
Abnormal adhesion and cell-cell interactions in sickle cell disease

1
S. PYOGENES SER/THR KINASE-REGULATED CELL WALL HYDROLASE IS A CELL DIVISION-PLANE-RECOGNIZING AND CHAIN-FORMING VIRULENCE FACTOR*
Vijay Pancholi* Gregory Boel¶, Hong Jin, From Department of Pathology1, The Ohio State University College of Medicine,
Columbus, OH 43210-1214 Running title: CdhA, a GAS chain-forming virulence factor
Address correspondence to:Vijay Pancholi, Ph.D., 420 W 12th Ave, TMRF-288A, Columbus, OH 43210-1214, Tel: 614-688-8053, Fax: 614-688-3192, Email:[email protected] Cell-division and -wall synthesis are closely linked complex phenomena and play a crucial role in the maintenance and regulation of bacterial virulence. Eukaryotic-type Ser/thr kinases reported in prokaryotes including GAS (SP-STK) regulate cell division, growth and virulence. The mechanism of this regulation is, however, not known. In this study, we demonstrated that SP-STK-controlled cell division is mediated under the positive regulation of secretory protein that possesses a CHAP-domain with functionally active cell wall hydrolase activity (henceforth named as CdhA (CHAP-domain-containing and chain-forming cell-wall hydrolase). Deletion of the CdhA-encoding gene, resulted in severe cell division and growth defects in GAS mutants. The mutant expressing the truncated CdhA (devoid of CHAP domain) although displayed no such defects became attenuated for virulence in mice and highly susceptible to cell wall-acting antibiotics as observed for the mutant lacking CdhA. When CdhA was overexpressed in the wild-type GAS as well as in heterologus strains; E. coli, and S. aureus, we observed a distinct increase in bacterial chain length. Our data reveal that CdhA is a multifunctional protein with a major function of is N-terminal region as a cell division-plane-recognizing domain and its C-terminal CHAP-domain as a virulence-regulating domain. CdhA is thus an important therapeutic target.
Group A Streptococcus (GAS, S.
pyogenes), a human pathogen, is known to cause the broadest range of diseases of any known pathogen (1). These diseases range from mild pharyngitis and impetigo to severe, debilitating and often fatal toxic shock syndrome and necrotizing fasciitis (2,3,4). Morbidity and mortality caused by GAS-associated autoimmune sequelae, rheumatic heart disease
(5,6) and glomerulonephritis (1,7), especially in underdeveloped and developing countries (8), continue to challenge us clinically and scientifically (9,10). GAS expresses an array of surface bound and secreted proteins, which either alone or in concert play important roles in bacterial adhesion, invasion and further proliferation in a variety of constantly fluctuating host environments (11). The expression of these proteins is also bacterial growth-dependent, and controlled by the two-component regulatory systems (TCSs) composed of an environmental sensor, histidine kinase, and a response regulator which serves as a transcription factor (12). In other words, TCSs play an important role in regulating genes required for successful colonization and infection by GAS. We, previously, identified a novel one-component signaling system that is structurally very similar to eukaryotic-type serine threonine kinase (ESTK) (13). GAS mutants lacking ESTK (SP-STK) are defective in cell division and growth but display increased production of capsule and hemolysin (13) indicating that SP-STK, which cotranscribes with serine threonine phosphatase (SP-STP), positively regulates bacterial growth and cell division, while negatively regulates bacterial capsule and hemolysin production (13), which is reminiscence of the CovR-mediated regulation (14). The phenotypic characteristics of SP-STK mutant mimic those reported for Gram-positive pathogen mutants that lack S. pneumoniae/S. agalactiae PscB orthologs (15,16,17). In GAS, PscB ortholog is SPy0019, a secreted protein of unknown function. The purified SPy0019 has been previously reported to possess a human immunoglobulin binding ability, and hence was termed as SibA (immunoglobulin-binding protein of group A Streptococcus) (18). Besides pneumococcal and group B streptococcal PcsB (15,16,17), SPy0019 is structurally similar to Streptococcal mutans
http://www.jbc.org/cgi/doi/10.1074/jbc.M110.153825The latest version is at JBC Papers in Press. Published on July 19, 2010 as Manuscript M110.153825
Copyright 2010 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
by guest on July 22, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

2
GbpB/SagA (19), Streptococcus thermophilus Cse (20), Enterococcus faecalis/faecium (SalB/SagA) (21,22) and Listeria monocytogenes P45 protein (23). All these proteins are involved in some way in the regulation of cell division. However, the exact function of these proteins in bacterial cell division and in particular SPy0019 in GAS cell division and pathogenesis is not known. Since GAS mutants lacking SP-STK display defective cell division (13), we hypothesized that SP-STK regulates GAS cell division by regulating the expression of SPy0019. Further, protein domain analysis of SPy0019 and its orthologs revealed the presence of CHAP [Cysteine, Histidine-dependent Aminohydrolases/Peptidases] domain at its C-terminal end (24). The ubiquitous presence of this domain and its possible cell wall hydrolase activity can be argued in favor of the fact that these structurally related proteins may have a specific role in cell wall synthesis. The latter is closely linked with cell division. However, the role of the CHAP domain in cell division, cell wall synthesis or bacterial pathogenesis has not been proven conclusively partly because of the fact that these proteins are found to be essential for bacterial survival. Available studies are therefore limited to conditional mutants or mutants with depleted protein expression (15,16). In this report, we characterized the structure and functions of SPy0019, phenotype of domain specific GAS mutants, and their possible roles in cell division and pathogenesis of GAS. In this study, we have further demonstrated the role of this protein in bacterial chain formation. Cumulatively, our findings indicated that SPy0019 by serving as a GAS cell- division-plane recognition protein, plays a critical role in the regulation of early events in cell division process which in turn help regulate cell wall synthesis and ultimately virulence. In particular we show that the N-terminal portion of SPy0019 plays a significant role in cell division and bacterial growth while the CHAP-domain plays a significant role in the regulation of GAS virulence but not in cell-division or -wall synthesis. We, therefore, preferred to term SPy0019 henceforth as CdhA (CHAP domain-containing and chain-forming cell wall hydrolase from group A Streptococcus).
Experimental Procedures
Bacterial strain, growth conditions, and cell culture-The wild-type GAS strain M1-SF370 (ATCC 700294) (25) and the derived mutants were grown at 37oC in Todd-Hewitt broth (Difco Laboratories) supplemented with 0.5% (w/v) yeast extract (THY) or on proteose peptone blood agar plates with or without spectinomycin (500 μg/ml) (13). Staphylococcus aureus RN4220 was grown in trypticase soy broth (TSB) or agar (TSA) as described previously (26). Escherichia coli strain XL1-Blue and BL21 (DE3 pLysS), were grown in Luria-Bertani (LB) broth or agar at 37oC. The N-terminal His-tagged expression vector pET14B (Novagen) was used to make recombinant 6xHis-tagged proteins. Vector pFW5 containing the spectinomycin gene (aad9) was used to transform M1-SF370 as described before (27,13,28). Vector pDC123 (29) and pCN40tet (26) were used for the complementation experiments within M1-SF370 and S. aureus RN4220, respectively. Culture plates and media supplemented with ampicillin (100 μg/ml), spectinomycin (500 μg/ml), chloramphenicol (10 μg/ml) and tetracycline (12μg/ml) were used for the selection of mutants or for construction of complemented mutant strains. Detroit 562 human pharyngeal cell lines (ATCC CCL138) were cultured and maintained in MEM with 10% FBS at 37 oC in a humidified CO2 incubator as described previously (13). Production of His-tagged recombinant CdhA protein and affinity purified anti-CdhA rabbit polyclonal antibodies-Recombinant CdhA and corresponding rabbit polyclonal antibodies were made essentially as described previously (13). Briefly, using M1-SF370 genomic DNA as template, the cdhA gene (encoding 1125 bp excluding the first 72 bp which encodes for a signal sequence) was amplified by PCR using gene specific primers (Primer-1 [GBSPy19-F] and Primer-2 [GBSPy19-R]) sequentially digested with XhoI and BamHI and cloned into pET14B vector digested with the same enzymes to obtain pETCdhA. E. coli XL1-Blue cells were transformed with pETCdhA, and the recombinant His-tagged CdhA (rCdhA) protein was expressed in E.coli BL-21 (DE3-pLysS) strain after IPTG induction and purified by Ni+2-NTA affinity column chromatography (Qiagen) under denaturing condition using 8M urea as per the manufacturer’s instructions. The purified
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

3
protein was then renatured by dialyzing against 50 mM ammonium bicarbonate pH 8.6, concentrated, and redialyzed against 50 mM Tris/HCl pH 8.0. Rabbit polyclonal antibody against rCdhA was custom-raised by Lampire Biological Laboratories (Pipersville, PA) and was purified by sequential Protein G and CdhA-DADPA affinity column chromatography as described previously (13). Fab-specific anti-CdhA antibody fragment after pepsin digestion was obtained as described previously (28). Construction of cdhA deletion mutants-From the wild-type M1-SF370 strain, two CdhA-specific isogenic mutant strains were created. To create a mutant lacking the entire CdhA (M1ΔCdhA), an upstream 930 bp PCR products (including first 61 nucleotides of SPy0019) obtained with Primer-3 (GB19Bam-UpNt) and Primer-4 (GB19Hin-UpCt), and a downstream 774 bp PCR product (including the last 267 nucleotides of SPy0019) obtained with Primer-5 (GB19NcoDnNt) and Primer-6 (GB19PstDnCt) were digested with BamH1/HindIII and NcoI/PstI for upstream and downstream fragments, respectively (For all primer sequence see Table S1). They were inserted serially into the MCS-I and MCS-II of pFW5 digested with the same enzymes (27) to obtain plasmid pFW5ΔcdhA. The resultant plasmid was then used to transform the wild-type GAS strain M1-SF370 by electroporation to obtain M1ΔcdhA mutant as a result of a double crossover homologous recombination event which was confirmed by PCR and DNA sequencing using flanking primers of the genomic DNA (primer 7 [upNT-Pr3] and primer 8[dnCT-Pr6]) and aad9 specific primers (primer 9 and primer 10) in a primer pair combinations of 7/10 and 9/8. To elucidate the function of the C-terminal domain of CdhA, a mutant (M1CdhAΔC55, Table-1) expressing a truncated form of CdhA lacking the last 55 amino acids (the catalytic portion of the CHAP domain (24)) was constructed. An internal fragment of 1,075bp size (from nucleotide 157 to 1231 of cdhA) generated by PCR using Primer-11 (GB19C55-F) and Primer-12 (GB19C55-R) was cloned into BamH1/HindIII sites of MCS-I of pFW5. The resultant plasmid pFW5cdhAΔC55 was then used to transform the wild-type GAS strain, M1-SF370, by electroporation yielding M1CdhAΔC55 mutant (Table-S2). Integrity of
the mutants was verified by colony PCR employing primer pairs 7/10 and 9/8 or 9/2 as described above and further confirmed by DNA sequencing. Construction of plasmid for complementation of cdhA and over expression of CdhA-The wild-type cdhA gene along with its native promoter (including 100 bp upstream region) was amplified by PCR using Primer-13 (GB19PDCHin-F) and Primer-14 (GB19PDCKpn-R) and cloned into pDC123 (29) between HindIII and KpnI sites to obtain pDC123-cdhA. The insert was ligated in the plasmid in the opposite orientation of the cat gene to allow the expression of cdhA by its native promoter. To understand the role of CdhA in SP-STK-mediated regulation of cell division, the latter was used to transcomplement M1ΔSTK (13) as well as M1�CdhA to obtain M1ΔSTK::cdhA and M1ΔCdhA::cdhA respectively as described before (13). To understand the innate function of CdhA, the wild-type strain M1-SF370 (M1-WT) was similarly transformed with pDC123-cdhA to obtain M1-WT::cdhA. The effect of the over expression of CdhA on cell division in other species of gram-positive and gram-negative bacteria was studied by complementing cdhA in S. aureus RN4220 and E.coli BL21 respectively. For S. aureus, the wild-type cdhA gene along with 100 bp upstream region containing its native promoter was cloned into pCN40tet between XmaI and KpnI sites as described recently (26) (See Table S2). The effect of over expression of CdhA on cell division in E. coli BL21 cells under IPTG induction was measured as described above. Microscopy-Growth curve patterns of the wild-type, mutants, and complemented GAS strains were monitored for 16h in THY broth. At the late log phase of growth, the GAS strains were harvested, washed and subjected to light (Gram stain, phase-contrast) and fluorescence microscopy followed by scanning and transmission electron microscopy. For fluorescence microscopy, GAS strains were adsorbed onto the poly-L-lysine coated glass slides for 30 min, washed, and stained with CdhA-specific IgGFab (1:50 dil) followed by FITC labeled goat-anti rabbit IgG (Fab’-specific) (BioRad 1:50). Stained bacteria were
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

4
kept in Slow FadeTM equilibrium buffer and stained with DAPI (1μg/ml) for 5 min and were visualized by fluorescence microscopy (Nikon Eclipse E600). For electron microscopy, the wild-type, mutant, and complemented streptococcal strains were grown till late log phase in THY broth, washed once with phosphate buffer saline, and fixed in 0.1 M cacodylate buffer containing 2.5% glutaraldehyde and 4% paraformaldehyde for 12-16 h at 4 oC. Samples were then processed for transmission and scanning electron microscopy at the OSU Campus Microscopy and Imaging Facility equipped with NOVA nanoSEM 400 scanning electron microscope with secondary and low vacuum detectors and cryo-capable digital TEM (Technai G2 Spirit , EFI) as described previously (13). Human IgG-binding assay-To determine changes in human immunoglobulin binding activity as a result of the deletion of CdhA/SibA/SPy0019, hIgG-binding assay was carried out essentially as described previously with minor modifications (30). Briefly, wild-type, mutant and cdhA-complemented GAS strains were suspended in 0.05 M Tris/HCl buffer, pH8.0 containing 0.5 % BSA to an O.D.600 1.00. Equal volumes of the bacterial suspension and a serially diluted hIgG solution (Sigma, starting concentration 100 μg) were mixed in a final volume of 200 μl in 96-deepwell plates for 1 h followed by same volume of alkaline phosphatase-conjugated monoclonal antibody against anti-hIgG (Sigma, 1:3000) for 1 h. Bacteria were thoroughly washed with the suspension buffer after every step and finally resuspended in 0.05 M Tris/HCl pH 10.0 containing 5mM MgCl2. The contents of each well were then transferred to a new 96-well microtiter-plate and centrifuged. Indirect measurement of bacteria-bound hIgG was determined spectrophotometrically after adding 200 μl of ready-to-use phosphatase substrate (Sigma) in each well and incubating further for 1 h using a 96-well plate reader (PolarStar Galaxy, BMG). The binding of bacteria with different concentrations of hIgG was carried out in triplicates in three independent experiments. The values obtained for reactions carried out with only conjugate antibody were treated as back ground values and were subtracted from the test values. The results were statistically evaluated
by non-parametric 2-way ANOVA and ‘p’ values were determined using GraphPad-Prism 4 software. Streptococcal peptidoglycan preparation-Peptidoglycan from the over-night grown culture of the wild-type M1SF370 strain was extracted as described previously with some modifications (31,26). Briefly, the bacterial pellet obtained from the overnight grown culture was chilled and suspended in 4% SDS and boiled for 30 min and centrifuged at high speed. The pellet of crude cell wall was then broken using FastPrep20 (MP biomedical) and subjected to sequential treatment of amylase, DNAse/RNAse followed by trypsin to remove cell wall-associated carbohydrate, nucleic acid and surface proteins, respectively. Subsequently, by an additional step of boiling, all these enzymes were inactivated. Other cell wall-associated lipid/protein components were then released and removed by sequential treatment of 8M LiCl, EDTA, water, and acetone followed by lyophilization. The acetone powder of cell wall was then treated with hydrofluoric acid followed by alkaline phosphatase to remove lipoteichoic acid and boiling in water bath for 5 min. Peptidoglycan thus obtained was dried, stored at 4oC, till further use in subsequent zymogram assays. Zymogram assay-Cell wall hydrolase zymogram assay was performed using SDS-PAGE and renaturing SDS-PAGE essentially as described previously (32) with the following modifications. S. pyogenes M1-SF370 peptidoglycan (0.5% w/v) was included in 10% polyacrylamide gels for detection of cell wall hydrolase activity of CdhA in culture supernatants. Culture supernatants (1.6 ml) from each of the late log phase cultures of wild-type, CdhA-specific two mutants and cdhA-complemented M1ΔCdhA mutant strains were separately collected and precipitated overnight with 80 % of ammonium sulphate saturation at 4oC. The resulting precipitates were centrifuged (4oC, 10,000x g, 40 min) and solubilized in a fixed volume of sample loading buffer. Proteins were then resolved on the polyacrylamide gel containing S. pyogenes peptidoglycan. After electrophoresis, the gels were washed twice in distilled water at room temperature with gentle shaking and incubated at 37oC (with shaking) in
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

5
40 ml of renaturing buffer (50 mM MES [2-morpholinoethansulfonic acid]/NaOH, pH 6, 0.2% [w/v] Triton X-100) for 2 days with periodic buffer replacement after every 5-6 h. Gels were stained with 0.1% (w/v) methylene blue in 0.01%(w/v) KOH for 2 h at room temp with gentle shaking followed by destaining with distilled water as described previously (32). The clear band on a blue gel was perceived as cell wall hydrolase activity. Stained gels with clear bands were scanned and the images were inverted to visualize the clear bands as dark bands, using Photoshop software (Adobe). Bacterial adherence assays-GAS adherence to Detroit 562 human pharyngeal cells (ATCC CCL138) was determined essentially as described previously (13). Briefly, adherence assays were performed for a period of 3h using confluent culture of Detroit 562 cells (grown in 24-well tissue culture plates) which were infected with the wild-type and mutant strains (multiplicity of infection, 50:1::bacteria:cell). Before infecting the confluent cell culture, washed bacteria were washed sonicated gently on ice to disrupt the aggregates. The number of colony forming units in this preparation was determined prior to performing this experiment. Adherent bacteria were removed by repeated washing and were counted on proteose peptone blood agar plates as colony forming units (CFUs). Phagocytosis/bactericidal assays-GAS ability to resist phagocytosis and survive in human blood was analyzed in bactericidal assay as described previously (13). Briefly, 50 μl (~150-200 CFUs) GAS grown at the late log-phase was added to freshly drawn heparinized blood (1 ml) in sterile glass tube and incubated at 37oC for 3h under constant gentle mixing. The number of CFUs in the inoculum and those obtained after incubation in human blood were analyzed by the pour plate method using sheep blood agar plate. M1Δemm1 mutant strain (13) lacking the M protein (SPy2018) was used as a positive control. Bacterial growth (multiplication factor, MF) was calculated as the mean number of CFU after 3h divided by the initial mean number of CFU (t=0).
Approved Ohio State University IRB protocol was obtained to draw blood from healthy non-smoking individuals/volunteers (18-
65 yrs of age). A written informed consent was provided by study participants. Estimation of hyaluronic acid of GAS capsule-The amount of hyaluronic acid capsule of the wild-type and mutant GAS strains was measured spectrophotometrically (640nm) using the standard curve generated with known concentrations of hyaluronic acid and Stain-All chromogenic reagent (Sigma, 1-ethyl-2-[3-(1-ethyl-naptho-[1,2-d]thiazolin-2-ylidene)-2-methylpropenyl]-naptho-[1,2-d]thiazolium bromide) in a final volume of 50μl as described earlier(13). Antibiotic sensitivity assay-Susceptibility of the wild-type strain and the isogenic M1ΔCdhA and M1CdhAΔC55 to various antibiotics was determined using the E-TestR method (AB BIODISK North America inc. Piscataway, NJ). For this, antibiotic-specific Etest strips containing gradient of five representative antibiotics with MIC range as indicated (benzylpenicillin-PG [MIC 0.002-32 μg/ml], cefotaxime-CT [MIC 0.016-256 μg/ml], erythromycin-EM [MIC 0.016-256 μg/ml], and clindamycin-CM [MIC 0.016-256 μg/ml], ciprofloxacin-CI [MIC 0.002-32 μg/ml]) were used. Inoculum preparation, media, usage of E-test antibiotic strips, incubation time, and interpretation of the results obtained for bactericidal and bacteriostatic antibiotics, were according to the manufacturer’s instructions and essentially as described (26). Bacterial virulence assay-In vivo virulence of the wild-type and mutant strains was measured in the mouse peritonitis model. A group of ten mice (BALB/C, 5-week old, 20-22 g, Charles River Laboratories) were injected intraperitoneally with 2x108 CFU. Morbidity and mortality in the infected animals were observed twice daily for 10 days. The data presented in mortality/survival curve were statistically evaluated using the Log-rank test using GraphPad Prism4 software. The dissemination of GAS in liver, spleen, kidney and lungs was determined by enumerating the CFUs on blood agar plates as beta-hemolytic colonies. All animal experiments were performed per the OSU-IACUC-approved protocol. The infected animals developing the sign and symptoms of extreme distress (e,g. due
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

6
to to increased bacteremia after I/P infection), such as gasping, asyphyxia, apnea, curved body posture with limited movements, low heart beats, within 24 hr post-or during time of observation after infection were euthanized for further study. SDS-PAGE and Western blotting- Procedures of SDS-PAGE of whole bacterial extracts and electro-transfer of resolved proteins on PVDF membranes were performed as described (13). Western blots were developed using Protein-G affinity-purified rabbit polyclonal anti-CdhA and anti-M6 (10B6) mouse monoclonal antibody (33) followed by corresponding alkaline phosphatase-labeled goat anti rabbit/mouse conjugate antibodies to detect the expression level of CdhA and the M protein in culture supernantants/cell walls of GAS obtained at early, mid, and/or late-log phases. Specific-reactive bands were visualized using alkaline phosphatase-specific chromogenic substrates (13). Microarray analyses and quantitative real-time PCR-The complete/whole M1 genome (25) oligonucleotide-based microarray was designed, synthesized and made by Illumina Inc. Oligonucleotides (50-60 mer) were designated to hybridize 1,769 ORFs in the genome of S. pyogenes type M1 strain SF370 (25). The oligonucleotides were spotted onto poly L-lysine microarray slides by a robotic microarrayer (Omni GRAD 100 microarrayer) of the Center for Applied Genomics (CAGE) at the Public Health Research Institute/UMDNJ, Newark, NJ.
Microarray analyses were performed to determine relative transcript abundance of M1ΔCdhA and M1CdhAΔC55 with M1SF370. Bacteria were grown in Todd-Hewitt broth till late log phase, harvested by centrifugation and washed twice with sterile PBS. Total RNA was isolated from these washed streptococcal strains using Qiagen RNeasy Mini kit. High purity and integrity of RNA was confirmed by Agilent 2100 Bioanalyzer (Agilent technologies, Palo Alto, CA). Synthesis and labeling of cDNA, was carried out using 20 μg of total RNA, 1.5 μg of random hexamer, 0.5mM dNTPs (except that 0.2mM of dTTP was replaced by the same amount of amino-allyl dUTP to incorporate dUTP into first-strand cDNA) were used in each
reverse transcriptase reaction (Superscript II Reverse Transcriptase, Invitrogen).
Purified cDNA preparations from the wild-type, M1ΔCdhA, and M1CdhAΔC55mutant strains were labeled with either Alexafluor-555 or Alexafluor-647 depending on the experimental design (i.e. dye swap experiment). Differentially labeled probes were then combined and purified. Using three independently isolated RNA preparations from each mutant (biological replicates), a total of eleven experiments for M1ΔCdhA strain (incorporating dye swaps) and seven experiments for M1CdhAΔC55 strain were performed. Signals of the bound reagents on the microarray spots in terms of relative fluorescence values were measured and quantified by a laser scanner (GenePix 4100) at 10μm/pixel resolution. The resulting images were processed using Gene Pix Pro software (version 4.0, Axon instruments).
The web application CARMAweb (Comprehensive R based microarray analysis web service) was used for the normalization and analysis of microarray data (34). All raw data (in the form of *. GPR files) were uploaded to the web application in the data directory. Using appropriate navigation tree, background correction from the foreground signal was applied and within microarray normalization was achieved using the Lowess method. Genes flagged as bad spots by the scanning software were excluded from the analysis and all flagged spots were given a weight of zero. The normalized data were then subjected to fold-change analysis and t-statistics using Bioconductor multtest package. Differentially expressed genes were defined based on cut-off value >1 Log2< with P < 0.05. All microarray data (intensity and expression ratio) for 11 M1ΔCdhA experiments and 7 M1CdhAΔC55 experiments are deposited in GEO database (Http://www.ncbi.nlm.gov/geo) (accession number for M1ΔCdhA-GSE15546, M1CdhAΔC55- GSE15598).
The expressed mRNA levels from microarray analysis were confirmed by real-time PCR using 1μg of total RNA in reverse transcriptase reaction to synthesize first-strand cDNA according to the manufacturer’s instruction (Roche RT kit). Real-time PCRs for selected genes were performed using Brilliant SYBR Green QPCR Master Mix (Roche) and
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

7
specific primers (Table- S3) and conducted using LightCycler®480 (Roche) real-time PCR instrument. The copy numbers for all the genes were normalized with gyrA using at least three biological replicates and results were analyzed using Exor4 software (Roche) (35). Statistical analysis Unless otherwise indicated, statistical analysis was carried out from at least three independent experiments and based on nonparametric two-tailed student’s t-test with Welch’s correction using GraphPad prism-4 software.
RESULTS SP-STK positively regulates CdhA, a CHAP-domain containing streptococcal protein-Based on our earlier reports, GAS lacking a eukaryotic-type serine threonine kinase (M1ΔSTK) is defective in cell division (13). Phenotypic characteristics of this mutant were found to mimic growth- and cell division-defective S. agalactiae (group B Streptococcus, GBS) and S. pneumoniae mutants lacking PcsB protein (36,15), S. mutans lacking GbpB protein (19,37) E. faecalis/faecium lacking SagA/SalA(22,21). PcsB, GbpB, and Saga/SalA are orthologs and reported to possess similar structural characteristics including the CHAP-domain at their C-terminal ends (24,38,39). Based on this previous study, we hypothesized that the cell division defects observed in the M1ΔSTK mutant are likely due to the down-regulation of a PcsB ortholog SPy0019/CdhA. To confirm this, first, we complemented M1ΔSTK GAS mutant with the wild-type spy0019 and observed by light microscopy that the cell division defects of M1ΔSTK were restored to the wild-type chain-forming strain. The complemented strain displaying normal chain forming ability indicated that SP-STK directly or indirectly regulates the expression of CdhA. In the subsequent experiments, by employing real time qRT-PCR, we observed a significant decrease in the cdhA transcript abundance in the M1ΔSTK mutant strain (Fig. 1) and thus established a direct link between SP-STK and CdhA. Earlier on CdhA/SPy0019 was identified as SibA because of its ability to bind to human immunoglobulin (18). However, apart from this function, the physiologically relevant role of this protein in GAS has not been described.
CdhA regulates GAS growth-Based on the structural similarity of CdhA with six other Gram-positive orthologs (36,15,19,37,22,21), we predicted that the N-terminal coiled-coil portion may have a role in cell division/separation/aggregation and the C-terminal containing CHAP domain may have a cell wall-hydrolysis /synthesis function. CdhA possesses a typical motif of the CHAP-domain at its C-terminal end ( SS-7X-C-13X-G-30X-G-10X-G-5X-YGH*VXVV, where X represents non-conserved amino acid and H* is a putative catalytic histidine residue) (24,38,39). Since the precise functional roles of these regions in Gram-positive pathogens in general and GAS physiology or pathogenesis in particular are not known, we created two mutants using pFW5 suicide vector; one with complete deletion of the cdhA gene (M1ΔCdhA) and the second with deletion of the region corresponding to the C-terminal CHAP domain (M1CdhAΔC55). The latter expressed truncated CdhA (aa1-343) lacking the last 55 amino acids constituting the conserved and putative catalytic histidine residue (His354) responsible for conferring the cell wall hydrolase activity (24). Growth characteristics of the mutants (M1ΔCdhA, M1CdhAΔC55, and M1ΔSTK) and the cdhA- complemented strains (M1ΔCdh::cdhA, M1-WT::cdhA, M1ΔSTK::cdhA) determined over a period of ~17 h revealed unique patterns (Figs. 2A and 2B). Among all mutants, M1ΔCdhA exhibited retarded growth attaining a maximum O.D.600 of 0.62 in ~8 h. This mutant was unable to grow further and in fact showed a decrease in the O.D. (0.42) after an overnight incubation (~17 h), indicating a progressive lysis of the mutant. The corresponding complemented strain (M1ΔCdhA::cdhA) grew faster and displayed a maximum O.D. of 1.0 (vs. wild-type O.D.600nm ~1.3) indicating no degradation in the overnight grown culture and hence the restoration of CdhA function. The growth patterns of mutants expressing the truncated CdhA (lacking the CHAP-domain containing C-terminal 55 residue (M1CdhAΔC55), and that of the wild-type strain were comparable. M1CdhAΔC55, despite being devoid of CHAP domain with putative cell wall synthesis function, in fact, grew faster than the CdhA-overexpressing complemented strain, M1-WT::cdhA. This indicated that the growth defect observed in M1ΔCdhA is primarily due to the
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

8
absence of the N-terminal region of CdhA. The growth pattern of M1ΔSTK, which displayed slow growth as reported previously (13), when complemented with cdhA, grew as fast as the wild-type strain indicating that the slow growth observed of the M1ΔSTK mutant (13)could be due to the down regulation of cdhA. Western blot analysis of the recombinant CdhA protein and the proteins present in the whole cell lysates of the strains described above (Fig. 2A-2B) using affinity purified monospecific anti-CdhA IgG showed the presence of 43kDa CdhA-specific reactive protein band in the wild-type and the cdhA-complemented strains and its absence in the M1ΔCdhA and M1ΔSTK mutant strains, thus confirming the integrity of these strains also at the protein levels (Fig 2C). The truncated CdhA found in the M1CdhADC55 mutant despite the loss of the C-terminal 55 residue migrates similar to wild-type CdhA (Fig 2C, lane 4). The recombinant truncated CdhA (aa 1-270) also migrates similar to the wild-type (data not shown) indicating that the CHAP-domain of CdhA does not contribute to the migration of the whole protein in the SDS-PAGE. CdhA regulates GAS cell division and belongs to a class of proteins that participates in the formation of cell septum and determines bacterial chain length.-Indirect immunofluorescence-microscopy of the wild-type and M1ΔCdhA mutant strains using FITC-labeled affinity purified CdhA-specific IgG (Fab’-fragment) revealed localization of CdhA protein primarily in the septum (Fig. 3A, see inset panel I ). Further, the absence of CdhA resulted in aggregation of the bacterial cells as shown in Fig. 3A (DAPI stain panel II). Electron microscopy of M1ΔCdhA clearly revealed defective cell division resulting in the formation of incomplete and unorganized septa and in turn bacterial aggregates (Fig.3B). This random septa formation resulted in the generation of ghost cells devoid of the genetic content. These cells were confirmed as dead by fluorescence microscopy using live-dead bacterial stain (Fig. 3 A, panel IV). As shown in Fig. 3B, M1CdhAΔC55 mutant strain expressing truncated CdhA did not display defect in cell division and the cdhA-complemented mutant strain (M1ΔCdhA::cdhA) reverted back to the
wild-type phenotype supporting our hypothesis that the cell division function is restricted to the N-terminal region. Further, as was also reported for PscB protein of S. pneumoniae (15,16) CdhA-overexpressing M1-WT::cdhA strain displayed increased in chain length confirming that CdhA indeed plays an important role in the chain formation in GAS (Fig 4 A). However, the SEM- and TEM-based analyses of M1ΔSTK::cdhA revealed striking differences in the cell division restoration pattern. M1ΔSTK::cdhA strain primarily showed increase in only chain length, however the organized cell septation was not completely restored indicating the primary function of SPy0019/CdhA is more likely in the recognition of the division plane but not the whole complex phenomenon of cell division governed by a battery of proteins (40,41,42) and possibly governed directly or indirectly by SP-STK (13). To confirm this in heterologous bacterial species, we examined the CdhA-over expressing E.coli BL21 strain before and after IPTG induction. It was evident that upon CdhA expression, the cococbacillary form of uninduced E. coli was indeed transformed into the long chain-forming bacilli (Fig. 4B). Further, the cdhA-complemented S. aureus RN4220 (which is known to divide in X, Y, and Z planes and form clusters) using a S. aureus-compatible complementation vector, pCN40tetR (26) also formed chains with intermittent kinks (Fig. 4C). Together, light microscopy, SEM and TEM of the wild-type, mutant, and complemented strains confirmed unequivocally that N-terminal domain of CdhA is responsible for the cell-division plane-recognition and chain-formation. CdhA does not contribute significantly to the overall GAS ability to bind human immunoglobulins (hIgG)-SPy0019 is a secretory protein and was previously termed as SibA based on its ability to bind human immunoglobulin-(hIgG, hence named SibA) (18). Fluorescence microscopy with anti-SPy0019 antibody indicated that SPy0019/SibA is located both at the septum and on the surface. We, therefore, hypothesized that hIgG-binding ability of GAS may be attributed to SPy0019 and the corresponding mutant lacks hIgG-binding ability. In fact, to our surprise, the bacterial-ligand binding assays revealed significantly increased hIgG-binding activity in
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

9
the mutant strains as compared to that in wild-type (P<0.0001) increased as compared to that of the wild-type indicating that the absence of CdhA on the surface somehow expose the native GAS hIgG binding activity (Fig. 5). This hIgG-binding activity of the mutant was not reduced by upon complementation with the wild-type cdhA gene possibly due to homologous functions possessed by CdhA and other bacterial immunoglobulin binding protein. Together these results indicate that while CdhA is indeed a hIgG binding protein, it does not primarily contribute to the overall GAS hIgG binding activity. CdhA hydrolyzes GAS cell walls- The CHAP domain-specific cell-wall hydrolase/amino peptidase activity for CdhA orthologs (15,18,19,21,22) in Gram-positive organisms has not been reported so far. To determine the putative peptidoglycan hydrolase activity of CdhA, the recombinant CdhA protein and ammonium sulphate precipitates of culture supernatants were subjected to renaturing zymogram analysis using purified M1SF370 peptidoglycan, since our initial attempts using acetone powder of Micrococcus luteus and S. pygenes M1SF370 did not yield detectable activity (data not shown). As shown in Fig 6, the recombinant CdhA protein, and the ammonium sulphate precipitates of culture supernatants of the wild-type (M1-WT, M1ΔCdhA::cdhA, and M1-WT::cdhA showed 43 kDa protein-associated hydrolysis of the GAS peptidoglycan. The absence of similar activities in M1ΔCdhA and M1CdhAΔC55 supernatants clearly indicated that the CHAP-domain of CdhA indeed possesses cell wall hydrolase activity. Absence of CdhA adversely affects GAS adherence to human pharyngeal Detroit 562 cells-Electron microscopy of M1ΔCdhA and M1CdhAΔC55 revealed significant reduction in the size of surface electron dense fuzzy layer (Fig. 3B). The presence of the latter is essentially attributed to many surface proteins (43,44) that play a crucial role in GAS adherence to host cells (45). We therefore speculated that the reduced expression of crucial surface proteins may significantly affect the bacterial adherence ability to host cells. Direct bacterial adherence assays, using Detroit 562
pharyngeal carcinoma cell lines, detected significantly reduced (CFU/well, P<0.001) adherence property of M1ΔCdhA mutant strain in comparison to the wild-type strain (Fig. 7). The mutants expressing truncated CdhA (M1CdhAΔC55), which although adhered better (CFU/well) than M1ΔCdhA to pharyngeal cells and displayed their surface structure nearly normal, also adhered significantly less efficiently when compared with the wild-type strain. This indicated that CdhA directly or indirectly controls expression of certain surface proteins. Number of CFUs in control wells containing minimal essential medium without pharyngeal cells remained the same at the end of each bacterial adherence assay, indicating that bacterial cell death or multiplication did not contribute to these changes. Lack of CdhA abrogates antiphagocytic function-Since the M protein is a major virulence factor and an important antiphagocytic molecule that contributes significantly to the external electron dense layer of GAS (44), we predicted that the loss of electron-dense surface layer in the M1ΔCdhA mutant may in part attributed to the loss of the M or M-like proteins. Alternatively, we also predicted that the aggregated forms of the mutant might not be efficiently phagocytosed because of their size limitation. To determine whether the predicted loss of the M protein in the M1ΔCdhA mutant in fact affects bacterial ability to resist phagocytosis, whole blood phagocytosis assays were performed to determine the ability of GAS to survive and multiply. Both M1ΔCdhA and M1CdhAΔC55 mutant strains, in comparison to the wild-type strain, were readily phagocytosed like the Δemm1 mutant GAS strain (Fig. 8A), even though the M1ΔCdhA mutants showed tendency to form aggregates and M1CdhAΔC55 did not show any growth defect. The loss of ability to resist phagocytosis in these two mutants was therefore attributed to the loss of the M protein as determined by Western blot analysis and not simply due to some defects in the cell growth observed only for M1ΔCdhA mutant (Fig. 8B). These results corroborated with the observed 4 (M1CdhAΔC55) to 50 (M1ΔCdhA)-fold decrease in the emm1-specific mRNA level as determined by quantitative real-time PCR analyses (Fig. 8C). Further, both mutants displayed significantly reduced
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

10
(P<0.05) capsular formation as determined by hyaluronic acid contents (Fig. 8D). Almost 2-fold increased capsular contents in the cdhA-complemented mutant and the wild-type strains (M1ΔCdhA::cdhA and M1-WT::CdhA) suggested that CdhA somehow regulates the expression of the M protein and the capsule formation. Absence of CdhA increases bacterial susceptibility to antibiotics-On the basis of the functional properties CdhA to regulate cell division and cell wall hydrolase activity, we hypothesized that the lack of multifunctional CdhA (cell division and CHAP domain) may affect susceptibility to certain antibiotics that target peptidoglycan biosynthesis. To determine this, we measured minimum inhibitory concentrations of various antibiotics for the wild-type and M1ΔCdhA and M1CdhAΔC55 strains. We included two bacterial cell wall synthesis-targeting (Benzylpenicillin-PG and cefotaxime-CT), two protein synthesis-targeting (clindamycin-CM, and erythromycin-EM), and one DNA gyrase/topoisomerase IV-targeting (ciprofloxacin-CI) antibiotics using AB-BIODISK E-test. Significant reduction in MICs of penicillin G-PG (4-fold for M1ΔCdhA, 3.0-fold for M1CdhAΔC55) cefotaxime-CT (22.7-fold for M1ΔCdhA and 2.5 fold for M1CdhAΔC55) and clindamycin (5.4-fold for M1ΔCdhA, 3.9-fold for M1CdhAΔC55) and only marginal reduction (1.5-3-fold) in MICs of erythromycin and ciprofloxacin for both mutants indicated that presence of CdhA protein plays an important role in the maintenance of the cell wall integrity however, the hydrolase activity CdhA may have a limited contribution in the perceived role of controlling cell wall composition and modeling (Fig. 9A and B). SPy0019 mutants are attenuated for virulence in mice-Collectively, the growth pattern, in vitro adherence, and bactericidal/phagocytosis assays strongly suggest that CdhA could serve as a potential virulence factor and thus may have a direct or indirect role in GAS pathogenicity. To determine this, we employed the mouse intra-peritoneal infection model. As shown in Fig. 10, animals infected with the wild-type strain showed 50% mortality on day 4, while the M1ΔCdhA GAS mutant was completely attenuated for virulence as no morbidity or
mortality was observed in infected mice during the entire observation period. In contrast, M1CdhAΔC55, despite its similar growth pattern as the wild-type strain, displayed significantly reduced virulence as only 20% mortality was observed for the infected-mice by day 10 post infection. The virulence property of the complemented strain, M1ΔCdhA::cdhA, which was able to regain the normal wild-type growth function (Fig. 10A), was significantly restored (40 % mortality, P<0.05), although it was partial. Further significantly reduced bacterial load in the spleen, kidney and other visceral organs of the M1ΔCdhA-infected mice and increased bacterial load in the complemented strain indicated that the CHAP domain contribute significantly to the virulence regulating properties of CdhA (Figs. 10 A and B). Although truncated CdhA expressing M1CdhAΔC55 mutant was readily phagocytosed by human neutrophils, total bacterial load in the lungs of mice infected with M1CdhAΔC55 was found to be significantly higher than those in the wild-type GAS (M1SF370)-infected mice. The resulting partial loss of GAS virulence indicated that in the absence of CHAP-domain, the CdhAΔC55 protein somehow help modify the host response (more specifically associated with mouse lung or other organ macrophages) in a manner favorable for GAS (Fig. 10B). Thus, the CHAP domain of CdhA seems to play a critical role in GAS virulence. CHAP-domain of CdhA directly or indirectly regulates GAS virulence-Results described above indicated that CdhA is a multifunctional protein of which the cell division-plane recognition/chain-forming domain is located in the N-terminal 2/3 region of the protein and virulence is likely regulated by the CHAP domain. To determine the mechanism underlying the CHAP-domain-mediated regulation of virulence, we analyzed microarray-based differential global gene expression profiles for M1ΔCdhA and M1CdhAΔC55 mutants (Tables S4-S6), validated with qRT-PCR. Comparative analyses revealed several similarities in terms of genes belonging to specific functional categories that showed significant difference in their expression levels (> 2-fold, p<0.05). Out of 67 and 106 significantly differentiated genes in M1ΔCdhA
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

11
and M1CdhAΔC55, respectively, the 50 differentiated genes were identical in both strains (Table-1 and Tables-S4 and-S5). The most affected and notable categories of genes in this common group belonged to virulence (7/50), carbohydrate transport/metabolism (11/50), amino acid metabolism and transport (7/50), and energy production (putative V-type Na+ ATPase pump, 10/50). Among the 11 major virulence-related genes affected in M1ΔCdhA, 10 genes (SPy0165-nga, SPy0167-slo, SPy0738-sagA, SPy1302-amyA, SPy1983-scl, SPy2016-sic, SPy2018-emm1, SPy2039-speB, and SPy2043-mf, SPy2200-hasA) were severely down regulated (5 to 128-fold) and 3 genes (SPy1357-grab, SPy712-mf2, SPy0711-speC) were upregulated (3-4 fold) (Tables 1-2 and S4 –S6). In M1CdhAΔC55 expressing truncated CdhA lacking last 55 residues, six (scl, hasA, amyA, emm1, sagA, and mf) and two (mf2, speC) of the above mentioned virulence-related genes were down- and up-regulated respectively. These results concur with electron microscopy, and in vitro (adherence, antiphagocytic studies) and in vivo (experimental infection model) virulence studies.
DISCUSSION In the present study, we describe novel domain-specific functions of group A streptococcal secreted protein SPy0019/CdhA, which was described earlier as “SibA” based on the hIgG-binding ability of its recombinant form (18). Based on our earlier observations for the GAS mutants lacking SPy1625 or SP-STK (M1ΔSTK) showing cell-division defective phenotypes (13), the present study was carried out to address two important questions: (i) Are SP-STK-regulated cell division defects controlled by CdhA, as gram-positive pathogens lacking CdhA orthologs display similar cell division defects ? and (ii) What is the contribution of CdhA in GAS pathogenesis? The answer to the first question was readily addressable by complementing and restoring the cell division defects of the GAS mutant lacking SP-STK with the wild-type cdhA. We believe that unlike S. pneumonia PscB (15,36) and S. mutans SagA/GtfB (46), the dispensable nature of CdhA allowed us to further study its domain-specific biochemical and biological functions in GAS pathogenesis using domain-specific knockout mutants. Additionally, by
complementing heterologus wild-type strains, E. coli and S. aureu, and the mutant strain, M1ΔSTK with the wild-type cdhA, we also delineated the specific role of CdhA as cell division-plane-recognition protein. The latter property seemed to be utilized as one of the earliest events in bacterial chain formation attributed previously to streptococcal PcsB (17,16). The designation assigned to SPy0019 as CdhA in this study was thus based on its biochemical and biological functions. By light microscopy and complementation of M1ΔSTK mutant with the wild-type cdhA, we provided an initial proof that CdhA is regulated by SP-STK, since by increasing the expression of CdhA in M1ΔSTK mutant, it was possible to recover the wild-type chain forming characteristic, growth pattern and partial restoration of cell division defects observed in M1ΔSTK (13). These phenotypes looked similar to the M1ΔCdhA mutant complemented with wild-type cdhA. While SEM and TEM of the cdhA-complemented strain, M1ΔCdhA::cdhA, indeed confirmed the restoration of cell division defects, a similar study for M1ΔSTK::cdhA revealed restoration of chain formation but not the defective cell division. These results indicated that CdhA play an important role in certain aspects of the complex cell division process (40,41,42). In particular, the ability of CdhA to induce chain formation was confirmed when the CdhA-overexpressing wild-type M1-SF370 strain (M1-WT::cdhA) exhibited extremely long chain. The ability of CdhA to confer similar phenotypic effects in heterologous systems like E. coli (BL21::cdhA) and S. aureus (RN4220::cdhA) which do not normally form chains, unequivocally indicated that CdhA is a chain-forming protein. This is in contrast to PcsB from S. mutans and S. agalactiae which do not replace pneumococcal PcsB function (16).In fact, the underexpression and not the overexpression of PcsB in pneumococci leads to formation of long chains of cells and excessive septal wall synthesis (16). These contrasting results indicates that despite the structural similarities, their biochemical and biological activities are unique. Since S. pyogenes and E. coli divide in one plane, the long chain formation in these organisms was very obvious as an outcome of CdhA over expression. In S. aureus, however, complementation with wild-
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

12
type cdhA resulted in loosely formed clusters with short chains. Characteristically, these chains of S. aureus were, unlike other two strains, were with frequent kinks. We believe that the kink formation in S. aureus is due to its characteristic division pattern in three specific planes, X, Y, and Z (Fig. 4C) and a long chain in one plane is due to S. aureus obligation to divide under the influence of CdhA until the onset of “Y” and “Z”-specific second and third signals that will divide S. aureus in three different planes. Accordingly, it is likely that S. aureus may possess more than one plane recognizing proteins including an ortholog of CdhA. Although SA2093 (SSA) of S. aureus is a likely ortholog (47), its function has not been revealed so far. Although CdhA is a secreted protein, its role in cell division is underscored by its location in septum as seen by fluorescence microscopy (Fig. 3A). It is not known how CdhA is functionally integrated with the temporally regulated orchestration of several proteins, such as FtsZ, FtsA and/or DivA, that are involved in the complex cell division process to achieve specific shape and forms of bacteria (48,41,49). While this relation is presently under investigation, the results showing unregulated cell division with multiple septa in different planes in M1ΔCdhA and restoration of these defects in M1ΔCdhA::cdhA clearly indicated that the cdhA is a cell division-plane-recognition protein playing a crucial role in early events of cell division and ultimately chain formation in S. pyogene. Several decades ago, the induction of long chains in GAS was attributed to the presence of type M-specific antibody (50). However, the molecular and genetic basis of bacterial chain formation in GAS that is described in this study is distinct from the trypsin-sensitive, and type-specific antibody-mediated physical associations of more than one bacterial chains (50). Although the mechanisms behind the direct relationship of the absence and over expression of CdhA with up and down regulation of capsule formation is presently not known, a similar corroborating report for pneumococcal PcsB (51) suggest that certain capsule regulator(s) may potentially interact with CdhA and modulate their regulatory functions. CdhA belongs to a class of proteins that contains the CHAP domain (24,38). The CHAP-
domain-containing proteins are in general known to degrade the cell-wall cross-links, or as an amidase to disrupt acetylmuramyl bonds (24,38). Interestingly, none of the CdhA orthologs in gram-positive pathogens have been conclusively shown to possess either in vitro or in vivo hydrolase activity or specific biologically relevant functions. The ability of CdhA to reveal in vitro zymogram-based cell wall hydrolase activity only in the presence of purified GAS cell wall peptidoglycan (Fig. 6) is likely due either to uniform distribution of peptidoglycan in the SDS-gel or to specificity of CdhA-CHAP for M1SF370 peptidoglycan as earlier attempts failed when acetone powder of M. luteus and S. pyogenes were employed. Since cell wall synthesis and cell division are physiologically closely linked (40,49), we initially attributed the cell division regulatory function to the CHAP-domain of CdhA, which, unlike other orthologs from Gram-positive pathogens (19,16,22), is not essential for GAS survival. Hence, it was possible to derive GAS mutants lacking CdhA and that expressing truncated CdhA lacking C-terminal 55 residues encompassing the catalytic site of the CHAP domain and investigate their physiologically relevant functions based on phenotypic characteristics of the derived mutants. M1CdhAΔC55 mutant although did not display any in vitro hydrolase activity, to our surprise the mutants grew like the wild-type strain and did not show any defects in cell division indicating that the CHAP-domain is not involved in cell division or the cell division-plane recognition process as proposed before. To reveal the precise biological role of the CHAP domain that is directly relevant to GAS pathogenesis, we initially performed several in vitro virulence-determining assays. Significant reduction in the abilities of M1ΔCdhA as well as M1CdhAΔC55 mutants to adhere to human pharyngeal cells and resist phagocytosis most likely due to significant down regulation of the M-protein in the corresponding cell wall extracts and significantly reduced amount of hyaluronic acid clearly indicated that CdhA, and more specifically the CHAP domain, directly or indirectly regulate GAS virulence. The complete attenuation of M1ΔCdhA as determined by mouse virulence assays and a partial yet significant reduction in mouse mortality by i.p. injection of M1CdhAΔC55, substantiate results obtained by employing in vitro virulence factor
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

13
determining assays. These results conclude that the CHAP-domain of CdhA, although does not participate in cell division process and grow like the wild-type strain, plays a significant role in the GAS virulence. In other words, the growth defects observed in M1ΔCdhA (grows maximally at O.D 0.6) contribute minimally for the significantly decreased MF values in in vitro phagocytosis assays and in vitro virulence assays.
Since bacterial adherence and subsequent colonization play a primary role in bacterial virulence, the decreased virulence of M1ΔCdhA and M1CdhAΔC55 GAS mutants is also due to their significantly decreased ability to adhere to human pharyngeal cells in addition to loss of antiphagocytic activity. The mechanism underlying the ability of CdhA in controlling the expression of other surface proteins, which may play an important role in GAS adherence, is not apparent at present. We initially speculated that the incomplete septum formation might cause missorting of many surface proteins and virulence factors and even their spilling in the supernatant instead of linking covalently to the cell wall (43,52,53). The microarray analysis of the CdhA-specific mutants, however, offered more plausible explanation as, in addition to Emm1, at least three other well established surface proteins, ScpA (54), Sic (55), and Scl (56) were found to be significantly downregulated. This may result in cell walls without adhesins.
Functionally active CHAP-domain indicated that cell wall hydrolase activity of CdhA may play an important role in cell wall synthesis and modulation. However, the absence of the CHAP domain of CdhA does not influence growth and cell division. Further the increase in susceptibility to cell wall acting antibiotics (penicillin and cefotaxime) especially in the absence of the CHAP-domain was only limited to 2-3 fold. The antibiotic susceptibility pattern thus question whether the CHAP domain of CdhA cell plays a role in cell wall synthesis or modulation (24,38). Thus, defective cell walls of CdhA knockout mutants as revealed by high susceptibility to the cell wall-acting antibiotics may be an indirect outcome of improper cell division. The recently reported mass-spectometric analysis for a pneumococcal mutant lacking the PcsB protein (CdhA homolog) revealing apparently defective wall
without any change in the relative amount or composition of the peptidoglycan peptides lend support our hypothesis(16,51). The function of CHAP-domain could therefore be only to release muropeptides (57) during multiplication and until stationary phase and not for the modulation of cell wall. The precise role of the CHAP-domain of the bacterial peptidoglycan hydrolases in controlling bacterial virulence is presently not known. In this regard, in the present study, the results obtained from the animal experiments showing significantly increased streptococcal colony forming units in the lungs of M1CdhAΔC55-infected mice as compared to those found in the lungs of the wild-type strain- infected mice draw special attention with regard to the specific virulence-related role of the CHAP-domain in CdhA. The presence of large number of CFUs of M1CdhAΔC55 in lungs despite it being depleted of the M protein and capsule indicate that lung macrophages of the infected mouse could not phagocytose GAS expressing truncated CdhA or possibly they were phagocytosed but not killed. The CHAP-domain containing bacterial peptidoglycan hydrolases have been shown to modulate muropeptide relrease and host innate immune responses (57). Certain members of Nod-like receptor (NLR) family, which are generally expressed constitutively by antigen presenting cells (APCs, e.g. dendritic cells and macrophages) and epithelia, are capable to detect specific muropeptides. Signaling through NLR family results in the coordinated activation of cytoplasmic, multimolecular protein complexes (inflammasomes) which in turn orchestrates massive inflammation and tissue injury (58). Thus, the mutant strain expressing the truncated CdhA lacking the catalytic site of its CHAP-domain may not be able to release the required muropeptides from the GAS that has metastatically localized in lungs after experimental IP infection (Fig. 10B). This may not result in activation of NLR-associated inflammasomes in lung macrophages and killing of the M1CdhAΔC55 in lungs. While this could be a valid reason of significantly increased number of M1CdhAΔC55 in lungs and partial attenuation for mouse virulence (Fig. 10A), the proposed hypothesis needs experimental verification.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

14
In addition to apparent changes in the transcription profiles of several virulence-related genes, we also observed drastic changes in the carbohydrate, and amino acid metabolism related genes. This is in contrast to recently published results for S. pneumoniae mutants depleted of PcsB which showed changes in few genes belonging to the VicRK regulon, but surprisingly limited for other genes despite the depleted mutant showed defect in cell shape and morphology (51). In the present study, most notable differentially down-regulated carbohydrate genes (2-4 fold) in both M1ΔCdhA and M1CdhAΔC55 were the malM operon, lacA.1, lacA.2, lacB.1, lacD.2 and lacE. The down-regulation of these genes in GAS has recently been reported to inversely affect GAS virulence (59,60,61,62,63). Thus the attenuation of virulence in mutants as a result of a complete or partial deletion of the cdhA genes can also be attributed the down-regulation of carbohydrate metabolism genes. Both mutants, unlike the wild-type GAS strain, were unable to grow efficiently in the chemically defined medium supplemented with maltose/maltodextrin instead of glucose (Data not shown).
Transcription of PscB of S. pneumoniae R6 and GbpB (SagA) of S. mutans is positively regulated by their respective VicR/K TCRSs indicating that CdhA and its orthologs in other gram-positive bacteria could be a common target of VicR/K (64,16,15,46). In the present study we showed that SP-STK also somehow regulates the transcription of cdhA. While the function of VicR is regulated by its cognate histidine kinase,
VicK, the streptococcal VicR has been shown to function independent of its cognate VicK (64,15). It is therefore conceivable that VicR-mediated regulation of CdhA expression is fine tuned by SP-STK by phosphorylating VicR at its threonine residues. The mechanism of this regulation is presently under investigation. In summary, this is the first report showing CdhA is an SPy1625(SP-STK)-dependent cell wall-associated as well as secreted GAS protein, displaying multifunctional nature with a cell division-function located in its N-terminal two third portion and the cell wall hydrolase-function and major virulence functions located in the remaining C-terminal region. Since CdhA does not significantly contribute to the human immunoglobulin binding property of GAS, we have annotated SPy0019 as CdhA. The mechanism by which the CHAP-domain of CdhA controls GAS virulence and the N-terminal domain recognizes the division plane and orchestrates cell division are presently not known. While host innate immune responses may be a target for this protein for the observed virulence, the increase antibiotic susceptibility of GAS in the absence of SPy0019/SibA/CdhA and the virulence attenuation especially in the absence of its C-terminal region favor this protein being an important candidate/target for the development of newer immunotherapeutic as well as chemotherapeutic agents.
REFERENCES 1. Carapetis, J. R., Steer, A. C., Mulholland, E. K., and Weber, M. (2005) Lancet Infect Dis 5, 685-
694
2. Cone, L. A., Woodard, D. R., Schlievert, P. M., and Tomory, G. S. (1987) N Engl J Med 317, 146-149
3. Stevens, D. L. (1995) Emerg Infect Dis 1, 69-78
4. Johnson, D. R., Stevens, D. L., and Kaplan, E. L. (1992) J Infect Dis 166, 374-382
5. Steer, A. C., Carapetis, J. R., Nolan, T. M., and Shann, F. (2002) J Paediatr Child Health 38, 229-234
6. McDonald, M., Currie, B. J., and Carapetis, J. R. (2004) Lancet Infect Dis 4, 240-245
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

15
7. Rodriguez-Iturbe, B. , Musser, J. M. (2008) J Am Soc Nephrol 19, 1855-1864
8. Carapetis, J. R. (2008) Circulation 118, 2748-2753
9. Musser, J. M. , Krause, R. M. (1998), 185-218
10. Krause, R. M. (2002) Indian J Med Res 115, 215-241
11. Cunningham, M. W. (2000) Clin Microbiol Rev 13, 470-511
12. Stock, A. M., Robinson, V. L., and Goudreau, P. N. (2000) Annu Rev Biochem 69, 183-215
13. Jin, H. , Pancholi, V. (2006) J Mol Biol 357, 1351-1372
14. Churchward, G. (2007) Molecular Microbiology 64, 34-41
15. Ng, W. L., Robertson, G. T., Kazmierczak, K. M., Zhao, J., Glimour, R., and Winkler, M. E. (2003) Mol Microbiol 50, 1647-1663
16. Ng, W. L., Kazmierczak, K. M., and Winkler, M. E. (2004) Mol Microbiol 53, 1161-1175
17. Reinscheid, D. J., Gottschalk, B., Schubert, A., Eikmanns, B. J., and Chhatwal, G. S. (2001) J Bacteriol 183, 1175-1183
18. Fagan, P. K., Reinscheid, D., Gottschalk, B., and Chhatwal, G. S. (2001) Infect Immun 69, 4851-4857
19. Mattos-Graner, R. O., Jin, S., King, W. F., Chen, T., Smith, D. J., and Duncan, M. J. (2001) Infect Immun 69, 6931-6941
20. Borges, F., Layec, S., Thibessard, A., Fernandez, A., Gintz, B., Hols, P., Decaris, B., and Leblond-Bourget, N. (2005) J Bacteriol 187, 2737-2746
21. Le Breton, Y., Maze, A., Hartke, A., Lemarinier, S., Auffray, Y., and Rince, A. (2002) Current Microbiology 45, 434-439
22. Teng, F., Kawalec, M., Weinstock, G. M., Hryniewicz, W., and Murray, B. E. (2003) Infect Immun 71, 5033-5041
23. Schubert, K., Bichlmaier, A. M., Mager, E., Wolff, K., Ruhland, G., and Fiedler, F. (2000) Arch Microbiol 173, 21-28
24. Bateman, A. , Rawlings, N. D. (2003) Trends Biochem Sci 28, 234-237
25. Ferretti, J. J., McShan, W. M., Ajdic, D., Savic, D. J., Savic, G., Lyon, K., Primeaux, C., Sezate, S., Suvorov, A. N., Kenton, S., Lai, H. S., Lin, S. P., Qian, Y., Jia, H. G., Najar, F. Z., Ren, Q., Zhu, H., Song, L., White, J., Yuan, X. and et al (2001) Proc Natl Acad Sci USA 98, 4658-4663
26. Beltramini, A. M., Mukhopadhyay, C. D., and Pancholi, V. (2009) Infect Immun 77, 1406-1416
27. Podbielski, A., Spellerberg, B., Woischnik, M., Pohl, B., and Lutticken, R. (1996) Gene 177, 137-147
28. Boel, G., Jin, H., and Pancholi, V. (2005) Infect Immun 73, 6237-6248
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

16
29. Chaffin, D. O. , Rubens, C. E. (1998) Gene 219, 91-99
30. Jin, H., Song, Y. P., Boel, G., Kochar, J., and Pancholi, V. (2005) J Mol Biol 350, 27-41
31. de Jonge, B. L., Chang, Y. S., Gage, D., and Tomasz, A. (1992) J Biol Chem 267, 11248-11254
32. Lepeuple, A. S., Gemert, E. V., and Chapot-Chartier, M. P. (1998) Appl Environ Microbiol 64, 4142-4148
33. Jones, K. F., Khan, S. A., Erickson, B. W., Hollingshead, S. K., Scott, J. R., and Fischetti, V. A. (1986) J Exp Med 164, 1226-1238
34. Rainer, J., Sanchez-Cabo, F., Stocker, G., Sturn, A., and Trajanoski, Z. (2006) Nucleic Acids Res 34, W498-W503
35. Pfaffl, M. W. (2001) Nucleic Acids Res 29, 2002-2007
36. Reinscheid, D. J., Ehlert, K., Chhatwal, G., and Eikmanns, B. J. (2003) FEMS Microbiol Lett 221, 73-79
37. Smith, D. J., King, W. F., Barnes, L. A., Peacock, Z., and Taubman, M. A. (2003) Infect Immun 71, 1179-1184
38. Rigden, D. J., Jedrzejas, M. J., and Galperin, M. Y. (2003) Trends Biochem Sci 28, 230-234
39. Anantharaman, V. , Aravind, L. (2003) Genome Biol 4, R11-
40. Cabeen, M. T. , Wagner, C. J. (2008) Nature Rev 3, 601-610
41. Rothfield, L. I., Taghbalout, A., and Shih, Y.-L. (2005) Nat Rev Microbiol 3, 959-968
42. Lock, R. L. , Harry, E. J. (2008) Nat Rev Drug Discov 7, 324-338
43. Navarre, W. W. , Schneewind, O. (1999) Microbiol Molec Biol Rev 63, 174-229
44. Fischetti, V. A. (2006)2nd edition, 12-25
45. Pancholi, V. (2005)2nd edition (in press),
46. Senadheera, M. D., Guggenheim, B., Spatafora, G. A., Huang, Y. C., Choi, J., Hung, D. C., Treglown, J. S., Goodman, S. D., Ellen, R. P., and Cvitkovitch, D. G. (2005) J Bacteriol 187, 4064-4076
47. Dubrac, S. , Msadek, T. (2004) J Bacteriol 186, 1175-1181
48. Margolin, W. (2005) Nature Rev Mol Cell Biol 6, 862-871
49. Scheffers, D.-J. , Pinho, M. G. (2005) Microbiol Mol Biol Rev 69, 585-607
50. Stollerman, G. H. , Ekstedt, R. (1957) J Exp Med 106, 345-361
51. Barendt, S. M., Land, A. D., Sham, L. T., Ng, W. L., Tsui, H. C., Arnold, R. J., and Winkler, M. E. (2009) J Bacteriol 191, 3024-3040
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

17
52. Carlsson, F., Stalhammar-Carlemalm, M., flardh, k., sandin, C., Carlemalm, E., and Lindahl, G. (2006) Nature 442, 943-946
53. Scott, J. R. , Barnett, T. C. (2006) Ann Rev Microbiol 60, 397-423
54. Ji, Y., McLandsborough, L., Kondagunta, A., and Cleary, P. P. (1996) Infect Immun 64, 503-510
55. Hoe, N. P., Ireland, R. M., DeLeo F.R., Gowen, B. B., Dorward, D. W., Voyich, J. M., Liu, M., Burns, E. H., Culnan, D. M., Bretscher, A., and Musser, J. M. (2002) Proc Natl Acad Sci USA 99, 7646-7651
56. Lukomski, S., Nakashima, K., Abdi, I., Cipriano, V. J., Shelvin, B. J., Graviss, E. A., and Musser, J. M. (2001) Infect Immun 69, 1729-1738
57. Humann, J. , Lenz, L. L. (2009) J Innate Immun 1, 88-97
58. Stutz, A., Goldenbock, D. T., and Latz, E. (2009) J Clin Invest 119, 3502-3511
59. Shelburne, S. A., Keith, D., Horstmann, N., Sumby, P., Davenport, M. T., Graviss, E. A., Brennan, R. G., and Musser, J. M. (2008) Proc Natl Acad Sci U S A 105, 1698-1703
60. Shelburne, S. A., Davenport, M. T., Keith, D. B., and Musser, J. M. (2008) Trends Microbiol 16, 318-325
61. Loughman, J. A. , Caparon, M. G. (2007) Mol Microbiol 64, 269-280
62. Loughman, J. A. , Caparon, M. G. (2006) EMBO J 25, 5414-5422
63. Rosch, J. , Tuomanen, E. (2007) Mol Microbiol 64, 257-259
64. Liu, M., Hanks, T. S., Zhang, J., McClure, M. J., Siemsen, D. W., Elser, J. L., Quinn, M. T., and Lei, B. (2006) Microbiology 152, 967-978
FOOTNOTES *The authors thank Amanda Beltramini for S. aureus complement construct, and Shivani Agarwal, Shivangi Agarwal, and Haritha Adhikarla for critical reading of this manuscript and helpful discussion. This study was supported in part by AI64912 grant from National Institute of Health to VP. Supporting Information: Table S1. DNA sequences of primers used for constructing recombinant proteins and GAS mutants in the present study. Table S2. Strains and Plasmids used in the present study Table S3. Primers sequences used for qRT-PCR assays in the present study Table S4. Microarray-based differential gene expression analysis for M1ΔCdhA vs the wild-type M1-
SF370 (GSE#15546).
Table S5. Microarray-based differential gene expression analysis for M1CdhAΔC55 vs M1-SF370 (GSE#15598).
Table S6. Comparison of expression profiles of 18 relevant genes in M1ΔCdhA and M1CdhAΔC55 by qRT-PCR and microarray analyses.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

18
Abbreviations used are: GAS, Group A Streptococcus; CdhA, CHAP domain-containing and chain-forming cell wall hydrolase from group A Streptococcus; CHAP, Cysteine, Histidine-dependent Aminohydrolases/Peptidases; TCS,Two-component regulatory systems; ESTK, eukaryote-type Serine/Threonine kinase; SP-STK, Streptococcus pyogenes Ser/thr kinase. ¶Present address: Gregory Boel, Department of Biological Sciences, Columbia University, 702A Fairchild Center, 1212 Amsterdam Avenue, New York, NY 10027
FIGURE LEGENDS
Fig.1. Physiological relationship between SP-STK and SPy0019/CdhA . mRNA expression levels of spy0019/cdhA in the M1ΔSTK mutant strain in comparison to that in the wild-type M1SF370 strain. Error bars represent Mean + SEM of three independent experiments each in triplicate wells. Fig. 2. Growth curves and integrity of CdhA-specific mutant and complemented strains. Wild-type (M1SF370 or M1-WT) and various isogenic mutant strains and corresponding complemented strains were created with different strategies and were grown in THY broth as described in Materials and Methods. (A) Growth curves of the wild-type (M1-WT), isogenic mutant strains (M1ΔCdhA mutant lacking CdhA, M1CdhAΔC55 mutant expressing truncated-CdhA, and M1ΔSTK mutant lacking SP-STK (13)), (B) corresponding complemented strains (M1ΔCdhA::cdhA, and M1ΔSTK::cdhA (SP-STK-mutant complemented with cdhA), and CdhA overexpressing strain (M1-WT::cdhA) strains were monitored spectrophotometrically by measuring O.D.(600 nm) over a period of 18 h. Each data point represents an average of three independent readings. For clarity purpose error bars have been omitted. Arrows M, and L indicate mid and late log phase points for each growth curve. (C) Western blot analysis showing the presence and absence of CdhA expression in the whole lysates of mutant, wild-type and complemented strains using anti-CdhA IgG. (1) recombinant CdhA protein, and whole cell lysates of (2) wild type M1SF370, (3) M1ΔCdhA, (4) M1CdhAΔC55, (5) M1ΔCdhA::cdhA, (6) M1-WT::cdhA, (7) M1ΔSTK, and (8)M1ΔSTK::cdhA. Fig. 3. Light, scanning, and transmission electron microscopy of M1-SF370 wild-type and M1ΔCdhA, M1CdhAΔC55, and M1ΔCdhA::cdhA mutant GAS strains. (A) Indirect immunofluorescence microscopy of intact wild-type SF370 and the mutant M1ΔCdhA strain using anti-CdhA antibody, FITC-labeled conjugate antibody and DAPI (blue nuclear stain). CdhA is primarily located at the septum (Green fluorescence, Panel I) of M1-SF370. Inset in the first panel shows the magnification of a region localized with CdhA in one of the dividing bacteria. Panel II shows the M1ΔCdhA strain forming cluster and the absence of green fluorescence (only nuclear staining with DAPI). Panels III and IV were stained with “Live-Dead bacteria stain”. The presence of red stain in some portion of the clusters of M1ΔCdhA (Panel IV) indicates the presence of dead bacteria. (B) Images acquired for the M1-WT, M1ΔCdhA, M1CdhAΔC55, M1ΔCdhA::cdhA strains employing light (left panel), scanning (middle panel, SEM) and transmission electron microscopy (right panel, TEM). The indicated scale mark applies to both SEM and TEM for each strain. Light microscopy was carried using Gram’s Stain. Fig. 4. CdhA is a bacterial cell division-plane-recognizing and a chain-forming factor. (A) Light and electron microscopy (SEM and TEM) of M1ΔSTK and M1ΔSTK::cdhA and M1-WT::cdhA mutant strains. (B) Light microscopy of uninduced E. coli BL21 and IPTG-induced pET14B-His-CdhA-
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

19
transformed (His-CdhA overexpressing) BL21::cdhA strains. (C) S. aureus RN4220 wild-type and pCN40tetR-cdhA-transformed (CdhA-overexpressing) RN4220::cdhA strains. Fig. 5. Comparison of human immunoglobulin-binding property of CdhA mutants with that of the wild-type GAS strain. Ligand-binding assays were performed in 96-well formats as described in Materials and Methods. Each data point represents an average + SEM of three individual experiments performed in triplicates. Final readings represent values obtained after subtracting the average value obtained from 12 control wells. Fig. 6. In vitro cell wall hydrolyzing property of native CdhA expressed in GAS and recombinant CdhA. Zymographic analysis of GAS cell wall hydrolase activity of purified CdhA and ammonium sulphate precipitates of culture supernatants of the wild-type, CdhA-specific mutants, and cdhA-complemented GAS strains as indicated. Fig. 7. Lack of CdhA adversely affects bacterial adherence to human pharyngeal cells. Adherence index was determined using Detroit 562 human pharyngeal carcinoma cell lines and as percentage of initial inoculum (MOI 50:1::bacteria:cell, i.e. 5x107cfu/well). Error bars represent mean + SEM of three independent experiments each performed in at least triplicate wells. Fig. 8. Lack of CdhA abrogates GAS antiphagocytic function by down-regulating the M protein expression and capsule formation. (A) Phagocytic index of M1-WT, M1ΔCdhA, M1CdhAΔC55 and M1ΔEmm. Phagocytic index was measured as a multiplication factor (MF) indicating the ability of the GAS strain to multiply in the whole human defibrinted blood as described in Materials and Methods. M1ΔEmm lacks the anti-phagocytic M protein encoded by the emm1 gene and was used as positive control (13). (B) Quantitative real-time PCR to determine emm1-specific mRNA in the late log phase culture (growth level III) of M1-WT, M1ΔCdhA and M1CdhAΔC55 GAS strains in comparison to the internal control gene (proS) specific mRNA. (C) Western blot analysis to determine the expression of the M-protein in the (i) culture supernatants and (ii) cell wall extracts of (I) early, (II) mid, and (III) late log phase-grown cultures of (1) M1-WT, (2) M1ΔCdhA, and (3) M1CdhAΔC55 strains using the Emm1-reactive 10B6 monoclonal antibody (33). (D) Comparison of hyaluronic acid contents of the mutants M1ΔCdhA, M1CdhAΔC55, M1ΔCdhA::cdhA and M1-WT::cdhA strains with the wild-type M1SF370 (M1-WT) GAS strains. Vertical bars represent Mean + SEM of three independent experiments. * P<0.001. Fig. 9. Lack of CdhA affects susceptibility of GAS to various antibiotics. Comparison of antibiotic susceptibility profiles (changes in MICs) of M1ΔCdhA, M1CdhAΔC55, and M1ΔCdhA::cdhA with respect to those of M1-WT GAS strain. MIC values were determined by E-test AB biodisk-recommended procedures. (A) Representative antibiotic susceptibility profiles of benzylpenicillin (PG) and cefotaxime (CT) for the wild-type, mutant, and the cdhA-complemented strains. (B) Each bar represents an average reading obtained with three E-strips. The Fold-changes (# x) in MIC values are calculated as the ratio of MIC for the Wild-type/MIC for the mutant. PG- benzylpenicillin; CT-cefotaxime; EM-erythromycin; CL-clindamycin; CI-ciprofloxacin. Fig. 10. Absence of CdhA or its catalytic CHAP domain attenuates GAS for virulence in experimental mouse intraperitoneal infection model. (A) Effects due to the deletion of the entire cdhA gene or its CHAP-domain specific region (C-terminal 55 residue) on GAS virulence in mice were observed for M1-WT, M1ΔCdhA, M1CdhAΔC55, and M1ΔCdhA::cdhA GAS strains using the mouse intraperitoneal infection model. Survival/mortality curves for all strains were monitored for 10 days and statistically evaluated and plotted by the Log-rank test using GraphPad Prism 4 software. The SHAM-infected mice survived throughout the observation period (not shown). (B) Bacterial load determined in different organs (lung, liver, spleen and kidney) from three infected animals from each group (10 mice each) sacrificed on day 4. * P<0.001, ** P<0.05.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

20
TABLE-1: Global gene expression profiles for M1ΔCdhA and M1CdhAΔC55 and distribution of differentially expressed genes* into major functional categories including virulence-related genes. d as significantly differentiated gene. (See also Table S4 and S5 for more details).
Functional Categories M1ΔCdhA M1ΔCdhA & M1CdhAΔC55
M1CdhAΔC55
Up (7)
Down (60)
Total (67)
Up (3)
Down (47)
Total (50)
Up (6)
Down (100)
Total (106)
Amino Acid transport 1 6 7 0 5 5 0 7 7 Carbohydrate transport 1 11 12 0 11 11 0 28 28 Cell envelope Biogenesis, Outer membrane
0 2 2 0 2 2 0 2 2
Cell motility and secretion 1 0 1 0 0 0 0 1 1 Co-enzyme metabolism 0 0 0 0 0 0 0 3 3 DNA replication and recombination
0 1 1 0 1 1 0 1 1
Energy production and connversion
0 5 5 0 5 5 0 10 10
Unknown function including CdhA*
1 19* 20 1 13 14 2* 21 23
General functions only 0 2 2 0 1 1 1 8 9 Lipid metabolism 0 1 1 0 0 0 0 3 3 Post-translational modification/chaperones
0 1 1 0 1 1 0 3 3
Secondary metabolism 0 0 0 0 0 0 0 1 1 Signal transduction 0 2 2 0 2 2 0 2 2 Transcription and translation 0 1 1 0 1 1 0 4 4 Virulence 3 9 12 2 5 7 3 6 9
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

21
Table-2 Global gene expression profiles for M1ΔCdhA and M1CdhAΔC55 and distribution of differentially expressed virulence-related genes (Up or down)*
• Bold figures in parentheses denote x fold change (up or down) in expression levels of individual
genes.
Mutant Type M1ΔCdhA M1ΔCdhA & M1CdhAΔC55
M1CdhAΔC55
Expression profile Up*
Down* Up Down Up
Down
Virulence genes 3 9 2 5 3 6 NAD-glycohydrolase SPy
0165**(x25 )
Streptolysin-O-precursor SPy 0167 (x13.4)
Exotoxin C-SpeC SPy 0711 (x3.4)
SPy 0711
SPy 0711 (x3.1)
DNase/mitogenic factor SPy 0712 (x3.2)
SPy 0712
SPy 0712 (x3.1)
Streptolysin-S-sagA SPy 0738 (x2.3)
SPy 0738 (x5.7)
Cyclomaltodextrin glucanotransferase-amyA SPy 1302 (x6.5)
SPy 1302
SPy 1302 (x12.3)
GRAB SPy 1357 (x3.6)
DNase-mitogenic factor-3-Mf3 SPy 1436 (x4.4)
Collagen-like surface protein-Scl SPy 1983 (x42.2)
SPy 1983
SPy 1983 (x21.1)
M-protein-Emm1 SPy 2018 (x17.1)
SPy 2018
SPy 2018 (x7.5)
Exotoxin-B, SpeB SPy 2039 (x12.1)
Mitogenic factor SPy 2043 (x4.7)
SPy 2043
SPy 2043 (x3.2)
Hyaluronate synthetase-HasA SPy 2200 (x147)
SPy 2200
SPy 2200 (x24.5)
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

22
B
M1-WT M1ΔSTK
proS
-nor
mal
ized
m
RN
A Ex
pres
sion
ratio
for S
Py00
19
Figure. 1.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

23
Figure.2
0
0.2
0.4
0.6
0.8
1
1.2
0 200 400 600 800 1000
O.D
. 640
nm
Time (Min)
M1-WT M1CdhAΔC55
M1ΔSTK M1ΔCdhA
(A)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0 200 400 600 800 1000
M1-WT M1-WT::cdhA
M1ΔSTK::cdhA M1ΔCdhA::cdhA
Time (Min)O
.D. 6
40nm
(B)
M
L ML
kDa50
37
25
CdhA1 2 3 4 5 6 7 8(C)
Anti-CdhA antibody
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

24
(A)
(I) (II) (III) (IV)
(B)
2 μm
M1-WT M1ΔCdhA
M1-WT
M1ΔCdhA
M1CdhAΔC55
M1ΔCdhA::cdhA3 μm
2 μm
M1-WT M1ΔCdhA
Figure. 3
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

25
3μm
3μmM1ΔSTK
3μm
M1ΔSTK::cdhA
M1-WT::cdhA
E.coli BL21 S. aureus RN4220
WT
WT:
:cdh
A
(A)
(B)
WT
WT:
:cdh
A
(C)
Figure. 4
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

26
Figure. 5.
0 25 50 75 1000
1
2
3
4
M1-WT M1ΔCdhA
M1CdhAΔC55 M1ΔCdhA::cdhA
P<0.0001
Human Immunoglobulin (μg/well)
O.D
.450
nm
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

27
Figure. 6.
Cell wall hydrolase activity of CdhA
50
37
kDa recC
dhA
M1Δ
CdhA
M1Δ
CdhA
::cdh
A
10075
M1-
WT:
:cdh
A
M1-
WT
M1C
dhAΔ
C55
CdhA
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

28
M1-WT
CdhA
ΔM1
C55Δ
M1CdhA
0
10
20
30
40
P=0.004P=0.033
GAS strains
GA
S A
dher
ence
(% in
ocul
um, 5
x 1
07
CFU
/wel
l)
Figure. 7
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

29
(A) (B)
M1-WT
CdhA
ΔM1
C55Δ
M1CdhA em
m1
ΔM1
0102030405060
* * *
M.F
.
5037
5037
i
ii
I II III
1 2 3 1 2 3 1 2 3(C)
Anti-M6 mAb (10B6)M1-W
TCdhA
ΔM1
C55Δ
M1CdhA
CdhA::cdhA
ΔM1 M1-W
T::cdhA
0
5
10
15
20
25
P=0.003
P=0.036μ
g/m
l hya
luro
nic
acid(D)
M1-WT
CdhA
ΔM1
C55Δ
M1CdhA
0
250
500
750
1000
1250 P=0.0194
P=0.04
emm
1 m
RN
A(n
orm
aliz
ed w
ithpr
oS)
Figure. 8.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

30
M1-WT M1ΔCdhA M1CdhAΔC55 M1ΔCdhA::cdhA
PG
CT
3.0x
1.5x
2.04x
22.8x2.5x
5.4x
3.9x2.7x
2.7x
PG EM CT CM CI0.00
0.05
0.10
0.15
0.20
0.25
0.30 M1-WTM1ΔCdhAM1CdhAΔC55M1ΔCdhA::cdhA
M.I.
C ( μ
g /m
l)
4.0x
Antibiotics (E-test®)
(A)
(B)
Figure. 9.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

31
(A) (B)
* **
Figure. 10.
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from

Table S1:
DNA sequences of primers used for constructing recombinant proteins and GAS mutants in the present study.
Primer Name Sequence Restriction sites
1 GBSPy19-F CTGTACTCGAGGAGGATTTAAGTACTAAGATTGC XhoI
2 GBSPy19-R AAAGGATCCGTTAGTGTGGATAAATAAATGTAACAC BamHI
3 GB19Bam-UpNt TCAGGATCCAGCGTTACAGGCTCAAGTAAG BamHI
4 GB19Hin-UpCt TTTAAGCTTTCGCACCTACCATCGGCGTTG HindIII
5 GB19Nco-DnNt AAATCCATGGCTCCTTGGGCAGGAAA NcoI
6 GB19Pst-DnCt TGACTGCAGTTCAGCACTAGCACGC PstI
7 upNt-Pr3 GTCTAAGACGTTGGGGGAT
8 dnCt-Pr6 GCCGTAGTAAGGCATCAC
9 aad9-F TTGGATCAGGAGTTGAGAGTGGAC
10 aad9-R GCCACTGCATTTCCCGCAATATCT
11 GB19C55-F CTCGGATCCGTGGGCACTGAGTATGCCTATGG BamHI
12 GB19C55-R CCGAAGCTTATTAAGTTGTAGCTGCTCCGAGGG HindIII
13 GB19PDCHin-F TTCAAAGCTTGAAGGATAAGGTGTCTGAGTGAAG HindIII
14 GB19PDCKpn-R AACGGTACCGTCAATACTCTCAGAACTATTCC KpnI
15 AMB93SPy19RN-F TCCCCCCGGGATGAAAAAAAGAATTTTATCAGC XmaI
16 AMB94SPy19RN-R CGGGGTACCTTAGTGTGGATAAATAAATGTAAC KpnI
Bold underlined letters indicate the restriction enzyme sites.

Table S2: Strains used in the present study: ( two-pages)
Strain Characteristic/Description Reference/source
E. coli XL1-blue F’::Tn10proA+B+lacIq �(lacZ)M15/recA1endA1 gyrA96(Nal r)thi hsdR17 (rk- mk+) glnV44 relA1 lac
Stratagene
E. coli BL21(DE3)pLysS
E.coli B F-dcm ompT hsdS(rB-mB
-)gal λ(DE3)[pLysS Camr]
Novagen
E. coli MC1061 F’araD139 �(ara-leu)7696 �(lac)X74 galU galK hadR2(rk- mk+) mcrB1 rpsL (Str+)
From: Craig Rubens
(1)
Streptococcus pyogenes SF370
Wild-type strain ATCC 700294 (2)
Staphylococcus aureus RN4220
A derivative RN450 (ATCC8325-4) a Toxic-shock syndrome exotoxin-negative indicator strain
From Barry Kreisworth, PHRI
(3)
Plasmid
pET14B Vector used for making recombinant protein with His-tag at the N-terminal- AmpR
Novagen
pFW5 Suicide plasmid containing SpectinomycinR (aad9 resistance marker- SpcR) flanked by multiple cloning site (MCS-I) and MCS-II for constructing GAS mutant
Podbielski et al (4)
Jin and Pancholi (5)
pDC123 Plasmid used for complementation experiments-ChloramphenicolR (ChlR)
From: Craig Rubens (1)
pCN40tet
pCN40 ( E.coli-S. aureus shuttle vector) with ermC gene replaced by tet(M) from pCN36Plasmid used for complementing Staphylococcus aureus- tetracycline( TetR)
Charpentier et al (6)
Beltramini et al (7).
pET14B-cdhA pET14B plasmid with spy0019/cdhA insert without signal sequence
This study
pFW5-up19Ntct-1 pFW5 with upstream flanking region (930bp) in MCS-I(BamH1 and HindIII)
This study
pFW5-dn19Ntct-2 pFW5 with downstream flanking region (774bp) in MCS-II(NcoI and PstI)
This study
pFW5ΔcdhA pFW5 with flanking up and down stream regions (930bp and 774bp) in MCS-I and –II respectively
This study
pFWCdhAΔC55 pFW5 with 1075bp (157-1231) insert of cdhA in MCS-I This study
pDC123-cdhA pDC123 plasmid with cdhA insert between HindIII and KpnI
This study
pCN49-cdhA pCN49tet plasmid with cdhA insert between XmaI and KpnI
This study
Mutant strains
XL-1-cdhA XL-1 blue E. coli containing pET14B-cdhA plasmid This study

*References for Table S2
1. Chaffin D O, Rubens, C. E. (1998) Blue/white screening of recombinant plasmids in Gram-positive bacteria by interruption of alkaline phosphatase gene (phoZ) expression. Gene 219: 91-99.
2. Ferretti J J, McShan W M, Ajdic D, Savic D J, Savic G, Lyon, K. et al. (2001) Complete genome sequence of an M1 strain of Streptococcus pyogenes. Proc Natl Acad Sci USA 98: 4658-4663
3. Kreiswirth B, Lofdahl S, Betley M J, O'Reilly, M., Schlievert P M, Bergdoll Met (1983) The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 305: 709-712.
4. Podbielski A, Spellerberg B, Woischnik M, Poh B, Lutticken, R. (1996) Novel series of plasmid vectors for gene inactivation and expression analysis in group A streptococci (GAS). Gene 177: 137-147.
5. Jin H, Pancholi V (2006) Identification and biochemical characterization of a eukaryotic-type serine/threonine kinase and its cognate phosphatase in Streptococcus pyogenes: Their biological functions and substrate identification. J Mol Biol 357: 1351-1372.
6. Charpentier E, Anton A I, Barry P, Alfonso B, Fang Y, Novick R P (2004) Novel cassette-based shuttle vector system for Gram-positive bacteria. Appl Environ Microbiol 70: 6076-6085.
7. Beltramini A M, Mukhopadhyay C D, Pancholi V (2009) Modulation of cell wall structure and antimicrobial susceptibility by a Staphylococcus aureus eukaryotic-like serine/threonine kinase and phosphatase. Infect Immun 77: 1406-1416.
AmpR
BL21-cdhA BL21(DE3)pLysS E. coli containing pET14B-cdhA plasmid and expressing CdhA without its signal sequence. AmpR
This study
M1ΔSTK M1 SF370 lacking SPy1625 (SP-STK[eukaryotic-type serine/threonine kinase]) SpectinomycinR (SpcR)
Jin and Pancholi (5)
M1ΔSTK::cdhA M1ΔSTK complemented with wild-type cdhA- SpcR and ChloramphenicolR(ChlR)
This study
M1ΔCdhA M1 SF370 lacking cdhA- SpcR This study
M1CdhAΔC55 M1 SF370 expressing truncated CdhA without the C-terminal 55 residues that contributes to the catalytic site of the CHAP-domain- SpcR
This study
M1ΔCdhA::cdhA M1ΔCdhA mutant complemented with the wild-type cdhA- SpcR-ChlR
This study
M1-WT::cdhA Wild-type M1 complemented with cdhA resulting in over expression of CdhA-ChlR
This study
RN4220::cdhA Wild-type RN4220 complemented with cdhA resulting in over expression of CdhA-tetR
This study

Table S3 : Primers sequences used for qRT-PCR assays in the present study.
Gene ORF Gene Symbol
Sequence
Forward Reverse
SPy1152 gyrA TTCGTATGGCTCAGTGGTTTAGTT CTGGTTCTCTTTCGCTTCCATCGT
SPy1962 ProS GGGTGGTTCTTGACAAGTCTATTGCG TTCTGCCAAGGCATCTTCAGCA
SPy0019 cdhA/sibA TCTTGTAAGTGGTGTTACCCTCGG GCGCCTCAAGTTCTGTATTTCTTGCG
SPy0136 unknown TTGGGACGGTACAGTTCGTTCTGA AGTGTGTGATTGACGGCCATAGGT
SPy0148 ntpl GCCATCGACAACAAGCAGCTCAAA AGGAATCGTTCCAACTCTTGCCGA
SPy0167 slo AGCTCCGCCACTCTTTGTGAGTAA CTCTGCAGCATCTCCTCC
SPy0252 PTS AAATGCTCAAAGATGCTGGCGTCC TGGTCACCACCTTGACCATTAGCA
SPy0711 speC AAGCGGCAGAATCGAAATTGGCAC TCTAGTCCCTTCATTTGGTGAGTC
SPy0712 mf2 GCAATGACAGCCTGGCTAAACACA TCGAGGCACTACTTCATTGCCACT
SPy0738 sagA GCTGCTGTTGCTGCTGTACTACTT GCGTATAACTTCCGCTACCACCTT
SPy0855 PTS GCCATTTGTTATTGGTGGCGGGAT AGCTGATCATTTGGCACACCAAGC
SPy1058 PTS GACCAAAGAGTTAAATGCGCCGCT CTTGTCTTTGGCGCGTGGATCATT
SPy1294 malB TTCCTTTGAATCAGGACCGTGGGA TTTAGCTGGTGCTTGGTTGACTGC
SPy1357 grab ACTCACCTATCGAACAGCCTCGAA GCGTCTGTTTGATCCGCTAATGCT
SPy1917 lacE TTACCATGGGTGGTACTGGTGCAA CTGATGCACGTCCAATGGCTTTGT
SPy1983 scl AAATCTGGAATCAAGGGAGACCGTGG TTTCACCGCGTTCACCGGTTTCA
SPy2018 emm1 TTAGCGATAGACCAAGCGAGTCAG GCTTTGACGACTTGCGTCTGAGAT
SPy2019 mga GACCTTTCACGCTTGTTTCACCTC GTGACGCTGAGATTGAGCCTGATT
SPy2039 speB TTCTGCAGGTAGCTCTCGTGTTC TCGATAACAAAGGCATGTCCGCC
SPy2200 hasA GCCACATGACTATAAAGTTGCTGCTG TTGACAAGGGAACGGTGAACGA

Table S4: Microarray-based differential gene expression analysis for M1ΔCdhA vs the wild-type M1-SF370
(GSE#15546).

ORF#
M1-SF370 Gene Functional category
Log2-fold change
(M1ΔCdhA vs M1-WT)
¶ P value*
Amino acid transport and metabolism
SPy0184 opuABC putative glycine-betaine binding permease protein 2.05640 0.00598
SPy1541 arcC putative carbamate kinase -3.4416 0.00153
SPy1542 - putative Xaa-His dipeptidase -3.29318634 0.00626
SPy1544 arcB putative ornithine transcarbamylase -2.93909102 0.02302
SPy1547 sagP (arcA)
streptococcal antitumor protein (possible arginine deiminase) -2.75651843 0.02362
SPy2189 sdhB putative L-serine dehydratase beta subunit -1.63081526 0.00773
SPy2190 sdhA putative L-serine dehydratase alpha subunit -1.78756811 0.0179
Carbohydrate transport and metabolism
SPy0146 - putative regulatory protein -3.10918311 0.00079
SPy0255 - putative regulatory protein -3.58895686 0.00364
SPy1057 - Hypothetical protein -2.58079205 0.03288
SPy1058 - putative phosphotransferase system (PTS), enzyme II component B -2.95458551 0.00816
SPy1059 - putative phospotransferase system (PTS), enzyme II, component C -2.67254237 0.01141
SPy1060 - putative phospotransferase system (PTS), enzyme II component D -2.16720074 0.03663
SPy1292 malM putative 4-alpha-glucanotransferase -2.3532552 0.04502
SPy1294 - putative maltose/maltodextrin-binding protein -2.08709915 0.05454
SPy1301 malC maltodextrin transport system permease -2.72742748 0.02236
SPy1304 amyB putative cyclomaltodextrinase -2.58349588 0.01888
SPy1707 lacB.1 putative galactose-6-phosphate isomerase -2.80990347 0.00963
SPy1708 lacA.1 putative galactose-6-phosphate isomerase -2.88753155 0.00565
Cell envelope biogenesis, outer

membrane
SPy2201 hasB UDP-glucose 6-dehydrogenase -5.83464339 1.08E-11
SPy2202 hasC UDP-glucose pyrophosphorylase -4.7896666 5.12E-06
Cell motility and secretion
SPy0857 mur1.2 putative peptidoglycan hydrolase 1.775223248 0.04556
DNA replication, recombination and repair
SPy1563 - conserved hypothetical protein -2.0734014 0.02289
Energy production and conversion
SPy0149 ntpK V-type Na+ -ATPase subunit K -2.36833651 0.0096
SPy0150 ntpE putative V-type Na+ -ATPase subunit E -2.83273618 0.00809
SPy0154 ntpA putative V-type Na+ -ATPase alpha subunit -3.07025116 0.00224
SPy0155 ntpB putative V-type Na+ -ATPase subunit B -2.79904425 0.03036
SPy0157 ntpD putative V-type Na+ -ATPase subunit D -2.89979146 0.00292
Function unknown
SPy0019 - putative secreted protein/ CdhA/sibA -3.72812322 0.00025
SPy0136 - hypothetical protein -3.80404194 0.00027
SPy0166 - hypothetical protein -3.09748473 0.0004
SPy0170 - hypothetical protein -4.80814656 1.55E-05
SPy0256 - hypothetical protein -2.4527254 0.00655
SPy0428 - conserved hypothetical protein -3.35991921 0.00024
SPy0430 - hypothetical protein -3.6328223 2.23E-05
SPy0433 - hypothetical protein -2.62327812 9.64E-05
SPy0435 - hypothetical protein -2.79926332 0.0008
SPy0437 - hypothetical protein -1.38948653 0.0458
SPy0743 - hypothetical protein -1.37395731 0.05468
SPy0861 - hypothetical protein -2.24634217 0.00658
SPy1017 - hypothetical protein 1.708325383 0.02793
SPy1543 - conserved hypothetical protein -3.33026296 0.00269

SPy1562 - conserved hypothetical protein -3.06442454 0.00091
SPy1564 - conserved hypothetical protein -2.44132327 0.00335
SPy1565 - conserved hypothetical protein -2.9655306 0.00187
SPy1686 - hypothetical protein 1.562696323 0.03546
SPy1710 - putative PTS system, enzyme IIB component -2.00494928 0.02123
SPy1832 - hypothetical protein -1.53364716 0.02572
SPy1985 - hypothetical protein -1.9194945 0.03479
General function prediction only
SPy0742 - hypothetical protein -1.5674321 0.04718
Lipid metabolism
SPy1718 - putative esterase -2.13943137 0.01467
Posttranslational modification, protein turnover, chaperones
SPy0416 prtS putative cell envelope proteinase -2.92746688 0.00015
Signal transduction
SPy1062 - putative two-component response regulator -2.24109495 0.02052
Transcription
SPy1546 - hypothetical protein -2.78005908 0.01756
Virulence factors
SPy0165 nga NAD glycohydrolase precursor -4.70774684 2.27E-06
SPy0167 slo streptolysin O precursor -3.74007054 7.33E-05
SPy0711 speC pyrogenic exotoxin C precursor, phage associated 1.784534856 0.0182
SPy0712 mf2 putative DNase (mitogenic factor), phage 1.66593819 0.00257
SPy1302 amyA putative cyclomaltodextrin glucanotransferase -2.74293186 0.02244
SPy1357 grab protein GRAB (protein G-related ?2M-binding protein) 1.860433621 0.02545
SPy1983 scl collagen-like surface protein -5.45831621 5.50E-06
SPy2016 sic inhibitor of complement-mediated lysis -3.76662345 0.00078

¶-- Microarray analysis and Log2-fold changes were derived from 11 independent experiments including dye-swap experiments as described in Material Methods using the CARMAweb software.
*-- Differential gene expression value, + Log2>1 fold change with p< 0.05, was treated as significant change.
# -Open reading frame (ORF) numbers are based on the genome sequence data of type M1 GAS strain SF370 (2).
OPR # in bold letters are also found in M1CdhADC55 mutant (Table-S2)
SPy2018 emm1 M protein type 1 -4.16572585 0.00012
SPy2039 speB pyrogenic exotoxin B -3.59998351 0.00128
SPy2043 mf mitogenic factor -2.25560173 0.02301
SPy2200 hasA hyaluronate synthase -7.19307349 6.59E-13

Table S5. Microarray-based differential gene expression analysis for M1CdhAΔC55 vs M1SF-370 (GSE# 15598).
ORF#
(M1-SF370) Gene Functional category and Product
Log2-fold change
M1CdhADC55 vs.M1SF370¶
P Value*
Amino acid transport and metabolism
SPy0257 nanH putative acylneuraminate lyase -2.25493 0.042168
SPy1217 - putative glycine cleavage system H protein -1.58447 0.040303
SPy1541 arcC putative carbamate kinase -4.36467 7.71E-06
SPy1542 - putative Xaa-His dipeptidase -5.34734 1.97E-08
SPy1544 arcB putative ornithine transcarbamylase -4.76064 1.63E-06
SPy1547 sagP (arcA) streptococcal antitumor protein (possible arginine deiminase) -4.46374 1.23E-06
SPy2190 sdhA putative L-serine dehydratase alpha subunit -1.83436 0.013469
Carbohydrate transport and metabolism
SPy0146 - putative regulatory protein -2.70835 0.0014
SPy0251 - putative N-acetylmannosamine-6-P epimerase -2.87709 0.003248
SPy0252 - putative sugar transporter sugar binding lipoprotein -2.93048 0.000724
SPy0254 - sugar transport system (permease) -2.40877 0.00528
SPy0442 - putative glycerol-3-phosphate transporter -2.10287 0.008705
SPy0854 fruB putative fructose-1-phosphate kinase -3.43612 4.05E-05
SPy1057 - hypothetical protein -4.92344 4.41E-08
SPy1058 - putative phosphotransferase system (PTS), enzyme II component B -4.48982 1.10E-06
SPy1059 - putative phospotransferase system (PTS), enzyme II, component C -4.35334 1.11E-06
SPy1060 - putative phospotransferase system (PTS), enzyme II component D -3.52878 9.14E-05
SPy1292 malM putative 4-alpha-glucanotransferase -3.11974 0.00104
SPy1294 - putative maltose/maltodextrin-binding protein -3.50005 0.00015
SPy1295 malF putative maltose/maltodextrin ABC transport system (permease) -3.02496 0.004624

SPy1296 malG putative maltose/maltodextrin ABC transport system (permease) -1.82093 0.023356
SPy1299 malD putative maltodextrin transport system permease -2.53865 0.00837
SPy1301 malC maltodextrin transport system permease -3.62747 6.60E-05
SPy1304 amyB putative cyclomaltodextrinase -3.96239 3.67E-05
SPy1306 malX maltose/maltodextrin-binding protein -3.63956 0.001779
SPy1503 - putative phosphomannomutase -2.00506 0.00482
SPy1704 lacD.1 putative tagatose 1,6-diphosphate aldolase -2.07113 0.018109
SPy1707 lacB.1 putative galactose-6-phosphate isomerase -3.03622 0.001591
SPy1708 lacA.1 putative galactose-6-phosphate isomerase -2.45743 0.008592
SPy1709 - putative PTS system, enzyme IIC component -2.82759 0.003145
SPy1711 - putative PTS system, enzyme IIA component -2.49478 0.010317
SPy1922 lacB.2 putative galactose-6-phosphate isomerase (B subunit) -1.7812 0.036192
SPy1923 lacA.2 galactosidase acetyltransferase -2.52042 0.011223
SPy1973 dexB dextran glucosidase -2.20939 0.006815
SPy2096 dexS putative dextran glucosidase -1.6257 0.037289
Cell envelope biogenesis, outer membrane
SPy2201 hasB UDP-glucose 6-dehydrogenase -4.53654 9.53E-07
SPy2202 hasC UDP-glucose pyrophosphorylase -3.8781 9.81E-05
Cell motility and secretion
SPy1976 msmK multiple sugar-binding ABC transport system (ATP-binding protein) -2.61871 0.000912
Coenzyme metabolism
SPy1063 - putative periplasmic-iron-binding protein -2.00402 0.007379
SPy1214 lplA putative lipoate-protein ligase -1.58223 0.026625
SPy1220 - conserved hypothetical protein -1.52459 0.02995
DNA replication, recombination and repair
SPy1563 - conserved hypothetical protein -2.17083 0.00679
Energy production and conversion
SPy0148 ntpI V-type Na+ -ATPase subunit I -3.78279 4.95E-05

SPy0149 ntpK V-type Na+ -ATPase subunit K -3.76487 4.38E-05
SPy0150 ntpE putative V-type Na+ -ATPase subunit E -4.07994 3.00E-05
SPy0151 ntpC putative V-type Na+ -ATPase subunit C -3.7906 8.06E-05
SPy0154 ntpA putative V-type Na+ -ATPase alpha subunit -3.8587 5.44E-05
SPy0155 ntpB putative V-type Na+ -ATPase subunit B -4.13006 1.78E-05
SPy0157 ntpD putative V-type Na+ -ATPase subunit D -3.74782 2.40E-05
SPy1218 - conserved hypothetical protein -1.92363 0.010479
SPy1849 pfl putative pyruvate formate-lyase -2.32763 0.002428
SPy2049 pflD putative pyruvate formate-lyase 2 -1.44584 0.051227
Function unknown
SPy0019 - putative secreted protein—CdhA/SibA 1.76572 0.012404
SPy0136 - hypothetical protein -2.4309 0.014809
SPy0147 - hypothetical protein -3.60598 0.000571
SPy0170 - hypothetical protein -3.225 0.001498
SPy0256 - hypothetical protein -2.98326 0.000944
SPy0430 - hypothetical protein -2.8642 0.002072
SPy0433 - hypothetical protein -1.89022 0.020243
SPy0435 - hypothetical protein -1.54147 0.050185
SPy0739 sagB streptolysin S associated ORF -2.97651 0.00026
SPy0740 sagC streptolysin S associated ORF -2.77969 0.001431
SPy0741 - hypothetical protein -2.66414 0.001328
SPy0743 - hypothetical protein -2.44239 0.003764
SPy0861 - hypothetical protein -2.34387 0.033703
SPy1216 - hypothetical protein -1.99825 0.011072
SPy1298 malA putative maltodextrose utilization protein -2.15022 0.016366
SPy1543 - conserved hypothetical protein -4.54638 5.13E-07
SPy1562 - conserved hypothetical protein -1.97883 0.006385
SPy1565 - conserved hypothetical protein -2.31821 0.002193
SPy1680 - putative transcriptional regulatory protein -1.6107 0.020366

SPy1710 - putative PTS system, enzyme IIB component -2.48215 0.011916
SPy1985 - hypothetical protein -2.13957 0.007286
SPy2066 - putative dipeptidase -1.73751 0.024801
SPy2191 - hypothetical protein 1.753186 0.017852
General function prediction only
SPy0044 adhA putative alcohol dehydrogenase I -3.09639 0.00021
SPy0519 - conserved hypothetical protein -1.645 0.046764
SPy0742 - hypothetical protein -2.87017 0.000685
SPy0745 sagH ABC transporter (ATP-binding protein) - streptolysin S associated ORF -2.57412 0.001301
SPy0746 - putative ABC transporter (ATP-binding protein) -2.14013 0.002678
SPy0747 - conserved hypothetical protein -1.95705 0.010762
SPy1113 - putative acid phosphatase (class B) -1.7422 0.028163
SPy1567 - conserved hypothetical protein -1.73871 0.012842
SPy1693 - putative reductase / dehydrogenase -1.31522 0.039902
Lipid metabolism
SPy1751 fabK putative trans-2-enoyl-ACP reductase II 1.255345 0.04672
SPy0022 plsX putative fatty acid/phospholipid synthesis protein 1.398864 0.035703
SPy1753 acpP putative acyl carrier protein 1.482747 0.041113
Nucleotide transport and metabolism
SPy0830 pyrR putative pyrimidine regulatory protein 1.680411 0.025495
SPy0832 pyrB putative aspartate transcarbamoylase 1.811749 0.014822
SPy0900 pyrF putative orotidine-5'-decarboxylase PyrF 2.225126 0.008888
Posttranslational modification, protein turnover, chaperones
SPy0379 pflC putative pyruvate-formate lyase activating enzyme -1.73878 0.028716
SPy0416 prtS putative cell envelope proteinase -2.13093 0.003128
Secondary metabolites biosynthesis, transport and catabolism
SPy0744 - Putative ABC transporter (ATP-binding protein) -2.74699 0.000836
Signal transduction mechanisms

¶-- Microarray analysis and Log2-fold changes were derived from 7 independent experiments including dye-swap experiments using the CARMAweb software as described in Materials and Methods .
*-- Differential gene expression value, + Log2>1 fold change with p< 0.05, was treated as significant change.
# -Open reading frame (ORF) numbers are based on the genome sequence data of type M1 GAS strain SF370 (2).
OPR # in bold letters are also found in M1ΔCdhA mutant (Table S1).
SPy1061 - putative two-component sensor histidine kinase -2.43915 0.005105
Transcription
SPy0853 - putative transcriptional repressor -3.96736 8.99E-07
SPy1062 - putative two-component response regulator -2.2121 0.024562
SPy1546 - hypothetical protein -4.67337 1.47E-06
Translation
SPy0145 - conserved hypothetical protein -1.61106 0.024883
SPy1613 - conserved protein - function unknown -1.60667 0.021384
Virulence genes
SPy0711 speC pyrogenic exotoxin C precursor, phage associated 1.64463 0.032466
SPy0712 mf2 putative DNase (similar to mitogenic factor), phage associated 1.652306 0.026745
SPy0738 sagA streptolysin S associated protein -2.45486 0.004107
SPy1302 amyA putative cyclomaltodextrin glucanotransferase -3.63063 4.88E-05
SPy1436 mf3 putative deoxyribonuclease 2.130089 0.008555
SPy1983 scl collagen-like surface protei -4.48531 3.10E-06
SPy2018 emm1 M protein type 1 -2.90199 0.000887
SPy2043 mf mitogenic factor -1.67711 0.021443
SPy2200 hasA hyaluronate synthase -4.61878 4.43E-06
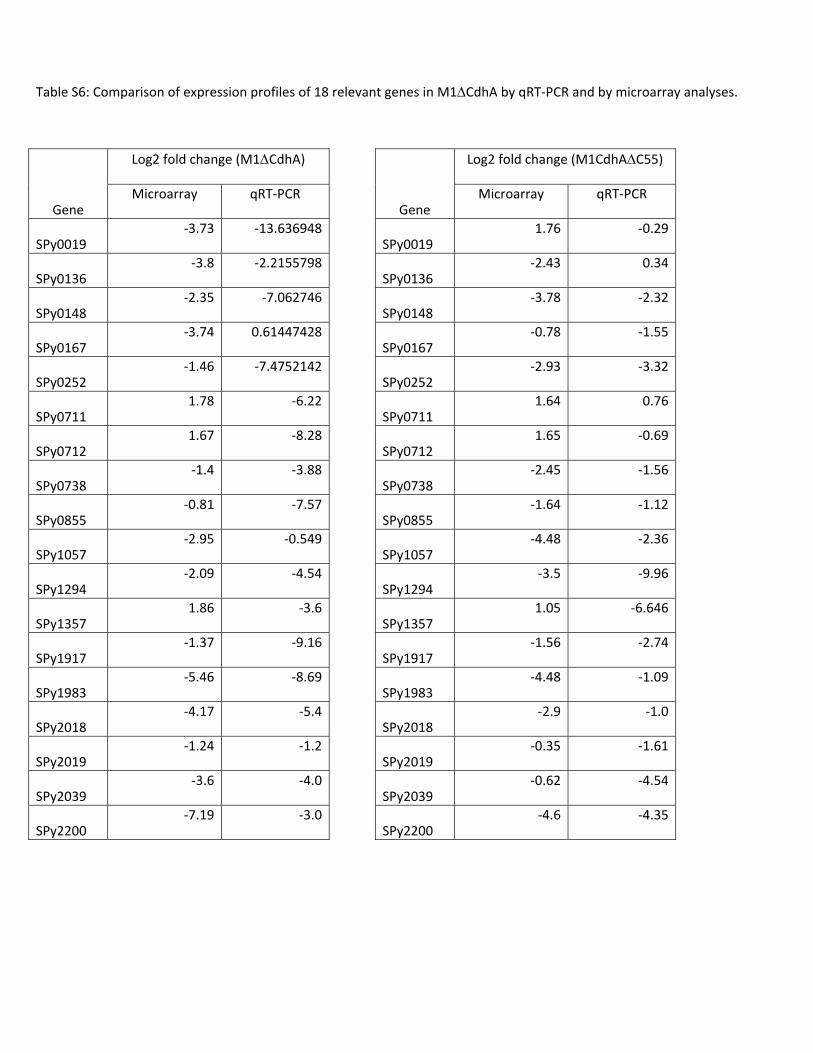
Table S6: Comparison of expression profiles of 18 relevant genes in M1ΔCdhA by qRT‐PCR and by microarray analyses.
Gene
Log2 fold change (M1ΔCdhA)
Microarray qRT‐PCR
SPy0019 ‐3.73 ‐13.636948
SPy0136 ‐3.8 ‐2.2155798
SPy0148 ‐2.35 ‐7.062746
SPy0167 ‐3.74 0.61447428
SPy0252 ‐1.46 ‐7.4752142
SPy0711 1.78 ‐6.22
SPy0712 1.67 ‐8.28
SPy0738 ‐1.4 ‐3.88
SPy0855 ‐0.81 ‐7.57
SPy1057 ‐2.95 ‐0.549
SPy1294 ‐2.09 ‐4.54
SPy1357 1.86 ‐3.6
SPy1917 ‐1.37 ‐9.16
SPy1983 ‐5.46 ‐8.69
SPy2018 ‐4.17 ‐5.4
SPy2019 ‐1.24 ‐1.2
SPy2039 ‐3.6 ‐4.0
SPy2200 ‐7.19 ‐3.0
Gene
Log2 fold change (M1CdhAΔC55)
Microarray qRT‐PCR
SPy0019 1.76 ‐0.29
SPy0136 ‐2.43 0.34
SPy0148 ‐3.78 ‐2.32
SPy0167 ‐0.78 ‐1.55
SPy0252 ‐2.93 ‐3.32
SPy0711 1.64 0.76
SPy0712 1.65 ‐0.69
SPy0738 ‐2.45 ‐1.56
SPy0855 ‐1.64 ‐1.12
SPy1057 ‐4.48 ‐2.36
SPy1294 ‐3.5 ‐9.96
SPy1357 1.05 ‐6.646
SPy1917 ‐1.56 ‐2.74
SPy1983 ‐4.48 ‐1.09
SPy2018 ‐2.9 ‐1.0
SPy2019 ‐0.35 ‐1.61
SPy2039 ‐0.62 ‐4.54
SPy2200 ‐4.6 ‐4.35

Vijay Pancholi, Gregory Boel and Hong Jindivision-plane-recognizing and chain-forming virulence factor
SER/THR kinase-regulated cell wall hydrolase is a cellS. pyogenes
published online July 19, 2010J. Biol. Chem.
10.1074/jbc.M110.153825Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/07/19/M110.153825.DC1.html
http://www.jbc.org/content/early/2010/07/19/jbc.M110.153825.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 22, 2016http://w
ww
.jbc.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















