
Comparison and differentiation of potyvirus isolates and ...

lable at ScienceDirect
Crop Protection 66 (2014) 120e126
Contents lists avai
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Sensitivity of Cercospora beticola isolates from Serbia to carbendazimand flutriafol
Dragana Budakov a, Nevena Nagl b, *, Vera Stoj�sin a, Ferenc Bagi a, Dario Danojevi�c b,Oliver T. Neher c, Ksenija Ta�ski-Ajdukovi�c b
a Faculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovica 8, 21000 Novi Sad, Serbiab Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbiac The Amalgamated Sugar Company, 1951 S. Saturn Way, Suite 100, Boise, ID 83709, USA
a r t i c l e i n f o
Article history:Received 11 April 2014Received in revised form15 September 2014Accepted 16 September 2014Available online
Keywords:Cercospora leaf spot (CLS)Fungicide resistanceCarbendazimFlutriafolCAPSFungicide efficacy
* Corresponding author. Tel.: þ381 (0)21 4898327.E-mail addresses: [email protected] (N. Na
(K. Ta�ski-Ajdukovi�c).
http://dx.doi.org/10.1016/j.cropro.2014.09.0100261-2194/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Cercospora beticola, causal agent of Cercospora leaf spot (CLS) of sugar beet, is primarily controlled byfungicides. Benzimidazole and demethylation inhibiting fungicides, including carbendazim and flutriafol,have been widely used in Serbia. Since these fungicide groups have a site-specific mode of action, there isa high risk for developing resistance in target organisms, which is the most important limiting factor inCercospora leaf spot chemical control. A rapid identification of flutriafol and carbendazim resistance canhelp researchers in examining the potential of different fungicide resistance management practices, aswell as in selection of fungicides for use in the areas where resistance has occurred. One hundred single-conidia isolates were collected from 70 representative locations of the sugar beet production region inSerbia. Evaluation of the isolates' sensitivity was based on the reduction of mycelial growth on mediumamended with 1.25 mg mL�1
flutriafol and 5 mg mL�1 carbendazim. Resistance to flutriafol and carben-dazim was detected in 16% and 96% of the tested isolates, respectively. All isolates resistant to flutriafolwere resistant to carbendazim as well, which is the first report of a double resistance to fungicides inC. beticola. Detection of the isolates resistant to flutriafol and carbendazim using Cleaved AmplifiedPolymorphic Sequence (CAPS) markers confirmed the results of the in vitro tests. The efficacy of car-bendazim, flutriafol, azoxystrobin, and tetraconazole at commercially recommended doses was evalu-ated in field trials where sugar beet plants in plots were inoculated with a mixture of isolates eithersensitive and/or resistant to flutriafol and carbendazim. Carbendazim and flutriafol efficacy was very lowin plots inoculated with isolates resistant to these fungicides. Presented results will contribute todevelopment of a pathogen population sensitivity monitoring strategy that could be used for an effectiveCLS management in the region.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Cercospora leaf spot (CLS), caused by Cercospora beticola Sacc., isthe most important foliar disease of sugar beet in warm and humidenvironmental conditions that often occur during summer monthsin the sugar beet growing regions in South-Eastern Europe. In theabsence of appropriate control measures in the areas with a highdisease incidence, severe epidemics of CLS can result in a significantreduction of root yield, recoverable sugar, sucrose concentrationand an increase in impurities leading to higher processing costs
gl), [email protected]
(Windels et al., 1998; Jacobsen and Franc, 2009). CLS is primarilycontrolled by fungicide applications, although it can also bemanaged by planting disease-tolerant varieties, crop rotation andtillage. In conditions of a high disease pressure, frequent re-applications of the same fungicide are common. This can selectfor resistance in the target organisms, which is the most importantlimiting factor for CLS chemical control. Several classes of protec-tive and systemic fungicides with various modes of action areavailable for use against this disease worldwide. In the West Bal-kans, benzimidazoles are among the fungicides registered for CLScontrol, while the sterol demethylation inhibiting (DMI) fungicideshave the highest number of registered active ingredients available.Since these fungicide groups have a site-specific mode of action,there is a high risk for developing resistance (Brent and Hollomon,2007).

D. Budakov et al. / Crop Protection 66 (2014) 120e126 121
Benzimidazoles act primarily by binding to fungal tubulin andinterfering with mitosis and the fungal cytoskeleton (Davidse,1986). The mechanism of resistance to benzimidazole fungicidesin C. beticola is based on a mutation in the b-tubulin gene, whichreduces benzimidazole binding (Davidson et al., 2006). Benzimid-azole fungicides have been used extensively in the management ofCLS, but resistance to them has become a major problem in manysugar beet growing areas (Georgopoulos and Dovas, 1973; Briereet al., 2001; Weiland and Halloin, 2001). Once resistance hasdeveloped, the frequency of benzimidazole-resistant strains re-mains stable or only drops slowly even in the absence of selectionpressure (Karaoglanidis and Ioannidis, 2010).
Sterol demethylation inhibiting (DMI) fungicides are inhibitorsof sterol C-14 alpha-demethylation during ergosterol biosynthesis,which is the main fungal sterol (Siegel, 1981). They represent themost important class of fungicides used to control CLS, providing anexcellent protective and curative activity against Cercospora spp.(Dahmen and Staub, 1992). In the United States, a reduction in theperformance of DMI fungicides has occurred over time as a result ofthe presence of C. beticola strains with a reduced sensitivity to DMIs(Secor et al., 2010). Population shift towards DMI resistance wasstep-wise, which indicates a polygenic control of the resistance(Brent and Hollomon, 2007). Resistance problems with the use ofDMIs have been reported in several phytopathogenic fungi and twodifferent resistance mechanisms have been identified: mutations inthe C-14 alpha-demethylase (CYP51) gene and overexpression ofthe CYP51 gene (Ma and Michailides, 2005). It has been suggestedthat overexpression of the C-14 alpha-demethylase (Cyp51) gene isthe mechanism of DMI resistance in C. beticola (Nikou et al., 2009;Bolton et al., 2013).
Northern Serbia is a major sugar beet producing region withabout 60,000 ha under this crop. Since a reduction in the perfor-mance of benzimidazole and some DMI fungicides has beenobserved (Mari�c et al., 1976; Bala�z et al., 1999; Budakov et al., 2010),the aims of this study were to: i) perform a sensitivity screening ofC. beticola isolated from the sugar beet growing areas in Serbia tobenzimidazole and DMI fungicides; ii) develop a CAPS (CleavedAmplified Polymorphic Sequence) marker for detection of theresistant and sensitive isolates to benzimidazoles; iii) test a
Fig. 1. Map of Northern Serbia showing locations w
previously developed CAPS marker for detection of the DMI resis-tant isolates; iv) field-evaluate fungicide efficacy on CLS control,caused by the isolates with known sensitivity levels to benzimid-azoles and DMIs; and v) evaluate how fungicide resistance deter-mined by in vitro tests and CAPS markers correlates with a reduceddisease control in the field.
2. Materials and methods
2.1. Sampling of leaves and collection of fungal isolates
Sugar beet leaves with typical symptoms of CLS were collectedduring the growing season of 2007, from 70 locations in the mainsugar beet growing region in Serbia (Fig. 1). A total of 100 isolateswere collected, with the number of isolates per location beingproportional to the acreage under sugar beet in that area. Allsampling sites had a history of benzimidazole and DMI fungicideuse, including carbendazim and flutriafol. Collected leaves werewashed under tap water and the lesions were cut out and placed inPetri dishes on a wet filter paper. After 24e36 h of incubation at25e26 �C, monosporial isolations were performed after conidia hadformed within lesions. Single conidia were lifted with a sterileentomological needle and placed on potato dextrose agar (PDA, CarlRoth GmbH). The cultures were transferred onto PDA slants andkept refrigerated at 4 �C until further analysis.
2.2. In vitro sensitivity testing
For in vitro fungicide sensitivity tests, we used two commerciallyavailable fungicide formulations containing flutriafol and carben-dazim as active ingredients: Takt (125 g a.i. L�1
flutriafol, Herbos,Croatia) and Galofungin (500 g a.i. L�1 carbendazim, Galenika,Serbia). Baseline sensitivities for flutriafol and carbendazim weredetermined by testing five references C. beticola isolates (CB 581, CB582, CB 586, CB 587 and CB 588) from beetroot and chard grown infields in which none of the fungicides examined had been used. Inorder to test the sensitivity to flutriafol, the concentrations of 0.3,0.6, 1.25, 2, 4, 8 and 16 mg mL�1 were applied in the medium. Anaverage EC50 was 0.8 mg mL�1, hence the next higher concentration
here Cercospora beticola isolates were collected.

D. Budakov et al. / Crop Protection 66 (2014) 120e126122
(1.25 mg mL�1) was a discriminative concentration. The sensitivityto carbendazimwas tested with a series of concentrations: 1, 2, 4, 5,8, 10, 16, 20, 32 and 50 mg mL�1. Since none of the tested isolateswas suppressed with tested concentrations, a discriminative con-centration of 5 mg mL�1 was used (Weiland and Halloin, 2001). Theautoclaved PDA was amended with fungicide aqueous solutions,while the control Petri dishes contained non-amended PDA. Thetests were performed in 90 mm-diameter Petri dishes with 3mycelial plugs per isolate (5 mm in diameter) turned upside downon the fungicide-amended PDA and the control Petri dishes. Eachisolate-fungicide combination was replicated 3 times. A colonydiameter was determined by averaging two perpendicular mea-surements of the diameter after 4 days of incubation at 25e26 �Cand subtracting the diameter of the mycelial plug. Obtained valueswere used to calculate the relative growth (RG). Based on the meanRG, the isolates were divided into four groups: i) sensitive(RG < 20%); ii) with a decreased sensitivity (RG ¼ 20e39.9%); iii)moderately resistant (RG¼ 40e69.9%); and iv) resistant (RG� 70%).
2.3. Molecular analysis
After the fungicide sensitivity was determined in vitro, 27 iso-lates with varying degrees of resistance to flutriafol were chosen forfurther analysis of the C-14 alpha-demethylase gene, while 23isolates were chosen for analysis of the b-tubulin gene, in order toidentify the carbendazim resistance-causing mutation.
Prior to DNA extraction, the isolates were grown on PDA for 10days at 25e26 �C, after which 500 mg of fungal tissue was har-vested by scraping off mycelia from the agar surface. The DNA wasextracted following the procedure described by Budakov et al.(2012).
In order to detect a mutation indicating the DMI resistant iso-lates, specific primers CYP51RT-F (50-AACTCCAAATTGATGGAGCA-30) andCYP51RT-R (50-CGGCTAGCAGTGTAAATGGT-30)wereused foramplification of the C-14 alpha-demethylase gene fragment (Nikouet al., 2009). Amplification started with an initial denaturation at94 �C for 4 min, followed by 38 cycles of denaturation at 94 �C for40 s, annealing at 52 �C for 35 s and extension at 72 �C for 45 s. Thefinal extension was at 72 �C for 10 min. Primers Bt512F (50-CCAGCTTTTCCGCCCAGACAAC-30) and Bt922R (50-ACGGCACCATGTTCACGGCAAGC-30) were used for amplification of the b-tubulingene fragment (Davidson et al., 2006). The amplification profile wasas follows: initial denaturation at 94 �C for 4 min, followed by 38cycles of denaturation at 94 �C for 40 s, annealing at 62 �C for 40 sand extension at 72 �C for 1min. The final extension lasted 10min at72 �C. All PCR reactions were carried out in a total volume of 25 mLcontaining: 30 ng of the DNA template, 2.5 mL of buffer, 0.2 mM ofeach dNTP, 2.5 mM MgCl2, 2 units of Taq polymerase (Fermentas)and 0.25 mM of each primer (Metabion). The reactions were per-formed in TPersonal and T1 thermocyclers (Biometra). The PCRproducts were separated on a 1% agarose gels containing 0.005%ethidium bromide and visualized under UV light. RestrictionenzymeAlw26I (BsmAI, Fermentas)was used todetect themutationin C-14 alpha-demethylase gene (Nikou et al., 2009). After restric-tionmapping of the b-tubulin gene sequence (Davidson et al., 2006),a mutation responsible for resistance to benzimidazoles at position198 was detected by digestion with Bsh1236I (BSTUI, Fermentas)restriction enzyme. The PCR products were digested in the manu-facturer's reaction buffer and electrophoresed on a 1.7% agarose gelcontaining 50% high resolution agarose (Carl Roth GmbH).
2.4. Field trials
Fungicides containing active ingredients carbendazim and flu-triafol were used in field trials to test their efficacy in controlling
the selected C. beticola isolates. In order to test for alternativespraying options in areas with a reported decreased sensitivity tocarbendazim and flutriafol, azoxystrobin (Quinone outsideInhibitors-QoIs)- and tetraconazole (DMIs)-based fungicides werealso included. Azoxystrobin was selected as a representative of anovel group of fungicides in Serbia that has already been widelyused in CLS control worldwide. Tetraconazole was chosen forseveral reasons: i) it belongs to the same chemical group as flu-triafol, but has been reported to have higher efficacy; ii) no cross-resistance with flutriafol has been reported yet; and iii) it is regis-tered for use in Serbia as a single ingredient in pesticide formula-tions. The isolates with known sensitivity levels to flutriafol andcarbendazim were selected previously in vitro and used as inoc-ulum for field trials. The inoculum was prepared in the form of aconidial suspension with conidia produced on Sugar Beet Leafextract Agar (SBLA) (150e200 g of fresh sugar beet leaves in 1 L ofdistilled water þ 20% Agar). Concentration of the suspension was500 spores per ml, which was determined by counting spores usinga haemocytometer.
Field trials were conducted on sugar beet cv. LARA, during thegrowing seasons of 2010 and 2011, on an experimental field of theInstitute of Field and Vegetable Crops, Novi Sad, Serbia, in acompletely randomized factorial design with four replicates. In2010 and 2011, the size of one plot was 12 m2 (3 rows of sugar beet,8 m long) and 24 m2 (6 rows of sugar beet, 8 m long), respectively.Individual plots were inoculated on July 8th 2010 and July 4th 2011,when the weather conditions were favorable for disease develop-ment, using a back-pack sprayer (Solo 456) and 200 L of water perha. The plots were inoculatedwith four isolatemixtures, containingthe isolates with a similar sensitivity to tested fungicides: a) sen-sitive to flutriafol and carbendazim (FSBS); b) moderately resistantto flutriafol and sensitive to carbendazim (FRBS); c) resistant toflutriafol and carbendazim (FRBR); and d) sensitive to flutriafol andresistant to carbendazim (FSBR) (Appendix B). Each inoculated plot,except for the non-treated control, was treated throughout thevegetation period with one of the following fungicides: Galofungin(Galenika, Serbia, carbendazim 500 g a.i. L�1), Takt (Herbos, Croatia,flutriafol 125 g a.i. L�1), Eminent (Isagro, tetraconazole 125 g a.i.L�1) and Quadris (Syngenta, azoxystrobin 125 g a.i. L�1). The firstfungicide treatment was applied seven days after inoculation, withsubsequent treatments on August 1st, August 20th and September4th in 2010 and July 20th, August 3rd and August 16th in 2011. Thefungicides were applied using a Solo 456 back-pack sprayer at400 L ha�1 of fungicide mixture. Disease intensity was evaluatedfour times until harvest on the same dates when the fungicidetreatments were applied. Evaluation was based on the percent ofleaf infection using a scale from 0 to 9, where 0 represents a healthyplant and 9 is a plant with 100% necrotic leaves. These values wereused to calculate the AUDPC (Area Under the Disease ProgressCurve) (Wolf and Veerret, 2002). Approximately two weeks afterthe last treatment in each year, the plants were harvested and theroot yield and sugar content were determined in a sugar beetautomatic laboratory (Venema Automation BV, The Netherlands).Fungicide efficacy was calculated on the basis of an averagepenultimate disease intensity evaluation.
2.5. Data analysis
Results of the field trials: AUDPC, root yield and sugar concen-tration were expressed as mean ± standard deviation. Statisticalanalysis was performed by analysis of variance (ANOVA) usingStatistica 12 software (StatSoft, Tulsa, OK, USA). Comparisons be-tween means were made with Fisher's least significant difference(LSD) test, P ¼ 0.05, among the isolates for each fungicidetreatment.

Table 1Sensitivity of C. beticola isolates and composition of isolate mixtures used in field trials.
Isolate Flutriafol Carbendazim Isolates used in field trials
RG (%) Sensitivity RG % Sensitivity
In vitro CAPS In vitro CAPS FRBR FRBS FSBR FSBS
CB 80 113.33 R R 121.90 R e þCB 254 112.30 R R 102.98 R R þCB 84 106.74 R e 101.00 R e þCB 546 106.62 R R 104.83 R R þCB 72 101.82 R e 98.58 R e þCB 253 101.81 R e 102.72 R e þCB 543 99.03 R R 105.07 R e þCB 551 96.77 R e 106.81 R e þCB 538 95.74 R R 97.59 R R þCB 179 92.86 R e 99.31 R e þCB 591 90.16 R R 106.03 R R þCB 412 89.66 R e 99.43 R e þCB 523 87.86 R e 91.62 R e þCB 605 86.00 R R 86.00 R R þCB 600 82.48 R e 98.60 R e þCB 504 75.27 R R 98.92 R R þCB 82 69.75 MR R 105.24 R RCB 10 69.22 MR S 103.26 R e
CB 603 68.53 MR S 2.80 S S þCB 3 63.25 MR S 96.14 R RCB 270 60.00 MR S 97.50 R e
CB 257 53.54 MR S 100.00 R RCB 152 50.00 MR S 112.38 R RCB 94 48.57 MR S 137.14 R RCB 515 47.90 MR S 10.86 S S þCB 322 46.75 MR S 94.08 R RCB 236 44.90 MR S 123.88 R RCB 50 32.00 DS e 102.83 R e þCB 382 24.42 DS S 93.02 R e
CB 561 24.11 DS e 104.56 R e þCB 133 23.30 DS S 98.92 R RCB 317 20.10 DS S 97.82 R RCB 526 19.87 S e 95.96 R e þCB 175 19.80 S e 99.42 R e þCB 74 19.74 S e 97.99 R e þCB 183 19.10 S e 94.24 R e þCB 319 19.05 S e 127.27 R e þCB 188 18.75 S e 102.74 R e þCB 609 18.39 S e 101.53 R e þCB 191 17.54 S e 100.78 R e þCB 316 17.14 S S 2.86 S S þCB 75 17.06 S e 102.78 R e þCB 577 16.84 S e 92.63 R e þCB 44 16.67 S e 104.66 R e þCB 112 16.51 S e 149.21 R e þCB 206 16.27 S e 100.02 R e þCB 20 15.48 S e 77.42 R e þCB 300 14.97 S e 113.26 R e þCB 292 14.33 S e 102.81 R e þCB 567 13.89 S e 102.07 R e þCB 135 13.79 S e 99.30 R e þCB 596 12.59 S e 128.67 R e þCB 165 11.67 S e 91.27 R e þCB 207 11.30 S e 100.33 R e þCB 115 10.98 S e 97.65 R e þCB 125 9.87 S S 0.00 S S þCB 33 9.50 S e 99.01 R R þCB 587 9.43 S S 84.18 R R þCB 157 8.33 S S 94.02 R R þCB 1 2.26 S e 113.56 R e þCB 171 0.00 S e 99.81 R e þCB 173 0.00 S e 93.43 R e þCB 204 0.00 S e 104.65 R e þCB 369 0.00 S S 95.27 R e þCB 388 0.00 S e 98.99 R R þCB 425 0.00 S e 97.05 R e þCB 440 0.00 S e 98.78 R e þ
RG-relative growth; S-Sensitive; DS-Decreased sensitivity; MR-Moderately resistant; R-Resistant; FRBR-resistant to flutriafol and carbendazim; FRBS-moderately resistant toflutriafol and sensitive to carbendazim; FSBR-sensitive to flutriafol and resistant to carbendazim; FSBS-sensitive to flutriafol and carbendazim.
D. Budakov et al. / Crop Protection 66 (2014) 120e126 123

D. Budakov et al. / Crop Protection 66 (2014) 120e126124
3. Results
3.1. In vitro sensitivity testing
A relative mycelial growth on the medium amended with flu-triafol in comparison to the non-amended control ranged from 0%to 113.3% (Table 1). Thirty six percent of isolates were sensitive toflutriafol, 38% had a decreased sensitivity, 10% were moderatelyresistant and 16% of the tested isolates were resistant to flutriafol.When the medium was amended with carbendazim, 96 isolateshad an equal growth on the fungicide-amended and non-amendedmedium (77.4e149.2%). In four isolates (CB125, CB316, CB515, andCB603), RG was suppressed and varied from 0 to 10.99% oncarbendazim-amendedmedium. After in vitro sensitivity testing, 16isolates resistant to flutriafol and carbendazim (FRBR), 2 moder-ately resistant to flutriafol and sensitive to carbendazim (FRBS), 36isolates sensitive to flutriafol and resistant to carbendazim (FSBR)and 2 sensitive to flutriafol and carbendazim (FSBS) were used forfurther investigation.
3.2. CAPS markers
Eight resistant (CB80, CB254, CB546, CB543, CB538, CB591,CB605, CB504), 11 moderately resistant (CB82, CB10, CB603, CB3,CB257, CB270, CB152, CB94, CB515, CB322, CB236) and 8 isolatessensitive to flutriafol (CB382, CB133, CB317, CB316, CB125, CB587,CB157, CB369) were chosen for further analysis of the C-14 alpha-demethylase gene using CAPS, while 19 resistant (CB254, CB538,CB546, CB591, CB82, CB152, CB257, CB322, CB33, CB504, CB157,CB388, CB587, CB3, CB133, CB317, CB236, CB605, CB94) and allisolates sensitive (CB125, CB316, CB515, CB603) to carbendazimwere chosen for detection of the mutation in the b-tubulin generesponsible for resistance (Table 1). After PCR reactions with C-14alpha-demethylase gene-specific primers, an amplification productof 200 bp in size was obtained in all isolates. After restriction withAlw26I enzyme, only one band of 200 bp was detected in theresistant isolates (RG � 70%) and the moderately resistant isolateCB82 (RG 69.35%), whereas two bands of 80 and 120 bp weredetected in the sensitive and other moderately resistant isolates(Fig. 2a). As a result of amplification with the b-tubulin specific
Fig. 2. Detection of flutriafol and carbendazim resistance in C. beticola isolates using CAPS. (aisolates were: (1) CB80, (2) CB254, (3) CB538, (4) CB543, (5) CB546, (6) CB591, (7) CB605, (8)(16) CB33, (17) CB157, (18) CB369, (19) CB388. M ¼ 100 bp GeneRulerTM, Fermentas, M1 ¼fragment with Bsh1236I. Tested isolates were: (1) CB504, (2) CB538, (3) CB543, (4) CB546, (5CB157, (14) CB587, (15) CB603, (16) CB125. M ¼ 100 bp GeneRulerTM, Fermentas.
primers, a 570 bp-long PCR product was obtained in all isolates. Asexpected, digestion of the amplification product with Bsh1236Ienabled differentiation between the resistant and the sensitiveisolates: three bands were detected in the resistant isolates (250,200 and 120 bp), whereas there were two bands in the sensitiveisolates (450 and 120 bp) (Fig. 2b).
3.3. Field trials
Based on the results of the in vitro sensitivity tests and the CAPSmarker analysis, C. beticola isolates were used for sugar beet in-oculations in field trials aimed at determining fungicide efficacy. Inboth 2010 and 2011, disease progressed quickly after inoculationdue to favorable environmental conditions, i.e. high temperaturesand humidity during July and August. The first symptoms wereobserved seven days after inoculation, at the moment of the firstfungicide treatment. Disease symptoms developed uniformly in allplots, with the spots being formed mainly on middle leaves withinthe rosette. At the second disease evaluation, a black pseudostromaformed within the spots. The disease progressed fastest in non-treated control plots and in plots inoculated with the resistantisolates treatedwith the fungicides corresponding to the resistance.In those plots, the leaf canopy completely decayed and regrowthoccurred by the middle and end of August. In both years, diseaseintensity (AUDPC) was highest in the non-treated control, while thelowest AUDPC occurred in all inoculated plots treated with azox-ystrobin (Fig. 3a). Disease intensity was higher in plots inoculatedwith the isolates resistant to flutriafol and carbendazim and treatedwith the fungicides corresponding to the resistance. We did notobserve a noticeable pattern in the effect of fungicide treatments onroot yield in plots inoculated with the selected isolates (Fig. 3b).Sugar concentration was lowest in plots treated with carbendazimand flutriafol and inoculated with isolates resistant to both fungi-cides (FRBR), while in 2010 this was also the case in plots inoculatedwith FRBS isolates and treatedwith flutriafol (Fig. 3c). In 2010, therewere no differences in sugar content in azoxystrobin and tetraco-nazole treatments, regardless of the sensitivity of the isolates usedfor inoculation. In carbendazim treatments, efficacy was very low inplots inoculated with isolates resistant to this fungicide (Fig. 3d). Influtriafol treatments, efficacy was very low in plots inoculated with
) Restriction of C-14 alpha-demethylase product with Alw26I restriction enzyme. TestedCB82, (9) CB587, (10) CB603, (11) CB94, (12) CB152, (13) CB257, (14) CB207, (15) CB322,DNA Ladder, Low Range, GeneRulerTM, Fermentas; (b) Restriction of b-tubulin gene
) CB605, (6) CB94, (7) CB33, (8) CB254, (9) CB591, (10) CB82, (11) CB257, (12) CB322, (13)
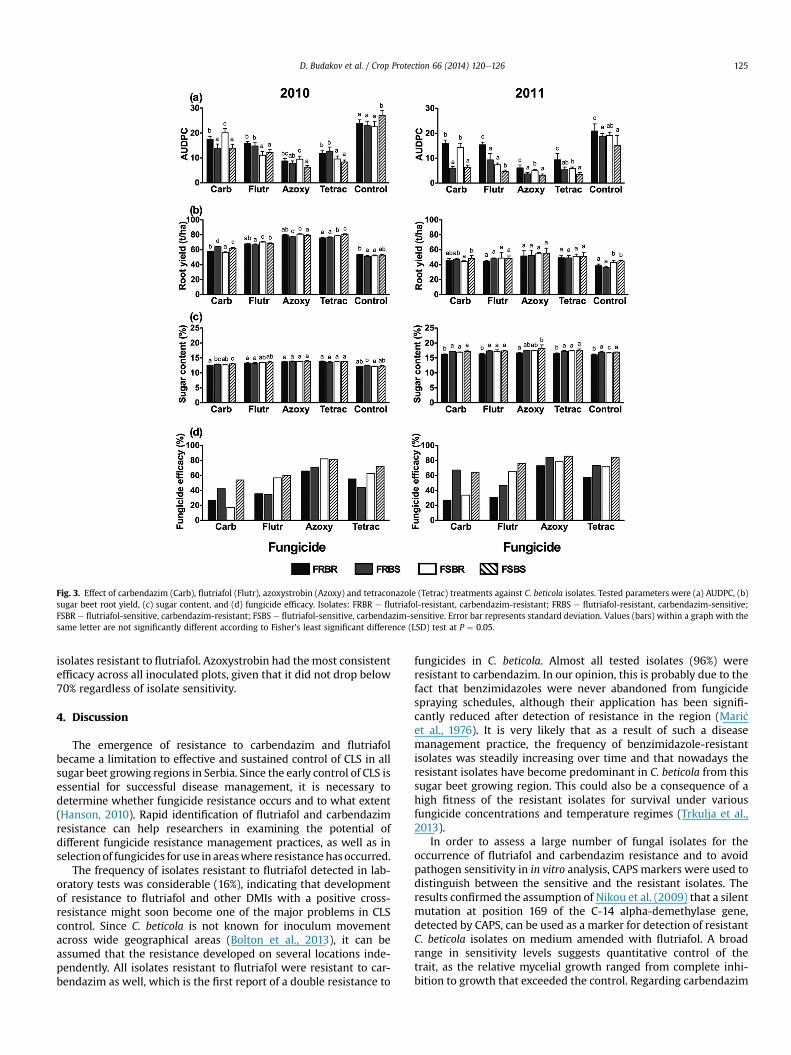
Fig. 3. Effect of carbendazim (Carb), flutriafol (Flutr), azoxystrobin (Azoxy) and tetraconazole (Tetrac) treatments against C. beticola isolates. Tested parameters were (a) AUDPC, (b)sugar beet root yield, (c) sugar content, and (d) fungicide efficacy. Isolates: FRBR e flutriafol-resistant, carbendazim-resistant; FRBS e flutriafol-resistant, carbendazim-sensitive;FSBR e flutriafol-sensitive, carbendazim-resistant; FSBS e flutriafol-sensitive, carbendazim-sensitive. Error bar represents standard deviation. Values (bars) within a graph with thesame letter are not significantly different according to Fisher's least significant difference (LSD) test at P ¼ 0.05.
D. Budakov et al. / Crop Protection 66 (2014) 120e126 125
isolates resistant to flutriafol. Azoxystrobin had the most consistentefficacy across all inoculated plots, given that it did not drop below70% regardless of isolate sensitivity.
4. Discussion
The emergence of resistance to carbendazim and flutriafolbecame a limitation to effective and sustained control of CLS in allsugar beet growing regions in Serbia. Since the early control of CLS isessential for successful disease management, it is necessary todetermine whether fungicide resistance occurs and to what extent(Hanson, 2010). Rapid identification of flutriafol and carbendazimresistance can help researchers in examining the potential ofdifferent fungicide resistance management practices, as well as inselectionof fungicides for use in areaswhere resistancehasoccurred.
The frequency of isolates resistant to flutriafol detected in lab-oratory tests was considerable (16%), indicating that developmentof resistance to flutriafol and other DMIs with a positive cross-resistance might soon become one of the major problems in CLScontrol. Since C. beticola is not known for inoculum movementacross wide geographical areas (Bolton et al., 2013), it can beassumed that the resistance developed on several locations inde-pendently. All isolates resistant to flutriafol were resistant to car-bendazim as well, which is the first report of a double resistance to
fungicides in C. beticola. Almost all tested isolates (96%) wereresistant to carbendazim. In our opinion, this is probably due to thefact that benzimidazoles were never abandoned from fungicidespraying schedules, although their application has been signifi-cantly reduced after detection of resistance in the region (Mari�cet al., 1976). It is very likely that as a result of such a diseasemanagement practice, the frequency of benzimidazole-resistantisolates was steadily increasing over time and that nowadays theresistant isolates have become predominant in C. beticola from thissugar beet growing region. This could also be a consequence of ahigh fitness of the resistant isolates for survival under variousfungicide concentrations and temperature regimes (Trkulja et al.,2013).
In order to assess a large number of fungal isolates for theoccurrence of flutriafol and carbendazim resistance and to avoidpathogen sensitivity in in vitro analysis, CAPS markers were used todistinguish between the sensitive and the resistant isolates. Theresults confirmed the assumption of Nikou et al. (2009) that a silentmutation at position 169 of the C-14 alpha-demethylase gene,detected by CAPS, can be used as a marker for detection of resistantC. beticola isolates on medium amended with flutriafol. A broadrange in sensitivity levels suggests quantitative control of thetrait, as the relative mycelial growth ranged from complete inhi-bition to growth that exceeded the control. Regarding carbendazim

D. Budakov et al. / Crop Protection 66 (2014) 120e126126
resistance, mutation of the b-tubulin gene at position 198 wassuccessfully detected by CAPS and enabled differentiation betweenthe sensitive and resistant isolates, as confirmed with other fungalpathogens (Canas-Gutierrez et al., 2006; Chung et al., 2010). Inselected isolates, sensitivity levels to carbendazim and, for highlyresistant phenotypes, to flutriafol, were detected by CAPS markers.These data completely correlate with the results of in vitroscreening, making CAPS markers a useful tool in monitoringC. beticola for fungicide resistance.
During the field trials in 2010, fungicide efficacy was relativelylow, even in fungicides with known high efficacy (flutriafol, azox-ystrobin and tetraconazole), which can be explained by extremelyfavorable environmental conditions for disease development dur-ing July and August in that particular year. Azoxystrobin treatmentwas highly effective against CLS, probably due to its multiple effectson this pathogen, such as inhibition of spore germination, as well aseradicative and curative effect (Anesiadis et al., 2003; Karaoglanidisand Bardas, 2006). Tetraconazole provided very effective CLS con-trol that resulted in high yield and high sucrose content, as previ-ously described (Khan and Smith, 2005). Although it is known thata cross-resistance between the DMI fungicides exists, various DMIscan have different resistance mechanisms in the target organism(Karaoglanidis and Ioannidis, 2010). In this experiment, tetraco-nazole had a higher efficacy than flutriafol in controlling flutriafol-resistant isolates, which can be explained by a relatively stablesensitivity of C. beticola to tetraconazole (Secor et al., 2010) and adifferential fungicide efficacy between members of the DMI group(Karaoglanidis and Ioannidis, 2010). Flutriafol efficacy was very lowin plots inoculated with flutriafol-resistant (FRBR) isolates and inplots inoculated with moderately resistant (FRBS) isolates. Efficacyof carbendazim was very low in plots inoculated with isolatesresistant to this fungicide, which is in agreement with the quali-tative nature of resistance to benzimidazoles.
Detection of carbendazim resistance using CAPS and in vitrolaboratory tests was also confirmed in field trials. According toNikou et al. (2009), a mutation in the C-14 alpha-demethylase genecan be used for detection of C. beticola isolates highly resistant toflutriafol. This, however, is not in accordancewith the findings fromBolton et al. (2012) that specific polymorphisms in or near theCbCyp51 gene are not associated with an enhanced resistance toDMI fungicides in C. beticola. In our current study, the use of thisparticular CAPS marker for detection of the abovementioned mu-tation resulted in a successful differentiation between the isolateswith a high level of resistance. Although the CAPS marker for flu-triafol resistance is not functionally connected to expression of theresistance itself, the results from our study indicate that it could bea useful tool for detecting resistance to flutriafol in C. beticola iso-lates from northern Serbia.
In summary, the detection and measurement of C. beticolaresistance to fungicides prior to actual control failures can help indeveloping strategies for the prevention of increased fungicideresistance and the successful management of CLS.
Acknowledgments
Funding for this research was provided by the Ministry of Ed-ucation and Science, Republic of Serbia, project number TR 31015.The authors wish to thank Dr. Bojana Stanic for her assistance inEnglish language editing.
Appendix A. Supplementary data
Supplementary data associated with this article can be found inthe online version, at http://dx.doi.org/10.1016/j.cropro.2014.09.010.
References
Anesiadis, T., Karaoglanidis, G., Tzavella-Klonari, K., 2003. Protective, curative anderadicant activity of the Strobilurine fungicide azoxystrobin against Cercosporabeticola and Erysiphe betae. J. Phytopathol. 151, 647e651.
Bala�z, F., Stoj�sin, V., Gavran-Starovi�c, M., 1999. Sensitivity of Cercospora beticola Sacc.in Vojvodina to some fungicides. Pesticides 14, 5e13.
Bolton, M., Birla, K., Rivera-Varas, V., Rudolf, K., Secor, G., 2012. Characterization ofCbCyp51 from field isolated of Cercospora beticola. Phytopathology 102, 298e305.
Bolton, M., Rivera, V., Secor, G., 2013. Identification of the G143A mutation associ-ated with QoI resistance in Cercospora beticola field isolates from Michigan,United States. Pest Manag. Sci. 69, 35e39.
Brent, K.J., Hollomon, D.J., 2007. Fungicide Resistance in Crop Pathogens: How can itbe Managed. Fungicide Resistance Action Committee, Brussels, Belgium.
Briere, S.C., Franc, G.D., Kerr, E.D., 2001. Fungicide sensitivity characteristics ofCercospora beticola isolates recovered from the high plains of Colorado, Mon-tana, Nebraska and Wyoming. 1. Benzimidazole and Triphenyltin hydroxide.J. Sugar Beet Res. 38, 111e120.
Budakov, D., Nagl, N., Ta�ski-Ajdukovi�c, K., Stoj�sin, V., Bagi, F., 2012. Evaluation ofRapid protocols for DNA isolation from Cercospora beticola Sacc. Field Veg. CropRes. 49, 202e207.
Budakov, D., Stoj�sin, V., Bagi, F., Jani�cijevi�c, M., 2010. Managing resistance of Cercosporabeticola Sacc. as a basis for integral concept of disease control on sugar beet. In:Kastori, R., Kadar, I., Aleksi�c, N., Kozi�c, T., Grbi�c,M.,Ðuranovi�c, S., Samsonov,M. (Eds.),Proceedings of XIV International Eco-conference, Novi Sad, Serbia, pp. 169e173.
Canas-Gutierrez, G.P., Patino, L.F., Rodriguez-Arango, E., Arango, R., 2006. Molecularcharacterization of benomyl-resistant isolates of Mycosphaerella fijiensis,collected in Colombia. J. Phytopathol. 154, 403e409.
Chung, W.H., Chung, W.C., Peng, M.T., Yang, H.R., Huang, J.W., 2010. Specificdetection of benzimidazole resistance in Colletotrichum gloeosporoides fromfruit crops by PCR-RFLP. New Biotechnol. 27, 17e24.
Dahmen, H., Staub, T., 1992. Protective, curative and eradicant activity of difeno-conazole against Venturia inaequalis, Cercospora arachidicola and Alternariasolani. Plant Dis. 76, 774e777.
Davidse, L.C., 1986. Benzimidazole fungicides: mechanisms of action and biologicalimpact. Annu. Rev. Phytopathol. 24, 43e65.
Davidson, R.M., Hanson, L.E., Franc, G.D., Panella, L., 2006. Analysis of b-tubulinegene fragments from benzimidazole-sensitive and -tolerant Cercospora beticola.J. Phytopathol. 154, 321e328.
Georgopoulos, S.G., Dovas, C., 1973. Occurrence of Cercospora beticola strains resistantto benzimidazole fungicides in northern Greece. Plant Dis. Rep. 62, 321e324.
Hanson, L., 2010. Genetics of fungicide resistance in Cercospora and Mycosphaerella.In: Lartey, R., Weiland, J.J., Panella, L., Crous, P., Windels, C. (Eds.), CercosporaLeaf Spot of Sugar Beet and Related Species. APS Press, St. Paul, MN, USA,pp. 179e188.
Jacobsen, B., Franc, G., 2009. Cercospora leaf spot. In: Harveson, R.M., Hanson, L.E.,Hein, G.L. (Eds.), Compendium of Beet Diseases and Pests. APS Press, St. Paul,MN, USA, pp. 7e10.
Karaoglanidis, G., Bardas, G., 2006. Control of benzimidazol- and DMI-resistantstrains of Cercospora beticolawith strobilurin fungicides. Plant Dis. 90, 419e424.
Karaoglanidis, G.S., Ioannidis, P., 2010. Fungicide resistance of Cercospora beticola inEurope. In: Lartey, R., Weiland, J.J., Panella, L., Crous, P., Windels, C. (Eds.),Cercospora Leaf Spot of Sugar Beet and Related Species. APS Press, St. Paul, MN,USA, pp. 189e211.
Khan, M., Smith, L., 2005. Evaluating Fungicides for controlling of Cercospora leafspot on sugar beet. Crop Prot. 24, 79e86.
Ma, Z., Michailides, T.J., 2005. Advances in understanding molecular mechanisms offungicide resistance and molecular detection of resistant genotypes in phyto-pathogenic fungi. Crop Prot. 24, 853e863.
Mari�c, A., Petrov, M., Ma�sirevi�c, S., 1976. Occurrence of tolerance in Cercosporabeticola Sacc to benomyl in Yugoslavia and management practice against thispathogen. Za�stita bilja 27, 227e236.
Nikou, D., Malandrakis, A., Konstantakaki, M., Vontas, J., Markoglou, A., Ziogas, B.,2009. Molecular characterization and detection of overexpressed C-14 alpha-demethylase-based DMI resistance in Cercospora beticola field isolates. Pestic.Biochem. Phys. 95, 18e27.
Secor, G.A., Rivera-Varas, V.V., Gudmestad, N.C., Weiland, J.J., 2010. Sensitivity ofCercospora beticola to foliar fungicides in the Red River Valley of North Dakotaand Minnesota. In: Lartey, R., Weiland, J.J., Panella, L., Crous, P., Windels, C.(Eds.), Cercospora Leaf Spot of Sugar Beet and Related Species. APS Press, St.Paul, MN, USA, pp. 213e222.
Siegel, M.R., 1981. Sterol-inhibiting fungicides: effects on sterol biosynthesis andsites of action. Plant Dis. 65, 986e989.
Trkulja, N., Ivanovi�c, �Z., Pfaf-Dolovac, E., Dolovac, N., Mitrovi�c, P., To�sevski, I., Jovi�c, J.,2013. Characterization of benzimidazole resistance of Cercospora beticola inSerbia using PCR-based detection of resistance-associated mutations of the b-tubulin gene. Eur. J. Plant Pathol. 135, 889e902.
Weiland, J.J., Halloin, J.M., 2001. Benzimidazole resistance in Cercospora beticolasampled from sugarbeet fields in Michigan, USA. Can. J. Plant Pathol. 23, 78e82.
Windels, C.E., Lamey, H.A., Hilde, D., Widner, J., Knudsen, T., 1998. Cercospora leafspot model for sugar beet: in practice by the industry. Plant Dis. 82, 716e726.
Wolf, P.F.J., Veerret, J.A., 2002. An Integrated Pest Management System in Germanyfor the control of fungal leaf diseases in sugar beet: the IPM sugar beet model.Plant Dis. 86, 336e344.
Copyright © 2022 FDOKUMEN




















