
Evidence for the use of an internal sense of direction in homing
Upload
independentCategory
view
0download
0
Exp. Anim. 57(4), 347–356, 2008
Role of MAdCAM-1 and Its Ligand on the Homing of Transplanted Hematopoietic Cells in Irradiated Mice
Tatsuya TADA1), Naoko INOUE1), Diah Tri WIDAYATI2), and Katsuhiro FUKUTA1)
1)Laboratory of Animal Morphology and Function, Graduate School of Bioagricultural Sciences, Nagoya University, Chikusa-ku, Nagoya 464-8601, Japan and 2)Faculty of Animal Science,
Gadjah Mada University, Yogyakarta, Indonesia
Abstract: We examined the expression of VCAM-1 and MAdCAM-1 after bone marrow transplantation (BMT). We also examined the influence of α4β7 integrin blockade on the homing of cells to the bone marrow and spleen. The expression of VCAM-1 and MAdCAM-1 by endothelial cells in the spleen and bone marrow was examined by immunoelectron microscopy using colloidal gold and was analyzed semi‑quantitatively. To examine the role of α4β7 integrin in donor cells, a homing assay was conducted following α4β7 integrin blockade in bone marrow‑derived hematopoietic cells or spleen colony cells. Immediately after BMT, the expression of VCAM‑1 and MAdCAM‑1 markedly decreased, but expression recovered significantly between 12 and 24 h after BMT. VCAM‑1 recovered more acutely than MAdCAM‑1 from 12 h onward. In the group transplanted with anti‑α4β7 integrin antibody-treated bone marrow cells, the numbers of homing cells in the spleen and bone marrow were significantly decreased in an antibody dose‑dependent manner. However, the number of homing cells was not different in either the spleen or bone marrow between anti-α4β7 integrin antibody treated and untreated spleen colony cells. It has been reported that α4β1 integrin and its receptor VCAM‑1 play major roles in the homing of hematopoietic cells to bone marrow. Our study indicates the importance of MAdCAM‑1 and its ligand, α4β7 integrin, in the homing of bone marrow‑derived hematopoietic cells, but not spleen colony‑derived cells, to both the spleen and bone marrow.Key words: α4β7 integrin, endothelial cells, homing, MAdCAM-1, VCAM-1
(Received 3 October 2007 / Accepted 17 December 2007)Address corresponding: K. Fukuta, Laboratory of Animal Morphology and Function, Graduate School of Bioagricultural Sciences, Nagoya University, Chikusa-ku, Nagoya 464-8601, Japan
Introduction
Interactions between hematopoietic tissues and he-matopoietic stem/progenitor cells (HSPCs) are essential in regulating the proliferation and differentiation of HSPCs [10, 13, 33]. This regulation appears to be medi-ated by mutual recognition between adhesion molecules expressed by stromal and vascular endothelial cells
(ECs) of hematopoietic tissues and their ligands on HSPCs [26, 29, 35, 37]. Moreover, it is well known that cell adhesion molecules and ligands regulate the migra-tion of HSPCs from the blood stream into the hematopoi-etic compartment [27]. After BMT following lethally irradiation, donor cells migrate, settle, and proliferate in hematopoietic tissues. In this process, interactions be-tween cell adhesion molecules on ECs and surface anti-

348 T. TADA, ET AL.
gens on donor cells play important roles in homing. Previous studies have shown that various cell adhesion pathways are related to the recruitment of blood cells into bone marrow parenchyma. For example, α4β1 in-tegrin of HSPCs and its receptor, vascular cell adhesion molecule-1 (VCAM-1), play key roles in the homing of HSPCs to bone marrow [15, 30]. Ligands of selectin-endothelial cell and selectin-platelet regulate the recruit-ment of neutrophils [8, 14, 25], and selectin-endothelial cell ligand and α4 integrins regulate hematopoietic stem cells [22]. Moreover, it has been reported that α4β1 in-tegrin, α5β1 integrin, β2 integrin, and sialophorin are expressed on the surface of HSPCs. These factors affect lymphocyte tethering and adherence to ECs in bone mar-row [3, 31] and thus may similarly mediate the homing of HSPCs in bone marrow. However, the changes of cell adhesion molecules on ECs and their ligands on donor cells following BMT remain unclear.
Mucosal addressin cell adhesion molecule-1 (MAd-CAM-1) is a homing receptor and is expressed in the high endothelial venules of the mesenteric lymph nodes and Peyer’s patch, where it plays a central role in leu-kocyte migration into these tissues [18–20]. In mice, MAdCAM-1 is expressed on the cells of the marginal sinus in the spleen [24]. The α4β7 integrin (LPAM-1; lymphocyte Peyer’s patch adhesion molecule-1), which is a ligand of MAdCAM-1, is expressed on leukocytes and mediates leukocyte homing to mucosal lymphoid tissues [5]. There have been few reports on the effects of MAdCAM-1 on the homing of hematopoietic cells to the spleen and bone marrow.
To examine the roles of MAdCAM-1 and VCAM-1 on the homing of HSPC, we investigated their expression in ECs of spleen and bone marrow from 1 to 24 h after bone marrow transplantation (BMT). In addition, we examined the influence of α4β7 integrin blockade on cell homing in bone marrow and spleen, using green fluores-cent protein (GFP) blood chimeric mice.
Materials and Methods
AnimalsAll studies were performed in accordance with the
guidelines for the care and use of experimental animals of Nagoya University. Male BALB/c mice (8–10 weeks
old) were purchased from a commercial breeder, Japan SLC (Hamamatsu, Japan), and C57BL/6N mice were purchased from CLEA Japan (Tokyo, Japan). Trans-genic mice with the enhanced green fluorescent protein gene (GFP mice) were a gift from Drs. Okabe and Ikawa, Osaka University, Japan [21, 28].
Experiment 1Bone marrow transplantation in BALB/c mice
As hematopoietic cells for transplantation, bone mar-row mononuclear cells (BMCs) were obtained from BALB/c mice by flushing the femoral and tibial bones with phosphate-buffered saline (PBS), and the suspen-sion was adjusted to 2 × 107 cells/ml (final concentra-tion). Recipient BALB/c mice were exposed to a cumu-lative dose of about 9.5 Gy of whole body irradiation, using an X-ray irradiator (MBR-1520, Hitachi, Japan). Within 6 h following irradiation, the mice were injected with 1 × 107 BMCs through the tail vein. The mice were sacrificed at 1, 3, 6, 12, and 24 h after BMT (4 mice at each point), and spleens and bone marrow were removed for immunoelectron microscopy.
Immunoelectron microscopyTo semi-quantitatively evaluate the expression of
VCAM-1 and MAdCAM-1 in vascular ECs, colloidal gold was used in the preparation of specimens for im-munoelectron microscopy. Tissues from the transplant-ed BALB/c mice were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer at 4°C. After fixation, the femurs were decalcified in 5% neutral EDTA solution. The specimens were embedded in OCT compound (Sakura Finetechnical, Tokyo, Japan) and frozen for cryostat sectioning. Sections (18 µm thick) were subjected to immunohistochemistry. Rat anti-mouse VCAM-1 anti-body (Santa Cruz Biotechnology, CA, USA) and rat anti-mouse MAdCAM-1 antibody (Santa Cruz Biotech-nology) were used as primary antibodies at dilutions of 1:400 and 1:200, respectively. Goat anti-rat IgG anti-body conjugated with 5-nm colloidal gold particles was used as the secondary antibody (BBInternational, Car-diff, UK). The small gold particles were then silver-enhanced for 5 min at room temperature, using an HQ silver enhancement kit (Nanogold reagent, Nanoprobe Inc., NY, USA). The cryostat sections were post-fixed

349ADHESION MOLECULES IN HEMATOPOIETIC CELL HOMING
with 1% OsO4 for 15 min, dehydrated through graded ethanol solutions, and embedded in Epon. Ultra-thin sections were stained with uranyl acetate for 20 min and observed using a transmission electron microscope (JEM-1210, JEOL, Tokyo, Japan).
Experiment 2Production of GFP+ spleen colonies
For transplantation, a suspension of GFP+BMCs was prepared by using PBS to flush the femoral and tibial bones of C57BL/6N mice carrying the GFP gene (GFP mice), and the suspension was adjusted to 4 × 105 cells/ml (final concentration). To produce GFP+ spleen colo-nies, 2 × 105 GFP+BMCs were injected into lethally ir-radiated (9.8 Gy) C57BL/6 mice within 6 h following irradiation. At 12 days after BMT, the spleens were removed for histological studies and for preparation of spleen cell suspensions. For histological studies, the removed spleens from 5 mice were observed under a fluorescence stereomicroscope (SZX12, Olympus, Ja-pan) to identify GFP+ spleen colonies, and these spleens were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer at 4°C, embedded in OCT compound (Sakura Finetechnical), and frozen for cryostat sectioning. Sec-tions (8 µm thick) were counterstained with propidium iodide (Sigma) and observed under a confocal laser scan-ning microscope (LSM5 PASCAL, Carl Zeiss, Germa-ny).
Preparation of GFP+BMCs and GFP+ spleen colony cell suspensions
GFP+BMCs were obtained by flushing the femoral and tibial bones of GFP mice with PBS, and the suspension was adjusted to 1 × 107 cells/ml (final concentration). GFP+ spleen colony cells (SCCs) were prepared by crushing spleens with GFP+ spleen colonies at 12 days, and the suspension was adjusted to 1 × 107 cells/ml (final concentration) with PBS.
α4β7 integrin blockade in donor cellsTo block the ligand of MAdCAM-1, rat anti-mouse
α4β7 integrin (LPAM-1) antibody (Southern Biotech, Birmingham, AL) was used. The prepared GFP+BMC and GFP+SCC suspensions were incubated with the anti-α4β7 integrin antibody (2 or 4 µg/107 cells) for 2 h at
4°C and these cells were washed with PBS to remove free antibody. Then, antibody-treated or untreated cells (5 × 106 cells) were transplanted into lethally irradiated (9.8 Gy) C57BL/6 mice. Four mice were used for each antibody concentration and 3 for untreated cells. At 12 h after transplantation, the spleens and femurs were re-moved for a histological homing assay.
Homing assay of cellsTissues from the C57BL/6 mice transplanted with
GFP+BMCs or GFP+SCCs were fixed in 4% paraform-aldehyde in 0.1 M phosphate buffer at 4°C. After fixa-tion, the femurs were decalcified in EDTA solution. The specimens were embedded in OCT compound and frozen for cryostat sectioning. Sections (8 µm thick) were counterstained with propidium iodide and observed un-der a confocal laser scanning microscope.
Spleen colony assayGFP+BMCs from GFP mice were adjusted to
4 × 105 cells/ml (final concentration) and incubated with anti-α4β7 integrin antibody (1.25 µg/105 cells) for 2 h at 4°C. Then, antibody-treated or untreated cells (2 × 105 cells) were transplanted into lethally irradiated (9.8 Gy) C57BL/6 mice (n=5 in each group). At 12 days after transplantation, spleen colonies were counted under a fluorescence stereomicroscope.
Statistical analysesColloidal gold particles on the ECs of spleen and bone
marrow were counted to evaluate the expression level of cell adhesion molecules. The data are presented as the number of gold particles/10 µm at the luminal surface of the endothelial cell; ten ECs at each stage were se-lected to determine the average number. For the homing assay, the number of GFP+cells was counted in 0.25 mm2 of spleen or bone marrow parenchyma. Eight sections were selected at random to determine the average num-ber. The changes in MAdCAM-1 and VCAM-1 expres-sion at time points from 1 to 24 h after BMT were as-sessed by one-way analysis of variance (ANOVA), followed by the Tukey-Kramer post hoc test. Other data were assessed using Student’s t-test.
350 T. TADA, ET AL.
Results
Expression of VCAM-1 and MAdCAM-1 in spleen and bone marrow ECs
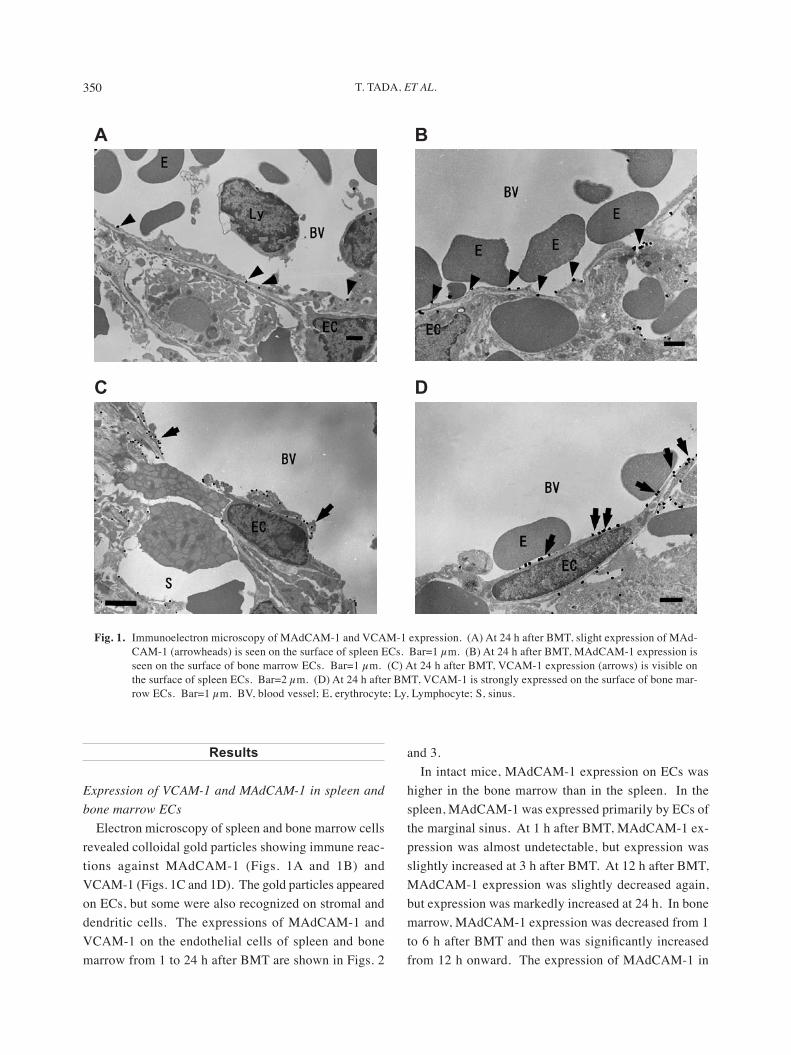
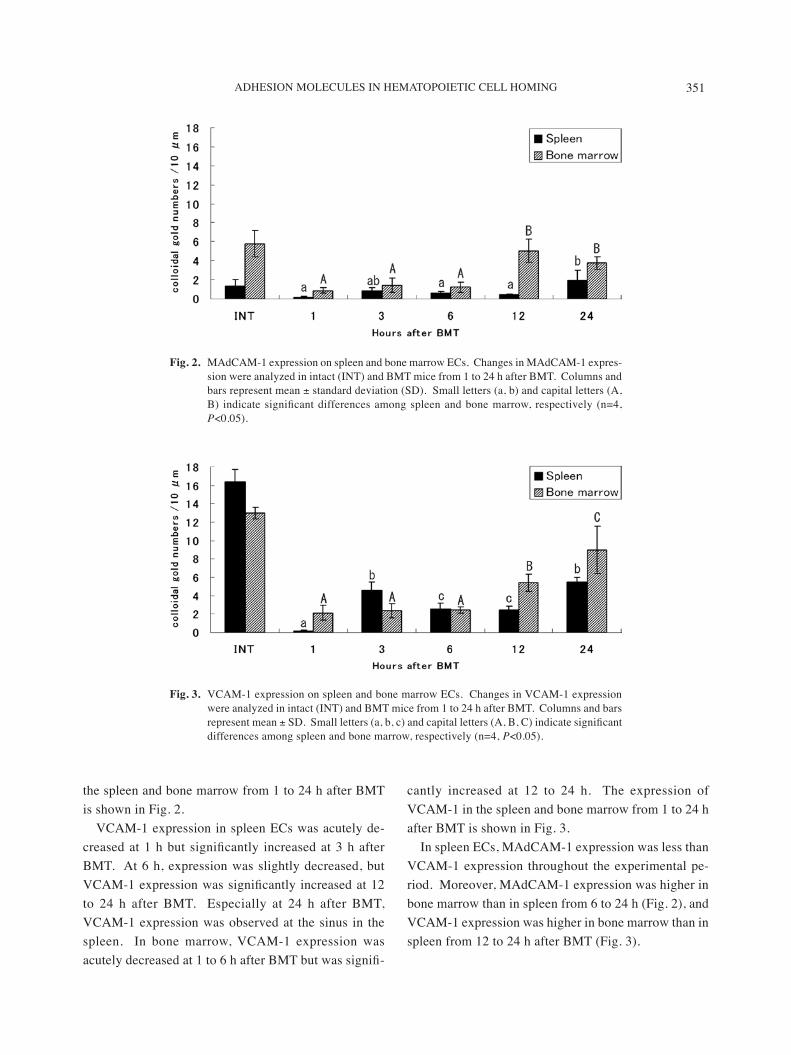
Electron microscopy of spleen and bone marrow cells revealed colloidal gold particles showing immune reac-tions against MAdCAM-1 (Figs. 1A and 1B) and VCAM-1 (Figs. 1C and 1D). The gold particles appeared on ECs, but some were also recognized on stromal and dendritic cells. The expressions of MAdCAM-1 and VCAM-1 on the endothelial cells of spleen and bone marrow from 1 to 24 h after BMT are shown in Figs. 2
and 3.In intact mice, MAdCAM-1 expression on ECs was
higher in the bone marrow than in the spleen. In the spleen, MAdCAM-1 was expressed primarily by ECs of the marginal sinus. At 1 h after BMT, MAdCAM-1 ex-pression was almost undetectable, but expression was slightly increased at 3 h after BMT. At 12 h after BMT, MAdCAM-1 expression was slightly decreased again, but expression was markedly increased at 24 h. In bone marrow, MAdCAM-1 expression was decreased from 1 to 6 h after BMT and then was significantly increased from 12 h onward. The expression of MAdCAM-1 in
A B
C D
Fig. 1. Immunoelectron microscopy of MAdCAM-1 and VCAM-1 expression. (A) At 24 h after BMT, slight expression of MAd-CAM-1 (arrowheads) is seen on the surface of spleen ECs. Bar=1 µm. (B) At 24 h after BMT, MAdCAM-1 expression is seen on the surface of bone marrow ECs. Bar=1 µm. (C) At 24 h after BMT, VCAM-1 expression (arrows) is visible on the surface of spleen ECs. Bar=2 µm. (D) At 24 h after BMT, VCAM-1 is strongly expressed on the surface of bone mar-row ECs. Bar=1 µm. BV, blood vessel; E, erythrocyte; Ly, Lymphocyte; S, sinus.
351ADHESION MOLECULES IN HEMATOPOIETIC CELL HOMING
the spleen and bone marrow from 1 to 24 h after BMT is shown in Fig. 2.
VCAM-1 expression in spleen ECs was acutely de-creased at 1 h but significantly increased at 3 h after BMT. At 6 h, expression was slightly decreased, but VCAM-1 expression was significantly increased at 12 to 24 h after BMT. Especially at 24 h after BMT, VCAM-1 expression was observed at the sinus in the spleen. In bone marrow, VCAM-1 expression was acutely decreased at 1 to 6 h after BMT but was signifi-
cantly increased at 12 to 24 h. The expression of VCAM-1 in the spleen and bone marrow from 1 to 24 h after BMT is shown in Fig. 3.
In spleen ECs, MAdCAM-1 expression was less than VCAM-1 expression throughout the experimental pe-riod. Moreover, MAdCAM-1 expression was higher in bone marrow than in spleen from 6 to 24 h (Fig. 2), and VCAM-1 expression was higher in bone marrow than in spleen from 12 to 24 h after BMT (Fig. 3).
Fig. 2. MAdCAM-1 expression on spleen and bone marrow ECs. Changes in MAdCAM-1 expres-sion were analyzed in intact (INT) and BMT mice from 1 to 24 h after BMT. Columns and bars represent mean ± standard deviation (SD). Small letters (a, b) and capital letters (A, B) indicate significant differences among spleen and bone marrow, respectively (n=4, P<0.05).
Fig. 3. VCAM-1 expression on spleen and bone marrow ECs. Changes in VCAM-1 expression were analyzed in intact (INT) and BMT mice from 1 to 24 h after BMT. Columns and bars represent mean ± SD. Small letters (a, b, c) and capital letters (A, B, C) indicate significant differences among spleen and bone marrow, respectively (n=4, P<0.05).
352 T. TADA, ET AL.
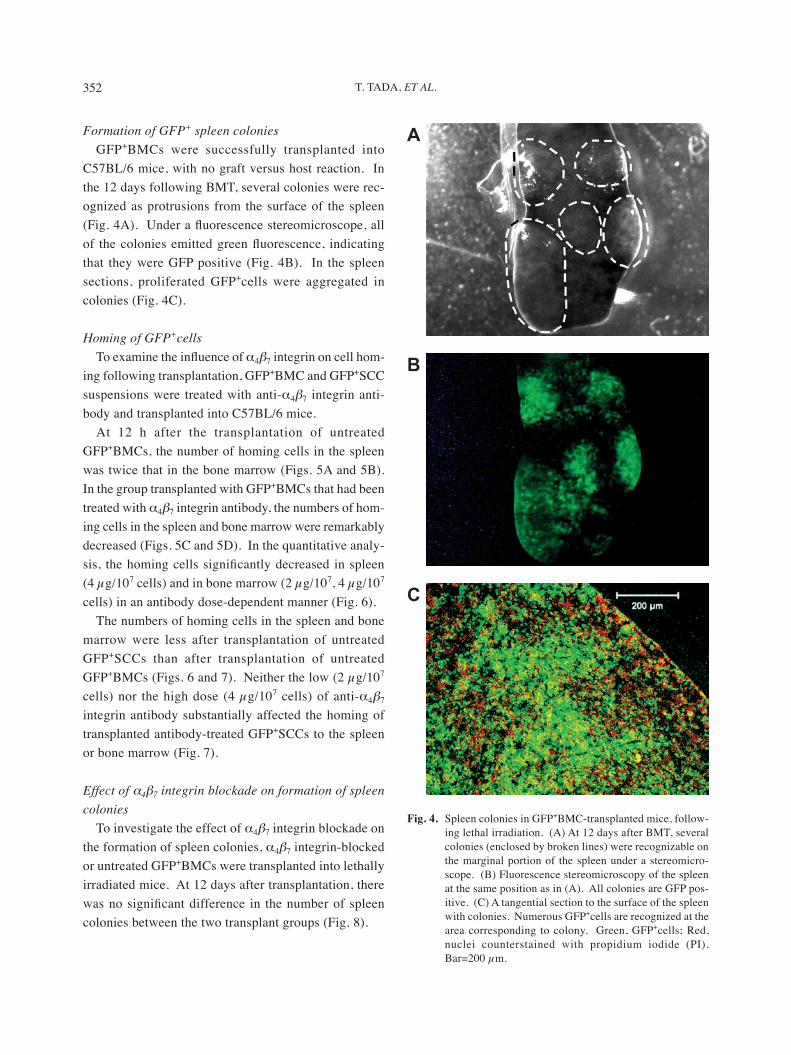
Formation of GFP+ spleen coloniesGFP+BMCs were successfully transplanted into
C57BL/6 mice, with no graft versus host reaction. In the 12 days following BMT, several colonies were rec-ognized as protrusions from the surface of the spleen (Fig. 4A). Under a fluorescence stereomicroscope, all of the colonies emitted green fluorescence, indicating that they were GFP positive (Fig. 4B). In the spleen sections, proliferated GFP+cells were aggregated in colonies (Fig. 4C).
Homing of GFP+cellsTo examine the influence of α4β7 integrin on cell hom-
ing following transplantation, GFP+BMC and GFP+SCC suspensions were treated with anti-α4β7 integrin anti-body and transplanted into C57BL/6 mice.
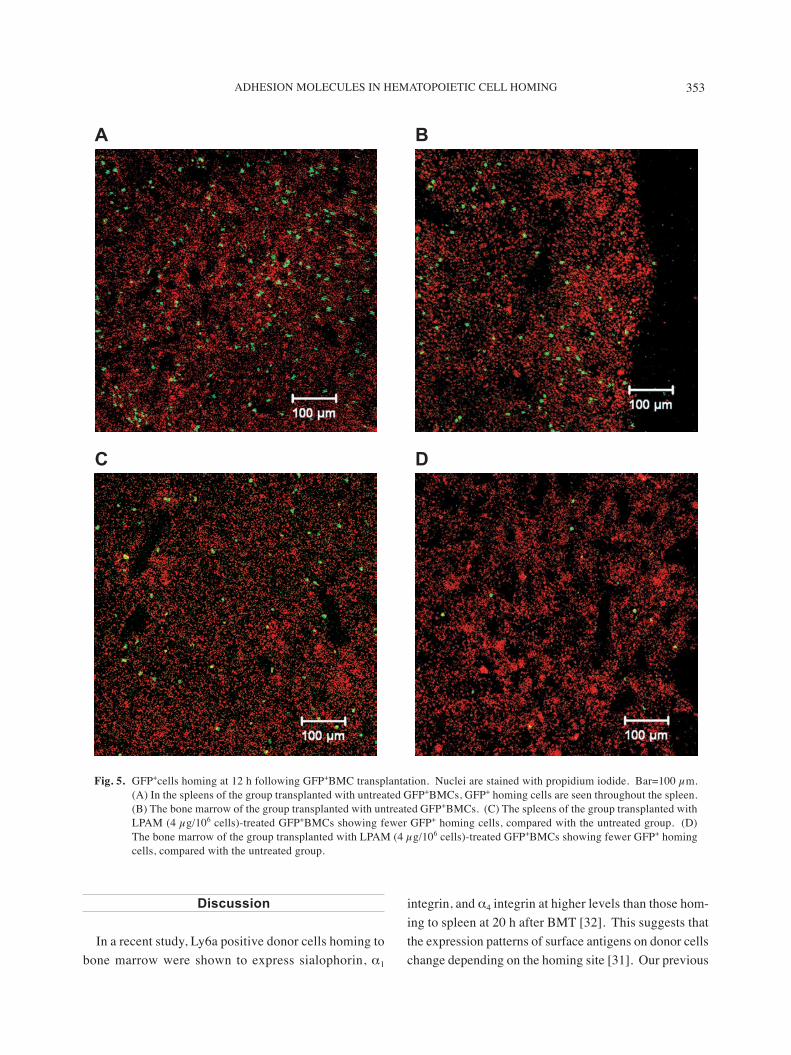
At 12 h after the transplantation of untreated GFP+BMCs, the number of homing cells in the spleen was twice that in the bone marrow (Figs. 5A and 5B). In the group transplanted with GFP+BMCs that had been treated with α4β7 integrin antibody, the numbers of hom-ing cells in the spleen and bone marrow were remarkably decreased (Figs. 5C and 5D). In the quantitative analy-sis, the homing cells significantly decreased in spleen (4 µg/107 cells) and in bone marrow (2 µg/107, 4 µg/107
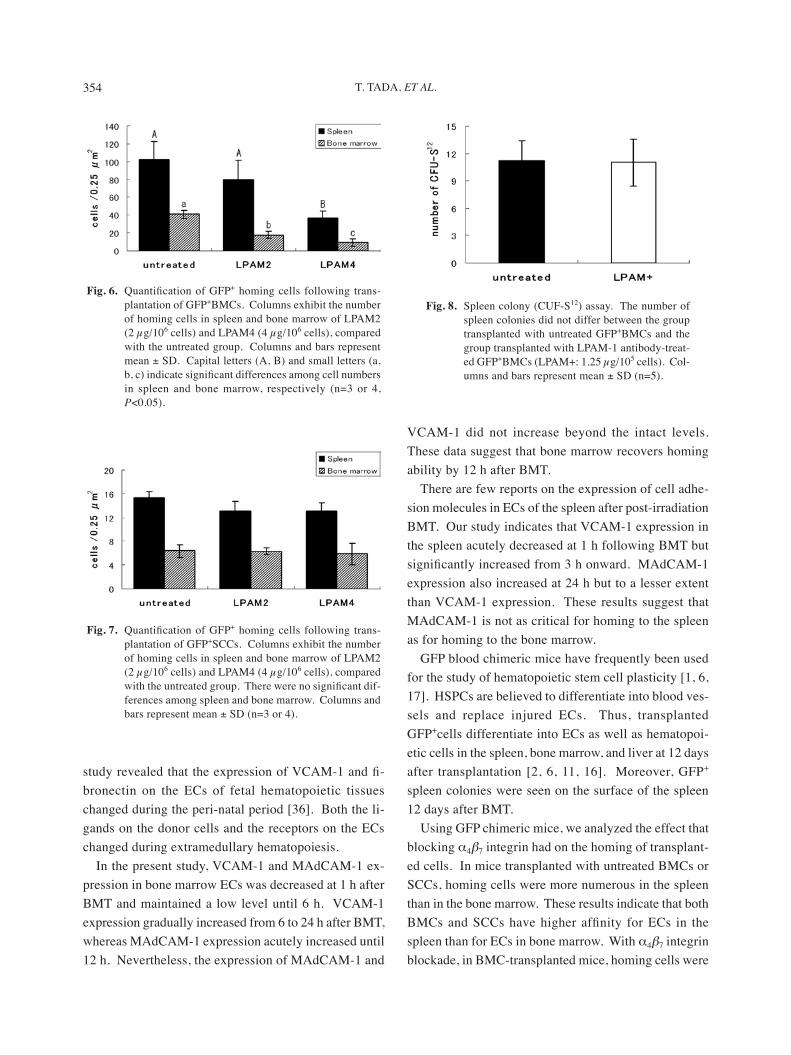
cells) in an antibody dose-dependent manner (Fig. 6).The numbers of homing cells in the spleen and bone
marrow were less after transplantation of untreated GFP+SCCs than after transplantation of untreated GFP+BMCs (Figs. 6 and 7). Neither the low (2 µg/107
cells) nor the high dose (4 µg/107 cells) of anti-α4β7 integrin antibody substantially affected the homing of transplanted antibody-treated GFP+SCCs to the spleen or bone marrow (Fig. 7).
Effect of α4β7 integrin blockade on formation of spleen colonies
To investigate the effect of α4β7 integrin blockade on the formation of spleen colonies, α4β7 integrin-blocked or untreated GFP+BMCs were transplanted into lethally irradiated mice. At 12 days after transplantation, there was no significant difference in the number of spleen colonies between the two transplant groups (Fig. 8).
A
B
C
Fig. 4. Spleen colonies in GFP+BMC-transplanted mice, follow-ing lethal irradiation. (A) At 12 days after BMT, several colonies (enclosed by broken lines) were recognizable on the marginal portion of the spleen under a stereomicro-scope. (B) Fluorescence stereomicroscopy of the spleen at the same position as in (A). All colonies are GFP pos-itive. (C) A tangential section to the surface of the spleen with colonies. Numerous GFP+cells are recognized at the area corresponding to colony. Green, GFP+cells; Red, nuclei counterstained with propidium iodide (PI). Bar=200 µm.
353ADHESION MOLECULES IN HEMATOPOIETIC CELL HOMING
Discussion
In a recent study, Ly6a positive donor cells homing to bone marrow were shown to express sialophorin, α1
integrin, and α4 integrin at higher levels than those hom-ing to spleen at 20 h after BMT [32]. This suggests that the expression patterns of surface antigens on donor cells change depending on the homing site [31]. Our previous
A B
C D
Fig. 5. GFP+cells homing at 12 h following GFP+BMC transplantation. Nuclei are stained with propidium iodide. Bar=100 µm. (A) In the spleens of the group transplanted with untreated GFP+BMCs, GFP+ homing cells are seen throughout the spleen. (B) The bone marrow of the group transplanted with untreated GFP+BMCs. (C) The spleens of the group transplanted with LPAM (4 µg/106 cells)-treated GFP+BMCs showing fewer GFP+ homing cells, compared with the untreated group. (D) The bone marrow of the group transplanted with LPAM (4 µg/106 cells)-treated GFP+BMCs showing fewer GFP+ homing cells, compared with the untreated group.
354 T. TADA, ET AL.
study revealed that the expression of VCAM-1 and fi-bronectin on the ECs of fetal hematopoietic tissues changed during the peri-natal period [36]. Both the li-gands on the donor cells and the receptors on the ECs changed during extramedullary hematopoiesis.
In the present study, VCAM-1 and MAdCAM-1 ex-pression in bone marrow ECs was decreased at 1 h after BMT and maintained a low level until 6 h. VCAM-1 expression gradually increased from 6 to 24 h after BMT, whereas MAdCAM-1 expression acutely increased until 12 h. Nevertheless, the expression of MAdCAM-1 and
VCAM-1 did not increase beyond the intact levels. These data suggest that bone marrow recovers homing ability by 12 h after BMT.
There are few reports on the expression of cell adhe-sion molecules in ECs of the spleen after post-irradiation BMT. Our study indicates that VCAM-1 expression in the spleen acutely decreased at 1 h following BMT but significantly increased from 3 h onward. MAdCAM-1 expression also increased at 24 h but to a lesser extent than VCAM-1 expression. These results suggest that MAdCAM-1 is not as critical for homing to the spleen as for homing to the bone marrow.
GFP blood chimeric mice have frequently been used for the study of hematopoietic stem cell plasticity [1, 6, 17]. HSPCs are believed to differentiate into blood ves-sels and replace injured ECs. Thus, transplanted GFP+cells differentiate into ECs as well as hematopoi-etic cells in the spleen, bone marrow, and liver at 12 days after transplantation [2, 6, 11, 16]. Moreover, GFP+
spleen colonies were seen on the surface of the spleen 12 days after BMT.
Using GFP chimeric mice, we analyzed the effect that blocking α4β7 integrin had on the homing of transplant-ed cells. In mice transplanted with untreated BMCs or SCCs, homing cells were more numerous in the spleen than in the bone marrow. These results indicate that both BMCs and SCCs have higher affinity for ECs in the spleen than for ECs in bone marrow. With α4β7 integrin blockade, in BMC-transplanted mice, homing cells were
Fig. 6. Quantification of GFP+ homing cells following trans-plantation of GFP+BMCs. Columns exhibit the number of homing cells in spleen and bone marrow of LPAM2 (2 µg/106 cells) and LPAM4 (4 µg/106 cells), compared with the untreated group. Columns and bars represent mean ± SD. Capital letters (A, B) and small letters (a, b, c) indicate significant differences among cell numbers in spleen and bone marrow, respectively (n=3 or 4, P<0.05).
Fig. 7. Quantification of GFP+ homing cells following trans-plantation of GFP+SCCs. Columns exhibit the number of homing cells in spleen and bone marrow of LPAM2 (2 µg/106 cells) and LPAM4 (4 µg/106 cells), compared with the untreated group. There were no significant dif-ferences among spleen and bone marrow. Columns and bars represent mean ± SD (n=3 or 4).
Fig. 8. Spleen colony (CUF-S12) assay. The number of spleen colonies did not differ between the group transplanted with untreated GFP+BMCs and the group transplanted with LPAM-1 antibody-treat-ed GFP+BMCs (LPAM+: 1.25 µg/105 cells). Col-umns and bars represent mean ± SD (n=5).

355ADHESION MOLECULES IN HEMATOPOIETIC CELL HOMING
markedly decreased in the spleen and bone marrow, whereas homing cells were unchanged in SCC-trans-planted mice. The MAdCAM-1/α4β7 integrin pathway regulates lymphocyte tethering and rolling on ECs of lymphoid tissues [4, 5, 34]. Katayama et al. reported that α4β7 integrin plays a key role in the initial steps of tethering and rolling but was not required for firm adhe-sion in the bone marrow microvasculature. Our data indicate that α4β7 integrin on the surface of HSPCs de-rived from bone marrow regulates tethering and rolling on ECs in the spleen and bone marrow. However, col-ony-forming unit-spleen homing is not regulated by the MAdCAM-1/α4β7 integrin pathway.
Treating bone marrow donor cells with anti-MAd-CAM-1 significantly reduced the number of cells homing to the bone marrow, whereas there was no significant reduction in cells homing to the spleen [23]. A recent study showed that colony-forming unit-spleen homing to bone marrow was unchanged after injection of an-ti-MAdCAM-1 in wild-type mice [38]. Our results sug-gest that α4β7 integrin is not important for the homing of SCCs to the bone marrow or spleen and that it does not function in forming spleen colonies.
Some reports have suggested the possibility that α4β7 integrin binds VCAM-1 because of the primary amino acid sequence similarity between VCAM-1 and MAd-CAM-1 [7, 9, 12]. This would mean that α4β7 integrin and α4β7 integrin may compensate between MAdCAM-1 and VCAM-1. Our data indicate that the regulation of homing to the spleen is mediated by a VCAM-1/α4β1 or α4β7 integrin pathway and that homing to bone marrow is mediated by a MAdCAM-1/α4β7 or α4β1 integrin, VCAM-1/α4β1 or α4β7 integrin pathway.
References
1. Aliotta, J.M., Passero, M., Meharg, J., Klinger, J., Dooner, M.S., Pimentel, J., and Quesenberry, P.J. 2005. Stem cells and pulmonary metamorphosis: new concepts in repair and regeneration. J. Cell Physiol. 204: 725–741.
2. Asahara, T., Takahashi, T., Masuda, H., Kalka, C., Chen, D., Iwaguro, H., Inai, Y., Silver, M., and Isner, J.M. 1999. VEGF contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells. EMBO J. 18: 3964–3972.
3. Asaumi, N., Omoto, E., Mahmut, N., Katayama, Y., Takeda, K., Shinagawa, K., and Harada, M. 2001. Very late antigen-5 and leukocyte function-associated antigen-1 are critical for
early stage hematopoietic progenitor cell homing. Ann. Hematol. 80: 387–392.
4. Berg, E.L., McEvoy, L.M., Berlin, C., Bargatze, R.F., and Butcher, E.C. 1993. L-selectin-mediated lymphocyte rolling on MAdCAM-1. Nature 366: 695–698.
5. Berlin, C., Berg, E.L., Briskin, M.J., Andrew, D.P., Kilshaw, P.J., Holzmann, B., Weissman, I.L., Hamann, A., and Butcher, E.C. 1993. Alpha 4 beta 7 integrin mediates lymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell 74: 185–195.
6. Brazelton, T.R., Rossi, F.M., Keshet, G.I., and Blau, H.M. 2000. From marrow to brain: expression of neuronal phenotypes in adult mice. Science 290: 1775–1779.
7. Briskin, M.J., McEvoy, L.M., and Butcher, E.C. 1993. MAdCAM-1 has homology to immunoglobulin and mucin-like adhesion receptors and to IgA1. Nature 363: 461–464.
8. Bullard, D.C., Kunkel, E.J., Kubo, H., Hicks, M.J., Lorenzo, I., Doyle, N.A., Doerschuk, C.M., Ley, K., and Beaudet, A.L. 1996. Infectious susceptibility and severe deficiency of leukocyte rolling and recruitment in E-selectin and P-selectin double mutant mice. J. Exp. Med. 183: 2329–2336.
9. Chan, B.M., Elices, M.J., Murphy, E., and Hemler, M.E. 1992. Adhesion to vascular cell adhesion molecule 1 and fibronectin. Comparison of alpha 4 beta 1 (VLA-4) and alpha 4 beta 7 on the human B cell line JY. J. Biol. Chem. 267: 8366–8370.
10. Cluitmans, F.H., Esendam, B.H., Veenhof, W.F., Landegent, J.E., Willemze, R., and Falkenburg, J.H. 1997. The role of cytokines and hematopoietic growth factors in the autocrine/paracrine regulation of inducible hematopoiesis. Ann. Hematol. 75: 27–31.
11. Crosby, J.R., Kaminski, W.E., Schatteman, G., Martin, P.J., Raines, E.W., Seifert, R.A., and Bowen-Pope, D.F. 2000. Endothelial cells of hematopoietic origin make a significant contribution to adult blood vessel formation. Circ. Res. 87: 728–730.
12. Day, E.S., Osborn, L., and Whitty, A. 2002. Effect of divalent cations on the affinity and selectivity of alpha4 integrins towards the integrin ligands vascular cell adhesion molecule-1 and mucosal addressin cell adhesion molecule-1: Ca2+ activation of integrin alpha4beta1 confers a distinct ligand specificity. Cell Commun. Adhes. 9: 205–219.
13. Dorshkind, K. 1990. Regulation of hemopoiesis by bone marrow stromal cells and their products. Annu. Rev. Immunol. 8: 111–137.
14. Frenette, P.S., Mayadas, T.N., Rayburn, H., Hynes, R.O., and Wagner, D.D. 1996. Susceptibility to infection and altered hematopoiesis in mice deficient in both P- and E-selectins. Cell 84: 563–574.
15. Frenette, P.S., Subbarao, S., Mazo, I.B., von Andrian, U.H., and Wagner, D.D. 1998. Endothelial selectins and vascular cell adhesion molecule-1 promote hematopoietic progenitor homing to bone marrow. Proc. Natl. Acad. Sci. U.S.A. 95: 14423–14428.
16. Grant, M.B., May, W.S., Caballero, S., Brown, G.A., Guthrie, S.M., Mames, R.N., Byrne, B.J., Vaught, T., Spoerri, P.E.,

356 T. TADA, ET AL.
Peck, A.B., and Scott, E.W. 2002. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nature Med. 8: 607–612.
17. Guthrie, S.M., Curtis, L.M., Mames, R.N., Simon, G.G., Grant, M.B., and Scott, E.W. 2005. The nitric oxide pathway modulates hemangioblast activity of adult hematopoietic stem cells. Blood 105: 1916–1922.
18. Hamann, A., Andrew, D.P., Jablonski-Westrich, D., Holzmann, B., and Butcher, E.C. 1994. Role of alpha 4-integrins in lymphocyte homing to mucosal tissues in vivo. J. Immunol. 152: 3282–3293.
19. Holzmann, B., McIntyre, B.W., and Weissman, I.L. 1989. Identification of a murine Peyer’s patch—specific lymphocyte homing receptor as an integrin molecule with an alpha chain homologous to human VLA-4 alpha. Cell 56: 37–46.
20. Hu, M.C., Crowe, D.T., Weissman, I.L., and Holzmann, B. 1992. Cloning and expression of mouse integrin beta p(beta 7): a functional role in Peyer’s patch-specific lymphocyte homing. Proc. Natl. Acad. Sci. U.S.A. 89: 8254–8258.
21. Ikawa, M., Yamada, S., Nakanishi, T., and Okabe, M. 1998. ‘Green mice’ and their potential usage in biological research. FEBS Lett. 430: 83–87.
22. Katayama, Y., Hidalgo, A., Furie, B.C., Vestweber, D., Furie, B., and Frenette, P.S. 2003. PSGL-1 participates in E-selectin-mediated progenitor homing to bone marrow: evidence for cooperation between E-selectin ligands and alpha4 integrin. Blood 102: 2060–2067.
23. Katayama, Y., Hidalgo, A., Peired, A., and Frenette, P.S. 2004. Integrin alpha4beta7 and its counterreceptor MAdCAM-1 contribute to hematopoietic progenitor recruitment into bone marrow following transplantation. Blood 104: 2020–2026.
24. Kraal, G., Schornagel, K., Streeter, P.R., Holzmann, B., and Butcher, E.C. 1995. Expression of the mucosal vascular addressin, MAdCAM-1, on sinus-lining cells in the spleen. Am. J. Pathol. 147: 763–771.
25. Labow, M.A., Norton, C.R., Rumberger, J.M., Lombard-Gillooly, K.M., Shuster, D.J., Hubbard, J., Bertko, R., Knaack, P.A., Terry, R.W., Harbison, M.L., Kontgen, F., Stewart, C.L., Mclntyre, K.W., Will, P.C., Burns, D.K., and Wolitzky, B.A. 1994. Characterization of E-selectin-deficient mice: demonstration of overlapping function of the endothelial selectins. Immunity 1: 709–720.
26. Masinovsky, B., Urdal, D., and Gallatin, W.M. 1990. IL-4 acts synergistically with IL-1 beta to promote lymphocyte adhesion to microvascular endothelium by induction of vascular cell adhesion molecule-1. J. Immunol. 145: 2886–2895.
27. Nilsson, S.K., Simmons, P.J., and Bertoncello, I. 2006. Hemopoietic stem cell engraftment. Exp. Hematol. 34:
123–129.28. Okabe, M., Ikawa, M., Kominami, K., Nakanishi, T., and
Nishimune, Y. 1997. ‘Green mice’ as a source of ubiquitous green cells. FEBS Lett. 407: 313–319.
29. Oostendorp, R.A. and Dormer, P. 1997. VLA-4-mediated interactions between normal human hematopoietic progenitors and stromal cells. Leuk. Lymphoma. 24: 423–435.
30. Papayannopoulou, T., Craddock, C., Nakamoto, B., Priestley, G.V., and Wolf, N.S. 1995. The VLA4/VCAM-1 adhesion pathway defines contrasting mechanisms of lodgement of transplanted murine hemopoietic progenitors between bone marrow and spleen. Proc. Natl. Acad. Sci. U.S.A. 92: 9647–9651.
31. Plett, P.A., Frankovitz, S.M., and Orschell-Traycoff, C.M. 2002. In vivo trafficking, cell cycle activity, and engraftment potential of phenotypically defined primitive hematopoietic cells after transplantation into irradiated or nonirradiated recipients. Blood 100: 3545–3552.
32. Plett, P.A., Frankovitz, S.M., and Orschell, C.M. 2003. Distribution of marrow repopulating cells between bone marrow and spleen early after transplantation. Blood 102: 2285–2291.
33. Rosendaal, M., Gregan, A., and Green, C.R. 1991. Direct cell-cell communication in the blood-forming system. Tissue Cell 23: 457–470.
34. Ruegg, C., Postigo, A.A., Sikorski, E.E., Butcher, E.C., Pytela, R., and Erle, D.J. 1992. Role of integrin alpha 4 beta 7/alpha 4 beta P in lymphocyte adherence to fibronectin and VCAM-1 and in homotypic cell clustering. J. Cell Biol. 117: 179–189.
35. Simmons, P.J., Masinovsky, B., Longenecker, B.M., Berenson, R., Torok-Storb, B., and Gallatin, W.M. 1992. Vascular cell adhesion molecule-1 expressed by bone marrow stromal cells mediates the binding of hematopoietic progenitor cells. Blood 80: 388–395.
36. Tada, T., Widayati, D.T., and Fukuta, K. 2006. Morphological study of the transition of haematopoietic sites in the developing mouse during the peri-natal period. Anat. Histol. Embryol. 35: 235–240.
37. Verfaillie, C.M., McCarthy, J.B., and McGlave, P.B. 1991. Differentiation of primitive human multipotent hematopoietic progenitors into single lineage clonogenic progenitors is accompanied by alterations in their interaction with fibronectin. J. Exp. Med. 174: 693–703.
38. Vermeulen, M., Le Pesteur, F., Gagnerault, M.C., Mary, J.Y., Sainteny, F., and Lepault, F. 1998. Role of adhesion molecules in the homing and mobilization of murine hematopoietic stem and progenitor cells. Blood 92: 894–900.
Copyright © 2022 FDOKUMEN




















