cross-resistance patterns of antimalarial drugs (Warsame et al., 1991). Testing in vitro is also useful in the evaluation of the efficacy of new drugs and in the investigation of bio- logical mechanisms of drug action and resistance (Des- jardins et al., 1979; Foote & Cowman, 1994). Despite the usefulness of the results of testing in vitro, the application of these methods is limited because of the requirement that samples be immediately cultured or cryopreserved for transport, the laborious adaptation of parasites to culture, the use of expensive and radio- active materials, and the evaluation of the parasite growth by cumbersome methods such as microscopy (Desjardins et al., 1979; Bruce-Chwatt et al., 1981). Due to the limitations of in vivo and in vitro meth- ods, genetic markers of resistance have been sought, and several polymorphisms have been associated with CQ resistance. The establishment of molecular markers strongly associated with resistance, would allow not only the definition of the rate of resistance in endemic *Deceased. Address for correspondence: Iveth J. Gonzfilez, Centro Intemacional de Entrenamiento e Invesfigaciones M~dicas (CIDEIM), Avenida 1N # 3-03, Cali, Colombia; phone +57 2 6682164, fax +57 2 6672989, e-mail [email protected]. areas with easily obtained and transported samples, but also the identification of putative molecular mechan- isms associated with or determining the resistant phe- notype (Cowman, 2001). During the last 2 decades, CQ resistance has been associated with polymorphisms in pfmdrl, cg2, and the more recently described pfcrt genes of P. falciparum (Foote et al., 1989; Suet al., 1997; Fidock et al., 2000a). The presence of these polymorphisms in sensitive as well as resistant para- sites, has ruled out a direct causal role in CQ resistance (Wellems et al., 1990; Fidock et al., 2000b; Chen et al., 2002). However, studies conducted in Africa and Asia concomitantly evaluating the 3 genes (Adagu & War- hurst, 2001; Babiker et al., 2001; Pillai et al., 2001), have suggested the possible association of more than 1 polymorphism in more than 1 gene as a determinant of the CQ-resistant phenotype. Chloroquine resistance was first described in Colombia and Thailand in about 1960 (Moore & Lanier, 1961; Payne, 1987). However, inadmissible levels of CQ resistance requiring changes in antimalarial drug policies in several countries with endemic P. falciparum malaria were not reported until the 1990s (Wernsdorfer, 1994). This delay in the spread of CQ resistance despite its use for more than 5 decades may reflect the complexity of the biological mechanisms of resistance requiring multiple genetic changes for its development (Warhurst, 2001; Wellems & Plowe, 2001). In 2000, when several studies in Colombia demon- strated CQ resistance in vivo of above 40% (Osorio et al., 1999; Castillo et al., 2002), CQ was replaced by amodiaquine (AQ) for the treatment of uncomplicated P. faleiparum malaria in all endemic areas of the country (Ministerio de Salud de Colombia, 2000). Notwith- standing the high levels of CQ resistance in Colombia and other South American countries, there is little information about genetic markers associated with CQ resistance in P. falciparum in this region (Mehlotra et al., 2001; Vieira et al., 2001; Cortese et al., 2002). This study determined the presence of polymorphisms in P. falciparum cg2 and pfcrt genes and their association with CQ resistance in vitro in Colombian parasites. In addi- tion, association of CQ resistance with resistance to other antimalarial drugs in vitro in Colombian isolates was examined. Materials and Methods Study sites Samples were collected in referral centres from dif- ferent endemic areas of Colombia (Fig. 1). Samples
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Polymorphisms in cg2 and pfcrt genes and resistance to chloroquine and other antimalarials in vitro...

TRANSACTIONS OF THE ROYAL SOCIETY OF TROPICAL MEDICINE AND HYGIENE (2003) 97, 318-324
Polymorphisms in co2 and pfcrt genes and resistance to chloroquine and other antimalarials in vitro in Plasmodium falciparum isolates from Colombia
I v e t h J. G o n z f i l e z 1 , R u b 6 n E . V a r e l a 1 , C l a r i b e l M u r i l l o 1 , B e a t r i z E . F e r r o 1, J e n n y S a l a s I, L u i s E . G i r a l d o ~*, M a r i a n o G. Zal is 2 a n d N a n c y G. S a r a v i a 1 ZCentro Internacional de Entrenamiento e Investigaciones NlJdieas, Cali, Colombia; 2 Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
A b s t r a c t Polymorphisms in Plasmodium falciparum cg2 and pfcrt genes and their association with chloroquine resistance in vitro in Colombian parasites were evaluated in this study. Association of chloroquine resistance with resistance to other antimalarial drugs in vitro was also examined. Polymerase chain reactions (PCR) for tc and o2 cg2 regions and nested PCR and digestion with ApoI enzyme for K-76T pfcrt point mutation defined corresponding polymorphisms in 83 samples collected between 1995 and 1999. The isotopic microtest was used to evaluate sensitivity in vitro in a subgroup of 18 isolates. The predominant cg2 pattern observed was 13~/14c0 repeats (46/83 [55.4%]) and all samples presented the K-76T mutant allele. Seventy-eight percent of samples were resistant to chloroquine in vitro, 35.3% to amodiaquine, 16.7% to mefloquine, and 5.6% to quinine. Significant correlations ( P < 0.05) were observed between the ICs0s of chloroquine and arteether, and among ICs0s of arteether, mefloquine, and quinine. These results suggest the development of multiple and cross-resistance of Colombian P. falciparum isolates to second- and third-line antimalarials and new alternative drugs.
Keywords: malaria, Plasmodiumfalciparum, drug resistance, chloroquine, cg2, pfcrt, Colombia
I n t r o d u c t i o n Resistance of Plasmodium falciparum to chloroquine
(CO.) has been one of the principal contributing factors to increased morbidity and mortality due to malaria worldwide (Barat & Bloland, 1997; Marsh, 1998). Dif- ferent approaches have been developed to determine the extent of CQ resistance and to establish the bio- logical mechanisms whereby the parasite has been able to evade the action of the drug. The evaluation of CQ efficacy in the treatment of uncomplicated P. falciparum malaria has allowed the determination of thresholds of treatment failure that are critical to the adjustment of antimalarial drug policies (WHO, 1990). However, the risk of loss of patients during long follow-up periods and the logistical demands of clinical response studies in endemic areas have motivated the development of laboratory strategies for the identification of CQ resis- tance (WHO, 1994). The latter include drug sensitivity tests in vitro and the evaluation of molecular markers associated with drug resistance (Bickii et al., 1998; Warhurst, 2001).
Tests of antimalarial drug sensitivity in vitro provide information on the frequency of the resistant phenotype among the populations of parasites being transmitted and, at the same time, the possible cross-resistance patterns of antimalarial drugs (Warsame et al., 1991). Testing in vitro is also useful in the evaluation of the efficacy of new drugs and in the investigation of bio- logical mechanisms of drug action and resistance (Des- jardins et al., 1979; Foote & Cowman, 1994). Despite the usefulness of the results of testing in vitro, the application of these methods is limited because of the requirement that samples be immediately cultured or cryopreserved for transport, the laborious adaptation of parasites to culture, the use of expensive and radio- active materials, and the evaluation of the parasite growth by cumbersome methods such as microscopy (Desjardins et al., 1979; Bruce-Chwatt et al., 1981).
Due to the limitations of in vivo and in vitro meth- ods, genetic markers of resistance have been sought, and several polymorphisms have been associated with CQ resistance. The establishment of molecular markers strongly associated with resistance, would allow not only the definition of the rate of resistance in endemic
*Deceased. Address for correspondence: Iveth J. Gonzfilez, Centro Intemacional de Entrenamiento e Invesfigaciones M~dicas (CIDEIM), Avenida 1N # 3-03, Cali, Colombia; phone +57 2 6682164, fax +57 2 6672989, e-mail [email protected].
areas with easily obtained and transported samples, but also the identification of putative molecular mechan- isms associated with or determining the resistant phe- notype (Cowman, 2001). During the last 2 decades, CQ resistance has been associated with polymorphisms in pfmdrl , cg2, and the more recently described pfcrt genes of P. falciparum (Foote et al., 1989; S u e t al., 1997; Fidock et al., 2000a). The presence of these polymorphisms in sensitive as well as resistant para- sites, has ruled out a direct causal role in CQ resistance (Wellems et al., 1990; Fidock et al., 2000b; Chen et al., 2002). However, studies conducted in Africa and Asia concomitantly evaluating the 3 genes (Adagu & War- hurst, 2001; Babiker et al., 2001; Pillai et al., 2001), have suggested the possible association of more than 1 polymorphism in more than 1 gene as a determinant of the CQ-resistant phenotype. Chloroquine resistance was first described in Colombia and Thailand in about 1960 (Moore & Lanier, 1961; Payne, 1987). However, inadmissible levels of CQ resistance requiring changes in antimalarial drug policies in several countries with endemic P. falciparum malaria were not reported until the 1990s (Wernsdorfer, 1994). This delay in the spread of CQ resistance despite its use for more than 5 decades may reflect the complexity of the biological mechanisms of resistance requiring multiple genetic changes for its development (Warhurst, 2001; Wellems & Plowe, 2001).
In 2000, when several studies in Colombia demon- strated CQ resistance in vivo of above 40% (Osorio et al., 1999; Castillo et al., 2002), CQ was replaced by amodiaquine (AQ) for the treatment of uncomplicated P. faleiparum malaria in all endemic areas of the country (Ministerio de Salud de Colombia, 2000). Notwith- standing the high levels of CQ resistance in Colombia and other South American countries, there is little information about genetic markers associated with CQ resistance in P. falciparum in this region (Mehlotra et al., 2001; Vieira et al., 2001; Cortese et al., 2002). This study determined the presence of polymorphisms in P. falciparum cg2 and pfcrt genes and their association with CQ resistance in vitro in Colombian parasites. In addi- tion, association of CQ resistance with resistance to other antimalarial drugs in vitro in Colombian isolates was examined.
M a t e r i a l s a n d M e t h o d s Study sites
Samples were collected in referral centres from dif- ferent endemic areas of Colombia (Fig. 1). Samples

GENETIC MARKERS AND ANTIMALARIAL DRUG RESISTANCE IN COLOMBIA 319
from Buenaventura (Pacific coast) and Florencia (Amazon region) were collected in 1995 during a study on molecular markers in P. falciparum; samples from Quibd6, Tumaco, E1 Chareo (Pacific coast), and Leti- cia (Amazon region) were collected between 1999 and 2001 within the framework of a treatment efficacy assessment of AQ and sulfadoxine-pyrimethamine (SP); and samples from Timbiqui (Pacific coast) and Puerto Asis (Amazon region), were collected in Cali between 1998 and 1999 during a CQ efficacy study in patients who acquired their infection in the south- western region of Colombia (Castillo et al., 2002). All samples were obtained from patients diagnosed with uncomplicated P. falciparum malaria before treatment. Patient permission was substantiated by signed in- formed consent. Study protocols were approved by the Centro Internacional de Entrenemiento e Investiga- ciones M6dicas (CIDEIM) Institutional Review Board, Call, Colombia, for research involving human subjects in accordance with national and international guide- lines for the protection of research participants. La- boratory procedures were performed between 2000 and 2001.
Collection of samples Blood samples for molecular analysis were collected
on filter-paper (Schleicher and Schuell, Keene, NH, USA and Whatman 3MM, Maidstone, UK) and when dried, stored in individual plastic bags at room tem- perature until their analysis. Blood samples for sensitiv- ity tests in vitro were collected in citrate phosphate dextrose anticoagulant. When feasible, samples were transported at room temperature and adapted to cul- ture conditions within 24 h. Some samples were mixed at a 1:1 proportion with a cryopreservation mixture (0.9% NaC1, 4.2% sorbitol, and 28% glycerol) and transported and stored in liquid nitrogen until their analysis.
DNA extraction DNA was extracted using a modified Saponin/Che-
lex-100 method (Giraldo et al., 1998). Briefly, approxi- mately 1 cm 2 of filter-paper containing the dried blood sample was removed and cut into smaller pieces that were incubated in 1 mL 1% saponin (Sigma, St Louis, MO, USA) at 4 °C for 1 h and then washed 3 times with 1 mL phosphate-buffered saline (PBS), pH 7.2. After washing, 200 pL 5% Chelex-100 (Bio-Rad, Her- cules, CA, USA) was added to the samples and incu- bated at 56°C for 15min and then at 100°C for 10 rain. Samples were macerated and centrifuged at 16 000 g for 5 min. Supematants were transferred into 1.5-mL tubes and stored at - 2 0 °C until their use in polymerase chain reactions (PCR).
PCR for cg2 ~: and eo repeat regions Specific PCRs for ~c and co repeat amplification were
performed using 5 oL of the extracted D N A and 50 pL reaction volume with the primer pairs KCG2P1/ KCG2M7 and OCG2M9/OCG2P41, respectively (Table 1). The amplification reaction was performed using a PTC-100 T M thermocycler (MJ Research Inc., Watertown, MA, USA) with the amplification condi- tions detailed in Table 1. Electrophoresis on a 4% agarose gel (Gibco, Grand Island, NY, USA) stained with 0.2 ~g/mL ethidium bromide (Sigma) was used to visualize and compare the sample bands with the bands obtained with reference strains (Table 1).
Nested PCR and restriction digestion for the detection of pfcrt K-76T poim mutation
The detection of the 2 alleles ofpfcrt codon 76 was performed as described previously by Djimd~ et al. (2001). Briefly, the first PCR reaction (PCR1) ampli- fied a 537 bp region around the mutation K-76T using the primers CRTP1 and CRTP2 (Djimd6 et al., 2001). The PCR product was diluted 1:5000 in distilled water
/3
L~
Panamfi
Pacific ~ OceaH
Q u i b d 6 ,
B u e n a v e n t u r a !
T i m b i q u i
E1 C h a r c l ~ / 4 T u m a c o I
Ecuador
Can'bbean Sea f ~ ' ~
~ / " ~ I " - ~ - - / / ~ I~f~ Pacific coast
/ • Amazon region
'l Venezuela
+ BOGOTa
I CALI ~ Q~
b h 4
Brazil
/ J
Peril ~ L c t i c i a
Fig. 1. Sites where Plasmodium falciparum samples were collected for analysis of cg2 and pfcrt gene polymorphisms and determination of sensitivity to antimalarial drugs in vitro, Colombia.

3 2 0 I . J . G o N Z A L E Z E T A L .
$
o
O
e9
~9 tXg .~ . .
0 0
R
n:J ' ~ ,,0
.~ o o o "~, e t m m
~- p p p p ~
~ ~ . ~
m e
S • r.)
O O
P P P P ~
< < WW < b
kc31m
. 2 ~
$
(3
. 9
o
e~
o
o
<
gN-~
and 5 pL of the final dilution was used for the second PCR (PCR2). The PCR2 amplified a 134 bp segment around the mutation using the primers CRTD1 and CRTD2 (Djimdfi et al., 2001). Five microlitres of the amplicon from the PCR2 was digested using 4 U of the ApoI restriction enzyme (New England Biolabs, Bever- ley, MA, USA) with overnight incubation at 50 °C according to the manufacturer 's protocol. The ApoI restriction enzyme cut samples with the wild-type allele into 2 fragments of 34 bp and 100 bp while samples with the mutant allele yielded a single band of 134 bp. Fragment sizes were determined by comparison with the standard ~X174RFDNA/Hse I I fragments (Gibco) and with the reference strains INDO (mutant) and HAITI (wild-type). Samples that presented the mutant genotype by enzyme digestion were analysed by nested PCR to establish the presence or absence of the specific T76 mutant allele. Five microlitres of the 1:5000 diluted PCR1 product was used in a third PCR reac- tion (PCR3) using the primers CRTP3 and CRTP4m (Djimdfi et al., 2001)• Samples with the T76 mutant allele presented a 366 bp band. The absence of amplifi- cation product suggested the presence of a different mutation in the same position.
Sensitivity tests in vitro Samples transported at room temperature were cen-
trifuged at 200 g and 25 °C and washed 3 times with RPMI-1640 (Gibco) to separate infected red blood cells from plasma and white blood cells• Samples stored in liquid nitrogen were thawed according to the method described previously by Cerutti et al. (1999). All sam- ples were suspended in RPMI-1640 supplemented with 10% heat-inactivated non-immune A-positive serum, 25 mM Hepes (Sigma), 25 mM NaHCO3, and 0•86 gg/ mL reduced glutathione (Sigma)• Haematocrit of 5% was adjusted with a 50% suspension of non-infected A- positive red blood cells• Samples were cultured by incubation at 37 °C in 5% 02 and 5% CO2 atmosphere and daily change of culture medium (Cerutti et al., 1999). Sensitivity tests in vitro were performed as described previously (Desjardins et al., 1979) when P. falciparum cultures reached parasitaemias > 1%. Briefly, cultures were diluted to parasitaemias between 0.5 and 1% and haematocrit of 1%. The culture suspensions were distributed in 96-well culture plates pre-dosed with different final concentrations of chloroquine di- phosphate (3.4-2500 ng/mL), AQ (0.34-250 ng/mL), quinine sulfate (6.8-5000 ng/mL), mefloquine (MQ; 0.34-250 ng/mL), and arteether (AR; 0.034-25 ng/ mL). Drugs were kindly provided by the Walter Reed Army Institute of Research, USA. Each isolate was tested in duplicate for all drugs. Wells without a drug were filled with either infected (positive controls) or non-infected (negative control) red blood cells sus- pended at 1% in complete RPMI-1640 medium. Cul- ture plates were incubated at 37 °C in 5% O2 and 5% CO2 atmosphere for 24h, and then pulsed with 0.25 ~Ci/well [3H]Hypoxanthine (Amersham Interna- tional, Buckinghamshire, UK). After a second period of incubation of 24 h, red blood cells were lysed by freezing and thawing culture plates and harvested on a Semiautomatic Cell Hat~gester (Skatron Instruments, Lier, Norway). The [3H]Hypoxanthine incorporation was quantified using a Beckman LS7500 scintillation counter (Beckman Scientific Instruments Division, Irvine, CA, USA). The 50% inhibitory concentration (ICs0), defined as the concentration of drug producing a 50% inhibition of [3H]Hypoxanthine uptake by para- sites in the drug-free control wells, was determined by non-linear regression function using N F I T software (University of Texas, Medical Branch, Galveston, TX, USA). Field strains were considered resistant to any of the drugs if ICs0s were above the following values: CQ > 45.5 ng/mL (~100 nM), AQ > 7 ng/mL (~20 nM), quinine (QN) > 275 ng/mL (~500 nM), and M Q > 10

GENETIC MARKERS AND ANTIMALARIAL DRUG RESISTANCE IN COLOMBIA 321
ng /mL (~24 nM) (Childs et al., 1989; Cerutt i et al., 1999). T o date, no specific IC50 value has been estab- lished to define A R resistance. Reference strains W2 (CQ-resistant) and D6 (CQ-sensit ive and MQ-resis- rant) were tested in each assay to validate reproducibil- ivy.
Statistical analysis Data entry and descriptive analysis were performed
using Epi-Info 6.04 ( C D C , Atlanta, GA, USA) and SPSS 7.5 (SPSS Inc., Chicago, IL, USA). Median ICsos and their ranges were determined for each tested drug. Associations between alleles and resistance in vitro were analysed using Fisher 's exact test. Spearman 's correla- t ion index was used to examine the association between IC50 levels of the evaluated drugs. A two-tailed P-value of < 0.05 was considered significant.
Results Characteristics of patient samples
One sample from each of 83 patients diagnosed with
uncomplicated P. falciparum malaria was collected for molecular analysis. Parasitaemias ranged from 90 to 34 400 asexual forms/pL with a geometr ic mean of 5236 asexual forms/pL. Fifty-nine samples (71.1%) were from different locations along the Pacific coast: 20 from T u m a c o (Narifio), 17 from E1 Charco (Narifio), 12 from Quibd6 (Choc6), 8 from Buenaventura (Valle), and 2 from Timbiqu i (Cauca); and 24 (28.9%) from the Amazon region: 16 from Florencia (Caquet/t), 7 from Leticia (Amazonas), and 1 from Puerto Asis (Pummayo) (Fig. 1).
cg2 and pfcrt gene polymorphisms The predominant cg2 K and ~o repeats observed were
13 (78/83 [94%]) and 14 (51/83 [61.5%]), respectively (Table 2). Twenty-four (28.9%) of the total samples presented a band of less than 14 repeats, and 8 (9.6%) samples presented diverse patterns of repeats involving the presence of more than 1 band in the co region (Fig. 2). Five samples from Leticia, a city located on the border with Brazil, presented a 12v, J14o3 pattern, which
T a b l e 2. cg2 ~: a n d to repeats found in Colombian Plasmodiumfalciparum samples
cg2 Gene
~: Region n (%) co Region n (%)
Origin 12 repeats 13 repeats < 14 repeats 14 repeats Others"
Pacific coast (n = 59) 0 59 (100) 17 (28.8) 35 (59.3) 7 (11.9) Amazon region (n = 24) 5 (20.8) 19 (79.2) 7 (29.2) 16 (66.7) 1 (4.1) Tota l (n = 83) 5 (6) 78 (94) 24 (28.9) 51 (61.5) 8 (9.6)
~Samples with co repeats different from < 14 and 14, and with more than 1 type of repeat in the same sample: 15 repeats (n = 1), < 14 and 16 repeats (n = 1), 14 and 16 repeats (n = 3), 14 and 16 to 19 repeats (n = 1), and 14 and 19 repeats (n = 2).
cg2 gene
Repeats in ~c region Repeats in co region
v
V
03 t--
pfcr~ gene
Digestion with ApoI
¢¢3 t ~ t'q
Fig.. 2. Detection of sequence repeats in K and co regions of the cg2 gene by polymerase chain reaction amplification, and the K-76T point mutation in the pfcrt gene by DNA digestion with ApoI enzyme in Plasmodium falciparum samples from Colombia. Samples P2611, P3034, and P 1276 are from the Pacific coast and sample L2028 is from Leticia in the Amazon region.
322 I.J. GONZALEZ ETAL.
has been described previously for the Brazilian refer- ence strain 7G8 (Su et al., 1997) and for samples from the Brazilian Amazon region (Vieira et al., 2001). Eighty samples were amplified for the pfcrt gene and all of them presented the K-76T point mutation based on enzyme digestion with ApoI (Fig. 2). Two (2.5%) of the 80 amplified samples, 1 from Quibd6 and 1 from Leticia, did not yield a product with the specific primer for the T76 mutation suggesting the presence of a different allele in the same codon.
Drug sensitivity to chloroquine in vitro and association with response to other drugs in vitro
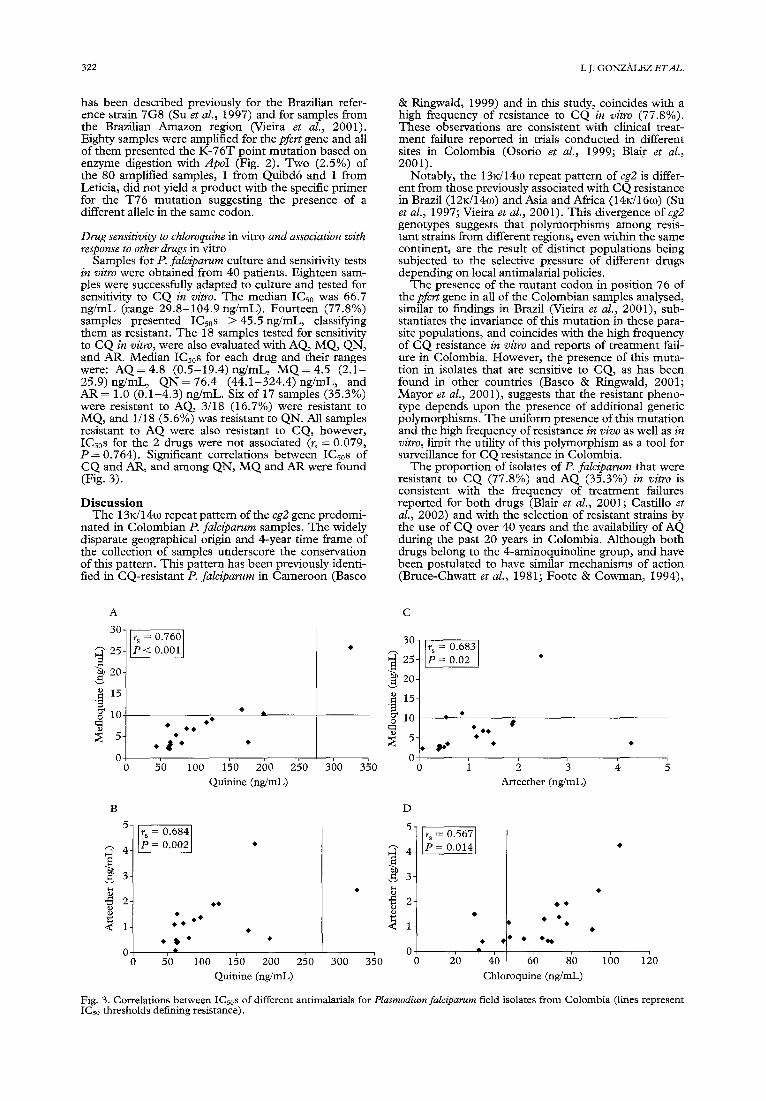
Samples for P. falciparum culture and sensitivity tests in vitro were obtained from 40 patients. Eighteen sam- ples were successfully adapted to culture and tested for sensitivity to CQ in vitro. The median ICs0 was 66.7 ng/mL (range 29.8-104.9 ng/mL). Fourteen (77.8%) samples presented ICs0s > 4 5 . 5 n g / m L , classifying them as resistant. The 18 samples tested for sensitivity to CQ in vitro, were also evaluated with AQ, MQ, QN, and AR. Median ICs0s for each drug and their ranges were: AQ = 4.8 (0.5-19.4) ng/mL, M Q = 4.5 (2.1- 25.9) ng/mL, QN = 76.4 (44.1-324.4) ng/mL, and AR = 1.0 (0.1-4.3) ng/mL. Six of 17 samples (35.3%) were resistant to AQ, 3/18 (16.7%) were resistant to MQ, and 1/18 (5.6%) was resistant to QN. All samples resistant to AQ were also resistant to CQ, however, ICs0s for the 2 drugs were not associated (r~ = 0.079, P = 0.764). Significant correlations between ICsos of CQ and AR, and among QN, M Q and AR were found (Fig. 3).
D i s c u s s i o n The 13n/14~o repeat pattern of the cg2 gene predomi-
nated in Colombian P. falciparum samples. The widely disparate geographical origin and 4-year time frame of the collection of samples underscore the conservation of this pattern. This pattern has been previously identi- fied in CQ-resistant P. falciparum in Cameroon (Basco
& Ringwald, 1999) and in this study, coincides with a high frequency of resistance to CQ in vitro (77.8%). These observations are consistent with clinical treat- ment failure reported in trials conducted in different sites in Colombia (Osorio et al., 1999; Blair et al., 2001).
Notably, the 13~14co repeat pattern of cg2 is differ- ent from those previously associated with CQ resistance in Brazil (12K/14c0) and Asia and Africa (14rJ16c0) (Su et al., 1997; Vieira et al., 2001). This divergence of cg2 genotypes suggests that polymorphisms among resis- tant strains from different regions, even within the same continent, are the result of distinct populations being subjected to the selective pressure of different drugs depending on local antimalarial policies.
The presence of the mutant codon in position 76 of the pfcrt gene in all of the Colombian samples analysed, similar to findings in Brazil (Vieira et al., 2001), sub- stantiates the invariance of this mutation in these para- site populations, and coincides with the high frequency of CQ resistance in vitro and reports of treatment fail- ure in Colombia. However, the presence of this muta- tion in isolates that are sensitive to CQ, as has been found in other countries (Basco & Ringwald, 2001; Mayor et aL, 2001), suggests that the resistant pheno- type depends upon the presence of additional genetic polymorphisms. The uniform presence of this mutation and the high frequency of resistance in vivo as well as in vitro, limit the utility of this polymorphism as a tool for surveillance for CQ resistance in Colombia.
The proportion of isolates of P. falciparum that were resistant to CQ (77.8%) and AQ (35.3%) in vitro is consistent with the frequency of treatment failures reported for both drugs (Blair et al., 2001; Castillo et al., 2002) and with the selection of resistant strains by the use of CQ over 40 years and the availability of AQ during the past 20 years in Colombia. Although both drugs belong to the 4-aminoquinoline group, and have been postulated to have similar mechanisms of action (Bruce-Chwatt et al., 1981; Foote & Cowman, 1994),
A
30-
25-
"~ 20-
.~ 15-
10 O
5-
0 0
~ = 0.760 < 0.001
5'0 160 150 260 Quinine (ng/mL)
2~o 360
C
30-
~ 25-
,~20-
.~ 15-
10
350 0
~ 2 0.683 0.02
Arteether (ng/mL)
B
5-
4 ̧
3
"5 2
~ - 0.684 0.002 *
D
~11 • #
5'0 160 150 260 Quinine (ng/mL)
0.567 0 0 1 4
300 350 0 2'0 0 •
0 250 4'0 6'0 8'0 Chloroquine (ng/mL)
#
160 1½0
Fig. 3. Correlations between ICs0s of different antimalarials for Plasmodiumfalciparum field isolates from Colombia (lines represent ICs0 thresholds defining resistance).

GENETIC MARKERS AND ANTIMALARIAL DRUG RESISTANCE IN COLOMBIA 323
we found no correlation between ICs0s for these 2 drugs in Colombian isolates. Similar results have been previously reported in studies where ICs0s of CQ were more correlated with that of desethylamodiaquine (DEAQ), which despite being less potent than AQ in vitro, is its principal active metabolite (Childs et al., 1989; Ringwald et al., 1998). The absence of a correla- tion between the ICs0s of CQ and AQ together with the low frequency of therapeutic failure of AQ reported in areas of Colombia with high CQ resistance (Blair et al., 2001) indicate that resistance to these 2 drugs is more likely to depend on drug pressure generated by their use in endemic areas than shared mechanisms of action or resistance. The disparity between sensitivity to CQ and AQ in vitro concurs with the demonstrated efficacy of AQ in areas of high CQ resistance and supports the use of AQ as an alternative drug for the management of uncomplicated P. falciparum malaria (Ringwald et al., 1998; Brasseur et al., 1999).
A study conducted in 1985 evaluating sensitivity of Colombian P. falciparum isolates to antimalarial drugs in vitro demonstrated the absence of resistance to QN (Espinal et al., 1985). In the present study, 1/18 (5.6%) isolates was resistant to this drug in vitro. The presence of resistance to QN in the border region of Brazil and Colombia (Zalis et al., 1998; Cerutti et al. 1999) and the finding of a Colombian isolate resistant to QN, draw attention to the need to more fully evaluate the sensitivity of P. falciparum to this drug in Colombia, where its use is restricted to hospitals for the treatment of severe malaria and cases of therapeutic failure with other antimalarials.
The finding ofP. falciparum resistance to M Q in vitro (16.7%) is in agreement with previous studies reporting MQ resistance in 13-19% of isolates from the Amazon region (Espinal et al., 1985; Blair, 1986). Due to the fact that M Q has not been commercially available in Colombia, resistance to this drug is not attributable to the selective pressure of its routine use. Mefloquine resistance in Colombia may be due to the introduction of resistant strains from Venezuela or Brazil where resistance to this drug has been demonstrated (Mock- enhaupt, 1995).
The correlation between sensitivity to QN and MQ in vitro has also been encountered in some studies of P. falciparum from other regions (Kremsner et al., 1989; Cerutti et al., 1999). Resistance to these two drugs has been associated with the presence of poly- morphisms in the pfmdrl gene (Reed et al., 2000), which encodes the Pghl protein that, like its homo- logue in turnout cells, mediates the transport of a great variety of chemical compounds across the cell mem- brane of the parasite. Changes in the gene product as a result of gene polymorphisms could reduce the entry or increase the efflux not only of these antimalarial drugs but also others, including derivatives of artemisinin, resistance to which has already been associated with polymorphisms of pfmdrl (Foote & Cowman, 1994; Reed et al., 2000).
The ICs0s to AR of Colombian field isolates were signifcantly correlated with ICs0s to CQ, MQ, and QN. Cross-resistance between quinolines and deriva- tives of artemisinin has also been demonstrated in previous studies (Ringwald et al., 1999; Noedl et al., 2001); however, the mechanisms have not yet been defined. Since the mechanism of action of the deriva- tives of artemisinin has been previously described and is different from that of quinolines (Ringwald et al., 1999), cross-resistance has also been attributed to shared mechanisms of transport mediated by the pfmdrl gene (Reed et al., 2000). The detection of field isolates resistant to derivatives of artemisinin, and dis- cernment of the mechanisms of resistance to these drugs, constitute priority concerns since emergence of resistance to them would be catastrophic for the treat- ment of multidrug-resistant malaria.
Notwithstanding the importance of detection of re- sistance to first-line antimalarial drugs, the results of this study indicate the need to concurrently investigate the emergence and mechanisms of resistance to sec- ond- and third-line antimalarials and new alternative drugs. Resistance to antimalarial drugs in vitro may be an early indicator of emerging treatment failures. The availability of research tools like bioinformatics, micro- array and proteomics technology, and genetic mapping (Cowman, 2001; Rathod et al., 2002) should permit the identification of the genes determining resistance. Although the investigation of CQ resistance is impor- tant to the understanding of the mechanisms of parasite resistance and the development of control tools, efforts should now be directed at the investigation of resistance to other drugs for the treatment of malaria.
Acknowledgements We are very grateful to the inhabitants and the health work-
ers of the Health Services in the endemic areas for their collab- oration in the conduct of this study and to Carmen Castillo for her technical assistance. We thank Jos6 R. Tovar for conduct- ing the statistical analysis, and Ricardo Palacios and Lyda Osorio for their permanent support and collaboration. This investigation received financial support from the UNDP/ World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR) project ID: 981015; the Pan American Health Organization (PAHO) project ID: ACS-9800127-0; and the Colombian Research Council (COLCIENCIAS) project IDs: 2229-04-165-97 and 2229- 04-1017-98.
This paper is dedicated to the memory of Luis E. Giraldo.
References Adagu, I. S. & Warburst, D. C. (2001). Plasmodiumfalcipar-
urn: linkage disequilibrium between loci in chromosomes 7 and 5 and chloroquine selective pressure in Northern Niger- ia. Parasitology, 123, 219-224.
BaNker, H. A., Pringle, S. J., Abdel-Muhsin, A., Mackinnon, M., Hunt, P. & Walliker, D. (2001). High-level chloroquine resistance in Sudanese isolates of Plasmodium falciparum is associated with mutations in the chloroquine resistance transporter gene pfcrt and the multidrug resistance gene pfmdr l . Journal of Infectious Diseases, 183, 1535-1538.
Barat, L. M. & Bloland, P. B. (1997). Drug resistance among malaria and other parasites. Infectious Disease Clinics of North America, 11, 969-987.
Basco, L. K. & Ringwald, P. (1999). Chloroquine resistance in Plasmodium falciparum and polymorphism of the cg2 gene. Journal of Infectious Diseases, 180, 1979-1986.
Basco, L. K. & Ringwald, P. (2001). Analysis of the keypfert point mutation and in vivo and in vitro response to chlor- oquine in Yaound6, Cameroon. Journal of Infectious Diseases, 183, 1828-1831.
Bickii, J., Basco, L. K. & Ringwald, P. (1998). Assessment of three in vitro tests and an in vivo test for chloroquine resistance in Plasmodium falciparum clinical isolates. Journal of Clinical Microbiology, 36, 243-247.
Blair, S. (1986). Resistencia del P. falciparum a drogas en Colombia. Biomedica, 6, 95-100.
Blair, S., Lacharme, L. L., Carmona, J. & Tob6n, A. (2001). Resistencia de Plasmodium falciparum atres ffirmacos anti- malfiricos en Turbo (Antioquia, Colombia), 1998. Pan American yournal of Public Health, 9, 23-29.
Brasseur, P., Guiguemde, R., Diallo, S., Guiyedi, V., Kombi- la, M., Ringwald, P. & Olliaro, P. (1999). Amodiaqine remains effective for treating uncomplicated malaria in West and Central Africa. Transactions of the Royal Society of Tropical Medicine and Hygiene, 93, 645-650.
Bruce-Chwart, L. J., Black, R. H., Canfield, C. J., Clyde, D. F., Peters, W. & Wemsdorfer, W. H. (1981). Chemo- therapy of malaria. WHO Monograph Series, 27, 211-224.
Castillo, C. M., Osorio, L. E. & Palma, G. I. (2002). Assess- ment of therapeutic response of P. vivax and P. faleiparum to chloroquine in a malaria transmission free area in Colom- bia. Memorias do Instituto Oswaldo Cruz, 97, 559-562.
Cerutti, C. Jr, Marques, C., Alencar, F. E., Durlacher, R. R., Alween, A., Segurado, A. A., Pang, L. W. & Zalis, M. G. (1999). Antimalarial drug susceptibility testing of Plasmo- dium falcipamm in Brazil using a radioisotope method. Memorias do Instimto Oswaldo Cruz, 94, 803-809.
Chert, N., Russell, B., Fowler, E., Peters, J. & Cheng, Q. (2002). Levels of chloroquine resistance in Plasmodium

324 I.J. GONZALEZ ETAL.
falciparum are determined by loci other than pfcrt and pfmdrl. Journal of Infectious Diseases, 185, 405-406.
Childs, G. E., Boudreau, E. F., Milhous, W. K., Wimonwat- tratee, T., Pooyindee, N., Pang, L. & Davidson, D. E. (1989). A comparison of the in vitro activities of amodia- quine and desethylamodiaquine against isolates of Plasmo- dium falciparum. American Journal of Tropical Medicine and Hygiene, 40, 7-11.
Cortese, J. F., Caraballo, A., Contreras, C. E. & Plowe, C. V. (2002). Origin and dissemination of Plasmodium falciparum drug-resistance mutations in South America. Journal of Infectious Diseases, 186, 999-1006.
Cowman~ A. F. (2001). Functional analysis of drug resistance ha Plasmodium falciparum in the post-genomic era. Interna- tional ffournal for Parasitology, 31, 871-878.
Desjardins, R. E., Canfield, C. J., Haynes, D. & Chulay, J. D. (1979). Quantitative assessment of antimalarial activity in vitro by a semiautomated microdilution technique. Antimi- crobial Agents and Chemotherapy, 16, 710-718.
Djimd~, A., Doumbo, O. K., Cortese, J. F., Kayentao, K., Doumbo, S., Diourtr, Y., Dicko, A., Su, X. Z., Nomura, T., Fidock, D. A., Wellems, T. E. & Plowe, C. V. (2001). A molecular marker for chloroquine-resistant falciparum ma- laria. New England Journal of Medicine, 344, 257-263, (http://medschool.umaryland.edu/CVD/plowe.html).
Espinal, C. A., Cortes, G. T., Guerra, P. & Arias, A. (1985). Sensitivity of Plasmodiumfalciparum to antimalarial drugs in Colombia. American Journal of Tropical Medicine and Hy- giene, 34, 675-680.
Fidock, D. A., Nomura, T., Talley, A. K., Cooper, R. A., Dzekunov, S. M., Ferdig, M. T., Ursos, L. M., Sidhu, A. S., Naud~, B., Deitsch, K. W., Su, X. Z., Wootton, J. C. & Wellems, T. E. (2000a). Mutations in the P. falciparum digestive vacuole transmembrane protein PfCRT and evi- dence for their role in chloroquine resistance. Molecular Cell, 6~ 861-871.
Fidock, D. A., Nomura, T., Cooper, R. A., Su, X. Z., Talley, A. K. & Wellems, T. E. (2000b). Allelic modifications of the cg2 and cgl genes do not alter the chloroquine response of drug-resistant Plasmodium falciparum. Molecular and Bio- chemical Parasitology, 110, 1-10.
Foote, S. J. & Cowman, A. F. (1994). The mode of action and the mechanism of resistance to antimalarial drugs. Aeta Tropica, 56, 157-171.
Foote, S. J., Thompson, J. K., Cowman, A. F. & Kemp, D. J. (1989). Amplification of the multidrug resistance gene in some chloroquine-resistant isolates ofP. falciparum. Cell, 57, 921-930.
Giraldo, L. E., Acosta, M. C., Labrada, L. A., Praba, A., Montenegro-James, S., Saravia, N. G. & Krogstad, D. J. (1998). Frequency of the Asn- 108 and Thr- 108 point muta- tions in the dihydrofolate reductase gene in Plasmodium falciparum from southwest Colombia. American Journal of Tropical Medicine and Hygiene, 59, 124-128.
Kremsner, P. G., Zotter, G. M., Feldmeier, H., Graninger, W., Kollaritsch, M., Wiedermann, G., Rocha, R. M. & Wemsdorfer, W. t-I. (1989). In vitro drug sensitivity of Plasmodium falciparum in Acre, Brazil. Bulletin of the World Health Organization, 67, 289-293.
Marsh, K. (1998). Malaria disaster in Africa. Lancet, 352, 924. Mayor, A. G., Grmez-Oliv~, X., Aponte, J. J., Casimiro, S.,
Mabunda, S., Dgedge, M., Barreto, A. & Alonso, P. L. (2001). Prevalence of the K76T mutation in the putative Plasmodium falciparum chloroquine resistance transporter (pfcrt) gene and its relation to chloroquine resistance in Mozambique. Journal of Infectious Diseases, 183, 1413-1416.
Mehlotra, R. K., Fujioka, H., Roepe, P. D., Janneh, O., Ursos, L. M., Jacobs-Lorena, V., McNamara, D. T., Bock- arie, M. J., Kazura, J. W., Kyle, D. E., Fidock, D. A. & Zimmerman, P. A. (2001). Evolution of a unique Plasmo- dium falciparum chloroquine-resistance phenotype in asso- ciation with pfcrt polymorphism in Papua New Guinea and South America. Proceedings of the National Academy of Sciences of the USA, 98, 12689-12694.
Ministerio de Salud de Colombia (2000). Guia de atenci6n mrdica para el diagndstico y tratamiento de la malaria. Resolu- ciSn N ° 0412, cap. 37 (htrp://www.minsalud.gov.co/l'qew Site/MScContact).
Mockenhaupt, F. P. (1995). Mefloquine resistance in Plasmo- dium falciparum. Parasitology Today, 11, 248-253.
Moore, D. V. & Lanier, J. E. (1961). Observations on two Plasmodiumfalciparum infections with an abnormal response to chloroquine. American Journal of Tropical Medicine and Hygiene, 10, 5-9.
Noedl, H., Werusdorfer, W. H., Krudsood, S., Wilairatana, P., Viriyavejakul, P., Kollaritsch, H., Wiedermann, G. & Looareesuwan, S. (2001). In vivo-in vitro model for the assessment of clinically relevant antimalarial cross- resistance. American Journal of Tropical Medicine and Hy- giene, 65, 696-699.
Osorio, L. E., Giraldo, L. E., Grajales, L. F., Arriaga, A. L., Andrade, A. L., Ruebush, T. K. & Barat, L. M. (1999). Assessment of therapeutic response of Plasmodium falcipar- urn to chloroquine and sulfadoxine-pyrimethamine in an area of low malaria transmission in Colombia. American Journal of Tropical Medicine and Hygiene, 61, 968-972.
Payne, D. (1987). Spread of chloroquine resistance in Plasmo- dium falciparum. Parasitology Today, 3, 241-246.
Pillai, D. R., Labb~, A. C., Vanisaveth, V., Hongvangthong, B., Pomphida, S., Inkathone~ S., Zhong, K. & Kain, K. C. (2001). Plasmodiumfalciparum malaria in Laos: chloroquine treatment outcome and predictive value of molecular mar- kers. Journal of Infectious Diseases, 183, 789--795.
Rathod, P. K., Ganesan, K., Hayward, R. E., Bozdech, Z. & DeRisi, J. L. (2002). DNA microarrays for malaria. Trends in Parasitology, 18, 39-45.
Reed, M. B., Saliba, K. J., Caruana, S. R., Kirk, K. & Cow- man, A. F. (2000). Pghl modulates sensitivity and resis- tance to multiple antimalarials in Plasmodium falciparum. Nature, 403, 906-909.
Ringwald, P., Bickii, J. & Basco, L. K. (1998). Amodiaquine as the first-line treatment of malaria in Yaoundr, Camer- oon: presumptive evidence from activity in vitro and cross- resistance patterns. Transactions of the Royal Society of Tropical Medicine and Hygiene, 92, 212-213.
Ringwald, P., Bickii, J. & Basco, L. K. (1999). In vitro activity of dihydroartemisinin against clinical isolates of Plasmodium falciparum in Yaounde, Cameroon. American Journal of Tropical Medicine and Hygiene, 61, 187-192.
Su, X. Z., Kirkman, L. A., Fujioka, H. & Wellems, T. E. (1997). Complex polymorphisms in a ~300 kDa protein are linked to chloroquine-resistant P. falciparum in southeast Asia and Africa. Cell, 91, 593-603.
Vieira, P. P., Alecrim, M. G., Da Silva, L. H., Gonzfilez- Jimrnez, I. J. & Zalis, M. G. (2001). Analysis of the pfCRT K76T mutation in Plasmodium falciparum isolates from the Amazon region of Brazil. Journal of Infectious Diseases, 183, 1832.
Warhurst, D. C. (2001). A molecular marker for chloroquine- resistant falciparum malaria. New England Journal of Medi- cine, 344, 299-302.
Warsame, M., Wernsdorfer, W. H., Payne, D. & Bj6rkman, A. (1991). Susceptibility of Plasmodium falciparum in vitro to chloroquine, mefioquine, quinine and sulfadoxine/pyri- methamine in Somalia: relationships between the responses to the different drugs. Transactions of the Royal Society of Tropical Medicine and Hygiene, 85, 565- 569.
Wellems, T. E. & Plowe, C. V. (2001). Chloroquine-resistant malaria. Journal of Infectious Diseases, 184, 770-776.
Wellems, T. E., Panton, L. J., Gluzman, I. Y., Do Rosario, V. E., Gwadz, R. W., Walker-Johan, A. & Krogstad, D. J. (1990). Chloroquine resistance not linked to mdr-like genes in a Plasmodium falciparum cross. Nature, 345, 253-255.
Wernsdorfer, W. H. (1994). Epidemiology of drug resistance in malaria. Acta Tropica, 56, 143-156.
WHO (1990). Practical Chemotherapy of Malaria. Geneva: World Health Organization, Technical Report Series, No. 805.
WHO (1994). Antimalarial Drug Policies: Data Requirements, Treatment of Uncomplicated Malaria and the Management of Malaria in Pregnancy. Geneva: World Health Organization, WHO/MAL/94.1070.
Zalis, M. G., Pang, L., Silveira, M. S., Milhous, W. K. & Wirth, D. F. (1998). Characterization of Plasmodium falci- parum isolates from the Amazon region of Brazil: evidence for quinine resistance. American Journal of Tropical Medicine and Hygiene, 58, 630-637.
Received 27 June 2002; revised 25 November 2002; accepted for publication 28 November 2002




















