
Scent marking in free-ranging tigers, Panthera tigris
10
Anim .Behav., 1989,37, 1 -10 Scentmarkinginfree-rangingtigers, Pantheratigris JAMESL .DAVIDSMITH,CHARLESMcDOUGAL*&DALEMIQUELLEt DepartmentofFisheriesandWildlife,UniversityofMinnesota, StPaul,MN55108,U .S.A . Abstract . Datawerecolletedonthescent-markingpatternsofradio-collaredandvisuallyidentifiable tigersfor4yearsinRoyalChitwanNationalPark,Nepal .Fivecategoriesofmarkingwererecorded :urine spraying;scrapingwithdepositsofurinefaeces,andanalglandsecretions ;clawing;cheekrubbing ;and vegetationflattening .Urinesprayingandscrapingwerethepredominantformsofmarkinginthis population .Tigersmarkedmoreheavilyatterritorialboundariesthanintheinteriorofterritories . Furthermore,inborderareasmarkswerehighlyclumpedatcontactzoneswheremajorroutesoftravel approachedterritorialboundaries .Thispatternappearstobearesultofthedensityofvegetationwhich channelstravel.Theintensityofmarkinginthesezonesrepresentedahigherfrequencyofmarkingrather thananincreaseintimespentintheseareas .Markingwasmostintensivewhentigerswereestablishing territories,andanimalsonadjacentterritoriesappearedtomarkinresponsetoeachother .Females markedintensivelyjustpriortooestrus ;thisbehaviourwasreducedduringoestrus .Malesmarkedmore frequentlywhenfemaleswereinoestrusthanduringotherstagesofthefemales'cycle .Amodelis proposedwhereanodourfieldsignalstheriskofencounteringaconspecific,thusallowinganimalsto comparethecostsofpossibleencounterswiththebenefitsofuseofagivenarea . Behaviouralstudieshavedemonstratedtheimpor- tantrolethatchemicalcommunicationplaysin socialorganizationandpartitioningofspaceand resourcesinmammals(Rails1971 ;Eisenberg& Kleiman1972 ;Muller-Schwarze1977 ;Muller- Schwarze&Silverstein1979 ;Brown&Macdonald 1986) .Amongthecarnivores,therehavebeen severalrecent,analyticalfieldstudiesofchemical communication :incanids(Peters&Mech1975 ; Henry1977 ;Macdonald1979a,b,1980 ;Rothman &Mech1979 ;Whittenetat .1980;Wells&Bekoff 1981),andinhyaenas, Hyaenabrunnes (Millsetal . 1982 ;Gorman&Mills1984).Althoughmarking behaviourhasalsobeendescribedforfree-ranging felids(Schaller1967,1972 ;Hornocker1970 ;Sei- denstickeretal .1973 ;Bailey1974),thisstudyof chemicalcommunicationintigers, Pantheratigris, isthefirstsystematicanalysisoffelidsthatalso includesparallelinformationonsocialorganiza- tion . Ourresearchwaspartofa10-yearinvestigation oftigerbehaviourandecologyinRoyalChitwan National Park, Nepal(Seidensticker 1976; McDougal1977;Sunquist1981 ;Tamang1982 ; Smith1984).Wehadtheopportunitytoanalyse scentmarkinginrelationtothespacingbehaviour *Presentaddress :SmithsonianAssociate, P.O .Box242, Kathmandu,Nepal . tPresentaddress :Department of WildlifeResources, University of Idaho,Moscow,ID 83843,U .S.A . of33regularlymonitoredindividuals .Earlier phasesofthetigerstudiesestablishedthatthe populationhasapolygamousmatingsystemwith intrasexualterritoriality(McDougal1977 ;Sun- quist1981 ;Smith1984 ;Smithetal .1987) .Females havecontiguousterritorieswhichappeartobe resourcebased,i .e .theyprovideadequatefoodand coverforraisingyoung,anJmaleterritoriesover- lapthoseoftwoormorefemales .Initially,data werecollectedonthecategories,locationsand frequencyofmarkingbehaviourthroughoutthe studyarea .Asthemarkingpatternsofindividual tigersemerged,weestablishedtransectsthatwere monitoredregularlytoobtainsystematicinfor- mation .Datawereanalysedtoexaminetwo hypotheses :(1)markingfunctionstoestablishand maintainterritorialboundaries,and(2)marking advertisesfemalereproductivestatus . STUDYAREAANDMETHODS RoyalChitwanNationalPark,Nepal,covering 1040km 2 , isoneofthelargesttigerreservesonthe Indiansubcontinent .ItliesintheTerai,asubtropi- calareaoflowelevationjustnorthoftheGangetic PlainofIndia .Thevegetationalzonesincludeflood plain,composedofinterspersedriverineforests andtallgrasses,opensalforests, Shorearobusta, withatallgrassunderstory,andlow,ruggedhills 0003-3472/89/010001+10$03 .00/0 (7 1989 TheAssociation for theStudy of AnimalBehaviour 1
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Scent marking in free-ranging tigers, Panthera tigris

Anim. Behav., 1989, 37, 1 -10
Scent marking in free-ranging tigers, Panthera tigris
JAMES L . DAVID SMITH, CHARLES McDOUGAL* & DALE MIQUELLEtDepartment of Fisheries and Wildlife, University of Minnesota, St Paul, MN 55108, U.S.A .
Abstract . Data were colleted on the scent-marking patterns of radio-collared and visually identifiabletigers for 4 years in Royal Chitwan National Park, Nepal . Five categories of marking were recorded : urinespraying; scraping with deposits of urine faeces, and anal gland secretions ; clawing; cheek rubbing; andvegetation flattening . Urine spraying and scraping were the predominant forms of marking in thispopulation . Tigers marked more heavily at territorial boundaries than in the interior of territories .Furthermore, in border areas marks were highly clumped at contact zones where major routes of travelapproached territorial boundaries . This pattern appears to be a result of the density of vegetation whichchannels travel. The intensity of marking in these zones represented a higher frequency of marking ratherthan an increase in time spent in these areas . Marking was most intensive when tigers were establishingterritories, and animals on adjacent territories appeared to mark in response to each other . Femalesmarked intensively just prior to oestrus ; this behaviour was reduced during oestrus . Males marked morefrequently when females were in oestrus than during other stages of the females' cycle . A model isproposed where an odour field signals the risk of encountering a conspecific, thus allowing animals tocompare the costs of possible encounters with the benefits of use of a given area .
Behavioural studies have demonstrated the impor-tant role that chemical communication plays insocial organization and partitioning of space andresources in mammals (Rails 1971 ; Eisenberg &Kleiman 1972; Muller-Schwarze 1977; Muller-Schwarze & Silverstein 1979; Brown & Macdonald1986). Among the carnivores, there have beenseveral recent, analytical field studies of chemicalcommunication : in canids (Peters & Mech 1975 ;Henry 1977 ; Macdonald 1979a, b, 1980 ; Rothman& Mech 1979 ; Whitten et at . 1980; Wells & Bekoff1981), and in hyaenas, Hyaena brunnes (Mills et al .1982; Gorman & Mills 1984). Although markingbehaviour has also been described for free-rangingfelids (Schaller 1967, 1972 ; Hornocker 1970 ; Sei-densticker et al . 1973 ; Bailey 1974), this study ofchemical communication in tigers, Panthera tigris,is the first systematic analysis of felids that alsoincludes parallel information on social organiza-tion .
Our research was part of a 10-year investigationof tiger behaviour and ecology in Royal ChitwanNational Park, Nepal (Seidensticker 1976;McDougal 1977; Sunquist 1981 ; Tamang 1982;Smith 1984). We had the opportunity to analysescent marking in relation to the spacing behaviour* Present address : Smithsonian Associate, P.O. Box 242,Kathmandu, Nepal .
t Present address : Department of Wildlife Resources,University of Idaho, Moscow, ID 83843, U .S.A .
of 33 regularly monitored individuals. Earlierphases of the tiger studies established that thepopulation has a polygamous mating system withintrasexual territoriality (McDougal 1977 ; Sun-quist 1981 ; Smith 1984 ; Smith et al . 1987) . Femaleshave contiguous territories which appear to beresource based, i .e . they provide adequate food andcover for raising young, anJ male territories over-lap those of two or more females . Initially, datawere collected on the categories, locations andfrequency of marking behaviour throughout thestudy area. As the marking patterns of individualtigers emerged, we established transects that weremonitored regularly to obtain systematic infor-mation. Data were analysed to examine twohypotheses : (1) marking functions to establish andmaintain territorial boundaries, and (2) markingadvertises female reproductive status .
STUDY AREA AND METHODS
Royal Chitwan National Park, Nepal, covering1040 km 2 , is one of the largest tiger reserves on theIndian subcontinent. It lies in the Terai, a subtropi-cal area of low elevation just north of the GangeticPlain of India . The vegetational zones include floodplain, composed of interspersed riverine forestsand tall grasses, open sal forests, Shorea robusta,with a tall grass understory, and low, rugged hills
0003-3472/89/010001 + 10 $03 .00/0
(7 1989 The Association for the Study of Animal Behaviour1

2
covered with mature sal (Mishra 1982 ; Tamang1982; Smith 1984) .
Data were collected from spring 1978 throughspring 1982 on 27 tigers fitted with radio collars(Smith et al . 1983) and on six additional animalsidentified by patterns of facial markings andunique characteristics of their tracks (McDougal1977). Scent posts and their associated visualmarks were located by searching throughout thePark. A mark was any sign that could be visually orolfactorally discerned ; these were scrapes, scats,anal gland secretions, urine stains on trees orshrubs, claw rakings and cheek rubbing sites . Theanal gland secretions were dark viscous depositswith a distinct odour. During the study we were notcertain of their origin, but Smith later confirmedtheir identity when working with immobilizedtigers at the Minnesota Zoo. Although no studieshave demonstrated that sites of flattened vege-tation, claw raking and cheek rubbing have achemical as well as a visual signal, we believe, alongwith other authors (Wemmer & Scow 1977 ; Reiger1978) that they may deposit odours, so we haveincluded these marks in this study . Scats, analgland secretions, scrapes and flattened vegetationwere found on the ground ; claw raking and cheekrubbing sites were always located on trees . Thelatter were detected by the presence of facial hair onthe trunks of trees . Urination sites were found onshrubs and trees, and also on the ground inassociation with scrape marks . Sprays could belocated by stains and by odours which could bedetected up to 3 weeks and, when fresh, at distancesup to 3 m. Marks of tigers were distinguished fromthose of leopards, Panthera pardus, by examiningthe size and shape of associated tracks or by the sizeof the marks themselves . A mark was judged tohave been inspected by an animal when it deviatedfrom its line of travel and approached to within0 . 5 m of a mark . Linear dimensions of marks andtheir distance from the tiger's reconstructed line oftravel were recorded . Height above ground wasmeasured for urinations, claw rakings and cheekrubbings. Sex of the marking or inspecting animalwas determined by the size and shape of the tracks(McDougal 1977) .
For a continuous, uniform record in an areawhere spatial distribution of local breeding resi-dents was especially well known, two transects wereestablished . The Tree Transect was a 500-m seg-ment of the major road in the Park which was nearthe boundary between two females . Daily for
Animal Behaviour, 37, 1
2 months, we brushed the bare surface soil aroundthe base of all trees that lay within 3 m of the roadedge and had greater than 10 cm diameter at breastheight (N=54). The Trail Transect was a 7 .9-knmsegment of an animal trail which crossed the entirelongitudinal axis of a female's territory ; it wasmonitored twice a week for 12 months to determinethe distribution of marks within a territory . Thisstudy was possible because of the nature of thesubstrate in the study area ; new tracks were clearlyvisible in the loose soil along the transects and wewere able to determine the details of an animal'smovement through each transect including whichtrees it inspected and marked .
RESULTS
Characteristics of Scent MarkingTigers mark recurrently along routes of travel as
well as elsewhere in their territories . Of the fivecategories of marking behaviour we recorded,urine spraying and scraping predominated ; forexample, of the 783 marks recorded over 6 monthsalong the Trail Transect 78 .2% were sprays, 21 . 3%were scrapes, and 0 .5% were other marks .
Urine sprayingTracks showed that as animals moved along
trails and roads they deviated from the line of travelto mark or inspect sites ; they rarely backtracked toreinspect or spray a tree they had already visited . Asample of the marks was measured . Male marksranged from 91 to 140 cm (X±sD=126± 10 cm,N= 58) from the ground to the top of the mark andfrom 10 to 27 cm (X±sD=16±3 cm) in width ;female marks ranged from 43 to 127 cm (T±sD=83 ± 16 cm, N=143) in height and 8 to 34 cm(T±sD=15±3 cm) in width . Because there wasoverlap between the sexes in height and width ofmarks we could not use the height of spray toidentify the sex of a marking animal without theadditional evidence of tracks .
On the Tree Transect, trees were sprayed 1131times in 20 months of observation ; 89`% of thespraying was concentrated on 19 trees . One species,Bauhinia malabaricum, comprised 6% of the treeson the transect, but was sprayed on only threeoccasions (0 . 3%) . In contrast, four out of fiveAdinia cordifolia were among the 10 trees sprayedmost frequently. The size of a tree and its angle oflean may have been important factors determining

preference. The mean diameter at breast height ofthe 10 most frequently sprayed trees was 62 . 3 cm,and seven of these leaned from 8 to 24 degrees .Leaning trees were sprayed on the underside ;Brahmachary & Dutta (1981) believe tigers mayselect these trees because the mark is protectedfrom rainfall .
Two females who visited the transect over a totalof 28 months marked 25 . 8 trees per month on 5 . 1visits per month ; three males who visited during20 months marked 13 . 0 trees per month during 4 . 4visits per month . The more frequent marking byfemales may reflect that the transect coincided withthe boundary between the territories of the twofemales (Smith et al . 1987) . In contrast, on the TrailTransect a male marked 2 . 4 times per km whilewalking 155 . 7 km in 6 months and a female marked1 . 8 times per km while walking 147 . 7 km .
ScrapingScrapes were made on a variety of substrates
from soft sand to rocky dry stream beds . Malescrapes ranged from 33 to 53 cm (X±sD=60±12 cm, N=45) in length and ranged from 19 to46 cm (X±sD = 30 ± 7 cm) in width; female scrapesranged from 41 to 91 cm (X±sD=59± 10 cm,N=43) in length and 17 to 37 cm (X±sD=28±4 cm) in width . The odour of urine was detectableat 54% of the marks, 17% had deposits of faecesand/or anal gland secretions, and 19% had nodetectable odour . Scrapes ranged from I to 187 cm(X±sD=62± 12 cm, N=88) from the centre of thepath and were usually parallel to it .
Vegetation flattening, clawing and cheek rub-bing accounted for a small percentage of the markswe recorded . Their signal function is poorly under-stood, but the interest other tigers showed in thesesites suggested to us that they have a communica-tion function .
Vegetation flatteningOn three occasions we found patches of vegeta-
tion which had been flattened by males, apparentlyby body rolling, a behaviour reported in manycarnivores (Reiger 1978) . These patches, whichwere approximately 2 . 5 x I m, were parallel to andadjacent to each animal's line of movement .
ClawingClawing was observed in both sexes ; claw marks
were found on eight trees and were 2-3 m above the
Smith et al . : Scent marking in tigers
I
ground . A leaning tree on the territorial boundaryof two males was clawed regularly by both indi-viduals over a period of a year .
Cheek rubbingTigers usually rubbed previously sprayed trees
by bringing their cheek in contact with the urinestain or a site just above it . Based on the sequenceof tracks, we also determined that individualstypically sprayed urine after they rubbed theircheeks at a site . In 89 days two males rubbed onetree on six of 20 marking visits ; the height of thecheek rubs ranged from 1 . 20 to 2 . 16 m .
Marking and TerritorialityTo examine the relation between marking and
territorial behaviour, we asked the following ques-tions : (I) is marking concentrated in contact zonesor is it distributed randomly throughout a terri-tory, (2) do tigers of the same sex mark in responseto each other, and (3) does the frequency ofmarking increase during the establishment of aterritory?
Marking in contact zonesWe recorded five situations where the frequency
of marking was higher in contact zones than inadjacent areas.
Case 1 . Marking occurred frequently along astretch of road that formed a contact zone betweenthe territories of F115 and Fl 18 (Fig . 1) . During 5weeks, 80 scrape marks were recorded within a 520-m section of road in the boundary area . Six weekslater we recorded 105 scrapes in this same area . Incontrast, only two marks were encountered in theadjacent 520-m section located to the west of thecontact zone .
Case 2 . When two dispersing males, M 110 andM 117, became localized next to each other theboundary between their territories coincided with anorth-south forest road (Smith 1984) . Althoughwe found 46 scrape marks in a 1 . 3-km stretch of thiscontact zone (Fig. 2), there were no marks alongthe intersecting east-west road .
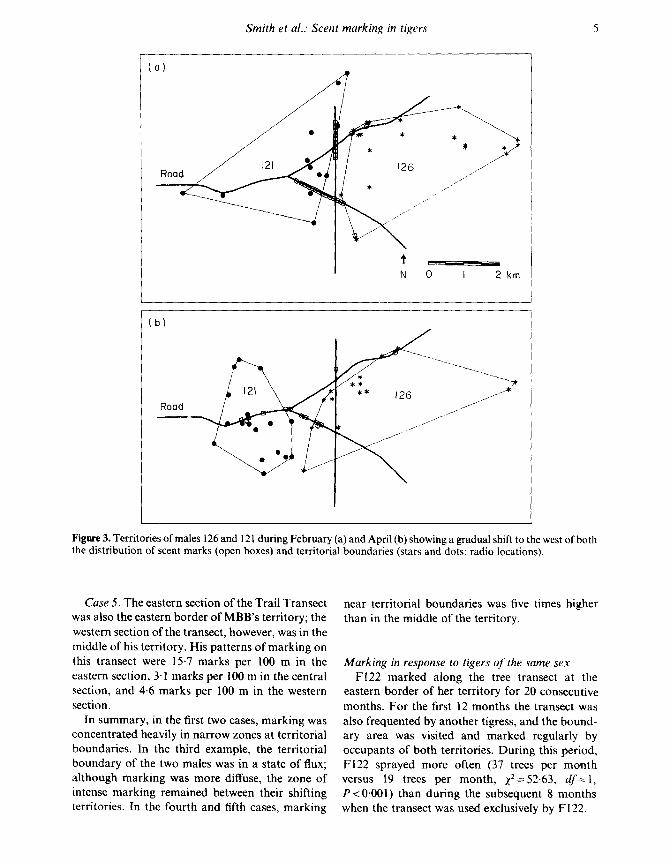
Case 3. Between November 1979 and April 1980 .dispersing M121 became localized, adjacent andpartly within the area where M126 had begun toestablish a territory during the monsoon (July-August). In November, the month that M121arrived, there was considerable overlap of theirradio locations, but from December through Maytheir ranges were mutually exclusive. During thisperiod, their territorial boundary gradually shifted

4
Animal Behaviour, 37, 1
Figure 1 . Territories of females 115 and 118 showing zone of concentrated marking at mutual territorial boundary (starsand dots : radio locations) .
Figure 2 . Territories of males 110 and 117 showing zone of concentrated marking along mutual territorial boundary(stars and dots : radio locations) .
westward as M126 expanded his territory . Thezone of marking, although more diffuse than theprevious cases, shifted correspondingly (Fig. 3) .
Case 4 . Marks were recorded along the TrailTransect that ran the entire longitudinal axis of theterritory of F122 . The trail was divided into aneastern section (900 m) that corresponded to hereastern territorial border, a central section (6200m), and a western section (800 m) that crossed herwestern border. F 122 marked 1 I •1 times per 100 m
in the eastern section, 9 .4 times per 100 m in thewestern section, but only 1 . 6 times per 100 m in thecentral zone. The higher incidence of marking per100 m of trail near territorial borders was areflection of the animal marking at a higher ratewhile walking in those areas . F122 marked at a rateof 4. 8 marks per km travelled on her easternborder, 4 . 4 marks per km travelled on the westernborder, and 0 . 85 marks per km travelled in thecentral part of her territory .
(b)
Road
Case 5 . The eastern section of the Trail Transectwas also the eastern border of MBB's territory ; thewestern section of the transect, however, was in themiddle of his territory . His patterns of marking onthis transect were 15 . 7 marks per 100 m in theeastern section, 3 . 1 marks per 100 m in the centralsection, and 4. 6 marks per 100 m in the westernsection .
In summary, in the first two cases, marking wasconcentrated heavily in narrow zones at territorialboundaries . In the third example, the territorialboundary of the two males was in a state of flux ;although marking was more diffuse, the zone ofintense marking remained between their shiftingterritories . In the fourth and fifth cases, marking
Smith et al. : Scent marking in tigers 5
Figure 3. Territories of males 126 and 121 during February (a) and April (b) showing a gradual shift to the west of boththe distribution of scent marks (open boxes) and territorial boundaries (stars and dots : radio locations) .
near territorial boundaries was five times higherthan in the middle of the territory .
Marking in response to tigers of the same sexF122 marked along the tree transect at the
eastern border of her territory for 20 consecutivemonths. For the first 12 months the transect wasalso frequented by another tigress, and the bound-ary area was visited and marked regularly byoccupants of both territories . During this period,F122 sprayed more often (37 trees per monthversus 19 trees per month, X 2 =52 . 63, df=1,P<0-001) than during the subsequent 8 monthswhen the transect was used exclusively by F122 .

6
Marking in relation to territorial establishmentObservations of seven individuals show that
marking was associated with territorial establish-ment. Three 35- to 37-month-old females beganmarking intensively when they settled in a specificarea and began to breed . F122 began exclusive useof an area in the eastern portion of her natal areawhen she was 36 months old . Over the next fewmonths she gradually established a territory, dis-placing her mother, FBB, from part of the latter'sterritory ; she also encroached on F1 15's territory .The contact zone between F122 and F115 was atthe tree transect, and beginning in September 1980both females visited and marked there regularly . InNovember, F115 visited the transect twice andsprayed only three trees . During this month, F122sprayed 16 trees during seven visits and inspected88 other trees . Subsequently, from Decemberthrough March, F122's visits to the transectincreased; she used it nearly twice as often as Fl 15(8 visits per month versus 4 . 3 visits per month) andmarked almost twice as many trees (48 . 8 per monthversus 24 . 8 per month) . By April 1980, F] 15stopped visiting the transect and F122 was theterritorial resident .
Another female, Fl 11, established a territory atthe end of the 1979 monsoon when she was 36months old; she occupied an area that had beenvacated by a resident female who disappearedduring the monsoon . 17111 marked intensivelyduring the next 3 months and at the end of thatperiod, 53 marks were recorded in a 492-m stretchof an animal trail she was using (one mark per9 . 3 m). At the same time she began to associatewith the resident male with whom she subsequentlycopulated .
A third female, F118, also began marking inten-sively when she began exclusive use of an area at37 months of age ; over a 6-week period she made105 marks in a 520-m section of road .
Three males followed the same pattern of inten-sive marking with the onset of territorial beha-viour. M110, M117 and M121 began markingbetween 29 and 32 months of age when they settledafter dispersal (Smith 1984) . Another male, MMB,who had disappeared at approximately 2 years ofage, returned to his natal area after an absence ofseveral months when he was approximately 35months old. Four months later he began markingin an area that became his contact zone with theadjacent resident, M 105 . In contrast, we discoveredthat young predispersal subadults marked infre-
Animal Behaviour, 37 . 1
quently. During a 10-month period, two juvenilemales together marked only 15 trees along the TreeTransect, whereas the two residents that used thearea consecutively during the same months marked17 trees .
Marking in Relation to the Oestrous Cycle
To determine if marking advertises female repro-ductive status we recorded the frequency of mark-ing by a female in relation to phases of herreproductive cycle and examined variation in malemarking behaviour in relation to the reproductivecycles of nearby females .
We classified females as being in oestrus whenthey associated with a male continuously for 1-2days or when we heard them roaring or moaningrepeatedly . Kleiman (1974) and Tilson (personalcommunication) reported that the same vocaliza-tions signal the onset of oestrus in captive tigers .Tilson (personal communication) found that theonset of repeated vocalization was correlated withblood hormone levels (Seal et al. 1985) . Severalobservations suggested that marking by femalesbecomes more frequent a few days prior to oestrusand is reduced during oestrus . For example, Fl 15marked 9 . 0 trees per visit on 23 and 25 February incontrast to her overall mean marking of 4 . 3 treesper visit . Two days later, while in association withM 123 and presumed to be in oestrus, she visited thetransect but did not mark . In another case, F122marked on 9 and 10 October, but later, while inassociation with MMB from 13 to 17 October, shemarked only on the final day . On another occasionF122 marked on 8. 9, and the morning of IINovember, but did not mark during her associ-ation with MBB on the afternoon of I I through 13November. These observations suggest that mark-ing advertises the onset of oestrus but is suspendedduring the period of association .
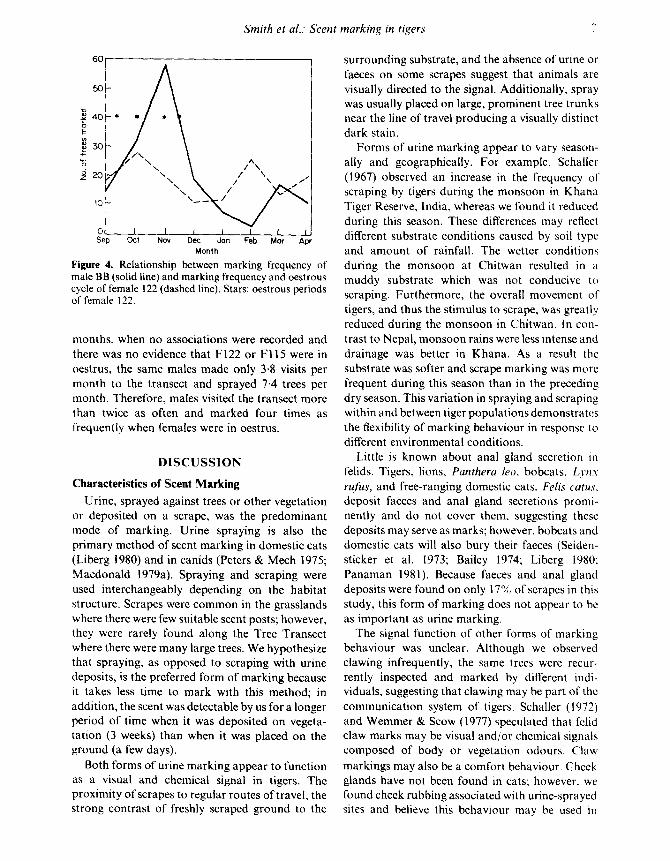
Males visited and marked the transect morefrequently when females were in oestrus than atother times. During 4 months when F122 wasobserved in association with MBB, he marked andvisited the transect more often than during the 4months when she was not in oestrus and did notassociate with him (Fig . 4) . F115 associated withM 123 twice in February 1981 and once with MSB amonth later . During those 6 months when femalesassociated with males, the three adult males made8 . 2 visits per month to the transect and sprayed31 . 7 trees per month. During the remaining 16
01111111Sep
Oct
Nov
Dec
Jan Feb Mar AprMonth
Figure 4. Relationship between marking frequency ofmale BB (solid line) and marking frequency and oestrouscycle of female 122 (dashed line) . Stars : oestrous periodsof female 122 .
months, when no associations were recorded andthere was no evidence that F 122 or F 115 were inoestrus, the same males made only 3 . 8 visits permonth to the transect and sprayed 7 . 4 trees permonth. Therefore, males visited the transect morethan twice as often and marked four times asfrequently when females were in oestrus .
DISCUSSION
Characteristics of Scent MarkingUrine, sprayed against trees or other vegetation
or deposited on a scrape, was the predominantmode of marking . Urine spraying is also theprimary method of scent marking in domestic cats(Liberg 1980) and in canids (Peters & Mech 1975 ;Macdonald 1979a) . Spraying and scraping wereused interchangeably depending on the habitatstructure . Scrapes were common in the grasslandswhere there were few suitable scent posts ; however,they were rarely found along the Tree Transectwhere there were many large trees . We hypothesizethat spraying, as opposed to scraping with urinedeposits, is the preferred form of marking becauseit takes less time to mark with this method; inaddition, the scent was detectable by us for a longerperiod of time when it was deposited on vegeta-tation (3 weeks) than when it was placed on theground (a few days) .
Both forms of urine marking appear to functionas a visual and chemical signal in tigers . Theproximity of scrapes to regular routes of travel, thestrong contrast of freshly scraped ground to the
Smith et al . : Scent marking in tigers
surrounding substrate, and the absence of urine orfaeces on some scrapes suggest that animals arevisually directed to the signal . Additionally, spraywas usually placed on large, prominent tree trunksnear the line of travel producing a visually distinctdark stain .
Forms of urine marking appear to vary season-ally and geographically . For example, Schaller(1967) observed an increase in the frequency ofscraping by tigers during the monsoon in KhanaTiger Reserve, India, whereas we found it reducedduring this season. These differences may reflectdifferent substrate conditions caused by soil typeand amount of rainfall. The wetter conditionsduring the monsoon at Chitwan resulted in amuddy substrate which was not conducive toscraping . Furthermore, the overall movement oftigers, and thus the stimulus to scrape, was greatlyreduced during the monsoon in Chitwan . In con-trast to Nepal, monsoon rains were less intense anddrainage was better in Khana. As a result thesubstrate was softer and scrape marking was morefrequent during this season than in the precedingdry season. This variation in spraying and scrapingwithin and between tiger populations demonstratesthe flexibility of marking behaviour in response todifferent environmental conditions .
Little is known about anal gland secretion infelids . Tigers, lions, Panthera leo, bobcats, Lynxrufus, and free-ranging domestic cats, Felis catus,deposit faeces and anal gland secretions promi-nently and do not cover them, suggesting thesedeposits may serve as marks; however, bobcats anddomestic cats will also bury their faeces (Seiden-sticker et al. 1973, Bailey 1974; Liberg 1980 ;Panaman 1981) . Because faeces and anal glanddeposits were found on only 17% of scrapes in thisstudy, this form of marking does not appear to beas important as urine marking .
The signal function of other forms of markingbehaviour was unclear. Although we observedclawing infrequently, the same trees were recur-rently inspected and marked by different indi-viduals, suggesting that clawing may be part of thecommunication system of tigers . Schaller (1972)and Wemmer & Scow (1977) speculated that felidclaw marks may be visual and/or chemical signalscomposed of body or vegetation odours. Clawmarkings may also be a comfort behaviour . Cheekglands have not been found in cats ; however, wefound cheek rubbing associated with urine-sprayedsites and believe this behaviour may be used in

8
intraspecific communication . Reiger (1978) sug-gested that cheek rubbing is not a form of marking,but instead speculated that this behaviour enablesthe animal to detect and learn scents of individualswith whom it comes in contact .
The function of vegetation flattening is equallyobscure . In his review of body rolling in carnivores,Reiger (1978) suggested that this behaviour mayhave a signal function, but no data are available tosupport his hypothesis .
Marking and TerritorialityOur data support the hypothesis that marking
plays a role in the establishment of territories . Burt(1943) defined territoriality as the mutually exclu-sive use of defended areas . Defence of territoriesmay include mechanisms that are direct, such asfighting, or indirect behaviour that includes chemi-cal and visual marking (Ewer 1973) . We found thatalthough direct aggression leads to territorial shiftsin tigers (Smith & McDougal, unpublished data),marking appears to be the primary means forshaping and maintaining territorial boundaries(Gosling 1986) .
Leyhausen (1965) stated that marking behaviourin felids cannot be interpreted to function as a formof territorial defence unless a potential intruder isobserved to encounter a mark and then turn back .However, Schaller (1972) argued that becauseresponses to a mark may be delayed, measuringimmediate response may not be an accurate indexof a function of a mark . We believe that immediateresponses should be expected only if the risk ofignoring a single mark is extremely high . Analternative hypothesis is that marks may communi-cate the strength of an aggression field (Waser &Wiley 1980) . Based on this idea we propose thefollowing model. At each encounter with an animalof the same sex there is some risk of injury . Theprobability of encountering another individual in agiven area is a function of the local density and ageof each individual's marks ; an animal can evaluatethe probability of encountering another individual,and ultimately the risk of potential injury based onthe strength of the local odour field . Therefore, thebenefit of using a given area is that which isachieved by incorporating an area into one'sterritory minus a cost in travel time based on thedistance from other parts of the territory and intime spent away from other resources . When theprobability of encounter and thus the risk of injury
Animal Behaviour, 37, 1
exceeds the benefit to be accrued from using anarea, an animal should cede territory without afight . When resources in another part of an indi-vidual's territory are abundant or travel is difficult,visits to and marking in an area should be reduced .Such a reduction in visits in turn signals toneighbours that the probability of an aggressiveencounter is low. This type of system should berelatively protected from cheaters ; an animal mustbe using an area frequently to establish a strongclaim (Gosling 1982, 1986) .
Pro-oestrous Marking
Our observations of a higher rate of pro-oestrouscompared to oestrous marking suggests that mark-ing during pro-oestrus signals the onset of oestrus .Choudhury (1979) observed the same pattern in atame but free-ranging tigress who invariablystopped marking during oestrus, but sprayed ex-tensively preceding its onset. Kleiman (1974) madesimilar observations on captive tigers . Shumake(1977) found that male rats, Rattus rattus, respondmost strongly to pro-oestrous urine . Similar experi-mental testing of tiger urine from pro-oestrous,oestrous and di-oestrous females would help eluci-date the role of scent marking in reproductivebehaviour .
We observed that males marked more frequentlyand at a higher rate during months when femaleswere in oestrus. Schaller (1972) noted that malelions commonly mark beside oestrous females .Perhaps males mark to advertise their exclusiverights to an oestrous female; this hypothesis issupported by our observation that most agonisticencounters between males occurred when they werein the vicinity of an oestrous female (unpublisheddata) .
A Comparison of Marking Strategies
Tigers marked along a network of trails, roads,dry streambeds and ridgetops that are used fortravel through their territories . Although markingoccurred throughout this network, it was moreintense at potential contact zones where majorroutes of travel approached territorial borders .Several studies (e .g. Peters & Mech 1975; Rothman& Mech 1979; Macdonald 1979a ; Harrington &Mech 1983) found that wolves and foxes also markextensively along the entire network of trails withintheir territories . They noted a similar increased

frequency in marking in the border area, but incontrast to tigers in Chitwan the entire boundary ofterritories was marked, creating an olfactory bowl .
Differences in patterns of marking appear to berelated to the distribution of cover . Chitwan tigersmove along regular routes between dense patchesof vegetation in which they hunt by stalking theirprey. Travel was very channelled, and marking wasconcentrated at sites where major routes of travelapproached territorial boundaries and the proba-bility of contact was maximized . This pattern ofhighly clumped signals may be a common markingpattern in other carnivores that occupy densehabitats where surveillance of entire borders isuneconomical . For example, Macdonald (1979b)reported that a group of jackals, Canis mesomelas,living in open habitat, marked the entire perimeterof their territory . In an adjacent group, whoseterritory was characterized by dense undergrowththat provided few access routes for intruders,marking was confined to major trails . This flexibi-lity in marking behaviour in response to theenvironment emphasizes the importance of com-parative intraspecific studies to an understandingof behavioural patterns and social organization .Additional research on tiger communication andterritorial behaviour needs to be conducted in moreopen habitats; such investigations will contributeto understanding the modes and distribution ofmarking and the extent to which both habitatstructure and marking influence territoriality .
ACKNOWLEDGMENTS
We are grateful to His Majesty's GovernmentDepartment of National Parks and Wildlife Con-servation for assistance and encouragementthroughout the duration of this project . We alsothank the staff of Tiger Tops Lodge for theirhospitality and help on numerous occasions . Manypeople read and provided comments on earlierdrafts of this manuscript ; we thank F . J . Cuthbert,F. W. Harrington, P . A. Jordan, D. G. Kleiman,D. F. McKinney, L. D. Mech, G. B . Schaller, C .Wemmer and an anonymous reviewer . Financialsupport for the project was provided by Smithson-ian Institution and by World Wildlife Fund-U .S .Appeal. The final manuscript was prepared whilethe senior author was supported by a grant fromthe Minnesota Agricultural Experimental Station .
Smith et al. : Scent marking in tigers
REFERENCES
Bailey, T. N. 1974 . Social organization in a bobcatpopulation . J. Wildl. Mgmt, 38,435-446 .
Brahmachary, R. L . & Dutta, J . 1981 .On the pheromonesof tigers: experiments and theory . Am. Nat., 118,561-567 .
Brown, R . E . & Macdonald, D . W. 1986 . Social Odours inMammals. Vol. 2 . Oxford: Clarendon Press .
Burt, W . H . 1943 . Territory and home range concepts asapplied to mammals . J. Mammal., 24, 346-352 .
Choudhury, S. R. 1979 . Olfaction ecology of peakcubbing in similipal tigers . Indian Forester, 103, 577-588 .
Eisenberg, J . F. & Kleiman, D. G . 1972 . Olfactorycommunication in mammals . A . Rer . Ecol . Svst ., 3, 1 -32 .
Ewer, R . F . 1973 . The Carnivores . Ithaca, New York :Cornell University Press .
Gorman, M . L . & Mills, M . G . L . 1984. Scent markingstrategies in hyaenas (Mammalia) . J . Zool., Lond., 202,535-547 .
Gosling, L . M. 1982 . A reassessment of the function ofscent marking territories . Z. Tierpsychol., 60, 89--118 .
Gosling, L . M. 1986. Economic consequences of scentmarking in mammalian territoriality . In: ChemicalSignals in Vertebrates 4 (Ed . by D. Duvall & D . Muller-Schwarze), pp. 385-395 . New York : Plenum Press .
Harrington, F. W. & Mech, L. D. 1983 . Wolf packspacing: howling as a territory independent spacingmechanism in a territorial population . Behav. EcolSociobiol ., 12, 161-168 .
Henry, J . D. 1977. The use of urine marking in thescavenging behaviour of the red fox (Vulpes vulpes)-Behaviour, 61, 82-105 .
Hornocker, M . G . 1970 . Winter territoriality in mountainlions . J. Wildl. Mgmt, 33, 457-464 .
Kleiman, D . G. 1974 . The estrous cycle in the tiger(Panthera tigris) . In : The World's Cats . Vol. 2 (Ed . byR . L . Eaton), pp . 60--75 . Seattle : Woodland Park Zoo
Leyhausen, P. 1965 . The communal organization ofsolitary mammals . Symp. Zool . Soc . Lond., 14, 249 -263 .
Liberg, O . 1980 . Spacing behavior in a population of ruralfree roaming domestic cats . Oikos, 35, 336-349 .
Macdonald, D. W. 1979a . Some observation and fieldexperiments on the urine marking behavior of the redfox, Vulpes vulpes L. Z. Tierpsychol., 51, 1-22 .
Macdonald, D. W . 1979b . The flexible social system ofthe golden jackel, Canis aureus . Behav . Ecol. Sociobiol ,5, 17-38 .
Macdonald, D. W . 1980 . Patterns of scent marking withurine and faeces amongst carnivore communities .Symp. Zool. Soc . Lond., 45, 107-139 .
McDougal, C . W . 1977 . The Face of the Tiger . London :Rivington Books and Andre Deutsch .
Mills, M . G . L ., Gorman, M. L. & Mills . M . E . J . 1982 .The scent marking behavior of the brown hyaenaHyaena brunnes . S. Afr . J . Zoo! ., 15, 240-248 .
Mishra, H. R . 1982. The ecology and behavior of chital(Axis axis) in the Royal Chitwan National Park, Nepal .Ph .D . thesis, University of Edinburgh .
9

1 0
Muller-Schwarze, D . 1977 . Complex mammalian beha-vior and pheromone bioassay in the field . In : ChemicalSignals in Mammals (Ed. by D . Muller-Schwarze &M. M. Mozell), pp . 413-433 . New York : Plenum Press .
Muller-Schwarze, D . & Silverstein, R . M . 1979 . ChemicalSignals in Vertebrates and Aquatic Invertebrates . NewYork: Plenum Press .
Panaman, R . 1981 . Behavior and ecology of free rangingfemale farm cats (Felis catus) . Z . Tierpsychol., 56, 59-73 .
Peters, R. & Mech, D . 1975 . Scent marking in wolves . Am .Scient ., 63, 628-637 .
Rails, K . 1971. Mammalian scent marking . Science, N . Y .,171,443-449 .
Reiger, 1 . 1978 . Scent rubbing in carnivores . Carnivore, 2,17-25 .
Rothman, R . J . & Mech, L . D . 1979 . Scent-marking inlone wolves and newly formed pairs . Anim . Behav., 27,750-760 .
Schaller, G. B . 1967 . The Deer and the Tiger . Chicago :University of Chicago Press.
Schaller, G. B . 1972 . The Serengeti Lion . Chicago :University of Chicago Press .
Seal, U . S ., Plotka, E . D ., Smith, J . L . D ., Wright, F . H .,Reindl, N. J ., Taylor, R . S . & Seal, M. F. 1985 .Immunoreactive lutenizing hormone estradiol, proges-terone, testosterone, and androstenedione levels duringthe breeding season and anestrus in Siberian tigers .Biol. Reprod., 32, 361-368 .
Seidensticker, J. 1976. On the ecological separationbetween tigers and leopards . Biotropica. 8, 225-234.
Seidensticker, J., Hornocker, M . G ., Wiles, V. W. &Messick J . P . 1973 . Mountain lion social organizationin the Idaho Primitive Area . Wildl. Monogr., 60, 1-60.
Shumake, S . A. 1977 . The search for applications ofchemical signals in management . In : Chemical Signalsin Mammals (Ed. by D . Muller-Schwarze & M . M .Mozell), pp . 357-375 . New York : Plenum Press .
Animal Behaviour, 37, l
Smith, J . L . D. 1984 . Dispersal, communication, andconservation strategies for the tiger (Panthera tigris) inRoyal Chitwan National Park, Nepal . Ph.D . thesis,Uniersity of Minnesota, St Paul .
Smith, J . L . D ., McDougal, C . & Sunquist, M. E. 1987 .Female land tenure system in tigers . In : Tigers q/ theWorld: the Biology, Biopolitics, Management, andConservation of an Endangered Species (Ed. by R . L .Tilson & U . S . Seal), pp . 97 109 . Park Ridge, NewJersey : Noyes .
Smith, J . L . D., Sunquist . M. E., Tamang, K . M . & Rai,P . B . 1983 . A technique for capturing and immobilizingtigers . J. Wild. Mgmt, 47, 255 -259 .
Sunquist, M . E . 1981 . The social organization of tigers(Panthera tigris) in Royal Chitwan National Park,Nepal . Smithson . Contrite . Zool. 336, 1--99 .
Tamang, K . M. 1982 . Population characteristics of thetiger and its prey . Ph .D. thesis, Michigan State Univer-sity, East Lansing .
Waser . P. M. & Wiley, R . H. 1980 . Mechanisms andevolution of spacing in animals . In : Handbook ofBehavioral Neurobiology (Ed. by P . Marler & J. G .Yanderbergh), pp . 159-223 . New York : Plenum Press .
Wells, M . C . & Bekoff. M . 1981 . An observational studyof scent marking in coyotes . Canis latrans . Anim .Behav ., 29, 332-350 .
Wemmer, C. & Scow, K . 1977. Communication in thefelidae with emphasis on scent marking and contactmarking and contact patterns . In : How AnimalsCommunicate (Ed. by T . A. Sebeok), pp . 749-766 .Bloomington : Indiana University Press .
Whitten, W. K ., Wilson, M . C ., Wilson, S. R . . Jorgenson,J . W., Novotny, M . & Carmack, M . 1980 . Induction ofmarking behavior in wild red foxes (Vulpes vulpes) bysynthetic urinary constituents . J . Cheer . Ecol., 6,49- 55 .
(Received 20 February 1987 ; revised 3 January 1988,MS. number! A4977)




















