
Sapropel burn-down and ichnological response to late Quaternary sapropel formation in two ∼ 400 ky...
20
Sapropel burn-down and ichnological response to late Quaternary sapropel formation in two ∼ 400 ky records from the eastern Mediterranean Sea L. Löwemark a, ⁎ , Y. Lin a , H.-F. Chen b , T.-N. Yang c , C. Beier d , F. Werner d , C.-Y. Lee a , S.-R. Song a , S.-J. Kao e a Department of Geosciences, National Taiwan University, P.O. Box 13-318, Taipei 106, Taiwan ROC b Institute of Applied Geosciences, National Taiwan Ocean University, 2, Pei-Ning Road, Keelung, Taiwan 20224, ROC c Institute of Earth Sciences, Academia Sinica, P.O. Box 1-55, Nankang, Taipei 115, Taiwan ROC d Institute of Geosciences, Kiel University, Olshausenstr. 40, 24118 Kiel, Germany e Research Center for Environmental Changes, Academia Sinica, P.O. Box 1-55, Nankang, Taipei 115, Taiwan ROC Received 15 September 2005; received in revised form 25 January 2006; accepted 16 February 2006 Abstract Two approximately 400 ky long sedimentary records from the eastern Mediterranean Sea were used to study the factors determining why some sapropels are more or less completely erased while others are preserved. X-ray radiographs, bulk density, organic carbon content, Fe / Al and Mn / Al ratios were used to reconstruct initial thickness and burn-down ratios of sapropels S1 to S11. The results show that, at the low-sedimentation rate, open ocean settings studied, initial carbon content of the sapropel, as well as post-sapropel sedimentation rate and carbon flux have had only limited influence on the preservation of sapropels. In contrast, the regional position relative to the deep-water source controlling bottom water current intensity and bottom water oxygenation played a major role in the preservation of the sapropels. Furthermore, orbitally controlled variations in the intensity of subsequent summer insolation maxima also seem to play an important role in the preservation potential of sapropel layers. Subsequent strong insolation maxima resulting in thick sapropels tend to act as a lid, preventing further burn-down of the older sapropels below. The long and well-dated records also offer an opportunity to study the response of the benthic fauna to the drastic environmental changes related to the formation of sapropels. The sediment in the two cores is generally characterized by mottled burrows, with only scarce occurrences of distinct trace fossils such as Scolicia, Thalassinoides, Chondrites, and Trichichnus. The meager ichnofauna is attributed to the well-oxygenated and warm bottom waters in an oligotrophic environment typical for non-sapropel times. The sparse ichnofauna and the lack of tiered ichnocoenosis render existing models of bioturbational response to changes in bottom water oxygenation unsuitable for this kind of environments. However, observations on individual traces support earlier studies that suggest that the trace fossils Trichichnus and Chondrites are the result of chemosymbiotic activities that benefit from reducing conditions in the pore water, and that the producers of Scolicia have a preference for well-oxygenated conditions and sandy substrates. © 2006 Elsevier B.V. All rights reserved. Keywords: Bioturbation; Sapropel; Eastern Mediterranean; Pleistocene; Anoxia Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406 – 425 www.elsevier.com/locate/palaeo ⁎ Corresponding author. Tel.: +886 2 33662944. E-mail address: [email protected] (L. Löwemark). 0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2006.02.013
Transcript of Sapropel burn-down and ichnological response to late Quaternary sapropel formation in two ∼ 400 ky...

alaeoecology 239 (2006) 406–425www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Sapropel burn-down and ichnological response to late Quaternarysapropel formation in two ∼400 ky records from the eastern
Mediterranean Sea
L. Löwemark a,⁎, Y. Lin a, H.-F. Chen b, T.-N. Yang c, C. Beier d, F. Werner d,C.-Y. Lee a, S.-R. Song a, S.-J. Kao e
a Department of Geosciences, National Taiwan University, P.O. Box 13-318, Taipei 106, Taiwan ROCb Institute of Applied Geosciences, National Taiwan Ocean University, 2, Pei-Ning Road, Keelung, Taiwan 20224, ROC
c Institute of Earth Sciences, Academia Sinica, P.O. Box 1-55, Nankang, Taipei 115, Taiwan ROCd Institute of Geosciences, Kiel University, Olshausenstr. 40, 24118 Kiel, Germany
e Research Center for Environmental Changes, Academia Sinica, P.O. Box 1-55, Nankang, Taipei 115, Taiwan ROC
Received 15 September 2005; received in revised form 25 January 2006; accepted 16 February 2006
Abstract
Two approximately 400 ky long sedimentary records from the eastern Mediterranean Sea were used to study the factorsdetermining why some sapropels are more or less completely erased while others are preserved. X-ray radiographs, bulk density,organic carbon content, Fe / Al and Mn / Al ratios were used to reconstruct initial thickness and burn-down ratios of sapropels S1 toS11. The results show that, at the low-sedimentation rate, open ocean settings studied, initial carbon content of the sapropel, as wellas post-sapropel sedimentation rate and carbon flux have had only limited influence on the preservation of sapropels. In contrast,the regional position relative to the deep-water source controlling bottom water current intensity and bottom water oxygenationplayed a major role in the preservation of the sapropels. Furthermore, orbitally controlled variations in the intensity of subsequentsummer insolation maxima also seem to play an important role in the preservation potential of sapropel layers. Subsequent stronginsolation maxima resulting in thick sapropels tend to act as a lid, preventing further burn-down of the older sapropels below. Thelong and well-dated records also offer an opportunity to study the response of the benthic fauna to the drastic environmentalchanges related to the formation of sapropels.
The sediment in the two cores is generally characterized by mottled burrows, with only scarce occurrences of distinct tracefossils such as Scolicia, Thalassinoides, Chondrites, and Trichichnus. The meager ichnofauna is attributed to the well-oxygenatedand warm bottom waters in an oligotrophic environment typical for non-sapropel times. The sparse ichnofauna and the lack oftiered ichnocoenosis render existing models of bioturbational response to changes in bottom water oxygenation unsuitable for thiskind of environments. However, observations on individual traces support earlier studies that suggest that the trace fossilsTrichichnus and Chondrites are the result of chemosymbiotic activities that benefit from reducing conditions in the pore water, andthat the producers of Scolicia have a preference for well-oxygenated conditions and sandy substrates.© 2006 Elsevier B.V. All rights reserved.
Keywords: Bioturbation; Sapropel; Eastern Mediterranean; Pleistocene; Anoxia
⁎ Corresponding author. Tel.: +886 2 33662944.E-mail address: [email protected] (L. Löwemark).
0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2006.02.013

407L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
1. Introduction
One of the most prominent features of Mediterra-nean sediments is the periodical occurrence of cm to mthick, organic rich, black sediment layers intercalatedin the organic-carbon poor, reddish brown pelagicsediments. These organic-rich layers are defined assapropels when their thickness exceeds 1 cm and thetotal organic carbon (TOC) content exceeds 2% (Kiddet al., 1978). In many cases the original organic-richlayer has been partially or completely oxidized due tosubsequently increased bottom water oxygen levels, aprocess often referred to as burn-down. Dark sedimentlayers where the TOC level is below 2% usually arereferred to as sapropelic layers whereas completelyburn-downed sapropels, only detectable through X-rayradiographs and geochemistry, are referred to as“ghost” sapropels. The formation of sapropels hasattracted much attention over the years, and twodifferent models have been proposed. Initial worksfavored the stagnation model (e.g., Olausson, 1961;Cita et al., 1977; Vergnaud-Grazzini et al., 1977;Thunell and Williams, 1989; Aksu et al., 1995), whichpostulates that a decreased or halted deep waterformation, due to stratification of the water column,led to anoxic conditions at the sea floor and thepreservation of organic material. A second model, theproductivity model (e.g., De Lange and ten Haven,1983; Pedersen and Calvert, 1990; Castradori, 1993;Sancetta, 1994), proposes elevated primary productiv-ity due to enhanced nutrient availability in the photiczone as the primary factor contributing to the increasedcarbon content in the sapropel layers. Recent studiestend to favor a combination of the two processes (e.g.,Rohling and Gieskes, 1989; Rohling, 1994; Emeis etal., 2000a,b; Warning and Brumsack, 2000). Thesapropel formation in the eastern Mediterranean isbelieved to be linked to variations in the intensity ofthe African monsoon. During Earth axis precessionminima, when summer insolation in the northernhemisphere reached its maximum, the increasedmonsoonal precipitation over northern Africa led toenhanced riverine fresh water input to the Mediterra-nean Sea (e.g., Rossignol-Strick et al., 1982; Ros-signol-Strick, 1999). The increased fresh water inputlowered the salinity of the surface waters causing thethermohaline ventilation of the deeper parts of theMediterranean basin to slow down or even stop,leading to anoxic bottom water conditions ideal for thepreservation of organic material. At the same time, theincreased riverine input combined with a shift of thenutricline into the photic zone led to enhanced primary
productivity and therefore an increased export oforganic material to the sea floor (Rohling and Gieskes,1989; Rohling, 1991; Corselli et al., 2002). Althoughthere is consensus about the overall understanding ofthe sapropel formation process, there are still dis-crepancies between the individual records and uncer-tainties in terms of onset of the sapropel formationconditions, sapropel thickness and preservation, aswell as variations in paleocirculation patterns duringsapropel formation.
The aim of this study is to examine the mechanismscontrolling the preservation of the individual sapropelsfound in two gravity cores from the eastern Mediterra-nean Sea with respect to the questions: why did somesapropels survive re-oxygenation while others weremore or less completely erased, and which factorsgovern the differences in sapropel development andpreservation between different sites? The two coressituated in different depths and positions relative to thedeep water source region offer a possibility to study theinfluence of local and regional factors on sapropeldevelopment, burn-down, and variations in preservationbetween different sapropels as well as between differentlocations.
The two eastern Mediterranean cores studied, whichcomprise approximately 4 glacial–interglacial cyclesand both contain well preserved, partially oxidized, aswell as completely erased sapropels, furthermore offer aunique opportunity to compare the observed changes intrace fossil associations with different ichnologicalmodels. The well defined age models and the wellknown environmental conditions also make these twocores ideally suited to study how changes in bottomwater circulation and oxygenation affected individualtrace fossil. These observations might form valuableanalogs for how the benthic fauna responded to anoxiaduring black shale formation.
2. Hydrographic setting
The Mediterranean Sea is a semi-enclosed basin withits own thermohaline circulation, making it a naturallaboratory to study how climatic changes influenceocean circulation and sedimentation patterns. Thedifferent climatic zones surrounding the basin makethe Mediterranean particularly susceptible to climaticvariations. Late Cenozoic sediment from the easternMediterranean Sea typically contains rhythmicallydistributed, conspicuous dark layers of organic-richsediment referred to as sapropels. Today, the deepwaters of the Mediterranean form by a series ofprocesses starting with the inflow of Atlantic surface

408 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
water. This inflow is driven by the water deficit causedby the excess evaporation of the warm Mediterraneansurface waters. As the Atlantic surface water moveseastward, evaporation drastically increases its salinity.The density increase gained by the evaporation isoffset in summer by high water temperatures prevent-ing the formation of deep waters. Only during winter,when cold winds from the Anatolian mountains coolthe surface waters in the easternmost part of theMediterranean, are the surface waters sufficientlycooled to sink down. The water mass formed duringthis process is called the Levantine Intermediate Water(LIW) and forms a layer from about 200 to 600 mwater depth. The LIW then spreads into both theeastern and the western Mediterranean. During winter,cold winds from the northern hinterland cool thesurface waters in the northern basins (Gulf of Lion,Aegean and Adriatic Seas) enough to obliterate thedensity contrast with the underlying LIW. As thesewater masses mix, heavy, saline waters are generatedthat are dense enough to form true deep waters, theEastern and Western Mediterranean Deep Waters,respectively (e.g., Pinardi and Masetti, 2000; Rohling,2001; Tomczak and Godfrey, 2002). The easternMediterranean deep water typically has a salinity ofaround 38.7, an oxygen content around 5 ml/l and atemperature above 12 °C (Roether and Well, 2001;Tomczak and Godfrey, 2002). These deep watersfinally leave the Mediterranean together with portionsof the LIW through the Gibraltar Strait (Minas et al.,1991) where they enter the North Atlantic Ocean as asalt tongue detectable as far north as Ireland (Reid,1979) and as far west as the Caribbean Sea (Haddadand Droxler, 1996). The rapid deep water formationwith an overturn time of only about 100 years keepsthe bottom waters of the Mediterranean well oxygen-
Fig. 1. Bathymetric chart showing the Mediterranean
ated despite their higher oxygen demand resultingfrom the relatively high bottom water temperatures(Lacombe and Tchernia, 1972; Lacombe et al., 1981;Rohling, 2001). Little is known about the effect oflowered sea level on the deep water circulation of theMediterranean Sea. Especially the sapropels thatformed during times of low sea-level stand, such asS2 and S6, are interesting as the low sea level at thesetimes likely restricted the exchange through theGibraltar and Sicily straits and also drastically reducedthe size of the Adriatic Sea, today of major importanceas a source of Eastern Mediterranean deep water.
3. Materials and methods
3.1. Laboratory analysis
Sedimentological records and X-ray radiographsfrom two gravity cores from the eastern MediterraneanSea recovered during METEOR cruise M40/1 in 1997(Hieke et al., 1999) are used in this study. The core M40/1-10 (34°45.8′N, 19°45.7′E, 2972 m) was taken on thewestern flank of the Mediterranean Ridge about 300 kmto the west of Crete. Core M40/1-22 (33°39.5′N,24°41.2′E, 2004 m) was taken in the Olimpi area onthe southern flank of the Mediterranean Ridge, about120 km south of Crete (Fig. 1). X-ray radiographs forthe study of morphologic features of the traces wereproduced according to the method described by Bouma(1964), Werner (1967), and Löwemark and Werner(2001). Plastic boxes 25×10×1 cm were pushed intothe sediment, cut out with a nylon string and placed invacuum-sealed plastic pouches. The sediment slabswere placed directly on the X-ray film bags and exposedto 35 kV for 10–15 min. Total carbon content (TC) andtotal organic carbon content (TOC) were determined
Sea and the core locations in the eastern basin.
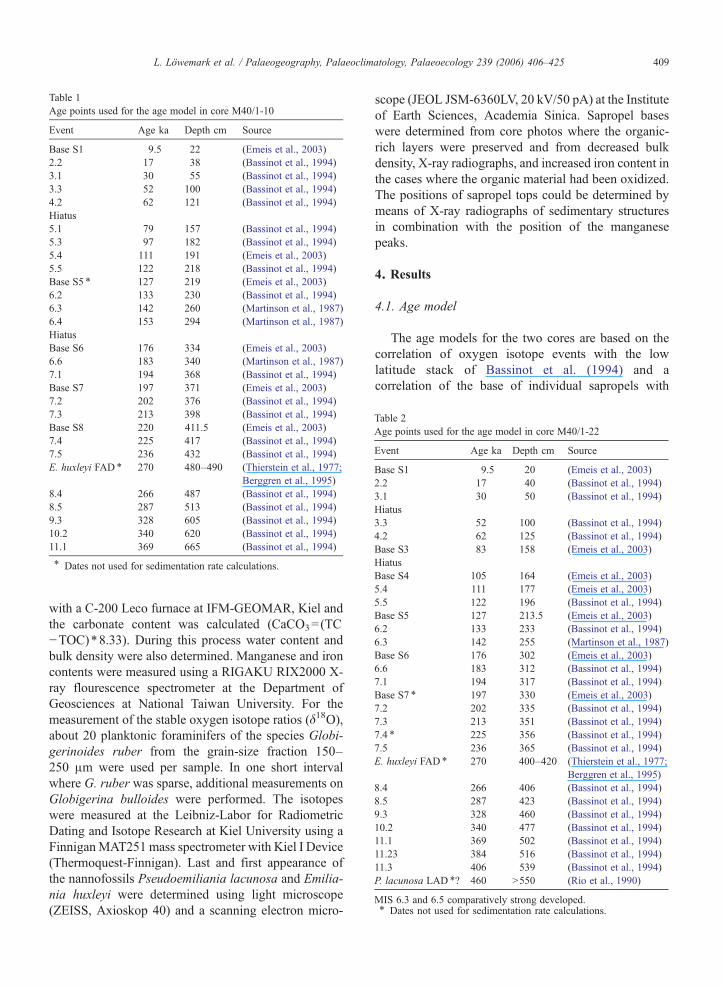
Table 1Age points used for the age model in core M40/1-10
Event Age ka Depth cm Source
Base S1 9.5 22 (Emeis et al., 2003)2.2 17 38 (Bassinot et al., 1994)3.1 30 55 (Bassinot et al., 1994)3.3 52 100 (Bassinot et al., 1994)4.2 62 121 (Bassinot et al., 1994)Hiatus5.1 79 157 (Bassinot et al., 1994)5.3 97 182 (Bassinot et al., 1994)5.4 111 191 (Emeis et al., 2003)5.5 122 218 (Bassinot et al., 1994)Base S5 ⁎ 127 219 (Emeis et al., 2003)6.2 133 230 (Bassinot et al., 1994)6.3 142 260 (Martinson et al., 1987)6.4 153 294 (Martinson et al., 1987)HiatusBase S6 176 334 (Emeis et al., 2003)6.6 183 340 (Martinson et al., 1987)7.1 194 368 (Bassinot et al., 1994)Base S7 197 371 (Emeis et al., 2003)7.2 202 376 (Bassinot et al., 1994)7.3 213 398 (Bassinot et al., 1994)Base S8 220 411.5 (Emeis et al., 2003)7.4 225 417 (Bassinot et al., 1994)7.5 236 432 (Bassinot et al., 1994)E. huxleyi FAD ⁎ 270 480–490 (Thierstein et al., 1977;
Berggren et al., 1995)8.4 266 487 (Bassinot et al., 1994)8.5 287 513 (Bassinot et al., 1994)9.3 328 605 (Bassinot et al., 1994)10.2 340 620 (Bassinot et al., 1994)11.1 369 665 (Bassinot et al., 1994)
⁎ Dates not used for sedimentation rate calculations.
Table 2Age points used for the age model in core M40/1-22
Event Age ka Depth cm Source
Base S1 9.5 20 (Emeis et al., 2003)2.2 17 40 (Bassinot et al., 1994)3.1 30 50 (Bassinot et al., 1994)Hiatus3.3 52 100 (Bassinot et al., 1994)4.2 62 125 (Bassinot et al., 1994)Base S3 83 158 (Emeis et al., 2003)HiatusBase S4 105 164 (Emeis et al., 2003)5.4 111 177 (Emeis et al., 2003)5.5 122 196 (Bassinot et al., 1994)Base S5 127 213.5 (Emeis et al., 2003)6.2 133 233 (Bassinot et al., 1994)6.3 142 255 (Martinson et al., 1987Base S6 176 302 (Emeis et al., 2003)6.6 183 312 (Bassinot et al., 1994)7.1 194 317 (Bassinot et al., 1994)Base S7 ⁎ 197 330 (Emeis et al., 2003)7.2 202 335 (Bassinot et al., 1994)7.3 213 351 (Bassinot et al., 1994)7.4 ⁎ 225 356 (Bassinot et al., 1994)7.5 236 365 (Bassinot et al., 1994)E. huxleyi FAD ⁎ 270 400–420 (Thierstein et al., 1977;
Berggren et al., 1995)8.4 266 406 (Bassinot et al., 1994)8.5 287 423 (Bassinot et al., 1994)9.3 328 460 (Bassinot et al., 1994)10.2 340 477 (Bassinot et al., 1994)11.1 369 502 (Bassinot et al., 1994)11.23 384 516 (Bassinot et al., 1994)11.3 406 539 (Bassinot et al., 1994)P. lacunosa LAD ⁎? 460 >550 (Rio et al., 1990)
MIS 6.3 and 6.5 comparatively strong developed.⁎ Dates not used for sedimentation rate calculations.
409L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
with a C-200 Leco furnace at IFM-GEOMAR, Kiel andthe carbonate content was calculated (CaCO3=(TC−TOC)⁎8.33). During this process water content andbulk density were also determined. Manganese and ironcontents were measured using a RIGAKU RIX2000 X-ray flourescence spectrometer at the Department ofGeosciences at National Taiwan University. For themeasurement of the stable oxygen isotope ratios (δ18O),about 20 planktonic foraminifers of the species Globi-gerinoides ruber from the grain-size fraction 150–250 μm were used per sample. In one short intervalwhere G. ruber was sparse, additional measurements onGlobigerina bulloides were performed. The isotopeswere measured at the Leibniz-Labor for RadiometricDating and Isotope Research at Kiel University using aFinniganMAT251 mass spectrometer with Kiel I Device(Thermoquest-Finnigan). Last and first appearance ofthe nannofossils Pseudoemiliania lacunosa and Emilia-nia huxleyi were determined using light microscope(ZEISS, Axioskop 40) and a scanning electron micro-
scope (JEOL JSM-6360LV, 20 kV/50 pA) at the Instituteof Earth Sciences, Academia Sinica. Sapropel baseswere determined from core photos where the organic-rich layers were preserved and from decreased bulkdensity, X-ray radiographs, and increased iron content inthe cases where the organic material had been oxidized.The positions of sapropel tops could be determined bymeans of X-ray radiographs of sedimentary structuresin combination with the position of the manganesepeaks.
4. Results
4.1. Age model
The age models for the two cores are based on thecorrelation of oxygen isotope events with the lowlatitude stack of Bassinot et al. (1994) and acorrelation of the base of individual sapropels with
)
410 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
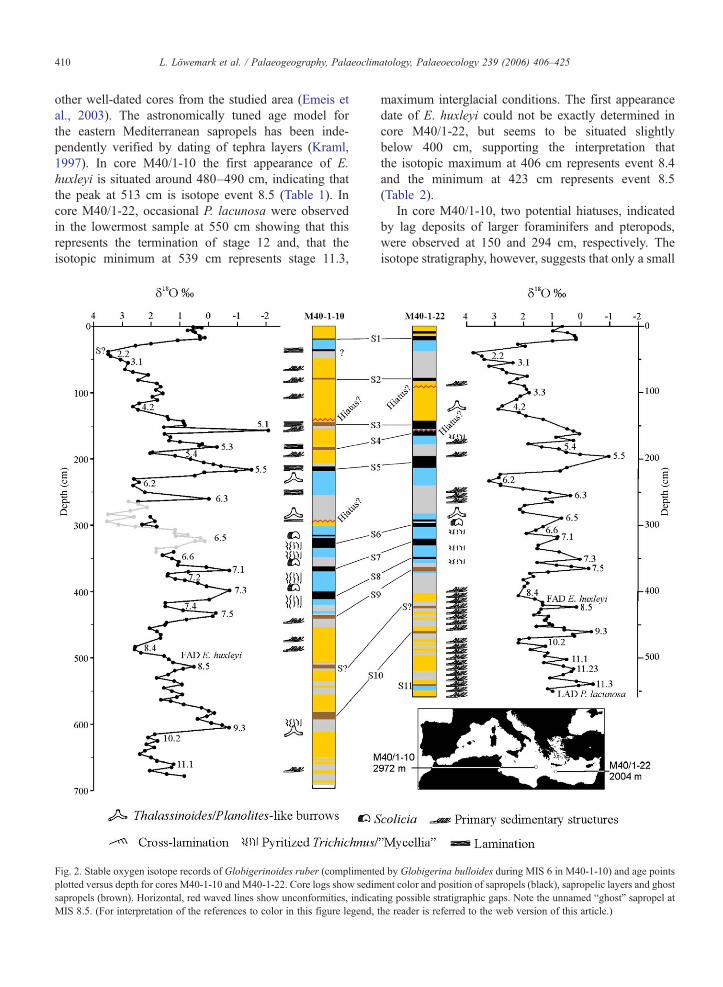
other well-dated cores from the studied area (Emeis etal., 2003). The astronomically tuned age model forthe eastern Mediterranean sapropels has been inde-pendently verified by dating of tephra layers (Kraml,1997). In core M40/1-10 the first appearance of E.huxleyi is situated around 480–490 cm, indicating thatthe peak at 513 cm is isotope event 8.5 (Table 1). Incore M40/1-22, occasional P. lacunosa were observedin the lowermost sample at 550 cm showing that thisrepresents the termination of stage 12 and, that theisotopic minimum at 539 cm represents stage 11.3,
Fig. 2. Stable oxygen isotope records of Globigerinoides ruber (complimenteplotted versus depth for cores M40-1-10 and M40-1-22. Core logs show sedimsapropels (brown). Horizontal, red waved lines show unconformities, indicaMIS 8.5. (For interpretation of the references to color in this figure legend,
maximum interglacial conditions. The first appearancedate of E. huxleyi could not be exactly determined incore M40/1-22, but seems to be situated slightlybelow 400 cm, supporting the interpretation thatthe isotopic maximum at 406 cm represents event 8.4and the minimum at 423 cm represents event 8.5(Table 2).
In core M40/1-10, two potential hiatuses, indicatedby lag deposits of larger foraminifers and pteropods,were observed at 150 and 294 cm, respectively. Theisotope stratigraphy, however, suggests that only a small
d by Globigerina bulloides during MIS 6 in M40-1-10) and age pointsent color and position of sapropels (black), sapropelic layers and ghost
ting possible stratigraphic gaps. Note the unnamed “ghost” sapropel atthe reader is referred to the web version of this article.)

411L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
amount of sediment is missing, while in core M40/1-22,the sediment between S4 and S3 has been largelyremoved by an erosional event. Assuming a duration ofabout 6 ky for sapropel S4 (Emeis et al., 2003), the ageof the remaining sediment just below the hiatus wouldbe approximately 99 ka, leaving a hiatus of 16 ky.Another possible hiatus is located just below S2 in the
Table 3Main ichnogenera found in the studied cores
Planolites Nicholson 1873Unlined, straight to gently curved, ±sediments. ∅=few mm to slightly m(Häntzschel, 1975; Pemberton and Frey(Wetzel, 1991; Bromley, 1996). The pinterpreted to indicate well-oxygenatedburrows point to lowered oxygen levels
After Wetzel (2002)
Nereites MacLeay in Murchison 1839“Selectively preserved, curved, windinhorizontal trails, consisting of a mediansediment” (Mangano et al., 2000). In mowell-oxidized sediment (Wetzel, 2002).
Thalassinoides Ehrenberg 1944Regularly branching cylindrical burroventilation shafts, ∅=1 to 20 cm, (Hänfeeding crustaceans. Thalassinoides haswaters but is essentially absent where d
After Bromley (1996).
Scolicia De Quatrefages 1849Scolicia is a 2–5 cm thick, unbranched,in longitudinal view and oval outline oftandWerner, 2000). Scolicia is one of theechinoids have been found at the end oproducers have been found to show asedimentation rates (Wetzel, 1991; Fuechinoids point to a preference for oxic
After Seilacher (1957)
Chondrites Sternberg 1833Consists of a root-like system of brancsurface (Häntzschel, 1975). When occChondrites most likely represents a c(Bromley and Ekdale, 1984; Savrda anmassively and more or less excludingoxygenated bottom waters (Ekdale et a
Trichichnus Frey 1970 and “Mycellia”Trichichnus (T) typically are vertically oseveral tens of cm in length and with diasediments generally pyritized below adiscernible in radiographs.“Mycellia” (M), not a formally namedcentimeters in length and tenths of mm tare pyritized below a certain sediment dVariations in abundance of these pyritiz(Thomsen and Vorren, 1984). Increasedoxygen levels, whereas periods characnumber of pyritized burrows.
same core but the isotope stratigraphy indicates minimalsediment removal (Fig. 2).
4.2. Sediment structures and composition
The cores consist of reddish-brown, light gray, orbluish gray hemipelagic muds with several intercalated
horizontally orientated burrow, infill differs from the surroundingore than 1 cm. Represents the active backfill of a deposit feeder, 1982; Keighley and Pickerill, 1995). Primarily occupies a middle tierresence of abundant large Planolites in shallow to middle tiers isbottom waters. In contrast, low abundances and small Planolitesin the bottom waters.
g to regularly meandering or spiralling, unbranched, predominantlybackfilled tunnel enveloped by an even to lobate zone of reworkeddern sediment from the South China Sea, Nereites occur exclusively in
ws forming horizontal networks connected to the surface throughtzschel, 1975; Bromley, 1996). Presumably dwelling burrows of filterbeen found in sediments overlain by oxic to slightly dysoxic bottomysoxic or anoxic conditions prevail (Bromley, 1996).
and mostly horizontal burrow characterized by thick meniscate backfillen with a pair of ridges at the base in transverse view (Wetzel, 1991; Fufew traces of which the trace maker is known; in several cases irregularf the burrows (e.g. Reineck, 1968; Bromley and Asgaard, 1975). Thepreference for well sorted coarse silts to fine sands and rather highand Werner, 2000). The shallow tier and the calcified skeleton of theenvironments.
hing tubes extending from a vertical tube connected to the sedimentupying the deepest tier, penetrating down into dysoxic sediments,ase of equilibrium specialization under well-oxygenated conditionsd Bottjer, 1986; Fu, 1991; Bromley, 1996). However, when appearingother traces from the fabric it is considered an indicator of poorlyl., 1984; Ekdale and Mason, 1988; Fu, 1991; Bromley, 1996).
riented cylindrical, thread-like, and sometimes branched burrows, up tometers normally less than 1 mm (Häntzschel, 1975). In modern marinesediment depth of 10–20 cm, and are therefore particularly well
trace fossil, are small, randomly orientated filaments up to severalhick (Blanpied and Bellaiche, 1981). Like Trichichnus, “Mycellia” alsoepth.ed burrows have been noticed to correspond to environmental changesnumbers of pyritized burrows during periods of reduced bottom waterterized by higher bottom-water oxygen levels displayed a decreased
412 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
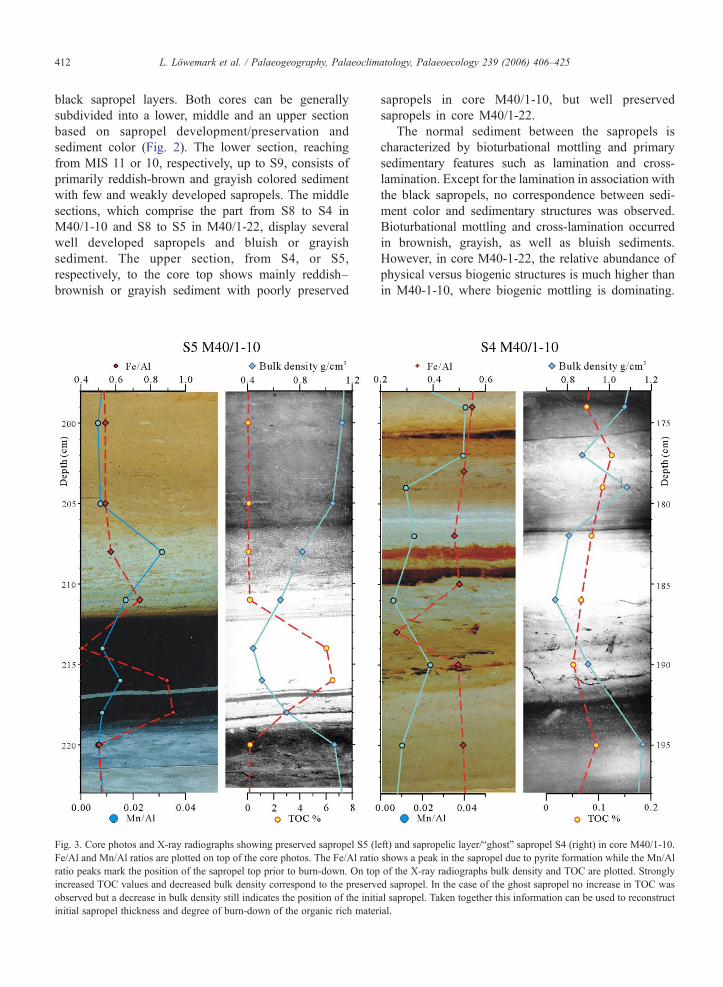
black sapropel layers. Both cores can be generallysubdivided into a lower, middle and an upper sectionbased on sapropel development/preservation andsediment color (Fig. 2). The lower section, reachingfrom MIS 11 or 10, respectively, up to S9, consists ofprimarily reddish-brown and grayish colored sedimentwith few and weakly developed sapropels. The middlesections, which comprise the part from S8 to S4 inM40/1-10 and S8 to S5 in M40/1-22, display severalwell developed sapropels and bluish or grayishsediment. The upper section, from S4, or S5,respectively, to the core top shows mainly reddish–brownish or grayish sediment with poorly preserved
Fig. 3. Core photos and X-ray radiographs showing preserved sapropel S5 (lFe/Al and Mn/Al ratios are plotted on top of the core photos. The Fe/Al ratioratio peaks mark the position of the sapropel top prior to burn-down. On topincreased TOC values and decreased bulk density correspond to the preservobserved but a decrease in bulk density still indicates the position of the initiinitial sapropel thickness and degree of burn-down of the organic rich mater
sapropels in core M40/1-10, but well preservedsapropels in core M40/1-22.
The normal sediment between the sapropels ischaracterized by bioturbational mottling and primarysedimentary features such as lamination and cross-lamination. Except for the lamination in association withthe black sapropels, no correspondence between sedi-ment color and sedimentary structures was observed.Bioturbational mottling and cross-lamination occurredin brownish, grayish, as well as bluish sediments.However, in core M40-1-22, the relative abundance ofphysical versus biogenic structures is much higher thanin M40-1-10, where biogenic mottling is dominating.
eft) and sapropelic layer/“ghost” sapropel S4 (right) in core M40/1-10.shows a peak in the sapropel due to pyrite formation while the Mn/Alof the X-ray radiographs bulk density and TOC are plotted. Strongly
ed sapropel. In the case of the ghost sapropel no increase in TOC wasal sapropel. Taken together this information can be used to reconstructial.

413L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
Both colors and sedimentary structures typically vary ona cm to dm scale.
4.3. Biogenic structures
The biogenic structures observed in the cores includemottled intervals, a few intervals dominated by largerThalassinoides–Planolites-like burrows, pyritized Tri-chichnus, and some isolated Scolicia and Chondrites(see Table 3). The traces in the mottled intervals in theupper part of M40/1-10 do show some similarities withthe modern Nereites described by Wetzel (2002) fromthe South China Sea. It is interesting to notice, that ashort interval containing Scolicia is situated just aboveS6 in both cores (Fig. 2). In core M40/1-10, Scolicia isalso found directly above sapropel layers S7 and S8. Inboth cores the sediment containing the Scolicia ischaracterized by elevated sand content (not shown),although microscopic evaluation of the sand fractionsuggest that the increased sand content is partly due tosecondary pyrite particles and do not necessarily reflectsediment conditions at the time of trace production. Thesegment between S6 and S5, corresponding to the MIS6.4 stadial, also contains an interval with a large numberof Thalassinoides–Planolites-like burrows (0.5–2 cm indiameter) in both cores. Thalassinoides–Planolites-likeburrows are also found in M40/1-10 below S10, in aninterval corresponding to interglacial stage MIS 9.3. Incore M40/1-10 a few small Chondrites were observed topenetrate down into laminated layers and sapropels. Inone case (M40/1-10, S10) Chondrites was founddirectly below the sapropelic layer, suggesting thatChondrites, which is known to be resistant to dysoxicconditions (e.g., Bromley and Ekdale, 1984; Fu andWerner, 1994), was the last trace in the sediment beforeanoxic bottom waters expelled all life. A large numberof pyritized microburrows, presumably Trichichnus and“Mycellia” (terminology of Blanpied and Bellaiche(1981), Wetzel (1983a), Werner (Werner, 1985), andBaldwin and McCave (1999)), occur in association withthe sapropel layers. Most Trichichnus are pyritized, butunpyritized burrows also occur. Other identifiable tracesare scarce. It may be noted that, contrary to what mightbe intuitively expected, the Fe/Al ratio does not increasein intervals with large numbers of pyritized burrows,indicating that these contribute little to the overall ironcontent.
4.4. Sapropels
The black sapropel layers show up clearly as layersof less dense material in the X-ray radiographs, as peaks
of strongly increased TOC, and as decreased bulkdensity. The sapropels are generally also characterizedby an increase in the relative Fe / Al and Mn / Al ratios(Fig. 3). Whereas the iron peak is related to pyriteformation in the sapropel, the manganese peak is usuallysituated above the visible black layers and is related tothe reoxygenation after the anoxic conditions, whichresulted in precipitation of dissolved manganese fromthe water column (Higgs et al., 1994; van Santvoort etal., 1996; Jung et al., 1997). These features allow therecognition of erased or oxidized sapropels, so called“ghost” sapropels that are not visible in the fresh cores(Fig. 3).
The oldest sapropel recovered by the cores is S11,which is visible in M40/1-22, but is not included in coreM40/1-10. Sapropels S10 and S9 are not visible in thecores but their positions can be inferred from X-rayradiographs, bulk density, Fe/Al, and Mn/Al ratios.Additionally, in association with MIS 8.5, bulk densityand Fe/Al and Mn/Al ratios indicate the remains of asapropelic layer (Fig. 4), although neither the fresh coresnor X-ray radiographs show any sign of increasedorganic material. Sapropels S8 to S5 are clearly visiblein both cores and show up in X-ray radiographs, TOC,bulk density, and by their Fe/Al ratios. In contrast, theMn/Al ratios only show small peaks in relation to thesesapropels. In the upper part of the core, containingsapropels S4 to S1, a larger difference between the twocores has been found. In M40/1-10, sapropels S4 and S3are not visible in the core, and S2 is only weakly visibleas a reworked rust brown layer. However, X-rayradiographs and measured sediment geochemical para-meters (Fig. 4) clearly indicate their positions. Incontrast, in M40/1-22, S4, S3 and S1 show up as thickblack layers with TOC content around 2% and distinctpeaks in bulk density and Fe/Al and Mn/Al ratios.Sapropel S2 appears as a thin brownish layer with TOCaround 2% and a small but distinct Mn-peak a few cmabove the sapropel. Core M40/1-22 generally displaysmore preserved primary sedimentary structures, espe-cially in the lowermost section up to below S9. In bothcores, there is a close correlation between isotopicallylight events and sapropels; most light peaks in theisotopic record are found to correspond to sapropels orghost sapropels, exceptions are the relatively smallpeaks MIS 3.3, MIS 6.3, MIS 11.1, and MIS 11.23.
4.5. Sapropel burn-down
Not only are some sapropels preserved and some aremore or less oxidized, there is also a large difference inthe total original thickness of the individual sapropels as

414 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
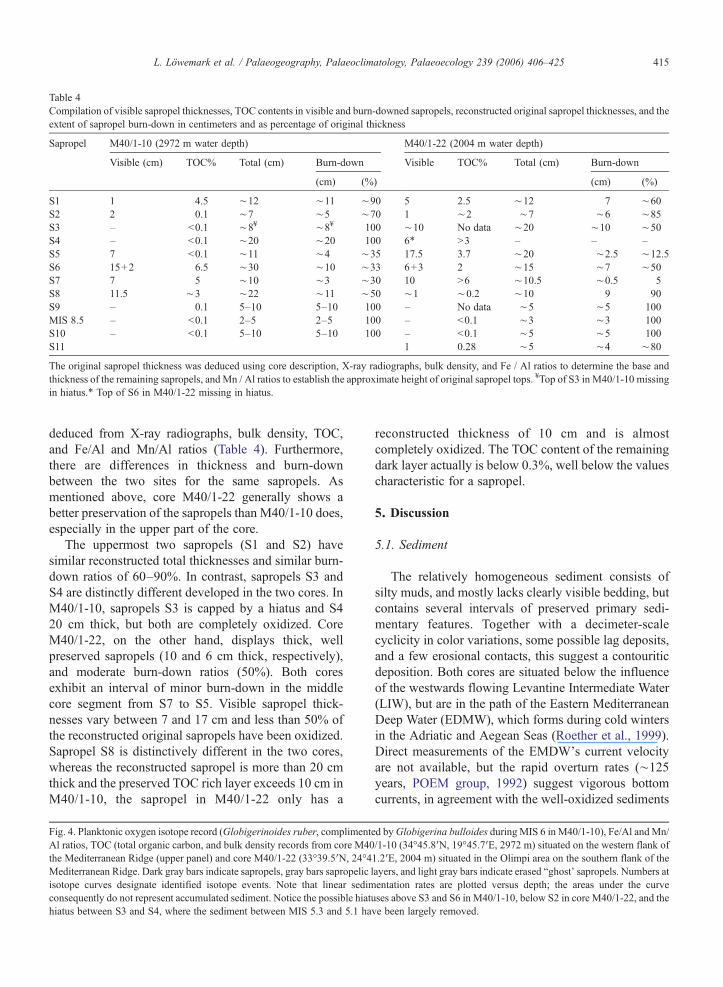
Table 4Compilation of visible sapropel thicknesses, TOC contents in visible and burn-downed sapropels, reconstructed original sapropel thicknesses, and theextent of sapropel burn-down in centimeters and as percentage of original thickness
Sapropel M40/1-10 (2972 m water depth) M40/1-22 (2004 m water depth)
Visible (cm) TOC% Total (cm) Burn-down Visible TOC% Total (cm) Burn-down
(cm) (%) (cm) (%)
S1 1 4.5 ∼12 ∼11 ∼90 5 2.5 ∼12 7 ∼60S2 2 0.1 ∼7 ∼5 ∼70 1 ∼2 ∼7 ∼6 ∼85S3 – <0.1 ∼8¥ ∼8¥ 100 ∼10 No data ∼20 ∼10 ∼50S4 – <0.1 ∼20 ∼20 100 6⁎ >3 – – –S5 7 <0.1 ∼11 ∼4 ∼35 17.5 3.7 ∼20 ∼2.5 ∼12.5S6 15+2 6.5 ∼30 ∼10 ∼33 6+3 2 ∼15 ∼7 ∼50S7 7 5 ∼10 ∼3 ∼30 10 >6 ∼10.5 ∼0.5 5S8 11.5 ∼3 ∼22 ∼11 ∼50 ∼1 ∼0.2 ∼10 9 90S9 – 0.1 5–10 5–10 100 – No data ∼5 ∼5 100MIS 8.5 – <0.1 2–5 2–5 100 – <0.1 ∼3 ∼3 100S10 – <0.1 5–10 5–10 100 – <0.1 ∼5 ∼5 100S11 1 0.28 ∼5 ∼4 ∼80
The original sapropel thickness was deduced using core description, X-ray radiographs, bulk density, and Fe / Al ratios to determine the base andthickness of the remaining sapropels, and Mn / Al ratios to establish the approximate height of original sapropel tops. ¥Top of S3 in M40/1-10 missingin hiatus.⁎ Top of S6 in M40/1-22 missing in hiatus.
415L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
deduced from X-ray radiographs, bulk density, TOC,and Fe/Al and Mn/Al ratios (Table 4). Furthermore,there are differences in thickness and burn-downbetween the two sites for the same sapropels. Asmentioned above, core M40/1-22 generally shows abetter preservation of the sapropels than M40/1-10 does,especially in the upper part of the core.
The uppermost two sapropels (S1 and S2) havesimilar reconstructed total thicknesses and similar burn-down ratios of 60–90%. In contrast, sapropels S3 andS4 are distinctly different developed in the two cores. InM40/1-10, sapropels S3 is capped by a hiatus and S420 cm thick, but both are completely oxidized. CoreM40/1-22, on the other hand, displays thick, wellpreserved sapropels (10 and 6 cm thick, respectively),and moderate burn-down ratios (50%). Both coresexhibit an interval of minor burn-down in the middlecore segment from S7 to S5. Visible sapropel thick-nesses vary between 7 and 17 cm and less than 50% ofthe reconstructed original sapropels have been oxidized.Sapropel S8 is distinctively different in the two cores,whereas the reconstructed sapropel is more than 20 cmthick and the preserved TOC rich layer exceeds 10 cm inM40/1-10, the sapropel in M40/1-22 only has a
Fig. 4. Planktonic oxygen isotope record (Globigerinoides ruber, complimenteAl ratios, TOC (total organic carbon, and bulk density records from core M40the Mediterranean Ridge (upper panel) and core M40/1-22 (33°39.5′N, 24°4Mediterranean Ridge. Dark gray bars indicate sapropels, gray bars sapropelic lisotope curves designate identified isotope events. Note that linear sedimconsequently do not represent accumulated sediment. Notice the possible hiatuhiatus between S3 and S4, where the sediment between MIS 5.3 and 5.1 ha
reconstructed thickness of 10 cm and is almostcompletely oxidized. The TOC content of the remainingdark layer actually is below 0.3%, well below the valuescharacteristic for a sapropel.
5. Discussion
5.1. Sediment
The relatively homogeneous sediment consists ofsilty muds, and mostly lacks clearly visible bedding, butcontains several intervals of preserved primary sedi-mentary features. Together with a decimeter-scalecyclicity in color variations, some possible lag deposits,and a few erosional contacts, this suggest a contouriticdeposition. Both cores are situated below the influenceof the westwards flowing Levantine Intermediate Water(LIW), but are in the path of the Eastern MediterraneanDeep Water (EDMW), which forms during cold wintersin the Adriatic and Aegean Seas (Roether et al., 1999).Direct measurements of the EMDW's current velocityare not available, but the rapid overturn rates (∼125years, POEM group, 1992) suggest vigorous bottomcurrents, in agreement with the well-oxidized sediments
d byGlobigerina bulloides duringMIS 6 inM40/1-10), Fe/Al andMn//1-10 (34°45.8′N, 19°45.7′E, 2972 m) situated on the western flank of1.2′E, 2004 m) situated in the Olimpi area on the southern flank of theayers, and light gray bars indicate erased “ghost’ sapropels. Numbers atentation rates are plotted versus depth; the areas under the curveses above S3 and S6 in M40/1-10, below S2 in core M40/1-22, and theve been largely removed.
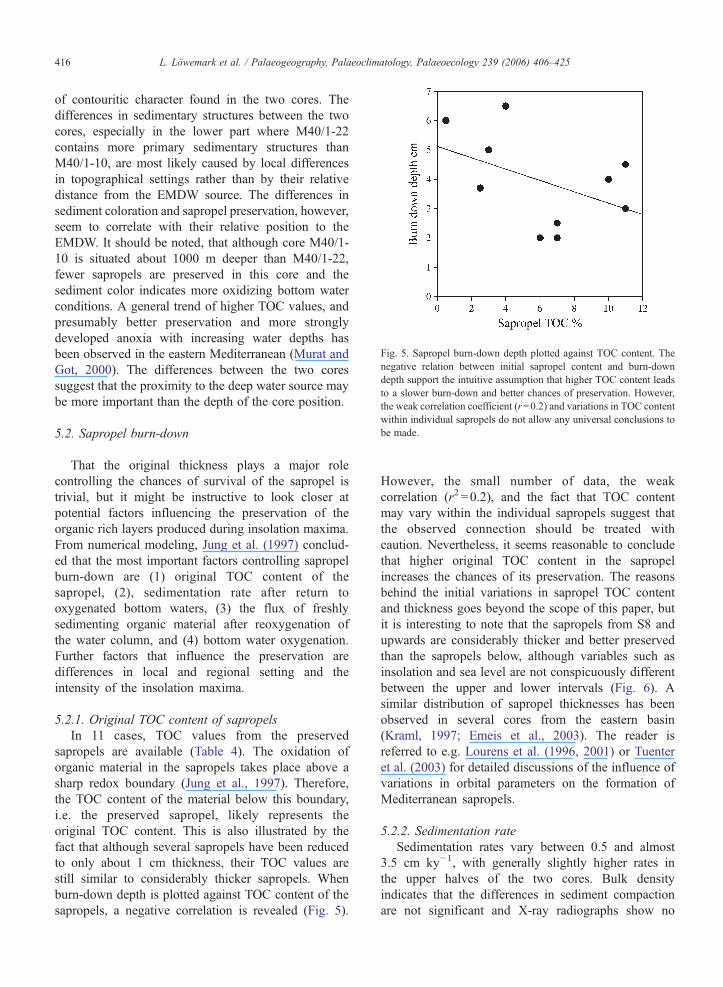
Fig. 5. Sapropel burn-down depth plotted against TOC content. Thenegative relation between initial sapropel content and burn-downdepth support the intuitive assumption that higher TOC content leadsto a slower burn-down and better chances of preservation. However,the weak correlation coefficient (r=0.2) and variations in TOC contentwithin individual sapropels do not allow any universal conclusions tobe made.
416 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
of contouritic character found in the two cores. Thedifferences in sedimentary structures between the twocores, especially in the lower part where M40/1-22contains more primary sedimentary structures thanM40/1-10, are most likely caused by local differencesin topographical settings rather than by their relativedistance from the EMDW source. The differences insediment coloration and sapropel preservation, however,seem to correlate with their relative position to theEMDW. It should be noted, that although core M40/1-10 is situated about 1000 m deeper than M40/1-22,fewer sapropels are preserved in this core and thesediment color indicates more oxidizing bottom waterconditions. A general trend of higher TOC values, andpresumably better preservation and more stronglydeveloped anoxia with increasing water depths hasbeen observed in the eastern Mediterranean (Murat andGot, 2000). The differences between the two coressuggest that the proximity to the deep water source maybe more important than the depth of the core position.
5.2. Sapropel burn-down
That the original thickness plays a major rolecontrolling the chances of survival of the sapropel istrivial, but it might be instructive to look closer atpotential factors influencing the preservation of theorganic rich layers produced during insolation maxima.From numerical modeling, Jung et al. (1997) conclud-ed that the most important factors controlling sapropelburn-down are (1) original TOC content of thesapropel, (2), sedimentation rate after return tooxygenated bottom waters, (3) the flux of freshlysedimenting organic material after reoxygenation ofthe water column, and (4) bottom water oxygenation.Further factors that influence the preservation aredifferences in local and regional setting and theintensity of the insolation maxima.
5.2.1. Original TOC content of sapropelsIn 11 cases, TOC values from the preserved
sapropels are available (Table 4). The oxidation oforganic material in the sapropels takes place above asharp redox boundary (Jung et al., 1997). Therefore,the TOC content of the material below this boundary,i.e. the preserved sapropel, likely represents theoriginal TOC content. This is also illustrated by thefact that although several sapropels have been reducedto only about 1 cm thickness, their TOC values arestill similar to considerably thicker sapropels. Whenburn-down depth is plotted against TOC content of thesapropels, a negative correlation is revealed (Fig. 5).
However, the small number of data, the weakcorrelation (r2 =0.2), and the fact that TOC contentmay vary within the individual sapropels suggest thatthe observed connection should be treated withcaution. Nevertheless, it seems reasonable to concludethat higher original TOC content in the sapropelincreases the chances of its preservation. The reasonsbehind the initial variations in sapropel TOC contentand thickness goes beyond the scope of this paper, butit is interesting to note that the sapropels from S8 andupwards are considerably thicker and better preservedthan the sapropels below, although variables such asinsolation and sea level are not conspicuously differentbetween the upper and lower intervals (Fig. 6). Asimilar distribution of sapropel thicknesses has beenobserved in several cores from the eastern basin(Kraml, 1997; Emeis et al., 2003). The reader isreferred to e.g. Lourens et al. (1996, 2001) or Tuenteret al. (2003) for detailed discussions of the influence ofvariations in orbital parameters on the formation ofMediterranean sapropels.
5.2.2. Sedimentation rateSedimentation rates vary between 0.5 and almost
3.5 cm ky−1, with generally slightly higher rates inthe upper halves of the two cores. Bulk densityindicates that the differences in sediment compactionare not significant and X-ray radiographs show no
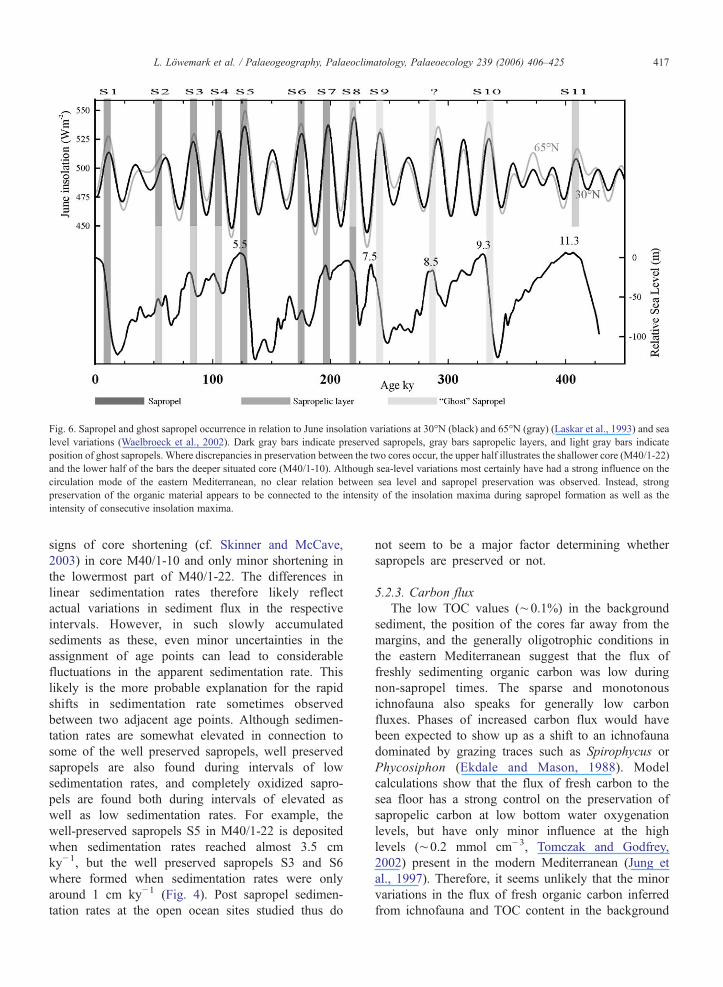
Fig. 6. Sapropel and ghost sapropel occurrence in relation to June insolation variations at 30°N (black) and 65°N (gray) (Laskar et al., 1993) and sealevel variations (Waelbroeck et al., 2002). Dark gray bars indicate preserved sapropels, gray bars sapropelic layers, and light gray bars indicateposition of ghost sapropels. Where discrepancies in preservation between the two cores occur, the upper half illustrates the shallower core (M40/1-22)and the lower half of the bars the deeper situated core (M40/1-10). Although sea-level variations most certainly have had a strong influence on thecirculation mode of the eastern Mediterranean, no clear relation between sea level and sapropel preservation was observed. Instead, strongpreservation of the organic material appears to be connected to the intensity of the insolation maxima during sapropel formation as well as theintensity of consecutive insolation maxima.
417L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
signs of core shortening (cf. Skinner and McCave,2003) in core M40/1-10 and only minor shortening inthe lowermost part of M40/1-22. The differences inlinear sedimentation rates therefore likely reflectactual variations in sediment flux in the respectiveintervals. However, in such slowly accumulatedsediments as these, even minor uncertainties in theassignment of age points can lead to considerablefluctuations in the apparent sedimentation rate. Thislikely is the more probable explanation for the rapidshifts in sedimentation rate sometimes observedbetween two adjacent age points. Although sedimen-tation rates are somewhat elevated in connection tosome of the well preserved sapropels, well preservedsapropels are also found during intervals of lowsedimentation rates, and completely oxidized sapro-pels are found both during intervals of elevated aswell as low sedimentation rates. For example, thewell-preserved sapropels S5 in M40/1-22 is depositedwhen sedimentation rates reached almost 3.5 cmky−1, but the well preserved sapropels S3 and S6where formed when sedimentation rates were onlyaround 1 cm ky−1 (Fig. 4). Post sapropel sedimen-tation rates at the open ocean sites studied thus do
not seem to be a major factor determining whethersapropels are preserved or not.
5.2.3. Carbon fluxThe low TOC values (∼0.1%) in the background
sediment, the position of the cores far away from themargins, and the generally oligotrophic conditions inthe eastern Mediterranean suggest that the flux offreshly sedimenting organic carbon was low duringnon-sapropel times. The sparse and monotonousichnofauna also speaks for generally low carbonfluxes. Phases of increased carbon flux would havebeen expected to show up as a shift to an ichnofaunadominated by grazing traces such as Spirophycus orPhycosiphon (Ekdale and Mason, 1988). Modelcalculations show that the flux of fresh carbon to thesea floor has a strong control on the preservation ofsapropelic carbon at low bottom water oxygenationlevels, but have only minor influence at the highlevels (∼0.2 mmol cm−3, Tomczak and Godfrey,2002) present in the modern Mediterranean (Jung etal., 1997). Therefore, it seems unlikely that the minorvariations in the flux of fresh organic carbon inferredfrom ichnofauna and TOC content in the background

418 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
sediment should exert more than only minimalinfluence on the preservation of sapropels in the twostudied cores.
5.2.4. Bottom water oxygenationThe sapropel burn-down generally seems to have
been more efficient in M40/1-10, closer to the source ofdeep water formation, despite this core being situated ata larger water depth. Even where organic carbon contentin remaining sapropels is higher in M40/1-10 than inM40/1-22, the burn-down has been more efficient in thesite closer to the deep water source (Table 4). A similartendency of growing preserved sapropel thicknesseswith increasing distance from the deep-water source canbe observed in four cores studied by Emeis et al. (2003).Model calculations by Jung et al. (1997) clearlydemonstrate the importance of bottom water oxygenlevel in determining the efficiency of carbon oxidation.High bottom water oxygen levels in combination withvigorous bottom water currents that prevented theformation of bottom nepheloid layers probably are themost important factors for an efficient burn-down of thesapropels.
5.2.5. Intensity of insolation maximumModeling results show that the burn-down is fastest
in the first few thousand years (Jung et al., 1997).After this initial phase the burn-down continues at aconsiderably slower rate (0.5–1 cm ky−1). Theoreti-cally, the oxidation would go on indefinitely ifsedimentation rates and flux of fresh carbon weresufficiently low (Jung et al., 1997). The preservationpotential of the sapropel therefore is not onlydepending on initial thickness and carbon content,which are directly related to the intensity of theinsolation maximum during sapropel formation, but isalso depending on the strength of the next monsoonalmaximum (Fig. 6). A strong maximum results in awell developed sapropel that acts as a shield for theunderlying sapropel, protecting it against furtheroxidation. The results show that the best preservedsapropels occur where a series of strong insolationmaxima come clustered. In contrast, moderate or weakinsolation maxima followed by weak insolationmaxima show the poorest preserved sapropels, oftenonly detectable as “ghost” sapropels through geo-chemical analysis.
The observed tendency of strong manganese peaksmarking the top of intensely or completely overprintedsapropels, but small or hardly noticeable peaks abovesome of the best preserved sapropels probably alsorelates to the intensity of the subsequent insolation
maxima. If the following insolation maximum is strong,then it will effectively seal off the underlying sapropelbefore its carbon has been oxidized. The oxygen in thepore water between the two sapropels presumably willbe rapidly consumed, anoxic pore waters will prevail,the manganese will become mobile and the peaksmarking the top of the sapropels will diffuse awaytowards areas of lower manganese ion concentration.Extreme examples of this phenomenon are found in S7in M40/1-10 and S7 and S5 in M40/1-22.
5.3. Ichnological response to variations in bottomwater oxygenation
5.3.1. Ichnological modelsBurrows left in the sediment by benthic organisms
reflect the behavior of the producing animal. Thebehavior, in turn, is a response to variations in substrateand different environmental conditions (e.g., Bromley,1996). In the deep-sea, variations in substrate conditionsare usually limited to a smaller range than in shelf seas.Consequently, a number of studies of deep-sea macro-benthos communities have revealed that there is nogeneral correlation between community patterns andphysical sediment parameters. Instead, large-scale varia-tions in hydrography, food flux and bottom wateroxygenation are thought to be responsible for thevariations observed (Flach et al., 1998; Schnack, 1998;Levin et al., 2000; Flach et al., 2002; Lavaleye et al.,2002). Therefore, the variations in trace fossil compo-sition in the stratigraphic column primarily comprise arecord of the response of the benthic community toenvironmental changes. In the following two models ofichnological response to lowered oxygen levels aredescribed.
1) Based primarily on material from Cretaceous chalks,Savrda et al. (1984) and Savrda and Bottjer (1986,1989, 1991) developed a model for the ichnologicalresponse to lowered bottom water oxygenation(summarized in Savrda, 1992). Basically, the modelcompares the preserved tiering sequence fromgradually and rapidly decreasing bottom wateroxygenation events with the normal tiering sequence.It also investigates the differences between a gradualand a rapid return to well oxygenated conditions afteran anoxic or dysoxic event. With decreasing bottom-water oxygen levels, changes in the benthic commu-nity can be expected. A gradual decrease in bottomwater oxygenation causes a rise of the redox potentialdiscontinuity (RPD) in the sediment. The living-space for the endobenthos thus decreases, and a

419L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
telescoping and upward movement of the tiersfollows. As the decrease in oxygen level continues,the upper tiers become replaced by deeper tiers. Thisprocess continues until the RPD reaches the sediment/water interface. When the bottom water also becomesanoxic, all bioturbation ceases. Because larger andmore active animals consume more oxygen, adecrease in animal size and thus burrow diameter aswell as a decreased burrow depth by the individualtrace makers is registered. In contrast, a rapiddecrease in bottom water oxygenation leaves notime for an adjustment in burrow depth of the fauna.In this case, the burrows in the mixed and transitionlayers are preserved as “frozen tiers” beneath thelaminated sediments deposited during the anoxicevent. Gradually rising oxygen levels after anoxicconditions result in the destruction of laminatedsediments, and in the development of a tieredsequence. Progressively larger and more oxygen-demanding traces will be introduced to the sequenceresulting in partly reversed crosscutting relations. Ifoxygen increases rapidly, the uppermost laminatedlayers become destroyed by the development of themixed layer. From this layer, well defined burrows arepiping down into the underlying laminated sediments(Savrda, 1992).
2) Another approach to ichnological reconstruction ofbottom water conditions was followed by Ekdale andMason (1988). Based on Carboniferous sedimentsfrom the Oquirrh Basin (northern Utah, USA), theyproposed a model where four distinctly different tracefossil associations are interpreted to represent differ-ent levels of bottom water oxygenation. (A) At fullyoxygenated bottom and interstitial-water conditions,there is little food in the sediment and the trace fossilassociation is dominated by dwelling traces (Dom-ichnia) such as Skolithos. (B) As the interstitial waterbecomes dysoxic, but bottom water still remains oxicor dysoxic, a shift toward grazing traces (Pascichnia)occurs, indicating the presence of food in thesubstrate but also adequate oxygen levels in theinterstitial waters. (C) When the pore water becomesanoxic at still oxygenated bottom waters, changetowards a Zoophycos and Chondrites dominatedassociation occurs. Ekdale and Mason (1988) assumethat they represent an opportunistic behavior oforganisms exploiting deeply situated food resources.However, more recent studies suggest more compli-cated ethological explanations, including chemosym-biotic gardening and cache behavior for these tracefossils (cf. Seilacher, 1990; Bromley, 1991; Fu, 1991;Löwemark and Schäfer, 2003). (D) When both
interstitial and bottom waters have become anoxic,all higher life ceases and laminated sediments arepreserved.
Although the rhythmic shifts between anoxic andoxic environments would seem ideal to record ichno-logic responses such as described by Savrda and Bottjer(1986), Savrda (1992) or Ekdale and Mason (1988),none of the predicted responses were observed. The welloxidized sediments between the sapropels, where welldeveloped tiering (Savrda, 1992) or dwelling traces(domichnia) (Ekdale and Mason, 1988) supposedlyshould be prevailing, are dominated by a mottlingbioturbation and only in a few limited intervals tracefossils possibly referable to Thalassinoides were seen.The ichnology of the studied cores also differ from thetrace fossils described from a Cretaceous pelagicsequence of marls, marly clays and marly limestoneswith rhythmically intercalated black shales (Erba andPremoli Silva, 1994). Although sediment composition,oceanic setting and the repeated occurrence of organic-rich layers are similar to the studied Mediterraneancores, the Cretaceous sequence contains a different setof trace fossils with abundant Chondrites and cyclicoccurrences of Zoophycos and Teichichnus.
These discrepancies most likely can be ascribed to theconfined nature of the eastern basin and the wayMediterranean deep waters are formed. Because of thehigh intermediate and deep water temperatures, causinga rapid degradation of sinking organic matter, the benthicenvironment is characterized by low food flux andconsequently by a benthic fauna of low diversity and lowdensity (cf. Jumars andWheatcroft, 1989;Wetzel, 1991).These oligotrophic conditions are also reflected by athinner than usual mixed layer in the eastern Mediter-ranean basin (Basso et al., 2004). The ichnofauna justbelow the sapropels also differ from what would beexpected from the models. No development of thepascichnina (grazing) or fodinichnia (deposit feeding)dominated trace fossil associations described by Ekdaleand Mason (1988) was observed below the sapropel.Neither was the upwards telescoping of the tiers norfrozen tiers (cf. Savrda, 1992) observed. The lack ofgradual changes in the ichnofauna below the sapropelssuggests a rapid onset of anoxic conditions.
The conditions leading to sapropel formation alsofavor a rapid onset of anoxic conditions. Becausesapropels form during insolation maxima, when seasurface temperature and monsoonal precipitationincreases, the surface waters quickly become lessdense than the colder and more saline deep waters(Rohling, 1994; Emeis et al., 2003). The halted deep

420 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
water formation combined with increased productivityquickly led to anoxic conditions of the deep waters(Mangini and Schlosser, 1986). Because the transitionfrom oxygenated to anoxic conditions was swift, therewas no time for an ichnofauna adjusted to highertrophic levels to develop. Furthermore, as the foodflux was too low to allow the development of tieredtrace fossil assemblages during well-oxygenated times,there were no trace fossils that could be preserved asfrozen tiers below the sapropel. Accordingly, neitheran upwards shift of deeper tiers nor frozen tiers wereobserved below the sapropels.
In the studied cores, there were only few distincttrace fossils penetrating down into the sapropels fromabove, although the upper parts were generally erasedand the entire sapropels had been oxidized in severalcases. In a few overprinted sapropels, the laminations ofthe oxidized sapropels were still visible (e.g., S3 and S5in M40/1-10), corroborating the view of Jung et al.(1997) that bioturbation by macroorganisms plays onlya minor role in the burn-down of sapropels.
When deep water formation commenced aftersapropel formation, the deep waters quickly becamewell oxygenated and oligotrophic again. The rapid returnto oligotrophic conditions may be one reason for thesparse ichnofauna encountered after sapropel conditionsended, although the organic matter in the sapropelsthemselves would have constituted a considerable foodsource. One might further speculate that the restrictedwater exchange with the open oceanmade recolonizationof the deep basins after the anoxic sapropel events slowerthan it would have been in the open ocean. It has beensuggested (Bouchet and Taviani, 1992) that the presentdeep-sea fauna of the Mediterranean actually consists ofa pseudopopulation of Atlantic species. Because of thehigh temperatures and salinities of the Mediterraneanbottom waters, many benthic species have difficultiesmaintaining their reproductive cycles. The Mediterra-nean fauna therefore is depending on a continuous inputof larvae from the Atlantic Ocean via Atlantic InflowWater entering through the Gibraltar straight. Similarconstraints on the diversity and density of the benthicfauna likely acted during previous non-sapropelic times,partially explaining the meager ichnodiversity observedin the cores.
In summary, the models proposed by Savrda andBottjer (1986) and Ekdale and Mason (1988) are oflimited value in explaining bottom water oxygenationrelated variations in the ichnology of semi-confined,generally warm, well-oxygenated deep basins such asthe Mediterranean Sea. However, the lack of adaptivechanges in the ichnofauna as described in their models
do suggest a very rapid switch from well-oxidized,oligotrophic conditions to high-productive surfacewaters and anoxic bottom waters.
5.3.2. Individual trace fossilsThe sparse occurrence of Chondrites, an otherwise
common trace in Late Quaternary deep-sea sediment(e.g., Wetzel, 1983a,b; Löwemark et al., 2004), isattributed to the generally low organic carbon content.The branched tube-system of Chondrites is believed tobe a sulphide well, where its producer lives inchemosymbiosis with H2S-oxidizing bacteria (Seila-cher, 1990; Fu, 1991). Highly oxic bottom waterresulting in low TOC values and oxic pore waterstherefore are unsuitable for this kind of behavior.Consequently, the few Chondrites found are associatedwith sapropelic and laminated layers indicating loweredpore water oxygenation and higher TOC values. Baas etal. (1998) found massive occurrences of Chondritesbelow the Heinrich layers in the eastern North Atlantic.These layers were deposited during halted or reducedthermohaline circulation leading to lowered oxygena-tion levels of the bottom waters. Also, on the northernslope of the Iceland–Færø-ridge, where sluggish bottomcurrents, poorly sorted, organic-rich sediments andoxygen deficiency prevails, Fu and Werner (1994)found Chondrites to be the dominating ichnogenus.Thus, our findings that Chondrites is absent under well-oxygenated, but occur (although sparse) under dysoxicto anoxic conditions, support the notion that theChondrites-producer has a strong preference for dys-oxic–anoxic pore waters, and that the trace maker undercertain circumstances may act as an opportunist (Ekdale,1985), being the first to colonize organic rich layerswhen the sea floor becomes habitable again after ananoxic event.
Although single Trichichnus burrows were sporad-ically present in the background sediment, most ofthem are strongly concentrated to the thickest and bestpreserved sapropels and in the sediments directly below(Fig. 7). This suggests that the producing organism hasa clear preference for reducing pore waters. Thesefindings are in agreement with earlier observationswhere the increased abundance in poorly oxygenatedenvironments (Thompson, 1980; Blanpied and Bel-laiche, 1981; Thomsen and Vorren, 1984; Werner,1985; Löwemark, 2003) and the deep penetration ofTrichichnus into anoxic environments (Weaver andSchultheiss, 1983; McBride and Picard, 1991) havebeen interpreted to indicate a chemosymbiotic behav-ior. Low oxygenated bottom- and pore waters com-bined with organic-rich sediments probably offer

Fig. 7. Large numbers of pyritized Trichichnus and “Mycellia” belowsapropel S7 in core M40/1-10. T = Trichichnus, M = “Mycellia”.
421L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
perfect conditions for chemosymbiotic activity; highlevels of H2S available for chemosymbiosis and fewcompeting organisms destroying the upper parts of theburrows.
The occurrence of Scolicia and Thalassinoides inboth cores just above sapropel S6 may be a coincidencerelated to similarities in substrate conditions, but it mightalso suggest a colonization event under conditionsfavorable to the producers of these traces. Manycrustaceans, echinoids, and worms have planktonicstages in their reproductive cycle, which may explainthe simultaneous occurrences of trace fossils such asScolicia and Thalassinoides in the two cores. Underparticularly favorable conditions, the larvae would beable to colonize large areas resulting in horizons ofcertain types of trace fossils that appear in the samestratigraphic position in both cores. This would be inagreement with results from benthic faunal studiesindicating mesotrophic conditions before and after thedeposition of sapropel S6 (Schmiedl et al., 2003). Incontrast to most other well-preserved sapropels thatformed under warm climatic conditions, sapropel S6formed during glacial condition with enhanced produc-tivity both before and after sapropel formation (Schmiedl
et al., 1998, 2003), creating favorable conditions forfilter feeders and deposit feeders. On the other hand,large Thalassinoides–Planolites-like traces are alsofound below the burn-downed sapropel S10, formedduring interglacial MIS 9 when oligotrophic conditionsprevailed. Furthermore, the producers of the Scolicia-trace in late Quaternary deep-marine sediments havebeen found to show a preference for well sorted coarsesilts to fine sands and rather high sedimentation rates(Wetzel, 1991;Werner, 1996; Fu andWerner, 2000). Theintervals in the two studied cores containing Scolicia areall characterized by enhanced levels of sand sizedmaterial, although not always by enhanced sedimenta-tion rates. Therefore, more cores covering a larger areaare needed to test the hypothesis that the observedvariations are due to opportunistic colonization by larvaefrom the Atlantic Ocean.
6. Conclusions
Variations in ichnology and in the sapropel develop-ment and preservation over the last 400,000 years havebeen reconstructed for two sediment cores from theeastern Mediterranean Sea. Based on the combinedstudy of TOC, bulk density, Mn / Al and Fe / Al ratios,trace fossils and sedimentary structures from X-rayradiographs, several conclusions can be drawn:
• Previously proposed ichnological models for recon-struction of paleo-oxygenation histories are of limiteduse in oligotrophic basins with warm and fast formingdeep waters. In the studied cores, only two majormodes of bioturbation exist, sparse mottling and nobioturbation at all.
• The lack of changes in the ichnofauna directlyunderneath the sapropels suggests a rapid onset ofanoxia, at least in the deeper parts of the basin.
• The results confirm a preference for anoxic porewaters by the producers of the trace fossils Chon-drites and Trichichnus, presumably because of theirchemosymbiotic use of H2S as an energy source.
• Our data tentatively confirm the trivial notion that theoriginal thickness and TOC content of the sapropelinfluences the burn-down rate, as suggested by modelcalculations (cf. Jung et al., 1997). However, the smalldifferences in sedimentation rates between wellpreserved and completely erased sapropels suggestthat sedimentation rate only plays a minor role incontrolling whether or not sapropels survivereoxygenation.
• In contrast, the local and regional setting in relation tothe deep-water source plays an influential role

422 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
determining the preservation potential of sapropels.Cores retrieved distal to the locus of well-oxygenateddeep water formation generally display better pre-served sapropels
• The development of subsequent sapropels also influ-ences the preservation of earlier sapropels. Stronginsolation maxima result in thick, TOC-rich sapropelsthat may act as lids, preventing downward oxygendiffusion and further oxidation of underlying sapropels.
One important general conclusion that can be drawnregarding sapropels is that the observed thickness of thepreserved sapropel layers does not necessarily corre-spond to the original sapropel thickness. The presentedmethod, using radiographs in combination with ele-ment–aluminium ratios of iron and manganese, is a fastand effective way to determine original thickness, burn-down rate, and even the positions of completely erased“ghost”-sapropels.
Acknowledgments
Niels Jöns (IfG, Kiel), Joachim Schönfeld and JuttaPagel (IFM-GEOMAR, Kiel) are thanked for help withTOC and carbonate measurements. Andrew K. Rinds-berg (Geological Survey of Alabama) is cordiallythanked for invaluable ichnological and semanticdiscussions. Yoshiyuki Iizuka (IESAS, Taipei) iscordially thanked for his help with the SEM. WernerHieke (TU München) has as chief scientist on CruiseM40/1 carefully and successfully selected the positionsof the investigated cores and gave many helpfulsuggestions. We thank Konstantinos I. Konstantinou(Academia Sinica) for help with the age modeling andHelmut Erlenkeuser and the team at the LeibnizLaboratory, Kiel for stable isotope measurements.Stephan Steinke (RCOM, Bremen), Teh-Quei Lee(IESAS, Taipei), and Priska Schäfer (IfG, Kiel) arethanked for valuable discussions. The two anonymousreviewers are cordially thanked for their constructivereviews. This work was funded by the DeutscheForschungsgemeinschaft (Grant Scha 355/15) to PriskaSchäfer (IfG, Kiel).
References
Aksu, A.E., Yasar, D., Mudie, P.J., 1995. Paleoclimatic andpaleoceanographic conditions leading to development of sapropellayer S1 in the Aegean Sea. Palaeogeography, Palaeoclimatology,Palaeoecology 116, 71–101.
Baas, J.H., Schönfeld, J., Zahn, R., 1998. Mid-depth oxygendrawdown during Heinrich events: evidence from benthicforaminiferal community structure, trace fossil tiering, and
benthic δ13C at the Portuguese margin. Marine Geology 152(1–3), 25–55.
Baldwin, C.T., McCave, I.N., 1999. Bioturbation in an active deep-seaarea: implications for models of trace fossil tiering. Palaios 14 (4),375–389.
Bassinot, F.C., Labeyrie, L.D., Vincent, E., Quidelleur, X., Shackleton,N.J., Lancelot, Y., 1994. The astronomical theory of climate andthe age of the Brunhes–Matuyama magnetic reversal. Earth andPlanetary Science Letters 126, 91–108.
Basso, D., Thomson, J., Corselli, C., 2004. Indications of lowmacrobenthic activity in the deep sediments of the easternMediterranean Sea. Scientia Marina 68 (3), 53–62.
Berggren, W.A., Hilgen, F.J., Langereis, C.G., Kent, D.V., Obrado-vich, J.D., Raffi, I., Raymo, M.E., Shackleton, N.J., 1995. LateNeogene chronology: new perspectives in high-resolution stratig-raphy. GSA Bulletin 107 (11), 1272–1287.
Blanpied, C., Bellaiche, G., 1981. Bioturbation on the pelagicplatform: ichnofacies variations as paleoclimatic indicator. MarineGeology 43, M49–M57.
Bouchet, P., Taviani, M., 1992. The Mediterranean deep-sea fauna:pseudopopulations of Atlantic species? Deep-Sea Research 39A(2), 169–184.
Bouma, A.H., 1964. Notes on X-ray interpretation of marinesediments. Marine Geology 2, 278–309.
Bromley, R.G., 1991. Zoophycos: strip mine, refuse dump, cache orsewage farm? Lethaia 24, 460–462.
Bromley, R.G., 1996. Trace Fossils: Biology, Taphonomy andApplications. Chapman and Hall, London. 361 pp.
Bromley, R.G., Asgaard, U., 1975. Sediment structures produced by aspatangoid echinoid: a problem of preservation. Bulletin of theGeological Society of Denmark 24, 261–281.
Bromley, R.G., Ekdale, A.A., 1984. Chondrites: a trace fossil indicatorof anoxia in sediments. Science 224, 872–874.
Castradori, D., 1993. Calcareous nannofossils and the origin ofeastern Mediterranean sapropels. Paleoceanography 8 (4),459–471.
Cita, M.B., Vergnaud-Grazzini, C., Robert, C., Chamley, H., Ciaranfi,N., D'Onofrio, S., 1977. Paleoclimatic record of a long deep seacore from the eastern Mediterranean. Quaternary Research 8,205–235.
Corselli, C., Principato, M.S., Maffioli, P., Crudeli, D., 2002. Changesin planktonic assemblages during sapropel S5 deposition: evidencefrom Urania Basin area, eastern Mediterranean. Paleoceanography17 (3) (1029/2000PA000536).
De Lange, G.J., ten Haven, H.L., 1983. Recent sapropel formation inthe eastern Mediterranean. Nature 305, 797–798.
De Quatrefages, M.A., 1849. Note sur la Scolicia prisca (A. De Q.),annélide fossile de la craie. Annales des Sciences Naturelles,Zoologie,série 3 12, 265–266.
Ehrenberg, K., 1944. Ergänzende Bemerkungen zu den seinerzeit ausdem Miozän von Burgschleinitz beschriebenen Gangkernen undBauten dekapoder Krebse. Paläontologische Zeitschrift 23,354–359.
Ekdale, A.A., 1985. Paleoecology of the marine endobenthos.Palaeogeography, Palaeoclimatology, Palaeoecology 50, 63–81.
Ekdale, A.A., Mason, T.R., 1988. Characteristic trace-fossil associa-tions in oxygen-poor sedimentary environments. Geology 16,720–723.
Ekdale, A.A., Bromley, R.G., Pemberton, S.G., 1984. Ichnology —The Use of Trace Fossils in Sedimentology and Stratigraphy. ShortCourse Notes, vol. 15. Society of Economic Paleontologists andMineralogists, Tulsa, Oklahoma. 317 pp.

423L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
Emeis, K.-C., Sakamoto, T., Wehausen, R., Brumsack, H.-J., 2000a.The sapropel record of the eastern Mediterranean Sea— results ofOcean Drilling Program Leg 160. Palaeogeography, Palaeoclima-tology, Palaeoecology 158, 371–395.
Emeis, K.-C., Struck, U., Schulz, H.-M., Rosenberg, R., Bernasconi,S., Erlenkeuser, H., Sakamoto, T., Martinez-Ruiz, F., 2000b.Temperature and salinity variations of Mediterranean Sea surfacewaters over the last 16,000 years from records of planktonic stableoxygen isotopes and alkenone unsaturation ratios. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology 158, 259–280.
Emeis, K.-C., Schulz, H., Struck, U., Rossignol-Strick, M., Erlenkeu-ser, H., Howell, M.W., Kroon, D., Mackensen, A., Ishizuka, S.,Oba, T., Sakamoto, T., Koizumi, I., 2003. Eastern Mediterraneansurface water temperatures and D18O composition duringdeposition of sapropels in the late Quaternary. Paleoceanography18 (1).
Erba, E., Premoli Silva, I., 1994. Orbitally driven cycles in trace-fossildistribution from the Piobbico core (late Albian, central Italy).Special Publication of the International Association of Sedimen-tologists 19, 211–225.
Flach, E., Lavaleye, M., Stigter, H.d., Thomsenc, L., 1998. Feedingtypes of the benthic community and particle transport across theslope of the N.W. European continental margin (Goban Spur).Progress in Oceanography 42, 209–231.
Flach, E., Muthumbi, A., Heip, C., 2002. Meiofauna and macrofaunacommunity structure in relation to sediment composition at theIberian margin compared to the Goban Spur (NE Atlantic).Progress in Oceanography 52, 433–457.
Frey, R.W., 1970. Trace fossils of Fort Hays Limestone Member,Niobrara Chalk (Upper Cretaceous), west-central Kansas. Univer-sity of Kansas Paleontological Contributions, Article 53 (Creta-ceous 2), 41 pp.
Fu, S., 1991. Funktion, Verhalten und Einteilung fucoider undlophocteniider Lebenspuren, Courier Forschungsinstitut Sencken-berg. Senckenbergische Naturforschende Gesellschaft Frankfurt a.M. 1–79.
Fu, S., Werner, F., 1994. Distribution and composition of biogenicstructures on the Iceland–Faeroe Ridge: relation to differentenvironments. Palaios 9, 92–101.
Fu, S., Werner, F., 2000. Distribution, ecology and taphonomy of theorganism trace, Scolicia, in northeast Atlantic deep-sea sediments.Palaeogeography, Palaeoclimatology, Palaeoecology 156,289–300.
Haddad, G.A., Droxler, A.W., 1996. Metastable CaCO3 dissolutionat intermediate water depths of the Caribbean and westernNorth Atlantic: implications for Intermediate water circulationduring the past 200,000 years. Paleoceanography 11 (6),701–716.
Häntzschel, W., 1975. Miscellanea, Supplement 1, Trace Fossils andProblematica. Treatise on Invertebrate Paleontology, Part W. TheGeological Society of America and the University of Kansas,Boulder, Colorado. 269 pp.
Hieke, W., Hemleben, C., Linke, P., Türkay, M., Weikert, H. (Eds.),1999. Mittelmeer 1997/98, Cruise No. 40, 28 October 1997–10February 1998. METEOR-Berichte, 99-2. Universität Hamburg.286 pp.
Higgs, N.C., Thomson, J., Wilson, T.R.S., Croudace, I.W., 1994.Modification and complete removal of eastern Mediterraneansapropels by postdepositional oxidation. Geology 22 (5), 423–426.
Jumars, P.A., Wheatcroft, R.A., 1989. Responses of benthos tochanging food quality and quantity, with a focus on deposit feedingand bioturbation. In: Berger, W.H., Smetacek, V.S. (Eds.),
Productivity of the Ocean: Present and Past. John Wiley andSons, New York, pp. 235–253.
Jung, M., Ilmberger, J., Mangini, A., Emeis, K.-C., 1997. Why someMediterranean sapropels survived burn-down (and others did not).Marine Geology 141, 51–60.
Keighley, D.G., Pickerill, R.K., 1995. The ichnotaxa Paleophycus andPlanolites: historical perspectives and recommendations. Ichnos 3,301–309.
Kidd, R.B., Cita, M.B., Ryan, W.B.F., 1978. Stratigraphy of easternMediterranean sapropel sequences recovered during Leg 42A andtheir paleoenvironmental significance. Initial Reports of the Deep-Sea Drilling Project 42A, 421–443.
M. Kraml. 1997. Laser-40Ar/39Ar-Datierungen an distalen marinenTephren des jung-quartären mediterranen Vulkansmus (IonischesMeer, METEOR-Fahrt 25/4). Dr Thesis, Albert-Ludvig-Universi-tät, Freiburg. 216 pp.
Lacombe, H., Tchernia, P., 1972. Caractères hydrologiques etcirculation des eaux en Méditerranée. In: Stanley, D.J. (Ed.), TheMediterranean Sea: A Natural Sedimentation Laboratory. Dowden,Hutchinson and Ross, Stroudsburg, pp. 26–36.
Lacombe, H., Gascard, J.C., Gonella, J., Bethoux, J.P., 1981. Responseof the Mediterranean to the water and energy fluxes across itssurface, on seasonal and interannual scales. Oceanologica Acta 4(2), 247–255.
Laskar, J., Joutel, F., Boudin, F., 1993. Orbital, precessional, andinsolation quantities for the Earth from −20 Myr to +10 Myr.Astronomy and Astrophysics 270, 522–533.
Lavaleye, M.S.S., Duineveld, G.C.A., Berghuis, E.M., Kok, A.,Witbaard, R., 2002. A comparison between the megafaunacommunities on the N.W. Iberian and Celtic continental margins— effects of coastal upwelling? Progress in Oceanography 52,459–476.
Levin, L.A., Gage, J.D., Martin, C., Lamont, P.A., 2000. Macro-benthic community structure within and beneath the oxygenminimum zone, NWArabian Sea. Deep-Sea Research II 47 (1–2),189–226.
Lourens, L.J., Antonarakou, A., Hilgen, F.J., Van Hoof, A.A.M.,Vergnaud-Grazzini, C., Zachariasse, W.J., 1996. Evaluation of thePlio–Pleistocene astronomical timescale. Paleoceanography 11(4), 391–413.
Lourens, L.J., Wehausen, R., Brumsack, H.J., 2001. Geologicalconstraints on tidal dissipation and dynamical ellipticity of theEarth over the past three million years. Nature 409, 1029–1033.
Löwemark, L., 2003. Automatic image analysis of X-ray radiographs:a new method for ichnofabric evaluation. Deep-Sea Research I.
Löwemark, L., Schäfer, P., 2003. Ethological implications from adetailed X-ray radiograph and 14C-study of the modern deep-seaZoophycos. Palaeogeography, Palaeoclimatology, Palaeoecology192 (1–4), 101–121.
Löwemark, L., Werner, F., 2001. Dating errors in high-resolutionstratigraphy: a detailed X-ray radiograph and AMS-14C study ofZoophycos burrows. Marine Geology 177 (3–4), 191–198.
Löwemark, L., Schönfeld, J., Werner, F., Schäfer, P., 2004. Tracefossils as a paleoceanographic tool: evidence from Late Quaternarysediments of the southwestern Iberian margin. Marine Geology204 (1–2), 27–41.
Mangano, M.G., Buatois, L.A., Maples, C.G., West, R.R., 2000. Anew ichnospecies of Nereites from Carboniferous tidal-flat faciesof eastern Kansas, USA: implications for the Nereites–Neonereitesdebate. Journal of Paleontology 74 (1), 149–157.
Mangini, A., Schlosser, P., 1986. The formation of eastern Mediter-ranean sapropels. Marine Geology 72, 115–124.

424 L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
Martinson, D.G., Pisias, N.G., Hays, J.D., Imbrie, J., Moore, T.C.,Shackelton, N.J., 1987. Age dating and the orbital theory of the iceages: development of a high-resolution 0–300,000-year chronos-tratigraphy. Quaternary Research 27, 1–29.
McBride, E.F., Picard, M.D., 1991. Facies implications of Trichichnusand Chondrites in turbidites and hemipelagites, Marnoso-arenaceaformation (Miocene), Northern Apennines, Italy. Palaios 6 (3),281–290.
Minas, H.J., Coste, B., Le Corre, P., Minas, M., Raimbault, P., 1991.Biogeological and geochemical signatures associated with thewater circulation through the strait of Gibraltar and in the WesternAlboran Sea. Journal of Geophysical Research 96 (C5),8755–8771.
Murat, A., Got, H., 2000. Organic carbon variations of the easternMediterranean Holocene sapropel: a key for understandingformation processes. Palaeogeography, Palaeoclimatology,Palaeoecology 158, 241–257.
Murchison, R.I., 1839. The Silurian system. Part 1. Founded ongeological researches in the countries of Solop, Hereford, Radnor,Montgomery, Caermarthen, Brecon, Pembroke, Monmouth, Glou-cester, Worcester, and Stafford; with descriptions of the coal-fieldsand overlying formations. John Murray, London, pp. i–xxxiii,1–578; part II. Organic remains, pp. 579–768.
Nicholson, H.A., 1873. Contributions to the study of the errantannelides of the older Paleozoic rocks. Royal Society of London,Proceedings 21, 288–290.
Olausson, E., 1961. Sediment cores from the Mediterranean Sea andthe Red Sea. In: Pettersson, H. (Ed.), Reports of the Swedish Deep-Sea Expedition 1947–1948, 8:4. Göteborgs Kungliga Vetenskaps-och Vitterhets-Samhälle, Göteborg, pp. 337–391.
Pedersen, T.F., Calvert, S.E., 1990. Anoxia vs. productivity: whatcontrols the formation of organic-carbon-rich sediments andsedimentary rocks. American Association of Petroleum GeologistsBulletin 74 (4), 454–466.
Pemberton, S.G., Frey, R.W., 1982. Trace fossil nomenclature and thePlanolites–Palaeophycos dilemma. Journal of Paleontology 56(4), 843–881.
Pinardi, N., Masetti, E., 2000. Variability of the large scale generalcirculation of the Mediterranean Sea from observations andmodelling: a review. Palaeogeography, Palaeoclimatology,Palaeoecology 158, 153–173.
POEM group, 1992. General circulation of the Eastern Mediterranean.Earth-Science Reviews 32, 285–309.
Reid, J.L., 1979. On the contribution of the Mediterranean outflow tothe Norwegian–Greenland Sea. Deep-Sea Research 26A,1199–1223.
Reineck, H.-E., 1968. Lebensspuren von Herzigeln. SenkenbergianaLethaea 49 (4), 311–319.
Rio, D., Raffi, I., Villa, G., 1990. Pliocene–Pleistocene calcareousnannofossil distribution patterns in the western Mediterranean. In:Kastens, K.A., Mascle, J., Auroux, C., Bonatti, E., Broglia, C.,Channell, J.E.T., Curzi, P., Emeis, K.-C., Glacon, G., Hasegawa, S.,Hieke, W., McCoy, F.W., McKenzie, J.A., Mascle, G., Mendelson,J., Mueller, C., Rehault, J.-P., Robertson, A.H.F., Sartori, R.,Sprovieri, R., Torii, M., Stewart, N.J. (Eds.), Proc. ODP Sci.Results, vol. 107, pp. 513–533.
Roether, W., Well, R., 2001. Oxygen consumption in the EasternMediterranean. Deep-Sea Research I 48, 1535–1551.
Roether, W., Beitzel, V., Sultenfuss, J., Putzka, A., 1999. The EasternMediterranean tritium distribution in 1987. Journal of MarineSystems 20 (1–4), 49–61.
Rohling, E.J., 1991. A simple two-layered model for shoaling of theeastern Mediterranean pycnocline due to glacio-eustatic sea-levellowering. Paleoceanography 6, 537–541.
Rohling, E.J., 1994. Review and new aspects concerning the formationof eastern Mediterranean sapropels. Marine Geology 122 (1–2),1–28.
Rohling, E.J., 2001. The Dark Secret of the Mediterranean — A CaseHistory in Past Environmental Reconstruction. http://www.soc.soton.ac.uk/soes/staff/ejr/DarkMed/dark-title.html.
Rohling, E.J., Gieskes, W.W.C., 1989. Late Quaternary changes inMediterranean intermediate water density and formation rate.Paleoceanography 4 (5), 531–545.
Rossignol-Strick, M., 1999. The Holocene climatic optimum andpollen records of sapropel 1 in the eastern Mediterranean, 9000–6000 BP. Quaternary Science Reviews 18, 515–530.
Rossignol-Strick, M., Nesteroff, W., Olive, P., Vergnaud-Grazzini, C.,1982. After the deluge: Mediterranean stagnation and sapropelformation. Nature 295, 105–110.
Sancetta, C., 1994. Mediterranean sapropels: seasonal stratification yieldshigh production and carbon flux. Paleoceanography 9 (2), 195–196.
Savrda, C.E., 1992. Trace fossils and benthic oxygenation. In: Maples,C.G., West, R.R. (Eds.), Trace Fossils. Short Courses inPaleontology. Paleontological Society, pp. 172–196.
Savrda, C.E., Bottjer, D.J., 1986. Trace-fossil model for reconstructionof paleo-oxygenation in bottom waters. Geology 14, 3–6.
Savrda, C.E., Bottjer, D.J., 1989. Trace-fossil model for reconstructingoxygenation histories of ancient marine bottom waters: applicationto Upper Cretaceous Niobrara formation, Colorado. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 74, 49–74.
Savrda, C.E., Bottjer, D.J., 1991. Oxygen-related biofacies in marinestrata: an overview and update. In: Tyson, R.V., Pearson, T.H.(Eds.), Modern and Ancient Continental Shelf Anoxia. SpecialPublication, vol. 58. Geological Society of London, London,pp. 201–219.
Savrda, C.E., Bottjer, D.J., Gorsline, D.S., 1984. Development of acomprehensive oxygen-deficient marine biofacies model: evidencefrom Santa Monica, San Pedro, and Santa Barbara basins,California continental borderland. The American Association ofPetroleum Geologists Bulletin 68 (9), 1179–1192.
Schmiedl, G., Hemleben, C., Keller, J., Segl, M., 1998. Impact ofclimatic changes on the benthic foraminiferal fauna in the IonianSea during the last 330,000 years. Paleoceanography 13 (5),447–458.
Schmiedl, G., Mitschele, A., Beck, S., Emeis, K.-C., Hemleben, C.,Schulz, H., Sperling, M., Weldeab, S., 2003. Benthic foraminiferalrecord of ecosystem variability in the eastern Mediterranean Seaduring times of sapropel S5 and S6 deposition. Palaeogeography,Palaeoclimatology, Palaeoecology 190, 139–164.
Schnack, K., 1998. Besiedlungsmuster der benthischen Makrofaunaauf dem ostgrönländischen Kontinentalhang. Berichte zur Polar-forschung 294, 1–124.
Seilacher, A., 1957. An-aktualistisches Wattenmeer? PaläontologischeZeitschrift 31 (3/4), 198–206.
Seilacher, A., 1990. Aberrations in bivalve evolution related to photo-and chemo symbiosis. Historical Biology 3, 289–311.
Skinner, L.C., McCave, I.N., 2003. Analysis and modelling of gravity-and piston coring based on soil mechanics. Marine Geology 199,181–204.
Sternberg, K.M.G.v., 1833. Versuch einer geognostisch-botanischenDarstellung der Flora der Vorwelt, part 5–6. Fr. Fleischer, Leipzig,1–80 pp.

425L. Löwemark et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 406–425
Thierstein, H.R., Geitzenauer, K.R., Molfino, B., 1977. Globalsynchroneity of late Quaternary coccolith datum levels: validationby oxygen isotopes. Geology 5, 400–404.
Thompson, B.E., 1980. A new bathyal Sipunculan from southernCalifornia, with ecological notes. Deep-Sea Research 27A,951–957.
Thomsen, E., Vorren, T.O., 1984. Pyritization of tubes and burrowsfrom Late Pleistocene continental shelf sediments off NorthNorway. Sedimentology 31, 481–492.
Thunell, R.C., Williams, D.F., 1989. Glacial–Holocene salinitychanges in the Mediterranean Sea: hydrographic and depositionaleffects. Nature 338, 493–496.
Tomczak, M., Godfrey, J.S., 2002. Regional Oceanography: AnIntroduction. Pergamon. 391 pp.
Tuenter, E., Weber, S.L., Hilgen, F.J., Lourens, L.J., 2003. Theresponse of the African summer monsoon to remote and localforcing due to precession and obliquity. Global and PlanetaryChange 36 (4), 219–235.
van Santvoort, P.J.M., De Lange, G.J., Thomson, J., Cussen, H.,Wilson, T.R.S., Krom, M.D., Ströhle, K., 1996. Active post-depositional oxidation of the most recent sapropel (S1) insediments of the eastern Mediterranean Sea. Geochimica etCosmochimica Acta 60 (21), 4007–4024.
Vergnaud-Grazzini, C., Ryan, W.B.F., Cita, M.B., 1977. Stableisotopic fractionation, climate change and episodic stagnation inthe eastern Mediterranean. Marine Micropaleontology 2,353–370.
Waelbroeck, C., Labeyrie, L., Michel, E., Duplessy, J.C., McManus, J.F., Lambeck, K., Balbon, E., Labracherie, M., 2002. Sea-level anddeep water temperature changes derived from benthic foraminiferaisotopic records. Quaternary Science Reviews 21, 295–305.
Warning, B., Brumsack, H.-J., 2000. Trace metal signatures of easternMediterranean sapropels. Palaeogeography, Palaeoclimatology,Palaeoecology 158 (3–4), 293–309.
Weaver, P.P.E., Schultheiss, P.J., 1983. Vertical open burrows in deep-sea sediments 2 m in length. Nature 301, 329–331.
Werner, F., 1967. Röntgen-Radiographie zur Untersuchung vonSedimentstrukturen. Umschau in Wissenschaft und Technik 16,532.
Werner, F., 1985. Sedimentary structures and the record of trace fossilsUpper Quaternary marine Skagerrak deposits. Norsk GeologiskTidsskrift 65 (1–2), 65–71.
Werner, F., 1996. Abschluβbericht zum DFG-ForschungsvorhabenMorphologie und Ökologie rezenter Lebenspuren in der Tiefsee.Az: We 564/10-Zeitraum 1991–1993. Geologisch-Paläontolo-gisches Institut, Kiel.
Wetzel, A., 1983a. Biogenic structures in modern slope to deep-seasediments in the Sulu Sea basin (Philippines). Palaeogeography,Palaeoclimatology, Palaeoecology 42, 285–304.
Wetzel, A., 1983b. Biogenic sedimentary structures in a modernupwelling region: northwest African continental margin. In:Thiede, J., Suess, E. (Eds.), Coastal Upwelling and its SedimentRecord, Part B, Sedimentary Records of Ancient CoastalUpwelling. Plenum, New York, pp. 123–144.
Wetzel, A., 1991. Ecologic interpretation of deep-sea trace fossilcommunities. Palaeogeography, Palaeoclimatology, Palaeoecology85, 47–69.
Wetzel, A., 2002. Modern Nereites in the South China Sea—ecological association with redox conditions in the sediment.Palaios 17 (5), 507–515.





![vf[ky Hkkjrh; vk;qfoZKku laLFkku] iVuk Corrigendum - AIIMS ...](https://static.fdokumen.com/doc/165x107/631a41301e5d335f8d0b7fb5/vfky-hkkjrh-vkqfozkku-lalfkku-ivuk-corrigendum-aiims-.jpg)





![pkS/ kjh j.kchj flag fo'ofo|ky;]thUn - CRSU](https://static.fdokumen.com/doc/165x107/63355175d2b728420307dd5d/pks-kjh-jkchj-flag-foofokythun-crsu.jpg)








