
Air Accident Investigation Sector Slow Cabin Depressurization ...
Upload
independentCategory
view
2download
0
http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/47/8/995The online version of this article can be found at:
DOI: 10.1177/002215549904700803
1999 47: 995J Histochem CytochemMoorman
Petra E.M.H. Habets, Diego Franco, Jan M. Ruijter, Anthony J. Sargeant, José A.A. Sant'Ana Pereira and Antoon F.M.Muscle Gene Expression
RNA Content Differs in Slow and Fast Muscle Fibers: Implications for Interpretation of Changes in
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Aug 1, 1999Version of Record >>
by guest on July 19, 2014jhc.sagepub.comDownloaded from by guest on July 19, 2014jhc.sagepub.comDownloaded from

© The Histochemical Society, Inc.
0022-1554/99/$3.30
995
ARTICLE
Volume 47(8): 995–1004, 1999The Journal of Histochemistry & Cytochemistry
http://www.jhc.org
RNA Content Differs in Slow and Fast Muscle Fibers: Implications for Interpretation of Changes in Muscle Gene Expression
Petra E.M.H. Habets, Diego Franco, Jan M. Ruijter, Anthony J. Sargeant,José A.A. Sant’Ana Pereira, and Antoon F.M. Moorman
Department of Anatomy and Embryology, University of Amsterdam, Amsterdam, The Netherlands (PEMHH,DF,JMR,JAAS’AP,AFMM); Neuromuscular Biology Group, Manchester Metropolitan University, Alsager,United Kingdom (PEMHH,AJS); Institute for Fundamental and Clinical Human Movement Sciences, Vrije Universiteit, Amsterdam, The Netherlands (AJS); and Department of Physiology, University of Wisconsin Medical School, Madison, Wisconsin (JAAS’AP)
SUMMARY
Quantification of a specific muscle mRNA per total RNA (e.g., by Northernblot analysis) plays a crucial role in assessment of developmental, experimental, or patho-logical changes in gene expression. However, total RNA content per gram of a particular fi-ber type may differ as well. We have tested this possibility in the distinct fiber types ofadult rat skeletal muscle. Sections of single fibers were hybridized against 28S rRNA as amarker for RNA content. Quantification of the hybridization showed that the 28S rRNA
content decreases in the order I
.
IIA
.
IIX
.
IIB, where Type I fibers show a five- to sixfoldhigher expression level compared to Type IIB fibers. Results were verified with an indepen-dent biochemical determination of total RNA content performed on pools of histochemi-cally defined freeze-dried single fibers. In addition, the proportion of myosin heavy chain(MHC) mRNA per
m
g of total RNA was similar in slow and fast fibers, as demonstrated byNorthern blot analysis. Consequently, Type I fibers contain five- to sixfold more MHC mRNAper
m
g of tissue than IIB fibers. These differences are not reflected in the total fiber proteincontent. This study implies that proper assessment of mRNA levels in skeletal muscle re-quires evaluation of total RNA levels according to fiber type composition.
(J Histochem Cytochem 47:995–1004, 1999)
I
t is widely recognized
that skeletal muscle fibersfall into several specialized classes, termed fiber types,which show differences in morphological, contractile,and biochemical properties (e.g., Moss et al. 1995;Schiaffino and Reggiani 1996). Type I fibers are slowcontracting and relaxing fibers, which are innervatedby slow motor neurons with a low-frequency firingpattern. To sustain the high workload imposed by theinnervating nerve, slow fibers have a high oxidativecapacity and are therefore not easily fatigued. The fastcontracting Type II fibers can be divided into severalsubgroups, depending on the species. For example, inadult rat hindlimb muscle, three subgroups can be dis-
tinguished: Types IIA, IIX, and IIB. The fatigue-resis-tant Type IIA fibers contain intermediate to high levelsof glycogenolytic enzymes, and in rodents these fibershave the highest oxidative capacity. Furthermore,Type IIA fibers have higher contraction and relaxationrates compared to Type I fibers. With respect to theabove-mentioned characteristics, Type IIA fibers arefollowed by Type IIX fibers and finally by Type IIB fi-bers. The latter have the highest contraction and re-laxation rates, a high glycolytic capacity, low levels ofoxidative enzymes, and are most easily fatigued. Thedifferences in contractile characteristics of the fibersresult mainly from the expression of different myosinheavy chain (MHC) isoforms, which appear to be theprimary determinant of functional variability (for re-view see Pette and Staron 1990; Schiaffino and Reggi-ani 1994,1996). In adult rat hindlimb muscle, fourdistinct MHC isoforms, Type I (
b
-cardiac) and TypesIIA, IIX, and IIB, can be distinguished, each of which
Correspondence to: Antoon F.M. Moorman, Dept. of Anatomyand Embryology, Academic Medical Center, Meibergdreef 15, 1105AZ Amsterdam, The Netherlands.
Received for publication October 5, 1998; accepted March 30,1999 (8A4797).
KEY WORDS
skeletal muscle
single fibers
RNA content
in situ hybridization
densitometry
by guest on July 19, 2014jhc.sagepub.comDownloaded from

996
Habets, Franco, Ruijter, Sargeant, Sant’Ana Pereira, Moorman
is encoded by a single gene (Mahdavi et al. 1987; DeNardi et al. 1993).
The several distinct MHC isoforms, which can evenbe co-expressed in the same fiber, provide the musclefibers with the ability to modulate their contractilecharacteristics by switching different isoform genes onand off. This property, commonly referred to as mus-cle plasticity, has been extensively studied in experi-mental and pathological conditions, mainly at the pro-tein level (Swyngedauw 1980; Pette and Staron 1990;Caiozzo et al. 1992,1996). The introduction of molec-ular biological techniques, in addition to biochemicaltechniques, enables the detection of changes in iso-form expression at the transcriptional level. The vastmajority of studies that assessed changes in gene ex-pression have quantified muscle mRNA in tissue ho-mogenates using Northern blot, dot-blot, RNase pro-tection, or RT-PCR techniques. As a consequence,cellular resolution is lost and the relative changes inspecific mRNAs were expressed in relation to theamount of total RNA (specific mRNA/total RNA).However, no account was taken of whether the totalamount of RNA per amount of tissue varied amongthe distinct fiber types or whether it varied under dif-ferent experimental and pathological conditions. Clearly,without taking into account such differences in totalRNA content, absolute increases in signal can be mis-leading with respect to the effect of experimental in-terventions, e.g., exercise, chronic stimulation, on dif-ferent populations of muscle fiber types. Therefore, inthis study we examined the total RNA content usingin situ hybridization (ISH) and densitometry on his-tochemically defined single fibers (Sant’Ana Pereira etal. 1995a,b; Jonker et al. 1997). The densitometric re-sults were compared to a biochemical determinationof total RNA content in the same fiber. ISH on whole-muscle cross-sections and Northern blot analysis werecarried out to illustrate the results from the single-fiberexperiments.
Our data show that Type I fibers have a five- to six-fold higher total RNA content than the fastest TypeIIB fibers, whereas the MHC mRNA content per
m
g oftotal RNA is similar. Consequently, slow fibers alsohave five- to sixfold more MHC mRNA per
m
g of tis-sue than Type IIB fibers.
Material and Methods
Animal Care
Wistar rats (250–300 g) were housed with a 12-hr light and12-hr dark cycle, with food and water available ad libitum.After decapitation, the medial gastrocnemius (MG), soleus,and extensor digitorum longus (EDL) muscles were carefullyexcised. The muscles were slightly stretched, stored for 15min at 4C, frozen in liquid nitrogen-cooled isopentane, andprepared for cutting of whole-muscle cross-sections as de-
scribed (Sant’Ana Pereira et al. 1995b). To dissect single fi-bers, frozen whole-muscle preparations were freeze-dried(
2
60C) and processed under standard conditions of temper-ature and humidity as described by Sant’Ana Pereira et al.(1995a). Each fiber was cut into two parts. The first partwas embedded in RNase-free gelatin solution and used tocut transverse serial sections (10
m
m), which were pro-cessed for histochemistry and ISH. The second part wasused for total RNA determination using the orcinol method(see below).
Histochemistry
Serial cryostat sections of whole-muscle and gelatin-embed-ded fibers were stained for acid- and alkaline-labile myo-fibrillar adenosine triphosphatase (mATPase) activity andfor an additional mATPase assay that solely differentiates fi-bers expressing MHC IIX (Sant’Ana Pereira et al. 1995b,1997). The combination of these staining procedures allowscorrect identification of pure and hybrid I, IIA, IIX and IIBfibers. Further analyses of single fibers were restricted topure fibers to rule out, as far as possible, differences causedby the co-expression of two or more distinct MHC isoforms.
Preparation of cRNA Probes
Single [
35
S]-CTP and double [
35
S]-CTP and [
35
S]-UTP-labeled sense and antisense cRNAs of rat
b
(Boheler et al.1992), IIA (De Nardi et al. 1993), IIX (De Nardi et al.1993), and IIB (Schiaffino et al. 1989a) MHC isoforms,mouse 28S rRNA, human
b
-MHC ATP binding site (Jaen-icke et al. 1990) and the mouse elongation factor-1
a
(EF-1
a
)were generated according to standard protocols (Melton etal. 1984). The EF-1
a
probe was isolated from an embryonicmouse cardiac cDNA library and corresponds to the bases230–1319 of EF-1
a
mRNA (accession number X13661). Tocompensate for the differences in length between the fastMHC isoform probes and the slow
b
-MHC probe (414 ntfor the
b
-MHC probe, and 120, 121, and 76 nt for the IIA,IIX, and IIB MHC probes, respectively), the fast MHC iso-form specific probes were double labeled and the exposuretime was twice as long compared to the single labeled
b
-MHC probe, which was exposed for 1 week. The 28SrRNA probe was a single labeled cRNA, transcribed fromthe 930-bp fragment of the mouse 28S rRNA cDNA corre-sponding to bases 3660–4590 of 28S rRNA (accession num-ber X00525). The human
b
-MHC ATP binding site probewas a double labeled cRNA transcribed from the 183-bpfragment of the human
b
-MHC cDNA corresponding to nu-cleotides 460–643 of
b
-MHC mRNA (accession numberM25137). The mouse EF-1
a
probe was a double labeledcRNA transcribed from the 1089-bp fragment of the mouseEF-1
a
cDNA.
In Situ Hybridization
Cryostat sections of whole-muscle preparations and gelatin-embedded single fibers were collected on RNase-free amino-propyltriethoxysilane (AAS)-coated slides and kept at
2
80C.After freeze-drying (
2
60C) for 2–3 hr, the sections werebrought to room temperature (RT), fixed for 20 min in 4%paraformaldehyde (PFA) in PBS, rinsed in PBS, dehydrated
by guest on July 19, 2014jhc.sagepub.comDownloaded from

RNA Content in Single Skeletal Muscle Fibers
997
through graded ethanol steps, and dried in a filtered airstream.
ISH was performed essentially according to Moorman etal. (1993). Briefly, sections were pretreated as follows: 20min 0.2 N HCl, 5 min bidistilled water, 2
3
SSC (SSC is0.15 M NaCl, 0.015 M sodium citrate, pH 7.2) for 10 minat 70C, 5 min in bidistilled water, 20 min digestion at 37Cin 0.1% pepsin dissolved in 0.01 N HCl, 30 sec in 0.2% gly-cine in PBS, twice for 30 sec in PBS, 20 min of postfixationin a 4% freshly made formaldehyde solution for 20 min, 5min in bidistilled water, 5 min in 10 mM EDTA, 5 min in 10mM dithiotreitol (DTT), and finally the sections were driedin a filtered air stream. The hybridization mixture contained50% formamide, 10% dextran sulfate, 2
3
SSC, 2
3
Den-hardt’s solution, 0.1% Triton X-100, 10 mM DTT, 200 ng/
m
l heat-denatured herring sperm DNA. Hybridization wasperformed overnight at 52C and the sections were washedas follows: a rinse in 1
3
SSC, 30 min in 50% formamide/1
3
SSC at 52C, 10 min 1
3
SSC at RT, 30 min at 37C in RNaseA (10
m
g/ml), 10 min in 1
3
SSC, and 10 min in 0.1
3
SSC.The sections were dried, dipped in an autoradiographicemulsion (Ilford Nuclear Research Emulsion G-5), and ex-posed for 2 days (28S rRNA), 1 week (
b
-MHC and MHCATP binding site mRNA), or 2 weeks (IIA, IIX, IIB MHCmRNA and EF-1
a
mRNA). Development time was set at4 min. Controls were performed on serial sections underidentical conditions either in the presence of the sense probeor after pretreatment of the sections with RNase.
Biochemical Determination of Total RNA
In addition to the ISH with the 28S rRNA probe, a biochem-ical method was used to determine the total RNA content.Fragments of histochemically characterized single fiberswere pooled according to their MHC composition (i.e., I,IIA, IIX, and IIB) and the total amount of tissue (200–300
m
g) was determined using a 0.1
m
g rated Sartorius-(S4) su-pramicrobalance. A scaled down version of the orcinol assay(Munro 1966) was used essentially as described by van denHoff et al. (1997). Briefly, the samples were homogenized inice-cold sterile bidistilled water. The nucleic acids were pre-cipitated in 0.5 M HClO
4
and hydrolyzed by incubation in0.3 M KOH at 60C for 1 hr. After removal of DNA by 0.5M HClO
4
precipitation, the samples were boiled for 30 minin 6 M HCl, 0.01% FeCl
3
, and 0.3% orcinol.Optical density (OD) was measured at a wavelength of
660 nm and RNA concentrations were calculated using acalibration curve with total adult liver RNA isolated by ce-sium chloride ultracentrifugation (Chirgwin et al. 1979).
Image Recording
Images of the fiber sections were recorded as described indetail by Jonker et al. (1997). Briefly, a Photometrics cooledCCD camera (Tucson, AZ) attached to an Axioplan micro-scope (Zeiss; Oberkochen, Germany) equipped with a stabi-lized power supply and a monochromatic filter was used torecord the images. The resolution of the images was set at512
3
512 pixels, which corresponds to 175
3
175
m
m
2
inthe object plane at
3
40 magnification. The acquired imageswere converted to OD images using the formula
ODx y, Ix y, IO,x,y⁄log–=
where I is the transmitted light (the acquired image) and I
O
the incident light (the image of the light source).
Image Analysis
Images were analyzed with the software package NIH-Image(v1.61, Rasband, National Institute of Health, Bethesda,MD; a public domain image processing and analysis pro-gram). First, a mask of the cell border was obtained by man-ual tracing of the fiber contour in a serial section used formATPase staining (Figure 1A). As a second step, this con-tour was manually positioned on the corresponding ISH sec-tion of the fiber (Figure 1B). Subsequently, a distance trans-formation was performed from the positioned contourinward to the core of the fiber and outward towards the pe-riphery of the image. This resulted in two distance-trans-formed images combined in Figure 1C.
From both distance-transformed images, consecutivezones of five gray values width (representing steps of 1.65
m
m) were selected. For reasons of clarity, only two zones arehighlighted in Figure 1D. Subsequently, the OD image of theISH section was masked with each zone (Figure 1E; as indi-cated for Figure 1D, masking is shown for only two zones).In this masked selection, the mean OD and the area of eachzone were measured, resulting in a graphic representation ofthe density distribution of each fiber (Figure 1F).
The calculation of the progressive mean OD, starting atthe outermost zone, was used to determine the backgrounddensity level. When the progressive mean OD increased bymore than 5% in one step, the previous calculated mean ODwas considered to be the background (indicated by the grayline in Figure 1F). Measured mean OD values per zone werecorrected by subtraction of the background and multiplica-tion by the area of the zone. The resulting OD values weresummed to calculate the integrated OD (IOD) per fiber.IOD values for each fiber were then divided by cross-sec-tional area to correct for differences in fiber size. The cor-rected IOD is related to the 28S rRNA concentration in thefiber, because IOD is linearly related to the amount of radio-activity bound to the section due to specific hybridization,which in turn is related to the amount of RNA in the section(Jonker et al. 1997).
Statistical Analysis
The data are presented as mean
6
SEM. A Kruskal–Wallisnonparametric one-way analysis of variance (ANOVA) fol-lowed by a multiple comparison of groups was used to testfor differences among the distinct fiber types (Conover1980). The significance level was set at
p
,
0.05.
Northern Blot Analysis
Total RNA was isolated from soleus (more than 90% Type Ifibers) and EDL (more than 90% Type II fibers) muscles ac-cording to Chomczynski and Sacchi (1987). Ten
m
g of eachtotal RNA preparation was electrophoresed, blotted onto anitrocellulose membrane (Hybond; Amersham, Poole, UK),and hybridized using a
32
P-labeled random-primed probe ofthe 183-nt ATP binding site corresponding to nucleotides460–643 of the human
b
-MHC cDNA (accession numberM25137) (Sambrook et al. 1989). After overnight hybrid-ization and washing of the membranes, blots were exposed
by guest on July 19, 2014jhc.sagepub.comDownloaded from

998
Habets, Franco, Ruijter, Sargeant, Sant’Ana Pereira, Moorman
in a Phosphor Image analyzer (Molecular Dynamics; Sunny-vale, CA) and analyzed using the Image Quant 3.3 program(Molecular Dynamics). Ribosomal RNA (18S and 28S)staining was used as the internal standard for the amount ofRNA loaded on the gel.
Determination of Protein Content
Whole-muscle cross-sections of EDL and soleus were used todetermine the protein content by a histochemical method us-ing naphthol Yellow S as described by Tas and James(1981). In addition, EDL and soleus muscle samples werehomogenized in 20 volumes of ice-cold bidistilled water andused for the colorimetric protein detection method based onbicinchoninic acid (BCA-kit; Pierce, Rockford, IL). Bovineserum albumin was used as a standard for the calibrationcurve.
Results
The ISH analysis using the Types I, IIA, IIX, and IIBisoform-specific MHC probe, is shown in Figure 2. A
strong hybridization signal is observed in slow Type Ifibers compared to the fast Type II fibers. Further-more, in the group of Type II fibers the hybridizationsignal is highest in Type IIA fibers and decreases to-wards Type IIX and IIB fibers. Hybridization of thesections to another MHC probe than its nominal one,e.g., fiber Type I to the probe for MHC IIA, IIX, orIIB did not reveal any signal (not shown).
A first conclusion from these observations was thatthe amount of MHC mRNA differs in the separate fi-ber types. This conclusion may imply a difference inMHC mRNA content in fibers that have equal totalRNA content. On the other hand, the observation canequally well be explained by assuming a difference intotal RNA content in fibers that have equal amountsof MHC mRNA per
m
g of total RNA. Obviously, amixture of both possibilities can be envisioned as well.To test whether different amounts of MHC mRNAper
m
g of total RNA are present in slow and fast fi-bers, a Northern blot analysis was carried out. Total
Figure 1 Analysis and quantification of ISH on serial cryostat sections of gelatin-embedded rat single-muscle fibers. A section used formATPase staining was used to trace the fiber contour (A). This contour was positioned manually on the corresponding ISH sections (B). Tak-ing the cell border as a starting point, distance transformations were performed towards the core of the fiber and towards the periphery ofthe image. During this procedure, the distance from the cell border is converted into consecutive gray values (C). Subsequently, successivezones of five gray values width were selected, two of which are highlighted (D). Each zone is masked with the OD image of the ISH section(E; masking is shown only for the same two zones presented in D). Mean OD and area of each zone are calculated and can be representedas a density distribution of the fiber (F), in which the white line represents the border of the cell. The gray and white bars represent theouter zones in D and E. The background level is indicated by the gray line.
by guest on July 19, 2014jhc.sagepub.comDownloaded from

RNA Content in Single Skeletal Muscle Fibers
999
RNA was isolated from soleus (more than 90% Type Ifibers) and EDL (more than 90% Type II fibers)muscle. Equal amounts of RNA were fractionized onsize by gel electrophoresis, blotted onto a nitrocellulosemembrane, and allowed to hybridize to a
32
P-labeledprobe specific for the ATP binding site of human
b
-MHC mRNA. This domain is highly conservedthroughout different myosin isoforms and thereforecan be used as a marker for the total MHC mRNAcontent. Figure 3 shows that the intensity of hybrid-ization was similar with both RNA preparations, indi-cating that equal amounts of MHC mRNA are presentper
m
g of total RNA.The same probe was then used to test whether it
could disclose differences in MHC mRNA content perfiber. To get a first indication of whether such differ-ences would exist, an ISH experiment was performed.Whole-muscle cross-sections from soleus and EDLmuscle were hybridized with the probe to the MHCATP binding site.
A high density of grains can be seen in Type I fibersfrom soleus muscle (Figure 4A) compared to the levelin the EDL muscle (Figure 4C), which hybridizes moreweakly and heterogeneously. This heterogeneity is dueto the presence of some Type I fibers that display highlevels of expression, as revealed by comparing the se-rial sections used for ISH and mATPase histochemis-try (Figures 4B, 4D, and 4E). It is clear that differencesin expression level between Type I and IIA fibers in so-leus muscle (Figure 4A) cannot be reliably discrimi-nated because the few IIA fibers present in this muscleare all surrounded by Type I fibers. In the EDL mus-cle, a distinction can be made because the Type IIA fi-
ber is surrounded by Type IIB fibers, which show avery low expression level (Figure 4C). In agreementwith the hybridization data shown in Figure 2, thesedata support the hypothesis that MHC mRNA con-tent is high in Type I fibers and low in fast fibers. Incombination with the Northern blot data, this impliesthat total RNA content differs in the separate fibertypes. This implication was confirmed by ISH on sec-tions from the EDL and soleus muscle in which acRNA probe for 28S rRNA was used as a marker fortotal RNA. The results observed in both muscle sec-tions are similar to the data described with the MHCATP binding site probe (not shown). To see whethermore general genes that are not tissue specific showsimilar pronounced differences among the distinct fi-ber types, a cRNA probe for elongation factor-1
a
wasused. Whole-muscle cross-sections from EDL and so-leus muscle were hybridized with this probe and ahigh density of grains was observed in the cross-sec-tion of the soleus muscle and in the few Type I fiberspresent in the EDL muscle. However, we were notable to show clear differences between the distinct fastfiber types present within the EDL muscle because ofthe overall low level of expression (not shown).
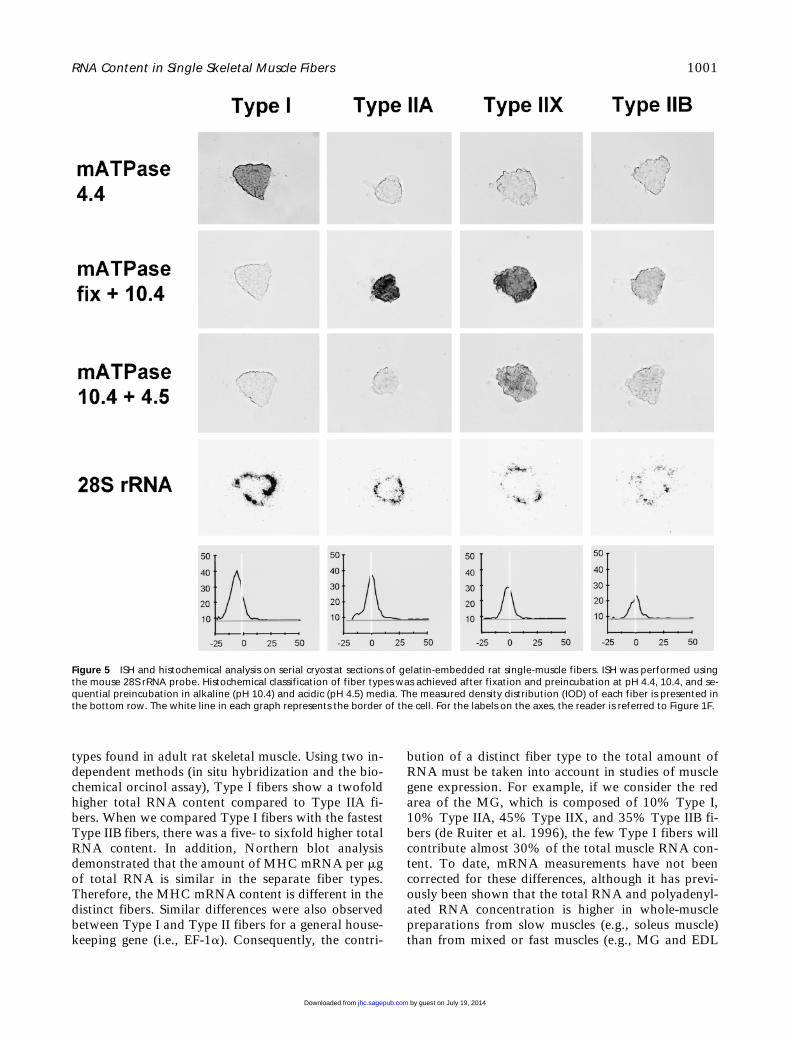
To test directly whether total RNA content differsin the distinct fiber types, a quantitative analysis of to-tal RNA was performed in single muscle fibers by ISHand densitometry in distance zones. As a marker forthe amount of total RNA, we used a cRNA probe for28S rRNA. Fibers were carefully classified as Types I,IIA, IIX, and IIB according to the combined stainingpatterns for the mATPase methods. As shown in Fig-ure 5, only fibers expressing Type I MHC stain afterpreincubation at pH 4.4. Fixation of the fibers fol-lowed by preincubation in alkaline media (pH 10.4)differentiated three fiber populations: Type I fibers(negative), Types IIA and IIX (black), and Type IIB fi-bers (gray). The double preincubation method allowsthe distinction of IIX fibers because this method stainsexclusively fibers expressing IIX MHC. The fourth
Figure 2 Expression of Types I, IIA, IIX and IIB MHC mRNA in his-tochemically characterized single rat muscle fibers by ISH with theisoform-specific MHC probe. A strong hybridization signal can beseen in the Type I fiber, and the signal decreases in the fast fibertypes.
Figure 3 Northern blot analysis using the MHC ATP binding sitecRNA probe in rat soleus and EDL muscle. Total RNA was isolatedfrom both muscles and 10 mg of total RNA was applied in eachlane. Equal amounts of MHC mRNA were observed in Lane 1 (so-leus muscle) compared to Lane 2 (EDL muscle). Ribosomal RNAstaining was used as the internal standard for the amount of RNAloaded on the gel.
by guest on July 19, 2014jhc.sagepub.comDownloaded from

1000
Habets, Franco, Ruijter, Sargeant, Sant’Ana Pereira, Moorman
row in Figure 5 shows the hybridization signal in ex-amples of these four distinct fiber types using the 28SrRNA probe. It is clear that Type I fibers have thestrongest expression of 28S rRNA. Among the fast fi-ber types the expression is highest in Type IIA fibersand decreases in IIX and IIB fibers. Both controls, ISHwith the sense RNA probe and pretreatment withRNase, resulted in nondetectable staining.
Quantification of the density of grains (IOD/area;mean
6
SEM) is summarized in Table 1 and shows asystematic decrease in the amount of rRNA peramount of tissue in the order I
.
IIA
.
IIX
.
IIB. Type Ifibers have twofold higher expression level comparedto Type IIA fibers, and when Type I fibers are com-pared with the fastest Type IIB fibers a five- to sixfold
higher expression level is observed in the Type I fibers.All differences observed were statistically significant(
p
,
0.05).To verify these results with an independent proce-
dure, the amount of total RNA in the distinct fibertypes was determined with the orcinol method (seeTable 1). The highest total RNA content is present inType I fibers. Among the fast fiber types, total RNAcontent is highest in IIA fibers and decreases towardsTypes IIX and IIB fibers, thus corroborating the dataof the 28S rRNA quantification. Furthermore, whenthe total RNA determination for the IIA, IIX, and IIBfiber types is expressed in relation to the value for theType I fibers, very similar values to those demon-strated by the 28S rRNA quantification are revealed(Table 1). The relative RNA content of the orcinolmethod is well within the 95% confidence interval ofthe values based on the ISH.
To assess whether the differences in total RNAlevel between slow fibers and fast fibers are reflectedin similar differences in total protein content, we per-formed the histochemical protein content analysis onsections. No differences among fiber types were de-tected. Subsequently, soleus (predominantly slow fi-bers) and EDL (predominantly fast fibers) muscleswere homogenized and used to determine total proteincontent by the colorimetric procedure. Again, no dif-ferences were found using this method (soleus 20.5
6
1.4
m
g/ml; EDL 20.9
6
1.7
m
g/ml).
DiscussionThe primary focus of this study was to determinewhether the amount of RNA varies among the distinctfiber types (I, IIA, IIX, and IIB) in adult rat skeletalmuscle. To our knowledge, this is the first report pre-senting data at the single-fiber level that quantifies asystematic difference among the four distinct fiber
Figure 4 ISH using the MHC ATP binding site cRNA probe inwhole-muscle cross-sections of soleus (A) and EDL (C) muscle. Type Ifibers in soleus and EDL muscle show a much higher expressionlevel compared to the level in the distinct Type II fibers. Fiber typ-ing was performed by staining transverse serial sections formATPase after preincubations at (B) pH 4.4, (D) pH 4.6, and (E) pH10.4 following formaldehyde fixation.
Table 1 Total muscle fiber RNA content assessed by 28S rRNA in situ hybridization and orcinol assaya
Fiber type
RNA content
28S rRNA in situ hybridization Orcinol method
IOD/areaRelativecontent mg/mg
Relativecontent
I (n 5 6) 861 6 108* 100 6 13 10.36 100IIA (n 5 11) 454 6 51.9** 53 6 4 5.45 53IIX (n 5 11) 327 6 82.5*** 38 6 7 3.67 35IIB (n 5 9) 131 6 29.6 15 6 3 2.07 20
aFor the in situ hybridization, IOD values are corrected for fiber cross-sec-tional area and are presented as mean 6 SEM.*Significantly higher compared to Types IIA, IIX, and IIB fibers; **significantlyhigher compared to Types IIX and IIB fibers; ***significantly higher comparedto Type IIB fibers (p,0.05). For the orcinol, assay total RNA content is ex-pressed as mg/mg dry weight. For both methods, the relative total RNA con-tent (6 SEM) for Types IIA, IIX, and IIB fibers was determined with the meanvalue for the Type I fibers set to 100.
by guest on July 19, 2014jhc.sagepub.comDownloaded from
RNA Content in Single Skeletal Muscle Fibers 1001
types found in adult rat skeletal muscle. Using two in-dependent methods (in situ hybridization and the bio-chemical orcinol assay), Type I fibers show a twofoldhigher total RNA content compared to Type IIA fi-bers. When we compared Type I fibers with the fastestType IIB fibers, there was a five- to sixfold higher totalRNA content. In addition, Northern blot analysisdemonstrated that the amount of MHC mRNA per mgof total RNA is similar in the separate fiber types.Therefore, the MHC mRNA content is different in thedistinct fibers. Similar differences were also observedbetween Type I and Type II fibers for a general house-keeping gene (i.e., EF-1a). Consequently, the contri-
bution of a distinct fiber type to the total amount ofRNA must be taken into account in studies of musclegene expression. For example, if we consider the redarea of the MG, which is composed of 10% Type I,10% Type IIA, 45% Type IIX, and 35% Type IIB fi-bers (de Ruiter et al. 1996), the few Type I fibers willcontribute almost 30% of the total muscle RNA con-tent. To date, mRNA measurements have not beencorrected for these differences, although it has previ-ously been shown that the total RNA and polyadenyl-ated RNA concentration is higher in whole-musclepreparations from slow muscles (e.g., soleus muscle)than from mixed or fast muscles (e.g., MG and EDL
Figure 5 ISH and histochemical analysis on serial cryostat sections of gelatin-embedded rat single-muscle fibers. ISH was performed usingthe mouse 28S rRNA probe. Histochemical classification of fiber types was achieved after fixation and preincubation at pH 4.4, 10.4, and se-quential preincubation in alkaline (pH 10.4) and acidic (pH 4.5) media. The measured density distribution (IOD) of each fiber is presented inthe bottom row. The white line in each graph represents the border of the cell. For the labels on the axes, the reader is referred to Figure 1F.
by guest on July 19, 2014jhc.sagepub.comDownloaded from

1002 Habets, Franco, Ruijter, Sargeant, Sant’Ana Pereira, Moorman
muscle). The reason may be the variability in muscletotal RNA concentrations presented thus far (e.g.,Goldberg 1967; Steffen and Musacchia 1984; Kurowskiet al. 1987; Hood and Simoneau 1989; Choo et al. 1992).
Because the majority of cellular RNA (80–85%) isribosomal, the rRNA content has been used as an in-dex of the protein synthetic capacity of the fiber (Mill-ward et al. 1973). It has been shown at the whole-muscle level that a slow-twitch muscle has a two- tothreefold higher protein synthesis rate than a fast-twitch muscle, which correlates well with the highertotal RNA content (e.g., Goldberg 1967; Flaim et al.1980; Hood and Terjung 1987). However, no data arepresently available with regard to protein synthetic ca-pacity or protein content at the single-fiber level. Oursingle-fiber data show systematic differences in totalRNA content for each fiber type and therefore suggestthat protein synthetic capacity and protein contentwould be highest in Type I fibers and would decreasetowards the faster fiber types. To this end, we deter-mined protein content in whole-muscle preparationsfrom soleus and EDL muscle. Surprisingly, no differ-ences were found with either method. This impliesthat no major differences in MHC protein content ex-ist among different fiber types despite large differencesin MHC mRNA content. This notion is underscoredby single-fiber analysis using SDS-PAGE (Sant’AnaPereira et al. 1995a). Assuming that the rRNA contentreflects the number of ribosomes, slow fibers have asixfold higher capacity to synthesize protein. Becausetotal protein content is not higher, either the potentialcapacity is not used or the protein turnover is higherin Type I fibers.
Because protein synthesis is a major determinant ofenergy turnover in the cell, the oxidative metabolic ca-pability of a fiber can be expected to be related to thetotal RNA content. Among the fast fiber types, a goodrelationship exists between oxidative capacity and to-tal RNA content. However, the correlation is less clearif we compare Type I vs Type II fibers. The highest to-tal RNA content was observed in Type I fibers. In ro-dents, however, succinate dehydrogenase (SDH) activ-ity, which has been used as a marker for oxidativecapability, is slightly higher in Type IIA compared toType I fibers (Schiaffino et al. 1989b; de Ruiter et al.1995; and our unpublished observations). Similarly,mitochondrial content has also been shown to behigher in Type IIA compared to Type I fibers. The ex-planation for this apparent discrepancy may lie in therelative balance between the energy turnover requiredfor protein synthesis and that required for the genera-tion of mechanical output by the different fiber types.Type I fibers, which have slow crossbridge detach-ment rates suited to a postural role, will have a mark-edly lower energy turnover compared with faster fiber
types, which are recruited in locomotive activities re-quiring active shortening and power production (deRuiter et al. 1996). Clearly, the balance of total pos-tural to locomotive activity may vary among speciesand hence may change the total demand for oxidativephosporylation in relation to that proportion attribut-able to protein synthesis.
The mechanism by which a higher total RNA con-tent is achieved can also be related to the number ofmyonuclei per fiber. Several studies demonstrated thatthe amount of myonuclei per mm of myofiber isgreater in Type I fibers compared to Type II fibers(e.g., Schmalbruch and Hellhammer 1977; Tseng et al.1994; Allen et al. 1996), although none of these stud-ies made further subdivision within the Type II fibers.If a correlation with myonuclei number holds true,rRNA content would be dependent on gene doses ratherthan on transcriptional rate or rRNA stability. Furtheranalysis needs to be done to test this possibility.
A second issue that deserves attention is the intra-cellular localization of the RNA. Our analysis of thedistribution of the silver grains reveals a symmetricdistribution of grains, with a peak density just beneaththe sarcolemma (Figures 1F and 5). Given the range ofthe b-particles of 35S (10 mm), we conclude that inadult rat muscle fibers the majority of RNA is subsar-colemmally localized. This conclusion is in line withthe localization of the ribosomes (e.g., Horne andHesketh 1990) and with ISH studies on whole-musclecross-sections (e.g., Dix and Eisenberg 1988; Aignerand Pette 1990). The symmetric distribution of grainsdoes not support an intermyofibrillar localization ofRNA as suggested by a study of Hesketh et al. (1991),who showed by ISH on longitudinal and transversesections of soleus muscle of young rats that the MHCmRNA was distributed throughout the myofibers.They found greater density of grains in the subsar-colemmal regions of the fibers but there was also aconsiderable number of grains in the core myofibrillarregion of the fibers. Their distribution of mRNAmight be a characteristic of immature fibers, and it isconceivable that the subsarcolemmal localization ofRNA in these fibers is acquired with their maturation.
In summary, this study demonstrates for the firsttime that the amount of total RNA per amount of tis-sue varies systematically among Types I, IIA, IIX andIIB fibers in rat hindlimb muscles. ISH on sections ofsingle fibers followed by quantification and indepen-dent biochemical method both demonstrate that theamount of total RNA decreases significantly fromI.IIA.IIX.IIB; slow Type I fibers show a five- tosixfold higher expression level compared to the fastestType IIB fibers. The MHC mRNA content per mg oftotal RNA is shown to be comparable in all fibertypes. Therefore, Type I fibers also contain five- to
by guest on July 19, 2014jhc.sagepub.comDownloaded from

RNA Content in Single Skeletal Muscle Fibers 1003
sixfold more MHC mRNA per mg of tissue comparedto the fastest Type IIB fibers. This implies that propermRNA quantification in slow vs fast muscle fibers andalso among fast Type II fibers requires correction forthe total amount of RNA per fiber type.
AcknowledgmentsSupported in part by NWO Grant number 902-16-219
(to DF). We gratefully acknowledge Piet de Boer for help with in
situ hybridization, J. Hagoort for image aquisition, Cars E.Gravemeijer and Cees J. Hersbach for excellent photogra-phy, and Dr Stefano Schiaffino and Dr Margaret Bucking-ham for kindly providing the IIA, IIX, and IIB MHCcDNAs.
Literature CitedAigner S, Pette D (1990) In situ hybridisation of slow myosin heavy
chain mRNA in normal and transforming rabbit muscles with theuse of non-radioactively labeled cRNA. Histochemistry 95:11–18
Allen DL, Yasui W, Tanaka T, Ohira Y, Nagaoka S, Sekiguchi C,Hinds WE, Roy RR, Edgerton VR (1996) Myonuclear numberand myosin heavy chain expression in rat soleus single muscle fi-bres after spaceflight. J Appl Physiol 81:145–151
Boheler KR, Chassagne C, Martix X, Wisnewsky C, Schwartz K(1992) Cardiac expression of a and b myosin heavy chains andsarcomeric a actins are regulated through transcriptional mecha-nisms. J Biol Chem 267:12979–12985
Caiozzo VJ, Haddad F, Baker MJ, Baldwin KM (1996) Influence ofmechanical loading on myosin heavy chain protein and mRNAexpression. J Appl Physiol 80:1503–1512
Caiozzo VJ, Ma E, McCue SA, Smith E, Herrick RE, Baldwin KM(1992) A new model for modulating myosin isoform expressionby altered mechanical activity. J Appl Physiol 73:1432–1440
Chirgwin JM, Przybyla AE, MacDonald J, Ruiter WJ (1979) Isola-tion of biologically active ribonucleic acid from sources enrichedin ribonuclease. Biochemistry 18:5294–5299
Chomczynski P, Sacchi N (1987) Single-step method of RNA isola-tion by acid guanidinium thiocyanate-phenol-chloroform extrac-tion. Anal Biochem 162:156–159
Choo JJ, Horan MA, Little RA, Rothwell NJ (1992) Anabolic effectsof clenbuterol on skeletal muscle are mediated by beta 2-adreno-ceptor activation. Am J Physiol 263:E50–56
Conover WJ (1980) Practical Nonparametric Statistics. 2nd ed.New York, Wiley
De Nardi C, Ausoni S, Moretti P, Gorza L, Velleca M, BuckinghamM, Schiaffino S (1993) Type IIX myosin heavy chain is coded bya muscle fibre type-specific and developmentally regulated gene.J Cell Biol 123:823–835
De Ruiter CJ, de Haan A, Sargeant AJ (1995) Physiological charac-teristics of two extreme muscle compartments in gastrocnemiusof the anaesthetized rat. Acta Physiol Scand 153:313–324
De Ruiter CJ, Habets PEMH, de Haan A, Sargeant AJ (1996) Invivo IIX and IIB fiber recruitment in gastrocnemius muscle of therat is compartment related. J Appl Physiol 81:933–942
Dix DJ, Eisenberg BR (1988) In situ hybridisation and immunocyto-histochemistry in serial sections of rabbit skeletal muscle to de-tect myosin expression. J Histochem Cytochem 36:1519–1526
Flaim K, Copenhower ME, Jefferson LS (1980) Effects of diabeteson protein synthesis in fast- and slow-twitch rat skeletal muscle.Am J Physiol 239:E88–95
Goldberg AL (1967) Protein synthesis in tonic and phasic skeletalmuscles. Nature 216:1219–1220
Hesketh J, Campbell G, Loveridge N (1991) Myosin heavy–chain
mRNA is present in both myofibrillar and subsarcolemmal re-gions of muscle fibres. Biochem J 279:309–310
Hood DA, Simoneau J (1989) Rapid isolation of total RNA fromsmall mammal and human skeletal muscle. Am J Physiol 256: 1029–1096
Hood DA, Terjung RL (1987) Leucine metabolism in perfused ratskeletal muscle fibres during contractions. Am J Physiol253:E636–647
Horne Z, Hesketh J (1990) Immunological localisation of ribo-somes in striated rat muscle. Biochem J 268:231–236
Jaenicke T, Diederich KW, Haas W, Schleich J, Lichter P, Pfordt M,Bach A, Vosberg HP (1990) The complete sequence of the hu-man beta–myosin heavy chain gene and a comparative analysisof its product. Genomics 8:194–206
Jonker A, de Boer PAJ, van den Hoff MJB, Lamers WH, MoormanAFM (1997) Towards quantitative in situ hybridization. J His-tochem Cytochem 45:413–423
Kurowski TT, Zak R, Hickson R (1987) Development of glucocor-ticoid-induced growth and atrophy in heart and skeletal muscle.J Cell Biol 105:11a
Mahdavi V, Izumo S, Nadal–Ginard B (1987) Developmental andhormonal regulation of sarcomeric myosin heavy chain genefamily. Circ Res 60:804–814
Melton DA, Krieg PA, Rebagliati MR, Maniatis T, Zinn K, GreenMR (1984) Efficient in vitro synthesis of biologically active RNAand RNA hybridisation probes from plasmids containing a bac-teriophage SP6 promoter. Nucleic Acids Res 12:7035–7056
Millward DJ, Garlick PJ, James WPT, Nnanyelugo DO, Ryatt JS(1973) Relationship between protein synthesis and RNA contentin skeletal muscle. Nature 241:204–205
Moorman AFM, de Boer PAJ, Vermeulen JLM, Lamers WH (1993)Practical aspects of radio-isotopic in situ hybridization on RNA.Histochem J 25:251–266
Moss RL, Diffee GM, Graeser ML (1995) Contractile properties ofskeletal muscle fibres in relation to myofibrillar protein isoforms.Rev Physiol Biochem Pharmacol 126:1–63
Munro NH (1966) The determination of nucleic acids. MethodsBiochem Anal 14:113–176
Pette D, Staron RS (1990) Cellular and molecular diversities ofmammalian skeletal muscle fibres. Rev Physiol Biochem Pharma-col 116:1–76
Sambrook J, Fritsch JE, Maniatis T (1989) Molecular Cloning: ALaboratory Manual. Cold Spring Harbor, NY, Cold Spring Har-bor Laboratory
Sant’Ana Pereira JAA, de Haan A, Wessels A, Moorman AFM,Sargeant AJ (1995b) The mATPase histochemical profile of rattype IIX fibres: correlation with MyHC immunolabelling. His-tochem J 27:715–722
Sant’Ana Pereira JAA, Ennion S, Sargeant AJ, Moorman AFM,Goldspink G (1997) Comparison of the molecular, antigenic andATPase determinants of fast MyHCs in rat and humans: a singlefibre study. Pflugers Arch Eur J Physiol 435:151–163
Sant’Ana Pereira JAA, Wessels A, Nytmans L, Moorman AFM,Sargeant AJ (1995a) New method for the accurate characterisa-tion of single human fibres demonstrates a relation betweenmATPase and MyHC expression in pure and hybrid fibre types. JMuscle Res Cell Motil 16:21–34
Schiaffino S, Gorza L, Sartore S, Saggin L, Ausoni S, Vianello M,Gundersen K, Lomo T (1989b) Three myosin heavy chain iso-forms in Type 2 skeletal muscle fibres. J Muscle Res Cell Motil10:197–205
Schiaffino S, Reggiani C (1994) Myosin isoforms in mammalianskeletal muscle. J Appl Physiol 77:493–501
Schiaffino S, Reggiani C (1996) Molecular diversity of myofibrillarproteins: gene regulation and functional significance. Physiol Rev76:371–423
Schiaffino S, Samuel JL, Sassoon D, Lompré AM, Garner I, MarotteF, Buckingham M, Rappaport L, Schwartz K (1989a) Nonsyn-chronous accumulation of a-skeletal actin and b-myosin heavychain mRNAs during early stages of pressure-overload-induced
by guest on July 19, 2014jhc.sagepub.comDownloaded from

1004 Habets, Franco, Ruijter, Sargeant, Sant’Ana Pereira, Moorman
cardiac hypertrophy demonstrated by in situ hybridization. CircRes 64:937–948
Schmalbruch H, Hellhammer U (1977) The number of nuclei inadult rat muscles with special reference to satellite cells. Anat Rec189:169–176
Steffen JM, Musacchia XJ (1984) Effect of hypokinesia and hypo-dynamia on protein, RNA and DNA in rat hindlimb muscles.Am J Physiol 247:R728–732
Swyngedauw B (1980) Developmental and functional adaptationsof contractile proteins in cardiac and skeletal muscles. PhysiolRev 66:710–771
Tas J, James J (1981) Quantitative cytochemistry of nuclear and cy-toplasmic protein using the naphthol Yellow S and dinitrofluo-robenzene staining methods. Histochem J 13:711–716
Tseng BS, Kasper CE, Edgerton VR (1994) Cytoplasm-to-myonu-cleus ratios and succinate dehydrogenase activities in adult ratslow and fast muscle fibres. Cell Tissue Res 275:39–49
van den Hoff MJB, Lekanne Deprez RH, Monteiro M, de Boer PAJ,Charles R, Moorman AFM (1997) Developmental changes in ratcardiac DNA, RNA and protein tissue base: implications for theinterpretations of changes in gene expression. J Mol Cell Cardiol29:629–639
by guest on July 19, 2014jhc.sagepub.comDownloaded from
Copyright © 2022 FDOKUMEN




















