
“Rol de GCSF-CHR19; CXCL8a y CXCL8b en la migración de ...
46
Universidad Andrés Bello Facultad de Ciencias Biológicas Escuela de Ingeniería en Biotecnología “Rol de GCSF-CHR19; CXCL8a y CXCL8b en la migración de neutrófilos a través de los vasos sanguíneos en pez cebra (Danio rerio)” Tesis presentada como parte de los requisitos para optar al Grado de Magister en Biotecnología Profesor patrocinante: Dra. Carmen Gloria Feijoo García Facultad de Ciencias Biológicas Universidad Andrés Bello Constanza Del Pilar Zúñiga Traslaviña Santiago, Chile Mayo, 2017
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of “Rol de GCSF-CHR19; CXCL8a y CXCL8b en la migración de ...

Universidad Andrés Bello
Facultad de Ciencias Biológicas
Escuela de Ingeniería en Biotecnología
“Rol de GCSF-CHR19; CXCL8a y CXCL8b en la migración de neutrófilos a
través de los vasos sanguíneos en pez cebra (Danio rerio)”
Tesis presentada como parte de los requisitos para optar al Grado de
Magister en Biotecnología
Profesor patrocinante: Dra. Carmen Gloria Feijoo García
Facultad de Ciencias Biológicas
Universidad Andrés Bello
Constanza Del Pilar Zúñiga Traslaviña
Santiago, Chile Mayo, 2017

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
1
Dedicatoria
A mis padres, Lincoyan Zúñiga y Christhal Traslaviña, por ser el pilar fundamental en todo lo que soy, en toda mi
educación, tanto académica, como de la vida, por su incondicional apoyo mantenido a través del tiempo. Todo este trabajo ha
sido posible gracias a ellos.
A mis hermanos, Christhal Zúñiga y Javier Zúñiga, por ser ejemplos de hermanos mayores y de quienes
aprendí aciertos y de momentos difíciles.
A mis abuelos, Guido Traslaviña, Mireya Farías y Raquel Urbina, a mis tíos y primos que me apoyaron directa o
indirectamente en la elaboración de esta tesis.
A Ángel Arias, mi compañero de vida, quien comprendió, comprendió y apoyó en este largo camino de estudio y
trabajo.
A mis amigas que me apoyaron en mi formación como persona, que me apoyaron y comprendieron hasta
ahora, seguimos siendo amigas: Camila Paul, Andrea Trullen y Constanza Vera.
Al laboratorio de inmunología en peces, quienes me apoyaron en mi formación profesional, a la Dra. Carmen Feijoo
quien me guio en todo este trabajo, y a mis compañeros de laboratorio especialmente a Karina Bravo con quien trabajamos a la
par.
¡Gracias por todo!

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
2
AGRADECIMIENTOS
Este trabajo fue financiado por el Fondo Nacional de Desarrollo Científico y Tecnológico
1171199 y 1150816, Dirección de Investigación UNAB DI-483-14/R, y Fondo de
Financiamiento de Centros de Investigación en Áreas Prioritarias 15110027.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
3
Índice
Abstract ....................................................................................................................... 5
Resumen ..................................................................................................................... 7
Introducción ................................................................................................................ 9
Marco teórico ........................................................................................................... 9
Sistema inmune ...................................................................................................... 9
Neutrófilos y respuesta inflamatoria ...................................................................... 10
Regulación de la migración de neutrófilos durante un proceso inflamatorio ......... 10
Regulación de la migración de neutrófilos en pez cebra ...................................... 14
Hipótesis ................................................................................................................ 16
Objetivo General ................................................................................................... 16
Objetivos específicos ........................................................................................... 17
Materiales y métodos ............................................................................................... 18
Mantención de peces ............................................................................................ 18
Daño estéril ............................................................................................................ 18
Expresión génica .................................................................................................. 19
Ensayos falta de función génica ......................................................................... 21
Cuantificación de neutrófilos ............................................................................... 23
Estadística ............................................................................................................. 23
Resultados ................................................................................................................ 24
Niveles transcripcionales de diferentes genes durante un proceso inflamatorio
luego de un daño severo ....................................................................................... 24
Niveles transcripcionales de diferentes genes durante un proceso inflamatorio
luego de un daño severo en ausencia de Gcsf-chr19 ........................................... 26
Niveles transcripcionales de diferentes genes en un proceso inflamatorio luego
de un daño leve ..................................................................................................... 28

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
4
Determinación del número de neutrófilos presentes en circulación y en zona
afectada luego de un daño severo ........................................................................ 30
Determinación del número de neutrófilos presentes en circulación y en zona
afectada luego de un daño leve ............................................................................ 32
Discusión .................................................................................................................. 34

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
5
Abstract Neutrophils migration is one of the characteristics that distinguish an
inflammatory process, and its deregulation occurs in several diseases with immune
component.
This thesis focused on studying the regulation, at a molecular level, of the
migration of neutrophils to blood vessels to reach the site of inflammation caused by
sterile damage. In mammals, specifically in mice, this process is regulated by the
CXCL1/ 2 cytokine pathway and its receptor CXCR2, and in turn are controlled via G-
CSF signaling. On the other hand, in humans, the main ligand of the CXCR2 receptor is
CXCL8, but it is not known if this signaling pathway regulates the migration of
neutrophils into the bloodstream. This deference between the murine model and human
prevents the development of studies of the mentioned problem in mice, so it is proposed
the use of zebrafish as an alternative study model. The zebrafish presents in its genome
two paralogs for Cxcl8, Cxcl8a and Cxcl8b. Available evidence indicates that Cxcl8a
regulates neutrophil migration during an inflammatory process and that Cxcl8b
increases its transcriptional levels in zebrafish. For its part, Gcsf-chr19 performs a
function similar to that described for its orthologous G-CSF in mammals.
Based on the above problem, the following hypothesis is proposed: "Cxcl8a
regulates the entry of neutrophils into the bloodstream, in response to mechanical
damage in zebrafish. This chemokine is regulated by Gsf-Chr19 ". Thus, the aim was to
characterize the function of Cxcl8a and Cxcl8b during the process of migration of
neutrophils to blood vessels triggered by sterile zebrafish damage and their relationship
to Gcsf-Chr19.
Using double transgenic larvae, Tg (BACmpx: GFP) i114 and Tg (fli1: EGFP) y1,
and two damage models with different intensities (severe and mild), which allowed
carried out in vivo monitoring of the migration of neutrophils in Cxcl8a and Cxcl8b lack of
function. In addition, measuring of the transcriptional levels of cxcl8a and cxcl8b, and
other genes involved in an inflammatory process such as: il-1b, il-6, cxcr1, cxcr2, sdf-1,
cxcr4, gcsf-chr12, gcsf-chr19 and myd88, in both types of damages and in severe
damage with lack of function of Gcsf-chr19.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
6
The results indicate that, in an inflammatory process triggered by severe sterile
damage, Gcsf-Chr19 represses the expression of sdf-1 and induces the expression of
Cxcl8b and thus favors the entry of neutrophils into the blood vessels. For its part,
Cxcl8a regulates the migration of neutrophils through the interstitial tissue.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
7
Resumen La migración de neutrófilos es una de las características que distinguen a un
proceso inflamatorio, y su desregulación se presenta en diversas enfermedades con
componente inmune.
Esta tesis se enfocó en estudiar la regulación, a nivel molecular, de la migración
de los neutrófilos a los vasos sanguíneos para llegar al lugar de inflamación causada
por un daño estéril. En mamíferos, específicamente en ratones, este proceso es
regulado por la vía de las citoquinas CXCL1/2 y su receptor CXCR2 y que a su vez
están controlados por vía de señalización G-CSF. Por su parte, en humanos, el
principal ligando del receptor CXCR2 es CXCL8, pero no se sabe si esta vía de
señalización regula la migración de neutrófilos a la circulación sanguínea. Esta
diferencia entre el modelo murino y humanos impide el desarrollo de estudios de la
mencionada problemática en ratones, por lo que se propone el uso de pez cebra como
modelo de estudio alternativo. El pez cebra presenta en su genoma dos parálogos para
Cxcl8, Cxcl8a y Cxcl8b. Antecedentes disponibles indican que Cxcl8a regula la
migración de neutrófilos durante un proceso inflamatorio y que Cxcl8b aumenta sus
niveles transcripcionales en pez cebra. Por su parte, Gcsf-chr19 ejerce una función
similar a la descrita para su ortólogo G-CSF en mamíferos.
Basado en la problemática antes descrita que se plantea la siguiente hipótesis:
“Cxcl8a regula el ingreso de neutrófilos al torrente sanguíneo, en respuesta a un daño
mecánico en pez cebra. Esta quimioquina a su vez es regulada por Gcsf-Chr19”. Así, el
objetivo general fue caracterizar la función de Cxcl8a y Cxcl8b durante el proceso de
migración de neutrófilos a los vasos sanguíneo gatillado por daño estéril en pez cebra y
su relación con Gcsf-Chr19. Utilizando larvas doble transgénicas Tg (BACmpx:GFP) i114 X Tg (fli1:EGFP)y1, se
llevaron a cabo dos modelos de daño con diferente intensidad (severo y leve), y se
monitoreo in vivo la migración de neutrófilos en condiciones de falta de función de
Cxcl8a y Cxcl8b. Además, se determinaron los niveles transcripcionales de cxcl8a y
cxcl8b, y otros genes involucrados en un proceso inflamatorio tales como: il-1b, il-6,
cxcr1, cxcr2, sdf-1, cxcr4, gcsf-chr12, gcsf-chr19 y myd88, en ambos tipos de daños y
en daño severo en ausencia de Gcsf-chr19.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
8
Los resultados obtenidos, indican que, en un proceso inflamatorio gatillado por un
daño estéril severo, Gcsf-Chr19 reprime la expresión de sdf-1 e induce la expresion de
Cxcl8b y con ello favorece la entrada de neutrofilos a los vasos sanguineos. Por su
parte, Cxcl8a regula la migracion de neutrofilos a traves del tejido intersticial.
.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
9
Introducción
En la actualidad existen diversas enfermedades relacionadas al sistema inmune
que afectan considerablemente la calidad de vida de las personas. En su mayoría,
éstas se deben a alteraciones en los distintos procesos que ocurren durante una
respuesta inmune (Licastro et al., 2005). En particular, la desregulación de la función de
los neutrófilos, células pertenecientes al sistema inmune innato, pueden causar
enfermedades tales como neutropenia, artritis reumatoide, enfermedad inflamatoria del
intestino y enfermedad obstructiva crónica de pulmón (Eyles et al., 2006). En
condiciones de homeostasis, los neutrófilos se encuentran retenidos en el tejido
hematopoyético, pero al generarse un estímulo inflamatorio se favorece su migración al
área afectada. Dicho proceso de migración consta de varias etapas; salida del tejido
hematopoyético, entrada a los vasos sanguíneos y circulación, salida de los vasos
sanguíneos y migración final al daño. En cada uno de estas etapas participan diferentes
genes regulando finamente el proceso.
Esta tesis se focaliza en el estudio de la regulación de la migración de neutrófilos
hacia los vasos sanguíneos y la acción de las citoquinas Cxcl8a, Cxcl8b y Gcsf-chr19
en este proceso en el modelo de estudio pez cebra (Danio rerio).
Marco teórico
Sistema inmune
El sistema inmune tiene como función proteger al organismo de diferentes
amenazas (patógenos, daño tisular, etc.) manteniendo las condiciones de homeostasis
durante su desarrollo y crecimiento (Klosterman, 2009). Éste se divide clásicamente en
inmunidad innata e inmunidad adaptativa (Magnadóttir, 2010). De estos dos tipos de
inmunidades, la presente tesis tendrá enfoque en la inmunidad innata.
El sistema inmune innato corresponde a la primera línea de defensa de los
organismos, consiste en un mecanismo celular y bioquímico que se encuentra listo para
responder rápidamente frente a una infección o daño (Abbas et al., 2015). En algunos
organismos, como los peces, el sistema inmune innato tiene mayor relevancia que el
sistema inmune adaptativo (Magnadóttir, 2006). Por ejemplo, en el caso de pez cebra
permite la sobrevivencia durante las primeras semanas de desarrollo, dado que el

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
10
sistema inmune adaptativo se encuentra activo a partir de la tercera semana posterior a
la fertilización (Kanther & Rawls, 2010).
La principal respuesta del sistema inmune innato es la respuesta inflamatoria, la
cual corresponde al proceso de migración y reclutamiento de células inmunes a la zona
afectada. Dicha respuesta puede ser activada por una infección o por un daño tisular
(Abbas et al., 2015).
Neutrófilos y respuesta inflamatoria
Dentro de los tipos celulares que participan en el proceso inflamatorio podemos
encontrar los neutrófilos (White et al., 1998). Éstos son células inflamatorias móviles, y
tienen la habilidad de fagocitar y destruir microorganismos como bacterias, hongos y
virus (Vento & Cainelli., 2003). La activación y consecuente migración de los neutrófilos
se considera un indicador de que está ocurriendo un proceso inflamatorio en el
organismo (Issekutz & Movat, 1980). Durante una infección, los neutrófilos migran a
esta zona para eliminar el agente infeccioso utilizando enzimas proteolíticas y proteínas
anti-microbiales (Palic et al., 2007). Por otro lado, se denomina inflamación estéril
cuando se genera un daño mecánico en un tejido. En Ambas situaciones se activan
vías de señalización de diferentes citoquinas que gatillan la migración de neutrófilos al
lugar afectado (Chen & Nuñez, 2010).
Regulación de la migración de neutrófilos durante un proceso inflamatorio
A pesar de que la entrada de neutrófilos a la circulación sanguínea es una etapa
crucial en el proceso inflamatorio, su regulación en humanos no se encuentra
completamente descrita. En mamíferos, en condiciones de homeostasis los neutrófilos
se encuentran unidos a los osteoblastos presentes en la médula ósea. Esta unión es
regulada por la interacción de interleuquina 12, también conocida como Factor 1
Derivado de Células Estromales (SDF-1), y su receptor de quimioquina C-X-C tipo 4
(CXCR4), además de moléculas de adhesión del tipo integrinas (Figura 1) (Teixidó et
al., 1992). Por otro lado, luego de un estímulo nocivo, los neutrófilos son liberados
desde la medula ósea y atraídos a la circulación sanguínea con el fin de favorecer su
rápida llegada al lugar afectado (Eash et al., 2010). Así, es necesario entonces que se

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
11
produzca una inhibición de la señal pro-retención de neutrófilos y un aumento de la
señal pro-migración para permitir la salida de neutrófilos desde la medula ósea. Ambos
procesos son regulados por la citoquina Factor Estimulante de Colonias de Granulocitos
(G-CSF por sus siglas en ingles), junto a su receptor G-CSFR (Layton & Hall, 2006;
Taga & Kishimoto, 1997). En condiciones de homeostasis, G-CSF se encuentra en
niveles muy bajos, pero durante un proceso inflamatorio sus niveles de expresión
aumentan drásticamente en diferentes tipos celulares, incluyendo monocitos,
fibroblastos y células endoteliales (Anderlini et al., 1996; Demetri & Griffin, 1991).
Figura 1. Modelo de ubicación de neutrófilos en homeostasis. Los neutrófilos son
retenidos en el tejido hematopoyético debido a la unión del ligando SDF-1 al receptor
CXCR4.
En el modelo murino, al aumentar los niveles de G-CSF se induce la disminución
de los niveles de SDF-1 en la médula ósea, además de reducir la expresión de CXCR4
en la superficie de las células mieloides permitiendo la salida de neutrófilos desde la
médula ósea hacia el torrente sanguíneo (Kyung et al., 2006; Nagase et al., 2002;
Semerad et al., 2002). Por otro lado, G-CSF también actúa a nivel de células

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
12
endoteliales para promover el aumento en la expresión de la interleuquina 1 (CXCL1) e
interleuquina 2 (CXCL2), además de aumentar la expresión del receptor de quimioquina
C-X-C tipo 2 (CXCR2), en neutrófilos. Dicha vía de señalización es la encargada de
dirigir la migración de los neutrófilos a los vasos sanguíneos (Nagase et al., 2002)
(Figura 2).
Figura 2. Modelo de Inflamación por daño. Los niveles de G-CSF aumentan
disminuyendo los niveles de SDF-1 y CXCR4 generando la liberación de los neutrófilos
y posterior entrada a los vasos sanguíneos, para finalmente dirigirse al lugar del daño
por señales quimioatractantes de CXCL1/2.
En humanos, los genes que participan en la regulación de la migración de
neutrófilos difieren de lo descrito en roedores. La diferencia radica en el ligando de
CXCR2 en humano, el cual es interleuquina 8 (CXCL8), y no CXCL1/CXCL2 (Van et al.,
1989). Además, se ha reportado que existe afinidad entre CXCL8 y el receptor de
quimioquina C-X-C tipo 1 (CXCR1) (Raghuwanshi et al., 2012).
Diversos antecedentes indican que al aumentar de manera exógena los niveles
de G-CSF en la sangre, se gatilla un aumento de los niveles endógenos de CXCL8 en

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
13
la sangre en conjunto con una mayor cantidad de células inmunes en circulación,
particularmente neutrófilos, sugiriendo que ésta quimioquina está regulada directa o
indirectamente por GCSF. Además, en este mismo estudio se muestra que al generar
un aumento de CXCL8 de manera exógena, existe una mayor movilización de
neutrófilos en la sangre (Watanabe et al., 1999). Por otro lado, en condiciones de falta
de función de CXCR2 se observa una disminución en la migración y quimiotaxis de los
neutrófilos al lugar del insulto (White et al., 1998).
Debido a las diferencias moleculares antes mencionadas en el proceso
inflamatorio entre ratones y humano, es que proponemos el uso pez cebra como
modelo alternativo y complementario para estudiar en detalle dicho proceso, ya que,
diversos antecedentes sugieren que al menos a nivel celular, el proceso inflamatorio en
pez cebra esta conservado con respecto a mamíferos (Deng Q et al., 2013; Galdames
et al., 2014).
Pez cebra (Danio rerio) es un pez teleósteo que se utiliza ampliamente en la
investigación biomédica debido a las grandes ventajas que presenta frente a otros
modelos de estudios, entre estas ventajas podemos encontrar que posee un alto grado
de similitud relacionado a los mecanismos moleculares como celulares con los
humanos, además de que el desarrollo embrionario ocurre de forma externa y rápida, lo
que permite que la manipulación se puede realizar desde el momento de la fecundación
y donde a las 24 horas post fertilización (hpf) se tiene un organismo funcional, también
son ópticamente trasparente lo cual facilita la observación de órganos internos, otra
ventaja es que se encuentra completamente secuenciado, y gracias a esto se pueden
utilizar técnicas de biología molecular como la generación de transgénico, knock out,
knock down, sobre expresiones, etc., finalmente posee un bajo costo de manejo y
mantención (Kari et al., 2007).
Proceso inflamatorio en pez cebra
Desde hace ya algunos años que el pez cebra se utiliza para estudiar la
respuesta inmune, sobre todo la respuesta innata (Novoa & Figueras, 2012). Esto,
debido a las ventajas comparativas que este modelo presenta, como la transparencia
de tejidos durante etapas embrionarias y larvales, además de la existencia de líneas

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
14
transgénicas donde células inmunes están fluorescentemente marcadas. Lo anterior
hace posible monitorear in vivo en un organismo completo la participación de
determinadas células en el proceso inflamatorio.
Regulación de la migración de neutrófilos en pez cebra
Se sabe que los neutrófilos permanecen retenidos en el tejido caudal
hematopoyético (TCH) debido a la señal pro-retención que es llevada a cabo por SDF-1
y CXCR4 (Walters et al., 2010). Posterior a un insulto se genera un incremento en los
niveles transcripcionales de Gcsf-chr19, el ortólogo de G-CSF, lo que favorece la
migración de neutrófilos desde el TCH hacia los vasos sanguíneos para posteriormente
migrar hacia la zona afectada (Galdames et al., 2014). Además, se ha demostrado que
concomitante con el aumento de Gcsf-chr19 ocurre un incremento significativo de la
expresión del transcrito de cxcl8a (Galdames et al., 2014), sugiriendo que Cxcl8a
estaría participando en conjunto con Gcsf-chr19 en el proceso inflamatorio generado
por daño estéril en pez cebra.
Se ha demostrado que Cxcl8a ejerce una función atractante sobre neutrófilos,
regulando su migración al tejido afectado (Oehlers et al., 2010; Sarris et al., 2012; Deng
et al. 2013). Por su parte, se ha reportado que Cxcl8b participa en el proceso de
migración con un aumento de sus niveles de expresión durante un daño mecánico (De
Oliveira et al., 2013).
En resumen, se sabe que en ratones la vía de señalización encargada de dirigir
la migración de neutrófilos a los vasos sanguíneos está compuesta por las citoquinas
CXCL1/2 y su receptor CXCR2, las que a su vez están reguladas por GCS-F. Esta
citoquina también disminuye los niveles de CXCL12 (Figura 3A). En cambio, en
humanos, esta función es llevada a cabo por CXCL8 y CXCR2, y es también regulada
por la vía de GCS-F (Figura 3B). En pez cebra, Gcsf-Chr19 regula la migración de
neutrófilos por los vasos sanguíneos, pero no se sabe si regula los niveles
transcripcionales de Sdf-1, Cxcl8a, Cxcl8b y Cxcr2 (Figura 3C).

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
15
Figura 3. Diagrama resumen de la problemática. (A) En ratones, la vía de G-CSF/G-
CSFr regula negativamente la unión de CXCL12/CXCR4, y positivamente los niveles de
CXCL1/2/CXCR2. (B) En humanos, la vía de G-CSF/G-CSFr regula negativamente la
unión de CXCL12/CXCR4, y positivamente los niveles de CXCL8/CXCR2. (C) En pez
cebra, si bien la vía de Gcsf-Chr19/Gcsfr regula la migración de neutrófilos a los vasos
sanguíneos, no se sabe si está controlando a Sdf1/Cxcr4b, y/o a Cxcl8a/b/Cxcr2.
Es importante mencionar que en pez cebra existen dos parálogos de CXCL8,
Cxcl8a (EH557944) y Cxcl8b (HF674400) ubicados en cromosoma 1 y 7
respectivamente, Cxcl8a posee una identidad del 94% con humano (NM_000584.3),
mientras que Cxc8b sólo de un 79%. En lo que respecta a GCS-F, este también se
encuentra en forma paráloga como Gcsf-Chr12 (FM174388.1) y Gcsf-Chr19
(EH451519.1), los cuales posee una identidad de 89% y 88% con respecto a humano
(NC_000017.11) respectivamente.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
16
En casi la totalidad de los trabajos disponibles en pez cebra, incluidos los
mencionados anteriormente, la migración de neutrófilos durante un proceso inflamatorio
ha sido estudiada como un proceso único, sin etapas, por lo que no se tiene mayores
antecedentes de cómo o cuando ocurre la migración de neutrófilos hacia los vasos
sanguíneos y que genes la regulan.
Con el fin de desarrollar nuestros objetivos, se llevaron a cabo dos tipos de
daños estériles, denominados severo y leve. El daño severo corresponde a una
amputación de aleta caudal incluyendo tejido muscular, mientras que el daño leve sólo
se realiza amputación del tejido de la aleta. Esta diferencia en la intensidad de los
daños gatilla procesos inflamatorios distintos. Entre las diferencias más importantes
encontramos que en el modelo de daño severo se gatilla migración de neutrófilos a la
zona afectada tanto a través de los vasos sanguíneos como por el tejido tisular, mientas
que en un daño leve la migración es exclusivamente por matriz extracelular (Galdames
et al.,2014). Así, la comparación de lo que ocurre a nivel molecular y celular entre estos
dos daños nos permitirá contestar nuestra pregunta: ¿Participa Cxcl8a o Cxcl8b o
ambas en la regulación de la migración de neutrófilos a los vasos sanguíneos gatillado
por un daño estéril, y son estas a su vez reguladas por Gcsf-Chr19?
Hipótesis “Cxcl8a regula el ingreso de neutrófilos al torrente sanguíneo, en respuesta a un
daño mecánico en pez cebra. Esta quimioquina a su vez es regulada por Gcsf-Chr19”.
Objetivo General Caracterizar la función de cxcl8a y cxcl8b durante el proceso de migración de
neutrófilos a los vasos sanguíneo gatillado por daño estéril en pez cebra y su relación
con Gcsf-Chr19.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
17
Objetivos específicos
1. Determinar los niveles transcripcionales de cxcl8a y cxcl8b, y otros genes
involucrados en un proceso inflamatorio tales como: il-1b, il-6, cxcr1, cxcr2,
cxcr4b, gcsf-chr12, gcsf-chr19 y myd88 después de un daño estéril severo en
presencia y ausencia de Gcsf-Chr19.
2. Determinar los niveles transcripcionales de cxcl8a y cxcl8b, y otros genes
involucrados en un proceso inflamatorio tales como: il-1b, il-6, cxcr1, cxcr2,
cxcr4b, gcsf-chr12, gcsf-chr19 y myd88 después de un daño estéril leve.
3. Caracterizar la migración de neutrófilos luego de un daño estéril leve y severo en
presencia y ausencia cxcl8a o cxcl8b.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
18
Materiales y métodos
Mantención de peces Se utilizaron peces cebra (Danio rerio) que se mantuvieron en el bioterio del
Laboratorio de biología del Desarrollo, Universidad Andrés Bello, de acuerdo a
protocolos previamente establecidos (Westerfield, 2009). Se usaron dos líneas
transgénicas, Tg(BACmpx:GFP)i114 (Renshaw et al., 2006) y Tg (fli1:EGFP)y1 (Lawson et
al., 2002). Tg(BACmpx:GFP)i114, es una línea transgénica de pez cebra que expresa
proteína fluorescente verde (GFP) bajo el promotor de mioeloperoxidasa (mpx)
específica de neutrófilos. Tg(fli1:EGFP)y1 es una línea transgénica que expresa EGFP
bajo el promotor del factor de transcripción Friend leukemia integration 1 (Fli1) que se
expresa en vasos sanguíneos (células endoteliales). Los embriones se recolectaron por
desove natural (Kimmel et al., 1995), y luego se mantuvieron en medio E3 (5 mM NaCl,
0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, pH 7.0) con Azul de metileno 0.01% (3
mL por 1 L de E3), en placas petri con no más de 50 embriones por placa en una
incubadora (VELP Scientifica) a 28°C. Los ensayos se realizaron 48 horas posteriores a
la fertilización (hpf).
Daño estéril Se llevaron a cabo dos tipos de daño, severo y leve. El daño severo corresponde
a una transección de la aleta caudal que incluye la amputación de tejido muscular
(Figura 4A), mientras que el daño leve corresponde a una transección de aleta caudal
que no incluye tejido muscular (Figura 4B). Ambos daños fueron realizados con bisturí
estéril. Previo a la realización de daños, los embriones se anestesiaron con tricaína
0.017% (Fluka Analytical) de acuerdo al protocolo descrito por Elks et al., 2011. Se
realizaron tres replicas biológicas de cada ensayo.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
19
Figura 4. Esquema de daños realizados. (A) Daño severo, cercano al loop circulatorio
y pérdida de tejido muscular. (B) Daño leve, perdida de tejido de la aleta sin músculo.
Expresión génica Se realizaron PCR de tiempo real (qPCR) para la determinación de la expresión
génica relativa de los distintos genes en estudio (cxcl8a y cxcl8b, il-1b, il-6, cxcr1, cxcr2,
sdf-1, cxcr4, gcsf-chr12, gcsf-chr19 y myd88) en las diferentes condiciones
experimentales con partidores que se especifican en la Tabla 1. Primeramente, se
extrajo el ARN utilizando TRIzol® Reagent (Life Technology) de acuerdo a las
instrucciones del fabricante, en los tiempos correspondientes de cada ensayo.
Posteriormente se sintetizó cADN utilizando la enzima Improm-II™ Reverse (Promega)
según las instrucciones del fabricante, con partidores oligo-dT. Finalmente se realizaron
qPCR utilizando un programa especificado en la Tabla 2, los resultados obtenidos se
estandarizaron con respecto a β-Actina. Se realizaron tres replicas bilógicas con cuatro
replicas técnicas cada una.
Los análisis de resultados de qPCR se realizaron, primero se normaliza al gen de
referencia calculando el ∆𝐶𝑡, para cada muestra obtenida, con la siguiente formula:
∆𝐶𝑡 = 𝐶𝑡%&')*+&,-.) − 𝐶𝑡%&'0&1&2&1&'3-4, luego se calculó la expresión con la siguiente
formula ∆𝐶𝑡&561&7-ó' = 2:∆;,, posteriormente se saca el promedio de las réplicas
técnicas y se calcula la desviación estándar, finalmente se normaliza al control
dividiendo los datos por el valor del control para dejarlo con valor igual a 1,

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
20
Tabla 1. Secuencia de partidores utilizados en los ensayos de qPCR.
Gen Partidor Secuencia 5’à3’
β-Actina Forward TTCTGGTCGTACTACTGGTATTGTG
Reverse ATCTTCATCAGGTAGTCTGTCAGGT
Cxcl8a Forward TGTGTTATTGTTTTCCTGGCATTT
Reverse GCGACAGCGTGGATCTACAG
Cxcl8b Forward CTACCGAGACGTGGGTGATT
Reverse GCTCGGTGAATGGTCATTTT
Cxcr2 Forward TGACCTGCTTTTTTCCCTCACT
Reverse TGACCGGCGTGGAGGTA
Cxcr4 Forward GCGACTTTTTTCACCATTCC
Reverse TTTCTCCAGACCCTGTTCC
Gcsf-chr12 Forward CTACTGGAGTCTGGTGATTGTGC
Reverse AGTTTCTGAGGAACAGGAGCAGA
Gcsf-chr19 Forward GTGAGTTCCAGATCCCGACG
Reverse TGTGATGAAGCTCCAGACCG
Il-1b Forward TGGACTTCGCAGCACAAAATG
Reverse GTTCACTTCACGCTCTTGGATG
Il- 6 Forward TCAACTTCTCCAGCGTGATG
Reverse TCTTTCCCTCTTTTCCTCCTG
MyD88 Forward TAAACGGCTAATCCCTGTCG
Reverse ATGGTCAGAAAGCGCAGAAT

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
21
Tabla 2. Programa qPCR. Segmento Número de ciclos Temperatura Tiempo 1 1 25 °C 1 segundo
2 1 95 °C 10 minutos
3 40
95 °C 15 segundos
Tm 15 segundos
72 °C 15 segundos
4 1
95 °C 15 segundos
25 °C 1 segundo
70 °C 15 segundos
95 °C 1 segundo
*Tm: temperatura específica para cada par de partidores.
Ensayos falta de función génica Se llevaron a cabo ensayos de falta de función génica utilizando morfolinos de
Splicing (Gene Tools) para Gcsf-Chr19, Cxcl8a y Cxcl8b (Tabla 3). Estos fueron
inyectados en embriones dobles transgénicos, obtenidos de la cruza entre la línea
Tg(Bacmpx: GFP) i114 y la línea Tg(fli1:EGFP)y1, un estadio de 1 célula de acuerdo al
protocolo descrito por Rosen et al. 2009. Para la selección de fenotipo morfante de
Cxcl8a y Cxcl8b se utilizó el criterio de disrupciones existentes en la vasculatura (Figura
5) descrita por Stoll at al. 2011, donde se utilizaron embriones con 2 o 3 disrupciones en
la prolongación de los vasos intersegmentales, y sin alteración en los vasos que se
encuentran en el tejido caudal hematopoyético. Para la selección de fenotipo morfante
ce Gcsf-chr19 se utilizó el criterio de Galdames et al. 2014, con una sección de
morfantes los cuales posee un número de entre 14 y 20 neutrófilos en el tejido caudal
hematopoyético. Como control de inyección de utilizaron morfolinos missences. Se
realizaron tres replicas biológicas de cada ensayo.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
22
Tabla 3. Secuencias de morfolinos
Nombre Tipo Secuencia 5’ à 3’ Concentración Gcsf-ch19 spli Splicing CAGGTGTTAGTGTGTATTTACCAGT 2,5 ng/embrión
Gcsf-ch19 ATG Traducción TAAAGAGCCTGATTGTTCAACTGGG 2,5 ng/embrión
Cxcl8a spli Splicing GGTTTTGCATGTTCACTTACCTTCA 10 ng/embrión
Cxcl8b spli Splicing TTAGTATCTGCTTACCCTCATTGGC 20 ng/embrión
Figura 5. Fenotipo morfante. (A, B, C) Imágenes representativas de los diferentes
fenotipos observados después de la microinyección de morfolino de Cxcl8a o Cxcl8b.
(A) Fenotipo control mostrando la vasculatura normal. (B) Fenotipo morfante usado en
los ensayos, de 2 a 3 disrupciones en la vasculatura, sólo en las venas
intersegmentales (fechas). (C) Fenotipo severo, con más de 3 alteraciones en la
vasculatura.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
23
Cuantificación de neutrófilos La cuantificación de neutrófilos se realizó en embriones controles e inyectados
con morfolino. En el del conteo de neutrófilos que llegaron al daño se realizó mediante
monitoreo in vivo a las 0, 1, 2 y 3 horas posterior al daño severo o leve (Figura 6A). En
el conteo de neutrófilos en sangre, se realizaron time-lapse de 5 minutos; a los tiempos
de 1, 2, 3 horas posterior al daño, severo o leve, además de un control sin daño. Los
conteos fueron realizados en la vena caudal (Figura 6B). Todas las cuantificaciones in
vivo y los time-lapse fueron realizados en una lupa estereoscópica de fluorescencia
(Olympus, SZX16). Se realizaron tres replicas biológicas de cada ensayo.
Figura 6. Esquema de las zonas de cuantificación de neutrófilos. (A) Área de
conteo en aleta caudal. (B) Área de conteo en circulación, vena caudal.
Estadística Los datos fueron analizados en el programa Prism 6 (GraphPad) usando Kruskal-
Wallis no-paramétrico, ANOVA de dos vías y test de múltiple comparación Dunn.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
24
Resultados Objetivo 1: Determinar los niveles transcripcionales de cxcl8a y cxcl8b, y otros genes
involucrados en un proceso inflamatorio tales como: il-1b, il-6, cxcr1, cxcr2, cxcr4b,
gcsf-chr12, gcsf-chr19 y myd88 después de un daño estéril severo en presencia y
ausencia de Gcsf-Chr19.
Con el fin de desarrollar el presente objetivo, se evaluaron los niveles
transcripcionales de distintos genes en un daño severo. Este daño corresponde a la
amputación de la aleta caudal incluyendo tejido muscular. Así, a embriones Tg(Bacmpx:
GFP)i114 de 48 horas post fertilización (hpf) se les realizó un daño severo y luego se
monitoreó los niveles de mRNA de un grupo de genes según se detalla a continuación.
Se evaluaron genes participantes en el proceso de liberación de neutrófilos del
tejido hematopoyético, Sdf-1, Cxcr4 y Gcsf-chr19, junto con su parálogo Gcsf-chr12.
También se evaluó el cambio en la expresión de los parálogos Cxcl8a y Cxcl8b, los
cuales corresponden a ortólogos de CXCL8 en humanos, además de los receptores
Cxcr1 y Cxcr2. Para verificar que el daño realizado efectivamente gatilló un proceso
inflamatorio se midieron las citoquinas pro-inflamatorias Interleuquina 1b (Il-1b) e
Interleuquina 6 (Il-6). Finalmente, para comprobar que el proceso inflamatorio fue
gatillado por un daño estéril y no por una infección se midieron los niveles del transcrito
del gen de respuesta primaria de diferenciación mieloide 88 (Myd88).
Niveles transcripcionales de diferentes genes durante un proceso inflamatorio luego de
un daño severo
Los resultados obtenidos, indican que, los niveles transcripcionales de il-1b e il-6
aumentan significativamente luego del daño. il-1b mantiene esta sobrexpresión durante
las tres horas posterior al daño (hpd), mientras que, il-6 vuelve a su estado basal a la
tercera hpd. Los niveles de mRNA de myd88 no presentan cambio significativo durante
el tiempo analizado (Figura 7A).
Los genes que participan en la liberación de neutrófilos desde el TCH, se pueden
observar que gcsf-chr19 se encuentra significantemente aumentado durante las tres
hpd, a diferencia de gcsf-chr12 que tiene una conducta oscilante, aumentando

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
25
significativamente su expresión durante la primera y tercera hpd, no así a la segunda
hpd. Por su parte, la expresión de sdf-1 disminuye significativamente durante la primera
hora de daño, para luego aumentar desde a la segunda hpd. Finalmente, los niveles de
cxcr4 no se ven afectados durante todo el tiempo analizado (Figura 7B).
Los niveles transcripcionales de los genes participantes en la migración de
neutrófilos, se puede observar que cxcl8a, cxcl8b y cxcr2 se encuentran
significativamente elevados durante las tres hpd, a diferencia de cxcr1 que no varía
(Figura 7C).
Figura 7. Niveles transcripcionales de genes relacionados a un proceso inflamatorio posterior a un daño severo. (A) Niveles de expresión relativos de genes
relacionados a un proceso inflamatorio en general. (B) Niveles de expresión relativo de
genes involucrados en la liberación de neutrófilos desde el tejido hematopoyético. (C)
Niveles de expresión relativo de genes relacionados a la migración de neutrófilos. Los
datos se presentan como el cambio en la expresión respecto a su control a 0 hpd y
están normalizados contra b-actina. *p-value < 0.05; **p-value < 0.01, ***p-value <
0.005.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
26
Niveles transcripcionales de diferentes genes durante un proceso inflamatorio luego de
un daño severo en ausencia de Gcsf-chr19
Dado que los antecedentes existentes indican que Gcsf-chr19 regula la entrada
de neutrófilos a la circulación y que en esta tesis se quiere determinar si esto lo hace
regulando la función de Cxcl8a o Cxcl8b, se analizaron los niveles transcripcionales de
los genes inmunes antes mencionados en ausencia de Gcsf-chr19. Para realizar la
inhibición de la función de Gcsf-chr19, se utilizó un morfolino (MO) contra dicho gen. Se
inyectaron embriones al estado de 1 célula con el MO y luego a las 48 hpf se realizó un
daño severo y se cuantificaron los niveles de mRNA de una serie de genes durante las
3 primeras horas posteriores al daño. Los resultados obtenidos muestran que tanto il-1b
e il-6 no se ven afectadas por la ausencia de Gcsf-chr19, manteniendo el mismo
comportamiento observado en presencia dicha citoquina. Es decir, il-1b esta
significativamente elevada durante las tres hpd analizadas e il-6 durante las dos
primeras hpd. Así mismo, los niveles de mRNA de myd88 no presentaron cambios
(Figura 8A).
Al analizar los genes relacionados con la salida de los neutrófilos del TCH,
pudimos observar que la ausencia de Gcsf-chr19 altera la expresión de gcsf-ch12,
aumentando significativamente desde la 0,5 hpd con un máximo de expresión a las 2
hpd (comparar Figura 7B con Figura 8B). Lo mismo ocurre con los niveles de sdf-1, los
que no se alteran en la ausencia de Gcsf-chr19 (comparar Figura 7B con Figura 8B). El
único gen que no altera su comportamiento de expresión es cxcr4, el cual no aumenta
sus niveles de expresión luego de un daño ni en ausencia como en presencia de Gcsf-
chr19. Finalmente, y a modo de control del experimento, los niveles transcripcionales de
gcsf-ch19 no aumentaron.
Por otra parte, al evaluar los niveles de mRNA de genes relacionados con la
migración de neutrófilos en ausencia de Gcsf-chr19, se pudo observar que su falta de
función altera la expresión de cxcl8b donde se observa que éstos no aumentan durante
un daño severo (Figura 8C), siendo que en presencia de Gcsf-chr19 presenta un
aumento significativo de los niveles de mRNA (Figura 7C). cxcl8a presentó un aumento
significativo en la segunda hora posterior al daño (Figura 8C), lo cual difiere con el
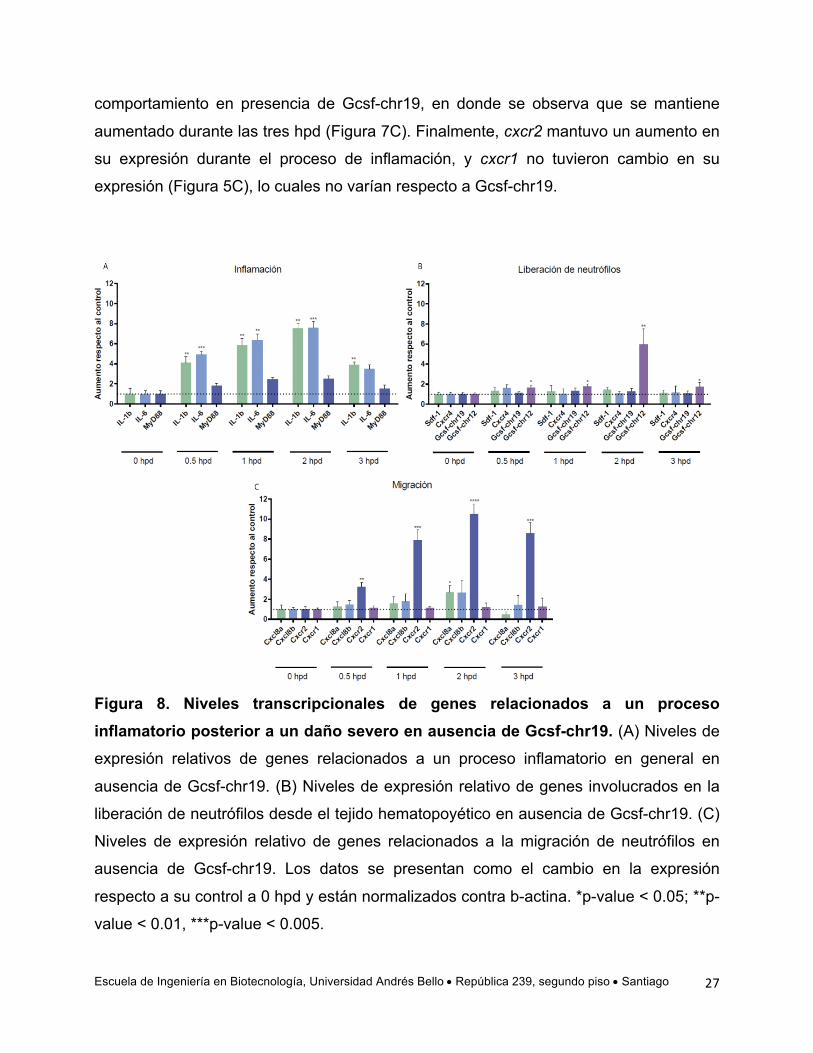
Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
27
comportamiento en presencia de Gcsf-chr19, en donde se observa que se mantiene
aumentado durante las tres hpd (Figura 7C). Finalmente, cxcr2 mantuvo un aumento en
su expresión durante el proceso de inflamación, y cxcr1 no tuvieron cambio en su
expresión (Figura 5C), lo cuales no varían respecto a Gcsf-chr19.
Figura 8. Niveles transcripcionales de genes relacionados a un proceso inflamatorio posterior a un daño severo en ausencia de Gcsf-chr19. (A) Niveles de
expresión relativos de genes relacionados a un proceso inflamatorio en general en
ausencia de Gcsf-chr19. (B) Niveles de expresión relativo de genes involucrados en la
liberación de neutrófilos desde el tejido hematopoyético en ausencia de Gcsf-chr19. (C)
Niveles de expresión relativo de genes relacionados a la migración de neutrófilos en
ausencia de Gcsf-chr19. Los datos se presentan como el cambio en la expresión
respecto a su control a 0 hpd y están normalizados contra b-actina. *p-value < 0.05; **p-
value < 0.01, ***p-value < 0.005.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
28
Objetivo 2: Determinar los niveles transcripcionales de cxcl8a y cxcl8b, y otros genes
involucrados en un proceso inflamatorio tales como: il-1b, il-6, cxcr1, cxcr2, cxcr4b,
gcsf-chr12, gcsf-chr19 y myd88 después de un daño estéril leve.
Para poder determinar si la intensidad del daño influye en el patrón de expresión
de los genes involucrados, y si éstos se expresan diferencialmente, es que se evalúa
los niveles de mRNA en un daño leve, el cual corresponde a una amputación de aleta
caudal sin tejido muscular.
Niveles transcripcionales de diferentes genes en un proceso inflamatorio luego de un
daño leve
Los resultados obtenidos muestran que tanto il-1b, il-6 como myd88 se
comportan de igual forma que en la situación de daño severo; il-1b e il-6 se ven
aumentadas significativamente respecto al estado inicial y myd88 que no presenta
cambio en ninguno de los tiempos analizados (Figura 9A).
Respecto a los genes que participan en el proceso de liberación de neutrófilos
TCH, ninguno de los genes evaluados; gcsf-chr12, gcsf-chr19, sdf-1 y cxcr4, aumenta
su expresión luego de realizado el daño leve (Figura 9B).
Finalmente, en los genes relacionados con la migración de neutrófilos, se
observa que cxcl8a y cxcr2 aumentan significativamente su expresión, no así de cxcl8b
y cxcr1. cxcl8a presenta un comportamiento oscilante, aumentando los niveles de
mRNA significativamente a las 0.5hpd y 3hpd. Por su parte, cxcr2 incrementa sus
niveles transcripcionales desde las 0.5hpd (Figura 9C).

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
29
Figura 9. Niveles transcripcionales de genes relacionados a un proceso inflamatorio posterior a un daño leve. (A) Niveles de expresión relativos de genes
relacionados a un proceso inflamatorio en general. (B) Niveles de expresión relativo de
genes involucrados en la liberación de neutrófilos desde el tejido hematopoyético. (C)
Niveles de expresión relativo de genes relacionados a la migración de neutrófilos. Los
datos se presentan como el cambio en la expresión respecto a su control a 0 hpd y
están normalizados contra b-actina. *p-value < 0.05; **p-value < 0.01, ***p-value <
0.005.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
30
Objetivo 3: Caracterizar la migración de neutrófilos luego de un daño estéril leve y
severo en presencia y ausencia cxcl8a o cxcl8b.
Dada la diferencia encontrada entre un daño severo, donde genes involucrados
con la entrada a los vasos sanguíneos, como Gcsf-chr19, aumentan su expresión, y un
daño leve, donde dichos genes no lo hacen, sumado a la diferencia encontrada en
relación a los niveles relativos de expresión de Cxcl8a y Cxcl8b, se decidió evaluar los
efectos de la falta de funcional de ambas proteínas en la migración de neutrófilos. Para
este propósito se utilizaron morfolinos para cada uno de los genes.
Determinación del número de neutrófilos presentes en circulación y en zona afectada
luego de un daño severo
En el modelo de daño severo, la ausencia de tanto Cxcl8a como Cxcl8b
disminuyen significativamente el número de neutrófilos presentes en la zona del daño
en comparación al control durante las tres horas de monitoreo. En el caso de Cxcl8a,
esta disminución es mayor a la tercera hpd (Figura 10B).
Dado que el número de neutrófilos presentes en el daño es el resultado de las
diferentes etapas de migración hacia éste, y que se postula que Cxcl8a estaría
relacionado al ingreso de neutrófilos a los vasos sanguíneos, es que se determinó el
número de neutrófilos presentes en el torrente sanguíneo durante el proceso
inflamatorio. Se observó que en larvas control sin daño, la cantidad de neutrófilos en
circulación se mantiene constante, siendo esta muy baja, cercana a 0 neutrófilo,
durante los 5 minutos analizados en cada hora (Figura 11A). Al realizar un daño severo,
se genera un aumento significativo y progresivo en el transcurso del tiempo de la
cantidad de neutrófilos que se encuentran en circulación, aumentando de 0 neutrófilos
en 5 min durante la primera hpd hasta 2 neutrófilos en 5 min luego a las tres hpd
(Figura 11B). En los embriones morfantes de Cxcl8a, el número de neutrófilos aumenta
significativamente luego del daño, tal como ocurre en el control con daño (Figura 11C).
Por otro lado, en los embriones morfantes de Cxcl8b el número de neutrófilos en
circulación permanece bajo y constante, similar a lo observado en el control sin daño
(Figura 11D).

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
31
Figura 10. Determinación del número de neutrófilos que llega a la zona dañada luego de un daño severo en ausencia de Cxcl8a o Cxcl8b. (A) Vista lateral de una
sección de la aleta caudal luego de 1, 2 y 3 horas de realizado el daño severo en larvas
control y morfantes. (B) Cuantificación de neutrófilos en el daño a las 1, 2 y 3 horas
posteriores al daño. *p-value < 0.05; **p-value < 0.01, ***p-value < 0.005.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
32
Figura 11. Cuantificación de neutrófilos circulantes luego de un daño severo en presencia y ausencia de Cxcl8a o Cxcl8b. (A) Cuantificación de neutrófilos
circulantes en el torrente sanguíneo en larvas sin daño; (B) con daño severo; (C) con
daño severo, en ausencia de Cxcl8a; (D) con daño severo ausencia de Cxcl8b. *p-value
< 0.05; **p-value < 0.01, ***p-value < 0.005.
Determinación del número de neutrófilos presentes en circulación y en zona afectada
luego de un daño leve
Los resultados obtenidos durante un proceso inflamatorio gatillado por un daño
leve, indican que sólo la ausencia de Cxcl8a afecta el número de neutrófilos que
migraron al daño, siendo significativamente menor respecto al control. En embriones
morfantes de Cxcl8b no se observa diferencia significativa con embriones control
(Figura 12C).

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
33
Figura 12. Determinación del número de neutrófilos que llega a la zona dañada luego de un daño leve en ausencia de Cxcl8a o Cxcl8b. (A) Vista lateral de una
sección de la aleta caudal luego de 1, 2 y 3 horas de realizado el daño leve en larvas
control y morfantes. (B) Cuantificación de neutrófilos en el daño a las 1, 2 y 3 horas
posteriores al daño. *p-value < 0.05; **p-value < 0.01, ***p-value < 0.005.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
34
Discusión Con el paso de los años y el avance en los estudios científicos, se ha podido
observar que aun cuando a nivel fisiológico los procesos son conservados, estos
pueden diferir a nivel molecular aún en especies evolutivamente muy cercanas. Un
ejemplo de esto, es la diferencia existente en relación a CXCL8, entre humanos y
roedores. Este hecho realza la importancia de conocer en profundidad la biología de
cada especie, y que no siempre es posible extrapolar mecanismos fisiológicos,
celulares y/o moleculares entre diferentes organismos.
En particular, esta tesis se centra en entender el mecanismo de regulación de la
migración de neutrófilos hacia los vasos sanguíneos en peces. Dada la problemática
antes expuesta y los resultados obtenidos durante el desarrollo de este trabajo, es que
se propone el pez cebra como animal modelo para estudios in vivo sobre la
movilización de neutrófilos durante procesos inflamatorios. Esto, tanto a nivel molecular
como celular, así también como para propósitos biomédicos.
Para determinar los roles de los parálogos Cxcl8a y Cxcl8b en la entrada de
neutrófilos a los vasos sanguíneos durante un daño mecánico estéril y su relación con
Gcsf-chr19, se llevó a cabo dos tipos de daños. En uno de ellos, el severo, en donde
existe migración de neutrófilos por vasos sanguíneos a diferencia del segundo, daño
leve, donde esto no ocurre (Galdames et al., 2014). La comparación de ambos
procesos a nivel molecular nos permitió identificar cuál de estos genes participa en la
entrada de neutrófilos a la circulación sanguínea. Por otro lado, ensayos de daño
severo en ausencia de Gcsf-chr19 nos permitió determinar si dicha citoquina reluga la
función de Cxcl8a y/o Cxcl8b.
En los ensayos de falta de función de Cxcl8a y Cxcl8b, a través de la acción de
un morfolino, se pudo observar que ambas citoquinas participan en la regulación en la
migración de neutrófilos en un modelo de daño severo. El número de neutrófilos
presentes en el daño disminuyó significativamente en larvas morfantes para Cxcl8a o
para Cxcl8b en comparación a larvas control. Este resultado concuerda con lo descrito
por De Oliveira et al., 2013, donde se demuestra que la migración de neutrófilos al lugar
del daño disminuyo en los knock-down de Cxcl8a como de Cxcl8b y que la presencia de
ambas citoquinas es necesaria para la mantención de comportamiento migratorio de

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
35
neutrófilos. Por otra parte, para poder determinar si los parálogos de Cxcl8 regulan la
entrada de neutrófilos al torrente sanguíneo, se realizó un conteo de neutrófilos
circulantes en la vena caudal. Los resultados obtenidos sugieren que Cxcl8b, no así
Cxcl8a, estaría regulando la migración de neutrófilo a través de los vasos sanguíneos.
Esta regulación de entrada de neutrófilos a los vasos sanguíneos que posee
CXCL8 en mamíferos, ha sido reportada anteriormente en ensayos clínicos con
humanos donde se observó un efecto de aumento de números de neutrófilos de en
ciclación al inyectar de forma exógena CXCL8, también se ha observado el mismo
efecto en estudios realizados en monos, conejos y ratones (Jagels & Hugl, 1992;
Laterveer et al.,1995; Van et al., 1992; Watanabe et al., 1999), la relevancia de los
resultados presentados es que al existir una duplicación de Cxcl8 en pez cebra, éstos
presentan una función complementaria, las cuales suplen la función completa que
posee CXCL8 en mamíferos.
En el caso de daño leve, los resultados obtenidos sugieren que Cxl8a, y no
Cxcl8b, regularía la migración final de neutrófilos al daño a través del tejido intersticial.
Esto, ya que en ausencia de Cxl8a, existió una disminución en el número de neutrófilos
presentes en el daño en comparación con el control, en cambio, Cxcl8b no presenta
variación significativa en el número de neutrófilos en el daño respecto al control.
El análisis transcripcional realizado luego de un daño severo como de uno leve
mostró que los niveles de las citoquinas pro-inflamatorias il-1b e il-6 aumentaron
indicando que ambos tipos de daños gatillaron un proceso inflamatorio (Ogryzko et al.,
2014; Varela et al., 2012).
En relación al proceso de liberación de neutrófilos desde el TCH se observó que,
durante las dos primeras horas de daño severo, sdf-1 disminuyó significativamente,
posiblemente debido a la acción que genera Gcsf-chr19 sobre éste ligando, lo cual
concuerda con lo descrito en ratones que indica que G-CSF inhibe la actividad de los
osteoblastos en la médula ósea, reduciendo la expresión de SDF-1 (Semerad et al.,
2005). Los resultados obtenidos en ausencia de Gcsf-chr19 apoyan esta hipótesis, ya
que los niveles sdf-1 no se vieron afectados respecto al control durante un daño severo.
Por su parte, los niveles de Cxcr4 no sufrieron cambios en ninguno de los daños
llevados a cabo, indicando que la función de este receptor de quimioquina no sería

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
36
regulada transcripcionalmente. Estos resultados no concuerdan con lo descrito en
mamíferos, donde se ha reportado existe una disminución de CXCR4 en células
mieloides tanto a nivel transcripcional como proteico, gatillada por la acción de G-CSF,
(Kyung et al. 2006; Levésque et al., 2003). Con estos resultados se puede inferir que en
pez cebra, la acción de Gcsf-chr19 en el proceso de liberación de neutrófilos desde el
TCH es mediada por Sdf-1.
Respecto a Gcsf-chr19, los ensayos de qPCR indican que éste únicamente
aumenta sus niveles de mRNA luego de un daño severo, confirmando que solo
participa en aquellos procesos inflamatorios donde ocurre migración de neutrófilos por
los vasos sanguíneos. El aumento significativo de gcsf-chr12 luego de un daño severo
en ausencia de Gcsf-chr19, posiblemente se debe a una función compensatoria. Esta
sería una nueva función de Gcsf-chr12, que se sumaría a su ya descrita participación
en el proceso de mielopoyésis (Liongue et al., 2009; Stachura et al., 2013).
En lo que respecta al proceso de migración de neutrófilos por los vasos
sanguíneos, de todos los genes evaluados, cxcr1 fue el único no que presentó cambios
en su expresión transcripcional, lo que indica que no estaría participando en el proceso,
o si lo hace, su función no es regulada transcripcionalmente. Por otro lado, los niveles
de mRNA de cxcr2 aumentan luego de un daño severo y de uno leve, además en
presencia y ausencia de Gcsf-chr19. Estos resultados sugieren que Cxcr2 participa
activamente en el proceso de migración de neutrófilos y que su función no depende de
Gcsf-chr19. Según lo descrito en la literatura, Cxcr2 participa específicamente en el en
proceso final de migración de neutrófilos, esto es en la llegada a la zona dañada en pez
cebra (Deng et al., 2013), en ratones se ha visto que CXCR2 estaría regulando los
niveles de GCSF y no GCSF los de CXCR2, esto a través de IL-17 con el objetivo de
lograr la homeostasis en los niveles de neutrófilos en circulación y así promover la
resolución de la inflamación (Mei et al., 2012). Con estos resultados no podemos
descartar que Cxcr2 también participe en la regulación de la migración de neutrófilos
por los vasos sanguíneos en pez cebra. Además, se observó que durante un daño
severo la expresión de cxcl8a y cxcl8b aumentó significativamente indicando que
participan en el proceso inflamatorio severo estéril. En cambio, en el daño leve sólo
cxcl8a presento una sobreexpresión de transcritos, sugiriendo que se encuentra

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
37
involucrado en la migración de neutrófilos a través de matriz extracelular y no por los
vasos sanguíneos, lo cual se estaría correlacionado con los resultados obtenidos con la
falta de función de ambos Cxcl8, donde se sugieren que Cxcl8b, no así Cxcl8a, estaría
regulando la migración de neutrófilo a través de los vasos sanguíneos.
En resumen, con los resultados obtenidos durante el desarrollo de esta tesis se
refuta una parte de la hipótesis planteada, aquella en donde se asevera que Cxcl8a
regula la entrada de neutrófilos a los vasos sanguíneos. Como ya se dijo, nuestros
resultados indican que es Cxcl8b. La segunda parte de nuestra hipótesis es verdadera
Gcsf-chr19, por medio de su regulación sobre Cxcl8b, controla la entrada de neutrófilos
a los vasos sanguíneos luego de daño severo.
Se plantea el siguiente modelo de migración: Durante un proceso inflamatorio
gatillado por daño estéril severo, Gcsf-chr19 disminuiría los niveles de Sdf-1,
favoreciendo la liberación de neutrófilos desde el tejido hematopoyético y al mismo
tiempo aumentaría la expresión de cxcl8b permitiendo la entrada a los vasos
sanguíneos. Es importante destacar que con nuestros resultados no podemos afirmar ni
descartar que la acción de Cxcl8b se llevase a cabo al unirse al receptor Cxcr2.
Finalmente, los neutrófilos serían atraídos a la zona del daño por la acción de Cxcl8a y
su receptor Cxcr2 (Figura 13).

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
38
Figura 13. Modelo final de migración de neutrófilos a una inflamación en pez cebra. (A) Los neutrófilos se mantienen retenidos en tejido hematopoyético debido a la
unión del ligando Sdf-1 al receptor Cxcr4. (B) Gsf-chr19 inhibe los niveles de Sdf-1
generando la liberación de los neutrófilos y aumenta los niveles Cxcl8b permitiendo el
paso a los vasos sanguíneos y finalmente dirigirse al lugar del daño por señales
quimioatractantes de Cxcl8a/Cxcr2.
Conclusión El trabajo realizado permite clarificar, al menos en parte, el mecanismo molecular
que controla la migración de neutrófilos a los vasos sanguíneos en pez cebra durante
un proceso inflamatorio. Específicamente, podemos concluir que:
1. En un proceso inflamatorio gatillado por un daño estéril severo, Gcsf-Chr19 regula
la expresión de sdf-1 y cxcl8b principalmente y con ello favorece la salida de neutrofilos
desde el territorio caudal hematopoyetico y su entrada a los vasos sanguineas,
respectivamente.
2. En un proceso inflamatorio gatillado por un daño estéril leve, no se requiere
liberación de neutrófilos desde el tejido caudal hematopoyético, por lo que neutrofilos
ubicados fuera de este territorio serían los que responde. La migración de dichos
neutrofilos es a traves del tejido intersticial y regulada por la vía de señalizacion
Cxcl8a/Cxcr2.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
39
Referencias
Anderlini P, Przepiorka D, Champlin R, Korbling M. (1996) Biologic and clinical effects of granulocyte colony-stimulating factor in normal individuals. Blood Journal
88:2819-2825.
Abbas A, Litchman A and Pillai S (2015) Basic Immunology: Functions and Disorders of the Immune System. 5a Ed. Elsevier, 1-3.
Chen G and Nuñez G (2010) Sterile inflammation: sensing and reacting to damage. Nature Reviews Immunology 10: 826-837.
Demetri G and Griffin J. (1991) Granulocyte colony stimulating factor and its receptor. Blood Journal 78:2791- 2808.
Deng Q, Sarris M, Bennin D, Green J, Herbomel P, Huttenlocher A. (2013) Localized bacterial infection induces systemic activation of neutrophils through Cxcr2 signaling in zebrafish. Journal Leukocyte Biology 93:761-9.
De Oliveira S, Reyes-Aldasoro C, Candel S, Renshaw S, Mulero V, Calado A. (2013)
Cxcl8 (IL-8) mediates neutrophil recruitment and behavior in the zebrafish inflammatory response. Journal of Immunology 190:4349-59.
Eash K, Greenbaum A, Gopalan P and Link D (2010) CXCR2 and CXCR4 antagonistically regulate neutrophil trafficking from murine bone marrow. Journal
Clinical Investigation 120: 2423-2431.
Elks P, Loynes A and Renshaw S (2011) Measuring inflammatory cell migration in the zebrafish. Cell Migration: Developmental Methods and Protocols. Chapter 18.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
40
Eyles J, Roberts A, Metcalf D and Wicks I (2006) Granulocyte colony-stimulating factor and neutrophils—forgotten mediators of inflammatory disease. Nature 2:
500-510.
Galdames J, Zuñiga-Traslaviña C, Reyes A and Feijoo C (2014) Gcsf-Chr19 Promotes Neutrophil Migration to Damages Tissue through Blood Vesselsin Zebrafish. Journal of Immunology 193: 372-378.
Issekutz A, Movat H (1980) The in vivo quantitation and kinetics of rabbit neutrophil leukocyte accumulation in the skin in response to chemotactic agents and Escherichia coli. Laboratory Investigation 42: 310-7.
Jagels M and Hugl T (1992) neutrophil chemotactic factors promote leukocytosis: A common mechanism for Cellular Recruitment from Bone Marrow. Journal of
Immunology 148: 1119-1128
Kanther M and Rawls J (2010) Host-microbe interactions in the developing zebrafish. Current Opinion in Immunology 22: 10-19.
Kari G, Rodeck U and Dicker A (2007) Zebrafish: An Emerging Model System for Human Disease and Drug Discovery. Clinical pharmacology & therapeutics 82: 70 -
80.
Kimmel C, Ballard W, Kimmel S, Ullmann B and Schilling T (1995) Stages of embryonic development of the zebrafish. Developmental Dynamic 203: 253-310.
Kyung K, De La Luz M, Cassin W, Gulino A and Tosato G (2006) G-CSF down-regulation of CXCR4 expression identified as a mechanism for mobilization of myeloid cells. Blood Journal 108:812-820.
Klosterman (2009). Immune System. Marshall Cavendish Benchmark. ISBN: 978-0-
7614-3054-4.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
41
Laterveer L, Lindley I, Hamilton M, Willemze R and Fibbe W (1995) Interleukin-8 Induces Rapid Mobilization of Hematopoietic Stem Cells with Radioprotective Capacity and Long-Term Myelolymphoid Repopulating Ability. Blood Journal 85:
2269-2275.
Lawson N and Weinstein B (2002) In vivo imaging of embryonic vascular development using transgenic zebrafish. Developmental Biology 248: 307-318.
Layton J, Hall N (2006) The interaction of G-CSF with its receptor. Frontiers
Bioscience 11:3181- 3189.
Licastro F, Candore G, Lio D, Porcellini E, Colonna-Romano G, Franceschi C and
Caruso C (2005) Innate immunity and inflammation in ageing: a key for understanding age-related diseases. Immunity & Ageing 2:8.
Liongue C, Hall C, O'Connell B, Crosier P and Ward A (2009) Zebrafish granulocyte colony-stimulating factor receptor signaling promotes myelopoiesis and myeloid cell migration. Blood Journal 113: 2535-2546.
Lévesque J, Hendy J, Takamatsu Y, Simmons P and Bendall L (2003) Disruption of the CXCR4/CXCL12 chemotactic interaction during hematopoietic stem cell mobilization induced by GCSF or cyclophosphamide. The Journal of Clinical
Investigation 111: 187-196.
Magnadottir B (2006) Innate immunity of fish (overview). Fish Shellfish Immunology
20: 137-151.
Magnadottir B (2010) Immunological Control of Fish Diseases. Marine Biotechnology
12: 361-379.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
42
Mei J, Liu Y, Dai N, Hoffmann C, Hudock K, Zhang P, Guttentag S, Kolls J, Oliver
P, Bushman F and Worthen G (2012) Cxcr2 and Cxcl5 regulate the IL-17/G-CSF axis and neutrophil homeostasis in mice. The Journal of Clinical Investigation 122: 974-
986.
Novoa B and Figueras A (2012) Zebrafish: model for the study of inflammation and the innate immune response to infectious diseases. Advances in Experimental
Medicine and Biology 253-275.
Oehlers S, Flores M, Hall C, O’Toole R, Swift S, Crosier K and Crosier P (2010)
Expression of zebrafish cxcl8 (interleukin-8) and its receptors during development and in response to immune stimulation. Developmental and Comparative
Immunology 34: 352-359.
Ogryzko N, Hoggett E, Solaymani-Kohal S, Tazzyman S, Chico T, Renshaw S, Wilson H
(2014) Zebrafish tissue injury causes upregulation of interleukin-1 and caspase-dependent amplification of the inflammatory response. Disease Model Mechanism
7:259-64.
Palic D, Andreasen C, Ostojic J, Tell R and Roth J (2007) Zebrafish (Danio rerio) whole kidney assays to measure neutrophil extracellular trap release and degranulation of primary granules. Journal of Immunology Methods 319: 87-97
Raghuwanshi S, Su Y, Singh V, Haynes K, Richmond A and Richardson R (2012) The chemokine receptors CXCR1 and CXCR2 couple to distinct G protein-coupled receptor kinases to mediate and regulate leukocyte functions. Journal of
Immunology 189: 2824–2832
Renshaw S, Loynes C, Trushell D, Elworthy S and Ingham P (2006) A transgenic zebrafish model of neutrophilic inflammation. Blood Journal 108: 3976-3978.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
43
Rosen J, Sweeney M and Mably J (2009) Microinjection of zebrafish embryos to analyze gene function. Journal of visualized experiments. JoVE Journal 25.
Sarris M, Masson J, Maurin D, Van der Aa L, Boudinot P, Lortat-Jacob H, Herbomel P
(2012) Inflammatory chemokines direct and restrict leukocyte migration within live tissues as glycan-bound gradients. Current Biology 22: 2375-82.
Semerad C, Liu F, Gregory A, Stumpf K and Link D (2002) G-CSF is an essential regulator of neutrophil trafficking from the bone marrow to the blood. Immunity
Journal 17: 413-423.
Semerad C, Christopher M, Liu F, Short B, Simmons P, Winkler I, Levesque J, Chappel
J, Ross F and Link D (2005) G-CSF potently inhibits osteoblast activity and CXCL12 mRNA expression in the bone marrow. Blood Journal 106: 3020-3027.
Stachura D, Svoboda O, Campbell C, Espín-Palazón R, Lau R, Zon L, Bartunek
P, Traver D (2013) The zebrafish granulocyte colony-stimulating factors (Gcsfs): 2 paralogous cytokines and their roles in hematopoietic development and maintenance. Blood Journal 122:3918-28.
Stoll S, Bartsch S, Augustin H and Kroll J (2011) The transcription factor HOXC9 regulates endothelial cell quiescence and vascular morphogenesis in zebrafish via inhibition of interleukin 8. Circulation Research. 108: 1367-1377.
Taga T and Kishimoto T (1997) GP130 and the interleukin-6 family of cytokines. Annual Reviews Immunology 15: 797-819.
Teixidó J, Hermier M, Greenberger J and Anklesaria P (1992) Role of B1 and B2 integrins in the adhesion of human CD34hl stem cells to bone marrow stroma. The
American Society for Clinical Investigation 90: 358-367.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
44
Van J, Decock B, Conings R, Lenaerts J, Opdenakker G and Billiau A (1989) The chemotactic activity for granulocytes produced by virally infected fibroblasts is identical to monocyte-derived interleukin 8. European Journal of Immunology 19:
1189–1194.
Van Z, Fischer E, Hawes A, Hebert C, Terre T, Baker J, Lowry S and Molldawer L
(1992) Effects of intravenous IL-8 administration in nonhuman primates. Journal of
Immunology 148:1746.
Varela M, Dios S, Novoa B, Figueras A (2012) Characterisation, expression and ontogeny of interleukin-6 and its receptors in zebrafish (Danio rerio). Developmental & Comparative Immunology 37: 97–106.
Vento S, Cainelli F (2003) Infections in patients with cancer undergoing chemotherapy: aetiology, prevention, and treatment. The Lancet Oncology 4: 595–
604.
Walters K, Green J, Surfus J, Yoo S and Huttenlocher A (2010) Live imaging of neutrophil motility in a zebrafish model of WHIM syndrome. Blood Journal 116: 2803-
2811.
Watanabe T, Kawano Y, Kanamaru S, Onishi T, Kaneko S, Wakata Y, Nakagawa R,
Makimoto A, Kuroda Y, Takaue Y and Talmadge J (1999) Endogenous Interleukin-8 (IL-8) Surge in Granulocyte Colony-Stimulating Factor–Induced Peripheral Blood Stem Cell Mobilization. Blood Journal 93: 1157-1163.
Westerfield M, Zon L, Detrich H (2009) Essential zebrafish methods: Cell & Developmental biology. Academic Press. ISBN: 9780123745996.

Escuela de Ingeniería en Biotecnología, Universidad Andrés Bello • República 239, segundo piso • Santiago
45
White J, Lee J, Young P, Hertzberg R, Jurewicz A, Chaikin M, Widdowson K, Foley J,
Marton L, Griswold D, Sarau H (1998) Identification of a Potent, Selective Non-peptide CXCR2 Antagonist that inhibits interleukin-8-induced neutrophil migration. Journal of Biological Chemistry 273; 10095-10098.




















