
River Fisheries.pdf - EIS
359
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/44535866 River fisheries / by R. L. Welcomme Article Source: OAI CITATIONS 2 READS 2,525 1 author: Some of the authors of this publication are also working on these related projects: I am retired from active work but am still continuing with reviewing and acting as Assistant Editor to Fisheries Management and Ecology View project Robin L. Welcomme Imperial College London 120 PUBLICATIONS 6,180 CITATIONS SEE PROFILE All content following this page was uploaded by Robin L. Welcomme on 25 June 2014. The user has requested enhancement of the downloaded file.
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of River Fisheries.pdf - EIS

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/44535866
River fisheries / by R. L. Welcomme
Article
Source: OAI
CITATIONS
2READS
2,525
1 author:
Some of the authors of this publication are also working on these related projects:
I am retired from active work but am still continuing with reviewing and acting as Assistant Editor to Fisheries Management and Ecology View project
Robin L. Welcomme
Imperial College London
120 PUBLICATIONS 6,180 CITATIONS
SEE PROFILE
All content following this page was uploaded by Robin L. Welcomme on 25 June 2014.
The user has requested enhancement of the downloaded file.

1
FAO FISHERIES TECHNICAL PAPER 262
RIVER FISHERIES
.
by
R.L.Welcomme Senior Fishery Resources Officer
FAO Fishery Resources and Environment Division
The designations employed and the presentation of material in this publication do not imply the
expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the
United Nations concerning the legal status of any country, territory, city or area or of its authorities, or
concerning the delimitation of its frontiers or boundaries.
M-42
ISBN 92-5-102299-2
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or
transmitted in any form or by any means, electronic, mechanical, photocopying or otherwise, without
the prior permission of the copyright owner. Applications for such permission, with a statement of the
purpose and extent of the reproduction, should be addressed to the Director, Publications Division,
Food and Agriculture Organization of the United Nations, Via delle Terme di Caracalla, 00100 Rome,
Italy.
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS Rome, 1985
© FAO
PREPARATION OF THIS DOCUMENT
This technical paper presents an updating and extension of material already published externally to
FAO in ‘Fisheries Ecology of Floodplain Rivers’ (Longman, 1979). It is intended as a general
summary of the current thinking based upon the literature on all aspects of river fisheries from the

2
physical and biological environment in which they are pursued to their management. As such it is of
interest not only to students, scientists and administrators working in the fisheries sector, but also to
biologists, ecologists and geographers working on aquatic organisms other than fish and more general
aspects of natural resources development and management.
The cover illustration shows a portion of the floodplain of the Magdalena River and illustrates the
complexity of the aquatic system in some floodplains.
Distribution:
FAO Fisheries Department
FAO Fisheries Regional Offices
Selector SI
CIFA
COPESCAL
EIFAC
For bibliographic purposes this document
should be cited as follows
Welcomme, R.L., River fisheries. FAO Fish.
1985 Tech.Pap., (262): 330 p.
SUMMARY
Rivers drain all but the most arid areas of the earth through channels that are regulated by physical laws that
impose on them certain forms. The ideal form is rarely encountered in practice and represents an end point to
which geographic process tend. In general a river may be divided into two principal zones, the steep and fast
flowing rhithron upstream and the sluggish and flat potamon downstream. While conditions in an individual
system are highly variable along its length, similar reaches of different rivers differ much less even between
continents and at different latitudes. All continents have a series of major river systems which consist not only
of the river channels but also the swamps, lakes and seasonally flooded lands associated with them.
Most rivers are highly conditioned by the patterns of precipitation in their basins. Differences in rainfall
intensity throughout the year generate a flood wave that progresses downstream in the majority of rivers (flood
rivers), although singular geographic circumstances may distribute discharge more evenly throughout the year in
some systems (reservoir rivers). The number of reservoir rivers is increasing through flow regulation and dam
building. Although the basic nature of the river is determined by the rocks over which it flows, the flood regime
seasonally modifies the physical and chemical conditions within the river particularly in the tropics. In higher
latitudes other features of climate, such as insolation or air temperature exert an increasing influence.
Seasonal changes in discharge, nutrient concentrations, pH, temperature and dissolved oxygen in their turn
influence the composition and abundance of the plant and animal communities inhabiting the river. These
changes are particularly marked in the floodplain area of the potamon where the rise in water during the floods
inundates extensive areas of land flanking the main channels. This increase in living space, together with the
release of nutrients associated with the submersion of the soil produces an annual surge of primary productivity
closely followed by an expansion in biomass of animal communities.
The number of fish species inhabiting rivers is a function of the size of the river, with larger basins such as
the Amazon having well over 1000 species. The individual species are highly adapted to the conditions in the
type of river reach in which they live. Such adaptations are not only morphological but also behavioural and
some species have developed extensive migrations to avoid adverse conditions or for breeding and feeding.
Alternatively elaborate breeding mechanisms have also evolved.
Other features of the biology of fishes are linked to the hydrological cycles within the river. Thus, the flood is
associated with spawning in the majority of species when the abundance of living space and food provides the
best conditions for the survival and growth of the young fish. Such is the influence of these factors that in years
of more intense flooding survival and growth are so improved that the total biomass of the fish community rises
and a strong year class is produced for transmission on to other years. In reservoir rivers and in the rhithron
seasonal and year-to-year differences of this type are not so marked.

3
Fish communities in rivers provide the basis for fisheries which are pursued with a great variety of gear.
Fishing intensity is also seasonal and is tied either to variations in temperature or to the flood. Because the fish
community can vary in abundance with the fluctuations in flood strength catch is similarly correlated with years
of high catch following after years of particularly good flooding. Fish communities respond to increases in
fishing pressure in a number of ways. In general catch in rivers having simple fish communities follow a typical
yield curve whereas those in which communities are complex show a plateau in catch which may persist over a
great range of effort. This plateau masks changes in the composition of the community with a drift from large,
slow growing to small, fast growing forms.
Rivers and their basins are used for many purposes other than fisheries. Many of these modify the quality or
quantity of water in the system and thus interact with the fish communities in the river to their detriment.
Management of the river for fisheries therefore becomes increasingly important as the intensity of use of the
river rises. Fisheries themselves also require management which may be accomplished either by direct
interventions on the fish stock or by legislative or economic activities on the fishermen themselves. As the river
becomes increasingly modified the capture fisheries originally pursued there become less viable. Although the
water courses may continue to provide food or recreation the major developmental emphasis is towards
replacement activities such as aquaculture on the former floodplain or the creation of new fisheries in the
reservoirs associated with the main channel.
Hyperlinks to non-FAO Internet sites do not imply any official endorsement of or responsibility for
the opinions, ideas, data or products presented at these locations, or guarantee the validity of the
information provided. The sole purpose of links to non-FAO sites is to indicate further information
available on related topics.
CONTENTS
INTRODUCTION
Chapter 1 - MORPHOLOGY OF RIVER SYSTEMS
FORM OF RIVER SYSTEM
TYPES OF RIVER
LONGITUDINAL PROFILE
TRIBUTARY DEVELOPMENT AND STREAM ORDER
Stream Order
Relation of River Length to Drainage Basin Area
MORPHOLOGY OF THE RHITHRON
MORPHOLOGY OF THE POTAMON
The Channels
The Floodplain
BRIEF REVIEW OF MAJOR RIVERS
AFRICA
AMERICA (NORTH)

4
AMERICA (SOUTH)
ASIA
EUROPE
AUSTRALIA AND NEW ZEALAND
Chapter 2 - PHYSICAL AND CHEMICAL PROCESSES
HYDROLOGY
FLOW
Variability of Flow Regimes
Velocity of Flow
ORIGINS OF FLOODING
WATER BALANCE ON FLOODPLAINS
CHEMISTRY
SEDIMENT LOAD AND TURBIDITY
NUTRIENT SPIRALLING
CARBON AND ORGANIC MATERIAL
IONIC COMPOSITION
HYDROGEN ION CONCENTRATION (pH)
TEMPERATURE
DISSOLVED OXYGEN
Chapter 3 - PRIMARY PRODUCTION IN RIVERS
THE RIVER CONTINUUM CONCEPT
MATERIAL OF ALLOCHTHONOUS ORIGIN
BACTERIA AND OTHER MICROORGANISMS
PHYTOPLANKTON
INFLUENCE OF ENVIRONMENTAL FACTORS
PRODUCTION
ATTACHED ALGAE
HIGHER VEGETATION
DISTRIBUTION AND ZONATION
SUBMERSED VEGETATION

5
FLOATING VEGETATION
FLOATING MEADOWS
FLOODPLAIN MEADOWS
THE ROLE OF HIGHER VEGETATION IN NUTRIENT BALANCE
Chapter 4 - SECONDARY PRODUCTION IN RIVERS
ZOOPLANKTON AND DRIFT
ANIMAL COMMUNITIES ASSOCIATED WITH FLOATING AND SUBMERSED
VEGETATION
BENTHOS
NEUSTON
VERTEBRATES OTHER THAN FISH
AMPHIBIA
REPTILES
BIRDS
MAMMALS
Chapter 5 - RIVER FISHES AND THE RIVERINE SYSTEM
NUMBERS OF SPECIES IN RIVER SYSTEMS
DIFFERENCES IN NUMBERS OF SPECIES BETWEEN SYSTEMS
RELATIVE ABUNDANCE OF SPECIES WITHIN ONE SYSTEM
SUB-POPULATIONS
SIZE OF SPECIES
DISTRIBUTION OF SPECIES IN RIVER SYSTEMS
DISTRIBUTION IN SPACE AND ZONATION
Zonation
Habitats of River Systems and Accompanying Floodplain
DISTRIBUTION IN TIME AND MIGRATION
Types of Migration and Movement
Migrations of Adult Fish
Movement of Juveniles
Distance and Speed of Movement
Timing of Migration

6
ADAPTATION TO EXTREME ENVIRONMENTAL CONDITIONS
LOW DISSOLVED OXYGEN CONCENTRATIONS
Adaptations to Air Breathing
Adaptations for Using the Surface Layer
Physiological Adaptations
ADAPTATIONS TO RESIST HIGH TEMPERATURE
ADAPTATIONS TO RESIST DESICCATION
ADAPTATIONS TO POOR LIGHT
ADAPTATIONS TO RESIST STRONG CURRENT
Chapter 6 - THE PRODUCTION BIOLOGY OF RIVER FISH
FEEDING
SOURCES OF FOOD
SPECIALIZATION AND RESOURCE PARTITIONING
Specialization
Resource Partitioning
SEASONALITY OF FEEDING
GROWTH
FACTORS AFFECTING GROWTH
MODELS OF GROWTH
YEAR-TO-YEAR VARIATIONS IN GROWTH
REPRODUCTION
SPAWNING SITES AND REPRODUCTIVE ADAPTATIONS
FECUNDITY AND SPAWNING PATTERNS
TIMING
THE INFLUENCE OF HYDROLOGICAL REGIME ON SPAWNING SUCCESS
MORTALITY
CAUSES OF MORTALITY
SEASONALITY OF MORTALITY
ESTIMATES OF MORTALITY RATES
MODELS OF MORTALITY

7
STANDING STOCK AND PRODUCTION
STANDING STOCK
Main River Channel
Backwaters
Standing Waters of Floodplains
Total System
PRODUCTION
Estimates of Production
Models of Fish Standing Stock and Production in Rivers
Chapter 7 - THE FISHERY
THE FISHERMEN
OCCASIONAL FISHERMEN
PART-TIME FISHERMEN
PROFESSIONAL FISHERMEN
FISHING GEAR
SEASONALITY
CAPTURE METHODS
Low Water
Fishing during Rising and Receding Floods
Fishing at Peak Flood
BOATS
PRESERVATION OF FISH
TYPES OF PRODUCT
Chilled Fish
Dried Fish
Salting
Smoking
Fish Meal
Fish Oil
Fermented Products

8
PROTECTION AGAINST INSECT INFESTATION
SPECIES CAUGHT
LATIN AMERICA
AFRICA
ASIA
EUROPE AND NORTH AMERICA
FISHERIES FOR JUVENILE FISH
CATCH
ANALYSIS OF CATCH IN DIFFERENT RIVERS
Catch as a Function of River Form
Catch as a Function of Fishing Intensity
Changes in Community Structure with Increasing Fishing Pressure
FLUCTUATION IN CATCH BETWEEN YEARS
Chapter 8 - MANAGEMENT OF RIVER FISHERIES
EFFECTS OF OTHER USES OF RIVERS AND THEIR BASINS ON FISHERIES
CHANGES IN FLOW
CHANGES IN SILT LOAD
CHANGES IN WATER QUALITY
INTERACTION WITH OTHER USES
Terrestrial Animals
Forestry
Agriculture
Urbanization
Hydraulic Engineering
DEVELOPMENT AND MANAGEMENT OF RIVER SYSTEMS FOR FISHERIES
OBJECTIVES AND STRATEGY
Objectives
Uses of Fish Resources
Developmental Stage
MANAGEMENT OF THE RIVERINE ENVIRONMENT

9
Preservation of the Natural System
Instream Improvement Structures
MANAGEMENT OF THE FISH STOCK
Introduction of New Species
Stocking
MANAGEMENT OF THE FISHERY
Regulation of Access
Increasing the Catch Capacity of Fishermen
Closed Seasons
Reserved Waters
Mesh Regulations
Banning of Certain Gears
Accessibility
DEVELOPMENT OF NEW OR ALTERNATIVE FISHERIES
Aquaculture
Reservoir Fisheries
ACKNOWLEDGEMENTS
BIBLIOGRAPHY
SPECIES INDEX
GEOGRAPHICAL INDEX
GENERAL INDEX

10
INTRODUCTION
Rivers have formed nuclei for human settlement from the origins of mankind. Many of the earliest
civilizations emerged upon the fertile floodplains and since about 5000 b.p., when the earliest
systematic colonization of the Nile, Mesopotamic, Indus and Chinese rivers occurred there has been a
concentrated effort aimed at the domination of the hydro-logical regimes for the benefit of agriculture.
The Roman culture and later that of Western Europe impounded many smaller rivers for water power.
In many of the more arid parts of the world streams and rivers were manipulated to provide water for
irrigation. These trends have increased until the present day efforts at impoundment, deviation and
canalization have left few rivers with undisturbed channels. To the environmental impacts of
hydraulic engineering must be added the hazards of the contamination of the waters with a variety of
agricultural, domestic and industrial chemicals. In addition bad, or non- existent basin management,
deforestation and farming of marginal hill slope lands has increased erosion and the silt loads of rivers
resulting in rapid modification of the lowland reaches of the river. These changes not only modify the
environment, depriving the fish of living space and access to parts of the river necessary for the
completion of their life cycles, but also changes in the quality and quantity of the water in which they
live.
The fish communities of rivers have provided the basis for fisheries presumably from the earliest
phases of human occupation of river valleys. The FAO Yearbook of Fishery Statistics (FAO 1984) the
shows the nominal catch of the worlds freshwaters to have grown from 7.1 million tons in 1977 to 8.9
million tons in 1983 representing 10.4 and 11.6 percent of total world catch respectively. The
relatively slow rate of increase of 3 percent per year would seem to indicate that catch levels are
reaching a maximum. Much of the present inland catch still derives from rivers or the seasonally
inundated ground associated with them especially in Latin America and South East Asia where large
lakes are rare. The increase in the number of uses competing with fisheries has brought about the
disappearance of some long established fisheries and others are on their way to extinction.
In general studies of fish communities have lagged behind those of lakes and reservoirs although there
has been an increase in interest in this topic in the last decade. Practical concern with the management
of rivers for fisheries began towards the end of the last century in North America and Europe and led
to research in such waters in support of stocking and physical improvement programmes principally
in support of sport fisheries for salmonids. These led to early classifications of rivers into zones in
Eastern Europe where commercial fisheries for coarse fish were also economically important. One of
the earliest systematic studies on large rivers were those of Antipa (1910) whose original work on the
Danube was continued by other workers until it has become one of the most extensively studied of the
world’s major rivers. Antipa's general conclusion that the fisheries production of the Danube was
directly proportional to the extent and duration of flooding in any particular year (Botnariuc, 1968)
has proved to be equally applicable to all other flood rivers investigated. The work on the Danube also
illustrated that the floodplain cannot be considered in isolation but must be treated as an integral part
of the larger system (Botnariuc, 1967; Balon, 1967). Somewhat later Russian workers commenced
studies on the Volga River although this work intensified only after the creation of the cascade of
reservoirs in that system and only part of the literature is available in translation. Detailed studies of
the Mississippi-Missouri system were further delayed and it is only in the last decade that
understanding of that system has been obtained. Modern work on these and other temperate rivers is
now highly complex and deals with many biological and ecological issues especially those connected
with the conservation of riverine habitats.
Systematic study of the fisheries ecology of tropical rivers began on the Niger when a laboratory was
set up in the Central Delta (Blanc et al., 1955) whose output through the numerous publications of
Daget clarified much of the taxonomy and biology of fish in that river several FAO projects have been
associated with the study of this river in Niger, Benin and Nigeria. The Nile Sudd in Sudan has been
studied by a series of missions including the Jonglei Investigation Team (1954) and Mefit Babtie
(1984). Intensive but short term duration studies on the Kafue River by the Universities of Idaho and

11
Michigan did much to shed light on the biology of the fishes of the Kafue flats. The ORSTOM team
studied the Yaeres floodplain of the Logone River during the 1970's and elsewhere workers have been
gathering information on the fisheries and general ecology of the Shire River, the Okavango delta
(Botswana Society, 1976) and components of the Congo River.
In South America most of the river systems have been examined to a certain extent. The numerous
works of Bonetto and his team have provided a great amount of information on the Parana River and
its tributaries while Godoy (1975) summarized an extensive amount of work on the Brazilian Mogi
Guassu tributary of the same system. In the Amazon staff of the Instituto Nacional de Pesquisas
Amazonicas has studied the area around Manaus and an FAO funded project enabled the Peruvian
authorities to collect information on the same river at the level of Iquitos. Work on the Orinoco was
begun by Mago-Leccia (1970) and has been continued by Novoa and his co-workers. Lowe-
McConnell carried out fundamental studies on the ecology of tropical river fish communities in the
Rupununi River. Surveys of the fisheries of the Magdalena River were started by INDERENA and
intensified through the activities of an FAO project.
Studies on Asiatic rivers have been somewhat more limited although work on the Mekong from
Chevey et Le Poulain (1940) onwards have contributed to the general understanding of large tropical
systems. Otherwise occasional studies have been carried out on rivers in Peninsular Malaysia, Borneo,
India, Sri Lanka and the Mesopotamic rivers. Doubtless there is abundant literature on the major
Chinese systems but this is not available in translation at this time although some Soviet studies on the
Amur River have been published.
All these works and many others combine to give a body of information on the fish and fisheries of
the world as presented in this Technical Paper. This information indicates that while there is a
considerable amount of diversity in rivers, some general conclusion can be drawn. Firstly, although
potamon and rhithron differ considerably the form and behaviour of these two major river zones
appears consistent irrespective of continent or latitude. In other words rhithron reaches resemble one
another wherever they are as do potamon reaches and each may be considered as forming a set
permitting information and data to be pooled irrespective of geographic origin. Secondly, the
dynamics and behaviour of fish communities in the potamon of flood rivers is different from those of
reservoir rivers (and those whose flows have been artificially modified by man). Thirdly, the biology
and ecology of many of the fishes in flood river is so finely attuned to the seasonal flood that
modifications to the hydrological regime produce changes in the composition and productivity of the
fish community consistent with the conversion from flood to reservoir conditions.
This technical paper summarizes the present state of knowledge on the fish and fisheries of river
systems although certain limitations are imposed by the need to contain the size of the document. It
emphasizes the role of rivers as food producers and, although other uses of the fish communities are
referred to, detailed analysis of the extensive literature on sport fishery management and practice is
omitted. The first section deals with the physical and chemical environment and briefly summarizes
those aspects of primary and secondary productivity of rivers in so far as they are significant for
fisheries. A complete treatise on the limnology of running waters is inappropriate here and would
require a volume this size for this topic alone. The analysis of the biology and ecology of the fish
concentrates on larger rivers as these are the locations of the major fisheries and, although mention is
made of the biology of rhithronic communities, the extensive literature on salmonid rivers is not
developed to the full as this has been published elsewhere. Consideration of the fisheries of rivers is
of necessity speculative as the quantity and quality of the statistical data available impose limits on
the depth of analysis. Nevertheless fishery scientists are increasingly called upon to make rapid
proposals for management strategies or evaluations of possible impacts of interventions by other users
within the river basins so some generalized models are needed. Clearly such models must be open to
criticism and eventual modification in the light of future experience. Similarly, the section on
management tends to examine the effects of management on the fish community and does not explore
fully the social and economic implications of the various approaches to the regulation and

12
improvement of fisheries in rivers, a more detailed treatment of these topics being found in more
specialized works.
CHAPTER 1
MORPHOLOGY OF RIVER SYSTEMS
FORM OF RIVER SYSTEMS
TYPES OF RIVER
Rivers are linear systems which serve to evacuate water falling on the continental masses towards the
oceans. This transfer involves the dissipation of the kinetic energy inherent in the water and the
morphology of river channels evolves so as to even out the loss of this energy along the length of the
river. The hydraulic processes arising from this loss act in a predictable manner within the river
channel, thus the forms taken by the various rivers of the world are closely comparable where
conditions of bed-rock, elevation and rainfall are similar (Leopold et al., 1964). In effect, greater
differences exist between the various zones of one river than between homologous zones of different
rivers. Thus biological studies on rivers tend to treat sub-sets of river systems, such as “trout streams”
or “potamon reaches”, rather than to consider the system as a whole from headwater to mouth.
However, such subdivisions are for convenience of study only and any river system should ultimately
be viewed as a continuum showing a succession of characters along its length.
Features of the geography of any particular river basin can impose certain characteristics on the river.
One useful distinction is between: (i) reservoir rivers, which have extensive lakes, swamps or
floodplains near their headwaters resulting in the gradual release of floodwaters and sustained flow
with only slight variations in rate; and (ii) flood rivers, where there are extremes of annual fluctuation
in water level from severe flood to sometimes complete desiccation in the dry season. The most
extreme form of flood rivers, i.e. those which frequently cease to flow or even dry out seasonally,
have been termed “sandbank” rivers by Jackson (1961a and 1963).
A second distinction originates from the type of landscape through which the river flows. Here (i)
tropical forest rivers have many of the characteristics of reservoir rivers in that variations of flow are
evened out by the retention of water in the flooded forest. Such rivers tend to have black waters with
low pH, low conductivity and ionic content, low silt load and high humic content; (ii) Savannah rivers
may be of either sandbank or flood type, depending on the form of their basins. The pH of their waters
is rarely extreme, varying from slightly acid to slightly alkaline, conductivities are often reasonably
high as are silt loads; (iii) Desert rivers, which receive no tributaries in their dry land course, tend to
conform to the flood type. They show greatly increased conductivity and alkalinity along their lengths
as the water becomes concentrated by evaporation, and in their more extreme form end up as salt
marsh or lake; (iv) Tundra rivers drain the arctic and sub-arctic regions northwards and tend to have
flow regimes that are dependent on winter freezing. Their ionic contents are frequently poor as the
lands over which they flow were denuded of topsoil during the glaciations. Mixed systems also occur,
and larger rivers, especially, may change their nature several times during their course. Equally,
developments within their basin may change what were once forest rivers into savannah rivers, and
eventually by erosion, siltation and water use into desert rivers.
Considerable modifications have been carried out in many river systems, particularly in the temperate
zone where there are few large water courses which now show all their original features, and the

13
numbers of reservoir rivers, in the form of regulated streams, are steadily increasing throughout the
world due to interventions in the aquatic system aimed at controlling river flow.
LONGITUDINAL PROFILE
Rivers tend to have longitudinal profiles which are concave open to the sky (Fig. 1.1). This means that
within any one river, there is typically a succession of types of water course with steep slopes near the
source to minimal slope near the mouth. This succession is by no means always adhered to, and many
major rivers, through accidents of terrain, alternate between fast-flowing, rocky and slow-flowing,
muddy stretches. Thus, after torrential upper courses, rivers such as the Niger (Fig. 1.1), the Congo or
the Danube show several successive reaches of floodplain and rapids along their length. The different
types of water course plainly support different communities of living organisms and this has formed
the basis for several systems of geographical and ecological zoning. Geographically, such zoning is
reflected in everyday speech, which distinguishes between a variety of types of water course,
including torrents, creeks, brooks, streams, rivers, etc. Ecologically, too, such distinctions have value
as they generally correspond to many differing conditions including flow, slope or bottom type, which
determine the types of plant and animal community living in them. Because of the variety of types of
flowing water that are recognized, it is not possible to produce a comprehensive classification of
rivers that is ecologically satisfying, although some authors such as Petts (1984) have recently
published such listings. However, a fundamental difference does occur between fast-flowing streams
and slow-flowing rivers. This corresponds to a classification proposed by Illies and Botosaneanu
(1963) after examination of the many schemes proposed for river zonation which divides the river
course into two main classes - the rhithron and the potamon.

14
Figure 1.1 Longitudinal profiles of river systems:
A. Theoretical ideal profile; B. The Niger River
The rhithron is defined as the region extending from the source to the point where mean monthly
temperatures rise to 20°C, where oxygen concentrations are always high, flow is fast and turbulent
and the bed is composed of rocks, stones or gravel with occasional sandy or silty patches. The
rhithron is subdivided into three zones - the epi-, meta- and hypo-rhithron covering a range of water
courses from strong streams to small rivers.

15
The potamon is the region where monthly mean temperatures rise to over 20°C, oxygen deficits may
occur, flow is slow and the bed is mainly sand or mud. Three sub-zones are distinguished - the
epipotamon, the metapotamon and finally, the hypopotamon, which is that brackish water zone
affected by marine waters.
Because under this system temperature is important in defining the various zones, the change-over
from rhithron to potamon tends to be at higher altitudes in the tropics, where at low altitudes a true
rhithron zone may be entirely absent. This accounts for the differences in emphasis in studies or river
biology in the temperate zone, which have almost completely addressed the rhithron and in the
tropics, which are much more preoccupied with the potamon. However, because of the fundamental
differences between animal and plant communities of torrential headwaters and slow-flowing lowland
rivers regardless of temperature, it is useful to conserve this distinction even for tropical systems.
Thus in this work I have adopted the term rhithron to cover the steeper, rocky, torrential upper reaches
of rivers and potamon to cover the slow, mature lowland reaches.
TRIBUTARY DEVELOPMENT AND STREAM ORDER
Stream order
The dendritic arrangement of the channels of a river throughout its drainage basin is well known.
Several suggestions for ranking streams forming this type of pattern have been proposed. The most
widely accepted is that whereby streams are categorized according to order in a hierarchy defined as
follows: first-order streams are those having no tributaries, second-order streams are formed by the
union of two first order streams, third order streams by the union of second order streams and so on.
In its original form the system provided for one stream, usually the longest, of each category to be
extended headwords in such a way that the main channel of the river extends continuously from
source to mouth (Horton, 1945) (see Fig. 1.2). Later modifications of the system suppressed this idea
in favour of the more simple classification of all streams of the same order into one class (Strahler,
1957).
For ecological studies of rivers, each system has its advantages. The former is of use when
considering the evolution of some characteristic, for example fish catch, along the whole length of the
river. The latter is a more natural grouping and is useful in generalized studies in that streams of any
particular order tend to form sets, members of which can be considered together. Sudden changes in
faunal abundance are not uncommon below the junction of streams, particularly those of similar
order, where abrupt differences in flow, sediment load and other hydrological factors produce
correspondingly gross changes in the channel of the river. These, in turn, lead to a shift in the
ecological factors favouring one species group over another.
Clear relationships emerge between the numbers and lengths of streams of each order, whichever
system of ordering is adopted. These show that the number of streams of different order in a
watershed increases with decreasing order, according to a logarithmic relationship of the form:
Ns = a.bs,
where Ns = Number of streams of any order and s = order.
The length of stream of any order (Ls) decreases with decreasing order (s) in a similar manner:
Ls = x.ys.
The factors for a, b, x and y will vary according to continent or climatic zone.

16
Figure 1.2 Different systems for ordering river systems:
A. Horton (1945); B. Strahler (1957); C. Strahler's system as applied to the Logone River at the
height of Moundou
These relationships show that there are a very large number of small tributaries whose combined
lengths make up a considerable percentage of the total in any system (Table 1.1). For example,
Leopold et al., 1964, estimated that, of the 5 million kilometres of United States' streams with a mean
length equal to or greater than 1.6 kilometres over 90% were located in streams of fourth-order or
less. The relevant formulae are:
Ns = 7 661 837 (-1.58s)
and
Ls + 0.697 (0.8320s)

17
Table 1.1
Number and length of river channels of various sizes in the USA (from
Leopold et al., 1964)
Order a Number Length km) Total length % Cu.
1 1 570 000 1.6 2 512 000 48.39 48.39
2 350 000 3.7 1 288 000 24.83 73.20
3 80 000 8.5 674 400 13.07 86.26
4 18 000 19.2 345 600 6.66 92.92
5 4 200 44.9 188 160 3.62 96.55
6 950 102.4 97 280 1.87 98.62
7 200 235.2 47 040 0.91 99.33
8 41 540.8 22 173 0.43 99.45
9 8 1 243.2 9 946 0.19 99.94
10 1 2 880.0 2 880 0.05 100.00
5 191 479
a According to Strahler
Low-order streams are often of a torrential, rhithronic nature, whereas potamon reaches tend to be
concentrated around higher-order rivers. Low-order tributaries in lowland areas tend to be potamonic
in character especially in the tropics.
Relationship between river length and drainage basin area
The area of drainage basins and the length of streams within them are also related. Thus for the fifty
largest rivers of the world, as ranked by mean annual discharge, a relationship:
L = 1.7084A0.5418 (r² = .70)
emerges, where L = the length of the main channel in kilometres and A = the area of the drainage
basin in km². When analysed by region the coefficient and the exponent of this relationship vary, as
shown in Table 1.2.
Table 1.2
Relationship of length of main river to basin area for several areas of the world
U.S.S.R. North flowing rivers L = 0.8747 A0.5901 r² = 0.97 n = 6
N. America, Canada L = 0.9618 A0.5789 r² = 0.81 n = 6
N. America, U.S.A. L = 1.2528 A0.5609 a
Europe L = 1.5421 A0.5420 r² = 0.56 n = 7
India L = 2.2475 A0.5138 r² = 0.61 n = 11
Asia, East flowing rivers L = 3.3608 A0.4849 r² = 0.37 n = 24
S. America L = 3.4641 A0.4843 r² = 0.93 n = 16
Africa L = 4.9500 A0.4521 r² = 0.76 n = 30
a Derived from Leopold et al., 1964
The coefficient (a) and exponent (b) are themselves related for this data set by the equation:

18
b = 0.5797a-0.1492
Similarly, Gregory and Walling (1973) summarized data on variations in drainage density for various
parts of the world where total channel lengths are related to area. These, together with the
relationships in Table 1.2 indicate that there are between 1.6 and 8 kilometres of channel per square
kilometre of land surface, depending on landform and rainfall.
MORPHOLOGY OF THE RHITHRON
The main morphological characteristic of rhithron reaches is the alternation of pools and riffles which
arise from changes in gradient (Fig. 1.3). The steeper epi-rhithron is dominated by rapids, waterfalls
and cascades, but as the river proceeds downstream, the proportion of pool-like reaches relative to the
riffles increases and eventually the hypo-rhithron merges into the potamon. Riffles are steep, shallow
zones having coarse bottoms of boulders, rocks or pebbles. Pools are flatter, deeper zones with
bottoms of finer material. They may be more complex in form, especially in the hypo-rhithron, with
diverticula and backwaters having muddy or detritus bottoms, giving greater ecological diversity.
According to Leopold et al., 1964, one pool-riffle sequence always occupies a length of channel
equivalent to between five and seven channel widths, irrespective of the form or geographic location
of the river. The ratios between the relative lengths of pool or riffle lying within this segment do,
however, differ considerably. The productivity of rhithron reaches is frequently expressed in terms of
pool-riffle ratios, in which a slight dominance of the riffle component is deemed favourable for
salmonids because of the increased invertebrate food to be found in these zones.
Historical evidence presented by Sedell and Luchessa, (1981), and the existing situation in streams in
some less settled parts of the world, indicate that the present appearance of upland streams throughout
the temperate and much of the tropical world is very different from that pertaining under regimes
unmodified by man. A considerable portion of the inhabited world was once forested and streams
running through such areas received large quantities of wood, vegetation, boulders and other flow
modifying structures such as beaver dams. Thus streams up to 7th order were, and still are in some
parts of the world, conditioned by such obstructions which produced much better regimes than are
common today, with extensive overbank flooding, greater retention of water outside the stream
channel, considerable instream floodplain storage of carbon and more diversified habitat structures,
(Swanson et al., 1982).
Few attempts have been made to classify the rhithron into morphological types other than pools and
riffles. The variability that exists within these two major habitat types is such that further division is
needed if the areas are to be adequately described. One attempt at such a division has been made by
Bisson and Sedell (1982), who, taking US salmonid streams as an example, produced a typology
which seems sufficiently flexible to encompass rhithronic streams in other areas of the world. This
classification subdivides the various habitats as follows:
Riffle: Low gradient riffles - shallow stream reaches with moderate current velocity and turbulence.
Substrate usually gravel, pebble and cobble sized particles.
Rapids - gradient greater than 4 percent with swiftly flowing water and considerable turbulence.
Substrate generally coarser with larger boulders.
Cascades - steep reaches with variations in gradient consist of alternating small waterfalls and
shallow pools. Substrate consists of boulders.
Pools: Secondary channel pools - often temporary pools remaining in areas flooded by freshets,
often associated with gravel bars; sand or silt substrate.

19
Backwater pools - found along channel margins caused by eddies from large obstructions. Often
shallow and dominated by fine-grained substrates.
Trench pools - long, often deep, slots arising downstream of major obstructions. Usually with
coarse-grained but stable bottoms.

20
Figure 1.3 The morphology of the rhithron showing the division of the channel into a sequence of
pools and riffles
Plunge pools - arise where a stream passes over a major obstacle and scours out the bottom.
Variable in depth and substrate.
Lateral scour pools - occur where flow is diverted by a large obstacle or obstruction.
Dammed pools - consist of water impounded upstream of a complete or nearly complete channel
blockage. Tends toward low current and finer substrates.
Glides: a third general habitat category consisting of moderately shallow reaches with even flow that
lack obstructions or pronounced turbulence. Bottoms usually gravel and small cobbles. Glides
are particularly common in large rivers passing through mountainous terrain where channels may
run relatively straight for many kilometres.
As rhithron reaches are for the most part situated in low-order streams, their flood regimes are apt to
be somewhat “flashy” with rapid changes in discharge. The water is usually confined within a well
defined bed, although peak spates may flood a narrow strip fringing the channel. During periods of
high discharge, the riffle pool complex tends to be completely submerged and from the surface the
two components may appear to lose their identity. At intermediate discharges the riffle has turbulent
flow while the pool has laminar flow, often with relatively calm areas at the banks. As discharge falls
the distinction between riffles and pools increases until eventually, when flow ceases completely, the
riffles are left dry and the pools retain water giving the channel the appearance of a string of beads.
At slightly lesser slopes for the same bankfull discharge braided channels arise. These consist of many
anastomosing anabranches, which may themselves meander, winding among rocky, sandy or
vegetated islands which are exposed at low water and flooded at high water (Fig. 1.4). Pool and riffle
systems may appear within the individual anabranches in steeper and stonier reaches.

21
Figure 1.4 An example of a braided channel; the Niger River at Ayourou
MORPHOLOGY OF THE POTAMON
The potamon of reservoir and regulated rivers consists only of the channel which may be meandrine
or braided in form. In flood rivers, however, there are two major components to the potamon: (i) the
channel, and (ii) the floodplain which correspond to the French terminology “lit mineur” and “lit
majeur” and represent the beds of the river in its two main phases - low water and flood.
The form of many of the world’s floodplains has undoubtedly been changed considerably by human
activities over the last few thousand years. Prior to human occupancy, it seems likely that most rivers
were associated with riparian gallery forests, and that, in more arid areas, the plains themselves were
covered with scrub of the bush savannah type. These conditions still persist in some Latin American
rivers, and give water regimes and channel forms which are more stable than those of the open
savannah rivers which have resulted from clearance of the trees for agriculture, grazing and firewood.
According to Leopold et al. (1964) floodplains will typically include the following features:
(a) the river channel;
(b) oxbows or oxbow lakes representing the cut-off portion of meander bends;
(c) point bars - loci of deposition on the convex side of curves in the river channel;
(d) meander scrolls - depressions and rises on the convex side of bends formed as the channel
migrates laterally down valley by the erosion of the concave bank;

22
(e) sloughs - areas of dead water formed both in meander-scroll depressions and along the valley
walls as flood flows move directly down valley scouring adjacent to the valley walls;
(f) natural levees - raised berms or crests above the floodplain surface adjacent to the channel,
usually containing coarser materials deposited as floods flow over the top of the channel banks.
They are most frequently found at the concave bank. Where most of the silt load in transit is fine-
grained, natural levees may be absent or nearly imperceptible;
(g) backswamp deposits - overbank deposits of finer sediments deposited in slack water ponded
between the natural levees and the wall or terrace riser;
(h) sand splay - deposits of flood debris usually of coarser and sand particles in the form of splays or
scattered debris.
These features, which are illustrated diagrammatically in Fig. 1.5, determine the quantity, distribution
and flow of water in the system throughout the year. Most of them are readily distinguishable in
moderate sized floodplains, but in smaller valleys are obscured by the rapidity with which changes
occur. Individual features of the floodplain are briefly described below (numbers in text refer to Fig.
1.5).
The channels - the lotic component
The main channel (1) or channels of the river and its anabranches (2) usually retain water, but not
necessarily flowing water, at all times of the year. As the river enters its alluvial plain it starts to
meander forming wide convoluted channels whose curves are proportional to river width (Leopold et
al., 1964). In some larger rivers, the Amazon and Congo, for example, braided channels occur in
which the islands form levees with depression lakes at their centre. Where such braided channels
occur the lateral floodplain is sometimes limited in width and its whole extent may come to be
contained within the main channel. In these instances the islands are analogous to the lateral
floodplain and fulfil a similar role in the biology of the fish (Svensson, 1933; Gosse, 1963).
In addition to the main channel or channels of the river, floodplains have a network of channels and
creeks which penetrate the levees to connect the main river with the back-swamps and meander scroll
lakes. Such channels may or may not retain water at all times of the year, but they represent the main
path of water and fish movement during the earlier periods of rising water and later phases of falling
water.
On many floodplains naturally occurring channels are supplemented by artificial canals constructed
for navigation, irrigation, drainage or even fisheries.
The floodplain
The floodplain is itself divided into two components: (i) the plain itself which is seasonally inundated,
but remains dry for at least part of the year, and (ii) the standing waters which remain in the plain
during the dry season.
The plain (seasonally inundated component)
The alluvial plain of a river can be divided into two main zones: Firstly, the levee regions (7), which
more or less follow the course of the river channel and its former beds, consist of raised areas that are
flooded for the shortest time annually; and secondly, the flats, which extend from the levee to the
terrace or plateau delimiting the plain. Exceptionally high levees sometimes occur immediately
adjacent to the channel which is only occasionally submerged by the highest of floods. Such areas,

23
together with the raised terraces bordering the plain are used for human habitation, or give island
refuges for cattle or wild game during the flood season. Levees may be much reduced or even
completely absent in rivers carrying very fine silt. Because coarser material is deposited early after the
slowing of the flow as a river enters the plain, there is a tendency for the raised areas bordering the
river channel to diminish in height downstream. In river basins with exceptionally high silt loads, for
example the Chao Phrya delta in Thailand prior to the installation of flood control structures (see Fig.
1.6), the process of levee building and of deposition within the main channel may raise the river
channel and levee high above the surrounding plain.
Figure 1.5 Diagram of the main geomorphologic features of a floodplain; numbers refer to
description in text

24

25
Figure 1.6 Plan and cross-sections of the Chao-Phrya delta, Thailand, before the installation of flood
control structures, showing channels raised within levees above the level of the surrounding
plain. (After Ohya, 1966)
The flats, which make up the greater proportion of the plain, show slight differences in relief due to
old depositional features. More depressed parts are interspersed with standing waters of various types.
The edges, adjacent to the terrace or valley wall delimiting the plain, may be more deeply excavated
where locally increased flow scours deposits to form lagoon or backswamp complexes (6).
Irregularities may also be formed where inflowing tributaries deposit alluvial fans. Both the levee and
the terraces may be covered with dense gallery forest (see Figs. 1.7a and 1.7b), but many floodplains
are occupied only with sparse scrub or are denuded of trees altogether (savannah plains).
Figure 1.7 Schematic profile of: A. The Amazon (after Sioli, 1964), and B. The Middle Parana (after
Bonetto, 1975), showing distribution of vegetation and main floodplain features

26
Standing waters (the lentic component)
Permanent or semi-permanent standing waters are left by receding floods in the form of sloughs in
oxbows (4), meander scroll depressions (5), backswamps (6), or the residual channels left by the
former course of the river (3). These water bodies expand and contract according to the annual flood
cycle (Fig. 1.8) and during the highest floods tend to merge into a continuous sheet of water covering
the whole plain.
Figure 1.8 Annual cycle of flooding of a typical floodplain depression of the Senegal River (Vindou
Edi). (After Reizer, 1974)
Distinction is often made between lakes, lagoons or pools on the one hand and swamps on the other.
Although the terms lake, lagoon and pool have been used interchangeably in the literature, and refer
more or less impartially to bodies of water of some depth and slight to moderate vegetation cover,
there is a useful distinction to be made between lakes, as large features of a floodplain system which

27
persist relatively unchanged over a number of years, and lagoons and pools as more transitory open
water feature. Lagoons remain connected to the river throughout the year, whereas pools are usually
smaller and more ephemeral bodies of water which become isolated and have a tendency to dry out in
the dry season. The term “swamp” is applied to those depression wetlands whose soil remains
saturated or more or less permanently covered with shallow waters and which support, as a result,
characteristic growths of vegetation which dominate the environment.
Permanent standing waters of the floodplain are generally shallow, rarely exceeding 4 m in depth and
may be in communication with the river. Alternatively, their deepest point may lie below the water
table of the plain enabling them to remain wet throughout the year.
Water bodies on the floodplain lose water by evaporation and to a lesser degree by filtration
throughout the dry season. This results in the contraction and eventual drying out of many water
bodies with a concomitant concentration of dissolved substances. In most river systems this does not
produce any appreciable result, as concentrated solutions are rapidly washed out by the next flood and
the average conductivity of the water remains low. However, in some regions of high salinity or one
way flow, temporary brackish pools or salt pans can result. The most extreme examples of such areas
are perhaps the one million hectare Plain of Reeds of the Mekong Delta, where there are
extraordinarily high concentrations of alum and the Ovambo floodplain which terminates in the saline
Etosha pan, (see Fig. 1.14) and the Helmand River in Afghanistan with its terminal Sistan Marshes.
Very large lakes or groups of lakes are associated with some floodplains. These are sometimes
distinct entities geologically, but ecologically they are usually completely integrated into the
river/floodplain system. The greatest of such lakes, the Grand Lac of the Mekong (Fig. 1.9) floods an
area of 11 000 km², most of which was forested, but reduces to about 2 500 km² of open water in the
dry season. It is connected to the main river by a broad channel with reversible flow, the Tonle Sap.

28
Figure 1.9 The delta of the Mekong River with the Grand Lac and Tonle Sap
A similar but smaller feature is found in the Senegal River valley where the Lac de Guiers is
connected to the main channel by the River Tawey. The lake has an extreme dry season area of 120
km² and expands to twice this when flood waters flow into it through the Tawey in August and
September. The lake district of the Niger Central Delta (Fig. 1.10) forms part of the general flood
system at high water, but breaks up into 18 major lakes with a combined area of 2 400 km² at low
water. The 11 000 km² Kamelondo depression floodplain of the Lualaba contains some 50 permanent

29
and semi-permanent lakes, having a total area of 1 545 km² and of which the largest, Upemba, has a
dry season area of 530 km². In the Magdalena River (Fig. 1.11) the waters are confined within some
800 lakes (cienagas) of varying size and permanence at low water. Their total area is about 3 400 km²,
although some individual water bodies, for example the Cienagas, Zapatosa (119 km²) and Ayapel
(123 km²) are of considerable size.
Figure 1.10 The internal delta of the Niger river and its tributary the Bani, in Mali, showing the
numerous lakes at the northern end of the plain
The varzea floodplain of the Amazon is similarly interspersed with lakes, some of which are very
large. There are 20 lakes of over 50 km² and the Lago Grande do Curuai covers 630 km². While these
resemble the floodplain lakes found in other systems a special type of water body is also found in the
Amazon. The clear water tributaries to the lower and middle reaches of the river have large widened

30
mouth bays formed originally from drowned valleys. Such “river-lakes” are the site of sedimentation
at their upstream ends where new varzea type floodplains are formed from the deposits.
The individual water bodies of the floodplain usually persist over long periods during which they age,
although particularly high floods can produce sudden changes through scour and deposition. The
ageing process consists of a natural succession from open water lake to dry land via the intermediate
stages of shallow lagoon and swamp ecosystems. Botnariuc (1967), in particular, has studied the
processes involved in the transformation of the lakes of the Danube floodplain. Here the transition
from lake (“Ghiol”) to reed-grown pond (“Japse”) is caused mainly by the growth of emergent
vegetation which slows water currents and accelerates siltation thus allowing a further extension of
the vegetated area, although the process is partially reversible when higher floods may scour deposits
and vegetation from the lake bottom. In the Amazonian floodplain too, the progression from water
bodies invaded by aquatic grasses, through shrub vegetation to different stages of floodplain forest,
noted by Junk (1983) is frequently prevented from reaching its climax through scour by exceptional
floods. Siltation can be detected in the deposition of new varzea in the mouth lakes of the Amazon or
in the change in form of lakes and channels viewed as a time series by remote sensing. Side looking
radar and satellite imagery show floodplains to be littered with the remains of old floodplain features
now silted over. The succession may also be reconstructed from series of lakes such as those
described by Green (1972) for the meander complex of the Suia Missu River in Brazil, or by Castella
et al. (1984) for the relic beds (“lones”) of the Rhone river in France. The disappearance of older
features of the floodplain in this way is compensated for by the generation of new bodies of water by
the constantly changing course of the main river channel. The speed with which this occurs is related
to the discharge of the river and its silt load, but it is to be supposed that, given relative constancy of
these parameters, the ratio of open water bodies on the plain to total area remains relatively
unchanged through time.

31
Figure 1.11 The internal delta of the Magdalena River at its confluence with the Cauca and San
Jorge rivers in Colombia showing the numerous floodplain lakes (cienagas)
Proportionality of floodplain features
The proportion of the floodplain which remains permanently under water is generally as difficult to
establish as is the total area submerged at the peak of the floods. There is for the most part a lack of
reliable maps showing floodplain features and even where they do exist the period of mapping rarely
coincides with the period of minimum water. However, the use of satellite imagery presents a possible
solution to this problem in areas where cloud cover remains within acceptable limits. Further
complications arise from the instability of floodplain features and the changes wrought by man
himself. In most populated areas the floodplain and its hydraulic regime have been considerably
modified by the digging of canals, the raising of artificial levees and the levelling of depressions.
Some information on relative areas is available from the best studied floodplains in Africa (Table 1.3).
As flood regimes vary in the intensity of both their flood and dry season components, values for high
water and low water areas fluctuate about a mean. The figures quoted in Tables 1.4 and 1.5 should be
treated as such averages, being only approximations derived from relatively small-scale maps.
In Africa the area of permanent water ranges from 5 to 59 percent of the total flooded area, with a
pronounced mode at between 10 and 20 percent. There is insufficient information to judge whether
significant differences exist between the flood ratios of the various categories of floodplain and these
may depend more on soil type or local climate. It is difficult to describe the simple ratio between

32
flooded and permanent water areas. Indeed the extent to which the various depositional features of the
plain can change from season to season makes such figures of only temporary value, except as
indicators. Table 1.5 shows, for some African plains, the proportion of the permanent waters that
remain on the floodplain as standing waters and those located in the divers river channels.
There are pronounced differences in this proportion and the smallness of the sample does not permit
definite conclusions to be drawn on the possible causes of this. However, it might be expected that on
floodplains where agriculture is intensively practised during the dry season, much of the standing
water and particularly the swamps will tend to disappear through drainage and fill. This, in fact,
appears to be the case for the cultivated Senegal, Ouémé and Pongolo plains as compared with the
Niger, Lualaba and Kafue plains, which are used primarily for cattle grazing. The main exception to
this is the Shire, in which a large amount of the standing water is classified as permanent swamp.
Unfortunately, the dividing line between swamp and lagoon is somewhat fine and vegetated areas of
lagoons are often placed in this category.
Table 1.3
Characteristics of some African floodplains
Floodplain Area at
peak flood
(km²) ‘A’
low water
(km²) ‘B’ B/A × 100 Authority
Senegal R. Mean for total system 5 490 800 15 OMVS
Niger R. Central delta 20 000 3 877 19 Raimondo, 1975
Fringing plain
Niger 907 270 30 FAO/UN, 1971
Fringing plain
Benin 274 32 12 FAO/UN, 1970
Fringing plain
Nigeria 4 800 1 800 38 FAO/UN, 1970
Benue R. Fringing plain
Nigeria 3100 1290 42 FAO/UN, 1970
Ouémé R. Coastal delta 1000 52 5 Pers. observations
Chari and Yaeres 7 000 NI - Ali Garam, pers.comm.
Logone R. Total system 63 000 6 300 10 Blache, 1964
Zambezi R. Barotse 10 752 537 7 FAO, 1969
Okavango Internal delta 17 000 3 120 20 Cross, pers. comm.
Pongolo R. 100 26 26 Coke and Pott, 1970
Kafue R. Kafue flats 4 340 1 456 27 Gay, pers. comm.
Shire R. Elephant and
Ndinde marshes 665 200 30 Hastings, pers. comm.
Total sytem 1 030 480 48
Luapula R. Kifakula (lagoon) 1984 195
depression (river) 1 500 75 13
Total 266
Lualaba R. Kamulondo
depression 11 840 7 040 59
Nile R. Sudd 31 800 16 300 51.2 Mefit-Babte, 1984
Volta R. Fringing plain
Ghana 8 532 1 022 12 Vanderpuye, pers.comm.
Ogun R. Fringing plain 43 25 59 Dada, pers. comm.
Oshun R. Fringing plain 37 20 73 Dada, pers. comm.
Masilli R. Fringing plain 15 2 13 Dada, pers. comm.

33
Table 1.4
Areas at peak flood of some Central and South American floodplains
Floodplain peak flood Area at high water (km²) Authority
Grijalva/
San Antonia R. Delta 8 000 USAF1
Atrato R. Delta 5 300 USAF
Magdalena R. Delta 20 000 Pardo, 1976
Catatumbo R. Delta 5 000 USAF
Orinoco R. Apure/Arauca
Internal delta 70 000 Matthes,
pers.comm.
Orinoco R. Coastal delta 20 000 USAF
Rupununi R. Internal flood zone 6 500 USAF
Amazon R. Central delta 50 000 Sioli, 1975
Amazon R. Coastal delta 25 000 Sioli, 1975
Paraguay R. Gran Pantanal 80–100 000 Bonetto, 1975
Parana R. Fringing plain 20 000 Bonetto et al.,
1 United States Operational Navigation Chart - Scale 1:1 000 000
Table 1.5
Composition by area of different types of permanent water on floodplains during the dry season
Standing waters
Floodplain
River and
channels (ha²) Lagoons (ha²) Swamps (ha²) Total (ha²) Total area
(ha²)
Niger R. Central delta 61 300(16) 300 400(77) 26 000 (7) 316 400(84) 389 700
Lualaba-Kamulondo 29 400 (4) 154 500(23) 480 000(72) 643 500(96) 663 900
Kafue-Kafue Flats 5 380 (4) 10 180 (7) 130 000(89) 140 180(96) 145 560
Shire 2 000 (4) 5 500(12) 40 599(84) 46 000(95) 48 000
Pongolo 392(14) N.D. N.D. 2 428(86) 2 820
Ouémé 1 402(27) 3 768(73) slight 3 768(73) 5 170
Senegal valley 28 100(57) 21 800(43) slight 21 800(43) 49 700
( ) Percentage of total
N.D. No distinction made
The various floodplain features are sufficiently important in the lives of the peoples inhabiting major
rivers for a particular terminology to have arisen in many languages. While by no means exhaustive,
Table 1.6 lists the main names that have been used in the literature by workers from various countries
to describe individual features.
Table 1.6
Glossary of terms for floodplain features in various regions of the world
Country

34
Levee Floodplain FEATURES Depression
lagoon or swamps Channel and side
arms
Benin Tikpa Ti
Brazil Varzea Lago de varzea Parana/Igarape
Cambodia Veal Beng Prek
Colombia Cienaga Cano
India Bheel Jheel
Romania Ghiol:Japse
Papua Roundwaters Barats
N. Guinea
Senegal Fonde Oualo Vindo Tiangol
Sri Lanka Villus
Sudan Toiche
Sumatra Lebaks (temporary)
Lebungs (permanent)
Congo Madzibe (large lagoon)
Edzibe (smaller Malala
oxbows)
Types of floodplain
It is perhaps hazardous to attempt any definitive classification of floodplains. However, three general
types can be discerned whose characteristics are sufficiently different that they may influence either
the behaviour of the fish populations inhabiting them, or the problems faced by a fishery.
Fringing floodplains: Nearly all tropical and sub-tropical rivers and many temperate ones have a
lateral flood zone. This takes the form of a relatively narrow strip of floodable land lying between the
river valley walls (Fig. 1.12). Fringing floodplains are normal developmental features of a river which
follow its course in all areas where the slope is favourable. They tend to increase in width the less the
slope of the river and this generally means a progressively greater elaboration of the floodplain along
the river's course to the sea.

35
Figure 1.12 Floodplain fringing the Niger river at the level of Gao in Mali
Internal deltas: Occasionally, river systems encounter geological features which cause them to spread
laterally over very large alluvial plains. Such features may be the site of a former lake filled with
alluvium, for instance the Yaeres of Lake Chad; the deltaic discharge of a small river into a larger
one, such as is found where the Apure flows into the Orinoco; or the backing up of the aquatic

36
systems by an obstruction downstream, the Kafue Flats for example. Such floodplains may occur at
any point along the course of the river. The main stream usually becomes divided into anabranches
which rejoin the main channel below the deltaic area, or several rivers flowing into the same plain
may interconnect with complex and shifting channels. In certain parts of the world, the Bahr Aouk
headwaters of the Chari system, the Gran Pantanal of the Paraguay River or the Apure-Arauca
tributaries of the Orinoco, for instance, the terrain is so flat that enormous areas of land are flooded to
a very shallow depth by rain-water as well as by overspill from the river. Such “sheet flooding” is
often dictated by land form processes other than those created by the rivers themselves, but the
network of channels and lagoons that arise by the erosion during drainage give such areas a deltaic
character.
Coastal deltaic floodplains: The terminal lateral expansion of the alluvial plain and the break-down of
the main river channel into distributaries produce the classic fan-shaped delta. Coastal deltaic
floodplains are influenced by the marine environment in that, in the dry season, sea water penetrates
the main channels as a saline tongue. Tidal effects are often transmitted far upstream even beyond the
limits of the saline tongue, but little invasion by salt water occurs during the floods, and only the
coastal fringe is submerged by tidal action.
Certain floodplains are intermediate between the internal delta and the coastal delta. These occur in
those rivers which discharge deltaically into either inland freshwater lakes, such as the Yaeres systems
located at the place of discharge of the Chari-Logone into Lake Chad, or into rivers much larger than
themselves. The principal ecological differences between fringing and internal deltaic floodplains on
the one hand, and coastal floodplains on the other are, firstly, the greater extent of the water body
available to fish in the dry season in the coastal floodplain and, secondly, in the penetration of sea
water and marine species into areas adjacent to the sea.
BRIEF REVIEW OF MAJOR RIVERS
This review is intended to highlight those features of the rivers of the world that are particularly
significant to fisheries. Thus less emphasis is placed on the numerucal3ly dominant small order
streams or upper torrential reaches, than on features such as the degree of modification or the
elaboration of the floodplains with their associated lakes and swamps.
It is difficult to define accurately the total number and lengths of the world's rivers, although by
simple extrapolation some estimate can be made. The relationship for river length as a function of
basin area for the world as a whole has a coefficient of 1.7, implying that for any one square kilometre
of land surface there are on average at least 1.7 km of channel. Taking the drained surface area of the
world as 1.1 × 108 km², a minimum length of channel of 1.8 × 108 km results. This is a minimum
estimate as the original equation relates only main channel length to basin area and thus omits any
secondary channels.
The drainage pattern of most continents is dominated by few very large river systems, the length,
basin area and mean annual discharge of the fifty largest of which are listed in Table 1.7.
AFRICA (Fig. 1.13)
Upland and torrential rivers are fairly common in Africa for, although there are few large mountain
chains, the elevated nature of the mass of the continent and the existence of individual massifs means
that most rivers take their source in highland regions. For example, many of the West African rivers,
including the Senegal and Niger, take their source in the Fouta Djallon mountains of Guinea; the
Benue and Logone rivers arise in the Mondara mountains of Cameroon; the north flowing tributaries
of the Congo in the highlands of Angola; the Blue Nile in the Ethiopian highlands, etc. However,
nearly all African rivers have extremely well developed fringing floodplains in their lowland courses
of which perhaps the best studied are the savannah plains of the Senegal and Niger rivers. The

37
Senegal flows through a broad valley which, during the dry season, retains about 500 km² of water
confined in “sloughs” of various kinds (66 km²), as well as in the Lac de Guiers (150 km²) and the
main river channel (281 km²). At peak floods the river covers 5 000 km² of valley. The Niger and its
tributary the Benue river, also cover an extensive lateral plain. In the Republics of Niger, Benin and
Nigeria, the Niger itself flooded 5 981 km² at high water, shrinking to about 35 percent of this area in
the dry season. Much of the Nigerian plain of this river has since been lost due to the flood control
effects of the Kainji dam and its reservoir. The Benue river, also in Nigeria, has a very impressive
fringing plain, broad for its length which has a flooded area of 3 100 km² and a dry season area of 1
290 km².
Table 1.7
50 large rivers of the world ranked by mean annual discharge at mouth
RIVER CONTINENT /
COUNTRY
MEAN
DISCHARGE
1000 m³ sec-1
DRAINAGE
AREA 1000
km²
LENGTH
km RANK BY
LENGTH
1. Amazon S.America: Brazil 212.5 5711 6437 2
2. Congo/Congo Africa: Congo 39.7 3968 4700 8
3. Yangtze Asia: China 21.8 1920 5980 4
4. Brahmaputra Asia: Bangladesh 19.8 924 2900 31
5. Ganges Asia: India 18.7 1047 2506 48
6. Yenisei Asia: USSR 17.4 2560 5540 5
7. Mississippi/
Missouri N.America: U.S.A. 17.3 3184 6020 3
8. Orinoco S.America: Venezuela 17.0 870 2151 60
9. Lena Asia: USSR 15.5 23.96 4400 10
10. Parana S.America: Argentina 14.9 2278 3998 18
11. St. Lawrence N.America: Canada 14.1 1274 4000 16
12. Irrawaddy Asia: Burma 13.5 424 2100 63
13. Ob Asia: USSR 12.5 2455 5410 6
14. Mekong Asia: Thailand 11.0 793 4000 16
15. Amur Asia: USSR 11.0 1822 4444 9
16. Tocantins S.America: Brazil 10.2 896 2700 38
17. Mackenzie N.America: Canada 7.9 1784 4241 13
18. Magdalena S.America: Colombia 7.5 238 1600 -
19. Columbia N.America: Canada 7.3 660 1954 76
20. Zambezi Africa: Mozambique 7.1 1280 3500 14
21. Danube Europe: Romania 6.2 806 2850 34
22. Niger Africa: Nigeria 6.1 1100 4200 14
23. Indus Asia: Pakistan 5.6 916 2900 31
24. Yukon N.America: Canada 5.1 921 2654 44
25. Pechora Europe: USSR 4.1 322 1809 88
26. Uruguay S.America: Uruguay 3.9 230 1612 -
27. Kolyma Asia: USSR 3.8 637 2513 47
28. Sankai Asia: China 3.6 117 1957 74
29. Godavari Asia: India 3.6 294 1440
30. Dvina Europe: USSR 3.5 355 726
31. Hwang-Ho Asia: China 3.3 665 4845 7
32. Frazer N.America: Canada 3.2 235 1360
33. Nile Africa: Egypt 2.8 2944 6650 1
34. Sao Francisco S.America: Brazil 2.8 665 2900 31
35. Neva Europe: USSR 2.6 279 -

38
36. Pyasina Asia: USSR 2.5 189 1056
37. Nelson N.America: Canada 2.3 1059 2570 43
38. Rhine Europe: Netherlands 212 143 1312
39. Krishna Asia: India 2.0 304 1120
40. Indigirka Asia: USSR 1.8 355 1725 95
41. Dnepr Europe: USSR 1.7 496 2200 58
42. Rhone Europe: France 1.7 94 816
43. Mobile/Tombigbee N.America: U.S.A. 1.6 107 598
44. Salween Asia: Burma 1.5 276 2400 52
45. Tigris/Euphrates Asia: Iraq 1.4 535 1900 81
46. Po Europe: Italy 1.4 69 648
47. Vistula Europe: Poland 1.1 194 1084
48. Susquehanna N.America: U.S.A. 1.1 71 710
49. Yana Asia: USSR 1.0 243 1067
50. Senegal Africa: Senegal 0.9 338 1633 100

39
Figure 1.13 Location of the major rivers and floodplains of Africa:
1. Senegal 7. Lualaba 13. Shire
2. Niger, Central delta 8. Barotse 14. Luapula
3. Ouémé 9. Okavango 15. Rufigi
4. Logone, Yaeres 10. Cunene/Ovambo 16. Ruaha
5. Chari 11. Pongolo 17. Nile, Sudd
6. Congo, Mbandaka 12. Kafue
The Zambezi has two plains on its upper course which flank the river for 240 and 96 km respectively.
They flood laterally for a greater distance than usual, penetrating up to 16 km² inland on either bank.
the combined area of these Barotse plains is over 10 000 km², but only 5 percent of this area remains
wet in the dry season.

40
The greatest of the African coastal deltas, that of the Nile whose floods were the basis of the wealth of
pharaonic Egypt, is no longer inundated seasonally. The fringing floodplains of the river have also
virtually disappeared due to the flood control measures of the series of dams barring the middle
course of the river. Savannah deltas occur in the Senegal river where nearly 8 000 km² are flooded
annually, and the Ouémé river, whose 1 000 km² delta terminates in the 180 km² brackish water Lac
Nokoue. The Niger delta covers 36 260 km² with a coastal fringe of saline mangrove swamps.
Internal deltas are common in Africa. The largest of these, that of the Niger (Figure 1.10), occurs
where sand blown from the Sahara has resulted in the deflection of the Niger river eastwards near
Timbuktu. A depositional plain has grown up behind this with lakes lying in the depressions between
rocky outcrops. It extends over 20 000–30 000 km² during the four to five month flood season, but its
area shrinks to 4 000 km² in the dry season, most of which is retained in the permanent lakes. The
Kafue river, backed up by a range of hills, which are now the site of the Kafue Gorge dam, forms an
alluvial plain of over 6 600 km² which is inundated almost completely during the rains. Only 1 456
km² of permanent waters remained throughout the year prior to the closing of the dam although the
duration and extent of inundation has been increased subsequently. The 7 000 km² Yaérés floodplain
is the site of the deltaic discharge of the Chari and Logone rivers into Lake Chad through the
Logomathia and El Beid rivers. The present lake and the floodplains of the lower Chari occupy the
site of a larger Paleo-Chad, now shrunk in size by the progressive desiccation of the Sahara Desert.
The Yaérés are only part of a much larger family of floodplains centred on the Chari and Logone
rivers. At least two other groups of plains subject mainly to sheet flooding by rainfall and local run-
off are found in the system. The largest of these extends from the Bahr Aouk and Bahr Salamat rivers
covering a considerable portion of southeast Chad and some 37 000 km² of northeastern Central
African Republic. The second group spreads between the Logone and Chari rivers and eastward along
the Bahr Erguig anabranch of the Chari. According to Blache (1964) the Chari/Logone basin
floodplains had a combined area of 90 000 km², of which about 70 percent were inundated at peak
floods (September-October) and only 7 percent remained wet in the dry months of April and May
during periods of normal rainfall.
Many African flood areas are associated with permanent swamp systems. The most famous of these
and the most extensive wetland in the continent is the Sudd of the river Nile. Located at the
confluence of the Nile and Bahr el Ghazal, the 16 300 km² permanent complex of papyrus swamp and
open water lagoon swells to twice this area when the floods of the Nile arrive from the south and
additional areas of variable extent are flooded by rainfall and local run-off. A large waterway, the
Jonglei canal, is currently being excavated to bypass the swamps and make more water available
downstream. The Elephant and Ndinde marshes of Malawi, which cover 673 km² when flooded,
shrink to 384 km² of swamps and lagoons in the dry season. They form part of the larger Shire river
system which covers a total flooded area of 1 400 km² or 480 km² in the dry season. A third large
swamp complex is centred on the internal delta of the Okavango river and the 800 km² Lake Ngami.
These swamps have a residual area of 3 120 km² and cover between 16 000 to 20 000 km² at high
water. The Kamulondo depression, one of the few true floodplain areas of the Congo, contains lakes
and swamps with a permanent water area of approximately 7 000 km², which nearly doubles during
the flood season. The Rufigi/Ruaha river system of Tanzania contains three large floodplains; the 1
451 km² fringing plain of the Rufigi river itself, the 4 400 km² Usungu plain of the Ruaha river which
includes the 518 km² Utungele swamp, and the 6 736 km² floodplain of the Kilombero tributary to the
upper Rufigi which contains the 89 km² Kibasira swamp. In common with most Africa wetlands the
extent of these large plains varies considerably according to the medium term climatic regime. The
sahelian drought of the 1970's and 1980's reduced the area of the plains of the Niger, Senegal and
Chari/Logone systems whereas the increased pluviosity of the 1960's in the Central Africa plateau
resulted in an enlarged Sudd. Indeed it is now suspected that the environment effects of such natural
variation exceed the results of such human interventions as the Jonglei canal.

41
Two smaller African plains resemble in some ways the cienaga floodplains of Latin America. The
Kifakula plain, formed by the deposition of alluvium by the Luapula river between the Johnson Falls
and Lake Mweru, covers 1 500 km² and is scattered with permanent lagoons. The Pongolo floodplain
is even smaller (130 km²), but is of interest in that its upstream end is blocked by the Pongolapoort
dam, which has permitted experiments on the discharge requirements for the maintenance of fisheries
in small floodplains.
True forested floodplains similar to the Amazonian Igapó and várzea forests are more or less confined
to the Congo basin and a few smaller river basins in Cameroon and Gabon. The course of the Congo
below Kisangani broadens to include an increasing floodable area. which culminates in the Bangala
swamps and the vast complex of flood-lands at the confluence of the Congo, Ubangui and Sangha
rivers. These areas are subject to bi-modal flooding and tend to retain their water to a great extent and
therefore differ very considerably from the highly seasonal type of plain. The true extent of this area
is difficult to assess and it is almost completely unpopulated.
An unusual type of flood wetland is formed by the overspill from the Cunene river in South Angola.
In the wet season this river discharges a considerable part of its flow southwards where it is contained
within a graben. As it flows to the south through numerous channels and pools the evaporation raises
the salt content until finally the system becomes dry at the Etosha pan. Some 10 000 km² of this
Ovambo floodplain consist of highly saline soils and water but conditions are less saline towards the
North where some 20 000 km² of waterways and floodable land are retained in a humid condition by
ground water seepage.
AMERICA (NORTH) (Fig. 1.14)
Many of the northern rivers of the continent consist of relatively short channels connecting the large
numbers of lakes of the Northern forest and tundra. The land is mainly flat with heavy flooding of
swampy areas during the spring when melting snow forms the major part of the run-off. Because of
the severe winters, streams and swamps freeze for up to six months of the year and the floodplain of
these are of the tundra type. Several large rivers drain the area, including the Mackenzie and the
Yukon. The Peace and Athabasca rivers together formed a 2 560 km² complex of flood lakes and
swamps at their entry to Lake Athabasca (Blench, 1972) but this environment has now been
superseded by a series of isolated mud flats following the closure of a dam on the Peace river. Within
the United States most rivers are highly modified by various flood control works. For instance, the
Colorado is now a cascade of reservoirs and abstraction of water from the system for irrigation is on
such scale that there is little outflow to the sea. Similarly the rivers of the Mississippi - Missouri
basin, which drain most of the United States, have also been extensively altered, the Missouri and
Mississippi mainstems having been converted into a series of pooled reaches. While many of the
rivers draining the Rockies and the Coastal ranges are turbulent and encased in steep gorges, most
streams are associated with floodplains and the 100 year flood area of the US is estimated by Sabol
(1974) at 543 000 km² or 6 percent of the total land area of the Nation. Most of Mexico is arid and
few large rivers drain the area. However the Grijalva, Usumacina, San Antonio and San Pablo rivers
of Tabasco state combine near their mouths into a series of anastomosing channels running through a
broad flood zone which covers an area of about 8 000 km² at maximum floods. The plain extends up
to 150 km inland and abundant floodplain water bodies are located along the main water courses. The
other rivers of Central America are short, often steep and swift flowing streams associated with the
mountainous terrain.
AMERICA (SOUTH) (Fig. 1.15)
The Chilean, Peruvian, Equadorian and Colombian rivers draining the Andes westwards to the Pacific
are all short and torrential. So too are the upper reaches of the North flowing Atrato, Magdalena and
Sinu rivers of Colombia, and the headwaters of those tributaries which drain the Andes eastwards to
the Orinoco, Amazon and Parana systems. The Marañon and Ucayali tributaries to the Amazon

42
particularly have long courses in the mountains. Other upland rivers are found in the various plateaux
highlands of the continent but on the whole Latin America is relatively flat and most rivers are of the
lowland type with extensive plains which are flooded by local rainfall as well as by river overspill.
Figure 1.14 Location of the major rivers and floodplains of North America:
1. McKenzie 5. Saskatchawan 9. Grande
2. Yukon 6. Fraser 10. Mississippi
3. Peace 7. Colorado 11. Missouri
4. Athbasca 8. Usumacinta/Grijalva 12. St. Lawrence
Several morphologically similar floodplains clustered around the Carribean consist of internal or
coastal deltas which contain enormous numbers of permanent or semi-permanent lakes, locally termed
‘cienagas’. The most important of these plains is that of the Magdalena river (Fig. 1.11), whose
internal delta with the San Jorge and Cauca rivers extends over 20 000 km² of savannah. Most of the
plain is well inland but extends seaward along the Canal del Dique on one hand and down the main
river channel to the coastal Cienaga Grande on the other. The whole area of 20 000 km² can flood for
up to a month, 16 000 km² for between 1 to 3 months, 13 000 km² for 3 to 6 months, and some 4 000
km² for periods varying between 6 to 8 months. When the floods recede completely about 800
cienagas, with an area of 3 260 km² remain. The Atrato river, also in Colombia, extends over a
forested alluvial plain for the last 100 km of its course. The plain, which covers 5 300 km², is
interspersed with numerous cienagas. Of similar size is the 5 000 km² of savannah flooded by the
Catatumbo river as it flows into Lake Maracaibo.
Several vast areas subject to sheet flooding are found on the continent. The greatest of these is
probably the Gran Pantanal of the Paraguay river whose shallow interconnecting complex of lakes

43
extends over 80–100 000 km² at peak floods. The savannah “llanos” of Colombia and Venezuela are
also subjected annually to shallow sheet flooding. This area is drained by the Meta, Arauca,
Capanaparo and Apure rivers which combine into a more deeply inundated deltaic floodplain of over
70 000 km² at their confluence with the Orinoco. The Rupununi river annually floods a very variable
area of savannah to a depth of 1 to 2 m. In exceptionally wet years the flooded area extends from the
headwaters of the Essequibo river to the Tokutu and Ireng rivers, which flow via the Rio Branco into
the Amazon. These sheet flooded plains have in common the shallowness and extensiveness of their
inundated area, their numerous anastomosing channels and small temporary lagoons.

44
Figure 1.15 Location of the major rivers and floodplains of South America:
1. Usumacinta/Grijalva 5. Orinoco 9. Paraguay
2. Atrato 6. Rupununi 10. Parana
3. Magdalena 7. Amazon 10. Parana

45
4. Catatumbo 8. Sao Francisco
The Amazon river and its major tributaries have densely forested floodplains for much of their length.
However, the main river channels are fringed by, and enclose, vast alluvial plains up to 50 km wide in
the upper reaches and 100 km wide further downstream. The Amazonian flood zone reaches its
greatest extent in the central delta located between the confluence of the Amazon and Tapajos. The
heart of this area, the Ilha Tupinamborana covers about 50 000 km². A similar combination of fringing
floodplain and internal delta is found along the Paraguay river below the Gran Pantanal and along the
Paraná river after its confluence with the Paraguay.
Coastal deltaic floodplains are uncommon in South America, although both the Orinoco and the
Amazon have densely forested terminal flood areas of 20 000 and 25 000 km² respectively.
ASIA (Fig. 1.16)
Asia is by far the largest and most geographically diverse land mass and its rivers cover a
correspondingly large number of types. The North flowing rivers of the Soviet Union, the Lena, the
Ob and the Yenisei, form a set of very large rivers which, despite having rapids in their upper reaches,
flow for most of their courses through plains which at their Northern extent lie within the permafrost
zone. These lower reaches remain frozen for 6–7 months of the year, and this produces a damming
effect where the still frozen lower reaches block waters released by the earlier break-up of the ice and
the snow melt in the upper reaches to cause extensive flooding. These floodplains, which may be
termed “tundra Floodplains”, differ from the temperate and tropical floodplains in that their water
bodies freeze completely for a considerable period of the year, and during the summer they retain a
marshy character.
The Tigris-Euphrates river system is isolated from the rest of Asia by the arid tracts of Iran and
Afghanistan. The upper courses of both rivers are torrential arising in the mountainous regions of
Turkey and Syria, but an extensive floodplain is present in the Mesopotamic region between them.
Here despite several thousand years of irrigation, associated with some of the most antique
civilizations, which have left many traces, flooding still continues over the 20,000 km² at high water
and the river channels are then confluent with the marshes. At low water, the lake area is reduced to
about 5 000 km². The largest lake in the system, the 5,200 km² L. Hammar, serves as a terminal drain
(Al Hamed, 1966).

46
Figure 1.16 Location of the major rivers and floodplains of Asia:
1. Amur 8. Indus 15. Irawaddy
2. Lena 9. Narmada 16. Chao Phrya
3. Yenisei 10. Ganges 17. Mekong
4. Ob 11. Krishna 18. Sui Chiang
5. Syr Darya 12. Godavari 19. Yangtze
6. Amu Darya 13. Brahmaputra 20. Hwang Ho
7. Tigris/Euphrates 14. Gangetic Delta
The hydrography of most of Eastern and South Eastern Asia is dominated by the central mountain
massif of the Himalayas and the Tibetan plateau. Most of the great rivers of China, India, and the
South East Asian Peninsula take their source in these highlands to flow out through a series of coastal
plains from China to Pakistan. In China, the Hwang Ho, Yangtze and Si rivers, drain the plateau
eastwards and flow across vast plains which were once liable to widespread and repetitive flooding.
The Yangtze in particular, is associated with an extensive complex of flood lakes. These systems are
now highly controlled by dams and other flood control devices so inundations are less common. The
Red, Mekong, Salween, Brahmaputra, Ganges and Indus rivers all rise on the plateau and have very
long torrential upper courses before debouching onto extensive lowland floodplains. The Indus river,
the site of very early civilization, has a wide braided channel for much of its course. This periodically
shifts its bed due to excessive silt deposition raising the river channel and its levees high above the
level of the surrounding plain. When it overspills its banks the course of the Indus may be displaced
laterally for up to 40 km, losing water by seepage and evaporation in the otherwise desertic region.
The river flows into the sea through a large but infertile deltaic plain. In the virtually rainless areas of
Pakistan, the Indus and its floods are essential for irrigated agriculture. Both the Ganges and the

47
Brahmaputra rivers also have unstable beds flowing through braided channels. The main fringing
floodplain of the Ganges is over 600 km long and between 16 and 80 km wide. The Brahmaputra has
an especially broad channel, up to 12 km in places. The two rivers combine to form an immense delta
liable to sheet flooding from rainfall as well as river discharge. The types of floodplain, classified by
the duration and depth of inundation of the portion of this plain lying in Bangladesh, are shown in Fig.
1.17. In fact most of the active part of this delta lies in this country as the western region, centred on
the Hooghly river in India, is silted and rarely floods through the now dead distributaries. In
Bangladesh alone, there are 93 000 km² of floodable land including 28 340 km² of paddy fields which
are inundated for 3–4 months of the year. In addition, there are an estimated 14,000 km² of permanent
open inland waters. The seaward parts of the plain have dense brackish water mangrove forests know
as “Sunderbuns” some 5 000 km² in area.
The 31 000 km² deltaic alluvial plain of the Irawaddy river is flooded both by rainfall and upper river
discharge. Submergence by local precipitation precedes the arrival of the river floods by at least a
month. Artificial levees constructed for flood control will eventually change the whole pattern of
flooding. A similar but smaller deltaic plain flanks the Chao Phrya river and its distributaries in
Thailand (Fig. 1.6). Here the main river channels are raised high above the surrounding backswamps
which are deeply flooded each year, although recently the construction of an upstream dam seeks to
control the timing and extent of the inundation. Another plain that is fast being altered by flood
control measures, irrigation and an ambitious programme of upstream dam construction is that of the
Mekong (Fig. 1.9) which is over 74 000 km² in total area. About 21 000 km² of its area are now no
longer flooded although 8 850 km of river, canals and irrigation channels take water to every part of
the plain. The area of permanent water is only 4 000 km², of which the majority is in the 2 500 km²
Grand Lac. In fact this body of water is one of the region’s most deeply affected by the modification
of the hydraulic regime of the river as its floods have been drastically curtailed.
The peninsular rivers of the Indian sub-continent, the Godavari, Krishna and Cauvery, once had
extensive floodplains as did the Mahaweli system in Sri Lanka. These systems have now been
modified by dams for power generation and irrigation.
Marshy flooded areas are common as the Indonesia island of Sumatra, Borneo and West Irian, and as
some of the Philippine islands. Fundamentally similar in nature these are located on the flat coastal
alluvial plains and are submerged and drained by numerous small rivers. In Sumatra, there are the
eastward flowing, Hari, Kampari, Rokan and Musi rivers; in Borneo the Southward flowing Kahajan,
Barito and Mendawa rivers and in New Guinea, the Fly, Diguil and Pulan rivers. The most complete
description of the fish and fisheries of such coastal flood rivers is that of Vaas et al. (1953) who make
it clear that, on the island of Sumatra at least, such areas are classic floodplains. In the Oga and
Komering rivers, which cover over 500 000 km at peak flood, the lateral plains of “Lebaks” are used
for both rice culture and fisheries.

48
Figure 1.17 The delta of the Ganges and Brahmaputra rivers in Bangladesh showing the major land
types classified by depth and duration of flooding
More typical seasonal floodplains, such as the 6 555 km² Kapuas lake district of West Borneo, the 7
000 km² lake district of the Mahakan River in East Borneo (Dunn and Otte, 1982) or the 7 500 km²
lateral plains of the Sepik River in Papua New Guinea, are also common on the larger islands.
EUROPE (Fig. 1.18)
Western Europe is drained by a number of relatively short streams although the Danube, Rhine,
Rhone and Vistula rivers have discharges sufficient to place them among the first 50 in the world. In
eastern Europe a number of larger rivers drain into the Black, Azov and Caspian seas including the
Dniester, Dnieper, Don and Ural. Torrential streams are present in the Alps, Pyrenees, Carpathian and
Caucasus Mountains, but on the whole European rivers are of the lowland type, particularly in the
East where the drainage is dominated by relatively few large rivers. On the whole European rivers

49
have been highly modified by canalization and dam building so that few retain much trace of their
original floodplains. For example, the largest river in Europe, the Volga, has been converted into a
cascade of reservoirs and only a very short terminal reach retains its original form, although even here
the flow from upstream is modified. The Danube and Ural are perhaps the only major rivers in which
some original characters survive (Liepolt 1967). The total area of the Danube floodplains was given
as 264 500 km² by Liepolt (1972), but recent damming, flood control and land reclamation schemes
have only left some 5 000 km² as liable to inundation in Romania. In the Czechoslovak-Hungarian
reach of the river 230 km² are regularly flooded from May to August and 30 km² remain as permanent
water in side arms and 86 km² in the main river channel. However, even these will disappear when
present plans for the hydraulic management of the system are completed.
Figure 1.18 Location of the major rivers and floodplains of Europe:
1. Vistula 6. Po 10. Dneister
2. Elbe 7. Rhone 11. Dneiper
3. Thames 8. Garonne 12. Don
4. Rhine 9. Danube 13. Volga
5. Loire
AUSTRALIA AND NEW ZEALAND (Fig. 1.19)
The hydrology of Australia is dominated by the aridity of its Central regions. Thus the Murray
Darling system which drains most of the inland South-Eastern Australia with a basin area of 1 029
000 km² has a remarkably small total annual discharge of 22 km² or 6% of the total discharge for the
continent. Its regime is very variable with long periods of low flow and occasional massive floods.
During its wetter phases the system is associated with a lowland marsh system which takes the form

50
of an internal delta with some permanent lakes, e.g., Lakes Meninolee and Tandou, although much of
the area may reduce considerably in size at times of drought. In contrast to this south flowing river
system there are several north flowing rivers which drain the more humid tropical areas of Northern
Australia and Queensland. These carry about 40% of the total discharge by Australian rivers and have
regular monsoonal regimes with annual floods which inundate the lowland plains associated with the
various rivers. New Zealand's rivers are mostly short and torrential particularly in the mountainous
South Island where they are often associated with deep narrow glacial lakes.
Figure 1.19 Location of the major rivers and floodplains of the East Indies and Australia:
1. Mahakan 4. Fly
2. Barito 5. Sepik
3. Murray/Darling 6. Kapuas

51
CHAPTER 2
PHYSICAL AND CHEMICAL PROCESSES
HYDROLOGY
FLOW
Variability of Flow Regimes
Flow regimes have their origin mainly in run-off from precipitation over the river basin. This may be
in the form of rainfall in warm temperate and tropical areas, in which case the response of the low
order streams to changes in precipitation is related directly to seasonality of pluviosity. In cold
temperate or high altitude regimes the precipitation is usually in the form of snow in which case
release of water into the stream channel is more closely related to seasonal patterns of snow-melt.
Retention and release of ground water, important factors in maintaining flow in the river channel
during the dry season, contribute less to spates and tend to smooth out flows, reducing the amplitude
but increasing the duration of a flood. The proportion of rain water that appears as run-off or as
ground water depends on the type of terrain and its vegetation cover. Local difference in these factors
can cause very different flood patterns even in two adjacent streams. Because of this, and because of
the rapidity with which run-off from local rain storms can affect low order streams, graphs of water
level or flow in the smaller water courses are apt to appear “spiky” with numerous rapid changes in
flood condition. As streams amalgamate into larger and higher order rivers their individual regimes
merge so the flow is smoothed (Fig. 2.1) and reflects the average of the precipitations over a
progressively larger area. The fate of the flow thereafter depends on the form of the river basin and
here reservoir rivers have internal features which reduce the variability of the flow, distributing it
more evenly throughout the year and suppressing pronounced flood peaks. Such features as extensive
swamps, forests, lakes or even floodplains themselves, may store water for later release. Alternatively
the cumulative discharge of many tributaries arising in different latitudes and having correspondingly
diverse flood peaks may also act in this way. The lower reaches of rivers, therefore, may tend to take
on “reservoir” characteristics, a good example being perhaps the Congo River below Kisangani. Even
here there are two pronounced flood peaks (Marlier, 1973). The existence of swamp, forest and lakes
in the river basin do not by themselves ensure the complete smoothing of flow and many flood rivers
have all these features.
Velocity of flow
Although Leopold et al. (1964) maintain that the rate of flow remains more or less constant as one
progresses downstream in rivers in the United States, it is the more general observation that flow rate
decreases with decreasing slope. Thus lower order rivers tend to have faster flows consistent with the
greater slope of upland streams, whereas the higher order streams have slow flows in the meandering
lowland course. Some higher order rivers however, such as the Magdalena of Colombia or the Sepik
of Papua New Guinea, do have considerable flow velocities in their potamon reaches with main
channel rates exceeding 2–3 m/sec at low water. By contrast flow may cease completely in low order
streams or even main channels during the dry season, and in many modified rivers abstraction of
water for irrigation has reached such a level that there is rarely any net outflow to the sea. Obviously
it is difficult to generalize as to the speed of current in various streams as it depends on many physical
and climatic variables, but the range of velocities quoted by Zhadin and Gerd (1961) for Middle Asian
rivers of the Soviet Union serves to illustrate the general slowing of the flow as the river proceeds
downstream. Here flow velocity were 5–6 m/sec in the upper reaches, 3–3.5 m/sec in the foothills and
2–2.5 m/sec in the flatland reaches. Furthermore, velocities change over relatively small stretches of
stream especially in the rhithron. For example, the rate of flow in Caucasian streams at low water was
1.25 m/sec over the riffles and only 0.6 m/sec through the succeeding pools.

52
Figure 2.1 Flood regime of rivers with different basin areas within the Chari-Logone river system
showing the increasing smoothing of the flood curve with increasing size of basin
It is very important for living aquatic organisms within the river channel that the velocity of the water
does not increase proportionally with discharge. Leopold and Maddock (1953) and Drury (1969) both
present graphs which show that the mean velocity at any point on a river only doubles for a tenfold
increase in discharge. Furthermore, since these figures apply to peak velocities at mid-channel the
increases in velocity at the bottom or sides of the channel would be even less.
Same idea of velocity of flow in any reach can be obtained from the size of particles in the stream
bed. Fig. 2.7 sets out the mean current velocity required to just move particles of various sizes along a

53
stream, thus the existing bed deposits should indicate that flows are just below the equivalent rate
listed. This serves only as a rough guide to the mean velocity which might be expected, as results
from various authors do not agree all that closely. Furthermore, resistance to movement is not entirely
a function of particle size for hard packed clay is far less easily entrained than is sand, even though its
particle size is far smaller.
Origins of flooding
Flooding originates from three sources: (i) overspill from the river channel; (ii) local rainfall; and (iii)
tides.
Overspill from the river channel
As flow increases a point is reached where the channel is no longer able to drain the volume of water
passing down the river. Further rises in discharge above this point (bankfull level) result in overspill
onto the floodplain. Here, because of the flatness of the terrain, increases in volume are achieved by
lateral expansion rather than by increases in depth, and the water spreads slowly and diffusely
outwards, hampered in its progress by the floodplain vegetation. Such flow is termed “creeping flow”
and is depositional (i.e., silt comes out of suspension onto the plain), as opposed to the erosive flow
where a strong directional current is confined within the main channel or along the terrace walls.

54

55
Figure 2.2 Flood regimes of typical river systems for various continents
Plots of the variation of water level or flow of a flood against time (flood curves) for large rivers have
the appearance of waves (Fig. 2.2), which usually are unimodal, although downstream of the
confluence of two systems having differing flood regimes, polymodal floods may occur. The flood
wave has amplitude (the total difference in level between maximum and minimum discharge) and
duration, (the time taken to pass from one low water stage to the next). It moves down valley at a
speed which may depend on many factors, including the slope and degree of enclosure of the river
valley, the type of vegetation and the intensity of the flood. In the Niger River, for instance (Fig.
2.3A), the flood crest takes over 100 days to move from Koulikoro to Malanville. As this is over 1
160 km, the average rate of travel is about 17 km/day. Similar speeds of transmission are found in the
Senegal, where the 620 km from Galougo to Dagana take over a month (17 km/day) (Fig. 2.3B) and
the Chari, where the 745 km between Mossala and N'Djamena are traversed in 42 days (18 km/day)
(Fig. 2.3C). The flood moves down-valley at about 13 km/day in the Brahmaputra, 29 km/day in the
middle Mekong and on the lower Mekong flood-plain 13 km/day. On the Kafue plain the speed of
transmission is 11 km/day and on the Central Delta of the Niger 13 km/day. These figures are
averaged over considerable distances and speed differs within various reaches of the same river. In
general, the more expansive the plain, the less its slope and the greater its vegetation cover, the slower
will be the velocity of the flood crest. The progression of a flood wave across a flood-plain is now
especially easy to illustrate using satellite imagery. For example, four phases in the passage of the
flood over the Yaeres floodplain are shown in Fig. 2.4 (after Quensiere, 1981).
Figure 2.3 Time taken for the flood wave to traverse the distance between selected points in three
African river systems. The time taken for the flood wave to cover the distance between fixed

56
points is indicated by the vertical lines showing the time of maximum flood and the line
transposed from the preceding point upstream as shown in the graph; figures refer to the
distance between one point and the next
In long rivers, where the timing of the rains is fairly uniform along their courses, the arrival of the
main flood crest in downstream reaches may be delayed until the dry season. Such a delay is typical
of the Niger River, where two floods occur at Malanville (Benin) (Fig. 2.3), one in August-November
caused by rainfall and drainage of local rivers and a second in December-March due to the arrival of
the flood from the headwaters of the river.

57
Figure 2.4 Successive stages in the flooding of the Yaeres by overspill from the Logone river (after
Quensiere, 1981). ---- : Limits of total area flooded
As the flood wave passes along the floodplain there is a tendency for it to decrease in amplitude and
increase in duration, provided no large amounts of water are added to it by other tributaries. This
process is best seen on a river such as the Niger where the channel runs for a considerable distance
through arid and semi-arid landscapes.
Local rainfall
Precipitation on the floodplain itself, or on the basins of the lower order streams immediately
surrounding it, saturates the soil and causes local flooding which often precedes the main flood by
some considerable period of time. Early work by Svensson (1933) showed that in the Gambia River
certain depressions are flooded only by local precipitation and are connected to the river and other
low lying swamps by a single channel or creek. Such swamps may be unaffected by variations in river
height, but fish may migrate to them by ascending the connecting channel. Early flooding of
floodplain depressions by rainwater has been noted from several systems. Elsewhere in Africa, Carey
(1971) described flooding of this type in the Kafue. The inundation of the Upper Chari (Bahr Salamat
and Bahr Azoum) appears to be almost entirely by local rainfall (Durand, pers.com.), and in the Nile
Sudd the swamps are filled one to two months before overspill from the river occurs. Extensive areas
of the Venezuelan Ilanos (Matthes, pers.comm.) and the Gran Pantanal or Paraguayan Chaco of the
Paraguay River flood in this manner, (Carter and Beadle, 1930). A similar type of flooding has been
recorded from the Irrawaddy River in Burma by Ohya, (1966), where submersion by rainwater
precedes the main floods by about one month. Flooding by local rainfall differs from overspill in that
the net flow of the water is from the floodplain depression to the river rather than the other way round.
Flooding with rainwater prepares the floodplain by saturating the soil and filling flood-plain
depressions and replenishing the ground water so that relatively little of the subsequent overspill flood
is absorbed by the ground.
Tides
Flooding caused by tidal action obviously affects only those parts of floodplains adjacent to the coast.
However, secondary effects are produced by the backing up of freshwater floods by high tidal levels.
Volker (1966) divided river estuaries into four reaches (Fig. 2.5); Zone A where vertical tides produce
a reversal of the direction of flow and the penetration of saline water into the river channel, or onto
the floodplain; Zone B where the river water remains fresh but tidal effects and current reversals still
occur; Zone C where the water levels are still affected by the tides giving rise to differences in current
velocity, although the flow always proceeds downstream and Zone D where water level and flow are
affected by upstream discharge only; this zone merges into the main river upstream.

58
Figure 2.5 Zones of interaction between marine and freshwater in estuaries and coastal deltaic
floodplains (after Volker, 1966)
Water balance on floodplains
The flood curve at any point in the river, results from discharge transmitted from upstream, input from
local tributaries and precipitation in the immediate area. This can be represented diagrammatically as
in Fig. 2.6 where the x axis is time. The y axis may represent any of the indicators of water quantity,
the most common of which are water level, discharge rate or storage volume. Although there is a
continuous fluctuation in water level the flood can be broken into two parts. In the first, the water is
confined in the river channel and alterations in discharge produce changes in depth and velocity.
Changes in volume are caused mainly by increase in depth so long as the flow remains confined in the
channel are caused mainly by increases in depth. This phase is delimited by the bankfull stage beyond
which the water overflows the channel limits and floods the plain. Thereafter changes in inflow
produce relatively little change in outflow, the excess water being temporarily stored on the floodplain
where the volume increases mainly from the extension of the water area. The bankfull level is usually
difficult to define precisely, as it varies from place to place in the river and also from year to year. The
detailed hydraulics of floodplains are extremely complex and beyond the scope of this discussion.
They have been the subject of many studies and attempts at mathematical modelling such as those
elaborated by Weiss and Midgley (1975 and 1976).

59
Figure 2.6 Diagram of (A) a generalized annual flood regime, and (B) probability of occurrence of
maximum and minimum water stages
The intensity of flooding varies from year to year in response to fluctuations in rainfall. Because of
this variability, a distribution of the frequency with which any flood height occurs can be established
over a number of years. Such distributions (Fig. 2.6) may be normal but are more often skewed. The
number of years which must elapse on average between a flood of a particular height and the next
flood of the same height is known as the return period. The 100 year flood, for example, is that level
of flooding which can be expected once in 100 years. The probability of occurrence of any flood is the
reciprocal of its return period; the 100 year flood having a probability of occurrence of 0.01 in any
year.
Not only does the intensity of flooding vary from year to year, but there is increasing evidence that the
variability is of a cyclic nature which is traceable to long-term global climatic patterns (Winstanly,
1975; Bryson, 1974). According to such cycles, years of exceptionally high flooding or of drought
tend to be grouped together so there is a good chance of two or more years of intense flooding to
appear in sequence or for one year of low flood intensity to be followed immediately by another poor
year. When the latter happens drought conditions can ensue such as was shown in the Sahelian rivers
between 1970 and 74 and again in 1980–85. The grouping of good and poor floods in this manner can
have severe consequences on the ecology of the fish as we shall see in the section on the dynamics of
floodplain communities. The variation in flood level is a normal phenomenon and knowledge of the
range as well as the mean value is essential to an understanding of the ecology of these systems.
Differences in the amplitude of the flood curves as measured by maximum and minimum discharge
rate are shown in Table 2.1. These are very variable and especially high values tending to infinity are
obtained in those rivers where flow virtually stops during the dry season. An example of the effects of
such variations on the actual areas flooded is that of the Senegal River as shown in Table 2.2.

60
Table 2.3 shows the mean rainfall over certain floodplains. In the tropics and sub-tropics, rainfall
makes only a moderate contribution to the water balance of the plain and is largely exceeded by the
evapotranspiration rates shown in Table 2.4. This excess of evaporation over precipitation causes the
net outflow from some floodplains to be very much lower than the inflow. Thus, of the 7.11 × 1010 m³
of water entering the Central Delta of the Niger through the Niger and Bani rivers in years of normal
flow, only 3.82 × 1010 m³ emerges at Dire, a net loss of 46 percent. Similarly, 2.7 × 1010 m³ of water
enter the Sudd at Bor and only 1.4 × 1010 m³ leave it at Lake No, a loss of 48 percent of the water. In a
few systems, the Okavango swamps for instance, evaporation almost balances inflow over the year so
that there is no appreciable outflow from the system.
Table 2.1
Mean annual maximum and minimum discharge rates of various rivers (from
figures compiled by Van der Leeden, 1975)
Discharge m³ sec.-1
Basin area Mean Mean Max/
(km²) max. min. min.
Amazon(Obidos) 4 688 000 207 400 89 300 2.3
Brahmaputra (whole basin) 580 000 72 460 2 680 27.0
Mekong (whole basin) 795 000 67 000 1 250 53.6
Ganges (whole basin) 977 500 61 200 1 170 52.3
Orinoco (Ciudad Bolivar) 850 000 46 200 7 520 6.1
Indus (whole basin) 970 000 31 200 490 63.7
Mississippi(Tarbet landings) 3 928 000 30 800 7 930 3.9
Paraná (whole basin) 2 039 000 25 130 6 300 4.0
Magdalena (Calamar) 238 000 10 000 3 900 2.6
Oubangui (Bangui) 500 000 9 640 1 070 9.0
Blue Nile (Khartoum) 325 000 5 950 138 43.1
Senegal (Bakel) 218 000 3 430 26 131.9
Chari (N'Djamena) 600 000 3 390 199 17.0
Chao Phrya (Nakhon Sawan) 110 371 3 363 32 105.1
Niger (Mopti) 281 600 2 820 67 42.1
Danube (Bratislava) 131 338 2 746 1 402 2.0
Tigris (Baghdad) 134 000 2 720 286 99.5
Benue (Garoua) 64 000 1 920 32 60.0
Ouémé(Pont du Save) 23 600 758 0.63 1203.0
Table 2.2
Areas flooded by the Senegal River for different flood regimes (after
OMVS)
Year Statistical significance Maximum area flooded (ha)
1964 Five-year strong flood 766 000
1969 Mean flood 549 000
1926 Five-year weak flood 215 000
1972 Weakest recorded flood 104 000

61
The high evaporation rates also mean that, to be permanent, pools or lagoons must either be filled
annually to a depth greater than the loss by evapotranspiration (between 1.2 and 2.2 m from the data
in Table 2.3), or must receive inflow from ground water.
Table 2.3
Precipitation at sample locations on various rivers (from Van der Leeden, 1975)
Annual precipitation (mm)
J F M A M J J A S O N D Mean
AFRICA
Mali (Bamako) - - 0.4 15.2 37.7 137.2 348.0 205.7 43.2 15.2 - - 1120.1
Sudan (Wau) - 5.1 22.9 66.0 134.6 165.1 190.5 208.3 167.6 124.5 15.2 - 1099.8
Botswana (Maun) 106.7 8.7 71.1 17.8 5.1 2.5 - - - 22.9 5.84 8.64 434.3
Zambia (Lusaka) 235.1 190.5 142.2 17.8 - - - - - 10.2 91.4 149.9 835.7
ASIA
Dem. Kampuchea 7.6 10.2 25.4 78.7 144.8 147.3 152.4 154.9 226.1 251.5 139.7 43.2 1391.9
(Phnom Penh)
Thailand 5.1 27.9 27.9 58.4 132.1 152.4 175.3 233.7 233.6 355.6 45.7 2.5 1470.7
(Bangkok)
Bangladesh 7.6 30.5 61.0 137.2 243.8 315.0 330.2 337.8 248.9 134.6 25.4 5.1 1877.1
(Dacca)
Pakistan 63.5 63.5 68.6 48.3 33.0 58.4 205.7 233.7 99.1 15.2 7.6 30.5 927.1
(Rawalpindi)
SOUTH AMERICA
Venezuela 35.6 20.3 17.8 25.6 96.5 139.7 160.0 180.3 91.4 101.6 71.1 33.0 972.8
(Ciudad Bolivar)
Colombia 10.2 - 10.2 22.9 86.4 86.4 76.2 18.2 12.9 274.3 226.1 114.3 934.7
(Cartagena)
Brazil 248.9 231.1 261.6 221.0 170.2 83.8 58.4 38.1 45.7 106.7 142.2 203.2 1811.0
(Manaus)
Argentina 78.7 78.7 99.1 124.5 66.0 30.5 30.5 40.6 61.0 71.1 94.0 114.3 889.0
(Parana)
Table 2.4
Annual evapotranspiration rates from various floodplain regions
Annual ETP mm
Mean Range
AFRICA
Niger delta (Mali) 2 150 1 900–2 450
Sudd (Sudan) 1 900 1 800–2 000
Okavango swamp (Botswana) 1 800 1 700–1 900
Kafue flats (Zambia) 1 650 1 550–1 750
ASIA

62
Mekong delta (Viet Nam) 1 600 1500–1 700
Chao Phrya (Thailand) 1 600 1 500–1 700
Ganges delta (Bangladesh) 1 500 1 400–1 600
Indus valley ± 30°N (Pakistan) 1 750 1 600–1 900
Indus valley ± 25°N 2 000 1 900–2 300
SOUTH AMERICA
Llanos (Colombia) 1 400 1 300–1 600
Magdalena ± 5°N (Colombia) 1 500 1 400–1 600
± 8°N (Colombia) 1 300 1 100–1 400
Amazon river - Manaus (Brazil) 1 300 1 200–1 400
Parana river (Argentina) 1 250 1 150–1 350
CHEMISTRY
SEDIMENT LOAD AND TURBIDITY
There is a clear relationship between the velocity of the current in a river and the amount of matter
carried suspended in its waters - the greater the current, the larger the load and particle size (see Fig.
2.7) that can be carried by the stream. This has many repercussions both for the morphology of the
river and in the biology of many of the organisms living therein. Morphologically, because any river
will tend to transport the maximum amount of material in suspension for any given current, sediment
will be picked up or deposited as the current and the existing load vary. Slowing of flow results in
aggradation, the deposition of part of the load leading to the development of many of the
morphological features of the river, including levees, point bars and floodplains. Acceleration of the
flow leads to the river picking up sediment causing erosion or degradation. During the flood cycle
these two processes tend to alternate. Thus during the rising flood rivers are heavily charged with silt,
but as the water spills over the flood-plain the general slackening of flow leads to a deposition of
suspended matter producing decanted waters of some clarity. At falling flood acceleration in water
velocity causes localized scour but as flow slackens progressively through the low water period the
standing waters of the floodplain, and frequently of the main channel itself, deposit most of their load.
The amount of suspended matter in the water affects the penetration of light into the water, for
example Bonetto (1980) found a relationship S = a e bh for the Parana river, where S is the light
penetration as measured by Secchi disk, h is the velocity of the current as measured by the height of
the river and a and b are constants.
Sediment yields of representative rivers of the world are shown in Table 2.5. The suspended matter
carried by a river relative to its discharge is indicative of the stability of the system. Rivers which
carry a large amount of sediment relative to their discharge, for example the Indus, tend to be unstable
with shifting beds and rapidly evolving successions of floodplain feature, whereas rivers with low
sediment yields relative to their discharge tend to change much more slowly.

63
Figure 2.7 The speeds of flow required to move mineral particles of various sizes (line plotted from
data in Nielson, 1950); terminology of particle sizes from Gregory and Walling (1973)
NUTRIENT SPIRALING
The idea of nutrient spiralling posits that a nutrient atom cycles through three generalized components
of the stream ecosystem (Newbold et al., 1981). Each component is transported downstream with
current so the path of the atom describes an imaginary spiral. The three components are: (a) the water
in which the nutrient atom is dissolved; (b) the particulate phase in which the atom is sorbed onto
particulate matter and is thus located in the bottom although downstream transport continues through
saltation, suspension, erosion, etc.; (c) the consumer phase in which the atom is incorporated into
living matter. During this phase it may pass directly through a series of organisms before its release
once more into the water phase. The total distance travelled downstream between one release into the
water and the next, is termed the spiralling length (S). This length may be taken as a measure of the
efficiency with which nutrients are used within any particular system, the shorter the spiralling length

64
the more efficient the use in that any nutrient atom is utilized more frequently for any given length of
stream.
During the consumer phase, most of the nutrients are held in micro-organisms, but invertebrates
grazing in these and on higher plants can influence the spiralling length through accelerated release of
nutrients. Collector and shredder organisms tend to shorten spiralling length whereas grazers may
increase it.
Table 2.5
Representative rivers of the world and their sediment yields
Drainage area Average annual suspended load
River km² Total t × 106 t/km² Load/discharge
Amazon 6 062 080 406.4 67 1.92
Negro 93 875 1.5 16
Congo 3 968 000 72.4 18 1.82
Mississippi 3 184 640 349.5 109 20.0
Missouri 1 352 192 243.4 180
Ohio 196 044 15.2 77
Nile 2 944 000 123.8 42 44.2
Parana 2 278 400 91.4 40 6.13
Uruguay 384 000 15.2 39
Volga 1 335 014 21.1 16
Indus 1 231 360 488.7 396 87.3
Niger 1 100 800 5.1 5 0.8
Murray-Darling 1 059 840 35.7 33
Ganges 1 047 552 1625.6 1551 86.9
Brahmaputra 552 960 812.9 1469 41.0
Yangtze 1 012 864 560.8 553 25.7
Orinoco 938 752 96.9 103 5.7
Danube 806 400 21.8 27 3.5
Mekong 386 560 190.0 491 17.3
Irrawaddy 362 752 336.3 927 24.9
Vistula 190 873 1.7 9 1.55
Euphrates 119 219 4.8 40 3.4
Chao Phrya 105 216 12.7 120
CARBON AND ORGANIC MATERIAL
Rivers transport large, but usually not accurately quantified, amounts of dissolved and particulate
organic material from the terrestrial component of the drainage basin to the oceans (See Fig. 2.8).
Direct inputs usually consist of coarse particulate matter (CPOM) in the form of leaves, fruit, flowers,
pollen, branches, etc. whereas indirect inputs consist of leachates and other dissolved material
including inorganic carbonates from rocks and soils. Biological processes downstream tend to degrade
the particulate organic matter from CPOM (particle size full size trees to 1 mm) through fine
particulate organic matter (FPOM - particle size 50 mm - 1 mm) to ultrafine particulate organic matter
(UPOM - particle size 0.5 - 50 mm). Dissolved compounds increase in concentration down-stream

65
and a wide range of amino acids, fatty acids, sugars, hydrocarbons and pigments have been recorded
by Visser (1970) and Balogun (1970), either free in the water or absorbed onto sediment particles. At
the same time living organism contribute to the CO2 concentration through respiration.
Particulate carbon and compounds absorbed onto sediment particles follow similar processes to
sediment and other nutrients in that they may be stored within the plain through deposition, may be
recycled by biological agents and may be reincorporated into the flow towards the oceans through
erosion, (Fig. 2.9). Storage rates may be quite high. For example, in the floodplains of small rivers of
the forested zones of North America storage can amount to between 48 and 96 g C/m/yr (Sedell et al.,
1974) and in mature floodplain swamps can be as high as the 113 g C/m/yr recorded from the
Atchafalaya river (Gagliano and Van Beek, 1975).

66
Figure 2.8 A general model of the biogeochemistry of carbon in rivers (from Richey, 1981)
IONIC COMPOSITION
The chemical composition of river water depends on a wide variety of physical, chemical and
biological features, but three basic mechanisms control surface-water chemistry: precipitation, the
nature of the bedrock and the evaporation-crystallization process (Gibbs, 1970) (Fig. 2.10). In most of
the large tropical rivers the ionic composition of the water derives primarily from the rain and the rock
or sediments over which the river flows. In rivers flowing through hot arid regions, the rate of
evaporation and crystallization of the concentrated salts exerts the main influence and flood areas
located in such zones eventually transform into salt marsh habitats. Secondary influences on the ionic
composition are exerted by macrophytes and phytoplankton and to an increasing extent by the various
industrial, agricultural and domestic human activities.
Figure 2.9 Organic carbon flow between a floodplain wetland and its bordering river (Mulholland,
1981)

67
Figure 2.10 Diagram of the basic mechanism controlling surface-water chemistry (Adapted from
Gibbs, 1970)
Total dissolved solids (TDS) may be taken as one convenient measure of the total ionic concentration
in water. Naturally, the concentration and relative abundance of ions in river water is highly variable,
although there is a tendency for larger rivers to average out the variability of the tributaries. Fig. 2.11,
taken from Holland (1978), shows clearly that, while there is considerable spread in TDS for any
particular runoff the increase in concentration with decreasing runoff is unmistakeable.

68
Figure 2.11 Total dissolved solids in some major rivers as a function of the mean annual runoff (after
Holland, 1978)
The mean composition of the river waters of the various continents of the world are shown in Table
2.6. Holland (1978) discusses in more detail the concentration and origin of most of the major ions
present in these rivers.
Table 2.6
Mean ionic composition of river water of the continent of the world (after Holland, 1978)
Continent HCO SO Cl NO Ca Mg Na K Fe SiO Total
America N. 68.00 20.00 8.00 1.00 21.00 5.00 9.00 1.40 0.16 9.00 142.00
America S. 31.00 4.80 4.90 0.70 7.20 1.50 4.00 2.00 1.40 11.90 69.00
Europe 95.00 24.00 6.90 3.70 31.10 5.60 5.40 1.70 0.80 7.50 182.00
Asia 79.00 8.40 8.70 0.70 18.40 5.60 9.30 - 0.01 11.70 142.00
Africa 43.00 13.50 12.10 0.80 12.50 3.80 11.00 - 1.30 23.20 121.00
Australia 31.60 2.60 10.00 0.05 3.90 2.70 2.90 1.40 0.30 3.90 59.00
World 58.40 11.20 7.80 1.00 15.00 4.10 6.30 2.30 0.67 13.10 120.00
Anions 0.96 0.23 0.22 0.02 1.43
Cations 0.75 0.34 0.27 0.06 1.43

69
Detailed lists of ionic content of individual tropical rivers are outside the scope of this book.
Compilations of such data based on the work of numerous authors exist for Africa (Welcomme, 1972)
and Latin America (Ardizzone and Ziesler, 1977). Observations from Asia have not been compiled
systematically, but Jhingran (1975) cited data from Indian rivers and Johnson (1967) gave details of
the chemistry of some Malaysian rivers. Studies on the significance of ionic transport on a world scale
have been made by Meybeck (1976 and 1979) and Martin and Meybeck (1976 and 1979).
In those regions where the rocks are impoverished and soils leached, the main supply of nutrients
comes from precipitation, such rivers being grouped in the lower right hand area of Fig. 2.10. In the
Amazon basin for instance, the chemical composition of the blackwater streams of the Hylaea, or of
the larger blackwater rivers, can resemble that of the rainwater. The frequent similarity of the
composition of rain and river waters indicates that such ions as are leached from the soil may be
balanced by those carried by the rain. Similarly the chemistry of the headwaters of the Paraná and
Paraguay rivers are dominated by atmospheric precipitation (Bonetto, 1975) and in the same general
area the conductivity of rainwater falling over the Mato Grosso, Brazil, is often higher than that of the
streams (Green, 1970). In Africa, Visser (1974) noted that the composition of river and lake waters
within a radius of 100 miles of Kampala indicated that rain contributed on average all of the sodium,
ammonium and nitrate, over 75 percent of the chloride and over 70 percent of the potassium. In the
Gombak River, Malaysia, Bishop (1973) considered precipitation as a very important factor in
determining ionic concentrations.
The rivers of the three main tropical rainforest regions arise on very poor, leached, podsolitic soils.
Sioli (1968) described such waters in the Amazon basin as more or less transparent, devoid of
significant amounts of inorganic particles, but with very poor light penetration because of the brown
colour imparted by dissolved humic substances. PH is usually very low and dissolved nutrient
concentration poor. More detailed analyses of the composition of blackwaters of L. Tupe of the Rio
Negro (Rai and Hill, 1981) show such waters to be biased to sodium and sulphate dominance. Similar
waters have been reported from the headwaters of the Orinoco (Edwards and Thorne, 1970) and have
also been described from both Africa (Matthes, 1964) and Malaysia (Johnson, 1968). Descriptions
and analyses from the three continents are shown in Table 2.7.
Table 2.7
Description of chemical composition of blackwaters from Asia, Africa and South Americaa
Congo (Tshuapa
R.) (Matthes, 1964) Amazon (Rio Negro)
(Ungemach, 1972) Malaysia
(Johnson, 1967) Amazonian rainwater
(Ungemach, 1971)
pH 4.0–6.5 4.6–5.2 3.6–5.9 4.0–5.4
Conductivity 24–32 6.8–10.4 1.5–20
TDS 30–65 mg 1-1
Fe fairly abundant 268–535 μg 1-1 10–100 μg 1-1
+Ca very low 232–450 μg 1-1 0–1 904 μg 1-1 20–500 μg l-1
+Mg very low 108–254 μg 1-1 182–17 867 μg 1-1 10–30 μg 1-1
+Na 435–1 358 μg l-1 2 092–22 480 μg1-
1
+K 235–601 μg 1-1 586–7 429 μg 1-1
NNO3 trace 15–53 μg 1-1 5–300 μg 1-1
NNO2 trace 0.035–1.27 ug 1-1
PPO4 trace 3.2–8.6 μg 1-1 1–3 μg 1-1
N organic 70–80 μg 1-1 209–478 μg 1-1 10–150 μg l-1
Cl 1 800–2 600 μg 1-1 283–30 631 ug 1-1 80–320 μg1-1
Si 2 000–2 740 μg 1-1 2 000–2 740 μg1-1
SO4 absent absent 4 803–216 903 μg
1-1

70
A These figures are often based on only a few observations and do not therefore necessarily reflect
the full range of variations of the parameters measured
Blackwaters from the three continental areas are similar in general composition, except that sulphate
is even more common in the Malaysian waters than in those of South America and Africa. In South
America it is virtually absent in many blackwaters and in Africa is present only in those waters
flowing from volcanic regions. Johnson (1968) described an aberrant type of blackwater from
Malaysia which, by flowing over limestone, acquires higher total ionic concentrations of between
150–700 mg/l, accounting for the relatively high maximum values in Table 2.7. The “Gelam” type
waters also had very low PH values of 3.5–4.9 showing considerable excess sulphate. A third series of
blackwaters have become enriched by organic pollution and their PH was raised to 6 and above. This
series may indicate how blackwaters on other continents will react to pollution from urban sources as
human settlement of these areas becomes more intense.
In addition to the blackwaters, the Amazon and Orinoco are fed by clear water rivers. They originate
in the case of the Amazon from the massifs of central Brazil and in the Orinoco from the Guianian
shield. These waters are as poor in nutrients as the black-waters and have similarly low PH values of
between 4.0 and 6.6. They do not, however, have the dark staining by humic matter. A third category
of Amazonian rivers, the white water rivers, more closely resemble the general pattern of other
tropical rivers as described in the rest of this section and are the main source of nutrients in the basin
where, by the deposition of their heavy silt loads, they maintain the productivity of the várzea flood-
plains.
The lowland equatorial environments of the Amazon and the Congo give these two rivers many
common features. However, there are considerable differences between the two systems which arise
from the altitude and relief of the basins as well as from the geological formation and the vegetation
(Marlier, 1973). The forest is less extensive in the Congo and the main channel of the river is divided
into quiet and rapid reaches. Flooding is also less severe than in the Amazon.
CONDUCTIVITY
Conductivity is another measure of the total amount of ions present in a body of water and is a useful
approximation to chemical richness. As a measure, however, it does not give an indication of the
actual ionic composition of the water and may, therefore, fail to convey information on limiting
factors such as the lack of essential nutrients.
River waters are less variable than those of lakes and the conductivities of most major whitewater
rivers tend to resemble one another, with a few exceptions. In fact variations between tributaries
within any one river system are apt to be greater than those shown between different main river
channels. This is because the lower order streams are more sensitive to local geological and
vegetational patterns; whereas higher order channels average the conditions from a range of lower
order ones (Table 2.8).
Conductivity changes throughout the year within any one system. The broad, ionic composition of the
water is largely determined by the three processes mentioned in the introductory passage to this
section. The concentration of dissolved substances which include organic compounds not measured
by conductivity is influenced by five factors: (i) dilution effects, whereby flood or rainwater with
weak ionic concentration reduces conductivity; (ii) solution effects, the “shore factor” of Braun
(1952), whereby salts locked on previously dry land by decaying vegetation, animal dung, ash from
burnt vegetation etc., enter solution as the flood waters extend over larger areas; (iii) wash-out effects
whereby nutrient rich ground water is displaced into the river channel by increased infiltration; (iv)
concentration by evaporation and (v) absorption by living components of the system. Normally, these
effects combined tend to produce higher conductivities in the dry season than in the wet, both in
lagoons and in the river channels, giving an inverse relationship between water depth and

71
conductivity. The same trends have been recorded from the Oshun river (Egborge, 1971), the Ogun
river (Adebisi, 1981), the Kafue river (Tait, 1967; Carey, 1971; and University of Idaho et al., 1971),
the Senegal (Reizer, 1974) and the Zambezi (FAO/UN, 1969) in Africa. However, wash out of
nutrient rich ground waters may cause local rises in conductivity in small streams during the rains.
Several of these authors have also noted a secondary maximum of conductivity (due to solution
effects) as water invades the floodplain but this is of short duration.
Table 2.8
Conductivity and pH of some tropical river systemsa
Conductivity
(k20 μmhos cm-1)
ASIA
Gombak 29–41 6.6–7.0 Bishop, 1973
Mekong c. 280–630 6.9
Comm. for Coordination of
Investigations of the Lower Mekong
Basin
Meklong 8.2 Sidthimunka, 1970
SOUTH AMERICA
Magdalena system
Magdalena: mouth 490–630 6.2–7.2 Ducharme, 1975
above confluence 131 6.7–7.4 Ducharme, 1975
Cauca 250–500 6.5–7.6 Ducharme, 1975
San Jorge 320–350 6.5–6.6 Ducharme, 1975
Amazon system
Amazon/Solimoes 28.1–83.8 6.8–7.5 Schmidt, 1972
Maranon 129 Gessner, 1960
Ucayali 150 Gessner, 1960
Negrob 8.4–8.6 4.4–4.7 Schmidt, 1972
Tapajos 11.9–14.9 4.5–5.3 Schmidt, 1972
La Plata system
Paraná/Paraguay
alto 31.1–72.1 7.0–7.9 Bonetto, 1975/76
Paraná medio 112.0–184.0 7.2–7.7 Bonetto, 1975/76
Paraguay 69.7–335.0 6.9–8.2 Bonetto, 1975/76
AFRICA
Konkoure 22.1 5.9–6.2 Livingston, 1963
Bandama 90–200 6.7–7.6 Welcomme, 1972
Chari 22–73 6.9–7.7 Welcomme, 1972c
Logone 41–82 Welcomme, 1972c
Congo/Congo system
Congo main stem 37.1–76.7 5.5–6.5 Gosse, 1963
Ubangi 19.4–56 6.2–6.7 Micha, 1973
Luapula 150–180 6.2 Soulsby, 1959
Ruzizi 628 Marlier, 1951
Lualaba 145–255
Lt. Scarcies 35–55 7.1–7.4 Welcomme, 1972c
Gt. Scarcies 60 7.1 Welcomme, 1972c
Moa 36 6.6 Welcomme, 1972c
Oshun 57–96 Egborge, 1971
Niger 31–70 6.7–7.2 Daget, 1957
Sokoto 6.9–8.1 Holden & Green, 1960

72
Nile system
White Nile 220–500 8.0–8.9 Hammerton, 1972
Blue Nile 140–390 8.2–9.1 Hammerton, 1972
Kagera 93–99 Talling & Talling, 1965
Sobat 112 702 Talling, 1957
Bugungu Stream 245–395 7.1–7.8 Welcomme, 1969
Bahr-el/Ghazal 550 7.8 Talling, 1957
(L.No)
Semliki 400–910 Beauchamp, 1956
Orange 159 7.7 Keulder, 1970
Ouémé 60 Welcomme, pers.cos.
Ruaha 32–136 6.3–7.9 Petr, 1974/76
Senegal 72 6.8–7.1 Reizer, 1971
Volta system
Black Volta 41–124 6.5 Welcomme, 1972c
Red Volta 62 6.5 Welcomme, 1972c
White Volta 119 7.2 Welcomme, 1972c
Zambezi system
Kariba Lake 50–96 7.4 Coche, 1968
Upper course 57–126 FAO/UN, 1969
(Barotse plain)
Shire 220–450 7.5–8.8 Hastings,1972
Kafue 130–320 7.5–8.8 FAO/UN, 1968
Lower course 108–153 7.7–8.0 Hall et al., 1977
(Mozambique)
a These figures are often based on only a few observations and do not, therefore, necessarily reflect
the full range of the parameters measured
B Blackwater River
c Welcomme, 1972 compiled data received in response to a circular FAO questionnaire
The situation for Latin American rivers is less clear. Bonetto et al., 1978, show that, as may be
expected, conductivity in the Laguna La Brava rose during the low water period. Also, when floods
invade isolated floodplain lakes, they dilute the dissolved solids that have been concentrated during
the dry season, (Bonetto, Dioni and Pignalberi, 1969). Schmidt (1972) noted consistently higher ionic
concentrations during the dry season in the Amazon river as compared to the rains, and Junk (1973)
remarked on a similar fluctuation between high conductivities in the dry season and lower
conductivities in the flood in some floodplain lakes. However, both he and Schmidt (1972a) have also
described lakes in which the conductivity is minimal during the dry season and maximal during the
first phase of river inflow. The explanation advanced for this was that initial flooding by nutrient -rich
whitewaters, and solution of salts from inundated lands, gives high ionic concentration during periods
of increasing level. During the non-flood season water is diluted by drainage of nutrient-poor
blackwater from the forest streams and by rain-water.
In the littoral zone of the floodplain, represented by the vanguard of the advancing flood water,
nutrient-rich soil is freshly inundated and the solution of the salts produces a local increase in
conductivity. This is undoubtedly of great significance in the productivity patterns of floodplains
during the earlier part of the flood phase, when such shallow littoral zones must represent a
considerable proportion of the total area of the aquatic system.

73
HYDROGEN ION CONCENTRATION (PH)
Differences in water chemistry including pH, lead to variations in the type of aquatic organisms which
inhabit various waters. For instance distinctive communities of fish are particularly found associated
with highly acidic waters; these often have only low resistance to more elevated pHs.
Forest rivers, with their characteristics blackwaters and abundant humic materials, are slightly to very
acid with pHs ranging from 4 to neutrality. Savannah rivers are usually neutral or slightly alkaline
(Table 2.8). Because there is decaying vegetation on the bottom of most standing waters of the
floodplain, slight gradients often exist with higher pH values at the surface (Bonetto, 1975; Carey,
1971; Schmidt, 1973). Gradients in pH are also common at the water’s edge where the newly flooded
soil may produce either a local drop in pH, as in some African swamps, or a brusque rise in others.
The sudden rise in the pH of floodwaters on the Central Delta of the Niger, to as high as 8, has been
attributed by Blanc et al. (1955) to the effects of dung entering solution. Experimental evidence
confirmed that cow droppings raise the pH of water rapidly. The effects of this are, however,
temporary and more acid conditions are quickly restored. Laterite, which is a common soil in tropical
regions, and the peaty types of soil found on the surface of floodplains are both capable of reducing
pH (Mizuno and Mori, 1970). There is a tendency for swamps to become more acid as the dry season
progresses; Carey (1971) and Holden and Green (1960) particularly having noted that pH is lower in
swamps than in the river at this time. Drops in pH have also been noted by D'Aubenton (1963) under
the floating vegetation mat of the flooded forest of the Tonle Sap. By contrast conditions in some
open water lagoons can become increasingly alkaline as calcium salts are concentrated by evaporation
(Holden and Green, 1960). The lower pH values in swamps usually cause a general drop in pH
throughout the system when the acid waters are flushed out by rain or flood water with poor buffering
capacity early in the flood season.
Diurnal fluctuations in pH are primarily associated with the respiratory and photosynthetic activities
of the phytoplankton. A decrease in pH in the euphotic zone during the night, with a minimum early
in the morning, is a common feature of Amazonian várzea lakes (Schmidt, 1973). Similar trends can
reasonably be expected to occur throughout the day in the majority of floodplain lakes.
TEMPERATURE
Latitude, altitude degree of insolation, substrate composition, turbidity, ground or rainwater inflows,
wind, and vegetation cover, can all influence the temperature of water in rivers and floodplain lakes.
Generally, surface water temperatures follow the ambient air temperature fairly closely although
under dry hot conditions this is more likely to be correlated with air temperature minima due to the
cooling effect of evaporation. In the normal annual cycle in the tropics dry season water temperatures
are higher than those of the wet season. In the temperate zone winter temperatures are lower than
those in the summer and rivers may freeze for a considerable part of the year at higher latitudes and
altitudes. At relatively low latitudes, or at high altitudes in the tropics, temperatures may still fall
sufficiently to give conditions which are cold enough to cause growth checks and even fish
mortalities. In the Paraná River, for example, a combination of sub-zero temperatures and low water
caused massive mortalities in 1962 (Vidal, 1964).
Water in the main channel of a river rarely stratifies, as good mixing is maintained by the turbulence
associated with river flow. On the other hand stratified conditions are common in even very shallow
floodplain water bodies. Hastings (1972), for instance, recorded diurnal stratification in lagoons of the
South Elephant marsh (Fig. 2.12) which were only 1.6 m deep. Similarly, Sanchez (1961) and Junk
(1973) found 1–2 m deep lagoons from the Peruvian and Brazilian Amazonian várzea to be stratified
at least temporarily, and Ducharme (1975) (Fig. 2.12), Mikkola and Arias (1976), Arias (1975) and
Zarate and Cubides (1977) have all recorded stratified conditions from various cienagas of the
Magdalena system ranging from between 1.5–6 m depth. In the dry season the Amazonian Lago do
Castanho is only about 1.5 m deep, but clear stratification develops on windless days (Schmidt, 1973).

74
In flooded condition the lake is up to 11 m deep and stratification is more persistent, although the
temperature profiles are highly variable from one day to the next (Fig. 2.13) due to local changes in
conditions. The main day-to-day fluctuations occur in waters of less than 3.5 m depth, indicating that
only water down to this depth is normally affected by daytime heating. In the Paraná River, which is
subject to lower temperatures especially in winter and during the night, inverse stratification has been
noted by Bonetto, Dioni and Pignalberi (1969).
Figure 1.12 Diurnal cycle of temperature and dissolved oxygen in the Cienaga de Ayapel
(Magdalena River) 24 October 1974 (After Ducharme, 1975)

75
Figure 1.13 Daytime temperature fluctuations and changes in the stratification of Lago do Castanho
within short-time intervals during May 1969 (high-water). (After Schmidt, 1973)
It may be concluded that floodplain lakes not covered by vegetation normally stratify thermally
during at least part of the year. In shallower lakes stratification due to surface heating reaches a
maximum between 14.00 and 18.00 hrs, but tends to disappear at night or under the influence of
strong or persistent winds. The difference in density per degree at high temperatures is greater than
that at low temperature, for instance the drop in density between 29 and 30°C is about the same as that
between 4 and 10°C. For this reason at high temperature density differences are quite large for even
slight differences in temperature and the resistance to mixing is correspondingly high. In deep lagoons
stratification may last for several weeks or even months.
Water shaded by flooded or fringing forest tend to be cooler and more thermally uniform than that
exposed to direct sunlight, although thermal stratification has been described both from the flooded
forest fringing the Tonle Sap (D'Aubenton, 1963) and from Igapó forests of the lower Rio Negro
(Schmidt, 1976 and Geisler, 1969). Floating vegetation on the other hand gives rise to more varied
temperature conditions. Because of the restricted evaporation and high absorption of sunlight in calm
areas very high temperatures can be attained within masses of floating vegetation. Bonetto, Dioni and
Pignalberi (1969) found exceptional temperatures of up to 40°C in the surface waters of an oxbow
lake covered with Salvinia (Fig. 2.14). Similar temperatures have been found on the surface of Lago

76
Calado in stands of Paspalum repens by Junk (1973). Temperatures fall off more rapidly than they do
in open water due to shading by the vegetation. Floating mats of vegetation also encourage the
maintenance of stratified conditions by damping wind and wave action.
Figure 2.14 An extreme case of temperature accumulation in the waters under a bed of Salvinia at
midday. Solid line = open water; dotted line = water under vegetation. The arrow marks the
air temperature. (After Bonetto et al., 1969)
An important but little investigated habitat is the shallow open water littoral at the fringe of the flood
zone. At the time of rising water this represents the first contact between the advancing water and the
nutrient-rich dry soil. Because of its shallowness, daytime water temperatures may be very high and
gradients are set up between the very edge of the zone and the cooler water offshore (Fig. 2.15).
DISSOLVED OXYGEN
The distribution of dissolved oxygen within the aquatic system is one of the main factors influencing
the distribution of fish. In the rhithron waters are usually well oxygenated due to the turbulent flow
across the riffles. However, in the tropics these zones are apt to become separated into a series of
pools during the dry season when flow is minimal. At this time the pools may become deoxygenated
or even anoxic due to the decomposition of organic material, the oxygen demand of the fish and to
increased temperatures. In the Potamon the situation is more complex. As floodwaters invade the
floodplain of the Parana River there is an initial rise in dissolved oxygen concentrations in open water
and under floating vegetation. Similar effects have been noted from many other systems, particularly
the Sokoto (Holden and Green, 1960) and the Kafue (University of Idaho et al., 1971) in Africa, the
Paraguayan Chaco (Carter and Beadle, 1930) and the Magdalena (Arias, 1975; Zarate and Cubides,

77
1977) in South America. During the earlier part of the floods, flood or rainwater may flush
deoxygenated water out of depression lakes and swamps remaining from the preceding dry season.
There is, therefore, a tendency for dissolved oxygen levels to be low during the earlier period when
water is rising, although they remain fairly high throughout the major part of the floods. Higher
dissolved oxygen levels during the flood season are ascribed mostly to the aerating action of wind and
the mixing brought about by turbulence. In some deeper lakes, however, stratified conditions with low
oxygen tensions near the bottom may persist throughout the year. Vertical oxygen gradients have also
been observed in flooded Igapó forests in the Rio Negro basin by Geisler (1969) and Schmidt (1976).
In deoxygenated water, H2S concentrations may build up through the decomposition of bottom mud
and detritus.
Figure 2.15 Changes in the physical and chemical conditions with distance from the water’s edge in
a floodplain lagoon: (x--x--x) at 0800 hrs •--•--•) at 1200 hrs. (After Welcomme, 1970)
In the dry season dissolved oxygen conditions are linked to a number of factors including the size of
the water body, biological oxygen demand (BOD) of organic detritus or pollutants, degree of thermal

78
stratification, vegetation cover, phytoplankton development and wind action. In general there is a
tendency for even comparatively shallow bodies of water to stratify with higher dissolved oxygen
concentrations near the surface. Wind action is generally insufficient to ensure complete mixing in
deeper lakes where strong thermal stratification also exists. In shallower water bodies which lack
vegetation cover, wind action may ensure mixing through the whole water column (Schmidt, 1973).
However, resistance to mixing is comparatively high for the reason given in the preceding section.
Ducharme (1975) considered that, whereas wind is the major oxygenator in large lakes, dissolved
oxygen concentrations in smaller pools are largely controlled by the photosynthetic activity of the
phytoplankton. This means that smaller water bodies show greater diurnal changes in dissolved
oxygen than do larger ones. Photosynthetic activity raises the oxygen content of the water during the
day, particularly near the surface, but dissolved oxygen is withdrawn from water by plants and the
biochemical oxygen demand of silt and suspended organic solids and levels drop during the night
(Fig. 2.16).
Mass fish mortalities have been noted from Brazil (Brinkmann and Santos, 1973) and the Kafue flats
(Tait, 1967). In both cases the mortalities were attributed to sudden reductions in temperature cooling
the surface water of floodplain lakes which, coupled with strong winds, caused a rapid breakdown of
stratification, abrupt deoxygenation and contamination with H2S to lethal levels. Mortalities on the
Apure river floodplain have also been attributed to deoxygenated conditions (Matthes, pers.com.).
These arise when depressions containing unburnt grasses are inundated with rainwater. Because of the
persistence of thermal stratification the upper layers of the water (epilimnion) retain oxygen and the
fish survive there. However, the oxygen rapidly becomes depleted in the lower layers and H2S
accumulates. Later the turbulence associated with the entry of river flood waters mixes the layers
rapidly killing the fish. Other massive fish kills caused by the abrupt purging of oxygen depleted
waters from swamps have been recorded by Bryan and Sabins (1979) for the Atchafalaya river, a
tributary of the Mississippi. Such mortalities appear widespread in tropical floodplains as they have
been reported from many systems. Similar mortalities have been reported from cold water floodplains
where high BOD originating from pollution reduce dissolved oxygen levels below ice cover in winter
to levels below those tolerated by fish.
Floating mats of vegetation are an extremely common feature of tropical rivers and floodplain lakes.
The effects of these are similar whatever species of plant forms the mat. Fig. 2.16 for instance shows
the effect of stands of Paspalum repens on oxygen concentrations in Lago Calado of the Amazonian
várzea, but similar graphs have been drawn for the water column below Eichhornia crassipes (Arias,
1975 and Zarate and Cubides, 1977), Salvinia herzogii (Bonetto, Dioni and Pignalberi, 1969), Leersia
hexandra (Junk, 1973), Cyperus papyrus (Carter, 1953) and, Vossia (Tait, 1967). The general effect
of plant cover is to reduce dissolved oxygen concentrations, frequently to zero (Beadle, 1974) with a
coincident development and release of H2S. Because of the large amount of decaying organic matter
trapped in the root masses of floating plants, the deoxygenation often reaches a maximum in this zone
and sometimes rises slightly with increasing depth thereafter. The degree of deoxygenation below the
floating mat depends on water movements, for in strong currents the oxygen gradient beneath the mat
may be eliminated. It is, therefore, to be suspected that conditions under vegetation mats are less
extreme during the floods, when the constant movement of water may maintain at least some level of
dissolved oxygen. The density of the plant cover also influences the extent to which water becomes
deoxygenated; the more dense the cover, the less oxygen below it. Plant cover contributes to low
dissolved oxygen conditions in two ways. Firstly, a considerable biochemical oxygen demand is
created by both trapping organic matter and by the decay of their own vegetative parts, thus removing
oxygen from the water. Secondly, re-aeration of the water is prevented by reducing wind and wave
action on the surface of the water, and by producing shaded conditions unsuitable for phytoplankton
growth.

79
Figure 2.16 Diurnal changes in dissolved oxygen content and temperature in open waters and under
floating vegetation (Paspalum repens) in Lago Calado, 8 September 1968. (After Junk, 1973)
Oxygen gradients also seem to exist in the shallow littoral zone (Fig. 2.15). Wave and turbulence at
the water’s edge may here assure a restricted belt of oxygenated water, especially at night, even
though the water a little further offshore becomes completely deoxygenated. This effect, if
widespread, may be of considerable importance in the ecology of the fish, as many of the juvenile
forms are to be found in this zone.

80
CHAPTER 3
PRIMARY PRODUCTION IN RIVERS
THE RIVER CONTINUUM CONCEPT
Many workers have noted the apparent succession in ecological states along a river associated with
changes in morphology. Thus a river was early classified as young in its headwaters, where erosion
processes are dominant, mature in its mid-section where erosion and deposition are balanced and old
in its lower reaches where deposition is the major process. Rejuvenation zones may also occur in
mature or old reaches where increased gradient may temporarily reverse the more normal succession.
These descriptions are based on sedimentology but corresponding biological changes are also very
marked and have been summarized variously by Illies and Botasaneanu (1963), Hynes (1970) and
Hawkes (1975) who demonstrate the existence of different associations of flora and fauna along the
river which usually take the name of the dominant faunistic element, often fish. The nature of the flora
and fauna depend on the slope, current and the type of bed material in the river. As a river ideally
follows an orderly parabolic profile, the succession of the various elements appears correspondingly
orderly. Recently the river continuum concept, introduced by Vannote et al. (1980) has attempted to
assemble the various morphological and biological changes along a river into a coherent description
of this progression.
This concept assumes that the geo-physical variables within a river system present a continuous
gradient from source to mouth. Communities of living organisms succeed each other along the length
of the river in such a way as to minimize energy loss. This requires striking a balance between the
most efficient use of available energy through specialization and a contrasting tendency towards a
uniform rate of energy processing throughout the year. The structure of such communities is
summarized in Fig. 3.1. In the temperate rivers examined by the river continuum group, the following
shifts in community structure can be identified. Riverine communities can be separated into three
main groups, headwaters (orders 1–3), medium-sized streams (orders 4–6) and large rivers (orders
>6). Headwater streams are heavily influenced by riparian vegetation, which is responsible for large-
scale inputs of allochthonous nutrients while at the same time hindering autotrophic production by
shading. In some types of river, particularly blackwater rivers of the equatorial rainforests
allochthonous inputs in headwater streams or on floodplains may represent a major source of new
nutrients to the system. As stream size increases allochthonous inputs become less important and the
aquatic communities tend to concentrate more on autochthonous processing of nutrient transported
from upstream. This transport, with the exception of new allochthonous material arising from
floodplains and feeder tributaries, is the basis for all subsequent living processes. The first effect of
this process is in the nature of the allochthonous material which degrades from coarse particulate
organic matter (CPOM) in the low order streams to progressively finer particulate organic matter
(FPOM), ultrafine particulate organic matter and eventually to molecular components, amino acids,
sugars, etc. as one proceeds downstream. The composition of the living aquatic communities reflects
these changes in the nature of the nutrient substrates and in the physical form of the river ecosystems.
Plants progress from anchored submerged macrophytes in upstream reaches through periphyton to
phytoplanktonic communities within the main channel. Infrasubstrate communities shift from
shredder-dominated areas in the low-order headwater streams through grazer-dominated areas in the
medium-sized streams, to collector-dominated communities in the higher order streams. Fish
communities also tend to undergo a similar transition from invertivorous predator dominance in low-
order streams, through grazer-dominated communities in medium-sized rivers to iliophagous
dominance in the potamon.
Since its proposal many workers have used the concept as a framework for the analysis of small river
systems. Hawkins and Sedell (1981) for example, confirmed the predictions of the concept in four
Oregon streams, and Culp and Davis (1982) similarly successfully applied it to the Oldman and South
Saskatchewan river system. The ecological changes envisaged by the river continuum concept are

81
mostly accomplished in the progression through smaller order streams and little change is predicted in
rivers from order 6 onwards. This means that it may be used as a descriptor for the first 200 km or so
of a river course during the transition from rhithron to potamon but once the stable potamonic phase is
reached little further change can be anticipated for several thousand kilometres. Furthermore, within
the potamon, the floodplain is somewhat of a special case, for here the recycling of nutrients and
organic matter through the growth and decay of plants parallels, to a certain extent, the situation
nearer the headwaters, with the renewed input of CPOM. However, the nutrient base for this
productivity may be regarded as autochthonous in that it arises from the nutrients deposited in river
borne silt.

82
Figure 3.1 conceptual relationships between stream size and the progressive shift in structural and
functional attributes of lotic communities. (After Vannote et al., 1980)
Work by Bruns et al. (1984) indicates that, while this theoretical continuum may be applied to the
spatial evolution of conditions within the main channel, it is strongly influenced by presence of
tributaries. The ways in which these inflowing rivers affect the progression depends on the size of the
tributary relative to the main channel. This explains the sometimes abrupt transitions that may occur
at the confluence of two large water courses. Similarly the continuum may be interrupted or even
reversed by geomorphological irregularities in the normal catenary shape of the profile.
MATERIAL OF ALLOCHTHONOUS ORIGIN
An important input into the aquatic system is the rain of small non-aquatic creatures and organic
matter from terrestrial sources. As the floods advance, invertebrates, especially ants and termites, are
caught by the rising flood and incorporated into the aquatic system. There is also a continuous input
of insects, seeds, leaves, pollen and other material from flooded forests and grasslands which either
enter the drift in flowing waters, or settles to the bottom where it is decayed by bacterial and fungal
activity.
Workers on inundated forest regions all consider nutrients of allochthonous origin to be the single
most important, if not the only, input into the system. Geisler, for example, has in preliminary
experiments found that up to 56 individual pieces of organic material fell onto a quarter m² glue board
in the course of one day (Geisler, et al., 1973). The extent of leaf fall is indicated by data from many
parts of the world. For example, Blackburn and Petr (1979) summarizing data from low order
temperate streams showed that averages of between 3.5 and 8 t of plant litter (leaf, bark and
branches)/ha fall into the water every year. Comparable figures from the tropics indicate leaf falls of 6
t or more/ha/yr. Standing crops may be equally high, for instance in one small stream from the new
territories Hong Kong, Dudgeon (1982) found the detrital standing crop to be one hundred times
greater than that of the periphyton and more consistent at 80–120 gm/m² (= 1 t/ha). About 60% of this
was organic matter and thus accessible to the food chain.
Studies on the flow of nutrient through the Volga system indicate that the nutrient cycle in the river
and its reservoirs is greatly influenced by external inputs. The main trophic flows are thought to
proceed from allochthonous organic matter via bacteria direct to invertebrates, primary production by
phytoplankton and higher vegetation playing a very secondary role.
Annual litter fall on floodplain wetlands is very high, although the precise classification of material
generated on the floodplain as allochthonous or autochthonous is arguable. Records from the United
States range from 350 g (dry wt.)/m²/yr for an Illinois swamp to over 700 g (dry wt.)/m²/yr in North
Carolina (Mulholland, 1981). In Amazonian rain forests litter fall may be as high as 1 000 g (dry
wt.)/m²/yr although Adis et al. (1979) calculated that in the poorer Rio Negro, floodplain forest values
are closer to 580–790 g (dry wt.)/m²/yr. Even in savannah floodplains the presence of gallery forest
and floodable scrub vegetation provides a substrate from which materials fall into the water.
BACTERIA AND OTHER MICROORGANISMS
Much of the initial breakdown of CPOM in headwater streams is carried out by fungi and bacteria
which are abundant in the leaf letter of these zones. Sesile bacteria also form an important part of the
biomass of the riffle zones where they form epilithic slimes. Bacteria may became detached during
period of high flow and between 0.4 an 9.2 × 106 cells have been found suspended in flowing waters
depending on discharge (Marxsen, 1980).
Little information is available on the abundance of microorganisms in unpolluted rivers and
floodplain lakes, although they are obviously of immense importance both in the breakdown of

83
vegetation debris, dung and other organic remains, as well as in the diet of many species of detritivore
fish. The distribution of such microorganisms as fungi, actinomycetes, or starch, pectine and hemi-
cellulose decomposers, which obtain their energy from organic substrates, as well as aerobic and
anaerobic nitrogen fixing bacteria, were substantially higher in the swamps now submerged by the
Kainii reservoir, than in the river (Imevbore and Bakare, 1974). For example, a total of 6.3 × 107
organisms/ml was estimated to be present in the surface water of the swamps and 5.1 × 1011/ml were
estimated in swamp mud. This contrasts with the 3.5 × 104 and 1.3 × 1011/ml found in river water and
mud respectively. Fairly constant amounts of bacteria, between 2.3 × 105 organisms/ml for the Rio
Negro and 5 × 105/ml for the Solimoes were found in the rivers of the Amazon basin (Schmidt, 1970).
Greater numbers were found in the Lago do Castanho, a representative várzea lake. Here total
estimates of up to 7.3 × 106/ml were made although there is considerable seasonal and spatial
variation in density (Schmidt, 1969). In the lake, bacterial activity closely follows that of algae and
seasonal maxima of algae are always associated with maxima in bacterial number. Similarly there is a
distinct vertical stratification in both algal and bacterial numbers, with differences of between 0.5–4 ×
106 organisms/ml. Maximum densities occurred at about 1 m depth and at the bottom. Rai (1979) also
found a strong correlation between bacterial counts and water level in four Amazonian lakes. The
numbers of saprobic bacteria in a blackwater lake, ranged from 4 × 10³ - 2.2 × 105/ml during high
water to 1.1 × 105 - 9 × 105/ml during low water, and total bacterial counts in varzea lakes ranged from
2.1 to 11.6 × 108/ml during high water and 4.2 to 15.6 × 109/ml during low water. Rai further
confirmed Schmidt's findings that bacterial maxima were strongly correlated with algal maxima.
These examples from tropical systems indicate the richness of the bacterial flora in the more sheltered
tropical waters. The values do not in fact differ greatly from those quoted for the Danube by Mucha
(1967), where 1.5–2.6 × 106 organisms/ml were present in the bacterial plankton in Czechoslovakia
and Yugoslavia. In the Soviet portion of the delta 3.2–23.6 × 106 organisms/ml have been noted from
the river and 3.6–12 × 106 organisms/ml from the standing water Kilia arm. The Danube, however, is
highly polluted, particularly with organic matter. Other Ponto-Caspian rivers show similar ranges of
bacterial plankton numbers: 9-23.7 × 106 organisms/ml from the Volga and 1–3 × 106 organisms/ml
from the Dniester (Gavrishova et al., 1982).
PHYTOPLANKTON
The contribution made by phytoplankton to primary production within rivers is generally regarded to
be low when compared to other types of aquatic systems. However, phytoplankton is present in rivers
and contributes to the nutrient balance and to the trophic requirements of some of the fish species.
INFLUENCE OF ENVIRONMENTAL FACTORS
The major factors determining the presence and abundance of phytoplankton are temperature, velocity
of the current, availability of nutrients and availability of light.
TEMPERATURE
In high latitudes temperature is one of the principal regulators of planktonic abundance with a well
defined seasonal cycle based on the alternation of winter and summer. Thus in temperate rivers there
is a minimum in phytoplankton production and biomass during the winter. Even in the lower Parana,
where large seasonal temperature differences are normal, the summer high water figures for
phytoplankton numbers are frequently higher than the low water figures suggesting that temperature is
the major factor influencing phytoplankton abundance in this portion of the river (CECOAL, 1977).
In tropical rivers temperature plays a much diminished role and the greatest densities of
phytoplankton coincide with low water.

84
CURRENT
Phytoplanktonic organisms are sensitive to velocity and turbulence of flow in rivers as the rapid
currents and mechanical stresses of rapids and waterfalls inhibit the development of new plankton and
rapidly suppress any existing organisms discharged from associated lentic waters. Thus the agitated
waters of the rhithron generally support little plankton, although some does develop in the occasional
quiet backwater and pool. In addition a drift is present in the main channel made up of primarily
bottom living algal forms which are dislodged by the rapidity of the flow.
In the potamon, studies confirm the strong influence of flow and, consistent with this, show that
phytoplankton is more common in the lentic components of the system than in the lotic.
Prowse and Talling (1958) demonstrated the strong correlation between phytoplankton growth and
current velocity in the Nile at the Gebel Aulia dam. The dam slowed the Nile current and produced a
rapid increase in planktonic concentration. When the dam was open the flow was faster and plankton
concentration dropped. The build up of phytoplankton from this source has been thought to account
for the majority of potamonic plankton; a conclusion justified by the progressive disappearance of
such phytoplankton downstream from the point of discharge. Storage and discharge of water from
mainstream or major tributary reservoirs can also alter the indigenous patterns of phytoplankton
abundance. This has occurred in the Volga River where the whole algal fauna has been considerably
modified following the conversion of the river into a cascade of reservoirs (Kuzmin in Mordukhai -
Boltovskoi, 1979). Before impoundment the spring flood peak of phytoplankton biomass was less
than the summer low water peak. After reservoir construction, species composition changed, with a
reduction in the number of taxa present in the unmodified lower floodplain and delta. Here the
seasonal cycle of abundance has remained relatively unmodified although now the spring peak in
biomass (6.3 g/m³), when diatoms predominate, is greater than the prolonged period of high biomass
through the summer low water (2.4 – 3.8 g/m³) when green and blue algae are most common. The
phytoplankton of the Dniester River likewise underwent qualitative and quantitative changes after the
closure of the Dubassery reservoir with increases in Cyanophyta downstream of the dam. The
phytoplankton biomass varied between 0.66–0.85 g/m³ before the dam was built but rose to about 4
g/m³ in the river after its construction.
Discharge is assigned the major role in regulating phytoplankton abundance in the Mississippi river
(Baker and Kromerbaker, 1979), although in the river seasonal variations of temperature are high and
influence the succession of the various components of the phytoplankton. Phytoplankton abundance,
as represented by chlorophyll ‘A’ concentrations, was less in the main channels of the river than in the
backwaters or in a river lake where the current was slowed. In the Missouri river, Berner (1951)
associated the low planktonic densities (0.067 cells/ml) with high current and turbidity and a lack of
subsidiary floodplain waterbodies to feed into the mainstream. The Illinois River, a tributary of the
Missouri/Mississippi system had much higher phytoplankton densities (about 400 cells/ml) when it
was still connected to its floodplain lake and backwaters (Kofoid, 1908). The literature of the
Mississippi system between St. Louis and Cairo has been summarized by Schrammn et al., 1974 who
confirm these findings and commented on the difference in species composition and density between
main channel, riverine backwaters and floodplain lakes. Diversity is far higher in the lentic
environments where chlorophycea and Cyanophycea are dominant than in the river where
Chrysophycea are the major element. Similar findings are recorded by Bryan et al. (1975 and 1976)
who commented that phytoplankton has a very characteristic distribution within the Atchafalaya
system. When the flood is in progress habitats are inundated and flushed out producing homogenous
community structures. After the floods recede communities differentiate and representative forms
characteristic of the different types of water body again exert themselves. Clearly if much of the main
stream phytoplankton originates from flushing and discharge from the lentic components of the
system, build up in numbers must occur where the flow of a river is slowed or halted in backwaters or
in the standing waters of the floodplain. Rzoska and Talling (1966) found phytoplankton to be much
more abundant in backwaters of the Nile Sudd than in the main channels and thus Rzoska (1974)

85
quoted values of between 40 to 140 cells/ml for the river, whereas densities in lagoons reached from 1
720–2 330 cells/ml at river post 12. Differences in the specific composition of the phytoplankton were
also common. Blue-green algae such as Anabeana and Lyngbya dominated in the standing waters,
whereas in the river the sparse flora consisted mainly of diatoms especially Melosira. Samples from a
small West Africa river, the Oshun, showed similar trends to occur there. The main river is inhabited
mainly by desmids and diatoms, and colonial chlorophycea were the first to colonize the backwaters.
Phytoplankton abundance is also associated with seasonal differences in flow. Densities generally
reach a peak in the dry season and diminish in the floods in both types of water unless otherwise
influenced by temperature. Thus Egborge (1974) found a good negative correlation in the Oshun
between phytoplankton abundance and both water level and current velocity, with maximum
abundance at times of low water, although even then the total numbers of organisms were very low.
Iltis (1982) also found that algal populations were maximal during low waters and that the floods
were characterized by very poor phytoplankton in the main channels of six rivers of the Ivory Coast.
Carey (1971) had earlier found phytoplankton densities to be less during the floods in the Kafue River
with dense blooms occurring in the river at Nampongwe and in Namatenga lagoon between August
and November when the floods had receded. Phytoplankton was generally scarce in the Sokoto River
at most times of year, but was maximal in the dry season between March and June, especially in a
floodplain lagoon. Holden and Green (1960) suggested that, although the relative abundance of
organisms in terms of numbers per unit volume is lower during the floods, the absolute abundance
may well remain the same due to the dilution of the number of organisms by the enormously
increased volume of water in the system.
A similar argument was proposed by Bonetto, Dioni and Pignalberi (1969) who remarked at the same
time on the generally low contribution made by phytoplankton to the primary production of the
Parana River. This is particularly small during the floods or in water bodies with dense vegetation due
to high turbidity and shading effects, but may rise in some lagoons during the dry season. For
instance, CECOAL (1977) showed cycles of phytoplankton abundance in the upper Parana which
fluctuated between about 25 cells/ml in the floods to over 250 cells/ml during low water. In the
Middle Parana values of 2 331 cells/ml (low water) and 44 cells/ml (flood) represent values midway
between those of the two confluent systems (Bonetto, 1982). The algal population of the main channel
of the Amazon was also found to be higher at low water (15 000 cells/ml) than at high water (3 000
cells/ml) (Schmidt, 1970), and the same held true in a varzea lake with 500 000 algal cells/ml at the
period of minimum water level. Much of the rise in the number of phytoplankton in the river was
attributed to discharge of algal rich waters from lagoons. This cyclic pattern of activity is by no means
universal in the Amazon system as the Rio Negro showed a remarkably constant regime of about 10
000 algal cells/ml, and in lake Redondo the peak of algal production was reached during the rising
waters when the nutrient-rich whitewaters were invading the lagoon (Marlier, 1967). Because of the
interaction between the various water types in the Amazon system the nutrient regimes are likely to
differ considerably from the more normal regimes of rivers with only one dominant water type, and in
certain circumstances generally nutrient concentrations may be greater in the floods when lagoons are
invaded by nutrient-rich waters than during the dry season when nutrients have been diluted by
rainfall and inflow of poorer ground waters.
AVAILABILITY OF NUTRIENTS
There are several pointers to the important role nutrient availability plays in determining the
abundance of the phytoplankton and in particular in limiting it development beyond a certain level. In
the Gebel Aulia Dam for example, Prowse and Talling (1958) attributed the failure of phytoplankton
to develop beyond a certain level at slack water to nutrient depletion, particularly of nitrates. Talling
(1957) had earlier traced the high negative correlation of phytoplankton abundance and nitrate
concentration in the Nile. On the other end of the scale, extraordinary high concentration of nutrient
associated3 with eutrophication may result in blooms in excess of the usual.

86
The normal cycle of abundance associated with water velocity is shown in the Danube, where in
Romania 0.8 cells/ml were found during the June floods and 4.0 cells/ml were found during the
October low water (Szemes, 1967). Similarly in the slow reaches of the Russian part of the delta, cell
counts ranged from 192 cells/ml in the floods to 2 621 cells/ml at low water. This pattern was
disturbed by heavy pollution in the upper reaches of the river, where in the fast flowing Austrian
stretch 300 cells/ml were already present. Densities of 10–15 000 cells/ml were attained in the highly
polluted Czechoslovakian and Hungarian reaches, where water blooms are common during the
autumn giving a foul taste to drinking water. Juris (1975) even records numbers as high as 20 000
cells/ml in periods of maximum development, which may rise in high as 50 000 cells/ml in years with
especially low water. Further downstream, in Yugoslavia, cell counts dropped as low as 320–1 060
cells/ml and continued to fall to the figures shown for the delta. The Danube pattern indicates that
when abundant nutrients are available, flow becomes a secondary consideration in limiting
phytoplankton number.
Elevated standing stocks of phytoplankton in the Laguna Gonzales (54 642 cells/ml - 252 906
cells/ml) as compared to other lagoons of the Riachuelo (Parana river) - Laguna Totoras 129–1 330
cells/ml Laguna La Brava 335–9 235 cells/ml, Laguna Sirena 194–434 cells/ml and Laguna Meritta
72–654 i/ml are traceable to the highly eutrophicated state of Laguna Gonzalez (Bonetto et al., 1978).
The actual families of algae comprising the plankton vary much with water quality and in the Danube
Blue-green algae predominated under eutrophicated conditions. In the tropics too the major forms
present differ, for instance desmids tended to dominate in the flora of black-water streams both in the
Kapuas R., Borneo (Vaas, 1953) and in the Amazon.
In clear white waters of neutral pH diatoms and green algae are more abundant and in eutrophicated
waters or those with high pH, blue-green algae are the more common and indeed are often the only
element of the flora.
AVAILABILITY OF LIGHT
The amount of suspended matter in the water affects the penetration of light into the water, for
example Bonetto (1980) found a relationship S = a e bh for the Parana river, where S is the light
penetration as measured by Secchi disk, h is the velocity of the current as measured by the height of
the river and a and b are constants. It would thus appear that in many cases the limitation of plankton
development in the main channel of swift flowing rivers stems not from the current per se but from
the low penetration of light in such waters. In the main channel of the Parana just below the
confluence with the Paraguay river, for instance, the primary production varied between 0–285 mg
C/m²/day and the number of organisms varied between 80–2 000 cells/ml. The maxima and minima
were very strongly correlated with flow (Bonetto et al., 1979), abundance and production during the
flood being reduced not only by the strong current but by the poor light penetration. In many Latin
American floodplain waters, and probably in African areas too the productive zone is limited to a
relatively thin layer near the surface. This rarely exceeds 3 m in the Amazonian Lago de Castanho
(Figure 3.2) or in the flood lakes of the Riachuelo river (Bonetto et al., 1978a and b), or 2 m in the
Cienagas of the Magdalena (Mikkola and Arias, 1976). Limitation of photosynthetic activity during
the rainy season occurs when rising waters bring silt into the lagoon. It may be restricted in a similar
manner during the period of low waters when wind induced turbulence resuspends bottom mud.

87
Figure 3.2 Vertical patterns of primary production by phytoplankton and secchi disc transparency
(vertical bar) in Lago do Castanho from August 1967 to October 1968. (After Schmidt,
1973b)
OTHER FACTORS
Higher vegetation may also influence plankton abundance. In Bangula lagoon in Malawi, the waters
within patches of Nymphaea supported some 16 731 = 5 512 algal units (cells, filaments or
colonies)/ml. Here shading effects possibly reduce plankton densities. Over submersed vegetation,
higher but very variable figures were obtained of 38 107 – 52 188 units/ml (Shepherd, 1976).
Concentrations of planktonic organisms were some 13 times greater in patches of open water within
floating vegetation in the Laguna la Brava of the Riachuelo River than they were in the open water of
the lagoons on the same date (Bonetto et al., 1978a). Reduced phytoplankton densities near emergent
and floating vegetation and higher values over submersed vegetation have also been remarked upon
from the Danube.
PRODUCTION
Bonetto (1982) has proposed a model linking transparency with solar radiation, temperature and the
existing plankton density to predict the productivity of phytoplankton in the Parana River:
P = [10 + 0.28 (AmaxF)]e-0.019S
where: A max = -0.018 + 0.002Q + 0.021T

88
and
P = Productivity per unit area (mgC m³/day)
T = Mean temperature (°C)
F = Density of phytoplankton
A = Photosynthetic activity as defined by (2)
Q = Daily radiant solar energy (Cal/m²)
S = Secchi disc reading (in cm)
Considerable work on primary productivity of floodplain lagoons has also been carried out by
Schmidt (1973a) on the Lago de Castanho of the Amazonian várzea. His estimates of biological
production ranged from 2.15 gC/m/day at the lowest water level to 0.32 gC/m³/day during the inflow
of new river water. The net annual production was 297 gC m equivalent to a gross productivity of 358
gC/m²/year. The algal biomass was 1.9 gC/m² or 17 kg/ha with a gross productivity of 1.1 gC/m²/day.
Production from the Rio Negro was found to be considerably lower, ranging from 0.030 – 0.434
gC/m³/day giving a net productivity of 0.063 gC/m³/day or a gross annual production of 23
gC/m²/year (Schmidt, 1976). Other workers have found values between these two extremes. Bonetto,
Dioni and Pignalberi (1969) obtained values between 0.050 gC/m²/day and 1.0 gC/m²/day from two
different lagoons in the Paraná floodplain and two other lagoons, L. Totoras and L. Gonzalez of the
Riachuelo tributary to the Parana give somewhat higher readings of 0.2 – 2.5 gC/m²/day and 0.45 –
2.46 gC/m²/day respectively (Caro et al., 1979). Marlier (1967) found productions of between 0.14
and 0.7 gC/m²/day on the Lago Redondo. Values for various Magdalena cienagas range from 0.16 to
1.77 gC/m²/day (Mikkola and Arias, 1967) (Table 3.1), with mean values of 0.09 gC/m³/h from 18
lagoons of the Magdalena system (Arias, 1977). Values for gross primary productivity in the main
stream of the Godavari River (India) ranged from 0.30 to 1.06 gC/m³/day (Rajalakshmi and
Premswarup, 1975). Maximum values were recorded during the post flood period as transparency
rises and flow rate falls. At this time there was a good phytoplankton bloom. A second bloom
appeared after the summer rains. These figures are influenced by the discharge of organic polluting
effluents at one of the sites where mean values of primary production were the highest and also by the
anicut weirs which slow the flow and even out water level fluctuations.
These figures from tropical systems compare with temperate zone production rates of between 0 – 15
g 02/m²/day (equivalent to 0–4.7 gC/m²/day tabulated by FAO/UN (1973), and show the productivity
of phytoplankton in rivers to be extremely low, although peak production at low water from isolated
floodplain pools may temporarily reach the order of magnitude of production from lakes.
Table 3.1
Primary production estimates for various rivers and floodplain lakesa
System Production
gC/m²/day Authority
Amazon: Lago do Castanho 0.82 Schmidt, 1973
Lago Redondo 0.29 Marlier, 1967
Rio Negro 0.063 Schmidt, 1976
Rio Tapajos (0.44–2.41) 1.366 Schmidt, 1982
Paraná: Lago Los Espejos 0.05 Bonetto, Dioni,
Pignalberi, 1969
Lago El Aleman 1.00 Bonetto, Dioni,
Pignalberi, 1969

89
Laguna Totoras 1.2 –1.6 Bonetto et al., 1978
0.2 –2.5 Cara et al., 1979
Laguna Gonzalez 0.8 –2.5 Bonetto et al., 1978
0.45–2.45 Cara et al., 1979
Magdalena: Cienaga Guajaro 0.67 Calculated from data
Cienaga Maria la Baja 0.34 Mikkola & Arias, 1975
Cienaga Carabali 1.77
Cienaga Palotal 0.16
Godavari R.: 0.30–1.06 Rajalakshmi-Premswarup, 1975
Paraguay. Laguna Herradura 0.14–4.50 Zalocar et al., 1981
a N.B. These figures are often based on only a few observations and do not therefore necessarily
reflect the full range of variations of the parameters measured
ATTACHED ALGAE
Production by algae, especially diatoms growing on rocks, submerged wood and floating or
submersed vegetation may well be more important than the phytoplankton. This is certainly true in the
rhithronic headwaters where phytoplankton is virtually absent but where the rocks of the riffles
support. Mats of epiliths which in turn, provide the substrate for the complex of organisms comprising
the “aufwuchs”. In the pools too, the floating leafed vegetation of the slacks are colonized by dense
aggregations of epiphytes. The ecology of such communities in the running waters of the temperate
zone has been surveyed by Hynes (1970).
Few quantitative data appear to be available on this community in the potamon, although several
authors have remarked upon the abundance of such organisms. Rzoska, (1974) described the stems
and surfaces of emergent, submerged and floating vegetation in the Nile Sudd as being covered with
epiphytes including the red algae Compsogon. Indeed, Mefit Babtie (1982) record epiphytes as
comprising 16.6% of the total dry weight of Naias pectinata where 0.17 g of epiphytes were found per
gram of Naias. In the flood the proportion dropped to only 2.6%. Epiphytic algae were also recorded
as being very abundant on the Kafue flats by Carey (1971). The littoral of the Lake Chilwa swamps,
which closely resemble these of riverine floodplains, also support considerable populations of
epiphytes on Typha stems and on floating dead plant material wherever there is sufficient light
(Howard-Williams and Lenton, 1975). In Latin American waters Ducharme (1975), and Mikkola and
Arias (1976) considered the production by periphyton to be considerably superior to that of the
phytoplankton in the cienagas of the Magdalena river floodplain. The epiphytic community has also
been considered very important in the Middle Parana because of the abundance and density of support
in the form of floating and emergent vegetation, (Bonetto, Dioni and Pignalberi, 1969).
Although few figures are available it does appear that biological production by epiphytes is very high
in most flood zones, especially at the periphery of vegetation masses where light is adequate for
growth. In Bangula lagoon, Malawi, for example, the number of cells loosely attached to
Ceratopyllum was estimated by Shepherd (1976) at 3.35 ± 2.10 ×106/g fresh wt. of Ceratophyllum.
This gives an extrapolated number of 45.21 ± 17.70 ×109 cells/m² of lagoon surface, which was
between 0.8 and 20 (mean 9) times the number of cells of the same organisms found free in the water.
Investigations using artificial substrates in the Danube (Ertl et al., 1972) showed that periphyton
abundance was limited by frequent fluctuations in water level and by scouring. These effects may
have been magnified by the fixed nature of the artificial substrates used, which more closely
simulated rocks or emergent vegetation, than floating vegetation which could move with the water
level. Because of these effects, spring maxima in periphyton biomass were low and annual maxima of
up to 650 g/m² were achieved in autumn, and often persisted until early winter when flows were
minimal. Similar conclusions on the correlation between periphyton abundance and the flood cycle

90
were reached by Iltis 1982, in some West African rivers. Here the abundance of periphyton per unit
area followed the same fluctuations as those of the phytoplankton with the peak in relative abundance
being reached at the end of the dry season. However, the greatly increased area of substrate available
during the floods in the form of root masses of floating vegetation probably compensates for the low
density and absolute abundance may well be maximal at this time.
Colonization with diatoms such is Melosira or Oscillatoria produce biomasses of 2–15 /m² in the
muddy bottoms of the Volga and in the Dnieper biomasses exceed these of the zoobenthos. There are
indications that the density of epiphytes decreases towards the shaded interior of stands and mats of
vegetation along with the rest of the Aufwuchs community. For instance, in the Danube, Juris (1969,
1973) found 7.7 × 106 cells/cm² were present in shaded areas. Similarly standing crops of periphyton
are reduced with depth (Fig. 3.3) both because the lower light penetration limits growth and because
predatory consumer populations increase slightly with depth.
Figure 3.3 Biomass (B) and chlorophyll (CH) of periphyton at different depths in the littoral of the
Danube River (Ertl et al., 1972)
Because of the concentrated nature of the periphyton it obviously is a major locus of production in the
aquatic system. However, epiphytes may also perform subsidiary roles in the nutrient ecology of the
system. Heeg and Breen (1982), for example, found that epiphyton on Potamogeton crispus had a
considerable nitrogen fixing capacity of up to 23 mg N per 24 hours or 1.27 mg N/m² during the
inundation of the Pongolo floodplain. It also forms the base of a particular community which is
associated with the periphytic and perilithic habit, including the complex known as Periphytic Detrital
Aggregate (PDA). In samples from L. Valencia, Venezuela, Bowen (1979) has shown PDA to contain
42.7% dry weight of organic matter. Algae, on the other hand, only contributed a small portion of the
percentage (0.2 – 2.8%) by dry weight of the same sample even though they contribute a greater
volume (17–279 mg/ml of sample).
HIGHER VEGETATION
Distribution and zonation
Higher plants provide the major biotic structural elements in fluvial ecosystems. Not only does their
distribution depend on the geology and morphology of the environment, but the presence of
vegetation can itself modify the form of the system.

91
Although only rarely used directly for food by fish, vegetation has a range of ecological values for
fish communities. It provides refuge, shade, a substrate for spawning and a support for many
organisms which are of dietary importance for fish.
Longitudinal zonation of vegetation within rivers is based mainly upon the related factors of depth,
flow, and mechanical stress. In torrential headwaters liverworts and mosses are the earliest forms to
appear on rocks, both submersed and in the splash zone. This type of vegetation persists into the rocky
riffles of the rhithron, but in these reaches the pools increasingly support rooted, floating leafed
species in the slacks and emergent vegetation along the banks as slope decreases. As the river enters
the more mature potamon, the ideal channel is fringed with emergent plants and floating grasses
which give way to floating leafed plants and submersed species as the depth increases toward the
centre of the river. Such a stable state is rarely attained in natural rivers, where one or other of these
elements may be lacking. Similar successions can be seen in islands within the river channel where
sand banks laid down by the current became colonised by plants which fix the bank and lead to
further siltation. Such islands eventually form part of the seasonally floodable area taking the form of
internalized floodplains. Gosse (1963) describes the distribution of vegetation on such islands in the
Congo R. (Fig. 3.4).
The river floodplain shows much more complex lateral successions based on the degree of flooding
such as that proposed by Adams, 1964 which contains the following zones:
(a) permanently flooded waters with submersed vegetation only (open waters);
(b) permanently flooded areas with rooted or floating emergent vegetation;
(c) regularly seasonally flooded areas with rooted and floating emergent vegetation;
(d) areas that are occasionally flooded (between mean flood and highest flood levels;
(e) areas that are not flooded but whose water table is influenced by the flood regime.
In the Shire river Elephant marshes, Howard-Williams (in Hastings, 1972) distinguished the following
zones:
(a) aquatic plant zone: with floating sudd islands composed of Echinochloa pyramidalis, Ludwigia
stolonifera and Ipomoea aquatica, together with true floating plants such as Azolla nilotica,
Salvinia hastata and Pistia stratiotes;
(b) a swamp zone with water between 50 cm and 2 m in depth which consists of floating meadows of
Echinochloa pyramidalis, Vossia cuspidata, Leersia hexandra, Cyperus papyrus, and
Echinochloa stagnina;
(c) flooded grassland 1.5–6 m deep, mainly dominated by Oryza barthii;
(d) shallow flooded grasslands and levees with depths of flooding between 0.25 and 25 cm with
tussocks of Setaria avettae, Vetivaria nigritana;
(e) floodplain margin regions with Hyparrhenia rufa, Panicum coloratum, Vetiveria nigritana and
Setaria sphacelata.

92
Figure 3.4 Successions of vegetation on islands in the Congo River (A) Young Island; (B) Older
Island (from J. Louis, 1947, quoted by Gosse, 1963)
Zonation of vegetation fringing the Parana River and its floodplain lakes is similar and also varies
according to the type of terrain as shown in Fig. 3.5.

93
Detailed analyses of the vegetational zonation of floodplains carried out by Schmid (1961) for the
Tonle Sap in Cambodia and Smith (1976) for the Okavango Delta, show relatively little difference in
the basic type of spatial zonation for tropical systems.
In the Danube the zonation of vegetation is based on a hydrographic index where hg = 0.1 (mean high
water level - mean low water level) and vegetation complexes are correlated with hg in the following
manner:
Hg 6–7 highest river banks forested with willows, poplars, and ash
Hg 5–6 water meadow pastures
Hg 8–5 emergent water plants, cat tails and reeds
Hg 3 well developed reed beds
Hg 0.3 backwaters, lakes with floating vegetation and submersed plants

94
Figure 3.5 Zonation of vegetation fringing the Parana River: A. Sand banks; B. Bordering deep
lagoons and channels; C. Shallow lagoons ad marshes. (After Franceschi and Lewis,
1979)
Because of the regular seasonal variations in water level in many river systems there is a temporal
succession as well as spatial one. In the long term the colonization of river channels, backwaters and
the various types of floodplain water body by floating and emergent vegetation accelerates siltation
and tends to catalyze the transition of such waters to dry land. This type of succession has already
been described for the Danube River in considering the morphology of the floodplain. More detailed
profiles of the shorelines of two of the numerous lakes from the Riachuelo river of the Parana system
(Bonetto et al., 1978a and 1978b) (Fig. 3.6) show the difference between Lago Totoras, a clear,
moderately eutrophicated lake with prairie like marginal vegetation which is presumably young in

95
temporal succession whereas L. la Brava, which is covered with floating emergent vegetation,
possibly represents an older and mature lake of the same system.
Figure 3.6 Zonation of littoral vegetation in two lagoons of the Riachuelo River A. Laguna La Brava;
B. Laguna Totoras. (After Bonetto et al., 1978a and b)
In the short term there is an annual sequence of replacement of one species by another in response to
the different degrees of flooding. In individual lagoons, vegetation increases in density and biomass

96
throughout the dry season until it occupies most of the water surface. As the water rises much of the
vegetation is washed out and biomass within the lagoon falls rapidly. Nieff (1975) has further defined
the changes in the relative abundance of the species found in the lagoon showing the progressive
occupation of the free waters by floating forms such as Azolla and Salvinia during rising waters. The
subsequent growth of Polygonium punctatum, Ludwigia peploides and Mycrophyllum brasiliensis
shades out the free floating forms and as water level falls, these plants are in turn replaced by
Nymphoides indica.
Riparian forest is an important element in the vegetation complex of rivers in that, in the natural state,
fallen wood structures the environment, leaf fall provides a major source of organic and nutrient
inputs and the overhanging vegetation gives a mosaic of light and shade which conditions the
distribution of many aquatic organisms. Originally many of the world's floodplains would appear to
have been forested, at least by a strip of gallery along the channels. However, the influence of man
and his domestic animals in colonizing the rich alluvial bottom lands has resulted in their clearance to
produce the type of agricultural or savannah plains so familiar throughout the world. Two examples of
this historical development are given by Yon and Tendron, 1981, for the alluvial forests of Europe
(Fig. 3.7) and by Van Leynsele (1979) for the Ngiri floodplain. The Ngiri, which lies in the densely
forested Congo system is kept free of trees by regular burning of vegetation.
Figure 3.7 Schematic historical development of the landscape and vegetation of a river valley in
Europe. Middle reaches (right); Upper reaches (left).
a. At the beginning of the Christian era; b. Circa 1000; c. Circa 1800; d. Circa 1900
1. beech groves; 2. mixed forest and oak stands; 3. alder stands; 4. afforestation conifers;
5. bushy willows; 6. other bushes; 7. water meadow; 8. fresh meadow; 9. dry meadow; 10.
crops; 11. slope loam; 13. more; 14. gravel; 15. other soil; 16. average level of ground
water; 17. average level at high water. (After Yon and Tendron, 1981)
Deforestation is still proceeding throughout much of the tropical world, but considerable areas still
remain in South America, Africa and Asia, where the floodplains are occupied by dense forests of

97
flood-resistant trees. These may be of two main kinds which are best seen in the Amazon system
where the ombrophilous lowland forests occupying the alluvial plains of the whitewater rivers are
known as várzea forests and the tropical evergreen peat forests occupying the floodable zones of the
blackwater rivers are known as Igapó forest. River-side or gallery forests tend to occupy the levees on
many wet savannah rivers and Bonetto (1975) described the Paraná as a corridor by which the
Amazonian forest is able to penetrate far to the south of its normal distribution. Profiles of forested
floodplains given by Sioli (1964) and Bonetto (1975) (Figs. 1.7a and b) describe the general
distribution of the major vegetational zones of the Amazon and the Parandá Dry savannah rivers are
frequently bordered by flood-resistant trees and scrub, mainly of the Acacia type, but also some palms
especially on dry terrain ridges and levees. In the Central Delta of the Niger, the boundary between
unshaded and gallery forested floodplain channels was distinguished by Daget (1954) as following the
1 000 m isohyet; to the north drainage channels were unshaded and to the south were forested. The
displacement southwards of the isohyet during the Sahelian drought has produced changes in the
distribution of the woodlands bordering the river channels.
Submersed vegetation
True submersed rooted aquatics form a high proportion of the macrophytes in temperate rivers and
according to the data summarized by Westlake (1975) there are numerous studies indicating dry
weight biomasses of around 5 000 kg/ha at fertile sites, such as backwaters and lagoons or within
stabilized river channels. They are however, rarer in tropical systems. This appears to be mainly due
to high turbidity or shading by floating meadows and other floating plants in the tropics, which
prevent the development of species with no aerial parts. Aquatics with floating leaves are commoner
in the slacks of pool systems, in quiet bays, openings and backwaters or just off the open waters fringe
of the floating vegetation. Thus various species of Ceratophyllum, Trapa, Naja and Nymphaea are
widely if sparsely distributed through quiet river channels and in most of the permanent waters of the
world's floodplains. They also appear temporarily in the seasonally inundated area where they are
concentrated in the major depressions and channels.
In the floodplain lakes of the Danube, Potomogeton perfoliatus, Valisneria spiralis, Ceratophyllum
sp. as well as Trapa natans and Nymphaea alba contribute a significant portion of the biomass and
total plant production (Academia Republicii Socialiste Romania, 1967). In Crapina lagoon, for
instance, Nicolau (1952) calculated a biomass of P. perfoliatus of 1 749 kg/ha². Estimates in
Academia Republicii Socialiste Romania (1967) indicated productions of between 2.52 and 4.55
gC/m². Production is high in June-July but had fallen by September. The lowest values were recorded
in December. In the Pongolo river considerable standing crops of Potomogeton crispus of up to 1100
kg/ha² (dry wt) were recorded at the end of the growing season by Heeg and Breen (1982) indicating
that submersed plants can also be important in semi-tropical systems.
Floating vegetation
One of the most conspicuous features of the tropical floodplain swamp communities are the vast areas
occupied by floating vegetation; this may take the form of free floating types or of Sudd and meadow
forming varieties.
Free floating forms. The same types of small free floating plant tend to recur throughout the world's
swamps. Principal among these are Eichhornia crassipes, Pistia stratiotes, Azolla sp. and Salvinia sp.,
which form extensive mats which may choke water ways and induce deoxygenated conditions under
them. They are influenced by the wind and current, and Bonetto (1975) has illustrated the manner in
which this type of floating vegetation can accumulate at the outlet of depression lakes, clogging
normal drainage until released into the main channel as “embalsados” (Fig. 3.8) Eichhornia crassipes,
can double in number every 8–10 days in warm nutrient-rich waters (Wolverton and McDonald,
1976), but normal production is possibly less than this in the nutrient-poor swamps of floodplains.

98
Dymond (in Westlake, 1963), for example, found a biomass corresponding to 1.4 kg dry weight/m²
which is equivalent to an annual organic production of 11–33 t/ha.
Salvinia sp. can also form a major nuisance when introduced into waterways from which they were
previously absent. In the Sepik River of Papua New Guinea, for instance, S. molesta occupied nearly
all the free water in the system after its introduction and attained densities up to 6.8 kg of living
material (fresh out) and 2.8 kg of dead and decomposing material/m² [equivalent to 96 t/ha of
biomass], Mitchell et al. (1980). The mats effectively cut off all light from the underlying waters and
also produce significant reductions in Dissolved Oxygen concentrations.
Floating meadows
Although floating vegetation fringes temperate river channels, it is in the tropics that it finds its
greatest expansion. In Africa four species dominate the more deeply inundated parts of the floodplains
and regularly form vast floating mats fringing river channels and floodplain lagoons. During the
floods portions of these mats are liable to break up to form floating islands or sudds. These are
Cyperus papyrus, Echinochloa pyramidalis, E. stagnina and Vossia cuspidata. Of these, only C.
papyrus is dependent on permanent water for its survival and reaches standing crops of between 10
and 34 t/ha dry (Thompson et al., 1979). In the Upemba basin production of C. papyrus ranged
between 50 and 94 t/ha/yr.

99
Figure 3.8 Mechanism of release of floating vegetation masses “embalsados” during the flood cycle:
(A) ponds with “embalsados” in flood condition; (B) drainage clogged by vegetation after
flooding; (C) the same situation during the rains; (D) release of the “embalsados” into the
river. (After Bonetto, 1975)
In the Amazon basin the varzea grasses desiccate during the dry season, although some may be semi-
aquatic and have alternative dry season forms. During rising water there is an explosive growth phase
which culminates after 4–6 months in flowering, followed by senescence and death. During the six
month growing season Junk (1970) estimated Paspalam repens to attain 6–8 t (dry weight)/ha with a
production surplus of 3–5 t/ha. Marlier (1967) estimated a mean standing crop of 96 t/ha fresh weight
equivalent to 9.1 t/ha dry weight in Lago Redondo. The flora of Amazonian varzea lakes having high
oscillations in water level (Fig. 3.9A) are composed mainly of P. repens and E. polystachya which die
back completely at times of low water. In lakes with less extreme variations, more permanent islands
of Leersia hexandra form and these become secondarily colonized with Cyperus sp. and eventually
small trees and other non aquatic plants (Fig. 3.9B).

100
Figure 3.9 Schematic distribution of vegetation of two types of Amazonian varzea lake: (A) with big
oscillations in water level; (B) with small oscillations in water level. (After Junk, 1970)
After the floods stranded floating vegetation decomposes extremely rapidly with 40–50% of dry
matter being eliminated in the first 14 days and 60–70% within 50 days of desiccation. The nutrients
concentrated within it are thus rapidly made available to other elements of the community,
particularly to detritus feeders of various sorts. According to Howard-Williams and Junk (1976) the
nutrient levels and nature of the decomposition litter varies very little with plant type, which means
that, irrespective of origin, the detritus becomes chemically more and more uniform as decomposition
proceeds.

101
Floodplain meadows
The majority of savannah floodplains are covered with various types of grassland which follow a
fairly typical annual course in all but the most highly cultivated plains. Junk illustrates such a
temporal succession for grasses from the Amazonian floodplain (Fig. 3.10) whereby the dominant
plants on the plain change annually in conjunction with the flood cycle. Most floodplain grasses are
rhyzomatous and after the floods subside the enormous elaboration of wet season growth is burnt off
either naturally or by man set fires. New growth of the dry season type is grazed by cattle or by wild
game and burning may occur at intervals throughout the dry season. Very high levels of production
have been recorded during this phase, of which the 23 kg/ha/day recorded by Heeg and Breen (1982)
for Cynodon dactylon on the Pongolo plain may be considered normal. Although much of this is
grazed some 825 kg/ha remain at the end of the dry season which means that in a single year some 34
tons of C. dactylon is submerged representing about 25 tons of wet organic matter as input to the
aquatic system. In the floods some of these grasses may take on a wet season form with floating nodes
which themselves develop roots, as in the case of Vossia and Echinochloa, or may remain rooted in
the bottom but increase their stem length. Oryza barthii for instance, stands about 50 cm above the
water surface irrespective of depth even when submerged under 2–3 m of water. Growth is very fast,
as much as 1 m in two weeks, and productions of up to 2.5 t/ha can be achieved in five weeks. Total
annual production can be reasonably high, and while dry grassland will not produce more than 2–3
t/ha/yr, productions of 10–20 t/ha/yr are not regarded as unreasonable for seasonally flooded
grasslands by Thompson (1976).
Figure 3.10 Temporal succession of grasses on Amazonian floodplains correlated with changes in
water level throughout the hydrological cycle. (After Junk, 1983)

102
The floodplain grasslands are highly modified both by the natural flood regime, which selects for
flood-resistant forms, but also by burning and grazing which prevent the recolonization of the plain by
flood-resistant scrub bushes (Greenway and Vasey-FitzGerald, 1969).
Emergent vegetation: Emergent aquatic vegetation, such as Typha, Scirpus and Phragmites are
widespread but localized in shallow muddy areas, and also tend to colonize sheltered alluvial banks of
rivers in both temperate and tropical areas. Certain alkaline soils seem to favour these forms; the plain
of reeds of the Mekong system where Eleocharis equisetima appears in great abundance (Le-Van-
Dang, 1970) is a good example for this.
Stands of emergent vegetation, one established, are a major modifier of the ecosystem. By changing
flow patterns they increase siltation accelerating the filling of floodplains depressions and
contributing to the accretion of silt on islands on the main channel. They buffer the effects of scour
and with their root and rhizome masses stabilize the river channel and its islands. On the temperate
floodplain of the Danube, Bothnariuc (1967) recorded 1.2 kg/m² (dry weight) of plants from a marshy
pond, and characterized the succession from ghiol to japse as a progression from a primary production
based on phytoplankton in the open water “ghiol” to one based on higher emergent vegetation in the
marshy “japse”.
Westlake (1963) placed tropical reed swamps as one of the most productive communities of plants
with organic productions of up to 75 t/ha/yr. Analysis of Cyperus papyrus has shown it to have about
20 kg total biomass/m² in dense stands, the aerial portions forming 60–70 percent of the biomass
(Thompson in Westlake, 1957), although 3–5 kg/m² were considered more likely over larger areas.
Typha domingensis reaches a similar total biomass of 4.4 kg/m² with 52 percent underground.
However, freshwater macrophytes in Malaysia are reported as having a standing stock of 370–520
g/m² (Wassink, 1975), a tenth of the values for African swamps.
The role of higher vegetation in nutrient balance
There seems no doubt that the major part of the primary production of the floodplain is concentrated
in the higher vegetation and principally in the perennial grasses. These die through desiccation, and by
decay, burning or digestion by grazing animals are returned to the dry soil as nutrient, ash or dung
ready for solution and utilization during the next flood phase. Floating weeds and sudd islands, which
are often present in huge amounts, are also a mechanism for the translocation of nutrients within the
system, representing a very real loss as they are swept downstream.
The remarks of Howard-Williams and Lenton (1975) summarizing the role attributed to higher
vegetation in the littoral flood zone of lakes, can equally be applied to the floodplains and swamp
vegetation:
(i) the vegetation provides a diverse habitat for animals and plants;
(ii) it acts as a filter and trap for allochthonous and autochthonous materials which in turn serve
as nutrients for the plant communities themselves or for the associated aufwuchs and fish
communities;
(iii) the nutrient pump effect of the emergent vegetation reputedly increases the concentration of
elements in littoral areas of lakes and almost certainly does so in newly flooded waters of the
floodplain;
(iv) it contributes to the autotrophic production in that as it decays it forms a rich detritus which
is utilized as food by may organisms;

103
(v) furthermore, Howard-Williams and Junk (1977), suggested that the aquatic plants of the
Amazon, and presumably of other nutrient poor systems, act as nutrient reservoirs and play a
major role in the biogeochemical cycling of nutrients within the aquatic (floodable
ecosystem) of the varzea. The vegetation which proliferates during the flood is deposited on
the floodplain, on the exposed banks of the channels or in the lagoons where it decays. This
material is then subjected to leaching and elution by tropical rainstorms and is stored in the
soil until the next rainy season. This results in the conservation of salts in the floodplain
rather than their being swept downstream dissolved in the main water mass.
In forested rivers and streams, riparian or floodplain vegetation contributes considerable amounts of
nutrients to the system. This has been best studied in low order temperate rivers where the autumn-
shed leaves form an important source of allochthonous organic matter as well as food for
invertebrates. Kaushik and Hynes (1971) have documented the decomposition of such leaves to show
the build up of protein, nitrogen and phosphorus in water where the decomposing leaves serve not
only to nourish invertebrates but to provide food for detritivore fish. Indeed in nutrient poor rivers the
leaf fall may be the biggest single source of nutrients to the aquatic system.
CHAPTER 4
SECONDARY PRODUCTION IN RIVERS
ZOOPLANKTON AND DRIFT
Two groupings of small animal forms are found within the flowing waters of main river channels; the
true zooplankton and the drift. It is often very difficult to distinguish between the two and in many
cases the organisms comprising them are fairly similar. In torrential and low order streams the
majority of the drift arises from the dislodging of aquatic benthic organisms and from terrestrial
insects dropping from overhanging vegetation or swept into the channel with surface flow. In larger
slow flowing potamonic rivers the role of these dislodged and entrained organisms diminishes, and
the role of a true zooplankton increases accordingly. Kammeyn and Novotny, (1977) identified four
components of the drift in the Missouri river: (i) particulate organic matter (POM); (ii)
macroinvertebrates; (iii) larval fish, and (iv) zooplankton. Concentrations of POM and detritus ranged
from 11 mg/m³ in anabranches through 783 mg/m³ at the main channel border to 2331 mg/m³ in the
main channel in channelized reaches. Unchannelized reaches had less of the material with 18–135
mg/m³ in anabranches, 64–111 mg/m³ at the main channel border and 111–505 mg/m³ in the main
channel. Although concentration of POM may increase in channelized reaches of the Missouri, Morris
et al., 1968 recorded an opposing trend where mean values of drift organisms in unchannelized
reaches were 1980 mg/m³ and only 230 mg/m³ in channelized reaches. These estimates indicate that
about 526 kg of drift organism flow past a fixed point during the day in an unchannelized reach and
204 kg/day in a channelized reach. As with other workers Morris et al. found the species composition
of the drift was very dissimilar from that of the benthos in large river channels and more closely
resembled the composition of the aufwuchs, although about 20% of the drift by weight were terrestrial
(allochthonous) forms. Numbers of drift organisms increased at night when densities were often
double those during the day, and a marked diurnal periodicity in the drift has previously been
described from many temperate rivers (see Hynes, 1970). In tropical waters too the number of
organisms increases at night and particularly at nightfall (Elouard et Leveque, 1977). The abundance
of larval and juvenile fish in the drift, which shows no such periodicity may, in part, be attributed to
the downstream movement of yolk-sac and post yolk-sac larvae, and in part to the presence of
individuals feeding on the drift organisms.

104
Information on zooplankton in rivers, particularly those of the tropics, is rather sparse but existing
studies indicate that similar factors influence zooplankton densities as those influencing
phytoplankton. The abundance of phytoplankton itself is possibly one of the major determining
features and peaks in zooplankton abundance are apt to follow peaks in phytoplankton as exemplified
for the Paranà river by Bonetto (1976) (Fig. 4.1).
Variations in zooplankton abundance have, however, been attributed primarily to differences in flow,
with a number of other factors including turbidity, dissolved oxygen concentration and conductivity
also playing a minor role. Under normal flow regimes only low densities of zooplankton are present in
the main channel of rivers. For example only about 5 250 individuals/m³ of protozoa, crustacea and
rotifers have been recorded from the Mekong river (Sidthimunkra, 1970). Rzoska (1974) also
remarked on the small quantities of zooplankton found in the Sudd although Holden and Green (1960)
had noted higher numbers of up to 16 000 individuals/m³ in the Sokoto river. In fact considerable
differences are to be found between the planktonic fauna of the main channel and that of the lentic
waters of side arms and floodplain and the zooplankton of both lotic and lentic waters fluctuates in
abundance according to season. Thus whilst only 1 000 individuals/m³ of zooplankton were found in
the Upper Paranà River at high waters 10 000 individuals/m³ were present at low water. Similarly in
the Blue Nile at Kartoum planktonic Crustacea were present at about 20 000 individuals/m³ during
periods of moderate flow but these rose to up to 100 000 individuals/m³ at low water (Tailing and
Rzoska 1967). In the Nile Sudd densities were also higher in the dry season (mean of 4 460
individuals/m³ for four sites) than in the wet (mean of 2 070 individuals from the same four sites). The
abundance of zooplankton in temperate rivers may be seasonally high relative to that in tropical
waters, as in the Danube where Enaceanu (1964) recorded only 550 individuals/m³ in winter which
rose to over 1 million individuals/m³ in the September peak. This superabundance of organisms may
have been due to enrichment of the Danube waters by domestic and industrial contaminants but there
is no doubt that, were flow conditions unfavourable, no such development would have occurred.

105
Figure 4.1 Variations in abundance of phyto- and zooplankton in the Parana River at the level of
Parana relative to temperature and river level. (After Bonetto, 1976)
Zooplankton is much more common in the floodplain pools and backwaters where its abundance per
unit volume is usually inversely correlated with the amount of water in the system. Vranovsky (1974),
for instance, found the average biomass of zooplankton to be 14–15 times higher in the Baka side arm
of the Danube than in the main stream (mean of 6.75 g/m³ for two years in the river). During periods
of low or nil flow, greatly increased biomasses were present, especially in the isolated side arms
where values of up to 30 times those of the main river were recorded. Similarly Sidthimunka (1970)
was able to record 131 000 individuals/m³ in the Mong river, while only about 5 000 were present in
the main stem of the Mekong. In the standing waters of tropical floodplains too, there is a widespread
tendency for greater concentrations of organisms to be present in the dry season, as is the case with
phytoplankton. Like phytoplankton, however, the decrease in relative density may only be a dilution
effect and the number of organisms over the floodplain as a whole may in fact be higher during the
wet season than during the dry. Bonetto (1975) recorded increases in numbers of zooplankters per unit
volume even during the floods of the Parana, although elsewhere in the system the lowest values were
found during the same period (Bonetto and de Ferrato, 1966). Increases in specific diversity often
occur simultaneously with increases in absolute number. Rivers subject to marked annual thermal
variations tend to have peaks of zooplankton abundance in the spring and summer months (Bonetto

106
and de Ferrato, 1966; Bonetto, 1975), or in the spring and autumn (Ertl, 1966; Osmera, 1973) and
these do not always coincide with the period of lowest water. The abundance of organisms varies and
considerable numbers are occasionally found in permanent lakes. A summary of estimates of
zooplankton abundance from floodplain standing waters is shown in Tables 4.1 and 4.2.
Table 4.1
Estimates of biomass of zooplankton organisms in standing waters of floodplainsa
River Biomass g/m³ Authority
Danube:
Dyje:
Locality 1 2.0–4.6 (471.6 max.) Osmera, 1973
Locality 2 7.75
Locality 3 7.5
Baka:
Side arms 6.75 (mean of 2 years) Vranovsky, 1974
Amazon:
Lago Redondo 1.61 Marlier, 1968
Nile Sudd:
Aljab 0.002–0.001 Monakov, 1968
Shambe 0.002–0.40
Jor 0.021–0.049
No 0.138–0.107
Atar 0.246
Magdalena: 0.92 (mean for 4 lagoons April)
1.95 (mean for 5 lagoons June) Mikkola & Arias, 1976
1.53 (mean for 6 lagoons August)
a These figures are often based on only a few observations and do not therefore necessarily reflect
the full range of variations of the parameters measured
Flow is also slowed where waters are impounded behind dams and concentrations of zooplankton
reach high levels in the reservoirs thus formed. This high concentration of organisms forms a locally
increased concentration of zooplanktonic organisms at the point of discharge from the dam which
diminishes slowly as the water mass moves downstream, Dzyuban in Mordukhai-Boltovskoi (1979)
was thus able to state that flow regulation following the construction of the Volgograd reservoir has
brought about great changes in the composition and abundance of the zooplankton in the lower Volga.
These are now determined by zooplankton discharged from reservoirs together with the tendency of
the river to restore a rheophilic type community. Thus zooplankton decreases in abundance
downstream of the discharge point. Similar changes have occurred in the delta, mainly because of the
decrease in the overall volume of water and the suppression of the spring flood. High concentrations
of zooplankton were much more common in the floodplain pools of the upper delta where species
with high capacities for parthenogenetic reproduction attained up to 3 g/m³. The life of these pools has
been shortened and many have disappeared with a consequent reduction in overall zooplankton
abundance. By contrast an increase in emergent vegetation has provided favourable habitats for
phytophylic, periphytic and benthic forms. Extensive work on the zooplankton of the Nile showed
similar changes below the various dams. For instance Brook and Rzoska's (1954) data for river
plankton above and below Gebel Aulia showed a change in dominance from Copepoda upstream of
the lake to Cladocera below the dam. With the addition of the Roseires and Aswan dams the Nile has
undergone multiple regulation turning it into a cascade system in which the zooplankton community
has changed from a riverine species dominated fauna to one more typical of lacustrine systems. In the
lower Nile below the Aswan dam zooplankton density varied between 12 000 and 133 000

107
individuals/m³ with a more or less steady increase in density downstream. Since the closure of the
dam, this lowest river reach has been converted to a semi-lenitic body of water.
Table 4.2
Population estimates of zooplankton organisms in standing waters on floodplainsa
River No. of individuals/m Remarks Authority
Danube:
Erec lagoon 110 000 Minimum: December Ertl, 1966
4 006 000 Maximum: April
Husie lagoon 616 000 Minimum: February
8 493 000 Maximum: June
Dyje: locality 1 10 000 Minimum: February Osmera, 1973
500 000 Maximum: May-July
locality 2 2 000 Minimum: Spring flood
10 000 000 Maximum: June
Missouri:
Backwaters 6 670 Mean: April-October Kallemeyn & Novotny, 1977
Dvina: 247 999 400 000 Zhadin and Gerd (1970)
Oka: 950 000 Spring
1 281 000 Summer
Parana:
Don Felipe lagoon 17 000 Minimum: June Bonetto & De Ferrate, 1966
1 200 000 Maximum: October
Los Espejos lagoon 26 000 Minimum: July
277 000 Maximum: December
Flores lagoon 12 000 Minimum: January
830 000 Maximum: September
Laguna Totoras 80 000 Minimum: Falling water
November Bonetto et al., 1948
930 000 Maximum: High water
August
Laguna Gonzalez 6 330 000 Minimum: Falling water
October Bonetto et al., 1948
24 000 000 Maximum: Low water
December
Laguna La Brava 536 000 Minimum: High water
May Bonetto et al., 1978
761 000 Maximum: Low water
Laguna Sirena 12 000 Minimum: Low water CECOAL, 1977
536 000 Maximum: High water
Magdalena:
Los Ponches 5 380 000 Maximum: Low water Arias, 1977
Machado 4 360 000 Maximum: Low water
Pinalito 25 300 Maximum: Low water
Sokoto: 27 000 Maximum: Dry season Holden & Green, 1961
Mekong:
Nong pla pak swamp 38 500 Random sample Sidthimunka, 1970
Ping and Nan Rivers 136 234 Outflow Junk, 1976
Bung Borapet 122 279 Inflow
Amazon:
Lago Preto de Eva 738 000 Mean Marlier, 1967

108
Lago Calado 107 087 Junk, 1976
Nile:
Aljab lagoon 180 January Monakov, 1968
3 000 May
Shambe lagoon 600 February
3 000 May
Jor lagoon 2 700 February
12 400 May
No lagoon 8 000 February
53 000 May
Atar lagoon 59 000 April
a These figures are often based on only a few observations and do not therefore necessarily reflect
the full range of variations of the parameters measured
Kallemeyn and Novotney (1977) also found that numbers of zooplankters declined downstream of the
major impoundments in the Missouri river where the presence of zooplankton is strongly correlated
with discharge from the Lewis and Clark lakes. A 51% decrease in abundance 16 km downstream of
the point of discharge, 61% at 38 km and 70% at 145 km was shown by Hesse et al. (1982). High
concentrations of zooplankton also arise from the injection of organisms into the main river channel
from chutes and backwaters. In the Missouri this resulted in greater numbers in unchannelized section
of the river where 6 670 individuals/m³ were present in backwaters, 12 110 individuals/m³ in chutes,
10 864 individuals/m³ in the main channel adjacent to the river bank and 12 245 individuals/m³ in the
main channel itself. There was a strong peak of abundance in May at 370 mg/m³ when the rains
flushed out the backwaters, diminishing to less than 20 mg/m³ in winter. Vranovsky (1974) noted the
same phenomenon in the Danube where maximum values of zooplankton biomass (300–600 mg/m³)
occurred in autumn and the average amount of zooplankton drifting past a fixed point reached as high
as 260 g/s in 1967 due to the wash out of organisms from the side arms.
Sudden seasonal pulses in total zooplankton numbers seem to arise mainly by increases in rotifers,
although the other major components of the zooplankton, the copepods and cladocera, also have
characteristic peaks. Nauplii are common and are often washed out of the standing waters into the
main channel during rising floods. Rhizopods have also been noted as an important element of the
plankton at this time (Holden and Green, 1960; Green, 1963).
Some correlation between conductivity and zooplankton numbers has been found in vegetation
flanking the main channel of the Zambezi River. Here maxima of both conductivity and zooplankton
numbers (110,000 Cladocera and Copepoda m³) occurred during the dry season. Both parameters
were minimal during the peak flood when zooplankton was nearly absent.
The degree of vegetation also appears critical and the greatest number of zooplanktonic organisms is
commonly found in waters with at least sparse vegetation (Bonetto, 1975). Their density also
increases locally at or near the open water/vegetation interface, under mats of floating vegetation and
associated with submersed plants. This implies that there is a succession of planktonic organisms
within the standing waters of the floodplain, which corresponds to the general evolution of such
bodies of water, from the open water to the heavily vegetated state, as discussed by Botnariuc (1967).
Comparative studies that demonstrate this are few, although Green (1972 and 1972a) has
demonstrated such a succession in the floodplain lakes of the River Suià Missú in Brazil. As the
vegetated areas are generally more productive than open water areas, there is presumably a
comparable evolution in productivity per unit area. This would, however, be compensated for by the
diminution in size of the individual water body as it progresses through silting from the open water to
the marshy state. Work on lagoons of the Danube floodplain by Ertl (1966) and the Elbe floodplain by
Novotna and Korinek (1966), indicates that the density of the fish population influences the
composition and abundance of the zooplankton. Detailed work on ecological inter-reactions of this

109
nature is unfortunately lacking in tropical lagoons, although it is to be suspected that similar
relationships may also be found there.
Zooplankton densities are usually considered to be low in the main mass of water on the floodplain,
but the shallow littoral of the flood zone may support considerable quantities f planktonic forms. The
higher conductivity and temperature, and local availability of oxygen can support blooms of
zooplankton that can be readily observed at the water’s edge. Unfortunately little quantitative
information supports this as such areas have been relatively little investigated, but the high values for
the zooplankton from the Baka and Dyje backwaters of the Danube in Tables 4.1 and 4.2 may be
attributed to the fact that these water bodies are mainly littoral in nature (less than 1.5 m deep) with
considerable submersed vegetation. In a comparison between littoral and pelagic zones in backwaters,
Osmera (1973) showed that mean biomass from the littoral zone was 7.75 mg/1 as compared with
1.64 mg/1 in the pelagic zone of the same lagoon.
Survival of the adverse conditions of the flood poses certain problems for planktonic organisms which
are readily washed away by the increased flows. Nevertheless, zooplanktonic organisms reappear
rapidly when more favourable low flow patterns are re-established. This has been attributed to
reservoirs of organisms in upstream habitats, but Moghraby (1977) considered the 50–100
organisms/m present in riverside pools of the Blue Nile as inadequate to serve as sources for the
regeneration of the population. Instead he produced evidence that the adults or the eggs of many
species enter a diapause as temperatures are lowered and silt concentrations increased during the
earlier part of the flood. He found abundant pockets of diapausing individuals in various types of
bottom deposit and showed experimentally that they were only released when silt concentrations
dropped and temperatures rose to the dry season norms.
ANIMAL COMMUNITIES ASSOCIATED WITH FLOATING AND SUBMERSED
VEGETATION
Floating vegetation supports rich and varied animal communities either as a free living mesofauna in
the open water among the plant stems or as an epifauna in the root masses of floating plants (i.e.,
pleuston). These have been studied in most detail in the root masses of the floating meadow grasses
Paspalum repens and Echinochloa polystachya of the Amazon system from where many species of
crustacea, insect nymphs, oligochaetes and molluscs were identified. Junk (1973a) distinguished three
major biotopes according to fauna and flora. The first of these, the flowing whitewater biotope-
consists of the stands of vegetation bordering the main river channel. Here faunal densities on the
exposed outer fringe were low, possibly due to current, which sweeps organisms away, and to large
amounts of inorganic sediment, which hinder feeding. Faunal abundance increased from the fringe
towards the centre of the stand or mat and total abundance at the centre could be as-high as 100 000
individuals/m² although the norm was around 50 000 individuals/m². Biomass increased from 0.3
g/m² (dry weight) or 1.5 g/m² (wet weight) at the fringes to 4.2 g/m² (dry weight) or 20 g/m² (wet
weight) in sheltered places within the stand. A second biotope, well-oxygenated, sedimented
whitewater lagoons, undergoes fairly wide annual fluctuations in level which periodically destroy the
aquatic vegetation. During the optimal growing period large numbers of individuals occurred,
abundance usually ranged between 100 000 to 300 000/m², although densities of up to 700 000/m²
were recorded. Faunal density was evenly distributed and biomass varied between 2.5 and 11.6 g/m²
(dry weight) or 12–62 g/m² (wet weight). Marlier (1967) found similar values ranging from 3.9 to
13.9 g/m² (dry weight) or 18.9–39.7 g/m² (wet weight) from three samples of floating vegetation in
another lake of this type. The third biotope - sedimented whitewater lakes - had thick stands of
vegetation supporting little or no dissolved oxygen. Water regimes are more stable than those of the
former type of lagoon. In the peripheral zone of the vegetation mats, faunal abundance was about the
same as in the well-oxygenated lake, but various groups of organisms rapidly disappeared due to
oxygen stress and abundance was very low (0.16–0.29 g/m² - dry weight) even only a few metres
from the outer edge.

110
This community is also very important in the middle Paraná River where the abundance and density
of Eichhornia, Pistia, Salvinia and Azolla supplies an sample substrate (Bonetto, Dioni and
Pignalberi, 1969). As examples Bonetto et al. (1978) found 17 000 individuals/m² of molluscs,
oligochaetes, leeches, mites, crustacea and insects in Salvinia herzogii during the summer
(corresponding to high water) and 9 200 individuals/m² during winter in Laguna La Brava of the
Upper Parana; Varela et al. (1978) describe in detail the floral and faunal associations in the floating
islands or “embalsados” of that lagoon; CECOAL (1977) found a maximum value of 20 000
individuals/m² in summer falling to 4 000 individuals/m² in winter for Azolla and Pistia. Also in the
Parana system over 100 taxa at specific or generic level were found in the submerged stoloniferous
masses of Eichhornia (Poi de Neiff and Neiff, 1980) where seasonal changes in the dominance of
organisms and differences between the fauna associated with plants in river and lagoon were also
observed. Here maxima reached 262 000 individuals/m² within the Paspalum repens/Salvinia
hertzogii complex (Poi de Neiff, 1981). Also in Latin America, Zarate and Cubides (1977) found a
range of biomass from 6.56 and 130.7 gm/m² wet weight (Mean 35.14 g/m²) for 17 stations on
lagoons of the Magdalena system. The authors identified 67 taxa of invertebrates in the root masses of
Eichhornia. Similar distributions of organisms have been found in African papyrus swamps where the
zone close to the interface with the open water was considered the richest habitat in the system by
Rzoská (1974). Many species of oligochaetes, Bryozoa, Protozoa, Crustacea, insects and molluscs
have been found among the papyrus, reeds, Echinochloa and Eichhornia of the Nile Sudd, Monakov
(1969). Few quantitative samples have been taken of this habitat, but 5 minute pond net sweeps
captured as many as 547 individual Crustacea (Rzoska, 1974). Monakov (loc. cit.), who also
attempted to quantify the biomass of the Entomostraca, found that the fringe was 10–100 times richer
than the open water of the Sudd.
Petr (1968) found between 5 000 and 16 000 individuals/m² in the roots of Pistia from Volta Lake. In
the Sudd, Rzoská (1974) found up to 300 individual animals per Pistia plant, although densities
increased in quiet stretches of the river, notably in the swamps and reached a peak in river lagoons.
Fewer individuals were noted in floating vegetation in Bangula lagoon by Shepherd (1976). Here a
mean of 388 individuals of insect, oligochaete, crustacea and molluscs were present per m² of
Nymphaea and a similar assemblage at 210 individuals/m² was found in Pistia.
The weight of animals associated with the roots of Eichhornia was also high in the cienagas of the
Magdalena river, where Kapetsky et al. (1977) reported a mean of 35.1 g/m² from 17 sample sites in
10 different water bodies. Values ranged from 6.9 to 130 g/m².
Investigations of submersed vegetation in two lakes of the maritime delta of the Danube show the
considerable differences that can arise in faunal densities depending on relative distribution of support
plants. In Porcu lake Ceratophyllum demersum covers about 64% of the bottom and Nitellopsis obtusa
about 4% whereas in Rosu lake C. demersum occupied 14% and N. obtusa 53% of the bottom. Both
plants form a similar type of substrate and host similar species of Oligochaeta, Trichoptera and
(Chironomida) yet total quantities in Porcu lake, at 1.8 kcal/m² (0.36 g/m²) in May and 93.4 kcal/m²
(18.7 g/m²) in August exceeded the 2.2 kcal/m² (0.44 g/m²) in May and a maximum of 18.7 kcal/m²
(3.74 g/m²) in June. The differences in biomass were attributed to the greater density of organisms
supported by Ceratophyllum (Izvoranu, 1982).
A particular community has been identified in the umbels of Cyperus papyrus (Thornton, 1957). This
consists principally of terrestrial forms which pass their lives in this specialized habitat, but which
contribute to the allochthonous food supply of the swamp waters.
BENTHOS
The literature on the benthos of running waters has been reviewed by Hynes (1970) who concluded
that similar elements of the fauna of hard substrates are common to streams and rivers all over the
world. The benthic fauna of hard, stony runs and riffles is richer than that of the silty reaches and

111
pools of the rhithron in both number of species and in total biomass. The communities of Soviet rivers
have been classified on the basis of both substrate and current velocity (Zhadin and Gerd, 1970) as
follows:
- Lithorheophilic communities inhabit solid substrates in flowing waters and consist mainly of large
insects.
- Psammorheophilic communities occur over sand bottoms in flowing waters and consist of small
arthropods and protozoons living in the interstices of the sand.
- Argillorheophilic communities inhabit clay substrates and are mostly sessile or burrowing forms.
- Pelorheophilic communities live in silt in flowing waters.
- Pelophylic communities are found in silt in still waters.
- Phytophylic forms live in backwaters rich in plants.
Benthic fauna is, as a rule, better developed on stable bottoms. On mobile bottoms sessile insect
larvae with their sensitive ducts or tissue structures and also moving molluscs are readily displaced.
Other more mobile forms such as Asellus or Gammarus may equally be at risk through the risks of
mechanical damage.
Biomass was thus found to be closely related to the type of community. For instance, in the Dnieper
Lithorheophils had a standing crop of about 1 kg/m², Pelorheophils about 10 g/m² and psalmophils
about 1 g/m² although isolated pelophils in cut-off lakes attained over 100 g/m².
In the Danube pelorheophilic (with 16.5–99.2 g/m²), psammopelorheophilic (with 22.7–69.2 g/m²)
and Argillorheophilic (with 71.6 g/m²) communities all and considerably higher standing crops than
the psamorheophilic community.
Russev (1967 and 1981) has also described the distribution of biomass according to location and
bottom type in the Danube with mean values of 72.5 g/m² over leaf litter, 47.4 g/m² over mud, 26.4
g/m² over clay and 0.24 g/m² over sand which is far less stable a substrate. Furthermore water speeds
in excess of 2–3 m/s¹ inhibit the development of benthos in the centre of the channel forcing it to
concentrate mainly in protected shingle areas was of the littoral (Russev, 1982). The mean biomass on
the Bulgaria reaches of the river was 35.22 g/m² of which 34.21 g/m² were molluscs. Similar mean
values of 38.9 g/m² (total) and 34.21 g/m² (molluscs) were obtained in the autumn. Biomass per unit
area varied considerably with water level. The mean biomass (without molluscs) was only 0.76 g/m²
at high water (April-August), whereas at mid-water it was 1.94 g/m² and at low water 6.34 g/m². The
much higher values obtained as water level decreased were attributed to the dislodging and transport
downstream of benthos at times of high flood, and the dispersion of benthic organisms over a wider
area during the flood period.
In tropical rivers Bishop (1973) commented on the similarity of rheophilic rainforest communities in
Africa, Sri Lanka, tropical S. America and the Gombak River Malaysia. The lowland reaches of rivers
share many features, but relatively little information is available3 on the benthic fauna of the slow-
flowing silt-laden rivers associated with floodplains.3 Such that does exist indicates a generally poor
fauna consisting of a relatively small number of species consistent with Zhadin and Gerds conclusions
of the low biomass of pelorheophils in Russian rivers. Bonetto and Ezcura (1964) reached similar
conclusions based on observations on the Parana River which showed the benthic fauna to diminish in
diversity and, abundance as the current slows. Maximum values may reach over 71 000
individuals/m² in the region of Yacireta as compared with 17 000 individuals/m² at Posadas, 23 000 at

112
Ituzaingo and 13 000 at Ita bate (CECOAL 1979). Data from the Amazon (Junk 1971) also indicate a
poor fauna in the main river, although experiments with artificial substrates where 12 900
individuals/m² settled in four days showed populations in blackwater rivers to be higher than
expected. Monakov (1969) also concluded that the bottom fauna is monotonous in the main channels
of the main channels of the Sudd where only small groups of Chironomids and Oligochaetes inhabit
the sand and mud bottoms. In general biomass ranged between 0 and 0.2 g/m² although Rzoska
questioned this conclusion on the basis of the small number of samples taken. Further downstream,
between the Aswan dam and the sea, the macrobenthos is dominated by Molluscs although some
Polychaetes and Oligochaetes were also present. Quantitatively biomass varied between 1–9 g/m³
rising to the unusual value of 53.61 g/m³.
Differences in species composition have also been recorded from some rivers, for instance the
mollusc fauna of the Niger consists mainly of abundant Aetheria elliptica, Aspatharia, Mutela dubia
and Viviparus unicolor in muddy reaches, contrasted with rare individuals of Corbula fluminialis,
Mutela rostrata, Caelatura aegyptica and Cleopatra bulminoides in sandy reaches (Blanc et al.,
1955). Large aggregations bivalve molluscs also occur in the Volga, Dneister, Dneiper and Don. In
the Volga Dreissena contribute local standing crops of several kg/m². Otherwise the benthos of the
river is poor in the main channel. A high benthic biomass of 400 g/m² has also been recorded from the
many tributaries of the Mekong (Sidthmunka, 1970) which again was almost entirely due to a
concentration of molluscs.
In the Missouri river there are no data to show the benthic populations before impoundment, but
studies by Morris et al. (1968) indicated that there is relatively little difference between the benthic
biomass per unit area in channelized and unchannelized reaches. In the first case mean biomass was
calculated as 0.08 g/m² and in the second at 0.07 g/m². These means, however, conceal considerable
variations with locality as within the river biomass varied between 0.019 (0.048) 0.99 g/m² in
unchannelized reaches of the main stream whereas in channelized reaches it varied between 0.002
(0.007) 0.022 g/m². On mud banks the biomass was higher in channelized reaches at O.058 (0.803)
2.678 g/m² than in unchannelized reaches where it was 0.111 (0.267) 0.349 g/m². The general scarcity
of benthos was attributed to the general nature of the Missouri river in that it has shifting substrates,
high silt load, fluctuating water levels and absence of aquatic vegetation, all of which militate against
high production of benthic organisms. However an overall loss of benthos occurred in channelized
reaches where the area available for colonization was reduced by 67% over the unchannelized
reaches. Earlier Berner (1951) had found similar values ranging between 0.024 g m-2 in the middle of
the channel to 0.258 g/m² nearer the edge, and had constrasted the low values from the Missouri with
the clearer, slower flowing Illinois where a mean of 29.23 g/m² was found by Richardson (1921) at a
time when the river still had its floodplain lakes. Schramm et al. (1974) summarized the data for
benthos from the Middle Mississippi which show similar differences between channelized and
unchannelized reaches. On the basis for this they emphasize the importance of the channel area for
benthic production.
Information obtained from backwaters and standing waters of the floodplain indicate somewhat
higher standing crops than in the main river. In the Romanian portions of the Danube floodplain a
mean biomass of 20.06 ± 9.35 g/m² was calculated from 12 lakes (Academia Republicii Socialiste
Romania, 1967) and in lakes of the Volga delta values of between 7–48 g/m² have been found.
Monakov (1969) recorded biomasses of zoobenthos consisting mainly of Oligochaetes, chironomids
and molluscs ranging from 0.58–4.7 g/m² from five of the floodplain lakes of the Sudd. The mean
values of 2.95 g/m² for the period of high but falling water and 2.07 g/m² for low water compared
with the 0–0.2 g/m² found for the main channel. Benthic biomass quoted for the Nong Pla Pak
swamps of the Mekong floodplain (Sidthimunka, 1970) were of the same order at 1.75 g/m² at a time
when, benthic biomass in the main river was only 0.12 g/m². Similarly a mean of 1.51 g/m² (range 0–
6.51 g/m²) of benthic organisms was found in cienagas of the Magdalena system by Mikkola and
Arias (1976). Here a mean of 1.9 g/m² was found in the open waters, of the Cienagas whereas the
more sheltered bay habitats had a mean density of 3.1 g/m². Arias (1977) found values of 1.76±1.81

113
g/m² for the open waters of 18 cienagas in this system when the bay areas had values of 2.96 ± 1.51
g/m². These variations in biomass were attributed to the often deoxygenated conditions in the deeper
waters of the cienagas.
The tendency for littoral waters to support more benthos than the deeper waters has been confirmed
by numerous other workers. Bonetto et al. (1978) furnished detailed information about the benthic
fauna of two lagoons of the Riachuelo River, a tributary of the Parana system. In the Laguna Totoras
benthos was maximal during low water when total numbers exceeded 114,000 individuals/m². In
summer, during high water, maximum abundance was much less at 46 500 individuals/m². In this
lagoon too, there were considerable differences between the various zones of the lake with maximum
abundance in the sub-littoral zone and the least in the centre. Similar differences between the littoral
and open water zones on one hand and between seasons on the other, were found in Laguna Gonzalez
which, because of its great eutrophication, supported less organisms. In Laguna La Brava the meso-
and macro-benthos contained between 17 500 and 95 300 individuals/m² at low water and 1 700–57
000 individuals/m² at high water depending on location in the lake. Particularly significant in this
water body were thecamoebans which numbers sometimes exceeded 1.5 × 106 cells/m² at low water
but only 0.6 × 106 cells/m² at high water.
Three temporary lagoons in the savannah floodplain of the Rio Branco, N. Brazil, supported mean
weights ranging from 0.16–0.88 g of benthos/m². There were some variations with depth and
individual samples gave values as high as 2.52 g/m². The 0.25 g/m² (dry weight) obtained from the
Lago de Rodondo appears consistent with the range of biomass found in the lagoons, despite Marlier
(1967) opinion that these were very low. Nevertheless higher values have been reported for some
Amazonian lakes, Eittkau et al. (1975) for example, found a mean annual weight of between 0.14 and
6.20 g/m² of benthos, and studies by Reiss (quoted in Sioli, 1975) also revealed biomasses up to 6.2
g/m² in the centre of Amazonian Varzea lakes. By contrast, samples from Lake Tupe, a typical river
lake, and some blackwater várzea lakes, led Reiss (1977) to conclude that the profundal zone of such
bodies of water supports the poorest benthic fauna of any Amazonian lacustrine biotope. Values from
the littoral zone are however much higher, reaching up to 104 g/m².
The range of densities from both African and Latin American floodplain lagoons suggests extreme
values ranging from 0 to about 6 g/m² and mean values of about 2 g/m² for tropical rivers. Rzoska
(1974), however, considered that values from such areas should be treated with caution as the cyclic
emergences of Ephemoptera, Chaoborus and chironomids can influence the standing crop and
biomass profoundly. Very dense localized populations of other organisms can also arise where
conditions are particularly favourable. Botnaruic (1967), for instance, recorded densities of molluscs
of up to 2 657 g/m² in the deep water of Crapina Lake of the Danube floodplain. Values are not
normally this high, and Russev (1967) quoted Russian work on the 8 580 km² Kiliya arm in the
Danube delta, where the mean biomass was 9.45 g/m². The same work indicated an annual production
of 19 235 t (224.19 g/m²) from the whole arm. Work by Lellak (1966) on the benthos of
Czechoslovakian backwaters showed this to be strongly affected by the presence or absence of fish.
Where fish were absent, population densities of Chironomus and Chaoborus larvae were very high.
Similarly, Enaceanu (1957) showed benthic biomass to be higher in enclosures from which fish were
excluded (226.12 kg/hm² mean July-October) than in bottoms freely accessible to fish (70.98 kg/hm²
for the same period) in the same lagoon. In floodplain pools where the composition and density of the
fish populations isolated by receding flood waters are likely to vary, differences in benthos are liable
to arise as a result of diversity in predation pressures. Although the authors quoted above have noted
seasonal fluctuations in the benthos of the standing waters of the floodplain, Bonetto (1975)
considered this the least affected by differences in water level of all communities inhabiting such
habitats.
Submersed vegetation, where present, also acts as a centre of concentration for benthic invertebrates.
Examination by Academia Republicii Socialiste Romania (1967) of 1 550 g of Potomogeton
perfoliatus, collected from an area of 0.5 m², showed 10.489 individuals of insect larvae, Crustacea

114
and molluscs to be present. The total weight of 87.3 g equivalent to 175 g/m² is reasonably high for
the benthic fauna of this system, which normally varies around a mean of 20 ± 9 g/m². Values from
the Bangula lagoon ranged between 0 005 g/m² under Pistia, 0.06 g/m² in open water, 0.378 g/m² in
Ceratophyllum and 0.977 g/m² under Nymphaea. The numbers of individuals present in
Ceratophyllum was still relatively low, 4 385 individuals/m² but was significantly greater than the
densities found in Nymphaea and Pistia. Very large concentrations of invertebrate, including acarine
mites, snails and phytophagous species of insect and mollusc were present in Egeria naias of the
Totoras lagoon where between 10 000 individuals per 1 000 g of plant were present in winter and 320
000 individuals per 1 000 g of plant were present in summer.
On the floodplain itself, where seasonal desiccation occurs, flood season populations of macro-
invertebrates are less well studied. Personal observations have shown enormous densities of
pulmonate snails to be present on the bottom and at the water surface of inundated areas. Carey (1967
and 1971) reported Ephemoptera nymphs, Trichoptera larvae, chironomid larvae, Hemiptera and
molluscs to be very abundant and widely distributed in the inundated zones of the Kafue River,
especially in submerged banks of Najas and Ceratophyllum. The profusion of molluscs on the
floodplain has also been noted on the Central Delta of the Niger by Blanc et al. (1955). In the Parana
River, Bonetto, Dioni and Pignalberi (1969) commented on the restricted number of species which
contributed to the great biomass. Particularly abundant were Unionacea, which made the most
important contribution, despite the low calcium content and relatively low pH. The fact that molluscs
can make a significant contribution to the benthos under these conditions is emphasized by their
presence in Amazonian blackwaters. Certain of these are, however, so poor in minerals that only
small Ferrissidae with conchiolin cases are present (Fittkau, 1967).
In the forest floodplains of the Amazon inundated with whitewaters, benthic organisms are common
and have two production peaks, one at the beginning of the inundation and a second one after
maximum high water in June/July. Reiss' (1977) work on the blackwater river lake Tupe, also
indicated two peaks of abundance for littoral benthos. One peak occurred on the rising flood when 2
559 individuals/m² were recorded, and a second peak occurred at low water when 1 248
individuals/m² were present. Minimum densities during falling water were about 623 individuals/m².
The faunal composition changed completely between the minimum and rising water level peaks. At
low water chironomidae dominated the fauna with minor representation by oligochaetes, Acari and
Corixidae. During rising water Chaoboridae and Ostracoda became steadily more important to the
exclusion of other groups. Blackwater flooded Igapo forests, however, are generally very
impoverished; Irmler (quoted by Sioli, 1975) found a benthic biomass of only 0.2 g/m² in such an
area.
In areas of rapidly fluctuating water level which are subject to seasonal desiccation, it is evident that
mechanisms must exist to survive the dry season. Fittkau et al. (1975) considered three possible
mechanisms: migration, dormancy and recolonization. Hynes (1975) also considered that, in view of
the high degree of adaptation required for resting eggs or burrowing and aestivating larvae,
recolonization is by far the easiest strategy for drought survival. This implies very rapid growth of
larvae, and in fact most larvae and nymphs in the Ghana River studied by him were fully grown after
a month. Whyte (1971) has even noted some chironomid species which reached full size in three
weeks. Studies on Rhodesian streams liable to seasonal desiccation have shown that the re-
establishment of the fauna after the annual resumption of flow can be very rapid (Harrison, 1966).
Oligochaetes, Crustacea and insect larvae appeared within ten days and there was a form of
succession whereby the species composition typical of pools was re-established and stabilizing within
a month. Harrison's conclusions also indicated that pulmonate snails and oligochaetes survived the
drought by aestivating. Some smaller crustacea may have had dormant eggs. Most insects recolonized
the area by the movement of flying adults from more permanent water bodies.
Decapod crustacea, which form part of the macro-benthic community, are a particularly important
element of tropical river fauna. Ecologically they may be considered together with fish on the basis of

115
their size, position in the food chain, behaviour and economic importance as food organisms. Species
of Macrobrachium particularly form the basis for fisheries in many tropical rivers (see Kensley and
Walker (1982) for the Amazon; Robertson (1983) for the Sepik River, FAO/UN (1971) for the Oueme
River and Inyang (1984) for the Lower Niger River) and have also been widely adopted for fish
culture. Similar activities are pursued with Astacus, Procamberus and Pacifastacus in the temperate
waters of USA and Europe.
NEUSTON
The neuston “community” is little discussed by workers on tropical floodplain ecology. Nevertheless,
personal observations have shown the abundance of forms living at the air water interface. Mosquito
larvae and various forms of pulmonate snails are extremely common, as are water striders (Hemiptera
and Coleoptera), mites and spiders, particularly in the sheltered water among the stems of the floating
vegetation. It is to be supposed that this community expands considerably during the flood season,
although as yet there are no recorded observations to support this assumption.
VERTEBRATES OTHER THAN FISH
AMPHIBIA
Both larval and adult stages of various species of frogs and toads are plentiful on floodplains. Most
fringing areas of the plain and all permanent and temporary swamps are colonized and in the most
deoxygenated areas, tadpoles are often the only form of vertebrate life. Similarly amphibians rapidly
appear in the most isolated of temporary pools. Little work appears to have been done on the ecology
and dynamics of amphibian populations of rivers although they undoubtedly form an important
component of the fauna of the floodplain, and probably contribute to the rapid recycling of detritus
and mud by converting it into flesh which is useable by predatory species of fish, reptiles and birds.
Frogs are exploited commercially for their legs particularly in China and Romania. However the
resources have been either over-exploited, or the stocks have been reduced by pollution in some areas,
for instance France, where they are mow protected.
REPTILES
Several families of reptiles have remained associated closely with water, and of these, four groups
(Crocodiles, monitors, iguanas and turtles) are regularly found in and around rivers and the permanent
and temporary standing waters of their floodplains. Certain species of snake too have adopted aquatic
or semi-aquatic habits. Various forms of crocodile are distributed around the tropical world, and have
been widely associated with flood rivers in the past. They have, however, been subject to widespread
slaughter and are now virtually absent from many parts of their former range. The economic
importance of the crocodile as a consumer of fish has been discussed, principally by Cott (1961).
Although crocodiles eat a large quantity of fish, they also prey on other organisms which themselves
are predators of fish. Their status as major competitors of man for fish is therefore somewhat obscure.
A second function of crocodilia in the ecology of some types of tropical water has been suggested by
Fittkau (1970 and 1973). In the nutrient-poor rain forest the larger elements of the community serve as
nutrient sinks, which slowly accumulate the few minerals available (Fittkau and Klinge, 1973). Such a
system depends much on the abundance of its species which are able to maximize the storage and
recycling of nutrients of allochthonous origin. A similar process is thought to occur on the equally
nutrient-poor blackwaters of the rainforest zones. In such aquatic systems the low level of nutrients
does not permit much primary production and the food chain originates mostly in the rain of
allochthonous material. Fish moving into the mouth lakes are allochthonous to that particular
ecosystem and are fed upon by a variety of large predators, of which caimans are perhaps the most
significant. A medium-sized caiman can eat between 0.6 and 0.8 percent of its body weight per day
and excrete about 0.20 to 0.27 percent of its body weight of nitrogen, phosphorus, calcium,
magnesium, sodium and potassium ions per day. Their contribution to the nutrient balance of such

116
environments has been estimated by Fittkau to be locally superior and complementary to the nutrients
derived from rain water, itself the major source of nutrients in the system as a whole. In places where
crocodiles have been eliminated, declines in fish production have been noted, possibly because of a
drop in the primary production based on the excreted nutrients.
BIRDS
Birds are a very conspicuous feature of the floodplain ecosystem. As with most wetlands various
kinds of waterfowl are extremely common, but it is doubtful whether most of these react directly with
fish populations. Piscivorous predators are also abundant, however, and represent possibly the greatest
source of pressure from outside the aquatic system. Reizer (1974) for instance listed 37 species of
ichthyopredator in the Senegal River and 19 species are listed by Shepherd (1976) from the Shire
River. Birds are capable of taking a wide range of species of all sizes and the piscivorous bird
community appears to be specialized toward the taking of particular sizes and types of prey.
In general, the life cycle of water birds is closely linked to the floods. Avian breeding seasons
coincide with those of the fish and the fledglings are being reared at just that time when small fish
suitable for their feeding are most abundant. There is a particular heavy predation during the period of
receding water when many fish are stranded in temporary pools (Lowe-McConnell, 1964; Bonetto,
1975).
The impact of birds on the fish population is potentially very large, and studies from Africa indicate
that the amount of fish taken by them can surpass the amount taken by the fishery. In the Senegal
River, for instance, Reizer (1974) presented figures by Morel of between 100–200 000 herons and
cormorants and 2 000 pelicans in the delta alone. Fish consumption was estimated at between 500–1
000 gm per day for the darter Anhinga rufa and for a heron, 1 000–2 000 gm per day for a pelican and
250 gm for the kingfisher, Ceryle rudis. On the basis of this birds take about 70 000 t/yr of fish as
compared to a fish catch of about 50 000 t/yr.
Bowmaker (1963) gave an estimate for the daily ration of the cormorant Phalacrocorax africanus of
78 gm/day leading to a consumption of 286 t/yr from Lake Bangweulu. This estimate may be low by a
factor of 10 in view of Reizer data and that of McIntosh (1978) who found that the European
cormorant (Phalacrocorax carbo L.) eats some 650 to 700 gms of fish/day. Bowmaker concluded that
the African cormorant is not harmful to the fishery, but could in fact be beneficial by its use of non-
commercial species, and by its excreta which fertilize the water. Other figures from cold water
streams indicate that kingfishers eat about 14 kg of fish/km, of stream terns consume about 63 kg of
fish/km of river and Mergansers take some 25 kg/km. These are equivalent to 2.3%, 14.1% and 4.2%
of total mortality respectively (Alexander, 1979).
MAMMALS
Several aquatic or semi-aquatic mammals figure in the ecology of rivers although their role is by no
means always clear. Beavers, which are confined to the north temperate zone, have had an enormous
influence on the aquatic system. Their habit of building dams resulted in the more or less permanent
inundation of large areas of floodplain, creating widespread swampy conditions in the valley bottoms.
The dams themselves affected flow characteristics of streams, and the organic debris increase habitat
diversity and cover for fish. Beaver populations have been drastically reduced in parts of Europe and
North America and their dams have been cleared from rivers in an attempt to improve drainage. This
process, together with the clearance of dead wood, stumps and other naturally occurring obstructions
from the stream bed, has produced much dryer conditions in the valley bottoms and has effectively
removed considerable areas of floodplain from lower order temperate streams. Otters are widespread
throughout temperate and tropical systems. They apparently prey heavily on fish, although population
densities are far from high so the impact of such predation is, in most cases, probably relatively low.
They, do, however, have a bad reputation with fishermen for their habit of robbing and damaging set

117
gear such as traps or gill nets. One record by Alexander (1979) shows otters to take about 13 kg of
trout, km of cold water stream against 25 kg/km taken by mink. These figures were equivalent to
3.0% and 5.6% of total mortality respectively.
In African rivers hippopotami contribute a large amount of fertilizer to the aquatic system by cropping
terrestrial vegetation and excreting it into the water in a form readily accessible to phytoplankton.
Densities of hippopotamus were at one time very high in most African inland waters and they must
have made a considerable contribution to nutrient inputs and recycling. They have, however, been
eliminated through much of the continent and their numbers have been considerably reduced even
within the confines of protected zones.
In Latin America and elsewhere, members of the family Sirenidae appear to play a similar role in
nutrient balances particularly in the black water rivers and mouthlakes of the Amazon. The dugong
(Dugong dugon) is distributed throughout the Indo-Pacific region (Husar, 1975). The African manatee
Trichechus senegalensis is distributed throughout the rivers of West Africa and is present in
considerable number in Cameroon, Gabon and in the Niger River System, (Nishiwaki et al., 1982).
Trichechus manatus is distributed from Florida to North Eastern Brazil (Maranhao) and Trichechus
inguinis is found throughout the Amazon River system. Studies on the feeding of Sirenidae (Best,
1981) show manatees to consume about 8% of their total body weight in aquatic plants daily whereas
the dugong eats about 14%. Their selected foods are higher plants not normally eaten by other
organisms and they are thus not in active trophic competition with other elements of the community.
Many large river systems have species of dolphin of the family Platanistidae. Platanista gangetica is
found in the Ganges, Brahmaputra and other rivers of the Ganges basin. Platanista minor inhabits the
Indus River and enters the Zambezi, Lipotes vexellifer occurs in the Yangtse, Inia geoffrensis is
distributed throughout the Amazon system together with Sotalia fluviatilis, the only freshwater
Delpinid, which is also found in the Orinoco River. Pontoporia blainvillei inhabits the lower reaches
of the Parana and Uruguay rivers, the estuary of the Rio de la Plata and the adjacent coasted waters.
Orcaella breviostris, which primarily inhabits shallow coastal waters of the S.E. Asian region, enters
many including the Irrawaddy, Ganges, Brahmaputra and Mekong as well as minor water bodies of
the Indonensian archipelago (Watson, 1981). All these dolphins feed on fish and Crustacea using their
long beaks to root in the bottom. In the Amazon, the two coexisting species are separated by very
different ecological characteristics (Ferreira, 1983) Sotalia is a social dolphin inhabiting the open
waters of the river channel and feeding on small sized schooling fish eaten whole. It consumes about
6.6% of its body weight daily. Inia is a more solitary animal that depends on large solitary bottom
dwelling fish and which are ripped apart before being consumed. It consumes 3.1% of its body weight
daily. There is some overlap of diet at the edges of the channel. Inia feeds mainly during rising river
levels as fish spread out onto the floodplain and has been recorded many kilometres from the river in
the Igapo. Sotalia feeds principally during low waters as pelagic schooling fish are concentrated into
the river. Extrapolating from what is known of the ecological role of other freshwater aquatic
mammals, dolphins may play two major roles within the ecosystem. Firstly, they may contribute to
the maintenance of the nutrient balance especially in nutrient poor rivers in much the same way as that
described for the Amazonian caimans. Secondly, they may consume significant amounts of fish thus
affecting the dynamic of the populations within the river. Little information is available at present
upon which to base any conclusions as to the role of these species, although indications from Ferreira
(1983) are that large quantities of fish are consumed seasonally. This may be especially significant for
fisheries in the case of Inia, which, by taking large and commercially preferred species, is in direct
competition with fisheries. The smaller schooling fishes preyed upon by Sotalia have are not yet
formed the subject of a fishery and are thus of little or no direct economic important. Despite this
competition Inia, in common with most dolphins in freshwaters, is strongly protected by tradition, and
is neither harmed nor eaten.
The floodplain is also used during the dry phase by a number of wild and domestic animals, but these
will be considered separately in a later chapter of this review.

118
CHAPTER 5
RIVER FISHES AND THE RIVERINE SYSTEM
NUMBERS OF SPECIES IN RIVER SYSTEMS
DIFFERENCES IN NUMBERS OF SPECIES BETWEEN SYSTEMS
The considerable differences in the numbers of species inhabiting the various river systems are largely
attributable to the size of the river as represented by its basin area or some correlate of it such as
length of main channel or stream order. A relationship of the form N = fAb (where N = Number of
species and A = basin area in km2) can be fitted to the dispersion in Fig. 5.1. When all points are
included a regression, N = 0.297 A0.477 is obtained. In fact there are differences between different
geographical areas which become apparent when individual blocks of data are analysed separately as
follows:

119
Figure 5.1 Number of species of fish present in different river systems plotted according to their
basin areas: (•) South America; (o) Africa; (▭) Asia; (x) Europe; ( ) North America
North flowing Siberian and European Soviet Rivers:
N = 2.76A0.19 (N = 6, r = 0.91)
South flowing European rivers:
N = 0.6A0.14 (n = 11, r = 0.72)
Africa:
N = 0.449A0.434 (n = 25; r = 0.91)

120
South America:
N = 0.169A0.552 (n = 11; r = 0.95)
Daget and Economidis (1975) have independently fitted similar log-log regressions to date from sets
of small rivers in Portugal and Greece obtaining:
Greece:
N = 2.319A0.24 (n = 12; r = 0.94)
Portugal:
N = 1.786A0.19 (n = 12; r = 0.92)
which agree well with the regression established for Europe. Similar scatter plots for Australian rivers
also conform to these general principles with the distribution for nine North flowing tropical rivers
being located above the world mean regression line and five of the six points for temperate rivers
being located below the line. (Bishop and Forbes, in press). These relationships enable us to conclude
that while species diversity increases with basin area at all latitudes, it does so faster as one
approaches the tropics - as indicated by the larger exponent at lower latitudes. Because a log-log
relationship does not completely describe the distribution of points, the intercept, which Daget uses as
a relative index of faunal richness, is probably not a reliable predictor for basins of extremely small
area. Many reasons have been advanced for the increased diversity at low latitudes and these are
discussed more fully by Lowe-McConnell (1975). As to the influence of size, the number of
ecological niches is probably greater in larger river systems than in small ones. Meandering creates a
regular series of habitats in the main course of the river and some floodplain lakes are often isolated
from the rest of the system for long periods of time. Furthermore, similar habitats in the sub-systems
are often separated by considerable distances of inimicable biotope, leading to the formation of
distinct groups of species adapted to similar conditions along the length of the river. Roberts (1973)
has described the influence of such factors on the faunas of the two largest rivers of the world, the
Amazon and the Congo. However, such differences do not occur solely in the larger systems for they
have also been noted in small rivers and streams of sixth order or lower by Kuehne (1962); Barrel,
Davis and Dorris (1967), Whiteside and McNatt (1972), and Platts (1979) from the United States, and
Bishop (1973) from the Gombak river in Malaysia. Gorman and Karr (1978) also found that fish
community diversity is correlated with stream habitat diversity in selected Indiana and Panama
streams, and Horwitz (1978) demonstrated similar relationships in various types of rivers in Illinois,
Missouri, Ohio and Wyoming. Furthermore, diversity of habitat increases with increasing stream
order.
The aquatic components of river basins resemble island fauna in their relative isolation one from
another. The mathematical relationship between the size of islands and the number and diversity of
species inhabiting them has been examined by McArthur and Wilson (1967), who concluded that the
equilibrium between the extinction of species and the colonization with new species differs according
to the size of system; larger systems favour higher diversity perhaps because of the type of
biogeographical factors mentioned above.
Although total numbers in large systems tend to be high, groups of species are often located in very
different portions of the system, thus only a certain percentage concerns the floodplain or its fishery at
any one point. Some species also are limited to the turbulent waters of rapids and headwater
tributaries and are rarely, if ever, found in the slower flowing floodplain reaches. Nevertheless
individual fisheries in larger rivers do have many more species available to them than those of smaller
streams and catches consisting of over fifty species in one type of gear are not uncommon in the

121
biggest systems. Figure 5.2 illustrates the diversity in form of representative species of various genera
of tropical river systems.

122

123

124
Figure 5.2 Representatives of genera of fish from tropical river systems

125
RELATIVE ABUNDANCE OF SPECIES WITHIN ONE SYSTEM
In his review of data covering a number of areas and taxa, Preston (1962 and 1962a) found that within
a particular taxon and region, the relative abundance of species followed a lognormal distribution (i.e.,
the logarithms of the species' abundance were normally distributed). These distributions were of
course truncated at very high and very low levels of abundance, but over the observed range
distribution of this type described most of the sets of data well. In theory there are an infinite number
of lognormal distributions that could describe the relative abundance of a given number of species,
but Preston found a very close empirical relationship between the number of species in an assemblage
and the precise form (i.e., the variance) of the observed lognormal distribution of relative abundance.
He proposed that distributions obeying this relationship be called “canonical distribution”. Daget
(1966) concluded from two samples from the flooded plain of the Benue River, that the distribution of
numerical abundance and rarity of fishes in the sample locality did indeed conform to the canonical
type. A similar conclusion was reached by Loubens (1970) on the basis of 15 samples from various
parts of the Lake Chad-Chari river complex. Here samples were distributed log normally, both when
analysed by number and by weight, although in the latter case the correlations were less exact. With
smaller sample areas a simpler exponential relationship between ranked species (R) and number (N)
of the form N = abR was equally applicable. The extension of this type of analysis to other series of
data from other water bodies throughout the world demonstrates the wider application of these
principles, both to areas sampled by complete fishing and to samples obtained with individual types of
gear. Preston (1969) expanded the idea of relative abundance of taxa in one place at one time to
suggest that commonness and rarity are also distributed log normally in space and in time.
These conclusions are of interest when considering the community dynamics of floodplain
ecosystems insofar as they affect the dominance of species especially favoured by the fishery. They
also imply that the fish catch will tend to be dominated by only a few species even in communities of
high specific diversity and abundance. For this reason changes in population structure under
exploitation deserve further investigation. Particularly interesting is Preston's contention that the
distribution of commonness in any sample of a population is a truncated lognormal, having the same
modal height and logarithmic distribution as the “universe” from which it is drawn. The abundance
structure of fish isolated within floodplain depressions and lagoons may thus be linked by similar
relationships to the abundance structure of the fish community in the river as a whole.
SUB-POPULATIONS
There is very little information on the fine structure of species distribution of fishes inhabiting rivers.
From the behaviour of freshwater fish elsewhere it might be expected that any one species is far from
homogeneous and the sub-populations or stocks might be set up within the system. In the Volga River
the existence of such populations was indicated by the continued segregation of a species (Abramis
brama) into a number of local stocks after impoundment to form a reservoir of the riverine type. The
individual stocks (see Fig. 5.3) maintained distinct, isolated reproductive areas and areas for feeding
and over wintering. The feeding sites were separated from the breeding sites by considerable distances
(Poddubnyi in Mordukhai-Boltovskoi (1979). In rivers such as the Niger, Congo or Zambezi where
floodplain reaches alternate with stretches of rapids, conditions seem particularly suitable for the type
of genetic isolation which leads to the setting up of separate breeding populations.
In the Mekong, Sao-Leang and Dom Saveun (1955) noted that populations north of Khone falls
migrate downstream of them, whereas fish from the southern reaches often move upstream of the
falls. Blackfish species, particularly those whose longitudinal migrations are minimal, seem likely
candidates for the evolution of sub-populations along the length of the river. The more mobile
whitefish, however, have much more opportunity for dispersal and mixing of genetic strains, but even
here there are some indications that sub-populations are formed. There are hints that some species
have a homing instinct. Bonetto found tagged individuals of Prochilodus platensis in the same
floodplain pool in successive years in the Parana River, as did Holden (1963) with individuals of

126
Lates niloticus and Hydrocynus lineatus in the Sokoto. This, of course, may mean simply that the fish
have not moved at all in the intervening flood period, but it does imply a certain degree of
territoriality in the species concerned. Furthermore, Godoy (1959 and 1975) removed tagged
individuals of Prochilodus scrofa from one site on the Mogi Guassu to place several hundred
kilometres away and even located on different branches of the same river system. Some fish returned
rapidly to the site from which they were captured originally, indicating the type of homing ability
which is often associated with distinct stocks of migratory species.
Figure 5.3 Feeding and breeding locations of different stocks of bream (Abramis brama) in a river and
riverine reservoir (Volga system). (After Poddubnyi, 1979)
The existence of separate sub-populations of species in river systems has also been proposed on other
grounds. According to Durand and Loubens (1969), Alestes baremoze has two populations, one of
which is migratory within the Chari and Logone rivers, the other resident in Lake Chad. A similar
separation has been attributed to Basilichthys bonariensis on ecological evidence. Here there is an
estuarine population in the Rio de la Plata, and a riverine population in the Parana. The riverine
population can also be distinguished by its faster and more consistent growth rate (Cabrera, 1962).
From the results of their tagging experiments, Bonetto and Pignalberi (1964) suspected the existence
of an upstream and downstream population of Prochilodus platensis. Prochilodus platensis also has
two body forms, ‘longilineas’ and ‘brevilineas’ which may depend on the early nutritional history of
the individual fish (Vidal, 1967). Studies on a more restricted area of the La Plata estuary have also
shown this species to have sub-populations with different growth characteristics (Cabrera and Candia,
1964), whereas Parapimelodus valenciennesi, a blackfish species from the same region, only has a
single homogeneous population (Cabrera et al., 1973; Candia et al., 1973). Bayley (pers.com.) has
also collected meristic evidence of the existence of several distinct sub-populations of Prochilodus
insignis in the Amazon. Separate sub-populations of migratory fish have also been identified with
differing life-history strategies adapted to the range of physical and climatic conditions in the streams
in which the fish spawn. The anadromous Alosa sapidissima studied by Legget and Carscadden
(1978), for example, shows a gradation in fecundity and migratory distance in sub-populations
associated with East Coast American rivers. These authors' arguments can probably be extended to
other species and may apply to other systems which are sufficiently geographically or spatially
diverse as to have a range of climatic conditions over their length.

127
There are indications that many riverine species exist in two behavioural phases which may
correspond to sub-populations. One phase is migratory showing characteristic separation of breeding
and feeding grounds. The other is static and is more inclined to form territories. Such behaviour has
been described for Hilsa ilisha (Pillay and Rosa, 1963) in India and for Gobio gobio and Rutilus
rutilus in Europe (Stott, 1967). The existence of such alternative forms would explain the ease with
which some species adapt to changes in flow within rivers and readily adopt more sedentary habits
after flood control or impoundment.
There appears to be good circumstantial evidence that geographic sub-populations exist in rivers and
there is support for this from other inland waters such as the Great Lakes of N. America (Loftus,
1976). However, there are only few cases where such subpopulations have been positively identified.
Whether or not a population of fish is disassociated into separate stocks at a sub-population level is of
great significance to the management and conservation of fisheries for that species. Where separate
subpopulations exist, local depletion of the fish fauna by overfishing or by other environmental
pressures is less likely to be compensated for by recolonization from larger populations
SIZE OF SPECIES
The species of fish inhabiting floodplain rivers cover a wide range of size as is illustrated by the
maximum recorded lengths of the individual species from communities inhabiting three typical rivers
in Fig. 5.4 in which the histograms represent the number of species classed according to their total
lengths. Sizes normally span about 3 orders of magnitude (i.e., from about 1.5 cm to 1 500 cm). There
tends to be a high proportion of fish of very small adult size (less than 10 cm) in rivers both in feeder
streams or rocky headwaters and in the potamon reaches of the river. Small size is advantageous both
in the riffles of the rhithron and in most floodplains, as pygmy species can mature more rapidly (often
within one year) and can seek refuge in the root masses of vegetation in the interstices of pebble
masses and other small crevices. Equally they can colonize the surface area of the water and more
readily exploit the neuston or allochthonous food sources found there. Most river systems have a few
species of truly gigantic size. In Latin America the characteristic giant species are Arapaima gigas,
Pseudoplatystoma spp. and Brachyplatystoma spp., in Africa, Lates niloticus and in the Mekong
Pangasianodon gigas, although these are by no means the only fish attaining lengths of 1.5 m or
more.

128
Figure 5.4 Histograms showing the proportion by number of species of different maximum lengths in
three typical tropical river systems
DISTRIBUTION OF SPECIES IN RIVER SYSTEMS
DISTRIBUTION IN SPACE AND ZONATION
Zonation
Many attempts have been made at establishing general principles regulating the zonal distribution of
living organisms in rivers. Illies and Botosaneanu (1963) summarized the schemes proposed by a
number of authors and themselves suggested a separation of the river into creon, rhithron and
potamon zones based on faunistic criteria: A specific rule describing the distribution of Northern
European species was advanced by Huet (1949) on the basis of the slope and width of any particular
reach of the river (Fig. 5.5). More recently Vannote et al. (1980) suggested the river continuum
concept which posits an orderly downstream progression of organisms as discussed in Chapter 3.
These attempts mostly originated in the North Temperate Zone and pre-suppose an idealized river
form with a continuously decreasing gradient from source to mouth. As such they are useful within
areas where the original climatic or morphological conditions prevail, but are transposed to the tropics

129
only with difficulty. The temperate pattern of zonation has been demonstrated by Bishop (1973) for
the Gombak river (Malaysia) and in other Malaysia rivers the cyprinids, at least, show a clear
succession from Tor tambroides and Acrossocheilis hexagonolepis in upper rapids reaches,
Leptobarbus hoeveni, Puntius bulu and P. daruphani in intermediate streams and P. jullieni only in
the lower potamonic reaches (Tan, 1980). Watson and Balon (1984a) also found highly structured
communities to exist in the Baram River of North Borneo. Here fish partitioned first into layers in the
water column, surface, pelagic, benthic and substratum niche types. A tendency was noted for there to
be an increase in the number of species inhabiting the bottom zone with increasing distance
downstream. Furthermore a related reduction in niche size with the increasing numbers of species was
indicated by a decrease in mean life span and adult size of the individual species.
Figure 5.5 Graphical representation of the relationship between slope of river bed and channel width
and fish community in European rivers. (After Huet, 1949)
In Africa the Illies-Botosaneanu classification has only been applied successfully in some South
African rivers (Harrison, 1965) or in the Luanza, a high altitude tributary of the Congo River system
(Malaisse, 1976) where there is a succession of rhithronic and potamonic reaches similar to those in
Europe. In most parts of the continent zonation is much more obscure as is shown by examples from
West Africa (Ogun river, Sydenham (1978); Bandama river, Merona (1981); Ebo Stream, Ghana,
Lelek (1968)) or from Central Africa (Kalomo river, Balon and Coche (1974) and the Luongo river,
Balon and Stewart (1983)).
Provided there are no geographical discontinuities such as large waterfalls, the increase in diversity
along many rivers tends to be by addition of species to those present rather than by the replacement of
species. This is particularly true of the tropics although Horwitz (1978) remarks on this aspect of the
increase in diversity of fish faunas in streams of the mid-Western United States. In Africa and parts of
Asia most large river systems have such discontinuities so that there are sufficient important
exceptions for this not to constitute a general principle. For example, the Zambezi River is divided

130
into three faunistic zones, the middle of which between the Cahora Bassa rapids and the Victoria Falls
is the poorest, or in the Nile, the Kabalega Falls between Lake Kyoga and Lake Mobutu, marks a
discontinuity between the generalized nilotic fishes downstream and a distinct riverine and lacustrine
fauna upstream. Instead it is possible from the work that has been done, to separate the causes of
species distribution in water courses under two main headings: geographical factors and
geomorphological factors.
Geographical factors influence the taxonomic differences that may be encountered within a river
basin. For instance, the isolation of sub-populations of species in small order streams can result in
specific divergence over a long time period. In this way it is possible to observe two similar species
occupying identical or similar niches in two separate streams of the same basin. This isolation is to a
certain extent reinforced by the behaviour of the species themselves, which are often small, sedentary
and have a short life cycle, which may contribute to a rapid rate of speciation. This tendency for
species to diverge in the lower order streams of a river basin could account for much of the increasing
diversity of species shown by larger river basins in Fig. 5.1 and for the relatively large proportion of
species of small size in river fish communities. A second mechanism influencing distribution is river
capture when species may be exchanged between systems. Mahon (1984) discusses some of these
mechanisms in connection with the differences between fish faunas of similar river systems in Europe
and N. America. In North America the more normal divergence has occurred with occupation of head
water streams by numerous small cyprinid and percid species. In Europe the rivers are still occupied
by larger cyprinid species which inhabit the potamon and only migrate to the rhithron to spawn. The
headwater streams there tend to be used mainly by the young cyprinids. This difference is attributed
mainly to the affect of glaciation.
Whereas geographical considerations may influence the distribution of fish species between river
systems, the distribution of the various types of species within any one system is more likely to be
controlled by the geomorphology of any particular river reach. Thus, it is useful to distinguish two
major fish communities:
(i) communities of rhithron-like or rapids zones; and
(ii) communities of the potamon,
Certain amount of interchange between these two communities may occur, particularly by certain
elements of the potamon fauna which enter rapids to breed. Rhithronic fishes tend to be relatively
static within their preferred zone, although juveniles of migratory potamonic species or even marine
anadromous forms, such as the salmon, form temporary elements of their communities. Potamonic
fishes can, however, be divided into two fairly distant groups on the basis of their behaviour.
(a) The first group of fishes avoid severe conditions on the floodplain by migration to the main river
channel and frequently by more extensive movements in the river beyond the floodplain area.
Members of this group are recognized a “Piracema” or “Subienda” species in Latin America, in the
Mekong system are termed “white” fishes, and in other river systems may be classified as rheophilic.
Species of Cyprinidae in Asia, Africa and Europe and Characoidei in Africa and South America are
conspicuously members of this group, sometimes undertaking very spectacular migrations. Some
siluroids and mormyrids are also migratory in behaviour. A few species are confined to the river
channel at all times and never penetrate the plain.
(b) The second group of fish consists of species which have considerable resistance to deoxygenated
conditions and which are termed “black” fish in Southeast Asia or limnophylic elsewhere. Their
movements are therefore more limited than those of the “white” fish. They frequently remain in the
standing waters of floodplain during the dry period and if they move to the river they remain within
the vegetated fringes or in the pools of the river bed as it dries. Most siluroids belong to this category
together with ophiocephalids (channids) anabantids, osteoglossids, polypterids and lung fishes.

131
Although it is very rough, the distinction between blackfish and whitefish is very useful as a first level
ecological classification of species and will be retained in this book for this level of discussion. The
response of the two groups to varying conditions on the floodplain is very different and has important
implications for the management of the ecosystem and the fish stock.
Because many tropical rivers pass through several successive alternations between calm and rapids
reaches, these two faunas likewise alternate along the length of the stream, which gives a type of
zonation based purely on the flow or bottom characteristics peculiar to the river in which they are
found as, for instance, in the Luanza River (Balon and Stewart, 1983). Species of several families
have become adapted to life in rapids and as Poll (1959; 1959a) points out for the Congo River, the
inhabitants of downstream rapids are generally representatives of families which are normally found
in the potamon, but which have occupied the rapids, rather than representatives of those families, such
as the Kneriidae or Amphiliidae which are native to the rapids of the headwater streams. Asian and
South American rivers tend to be of a more traditional form with steep mountainous headwater
streams giving way to flat lower courses which are sometimes of extreme length. In such rivers the
discontinuity between the rheophilic fishes of the headwater streams and the communities of the lentic
potamon downstream is apt to be particularly abrupt.
Conditions become increasingly estuarine toward the mouth of the river where saline waters penetrate
many kilometres upstream, particularly in lowland rivers of slight slope. A pronounced zonation of
species occurs according to their salinity tolerance, although the zones move up and down river as the
interface between saline and freshwater varies with the state of the tide and the flood regime of the
river. Three groups of fish are to be found in this transitional zone.
(i) Freshwater stenohaline species which enter the zone during the flood and retreat upstream at
low water according to the penetration of the saline tongue.
(ii) Marine stenohaline species which follow the influx of marine waters into the river for feeding.
(iii) Euryhaline species which move little but which adapt to the changing salinities of the water.
These may be of freshwater origin (e.g., members of Cichlidae, Cyprinodontidae and some
Siluroidae) or of marine origin (e.g., members of Clupeidae, Atherinidae, Mugilidae, Lutjanidae,
Sciaenidae, Ariidae, Pomadasydae, Gerridae, Carangidae, Centropomidae, Eleotridae and
Gobiidae).
Several species migrate between the river system and the sea either for breeding or feeding.
Anadromous forms, where the breeding cycle is completed in freshwater, include several estuarine
species of marine origin, the Eleotrids Batanga lebretonis, for example, which undertake limited
migrations upstream, as well as coastal marine species such as the clupeids which sometimes migrate
over long distances in the river. In the temperate flood rivers of Southern Europe the sturgeons
(acipenseridae) and salmonids are the main anadromous fishes. Truly catadromous species are
somewhat rarer in large tropical rivers, although eels are present in some systems, notably Anguilla
nebulosa in the Zambezi and its tributaries. Many normally marine species enter the lower reaches of
rivers to feed during the dry season and return to the sea during the rains.
Habitats of River Systems and the Accompanying Floodplains
A number of areas can be considered distinct habitats from differences in morphology, chemical and
physical conditions, presence or absence of vegetation, structured objects such as rocks, wood or other
cover and abundance and type of food. Table 5.1 based on such ecological characteristics, lists the
major habitats found in river systems.
Table 5.1

132
Major habitats of river systems
RIVER CHANNELS FLOODPLAIN 1. Main channels: rapid and turbulent 1. Flooded grassland
flow; fairly uniform; floating sudd
islands. A. Floating meadows - these are prob-
ably not uniform as there are slight dif-
2. Tributary streams ferences in bottom substrate and relief,
floral associations are variable.
A. Small rocky torrential streams
descending from unflooded terrace, or B. Open water.
upstream of main floodplain area.
C. Littoral fringes area at limit of
B. Small channels linking floodplain to advancing or retreating water, submerged
subsidiary marsh or lake areas above main grass; often low D.O. in sheltered areas;
floodplain level - Terra Firme lakes of higher D.O. in turbulent wave-washed
the Amazonas type or type 1 lakes areas.
(Svensson, 1933).
2. Lagoons and depressions
A. Open water
(i) mud bottom
(ii) sand bottom
B. Standing vegetation
C. Floating vegetation mats
D. Floating leafed vegetation
E. Submersed vegetation
3. Lakes (as above but with a greater
proportion of open water and deeper)
4. Flooded forest
A. Dense rain forest
B. Gallery or levee woodland
C. Acacia and bush scrub
5. Flood areas outside main flood area
Terra Firme lakes of the Amazonas or type 1 lakes
(Svensspn, 1933)
1. Rhithronic zone (may break up into an alternation of
pools and rocky riffles) 1. Floodplain pools
A. Pools which dry out completely
A. Pools (in extreme form pools become isolated and
deoxygenated)
B. Marshy pools (heavily vegetated with little
dissolved oxygen)
(i) mud bottom
(ii) sand bottom (i) surface film
(iii) leaf litter - forested or open (ii) deeper water
α floating vegetation fringe C. Shaded pools (in forested or wooded
β submersed vegetation areas)
γ emergent vegetation
(i) clear
B. Rock riffles (a variety of habitats under rocks or
on surface of rocks) (ii) with tree trunks and other debris
C. Tree trunks and other debris 2. Lagoons
2. Potamonic zone (meanders produce a A. Deeper open waters
regular succession of habitats of varying
depth and bottom type) (i) mud bottom
(ii) sand bottom
A. Shallows
B. Vegetated fringes
(i) mud bottom with no current
(ii) sand bottom or with slight α floating mats
(iii) leaf litter current β submersed vegetation
γ emergent vegetation

133
B. Deeps with slow or faster current-shaded by forest
or open
α floating vegetation 3. Large lakes (sub-habitats as for lagoons but more
inclined to set up permanent stratification, greater
depth, more open water relative to shoreline, often with
sheltered and exposed shores)
β emergent vegetation
BACKWATERS CONNECTED TO MAIN
CHANNEL DOWNSTREAM LARGER WATER BODIES
Lentic water regions, open to main channel but with
many of the characteristics of lagoons or lakes above,
may be:
Lakes, sea or larger river system.
A. Shaded - clear or with tree trunks and debris.
B. Open with
(a) deep water:
(i) mud bottom
(ii) sand bottom
(b) shallow water with:
α floating vegetation mats
β submersed vegetation
γ standing vegetation
δ floating leafed vegetation
(c) shallow littoral usually vegetated.
The main channel
Fish in the rhithronic reaches are distributed either among the riffles or in the deeper pools. Although
the riffles present a reasonably homogeneous environment they are structurally complex and require
specific adaptations either to resist the current and the turbulent conditions found on the surface or to
live in the crevices and holes in the rocks. Large, swift swimming, streamlined species sometimes
penetrate those turbulent waters but either pass through in an upstream migration or drop back to the
shelter of the pools. Riffles are also favoured environments for deposition of eggs of some species and
for the subsequent development of their larvae and fry. The adaptations to swift current also fit fish to
exploit the available food sources in the riffles. Swift swimming forms pick up allochthonous material
falling on the surface of the water or drift organisms. Fishes with adaptations to cling to the surface of
the stones often have sucker like mouths with which to feed on epilithic or epiphytic organisms
whereas the diminutive or elongated species that inhabit the interstices in the rocks are particularly
well situated to prey upon the numerous insect larvae crustacea that inhabit the bottom.
Pool habitats are much more varied and fish tend to segregate both by depth and by distance from the
shore. Three communities can be readily distinguished within the water column. A pelagic community
which tends to consist of small species, silvery in colour with upward facing mouths, a mid-water
community of larger silvery fishes streamlined with terminal mouths and a bottom living community
of drab coloured fishes with dorsally humped profiles and ventrally positioned mouths. These various
forms are well illustrated by the communities of cyprinids in a small Borneo stream as described by
Inger and Chin (1962) Fig. 5.6. The slacks near the bank are often heavily vegetated with floating and
emergent aquatic and ever-changing terrestrial plants. Here bottoms are usually composed of food
rich detritus from leaf fall and from silt deposition in the quieter waters.
The slack areas at the fringes of pools, the limited floodplains of the rapids reaches and perennial
streams of the lowland zone have similar faunas many of the elements of which recur within the
floodplains of the potamon. Because there are a variety of microhabitats in the slacks small size is a
great advantage. Spatial niche selection seems very highly developed and all levels of the water
column are occupied as in the example of the Borneo streams above. In a tropical African forest

134
stream several genera of cyprinodonts occupy the water surface, particularly Epiplatys and
Aphyosemion although some Characins such as Brycinus macrolepidotus are also found there. The
mid-waters support clouds of dwarf Barbus and Aphyosemion species which are more often than not
associated with the shade of floating lily leaves. The rich detritus of the bottom attracts several species
of small mormyridae, Neolebias, Barbus and Labeo together with various small or dwarf cichlids such
as Pelvicachromis, Thysia ansorgii, Hemichronis bimaculatus. Similar elongated fishes are found in
the rocks also occur in the floating vegetation at the fringes of the pools as the sinuous habit is equally
adapted to such conditions. Mastacembelus and Calamoichthys particularly are conspicuous in such
environments as they also frequent similar vegetation in the potamon.
Semi-permanent channels, which are more characteristic of low order streams, frequently become
broken-up into a series of rocky riffles, and pools which may become deoxygenated or totally anoxic
as the season advances. Very great densities of fish may be found in the pools, which essentially
represent the sump into which most of the population drains. Lowe-McConnell (1964), for instance,
found 870 fish belonging to 36 species in a pool of 19 m³. This she attributed to a three dimensional
use of space within the pool, together with an alternation of activity between two very different
nocturnal and diurnal fish faunas. The species composition of main river pools is rarely the same as
that of floodplain pools, and within the pools segregation appears to occur by bottom type. Lowe-
McDonnell recorded 44 species of fish in the pools of the Rupununi river, of which 37 (84%) were
found over only one type of substrate.
During the floods the main river channel is filled with rapidly flowing water, but at its margin the
floating vegetation fringe merges with the flooded banks of the plain itself. Large masses of
vegetation become detached and are swept downstream as floating islands. As a consequence the
channel is usually comparatively sparsely populated at this time, although some species, for instance
Petrocephalus bane or Hydrocynus forskahlii, never move out of the river on to the plain (Daget,
1954). In Latin American rivers, the upstream migration, “Piracema” or “Subienda”, of the major
characins during low water and the beginning of the flood also ensures that some species are present
in the channel during at least the earlier phases of rising water. The fringing vegetation and floating
sudd islands shelter juvenile fish or small vegetation dwelling species, which may become distributed
throughout the system in this way.

135
Figure 5.6 Segregation by depth of cyprinid species in a Borneo stream. (From data in Inger and Chin,
1962)
In the Potamon most species of fish move to the main channel during the dry season and settle in
different habitats along its length. Some dry season habitats can lie a considerable distance away from
the main floodplain, and may even be located in entirely different aquatic systems such as the sea or
major freshwater lakes.
In the permanent channels during the dry season, fish also separate by depth, type of bottom and
vegetation cover. Many smaller species inhabit the root masses of the floating vegetation at the edge
of the river, finding there both abundant food and shelter. Several species have specific adaptations to
this habitat. The “upside-down” swimming position of some Synodontis species enables them to
browse on the root fauna, and the serpentine shape of Mastacembelus or Calamoichthys enables them
to weave among the entangled stems of the plants.

136
Deeper areas of the main channel attract the larger fish species. Pangasius sutchi, for example,
migrates to the deepest reaches of the Mekong River between Sambor and Stung Tren during the dry
season (Sao-Leang and Dom Saveun, 1955). The larger individuals of Lates niloticus are reputed to
frequent the deeper parts of the African rivers within its range. Similarly Carey (1967a) noted that
individuals of several species were much larger in the Kafue River than in the adjacent lagoons.
Several species are pelagic within the main channel, and also in the larger permanent water bodies. In
African rivers, small groups of Brycinus macrolepidotus are to be seen cruising at the surface under
overhanging vegetation, and small clupeids characins or cyprinids occur in the surface waters of all
three continents. The surface film presents a specialized habitat occupied by small cyprinodonts
which are found in all quiet stretches of river and floodplain alike.
Three other special habitats have been noted by Lowe-McConnell (1964, 1967) and Mago-Leccia
(1970). Some fishes burrow into sand bottoms. Gymnorhamphichthys hypostomus of the Rupununi
and Venezuelan savannah rivers has an elongated snout which facilitates respiration while buried.
Potamotrygon hystrix and Xenogoniatus have also been recorded from the bottom of Venezuelan
streams. A somewhat similar habit has been noted by Daget (1954) and Cromeria nilotica which
burrows in sandy bottoms of the Niger River when alarmed.
Leaf litter on the bottom of forested streams and pools also yield a rich harvest of small species some
of which do not occur elsewhere in the system. Lowe-McConnell listed Agmus lyriformis and
Farlowella sp. to which Mago-Leccia added Aequidens, Apistogramma, Orinocodoras, Corydoras,
Agamyxis, Homodiaetus and Hyphessobrycon. The crevices and hollows of the mass of decaying
branches which also accumulate in such creeks are inhabited by several small types of fish. Lowe-
McConnell reported having collected 17 species and over 200 individuals in two hours from split logs
and branches. In the Rupununi these were mostly catfishes, such as Platydoras, Trachycorystes,
Pseudopimelodus, Hoplosternum or Ancistrus. In the Apure River some gymnotids also have been
recorded from the crevice dwelling habitat including Electrophorus electricus, Sternopygus macrurus
and Apteronotus albifrons, although catfishes such as Panaque nigrolineatus also occur there. Both
Lowe-McConnell and Mago-Leccia maintained that crevice dwelling is associated with nocturnal
habits. Specialized habitats of this type have only been studied in Latin America, but it seems
probable that they also exist in both African and Asian inland waters.
The blind arms and backwaters of the main channel remain in communication with the main river at
one end, but are otherwise lentic, having many of the characteristics of floodplain lagoons. They are
usually especially rich localities, as silt accumulates in them giving rise to plankton blooms and
increased primary production. They are sheltered and accumulate tree trunks, branches, rocks and
other types of cover providing material for shelter so many fishes enter them as a refuge from the
current of the main stream. As such backwaters are of particular importance as major concentrators of
ichthyomass, and in rivers such as the Danube and the Mississippi are the main surviving features of
the flood system. In the Mississippi particularly they are identified by Schramm et al. (1974) as
providing essential habitats for feeding and reproduction in at least 57% of the fish species recorded
from the reach between St. Louis and Cairo. Guillery (1979) also identified a movement of 15
mainstream species on to the floodplain coincident with flooding of which some adults and juveniles
remained trapped in sloughs and blind areas during low waters.
The floodplain
When inundated the plain has a rich mosaic of habitats although there is little information on the
distribution of fishes amongst them. A basic difference exists between forested and savannah plains,
although originally many plains that are now exposed supported greater tree cover. This means that
much of the present day fauna on the now denuded plains has become modified in recent times as
human activities have worked upon the environment. Flooded rain forests themselves appear to be
varied habitats.

137
Observations on the fish faunas of flooded forests are limited, but indications from Grand Lac of the
Mekong (Bardach, 1959), the Congo River (Matthes, 1964) and the Amazon (Roberts, 1973) attest to
the variety of species in such fish communities. Goulding (1980) particularly has investigated the fish
fauna inhibiting the Amazonian Igapo showing the importance of the fruit and seed bearing trees to a
variety of fishes, including Colossoma macropomum, C. bidens, Brycon spp., Myleus spp., and
Serrasalmus spp. Occupancy of the flooded forest by these species is closely allied to the distribution
of these favoured fruit bearing trees.
On floodplains with more restricted gallery forest or bush scrub, the submerged branches and roots
provide a feeding substratum and concealment for many species. Mago Leccia (1970) lists several
fishes from the Orinoco river which occur among flooded scrub. These include nocturnal armoured
catfishes such as Hypostomus and Pterygoplichthys and the gymnotids, Sternopygus macrurus,
Rhamphichthys rostratus, Adontosternarchus sachsi and Eigenmannia. Diurnal forms such as the
cichlids Astronotus ocellatus and a multitude of characids including Triportheus, Cynopotamus,
Astyanax, Moenkhausia and Thoracocharax were also found. The flooded scrub habitat is common on
most tropical plains, and as observed above, probably more nearly approximates to the original
condition of most of them. Thus, fish communities of this type are presumably widespread although
they have not been specifically described from other systems.
The floating meadows, which now seasonally cover many of the world's savannah floodplains, appear
at first sight to contain little variation. Closer examination reveals a fine texture allied to contour.
There are deeper places around lakes and depressions which are often free of vegetation or have a
flora more typical of permanently wet areas, and shallower places over the levees and terraces. Local
differences in vegetation and bottom type are associated with these features and it may be assumed
that fish segregate accordingly. There is no doubt that certain areas do have particular attraction for
characteristic species. The most important factors controlling such distribution are probably dissolved
oxygen concentration, depth, substrate and vegetation cover.
The littoral zone of the plain, that area of interface between land and water, is usually colonized by
young fish. In Africa these are almost entirely cichlids of the general Tilapia, and Sarotherodon or
Oreochromis and cyprinodonts, which have specific tolerance of the elevated temperature found
there.
On some of the most extensive plains, where sheet flooding is common, there is an intermediate zone
where the areas flooded by rainwater meet those inundated by the rising river level. This zone marks
the boundary between two types of water of different productivity and may limit the distribution of
fish on the plain. Blanc, Daget and D'Aubenton (1955) distinguished two zones on the Niger river
floodplain, a) a zone corresponding to the major bed of the river which was rich in fish and b) a
peripheral flood area which was less densely colonized. A similar distribution pattern has been
reported from Arauca-Apure flood system by Matthes (pers.com.), from the flooded forests of the
Mekong by Le Van Dang (1970) and from the Sudd of the Nile River (Mefit Babtie, 1983). Some of
the patchiness in fish distribution on other large floodplains may be attributable to such differences in
water quality.
In the dry season most of the plain is drained leaving only the network of depression pools, lagoons
and swamps, some of which dry out and some of which persist until the next flood. Most fish leave
the plain at this time, but a certain section of the community remains in these standing waters. Of the
fish remaining, a proportion is composed of species which would normally retreat to the river
channels, but which have become isolated in various depressions. The majority of these die through
desiccation of the water body in which they find themselves, through deoxygenated conditions, or
through exposure to excessive temperatures. Sane find their way to deeper lakes where they may
survive.

138
An assortment of fishes stay on the plain. Daget (1954) and Blanc, Daget and D'Aubenton (1955)
listed Marcusenius senegalensis, Pollimyrus isidori, Petrocephalus bovei (mormyrids), Gymnarchus
niloticus, Heterotis niloticus, Ctenopoma, Parachanna, Polypterus, Synodontis, Clarias, Hepsetus,
Auchenoglanis and Heterobranchus as comprising the characteristic fauna of the floodplain pools of
the Niger river. These genera recur on floodplains throughout their range in Africa, which in some
cases is very widespread. In the Mekong, the blackfish assemblage described by Sao-Leang and Dom
Saveun (1955) contains a similar group of fishes, many of which belong to the same families and
closely resemble the African species. Major examples such as Ophicephalus striatus, O. micropeltes,
Anabas testudineus and Clarias batrachus remain in depression pools throughout the dry season to
spread over the plain during the flood. Species recorded from the lagoons and pools of the Apure
river, Venezuela, by Mago-Leccia (1970) included Hoplias malabaricus, Serrasalmus notatus,
Callichthys callichthys, Hoplosternum littorale, Pseudoplatystoma fasciatum, Sorubim lima.
Pimelodus, Leporinus and Pimelodella. Similar assemblages, often involving the same species or
genera, have been described from the Rupununi by Lowe-McConnell (1964), from the Parana river by
Bonetto et al. (1969) and from the Magdalena by Kapetsky et al. (1976).
There are considerable differences in the species composition of fish population from floodplain pools
of the same system, as typified by the data given in Fig. 5.7. Attempts have been made to correlate
these with the number of variables. Welcomme (1975) traced the increased specific diversity of
populations with size of pool in the Ouémé system (Table 5.2). That size of pool can influence species
composition has also been found by Lowe-McConnell (1964) who found that larger species inhabited
the larger bodies of water. Holden (1963) found the same effect on the Sokoto River and noted that
individuals of the same species tended to be larger in the bigger pools.

139
Figure 5.7 Percentage representation of major fish groups in eight floodplain depression lakes of the
middle Parana system. (After Bonetto et al., 1969a)

140
Table 5.2
Differences in species composition of catches from permanent floodplain lagoon of
different areas of the Ouémé floodplain (FAO/UN, 1971)
Species
Percentage of species in lagoons
smalla mediumb largec
Normally swamp
dwelling species
with auxiliary
breathing organs
Clarias ebriensis 72.2 20.0 1.3
C. lazera 5.0 13.6 3.4
Habitually found
only on flood-plain
Ctenopoma kingsleyae 0.9 7.2 P
Gymnarchus niloticus P 2.1
Heterotis niloticus 26.0 2.6
Parachanna obscurus 23.8 27.2 1.6
Polypterus senegalus P 0.3 0.7
Protopterus annectens P 0.8 0.9
Xenomystus nigri P 0.2 P
Occasional swamp
dwelling species
without auxiliary
organs
Citharinus latus 0.1 1.2
Distichodus rostratus 0.7 8.1
Hepsetus odoe 2.3 2.6
Found in flood-
plain or river
Chromidotilapia guntheri P P P
Hemichromis spp. P P P
Tilapiine cichlids 1.6 2.2
Synodontis spp. 15.2
small mormyrids 18.4
Species normally
found in river
Hyperopisus bebe P 5.4
Mormyrops deliciosus 18.4
Labeo senegalensis P
Schilbe mystus 6.0
Lates niloticus 10.1
a up to 500 m² b 500–5 000 m² c over 5 000 m²
The influence of size of pool may depend on the heightened dissolved oxygen concentration arising
from lessened vegetation cover and improved aeration by wind. Such relatively favourable conditions
allow many more species to survive, including some which would normally return to the river channel
in the dry season. Greater area al so leads to a greater diversity of habitat, with more varied bottom
types, submersed, floating and emergent vegetation, and open water in the place of the densely packed
stands of aquatic plants found in the smaller pools. On the basis of samples from a selection of
floodplain pools from the Sokoto River, Holden (1963) concluded that several species were
distributed according to substrate. Although no species was confined to any one bottom type, certain
species, such as Alestes dentex, were more common over sand. Oreochromis galilaeus was found
more frequently over mud and Tilapia zillii over intermediate bottoms. There is little explanation for
the variability in species composition found by Bonetto et al. (1969) (Fig. 5.7) in the pools of the
Parana river, either in terms of vegetation cover or size of water body, although Cordiviola, de Yuan
and Pignalberi (1981) attribute the variations in these and other lagoons to a number of factors.

141
Firstly, geographical location in the river valley determines overall species composition with
Triportheus paranensis, Moenkhausia dichroura, Acestrorhynchus falcatus and to a certain extent
Prochilodus platensis being dominant to the North (Corrientes area) and P. platensis alone being
dominant to the South (Santa Fe area). Elsewhere Bonetto (1975) has stressed the abundance of mud
eating fish in lagoons whose bottom is of fine particulate organic material. Secondly specific content
of the catch may be generally related to a complex of topographical and biological factors related to
the biology of the species. Thirdly, while some species are of constant occurrence in the ponds over at
least five years the presence of other species appears much more haphazard. Depth and exposure may
also influence the distribution of species composition of the shallow sheltered bay habitat of
Magdalena river lagoons, where Potamotrygon and Pseudoplatystoma dominate, as opposed to the
open water where Hemiancistrus, Triportheus and Prochilodus are more cannon. Other species such
as Plagioscion are distributed more or less indifferently.
How much the distribution of species among the dry season habitats is a matter of chance, with only
those specially adapted species surviving adverse conditions, and how much is a matter of deliberate
selection of habitats is not clear. It seems likely that those blackfish species, which always remain on
the plain, seek out, or maybe never leave, the vicinity of the depressions where they pass their lives,
and the year-to-year recurrence of such species in any one lagoon would support this. With migratory
species, on the other hand, there would appear to be a considerable element of chance and the precise
composition of the fish fauna of the larger lagoons is probably determined by the haphazard trapping
of such individuals. The shoaling habit of many of these fishes would lead to them being either
isolated in large quantities, or not present at all, giving skewed distributions of abundance.
There is some evidence from the Rupununi swamps that species do select their dry season habitats
very closely. In Lowe-McConnell's list of 129 species, 79 percent were found in only one habitat
(excluding channels draining the floodplain which would be likely to have a high proportion of
transient species). More generally, however, distribution patterns do not appear to be so exclusive and
preferences are shown by greater concentrations in one locality rather than another. Such selection
may be by species, although different life stages or age groups of the same species may also segregate
in this way. Intraspecific resource partitioning of this type has been shown for fish from headwater
streams in U.S. rivers such as the pearl dace, Semotilus margarita where in summer the 0 year class
occupies shallow pools and deep channels and age 2+ fish occupy deep channels and pools. This
segregation is also reflected in differences in feeding pattern which may reduce competition between
age classes in this omnivorous species (Tallman and Gee, 1982). This type of segregation is especially
common in those species which utilize the rhithron either as a full time habitat or periodically for
breeding. There the use of riffles for depositing spawn, for the development of the fry and the growth
of juveniles is widespread, whereas the pools serve to house the larger individuals of the resident
species. Oreochromis species also have different habitats for feeding of juveniles (nurseries in the
littoral zone) and adults (the bottom in deeper water). They also segregate for breeding, with the
mature males being found over distinct nesting sites on the bottom, non breeding adults, both male
and female, staying in mid-water adjacent to the breeding sites, and brooding females carrying eggs in
their mouths being located in sheltered brooding sites near the nurseries. Not all fishes show such
complex distribution patterns, but in many, some phase of the life cycle (usually, but not always the
immature stage) is passed in a habitat other than that frequented by the other age groups of the
species.
DISTRIBUTION IN TIME AND MIGRATION
TYPES OF MIGRATION AND MOVEMENTS
Migrations of Adult Fish
Because the best breeding habitat rarely coincides with the best feeding habitat most species have two
distinct centres of concentration and fish have to travel, sometimes over long distances, between the
two. Two components of such movements have been recognized by Daget (1960) for tropical African

142
species. His categories of longitudinal and lateral migration are of general application to flood rivers
everywhere. Longitudinal migrations are those which take place within the main river channel, and
lateral migrations are those whereby fish leave the main channel and distribute themselves over the
floodplain.
Four phases of fish movement have been identified separately by Blache et al., (1964) and Williams
(1971) from the Chari and Kafue rivers respectively. Combining their two classifications, six main
phases in the distribution of fish emerge:
(a) longitudinal migrations within main channel: these are usually upstream, but not always so;
(b) lateral migrations on to the floodplain;
(c) local movements on the floodplain and distribution among flood season habitats;
(d) lateral migration from the floodplain towards the main channel;
(e) longitudinal migrations within the main channel: these are usually downstream but not always so;
(f) local movements within the dry season habitat: this may be the river, adjacent lake or in sane
cases the sea.
Although these are broadly applicable to the majority of individuals of most fish species inhabiting
flood river systems, some species are confined to one habitat only. It is also not certain that all
individuals of the mobile species do in fact undertake migrations every year. Within the above pattern
three distinct groups of freshwater species can be distinguished.
(i) The “blackfish” species, whose migrations between dry and wet season habitats are restricted and
which, at the most, undertake lateral migrations to the fringes of the main channel. These species are
more normally confined to the plain spreading over it during the floods from the residual pools and
lagoons which are the dry season habitat;
(ii) Those species which undertake moderate movements within the river, but which spawn on the
floodplain. The major migrations are for dispersal to dry season habitats such as those of the African
Brycinus leuciscus described by Daget (1952), or Crossocheilus reba (Tongsanga and Kessunchai,
1966), and other Mekong species (Bardach, 1959). Migrations to the favoured breeding places on the
floodplain may be either upstream or downstream and rarely involve the formation of large shoals.
Migrations of this type appear to be mainly for the avoidance of unfavourable conditions on the plain;
(iii) The “whitefish” species which undertake an upstream migration during the dry season or early in
the wet season. Such migrations are usually linked both to reproduction and to the need to escape the
adverse conditions of the downstream river channels and lakes in which water levels and dissolved
oxygen concentrations may become dangerously lowered for sensitive species.
The migrations of freshwater fish in the tropics are best known from the South American continent
where a wide variety of migrations occur. A number of medium sized systems seem to have migratory
fish populations showing similar characteristics with a single seasonal movement to and from a down
river feeding zone and an up river feeding zone. Such patterns have been reported from the Rupununi
river with Boulangerella cuvieri, Hydrolicus scomberoides and Myleus pacu (Lowe-McConnell,
1964), the Pilcomayo river with Prochilodus platensis (Bayley, 1973), the Sao Francisco river with
Prochilodus sp. and Duopalatinus emarginatus (Paiva and Bastos, 1982) and are presumably repeated
throughout most moderate size rivers of the continent which carry characin and siluroid populations.
The most complete studies of this pattern of migration are from the Mogi Guassu in which a

143
succession of fish species moves up-river at rising water (Godoy, 1975) (Fig. 5.8). A similar but more
complicated sequence of movements is shown by several species from the River Magdalena. Here
there are two seasons of activity, the first the major “Subienda” involves a migration from the
extensive deltaic downstream floodplain to the more shallow upstream reaches in February and
March. Prochilodus reticulatus is the main species involved, although it may be accompanied by
others including Brycon moorei, Pimelodus clarias and Pseudoplatystoma fasciatum. This is followed
by a downstream movement in April to June and a second, minor, upstream migration the “mitaca” in
July to September with a final downstream movement in October-December. Prochilodus reticulatus
is also involved in similar migration upstream from the floodplains of the Catatumbo River at its point
of inflow into L. Maracaibo.
Figure 5.8 migration of fish within the Mogi-Pardo-Grande river system. (After Godoy, 1975)
In larger rivers the migration patterns tend to become more complex as migrations within the tributary
rivers at the main river channel are mixed with movements between these various elements of the

144
system. Thus different but equally complicated patterns have been reported from the Parana River
system and from the Amazon with some observations from the Orinoco indicating an intermediate
level of complexity.
Two main species Prochilodus platensis and Salminus maxillosus has been studied in the Parana and
Uruguay rivers by Bonetto and Pignalberi (1964), Bonetto et al. (1971) and Bonetto et al. (1981)
although several other migratory fishes were also investigated by them. The studies which combined
observation and tagging experiments show the movements of fish in the Parana and its tributaries to
be elaborations of the simple upriver and downriver translocations between feeding and breeding
sites. Prochilodus shows fairly straight forward movements in the peripheral rivers such as the
Pilcomayo but in the main stream individual fish appear to move upstream or downstream more or
less indiscriminately. This seeming confusion arises from the size of the system, the thermal regime in
the South which is severe enough to influence behaviour, the assumed existence of separate upriver
and downriver populations and to the presence of several major tributaries. A further example,
Salminus maxillosus, moves downstream in the Parana River in October whereas the same species is
ascending the Bermejo River at the same time. In an attempt to resolve these complex patterns
Bonetto et al. (1981) proposes the diagram in Fig. 5.9 to represent current thinking on fish movements
in this river. In at least one species from the Parana, Luciopimelodus pati, migration occurs in late
summer on falling water when fish travel upstream for 400–600 km to arrive at the spawning sites just
before the next rise in water level. The curious feature of this migration lies in the behaviour of the
male fish which tend to remain upriver to an increasing degree as they grow older so that females
between 40–70 cm in length come to dominate the downstream populations.

145
Figure 5.9 Summary diagram of principal movements of fish within the Parana River. (Bonetto et al.
1981)
Migrations of fish in the various components of the Orinoco system are still very imperfectly
understood, although observations have been made on Semaprochilodus laticeps in the lower reaches
of the river from Caicara to the delta (Novoa, 1982). Behaviourally this species resembles P. platensis
in that eggs and young fish drift downstream from an upriver spawning site, move laterally into
lagoons where they grow and subsequently emerge to migrate up the main channel to spawn some
two to four years later. This pattern of movement produces the type of upstream increase in mean
adult size observed by Goulding (1981) for the Madeira and for male Luciopimelodus pati in the
Parana and would suggest that in the Orinoco at least one discrete stock of S. laticeps is involved.
In the Amazon several types of migrations appear to be undertaken by the same species. Many of the
larger characin genera, Colossoma, Brycon, Mylossoma, Triportheus, Leporinus, Schizodon,

146
Rhytiodus, Prochilodus, Semaprochilodus, Anodus and Curimatus show a pattern of migration
originally described as “Piracema” by Ihering (1930) and Geisler et al. (1973). Recent investigations
by Goulding (1980) in the Madeira river, Goulding and Carvalho (1982) in the Amazon and Ribeiro
(1983 and a) in the Rio Negro have shed more light on the nature of these movements which can be
broken down into three components (Fig. 5.10).
Figure 5.10 Diagram of assumed migration patterns of prochilodontid fishes in the Amazon and
Madeira rivers
A. The cycle starts with a migration downstream in a tributary river on the rising flood which
culminates in breeding at the mouth of the tributary. Eggs and young fish then drift downstream in
the main channel until of sufficient size to move into marginal vegetation of the floodplains or
other suitable nursery sites.
B. Adult fish then re-ascend the same tributary and disperse into the flooded forest to feed.
C. As water levels fall vast shoals of fish move down the tributary, up the main channel and into the
next tributary upstream. It is this phase of the migratory cycle which is referred to as the true
“Piracema”.
This type of migration results in a lack of juvenile fish in the tributaries and an accumulation of larger,
older fish as one moves towards the headwaters and is probably adaptive to the very poor trophic
conditions for juvenile fish in the flooded forests fringing the black-water tributaries.
In Africa characins and some siluroids are conspicuous among the migratory species, but the
cyprinids also show this type of behaviour. Potamodramous migrations, whereby adult fish leave
lakes and ascend rivers during the floods to spawn in the upstream swamps, have been described by
Whitehead (1959) for 18 migratory species in the Lake Victoria/Nzoia river systems, and De Kimpe
(1964) for some mormyrids, characins and cyprinids of the L. Mweru/Luapula river system.
Carmouze et al. (1983) distinguish three groups of migrants in the Lake Chad basin. True migrant

147
species, Alestes baremoze, Brachysynodontis batensoda, Distichodus rostratus, Marcusenius
cyprinoides, Petrocephalus bane and possibly Hemisynodontis membranaceus, Labeo senegalensis
and Hydrocynus brevis made large-scale longitudinal movements for breeding. Such movements were
usually between the lake and the Yaeres floodplain, but a further species A. dentex appeared to
reproduce in the upper reaches of the Chari and Logone rivers and hence made even more extensive
migrations. Mixed migrants made similar large scale longitudinal movements whose motives were
less certain as both adult and juvenile fish were involved. These species included Schilbe
uranoscopus, Synodontis schall, Hyperopisus bebe, Mormyrus rume and Eutropius niloticus. Finally a
well defined class of lateral migrants was present whose movements between the river channel and
the floodplain were mostly associated with feeding.
A similar group of species is involved in an annual migration up the Senegal River from the lower
reaches around Dagana to as far up river as Dioulde-Diabe where further movement is asserted by a
shallow rocky sill across the river bed. The fish then move laterally across the plain during the floods
and return to the river in falling water. The downstream movement was found by Reizer (1974) to
take the form of a drift produced by the failure of fish to combat the current in the main channel. As
larger fish are generally more capable of combating the current there is a tendency for them to be
swept less far downstream and a noticeable gradation in mean size results in smaller fisher nearer the
mouth of the river.
Long distance spawning migration has also been recorded in S.E. Asia where Probarbus jullieni
migrate up the Mekong River to spawning sites having shallow water, moderate current and sandy
bottoms. Sritingsook and Yoovetwatana (1976). Day (1958) notes that there are several components
of the fish communities of Indian rivers which move long distances to the spawning sites in the hill
tributaries of the major rivers. Species of the genus Tor are particularly famous for their ascent of the
Himalayan streams. Several cyprinid species migrate during the early part of the spring flood in the
Tigris/Euphrates system (FAO/UN, 1954). Notable amongst these are Barbus xanthopterus, B. grypus
and Aspius vorax whose adults spawn on upstream gravel beds, and whose young later drift
downstream to occupy the flood lands in the lower reaches of the river complex. Other cyprinid
species remain downstream to spawn in the river channel or in the swamps, e.g., Barbus sharpeyi or
B. luteus.
Not all migrations are for breeding, however, as Thynnichthys vaillantii appear to move mainly to
avoid adverse conditions in their downstream habitat during the dry season. This species migrates out
of the flood lakes of the Mahakan system of Borneo during falling water and moves progressively
upstream as water levels and dissolved oxygen concentrations fall downstream. The movement is
accompanied by gonadial ripening but spawning is not reported to occur until the fish have returned to
the lakes during the rising flood (Saanin, 1953). The abandoning of downstream lacustrine habitats
due to unfavourable conditions has also been recorded among the Mesopotamic fishes which also
move out of the marshes and floodplains of the Tigris/Euphrates at low water.
In European rivers many species are normally long distance migrants to the extent that Mahon (1984)
comments on the prevalence of this behaviour in European rivers as contrasted to the relatively low
percentage of migrant species in equivalent North American waters. Most of these species still
undertake seasonal migrations although the highly modified state of most waters has limited these
severely. Many records exist of migrant species which have disappeared following the damming of
rivers but Zambriborsch and Nguen Tan Chin (1973) cite examples of continuing semi anadromous
migrations by Aspius aspius, Blicca bjoerkna and Abramis brama in the Kiliya arm of the Danube,
and Belyy (1972) attributed similar behaviour to Lucioperca lucioperca from the Dneiper.
In estuarine deltas where sea water penetrates upstream in the dry season, freshwater species also
undergo local movements to avoid the saline conditions. Reizer (1974) noted that the more sensitive
species move as a wave for up to 100 km in front of the saline tongue. In the Senegal River these are
Hyperopisus bebe, Mormyrus rume, Mormyrops deliciosus, Marcusenius senegalensis, Alestes dentex,

148
Citharinus citharus, Labeo senegalensis and Schilbe mystus. Similar migrations undoubtedly occur in
all other rivers having a long zone of interaction between sea and fresh waters.
Marine and brackish water species also show a variety of migration patterns. Entry into the lower
reaches of the river with the saline waters during the dry season may be regarded as an extension of
the estuarine environment. However, penetrations into the freshwaters further upstream are also a
common feature which has been described from many systems. Fish of marine origin regularly move
many hundreds of kilometres up such rivers as the Niger in Africa (Reed et al., 1967), the Mekong in
Asia (Shiraishi, 1970) or the Magdalena in South America (Dahl, 1971). These penetrations of the
freshwater habitat may be for feeding or for breeding. Some notable anadromous migrations also
occur, for instance that of Hilsa ilisha, which moves up many Indian and Southeast Asian rivers to
spawn (Pillay and Rosa, 1963), the Golden perch, Macquaria ambigua, which migrates up the Murray
river in Australia (Butcher, 1967). The salmonids of Europe of N. America or the many migratory
species of the Volga river which moved into the headwaters of the river each year prior to the
construction of the dams there (Poddubnyi in Mordukhai-Boltovskoi (1979). Other less spectacular
breeding migrations are those of the eleotrids, such as Batanga lebretonis or Dormitator latifrons,
which enter rivers to spawn on the flooded vegetation fringes of the lower reaches.
Movement of Juveniles
Originally it was suggested that only the longitudinal component of migration was active, being
directed by physiological stimuli, and that lateral migration was more of a passive affair with fish
being swept by the rising floods on to the plain. It has since become apparent from the orderly
succession of the migrations, and from the way in which fish often swim against the current to gain
access to the plain, that this is not generally the case. In fact it would appear from the available but
rather circumstantial evidence that most movements of healthy adult fish are directed rather than
passive. Passive drifts do occur, nevertheless, where the eggs and larval forms of several species
profit by the current for transport from the spawning site. Observations from the Danube and the
Tigris/Euphrates systems and from various rivers in the United States (e.g., Muth and Schmulbach,
1984) have shown the earlier phases of return movements of younger stages to be by passive drift
with the current. Similarly Godoy (1959) noted that the eggs of Prochilodus scrofa are semi-pelagic
and are carried downstream by the current from the spawning sites which are in mid-channel. The
eggs, however, develop rapidly and the fry soon take refuge in flooded oxbows only to be washed out
later by falling water when they swim downstream to the major feeding grounds. In the Pilcomayo
river, lateral floodplains are absent in the upper courses and Bayley (1973) supposed that the eggs and
fry of P. platensis are swept directly downstream. Similar drifts are suggested by Novoa (1982) for
Semaprochilodus laticeps in the Orinoco, several species of characins in the Amazon (Goulding,
1980) and Hilsa ilisha in the Indus (Islam and Talbot, 1968). Reynolds (1983) also draws attention to
the differences in migration patterns in the Murray-Darling River as a correlate of reproductive
strategy. Here only those fish with buoyant eggs such as the golden and silver perches undertake
extensive upstream movements in order to allow the eggs space for their downstream drift. Some light
is shed on the nature of the passive larval drift through work in the Missouri river (Harrow and
Schlesinger (1980) where a rich and varied ichthyoplankton was composed of larvae of Aplidonatus
grunniens, catastomids, cyprinids, centrarchids, sturgeon and paddlefish. The larvae disappear from
the drift abruptly at about 8–12 mm in length either by catastrophic mortality or because they leave
the drift to occupy more protected and food rich habitats. The latter view is supported by the rapid
build-up of post-larval and juvenile fish in the backwaters of the river coincident with this
disappearance and would also indicate purposeful movements beyond a certain size. Similar
conclusions were reached by Nezdoliy (1984) for Lucioperca lucioperca, Perca fluviatilis and
Abramis brama fry in the Ili River when the initiation of active migration was marked by schooling
behaviour.
The mechanisms for “passive drift” movements of juvenile fish may be complex as Pavlov et al.
(1977) found in the Volga and Kuban rivers where juveniles of several cyprinid, clupeid and gobiid

149
species move downstream from the spawning grounds to the feeding grounds by drift. This occurs at
night in clear water rivers probably due to the loss of visual fix by the young fish, thus there is a
marked diurnal migration pattern. In the Kuban, a turbid river, the drift continued around the clock.
Vertical distribution of the young fish differ considerably during the 24 hour period and horizontal
distribution patterns appear to be species specific with some species located in the middle of the
channel and others along the banks. Vertical distribution is also apparently highly biased in some
species such as Catastomus commersoni which tend to occupy the surface of the water rather than the
bottom, particularly as individuals increase in size (Clifford, 1972). Such distributions are regarded by
Clifford and by Pavlov et al. as an active process controlled by definite behavioural reactions.
Therefore the downstream movement, although passive in the sense of propulsion, may involve some
active participation thereby differing from the distribution and transport pattern of inert matter or
plankton within the stream.
The larvae of some species which lay their eggs in floodplain lagoons congregate on the bottom and
show the peculiar behaviour of frequently ascending to the surface and then sinking back down again.
Daget (1957) noted this behaviour in Heterotis niloticus and interpreted it as a respiratory adaptation.
However, similar behaviour in Labeo niloticus (Fryer and Whitehead, 1959), and Lucioperca
lucioperca (Belyy, 1972) and several cyprinid species from the Lli River (Nezdoliy, 1984) is thought
to be to catch currents for transport to the river and eventually downstream. In fact Belyy found that
fry were moved considerable distances in this manner and it could be that the same behavioural
pattern serves the two functions equally.
The downstream movement of juvenile fish within the main channel is an important phase of the total
migratory pattern of long distance migrants. It ensures the return of stocks to areas vacated by the
adults, and in many cases, where the feeding grounds differ from the breeding grounds, enables the
young fish to reach sources of food. However, the passive drift type of behaviour contrasts with that
of fry spawned in the mountain headstreams of Indian rivers, which according to Day (1958) take
their time migrating downstream and are often halted for one or more dry seasons in the residual pools
of the main river channel. This type of behaviour would seem better adapted to continual survival in
torrential streams where the loss of control inherent in the transport by drift may rapidly carry the fry
outside the range of the preferred habitat.
DISTANCE AND SPEED OF MOVEMENT
The main long distance migrations of tropical freshwater fish are those undertaken by the South
American characin species. Tagging experiments, particularly those undertaken on the Parana system,
have yielded considerable information on the extent of such movements. Bonetto and Pignalberi
(1964) tagged 40 000 fish of which 70% were Prochilodus platensis. A second and third series of
tagging (Bonetto et al., 1971, 1981) gave more details of migration distances and routes for this and
other species. In this series of experiments the maximum distance travelled downstream by P.
platensis was 650 km at 3.3 km/day; although the mean migration speed was 7 km/day. In 1962 the
maximum distance covered downstream was 500 km at a mean speed of 5.8 km/day. A considerable
proportion of the fish did not migrate but stayed near the point of release. Salminus maxillosus tagged
in the same series of experiments travelled further, 1000 km in 60 days (16.7 km/day). This and other
results are summarized in Table 5.3.
Table 5.3
Upstream and downstream migration rates in km/day of some species from Parana River
(from data in Bonetto et al., 1981)
Species Maximum recorded distance Migration rate
travelled (km)

150
Upstream Downstream Upstream Downstream
Ageneiosus brevifilis 657 3
Hemisorubim platyrhynchus 635 7.3
Luciopimelodus pati 600
Paulicei lutkeni 706 14.4
Prochilodus platensis 455 939 0.5(3.2)8.77 0.3(2.8)18
Pseudoplatystoma coruscans 821 281 4–6 1
Pterodoras granulosus 308 1054 1–3 1–2
Rhaphiodon vulpinus 270 265 2 2.3
Salminus maxillosus 737 1000 0.9(9.3)21.5 0.4(2.5)3.9
Serrasalmus nattereri 260 2
In the Mogi Guassu and Rio Grande sub-system of the Parana, 17 species of characin migrate every
year over a total distance of 1228 km between the rhithronic upriver spawning sites and the potamonic
feeding sites downstream. Recorded speeds upriver ranged from 5–8 km/day for Prochilodus scrofa,
2.5–10.0 km/day for S. maxillosus and 3 km/day for Leporinus copelandii. After spawning fish moved
downstream at between 3–5 km/day.
Bayley (1973) also recorded migrations of P. platensis in the Pilcomayo tributary of the Parana. Here
he assumes the fish to travel at least 450 km from the downstream floodplains to the upriver spawning
site. Migrations of the same type were described by INDERENA (1973) from the Magdalena River,
Colombia, where the “Subienda” migration of Brycon moorei, Pimelodus clarias, Prochilodus
reticulatus and Pseudoplatystoma fasciatum, among other species, from the cienagas of the floodplain
to the breeding grounds in small Andean tributary streams is about 500 km.
Tagging experiments in the Sao Francisco River conducted by Paiva and Bastos (1982) showed that
distances travelled by different species ranged from 3 km (Hoplias malabaricus) to 530 km
(Duopalatinus emarginatus). Daily migration rates ranged from 0.03 km/day in Leporinus to 13.9
km/day in Duopalatinus and one specimen of Prochilodus travelled 250 km downstream in 30 days
(6.6 km/day).
In Africa dry season migrations are associated mainly with dispersal of fish in the river system. In the
Niger River, for instance, Daget (1952) studies Brycinus leuciscus which travelled at 1–1.5 km/hr up
to a maximum of about 9 km/day. Total distances traversed were as great as 400 km before the
construction of the Markala dam. Also in the Niger fishermen from the middle reaches between
Ayourou and Niamey in Niger maintain that many species move upstream at the beginning of the
rising flood. Fish are full of eggs at this time and it is suspected that the main destination is the
floodplain of the Central Delta some 400–650 km distant. The floodplains in the middle reaches of the
Niger are very poorly developed and probably do not provide an adequate area for breeding in those
species requiring this type of habitat. Many migratory species including Alestes baremoze and A.
dentex are indicated by Blache, Miton and Stauch (1962) and Carmouze et al. (1983) as undertaking
potamodromous movements of up to 650 km between Lake Chad and the upstream swamps. Reizer's
(1974) observations in the Senegal River also indicate maximum distances of 400 km for the main
migratory species which here include Alestes, Brycinus, Citharinus, Distichodus, Labeo, Lates and
Clarias. Other recorded African breeding migrations are of shorter, but still impressive lengths. The
ascent of species from Lake Victoria up to Nzoia river in Kenya during the flood were classed by
Whitehead (1959) as long duration (80 km or more), Barbus altianalis; medium duration (15–25 km),
Labeo victorianus and Schilbe mystus and short duration (up to 8 km), Brycinus nurse. Another
Labeo, L. altivelis migrated up to 150 km up the Luapula River from Lake Mweru at the beginning of
the floods. Williams (1971) tagged several species of fish as they left the Kafue river plains and found
that most undertook movements of up to 60 km upstream and 120 km downstream.
Among Asiatic species the Indian major carps appear to move only locally and mainly laterally on to
the floodplains within the Ganges and other river systems (Jhingran, 1968, for Catla catla, and Khan

151
and Jhingran, 1975, for Labeo rohita). Other species such as the Mahseers (Tor spp) undertake
extensive migrations within the Gangetic system running up the major tributaries to the foothills of
the Himalayas to spawn. In the Mekong several species have been recorded as covering large
distances. Shiraishi (1970) quoted migration distances of up to 1 000 km between the delta and
Vientiane during the wet season. There appears to be little evidence to support this, however, and
more reasonable estimates were given by Bardach (1959) for the movement of Pangasius sutchi from
the Grand Lac to the Khone falls, a distance of some 3–4 00 km. Pantulu (1970) did mention that
Pangasianodon gigas is suspected of spectacularly long migrations, although other species in the
Mekong, including P. pangasius, P. sanitwongsei, Cirrhinus auratus, Probarbus jullieni and
Thynnichthys thynnoides only undertake medium to long-range migrations. Another species of
Thynnichthys, T. vaillanti has been described as migratory up to 450 km upstream of its wet season
habitats at low water (Saanin, 1953). Long distance migrations also occur in Ctenopharyngodon idella
and Hypophthalmichthys molitrix in the Amur River which cover up to 1 200 km from downstream
feeding grounds to upstream breeding sites (Krykhtin and Gorbach, 1981).
The Golden Perch (Macquaria ambigua) of the Murray-Darling river system of Australian are
recorded by Butcher, (1967 and Reynolds (1983) as undertaking migration of 800 to 1 000 kms from
the lower reaches of these rivers to the headwaters to spawn. The average speed of the fish was 3
km/day and a maximum speed of 15.7 km/day. A fourth species from this system, the Silver Perch,
Bidyanus bidyanus was also found to move up to 520 km upstream.
Many anadromous fish travel considerable distances within freshwater for instance Huso huso and
Acipenser gueldenstaeti are both recorded to have moved up to 3000 km up the Volga river from the
Caspian sea. Such journeys within the rivers may represent only a small part of the total distance
travelled as species such as Salmo salar may have a preliminary route of several thousand kilometres
in the sea.
TIMING OF MIGRATION
The timing of the initiation of longitudinal migration varies considerably according to various groups
of species.
Movements of fish in the Mekong River, recorded from the commercial fishery at Khone falls
(Chanthepha, 1972) showed two separate groups. The first, consisting of cyprinids, passed upstream
in November to February and probably represented dispersal migrations after leaving the floodplain.
The second group, mainly of siluroids, passed up through the falls from mid April to July and were
possibly pre-spawning migrations. Alternations between dispersal and spawning migrations in the
same species have also been recorded in Latin America. For instance, Prochilodus mariae in the
Orinoco shows two phases of movement, one reproductive at low water (February-May) and one for
dispersal from September to December just after peak flood. In this the species is accompanied by
Brycon sp. Curimatus sp. and some predatory siluroids such as Pseudopimelodus and Sorubimichthys.
Other species adapt different timings in the same river. Thus Colossoma brachypomum migrates at
low water, while Semaprochilous laticeps and S. kneri emerge from the lagoons and migrate up the
main channel to breed early in the rising flood May to July. Similar plurimodality of migration is
demonstrated by the Prochilodus and Semaprochilodus of the Amazon. In most other systems in Latin
America the Prochilodontidae and certain siluroids complete their major upstream movements at low
waters. Thus in the Parana, Mogi Guassu and Magdalena rivers the fish time their movements so as to
arrive at the upstream spawning sites as the flood begins to rise. Goulding (1981) also describes the
migration of many species of catfish up the Teotonio cataracts of the Madeira River at low water, and
here only one species, Goslinia platynema, was observed to migrate at high water.
The pattern also appears in the Lake Chad basin where Alestes baremoze move up the Chari and
Logone rivers at low water, arriving at the entrance to the Yaeres floodplains at the beginning of the
flood (Stauch, pers.com.), in the Senegal river where the longitudinal migrations of Alestes during the

152
dry season bring them to the channels opening on to the floodplain as the floods rise and in the
movements of Brycinus leuciscus and B. nurse within the Central Delta of the Niger, although in these
species the migration is for dispersal rather than spawning (Daget, 1952).
Other families of migratory fish, particularly the mormyrids in Africa, some siluroids and most
cyprinids in Africa and Asia and the Golden perch in Australia normally initiate their riverine
spawning migrations as the floods appear. In some cases, for example, Labeo victorianus and other
small cyprinids in Lake Victoria, riverine migration may be preceded by a preparatory movement to
the river mouth in the lake itself (Cadwalladr, 1965). However, the arrival of the first freshets of the
floods in the river seems to trigger longitudinal migratory behaviour in the majority of species. This
migratory urge appears to be transformed into a general impulse for lateral movements as the bankfull
stage is reached and floods spill on to the floodplain. The observations of several workers that many
fish migrate actively against the current up channels where water is flowing out of the plain, rather
than to enter passively on inflowing currents, points to the physiological orientation of such
movements.
Migration on to the floodplain seems to develop as an ordered sequence of species. Sao-Leang and
Don Saveun (1955) described the movements of fish in the Mekong as a series of waves with siluroids
entering early in the sequence and whitefish later. In the Niger (FAO/UN, 1970), swamp tolerant
species which often stay in the larger lagoons over the dry season, tended to enter first from the river.
Earlier entries included species of Clarias, Distichodus, Citharinus and Labeo, whilst Brycinus,
Tilapia, mormyrids, Schilbe and Synodontis entered second. In the Kafue flats, Williams (1971) has
observed a different sequence that, to a certain extent, contradicts that observed above. Hare, Clarias,
Schilbe, Barbus and Tilapia migrated on to the plain while the water was still quite low and
Serranochromis and Haplochromis only moved on to the plain later. The early movement of Tilapia
and the clariids is confirmed by the University of Idaho et al. (1971) for the same area. Because of the
lack of information, which is difficult to collect, the situation with regard to movement during rising
water is far from clear, although the concept of phased migration of species on to the plain is probably
sufficiently well-established.
During normal flooding there appears to be adequate space on the floodplains to accommodate both
the resident black fishes and the migratory individuals of the more mobile species. Benech and
Quensiere (1983), however, concluded that, during periods of drought, the sedentary stocks can
restrain the annual colonization of the floodplain by riverine fishes. In the Yaeres, the disproportions
between resident and immigrant species increased in those years when floods were less intense and
was minimal during the period when virtually no wetting of the floodplain occurred. The effect may
be due in part to the prior occupation of such living space as is available on the plain, and in part to
the reduction in possible ingress channels with lessened flooding.
Movements off the plain are more easily studied and these too show that fish leave in a distinct
sequence. This appears to be roughly the reverse of the sequence with which fish enter the plain. In
the Mekong, studies by Blache and Goosens (1954) and by Sao-Leang and Dom Saveun (1955) from
trap catches of over 100 species showed that movement at the level of Quatre Bras occurs between
October and February. Here the sequence of migration was apparently conditioned primarily by the
size of the fish, the larger fish and larger species leaving first. Similar observations on size sequence
in migration have been made by Saanin (1953) in the Mahakan system where the larger individuals of
Thynnichthys vaillantii left the floodplain lakes and migrated upriver before the smaller fish and in the
Mogi Guassu where smaller species always precede the larger ones in the upstream movement.
Lunar phase also influenced the timing of the migration of whitefish in the Mekong which takes place
only during the second quarter to full moon each month. Thus there were monthly waves of migration
in which a few of the largest fish moved in October and November. In December to January mixed
groups of large fish including Cirrhinus auratus, Osteocheilus hasselti, Pangasius sutchi, Pangasius

153
larnaudi, Belodontichthys and Cyclocheilichthys were caught. In February smaller species such as
Thynnichthys thynnoides, Cirrhinus jullieni and Botia modesta dominated in the catch.
Studies by Durand (1970 and 1971) on the succession of species passing through the El Beid River
which drains the Yaeres, also pointed to there being a definite sequence of species which correspond
to different water masses. The first group, which moved at high water and had a very clear peak in
November and December, consisted of Marcusenius cyprinoides, Hyperopisus bebe, Alestes dentex
and Labeo senegalensis, preceded by a group of accompanying species, Alestes baremoze, Polypterus
bichir, Hydrocynus brevis and Lates niloticus and followed by Distichodus rostratus, Oreochromis
aureus, Pollimyrus isidori and Distichodus brevipinnis. A second well-defined group consisting of
Oreochromis galilaeus, Brienomyrus niger, Barbus and Clarias species was very abundant in January
and Ichthyoborus besse, Synodontis auritus, S. schall and Schilbe mystus appeared most strongly in
February. The first group corresponded to fish migrating at the end of the Logone floods, the second
and third groups move with the water draining off the flooded plain. More detailed descriptions of this
phenomenon by Benech and Quensiere (1982,1983 and 1984) confirmed the various species groups
and their order of migration and linked these to flood conditions on the plain. A strong diurnal pattern
was also demonstrated with diurnal, nocturnal and crepuscular migrants. Lunar phase influenced the
nocturnal migrants but this was considered of secondary importance to the hydrological phase.
Certain species may change their lunacity depending on the flood period. Thus Hyperopisus bebe,
which is purely nocturnal on the floodplain during the rising flood, is active both day and night at the
beginning of the flood and as the water is draining from the plain, i.e., at times of migration.
This example of the migrations from the Yaeres serves to illustrate the probable complexity of
migration patterns elsewhere and in some ways similar to the phased appearance of young fish on
shoreline nursery habitats in a North American stream studied by Floyd et al. (1984). Here an ordered
succession of species, from cottids through percids, catostomids, centrarchids to ictalurids, occupied
the habitats. The phasing of arrival on the shoreline was linked to the time of breeding of the species
in a somewhat extended season. Nevertheless, sufficient numbers of species were present in the
habitat at the same time as to call for their partitioning of the resource. Residence time on the
shoreline ranged from three to sixteen weeks depending on species and represented a way station
between the spawning habitat on the shoreline or the riffle systems and the adult habitat.
That adult fish leave the floodplain before the young-of-the-year is borne out by many authors. For
example, Motwani (FAO/UN, 1970) remarked that older age groups leave the swamps of the Niger
and Benue River first. The juveniles remain on the floodplain until the later stages of its emptying.
The University of Idaho et al. (1971) and Williams (1971) noted the same phenomenon in the Kafue
River. In the Yaeres, the breeding fish often do not return through the El Beid River, preferring to re-
enter the lake via the main channels of the Chari and Logone rivers. Thus Durand (1970) was able to
record that young fish make up 95 percent of the El Beid catch by number and weight. This
phenomenon is mainly caused by the peculiar flow patterns of this system (Fig. 5.11) which only
allow the adult fish to fight against the currents entering the plain through the levees and thus to
regain the main channel of the river. The young fish are not strong enough to do this and are more
strongly influenced by the current which directs them down the El Beid as it drains the plain. More
extreme examples of delayed migration are shown by the juveniles of Prochilodus platensis, which
remain in the floodplain pools of the Parana River during an entire dry season before entering the
river after the next flood (Bonetto, 1975). Prochilodus species in the Apure River show similar
behaviour (Matthes, pers.com.). That return migrations by juveniles are more complex than just a
passive movement under the influence of the movement of water as it leaves the plain is indicated by
the different behaviour of two very similar species of small cyprinid in a stream system flowing into
Lake Victoria (Welcomme, 1969). Here the adults of both species left the upstream swamps towards
the end of the flood. The juveniles of one species, B. kerstenii remained only a short time in the
swamps, moving quickly to the drainage channels and the river and migrating shortly after to the lake.
B. apleurogramma on the other hand tended to stay in the swamps until they were almost dry, moving
to the drainage channels and later to the river where it remains throughout the dry season. These

154
movements occurred at quite characteristic sizes. By contrast a third sequence of movement is shown
by a small cichlid which inhabited the same system and did not enter the lake.
External factors which might stimulate spawning migrations have been discussed for many years,
somewhat inconclusively. In view of the great differences in timing both of the initiation of
movement and of spawning among the many species inhabiting tropical river systems, it is likely that
there are a whole range of fine tuning stimuli which influence the various species in different ways.
Because the end of the flood tends to be less predictable than the beginning, and the behaviour of the
fish at this time is not apparently subject to a strong physiological impulse such as reproduction, it
seems reasonable to suppose that simple mechanical and chemical factors would be sufficient to
initiate movement out of the floodplain habitats. It is important to the survival of the species that such
signals should anticipate the onset of conditions that would be lethal to the majority of the population.
Nevertheless, it does seem that stimuli for return migrations are often less than totally effective in this
respect, as huge quantities of fish are stranded and die every year.
The generally noted tendency for both the larger species and the larger individuals of each species to
leave the floodplain earlier than the smaller fish is probably indicative that depth is one of the major
factors controlling this. The fact that the large fish often fail to move out of deeper floodplain pools
would also tend to support this supposition. Dissolved oxygen concentration and temperature also
seem to be of major importance in determining the distribution of fish within the system, and changes
in these factors are liable to provoke fish into leaving regions where conditions are less than optimal.

155

156
Figure 5.11 Migration patterns within the Yaeres-Lake Chad system. (Adapted from Durand, 1970)
Light plays a very strong role in regulating the time of migration. Several families move mainly at
night. In Asia these include siluroids and ophicephalids, in Africa siluroids, ophicephalids and
mormyrids, and in South America siluroids and gymnotiids. Other fishes including cichlids, cyprinids
and some characins, tend to move by day, yet others, such as the small mormyrids which move up the
Nzoia River from Lake Victoria to spawn concentrate their movements at dawn and dusk (Okedi,
1969). There is, in this manner a possibility for round-the-clock utilization of both the migratory
pathways and the slack water refuges and resting places. That diurnal sequences of migration may be
quite complex is evidenced by Lowe-McConnell (1964) observations on the Rupununi system. Here,
the periods for peak migration were more closely defined as follows: dawn and early morning -
Cichla ocellaris, Osteoglossum; daytime - Serrasalmus nattereri, Metynnis sp., Geophagus jurupari,
Cichla ocellaris, Cichlosoma severum, Leporinus friderici; late afternoon - Brycon falcatus, Metynnis
sp.; evening (just after dark) - Prochilodus insignis, Schizodon fasciatum; night - Pimelodus and other
catfishes.
The strong influence of lunar phase on the timing of the migration of whitefish in the Mekong has
already been mentioned. A similar phenomenon has been described for Brycinus leuciscus in the
Central Delta of the Niger by Daget (1952). This characin species forms extensive shoals during
moonlit nights for their upstream dispersal migrations. When there is no moon the shoals dissociate.
Migration is also initiated as soon after the draining of the floodplain as is consistent with lunar phase,
and because of the topography and drawdown regime of the central delta, four different migratory
groups are formed according to the successive coincidence of full moon and low water as the flood
recedes downriver. Other characin species, Brycinus nurse, Alestes dentex and A. baremoze also
migrate upstream but are not ordered by lunar phase. Upstream movements of B. nurse in fact always
precede movements of B. leuciscus from the same portion of floodplain.
Potamodramous migrations would appear to have several advantages for the fish species undertaking
them. Many species may migrate primarily to avoid unfavourable conditions in the lower reaches of
the river, but the majority of migrations seem directed at reaching localities suitable for reproduction
or feeding. By placing the young fish nearer the headwaters, or where appropriate in the main stem of
the river, their journey downstream can coincide with the flood wave over several hundred kilometres.
Juvenile fish can thereby arrive at floodplain reaches suitable for feeding grounds, which may differ
from the rocky, sandy and turbulent areas chosen for breeding, and thus ensure maximum exposure to
zooplankton in the floodplain pools. Furthermore, in forested nutrient poor rivers, such as the
Amazonian tributaries, the fringing flood forests are so lacking in nutrients that the young need to be
transported downstream to find suitable feeding and nursery habitats. This opinion is also advanced
by Krykhtin and Gorbach (1981) for Ctenopharynogodon idella and Hypophthalmichthys molitrix
from the Amur River. The eggs of these species originate from spawnings on upstream grounds more
than 1200 km from the mouth of the river, which distance rules out the possibility of eggs or larvae
being swept out to sea. Spawning at the earliest phase of the rising flood also ensures that the eggs
and larvae drift downstream and that the juvenile fish are thus positioned opposite suitable floodplain
foraging areas at the time of changeover from zooplankton to mixed feeding some four and a half
days after hatching. At the current velocity of 4.7 km/hr pertaining in the Amur even this short time
lapse may mean a drift over 500 km between hatching and seeking refuge in the side arms of the
floodplain.
A similar strategy, that of placing the young on the floodplain as early as possible also is followed by
these upstream migrants who time their arrival at the channels opening on to the floodplain to
coincide with the first flow of water on to the plain. In addition to the benefits for feeding noted
above, upstream migration by adult fish prior to spawning must have a role in counteracting
downstream drift of larvae and fry in those species that are obligate main channel spawners. Although
not analysed in detail by any studies so far it would appear that migration distances upstream as
described for the Pilcomayo (Bayley, 1973), the Parana (Bonetto, et al. 1971, 1981) or the Amur

157
(Krykhtin and Gorbach (1981) are of the same order as the distance drifted downstream by larvae and
fry given the current velocities of the rivers and development times of the fishes concerned.
Obviously during drift type movement there must be considerable dispersal of the population which
may ensure mixing of stocks over such a long river as the Parana. In other systems, such as the
Orinoco, it is, however, clear that separate sub-populations are maintained in different reaches of the
river.
There has been much discussion of the role of this potomodromous habit in African species, Jackson
(1961) maintaining that it is principally a device to protect the young from predation, whereas Fryer
(1965) considered it mainly as a mechanism to secure dispersal over the whole river course. As in so
many such arguments both participants are probably partly correct and there is no doubt that the use
of upstream swamps and spawning habitat does have the double advantage of presenting the young
fish with a rich habitat in which to start life, while at the same time giving considerable shelter from
the predation to which they would be exposed in the adult lacustrine habitat.
ADAPTATIONS TO EXTREME ENVIRONMENT CONDITIONS
Many of the habitats within river ecosystems have extreme physical or chemical conditions which call
for special adaptations on the part of the fish inhabiting them. Many of the adaptations are
behavioural, involving migrations or local movements whereby the adverse conditions are avoided. A
certain section of the fish fauna, however, has specific anatomical or physiological adaptations which
permit the species concerned to survive low dissolved oxygen concentrations or even complete
deoxygenation, high temperature, desiccation, poor light conditions or strong current.
LOW DISSOLVED OXYGEN CONCENTRATIONS
One of the main factors determining the distribution of fish within the river and floodplain system is
the availability of dissolved oxygen. As has been shown above low dissolved oxygen concentrations
or completely anoxic conditions are common on floodplains at certain times of the year. Fish have
become adapted to these conditions in a variety of ways but some species which inhabit the lentic
waters of the floodplain are strangely sensitive to low dissolved oxygen levels. Serrasalmus nattereri
and S. rhombeus for instance show the first symptoms of asphyxia when the oxygen falls below only
20 percent saturation. Such species are especially sensitive to sudden drops in dissolved oxygen and
are among the first to suffer catastrophic mortalities. There are two sources from which fish may
obtain supplementary oxygen in poorly aerated waters. These are:
(a) the air above the water, and
(b) the thin, but well oxygenated surface layer which is often only a few millimetres deep.
Many of the blackfish species inhabiting swamps have modifications which allow them to benefit
from one or another of these sources. For example, Carter and Beadle (1931) found that eight out of
twenty species occurring in the Paraguayan Chaco had anatomical respiratory modifications for air
breathing, whereas the rest used the surface layer as a source of oxygenated water.
Adaptations for Air Breathing
The development of specialized organs which enable fish to breath air has occurred independently in
many taxa and in all zoogeographic regions. Respiratory modifications have been centred on three
main anatomical systems, the mouth and digestive tract, the gills and branchial chamber, and the lung
or swim bladder. These are discussed in detail by Carter (in Brown, 1957), Norman (1975) and
various contributions in Hughes (1976) particularly that of Dehadrai and Tripathi.

158
Several species have become so dependent on air that they die if prevented from reaching the surface.
Thus the lung fishes are obliged to breathe at frequent intervals, and the paiche (Arapaima gigas)
needs air every 10–15 minutes when adult and more frequently when young (Sanchez Romero, 1961).
Modification of the digestive tract for air breathing is made possible by the stopping of feeding during
the dry season (Lowe-McConnell, 1967), at just those times when deoxygenation is most extreme.
Most parts of the alimentary canal have been modified in one family or another. The mouth cavity and
pharynx are highly papillated and well supplied with blood in Electrophorus electricus which surfaces
to gulp air. Air bubbles are passed backwards to lodge inside the heavily vascularized stomach of
Ancistrus, Corydoras, Plecostomus spp. and Hypostomus, the intestine of Hoplosternum or
Callichthys, or the rectum of Misgurnus fossilis.
The branchial or pharyngeal cavity has become modified by simple vascularization in Hypopomus and
in particular in the synbranchid eels such as Monopterus and Symbranchus marmoratus.
Mastacembelus spp. secrete a protective slime over the unmodified gills, which permits a limited
amount of aerial respiration. Three different families have developed diverticula of the branchial
cavity (Fig. 5.12). These are least developed in the Channidae whose supra-branchial chambers are
simply lined with a richly vascularized epithelium. The Anabantidae have labyrinth organs elaborated
from the first gill arch. In the clariidae the II and IV gill arches have become modified into
arborescent organs in many genera, and in Heteropneustes (Saccobranchus) fossilis the branchial
chamber is extended backwards along the body.
Figure 5.12 Adaptations for air-breathing in (A) Channidae, Channa; (B) Anabantidae, Ctenopoma;
(C) Clariidae, Clarias; (D) Heteropneustidae, Heteropneustes
Only the lung fishes (Dipnoi) and bichirs (Polypteridae) have true lungs, but several physostomous
families have modified swim bladders which act in an almost identical manner. These included
Osteoglossidae (Arapaima gigas), lepisosteidae (Lepisosteus sp.); Gymnarchus and several species of

159
Mormyridae, Erythrinidae (Erythrinus and Hoploerythrinus), Notopteridae, and Umbridae (Umbra).
Young forms of Lepidosirenidae, Polypteridae, Osteoglossidae and Gymnarchidae have external gills
which are resorbed during development when the lungs or swim bladder take over the main
respiratory function.
The ability to breathe atmospheric oxygen enables fish to colonize waters which would be otherwise
uninhabitable thereby reducing interspecific competition. Furthermore as comparatively few
ichthyophage predators have developed these mechanisms fish colonizing such deoxygenated waters
are relatively free of predation (Junk, Soares and Carvalho, 1983). It has the additional bonus that it
permits overland movement. Migration of fish over dry land, or at least over damp, swampy grounds
have been recorded in several species, particularly various clariids in Africa and Asia, but also in the
erythrinids and perhaps the callichthyids and loricariids of South America (Kramer et al., 1978). Such
mobility permits fish to escape desiccating water bodies and also to colonize pools isolated from the
main water mass.
Dehadrai and Tripathi (in Hughes, 1976) mention the energy cost of air breathing. Young
Ophicephalus punctatus kept in 40 cm of water surfaced 1 879 times per day at a cost of 161 cal/day.
In 2.5 cm of water the same species surfaces 482 times per day at a cost of 92 cal/day. Fish living in
deeper water consume about 1.5 times as much food as those from the shallower waters presumably to
compensate for the additional energy required. Another disadvantage of the air breathing habit is the
increased vulnerability to predation of the fishes by both aquatic and airborne predators during their
journey to and from the surface and at the surface itself (Kramer et al., 1982). Kramer and Graham
(1976) suggested that, in many species from many families this is to a certain extent mitigated by the
social behaviour of synchronous breathing by a number of individuals which space their breath
randomly. This type of behaviour has been observed in the laboratory in Ancistrus chagresi
(Loricariidae), Hoplosternum thoracicum (Callichthyidae), Piabucina festae (Lebiasinidae) and
Trichogaster leeri (Anabantidae), and in the field in Piabucina and Hoplosternum. Other species from
other families (Polypteridae, Lepisosteidae, Notopteridae and Megalopidae appear to behave in a
similar manner from descriptions in the literature, indicating that the adaptation may be widespread.
Adaptations for Using the Surface Layer
In species lacking specific physical or physiological adaptations the practice of utilizing the better
oxygenated surface layers of the water is common. For instance Kramer (1981) recorded 93% of non-
air breathing fishes in Panama as breathing at the surface layers and later Kramer and McClure (1982)
extended these observations to the Amazon where 31 species belonging to 14 families made
increasing use of the surface layer as oxygen concentrations dropped. Thus aquatic surface respiration
was considered to be a specific adaptation to hypoxia in fishes inhabiting shallow tropical waters.
Anatomical modifications which enable certain small species of fish to use the oxygenated surface
film of the water particularly efficiently have been described by Lewis (1970). These adaptations
include the small dorsally-oriented mouth and dorso-ventrally flattened head found in most
cyprinodonts. Lewis carried out experiments to show that in deoxygenated conditions Fundulus,
Poecilia and Gambusia all adopted a characteristic posture at the water surface where they can
survive indefinitely, provided a critical population density is not exceeded. Kramer and McClure
indicate similar morphology and behaviour in species of cichlids (Pterophyllum) and osteoglossid
(Osteoglossum). Some larger fishes too have adopted this strategy and in addition to their posture at
the surface Colossoma macropomum and Brycon melanopterus develop dermal enlargements of the
lower jaw during hypoxic conditions which facilitate the influx of surface water into the buccal
cavity. These enlargements are lost in oxygen rich water (Braum and Junk, 1982).
Physiological Adaptations

160
Physiological mechanisms in some species allow the fish to withstand low dissolved oxygen
concentrations, although not necessarily to survive under complete anoxia. For instance Blazka (1958)
found that Carassius carassius can tolerate anoxia for at least two months at low temperature. At
higher temperatures tolerance time is lessened. Much of this is due to the composition of the blood
which in some species, notably those inhabiting oxygen-poor waters, is relatively unaffected by
changes in 02 uptake produced by increases in CO2 tension (Lagler et al., 1977). There is in fact a
great diversity of response by fish to respiratory stress as described by Fry (in Brown, 1957). Work by
Powers et al., (1979) on Amazonian fishes has particularly drawn attention to differences in the
affinities of the blood of different species of fish for oxygen. Their work was summarized by a series
of curves (see Fig. 5.13) which show fish from running waters where there is much dissolved oxygen
to have blood of lower affinity than fish from still waters with low dissolved oxygen tensions. Fish
from the margins of rapids occupy an intermediate position. By extension fish occupying rapids which
dry up during the dry season may change the O2 affinity of their blood to correspond to prevailing
conditions.
ADAPTATIONS TO RESIST HIGH TEMPERATURE
Temperatures at the fringes of the flooded plain, and in shallow water bodies, may rise as high as
40°C. Certain species of fish, particularly the juveniles, tend to prefer such areas, using them as
nurseries. Here they benefit from the higher temperature and greater availability of food to grow
faster, and also as a partial refuge from predation (Welcomme, 1964). There is experimental evidence
that species inhabiting these warmer water areas have a much greater physiological resistance to the
effects of high temperatures than most other species. For example, Fig. 5.14 illustrates the difference
between the survival of Tilapia zillii and Haplochromis spp. under experimental high temperatures. T.
zillii survive indefinitely at 38°C, a temperature at which half of the sample of Haplochromis would
die within two minutes.
There are indications that such thermal tolerance may be cyclic, at least in some species. Johnson
(1976) has shown that there is a daily rhythm of thermal tolerance in Gambusia affinis which rises
some 3 degrees between morning and mid-day and falls away again towards evening.

161
Figure 5.13 A comparison of O2-Hb affinity for water breathing fish from lotic habitats in the
Amazon system. Thick solid lines enclose the range of blood equilibrium curves for species
from slow flowing zones; Thin solid lines those for species from rapidly flowing water;
Dashed lines those from rapid waters but which inhabit streams or river margins. (From
Powers et al., 1979)

162
Figure 5.14 Survival of Haplochromis species and Tilapia zillii in high temperatures when
acclimatized to 27°C
ADAPTATIONS TO RESIST DESICCATION
Very few species are adapted to survive desiccation and annual losses of fish trapped in temporary
water bodies is enormous. Some species however survive the dry period by cocooning. The African
lung fishes Protopterus annectens and P. aethiopicus burrow into the bed of a drying pool and secrete
a cocoon of hard slime in which they rest, coiled, so the mouth is upwards and connected to an air
passage. Fish have been recorded as having survived over a year of aestivation. Some murrels
(Ophicephalidae) are reputed to survive short periods of drought in a similar manner as do the
synbranchid eels of the Amazon (Kramer et al., 1978). Dehadrai and Tripathi (in Hughes, 1976) also
report that Clarias and Heteropneustes take refuge in soft mud in drying pools, and Donnelly (1978)
summarizes reports from Africa of Oreochromis mossambicus and Clarias gariepinus survival in wet
sand. Bruton (1979), in his review of the literature on survival of habitat desiccation by air breathing
clariids, supports the idea that Clarias species can survive for some time in burrows in damp mud or
wet sand. This ability, however, does not seem to extend to survival under totally dry substrates as in
the case of Protopterus to which the construction of a cocoon is unique.
Several species of cyprinodont in Africa and South America can maintain permanent populations in
temporary aquatic habitats. These are annual fishes which complete their life cycles in as little time as
a few weeks and which have several adaptations including drought resistant eggs, embryonic
diapause, rapid hatching responses to rainfall all of which adapt them to withstand prolonged
desiccation (Simpson, 1979).

163
The lowering of the water level in their environment, and possibly the associated physical and
chemical changes in water quality, appear to act as the stimulus for reproduction. The eggs are shed
on the bottom where they settle or are pushed into the mud by the parent fish which die shortly
afterwards. The eggs and growing embryos may stop their development for variable periods and such
arrests (diapauses) may occur at three stages, described by Wourms (1972) as Diapause I (dispersed
cell phase), Diapause II (long somite embryo) and Diapause III (pre-hatching). In some species the
arrest is facultative at Diapause I and II, but obligate at Diapause III. The different combinations of
Diapause can generate 8 different distributions of total development time as shown in Fig. 5.15. In
this way a single egg population can give rise to several sub-populations which allow for the repeated
loss of individual eggs under conditions which can start development but do not allow maturation and
successful reproduction. This “multiplier” effect guarantees that some portion of the egg stock will
survive to reproduce.
Figure 5.15 “Multiplier” effects of various combinations of diapause on egg stock of annual fish.
(After Wourms, 1972)
ADAPTATIONS TO POOR LIGHT
Levine et al. (1980) and Levine and MacNichol (1982) have drawn attention to the necessity for
fishes to have eyes which are fully adapted to the environments in which they live and to their own
particular position in that environment. The quality and quantity of light within river systems varies
considerably according to habitat. Thus in some mountain streams or clear water rivers the limpidity
of the water permits great light penetration and only a gradual modification of the spectrum. However,
more commonly, the highly turbid conditions of rivers and their floodplains leads to distinctly murky
conditions. Equally, the poor penetration and high spectral distortion of blackwaters requires
specialized visual adaptation. Differences in visual structures have been found among various species
from the Amazon which are directed towards life in the diverse habitats (Menezes et al., 1981).
Equally, specializations are found which correspond to the fish’s accustomed position in the water
column or to its preferred period of activity (Levine and MacNichol, 1982) which may be termed
“visual riches”. Some behavioural quirks may be explained in this manner although at present there is

164
insufficient evidence for or against such hypotheses. For instance, the migration of prochilodontid
characins from the turbid floodplains and white water rivers to the mouths of blackwater rivers for
breeding might be explained by differential visual requirements.
Many species either supplement, or completely supplant, vision with other organs, particularly in
those fishes which live at or near the bottom of the river. Thus barbels are found in most cyprinids and
siluroids, some of which are eyeless (e.g., the cetopsid catfishes). An alternative system is the
elaboration of electric organs to send impulses which presumably serve not only as communication
but also to define the environment. The gymnotids of South America and the mormyrids of Africa are
noted examples of this adaptation which in the mormyrids is coupled with greatly reduced eyes.
ADAPTATIONS TO RESIST STRONG CURRENT
Species of fish inhabiting the rocky riffles of the rhithron or the rapids reaches of larger rivers are
highly adapted to the turbulent conditions. There are three main groups of such fishes; (i) Those
which cling to the surface of vegetation and rocks, (ii) Those which take refuge from the current in the
crevices and holes between rocks and (iii) those which can swim sufficiently fast as to resist the
current.
The first group is particularly well represented by members of the family Amphiliidae in Africa
whose several genera are all adapted in various ways to life in strong currents. They are all elongated,
streamlined and with the slightly humped form that results in the fish being forced on to the bottom by
the flow. In addition the various genera possess a variety of suckers or enlarged fins with which they
cling to the substrate (Fig. 5.16a). For example, Amphilius spp. have sucker like mouths and stiff
pectoral spines. Doumea spp. have enlarged rigid pectoral spines and Phractura clauseni has been
observed to cling to the edges of leaves with suffered maxillary barbels. Similar structures are found
in the Mochokidae where Chiloglanis spp. have elaborate pectoral spines and in the Bagridae with
some Auchenoglanis spp. Sucker like mouths are also found among the Cyprinids in both Garra and
Labeo species. Numerous other families and genera have species with similar modifications
throughout the world - among which, the Gastromyzontidae, Cyprinidae, or Sisoriidae in Asia and the
Astroblepidae in South America.
Two particular adaptations fit fish for life in the interstices of rocks and anchored vegetation, sinuous,
serpentine form and small size (Fig. 5.16b). Both are found in rapids fauna where a number of genera
from various families, for instance the clariidae, with Gymnallabes and Clariallabes, the mormyridae
with Mormyrops, the mastacembelidae with Mastacembelus and the cichlids with Gobiocichla in
Africa. The cyprinids and the Gobiidae with a wide variety of species in Asia, Europe and N.
America, the Trichomycteridae in Latin America and the Galaxiidae in Australia have all developed
elongated or pygmy species.

165

166
Figure 5.16 Adoptions to swift current in some African rheophilic species.
(A) illustrates mouth suckers, stiffened barbels and stout pectoral spines;
(B) illustrates slim or serpentine form
CHAPTER 6
THE PRODUCTION BIOLOGY OF RIVER FISH
FEEDING
SOURCES OF FOOD
The richness and variety of riverine habitats provide a wide range of possible food organisms and
substrates. These originate either from within the aquatic system itself (autochthonous food sources)
or from outside the system (allochthonous food sources), although they are all ultimately dependent
on materials of external origin in the form of alluvial silt, dissolved nutrients, material washed into the
system with surface flow or decomposition products on inundated ground. These nutrients form the
basis for numerous sources of food as follows:
Autochthonous
Plankton community - phytoplankton
- zooplankton
- drift organisms
Benthic community - mud and associated microorganisms,
- coarse detritus, decomposing vegetable or animal remains
- insects and small crustacea
Plant community - plants including filamentous algae and submersed, floating or emergent
higher vegetation
Epilithic - Epiphytic - epiphytic or epilithic algae
community (“Aufwuchs”) - associated microorganisms, insects, crustacea, etc.
- this category can include the root flora and fauna of floating vegetation as
well as some detrital aggregate, that slimy coating found on submerged parts
of plants or rocks which consists of detritus, bacteria and algae.
Neuston community - surface living insects and larvae at the air/water interface
Fish - including eggs, larvae and juveniles
Other vertebrates - amphibia, reptiles, birds or small aquatic mammals
Allochthonous
Vegetable matter - leaves, roots, flowers, fruit and seeds of plants growing near the water or
overhanging the water course which contribute to the surface drift and to the
detritus
Animal matter - insects, arachnids, worms, etc., falling on to or washed into the water from
terrestrial environment

167
In rivers primary production is located more in the proliferation of higher plants than in the
phytoplankton. Epiphytic or epilithic algae are abundant only at the fringes of the vegetation mass or
on rocks and other supports. Higher plants are themselves useful for food only when young and
tender, although fruit and seeds do figure in the diet of some species. The major contribution of higher
plants to the nutrient flow is by decay and the consequent enrichment of the detritus. Because
acceptable primary plant foods are not common in river floodplain habitats, purely herbivorous
species are relatively rare. Species that do eat higher plants or phytoplankton usually have an
alternative source of food. In the case of higher plant browsers such as Tilapia zillii, the fish often
have recourse to vegetable detritus for the consumption of which they are equally well-adapted. The
scarcity of phytoplankton in flowing water systems means that this source of food, so common in
lakes, is also of minor importance.
The general absence of primary feeders means that other types of food dominate in the diet of riverine
species. Four categories emerge as of particular importance depending on locality within the river
system. These are:
Benthos which is particularly important in the headwater streams or in rejuvenated reaches. Benthic
organisms are particularly abundant in the rocky and often torrential low order streams but decline in
abundance downstream until, in the mesopotamon, they form a relatively minor part of the diet. Thus
food supply in such reaches tends to alternate between a variety of forms living in the bottom, usually
in the interstices of the rocks, in the riffles, and a drift of such autochthonous benthic organisms and
of allochthonous material in the pools. The supply of microbenthos in these spaces between the rocks
favours small size thus the young of many species of fish pass the earlier stages of their development
within the riffles.
Mud and detritus: Bottom deposits really represent two rather different kinds of food. The detritus
feeders rely on coarser decomposing plant material together with associated micro organisms and
animal communities. These comprise a high proportion of species particularly in headwater stream
and forested habitats where leaf fall accumulates in the slack of the pools or close to floating
vegetation where litter is also abundant. The resulting coarse detritus tends to be a feature of low
order streams and it becomes finer with progress downstream until, in the potamon, it forms fine
organic mud.
Mud itself contains amino acids and other organic products of decay which can be used by fish in
combination with the saprophytic bacterial and protozoan microorganisms. Bakare (1970) has
analysed this element of the diet of Citharinus and Labeo in the Niger River. The finer the particle the
greater its alimentary value, and the preferred particle size was between 0.10 and 0.05 mm, although
grains as large as 0.18 mm were taken. The finer fractions contained relatively larger amounts of
carbon and nitrogen than did the larger particles. The size of particle and the food content of the
deposit, which the fish seemed able to detect, appeared to be the major factors limiting the distribution
of these species. At the time of sampling about 70 percent of the bottom deposits were suitable for
food. Bakare noted that bottom deposits became progressively depleted of C and N during the flood
when C. citharus was actively feeding. Periodic drying of the mud may recharge the organic content
through the incorporation of dung and other decaying animal and vegetable matter. Similarly studies
by Quiros et al. (1981) in the artificial lake of Salto Grande, a river type reservoir on the Uruguay
river, showed strong correlations between both the total organic matter and organic nitrogen
concentrations in different locations throughout the lake and the catch of such iliophagous and
detritophagous species as Prochilodus platensis, Pseudocurimata gilberti, P. nitens, Curimatorbis
planatus, as well as Plecostomus and Loricaria anus. Further evidence for the ability of certain
species of fish to feed on amino acids present in bottom mud was produced by Bowen (1980) who
found that the substances were readily absorbed along the intestine of Oreochromis mossambicus
living in Lake Valencia, Venezuela. Unusual gastric juices are required to liberate protein from this
form of detritus and O. mossambicus is recorded as having gastric acid at an uncommonly low pH
(<1.5) for fish (Bowen, 1981). Bowen (1979a) had earlier established that benthic detrital-aggregate

168
contained rich organic residues (up to 45.7% carbohydrate and 1.8–14.2% protein) a large proportion
of which was in the form of non living amorphous material. Both Bakare and Sandon and Tayib
(1953) found a high proportion of mud feeders in the fish populations of the Niger and Nile rivers. In
the Niger 10 percent of the species feed exclusively on this source of food and 10 percent more
include it as a major element of the diet. The number of species, however, is little guide to the true
abundance of mud-eating fish. In the La Plata system, for instance, 60 percent of the ichthyomass of
the floodplain pools is located in the main mud-eating (iliophagous) species Prochilodus platensis
(Bonetto, Dioni and Pignalberi, 1969). Fish of the genus Prochilodus are widespread mud-eaters in
Latin America and are met in equal abundance in other systems such as the Mogi Guassu (Godoy,
1975) and the Magdalena (Kapetsky et al., 1976). Goulding (1980) also confirms that mud and
detritus eating fishes of the genus Prochilodus, Semaprochilodus and Curimatus account for a large
part of the biomass in the nutrient poor forested floodplains of the Amazonian rivers he studied.
Allochthonous material: Many workers here have remarked on the quantity of allochthonous
material consumed by fish in river systems. Not only is such external material one of the two major
food sources in headwater streams but on forested blackwater river floodplains the rain of animal and
vegetable matter from the overhanging vegetation is the only appreciable source of food and all food
chains start from it. Typical of the latter are the Amazon and Congo rivers from which Geisler et al.,
(1973), Roberts (1973) and Goulding (1980 and 1981) have noted that food from terrestrial sources is
particularly important. Similar observations have been made in the Mekong basin especially in the
flooded forest surrounding the Grand Lac. Most species in these habitats show great flexibility in the
type of allochthonous food taken, but some fish, such as the frugivores of tropical forest rivers have
specialized in using particular food items. Indeed the most exhaustive study carried out on such
species (Goulding, 1980) indicates that some Amazonian species of Characidae, Cynodontidae,
Anostomidae, Pimelodidae, Doradidae and Auchenipteridae specialize in fruit or seed eating in the
Amazon during high water to the extent that over 87% of the total food consumed by Colossoma,
Mylosoma, Myleus and Brycon in the wet season was fruit or seeds. The various species even show
preferences for particular types of fruit or seed correlated with their dentition. Most such species
either cease feeding during the dry season or turn to alternative food sources which in many cases
may consist of other allochthonous material such as leaves or flowers, but may as in the case of the
piranhas be of living animal origin. The frugivorous habit has been described, or hinted at, by workers
from other forested areas. Fruit, seeds and flowers form a component of the diet of Notopterus
notopterus, Paralaubuca typus and Clarias batrachus in the Mekong (Bardach, 1959); Distichodus
atroventralis and D. sexfasciatus of the Congo (Matthes, 1964); Leptobarbus melanotaenia, Puntius
bulu, P. binotatus, P. bramoides, P. sealei, Botrachocephalus mino and Chonerhinus modestus of
North Borneo (Inger and Chin, 1962); Leptobarbus hoeveni of Sumatra (Vaas et al., 1953). Tan
(1980) records Tor tambroides, Acrossocheilus hexagonolepis, Leptobarbus hoeveni, Puntus bulu and
P. daruphani as gathering around Ficus variegata, Eugenia sp., Diptocarpus oblongifolius,
Dypoxylon angustifolium and Elateriospermus tapus to eat the ripe fruit as it falls into the water.
Other species have become adapted to taking organisms from outside the aquatic system, the most
extreme example of this being found in the Archer fish (Toxotes) which shoots water droplets at
insects which settle on overhanging vegetation so as to knock them into the water. Fittkau (1973) has
illustrated the type of simplified nutrient-food-consumer cycles that are found in the mouth lakes of
the Amazonian tributaries (Fig. 6.1). The role of food coming from outside the aquatic system is not
confined to forested rivers, as some species inhabiting savannah plains also rely heavily on this source
(Kelley in FAO/UN, 1968a). Adaptations to this are such that Brycinus spp. have been recorded as
deliberately jumping against the stems of rice plants to bring down seeds for consumption (Matthes,
1977).

169
Figure 6.1 The nutrient cycle in the mouth lake of an Amazonian tributary. (After Fittkau, 1973)
Predation on fish and large invertebrates by other fish is relatively unimportant in small streams.
Indeed the smaller and more unstable a water course, the less likelihood that predation will
significantly affect its population structure. (Moyle and Li, 1979). In some small coldwater streams
more stable conditions allow predation by fish such as trout to play a more significant role. Trout for
instance accounted for 15% of the total mortality of other fish in the water course studies by
Alexander (1979). The relative importance of predatory species tends to increase downstream until
the point where Bayley (1983) found piscivorous predation to account for most of the productivity of
small and medium sized fishes in the forested Amazon. On savannah floodplains there is a tendency
for bottom-feeding forms to dominate by weight, although there are usually only a few species at this
trophic level. Furthermore in some systems decapod Crustacea perform the primary scavenging role
and fish are confined to the higher links of the food chain. In the Potamon community structures
usually contain a very high proportion of predatory species and piscivorous predators are generally
very common. The relative abundance of these elements of the community tends to increase during
the dry season giving the very high predator-forage fish ratio recorded by some authors. For example,
Mago-Leccia (1970) noted that up to 75 percent of the population of some floodplain pools of the
Orinoco consisted of fish eaters such as Hoplias malabaricus. Lowe-McConnell (1964) and Bonetto,
Dioni and Pignalberi (1969) equally commented on the abundance of piscivores and the absence of
small fish in flood pools of the Rupununi and Parana rivers respectively at the end of the dry period. I
have observed the same phenomenon in the Ouémé River and a number of other workers have
commented on it in passing with reference to other African systems. Strangely, prolonged and
stabilized high water in the normally fluctuating Everglade marshes also produced an increase in
predatory species according to Kushlan (1976). This change was due to the migration into the marsh

170
of large predatory species which are normally intolerant of the extreme swamp conditions. Similar
shifts may be anticipated where floodplains are impounded to increase the inundation time.
Lowe-McConnell (1975) concluded that there is a linear succession of dominant food sources in
streams and rivers. Fishes in headwater streams depend mostly on allochthonous foods. As the stream
enlarges grazers and generalized predators feeding on benthic invertebrates become more important.
Finally in the lower reaches, the accumulation of detritus and soft mud supports a number of mud-
eating species although piscivorous predators are also abundant. Nikolsky (1937) described this same
succession from the Amu Darya and Syr Darya and correlated with it the increase in length and
complexity of the guts of fish as one proceeds downstream in these rivers. This progression of feeding
types is in accordance with the predictions of the river continuum concept and also agrees with similar
progressions in other organisms.
SPECIALIZATION AND RESOURCE PARTITIONING Specialization
Feeding habits of individual fish species in rivers have been described by many authors. These have
generally been analysed as trophic “statics” which, by approaching diet in a manner limited in space
or time, tends to assign fish to trophic categories or niches. This has led to the assumption that the
considerable specializations of dentition, jaw structure, body form and alimentary tract, corresponds
to real feeding niches particular to the species. Closer examination of this assumption shows the
situation to be considerably more complex. In stable systems such as reservoir rivers it may be
supposed that specialization leads to the species adopting a fixed position in the community whereas
in fluctuating systems typified by flood rivers specializations may be more valuable in one or other of
the phases of the system. There has been some debate as to whether the advantage accrues during
times of abundance (usually high water) when adequate food of diverse nature is present or at times of
scarcity when specialization would sharpen the competitive capacity for those resources which are
available. In support of the latter hypothesis Zaret and Rand (1971) found trophic overlaps in a
Panamanian stream to be minimal during the dry season, when food abundance was at its lowest. For
example, Astyanax, which normally occupies the surface during the nutrient rich wet season, is forced
into the mid waters by the more specialized Gephyrocharax during the dry season. Here access to its
usual diet is prevented by Roeboides and as a consequence switches from its normal concentration on
allochthonous insects to a purely vegetarian diet. In more temperate waters, Angermeier (1982) also
found an increase in the variety of prey selected by nine species of United States stream fish during
times of scarcity whilst the same species all exploited virtually the same food resources in times of
abundance. Greater specialization has been found during the high water period by several other
authors. Matthes (1964) found greater degrees of specialization during the floods of the Congo River.
Lowe-McConnell (1964 and 1967) also found high water to be the period when fishes are most
segregated trophically in the Rupununi. This has also been illustrated for the frugivorous fishes of the
Amazon by Goulding (1980) where the various species concentrate on particular types of fruit and
seeds during the floods but are less selective during the dry season. In direct contrast to Zaret and
Rand's (1971) findings, Power (1984) established that species in a Panamanian stream concentrate on
the major substrates when food was abundant, but diversified their choice as food became scarce.
The habit of switching diet seasonally, noted by most of these authors, has also been commented on
by others. For example Cabrera et al., (1973) found that the diet of Basilichthys bonariensis in the
Plata system could be separated into three main components: constant elements (algae, mud,
vegetable remains); seasonal elements (cladocerans, copepods, diatoms, malacostraca, gasteropods
and fish all of which appeared mainly in the flood); occasional elements (rotifers and ostracods). This
pattern of feeding, where a basic major trophic category is at the same time flexible enough to take
advantage of other food items as or when they are available seems very common. Even such species
as piranha (Serrasalmus spp.) which are noted for their predatorial ferocity can switch to an
alternative diet of vegetable detritus or mud during the dry season (Mago-Leccia, 1970; Goulding,
1980). Some such species are clearly examples of an adaptation to one food type serving equally for

171
an alternative food, as in the case of mud-eating microphages such as Heterotis, Oreochromis
niloticus or Alestes spp which may adopt a planktonophage habit in lentic waters. Many species select
a succession of food types as the flood season progresses. Typical of such are the Brycinus species
studied by Daget (1952) that changed from a diet of insects and seeds or even higher vegetation
during the rising waters to feeding on phytoplankton as the waters begin to contract. Similarly,
Colossoma bidens or C. macropomum concentrate of allochthonous fruits during the flood in the
Amazon switch to autochthonous planktonic Crustacea at low waters (Honda, 1974). In the savannah
floodplain of the Apure River the same species shows a much greater range of diet including fish,
birds and other animal matter in the range of items eaten.
There is also a tendency for food preferences to change as individuals grow older, and the diet of
juvenile fish often differs widely from that of adults of the same species. Young Prochilodus
platensis, for example, feed on planktonic diatoms and crustacea, whereas the adults are uniquely mud
eaters (Vidal, 1967). Most extreme in this respect are the juveniles of the major piscivorous predators,
such as Lates niloticus or Hydrocynus which eat small Crustacea or even phytoplankton. Plankton
appears to play a very important role in the diet of the youngest fish and many of the movements
described under migration appear designed to bring the fry into contact with this food source.
That fish can respond to the concentration of their preferred food is illustrated by Quiros' findings
with iliophagous fishes in the Parana system. Power (1984) also found that concentrations of
Ancistrus spinosus and other loricariid catfishes in Panamanian streams corresponded to the
productivity of the algae upon which they graze. Here there were 6–7 times more fish in sunlit areas
of stream where algae productivities were up to 7 times faster than in shaded areas. Algal growth is
fastest in shallow waters (20 cm) but fish did not enter these waters because of risks of predators.
However, in other streams in Panama, Angermeier and Karr (1983) found that the distribution of
feeding guilds by biomass was not generally correlated with the availability of their major food
resources.
Resource Partitioning
Knoppel (1970), in his study of the nutrient ecology of 49 species from streams and lakes of the
Amazonian terra firma and floodplain was forced to conclude that there were no specialists in these
habitats thus the resource was virtually unpartitioned. Knoppel's study, however, was limited to a
small forested stream where all species concentrated on allochthonous items as the most abundant
food source and was also limited in season. In other systems it is generally both convenient and
possible to classify fish into broad categories or trophic guilds according to their predominant feeding
habits. For example, Matthes (1964) distinguished the following categories in the Congo River basin:
Mud feeders, which eat finely divided silt together with the microorganisms and organic decay
products it contains. In the floodplain pools this niche is filled by Phractolaemus ansorgii.
Detritus feeders, which ingest mainly vegetable debris, leaf litter and the associated animal
communities: e.g., Stomatorhinus humilior, Clarias buthupogon, Clariallabes variabilis, C.
brevibarbis, C. melas, Channallabes apus.
Omnivores, which are widely represented by all families and most genera in the floodplain water
bodies by Stomatorhinus fuliginosus and Ctenopoma fasciolatum.
Herbivores, which can be further separated into:
(i) microherbivores which eat algae and diatoms
(ii) macroherbivores which eat higher plants

172
These were unrepresented in the floodplain pools, but in adjacent floating prairies Neolebias gracilis,
Distichodus affinis and Synodontis nummifer ate this type of food.
Plankton feeders, which are rare due to the lack of plankton in the riverine environment, but which
are nevertheless represented by Aplocheilichthys myersi in the floating vegetation.
Carnivores, the most important group which subdivide into:
(i) Meso-predators which feed mostly on insects and crustacea, and which are either
(a) feeders on allochthonous matter or neuston, such as Pantodon buchholzi, Ctenopoma
nigropannosum or Ctenopoma ansorgei in the pools
(b) bottom feeders which eat insects and molluscs from the bottom, such as Polypterus
retropinnis, Stomatorhinus polli, Clarias submarginatus, Kribia nana
(c) carnivorous browsers which inhabit floating vegetation and feed on the small insects and
Crustacea found there, for example Xenomystus nigri, Nannocharax schoutedeni, Hemistichodus
mesmaekersi, Heterochromis multidens or Ctenopoma kingsleyae
(ii) Macro-predators
(a) generalized predators which feed on fish or larger invertebrates such as decapod Crustacea or
insect larvae. In floodplain pools Clarias platycephalus feeds in this manner.
(b) piscivorous predators which feed only on fish. Although Matthes did not record any such
species in the floodplain pools, fish such as Parachanna obscurus, Hydrocynus vittatus or Lates
niloticus have been recorded from such habitats elsewhere.
(c) fin nippers which are representative of specialized predators generally.
Similar tabulations have been presented by other authors, for example Marlier (1967) who
investigated the feeding habits of fish in the Lago Redondo of the Amazonian Varzea (Table 6.1) and
Vaas (1953) who classified the fish fauna of the Kapuas river, West Borneo (Table 6.2)
The trophic relationships of river and floodplain communities can be summarized as a generalized
food web of the type shown in Fig. 6.2. Not all elements of this diagram are necessarily present in all
environments. As we have seen the heavy bias towards allochthonous food in the forest environment
and low order streams favours sequences following from this source of nutrition and diminishes the
importance of phytoplankton.
Table 6.1
Fish fauna of the Lago Redondo classified according to their feeding habits (after
Marlier, 1967)
Unspecialized STENOPHAGES Specialized
Carnivores
Serrasalmus nattereri Piscivores Arapaima gigas
Serrasalmus elongatus Boulangerella cuvieri
Eigenmannia virescens Ageneiosus ucayalensis
Pimelodella cristata Symbranchus marmoratus
Plagioscion squamosissimus Cichla ocellaris

173
Geophagus surinamensis Insectivores Triportheus elongatus
Apistogramma taeniatum Oxydoras niger
Colomesus psittacus
(=asellus) Zooplankton-feeders Metynnis hypsauchen
Astyanax fasciatus
Hypophthalmus edentatus
Herbivores
Anodus laticeps Grass seeds Ctenobrycon
hauxvellianus
Cichlasoma bimaculatus Water grasses Metynnis maculatus
Cichlasoma festivum Leporinus maculatus
Fruit Colossoma bidens
Algae and Aufwuchs Poecilobrycon trifasciatus
Poecilobrycon
unifasciatus
Mud-eaters
Curimatus spp.
Prochilodus sp.
Pterygoplichthys multiradiatus
Potamorhina pristigaster
EURYPHAGES
Predominantly carnivores
Osteoglossum bicirrhosum
Serrasalmus rhombeus
Predominantly herbivores
Phytoplankton, zooplankton Anchoviella brevirostris
Grass leaves and seeds, insects Pyrrhulina brevis
Hyphessobrycon rosaceus
Hyphessobrycon callistus
Benthic and epiphytic diatoms and cladocera Hyphessobrycon sp.
Cheirodon piaba
Algae, grass and cladocera Metynnis lippincottianus
Aufwuchs Corydoras sp.
Seeds, molluscs Acarichthys heckeli
Littoral zooplankton and higher plants Pterophyllum scalare
Table 6.2
Fish fauna of the Kapuas river and adjacent lakes and river arms, classified according to their feeding
habits (after Vaas, 1953)
a
b
c
d
e
f
g
h
i
j
k
l
Plankton feeders
Helostona temmincki xx x
Thynnichthys thynnoides xx xx x x
Thynnichthys polylepis xx x x x
Dangila ocellata xx x x x
Dangila festiva xx x x x
Periphyton and vegetable feeders
Amblyrynchichthys truncatus x xx x x
Osteochilus melanopleura x xx x x xx
Osteochilus brevicaudata x xx x x xx
Osteochilus waandersi x xx x x xx

174
Osteochilus vittatus x xx x x xx
Vegetable feeders on submerged higher plants, inundated land
plants, fruits and seeds
Puntius waandersi x xx x
Puntius nini x x x xx x x x
Puntius bulu x xx x
Puntius schwanefeldi x xx x
Leptobarbus hoeveni x xx
Leptobarbus melanotaenia x xx
Pristolepis fasciatus x xx
Osphromenus gourami x x xx x
Omnivores feeding mainly on insects and larvae, zooplankton
Balantiocheilus melanopterus x xx x
Cyclochilus repasson x xx
Luciosoma trinema x x x xx
Rasbora argyrotaenia x x x xx
Rasbora vaillanti x x x xx x x
Eaters of insects at surface
Chela oxygastroides x xx x x
Toxotes chatareus x x x xx x
Omnivorous bottom feeders
Barynotus microlepis x xx x
Pangasius pangasius x x xx
Pangasius polyuranodon x x xx
Mastacembelus armatus fayus x x xx
Mastacembelus argus x x xx
Omnivorous predators
Macrones nigriceps x xx xx x
Macrones nemurus x xx xx x
Hemisilurus chaperi x xx
Hemisilurus scleronema x xx
Predators on small fish and small animals, insects, shrimps
Lycothrissa crocodilus x x xx
Kryptopterus cryptopterus x x xx
Kryptooterus schilbeides x x xx
Kryptopterus limpok x x xx
Kryptopterus micronema x x xx
Macrochirichthys macrochirus x xx xx
Setipinna melanochir x xx
Datnioides microlepis xx xx
Hampala bimaculata x x xx
Large predators eating fish of all sizes, shrimps, prawns and
crabs
Ophicephalus striatus x xx
Ophicephalus micropeltes x xx
Ophicephalus pleurophthalmus x xx
Ophicephalus lucius x xx
Notopterus chitala x xx
Wallago leeri x xx
Silurodes hypopthalmus x xx
Note:
x = additional food;
xx = main food
a Phytoplankton b Periphyton c Filamentous algae d Bottom algae

175
e Submerged plants inundated land plants, fruite seeds f Small zooplankton g Cladocera, copepods and rotifera h Insects and their larvae i Allochthonous insects j Shrimps k Insect larvae in the bottom, worms l Fish, prawns and crabs
When feeding habits are matched with habitats some complex relationships emerge, as is shown by
Matthes (1964) for Lake Tumba and the adjacent forested floodplains of the Ikela region. Table 6.3
illustrates this for the Lower Ouémé river and floodplain during the dry season, where one of the
major primary feeders on detritus are the decapod crustacea Macrobrachium macrobrachion and
Caridina sp. which, because of their size and abundance, are more closely allied to the fish
ecologically than they are to the other invertebrate fauna.
Fish in rivers, however, appear to be highly facultative in their feeding and with few exceptions may
move within the guild structure according to the species composition of the fish community, the time
of year and shifts in the non-biotic components of the ecosystem. Thus the assignment of species to
particular niches may be inappropriate. Indeed evidence from a variety of systems indicates that i) the
same food resource may be shared by numerous different species and ii) the same species may
successively exploit several different resources during the year. The long term persistence of fish
faunas consisting of many tens of species the generalized feeding patterns of the fish, the temporal
succession of dietry items and the consequent overlaps of trophic habit cause conceptual problems in
that some communities seem to violate the Gaussian principle of competitive exclusion. Alternative
interpretations of the trophic niche have therefore been sought. In stable systems such as reservoir
rivers, it is to be supposed that the range of specialization among the fish community leads to a more
or less defined partitioning of the available resources among the various species present and the
assignment of the species to more or less fixed niches. The available evidence would support this
view, as most observations of systems where more or less stable partitioning of the resource, whether
food or space, occurs, are from smaller streams and rivers with more or less stable flow regimes.
Despite the apparently greater stability of such systems there are still reasons to suppose that
considerable flexibility in feeding and spatial niche selection exists. Detailed studies of the cyprinid
communities of small relatively stable streams of Sri Lanka show that even within one family in
similar systems different levels of partitioning may be found. For example Puntius bimaculatus and P.
titteya co-occur but do not overlap in trophic niches (De Silva et al., 1977). Three species in the same
streams show considerable overlap in the choice of food items but avoid direct competition by
different preferred living spaces (De Silva and Kortmulder, 1977) and a further four species overlap in
diet and living space thereby competing directly but apparently co existing without problems (De
Silva et al., 1980). Where close apparent competition exists, investigations by Werner and Hall (1979)
indicate that a species may switch trophic (and spatial) habitats within a range of acceptable or
accessible food items depending on the relative profitability of the individual item. Thus a preferred
item under one community structure may be selected against if a more efficient competitor alters the
profitability balance. Such switches may occur from season to season as in the case of the Astyanax in
Panama (Zaret and Rand, 1971) or from year to year. Year to year changes in the trophic structure of a
community would then depend on the relative abundance of its component species which in turn is
determined by factors other than food availability. Here Schlosser (1982) considered that changes in
temporal reproductive success were more important than competitive exclusion or predation in
determining community organisation. Grossman et al. (1982) reached the similar conclusion that
“random” factors (e.g. environmental variables) rather than deterministic ones were responsible for
the lack of repeatability of community structures in the same Indiana stream over a twelve year
period. The term “Condominium” first suggested by Wynne-Edwards (1962) to describe associations
of nearly related species of similar habitats that can be united ecologically and are not in competition
for resources within the association could well be extended to include these grouping of river fish

176
species which coexist over long periods showing similar feeding, breeding and spatial distribution
patterns. The flexibility in trophic organization among stream fish communities implies that inter-
specific competition is not a major factor in regulating biomass and community structure. This
conclusion is to some extent supported by Bayley (1983) in his observations on Amazonian fishes.
Figure 6.2 Diagram of trophic relationships in a river-floodplain community. Broken line = influence;
Solid line = feeding interaction
Table 6.3
Main dry season habitats of fish species in the Ouémé River ordered by major trophic categories
Main river channel bottom Floodplain pools and lagoons
Trophic
category Surface Mud Sand
Bank
vegetation Surface Bottom
Vegetated
(swampy)
area
Mud and
detritus
feeders
Heterotis
niloticus
Synodontis
schall
Clarias
ebriensis
Heterobranchu
s longifilis
Clarias
ebriensis

177
Citharinus
latus
Labeo
senegalensis
Heterobranchu
s longifilis
Heterotis
niloticus
Neolebias
unifasciatus
Labeo
ogunensis
Auchenoglanis
occidentalis
Synodontis
schall
Phractolaemus
ansorgii
Citharinus
latus
Synodontis
schall
Herbivores
micro
Labeo
senegalensis
Synodontis
nigrita(Juv)
Oreochromi
s galileus
macro Distichodus
rostratus
Distichodus
rostratus
Tilapia
guineensis
Tilapia
guineensis
Zooplankton Pellonula
afzeliusi
Allochthonou
s and neston
feeders
Brycinus
longipinnis
Brycinus
macrolepidotus
Epiplatys
bifasciatus
Epiplatys
sexfasciatus
Omnivores Brycinus
nurse
Marcusenius
brucii
Clarias lazera Synodontis
nigrita
Clarias
lazera
Chrysichthys
auratus
Protopterus
annectens
Chrysichthys
walkeri
Synodontis
melanopteru
s
Synodontis
nigrita
Micropredator
s
Synodontis
sorex
Synodontis
sorex
Chromidotilapi
a guntheri
Chromidotilapi
a guentheri
Ctenopoma
kingslayae
Physailia
pellucida
Petrocephalu
s bane
Brienomyrus
brachyistius
Thysia ansorgii
Barbus
Hyperopisus
occidentalis
Petrocephalu
s bovei
Hemichromis
bimaculatus
callipterus
Pollimyrus
Mormyrus
rume
Cyphamyrus
psittacus
adspersus
Brienomyrus
niger
Eutropiellus
buffei
Pollimyrus
petricolus
Pollimyrus
adspersus
Generalized
predators
Schilbe
mystus
Chrysichthys
nigrodigitatu
s
Protopterus
annectens
Malapterurus
electricus
Calamoichthy
s calabaricus
Eutropius
niloticus
Calaraoichthys
calabaricus
Piscivores Hydrocynu
s forskahlii
Bagrus
docmac
Hemichromis
fasciatus
Hepsetus
odoe
Parachanna
africanus
Hemichromis
fasciatus
Lates
niloticus
Polypterus
senegalus
Polypterus
senegalus
Hydrocynu
s vittatus
Gymnarchus
niloticus
Gymnarchus
niloticus
Hepsetus odoe Parachanna
obscurus

178
Parachanna
obscurus
SEASONALITY OF FEEDING
In temperate rivers and streams the onset of winter usually marks a drop in overall productivity of
aquatic system and in the production of food organisms. Furthermore, the amount of food eaten by
fishes is closely related to temperature thus a general cessation of feeding occurs in most temperate
and arctic species during the winter months. In tropical waters the effects of temperature are clearly
less pronounced but since Chevey and Le Poulain (1940) remarked on the fact that fish did not feed in
the Mekong system during the dry season it has become generally accepted that feeding by fish in
tropical rivers is likewise highly seasonal all over the world. In flood rivers the feeding cycle is
clearly linked to two factors, firstly the food supply and secondly the population density. During the
flood the rapid increase in food organisms, together with the wide dispersal of fish over an extensive
biotope, favours intensive feeding. At low water, when the aquatic environment is contracted the fish
are concentrated in a few permanent reserves of water and food sources are limited or exhausted,
fasting therefore ensues. In the tropics this contrasts with the more or less continuous feeding of fish
in lakes; although in some species inhabiting rivers closely allied to lakes such as the Lake
Chad/Yaeres system the fish cease feeding at low water despite the adequate supply of food which
would enable them to continue feeding at all times of year. In reaches of Indian rivers having little or
no floodplain the seasonality of feeding may be reversed with more intense food intake during the dry
season. Bhatnagar and Karamchandani (1970) attributed this to the food being washed away by the
high current during the flood in the case of Labeo fimbriatus. Tor tor showed a similar pattern to L.
fimbriatus although in this case Desai (1970) correlated the lessened feeding with breeding. It would
seem that feeding stops just before and during breeding in flood and reservoir rivers alike. There are
nevertheless seasonal differences in the availability of food which depend on the morphology of the
river.
The intensive feeding by fish during the periods of abundance permits them to build up large stores of
fat which are sufficient, not only to tide the animals through the following barren winter or dry
season, but to elaborate gonadial tissue in preparation for breeding. Starvation during the winter or dry
season causes fish to lose condition. Daget (1956) for example traced this for Tilapia zillii in the
Niger. Fig.6.3 shows the variation in weight over the dry season using condition factor K (where K =
weight in gms x 105 over total length in mm³) as an index of change. In the river relatively little
change occurred over the dry season until the start of reproduction when there was a sudden reduction
in weight corresponding to about 10.7 percent for the whole period. In a floodplain pool fish ended
the flood in better condition but lost weight more evenly throughout the dry season: in October and
November - fish were fat and full of food (K = 4.52); in December - feeding was reduced (K = 4.68);
January - stomachs empty (K = 4.67); February - very little food (K = 4.39); March - even less food
(K = 4.48); April - only mud (K = 4.34); May (K = 4.16); June (K = 4.2). The net loss in weight over
five months was 11 percent. In a second, somewhat richer, pool the loss in weight was less rapid
(dashed line). Daget (1952) had previously noted similar seasonal changes in weight with Brycinus. In
the Amazon, Junk (in press) observed seasonal changes in the fat content of 40 species. The majority
of these showed a pronounced seasonality in chemical composition with peaks in fat content during
the falling flood and minimum fat content during the breeding season at the end of low water.
The pattern of abundant feeding during the flood and fasting during low water is, perhaps, not as
simple as it appears. Observations by Willoughby and Tweddle (1978) indicated that peak feeding
takes place at different times in different species (Fig. 6.4). The food consumption of Clarias
gariepinus in the Shire system, for instance, reached its maximum just before the flood peak, whereas
Oreochromis mossambicus fed more intensively as the floodplains were draining. A third species,
Clarias ngamensis, fed at a fairly constant rate for most of the year. In all three species food intake
was minimal at low water. There are also indications that certain categories of feeders continue to
feed throughout the dry season. Microvores such as Heterotis niloticus may feed throughout the year,

179
although only at maintenance level during the dry season (Daget, 1957a). Surface feeders also have a
continuing food source and Brycinus macrolepidotus continues to feed long after other species of
Alestes and Brycinus in the Niger. The change in predator-forage fish ratio throughout the dry season,
and the gradual disappearance of smaller fishes from floodplain pools, as noted by Lowe-McConnell
(1964) would suggest that predators may continue feeding well into the dry season. Goulding (1980)
also found that, whereas the majority of frugivorous Amazonian fish greatly reduced their feeding
during the low water seasons, the predatory characins were less inclined to do so in that roughly equal
proportions of such fish contained food in the dry and wet seasons. However, the nature of the food
changed from primarily plant foods (seeds and fruit) in the wet season to fish scales in the dry.
Neverthless, predators appear to stop feeding in other systems and Mago-Leccia (1970), in noting that
Piranhas turn to mud as a feeding substrate in the dry season, also remarked that small fish are not
eaten by predators at low water. Such continued feeding does appear to be somewhat exceptional and
most observers confirm the dry season fast.

180
Figure 6.3 Changes in condition factor between October and July for adult Tilapia zillii in: (A) the
Niger River, and (B) two floodplain pools (after Daget, 1956). Also shown are (C) changes in
condition factor of Brycinus leuciscus (after Daget, 1957a)

181
Figure 6.4 Seasonal variations in daily food intake by three species of fish from the Shire River.
(After Willoughby and Tweddle, 1978)

182
GROWTH
FACTORS AFFECTING GROWTH
Fish from most river systems show well-defined rings or annuli on their scales, bones or otoliths, a
fact which has been noted from tropical and subtropical systems as well as from temperate ones. The
rings of temperate salmonid and coarse fish species have been widely described. Chevey and Le
Poulain (1940) noted rings on the scales of cyprinid species in the Mekong, and rings have been
described from other cyprinids such as Labeo spp. in both the Gambia (Johnels, 1954) and Indian
rivers (Khan and Jhingran, 1975), or Catla catla from the Ganges (Natarajan and Jhingran, 1963).
Numerous characin species have been studied, Prochilodus scrofa (Godoy, 1975), P. platensis is
(Cabrera and Candia, 1964; Vidal, 1967) in South America, and Brycinus leuciscus (Daget, 1952) and
Alestes baremoze (Durand and Loubens, 1969) in Africa. Cichlids from both Africa (Oreochromis
spp., Dudley, 1972) and Latin American rivers (Cichlasoma bimaculatum, Lowe-McConnell, 1964)
have been recorded with annuli on their scales. Johnels (1954) also mentions rings on the scales of
Notopterus sp. and mormyrids. Other hard parts of the fish also show qrowth rings. Opercular bones
were used by Cordiviola (1971) for ageing Prochilodus platensis. The scaleless siluroids (Candia et
al., 1973) have shown clear rings in the otoliths and pectoral spines of Parapimelodus valenciennesi
and Fenerich et al., (1975) have demonstrated their existence in the otoliths of Pimelodus maculatus.
Pectoral spines and vertebrae were used to age Clarias spp. from the Kafue flats by the University of
Idaho et al. (1971). Other growth studies, such as those by Bayley (1983) for 12 species of the
Amazon system, have been based on length frequency analyses without recourse to structures.
The marks or rings have been correlated with the partial or complete cessation of growth during one
or more periods of the year. In the temperate zone these are clearly associated with the winter
cessation of growth. However, in the tropics more rings have often been recorded than would be
expected if ring formation depended solely on a regular seasonal event. Care therefore has to be taken
in interpreting rings in scales or other hard structures as indicators of age or time series. Nevertheless
at low water feeding either stops completely or is seriously reduced in most species. The fish live on
their fat reserves, sometimes losing condition to the point where resorption occurs at the margin of the
scales. Durand and Loubens (1969) made a useful distinction between growth in weight and growth in
length. The latter is a good indicator of long-term change, but as it depends mainly on skeletal
structures it is not so liable to modification during the growth arrest. Growth in weight is as much
through the addition of soft tissues including fat. These fat stores are liable to be rapidly modified
under adverse conditions, as has been shown for Tilapia zillii and Brycinus leuciscus (Fig. 6.3), and as
Durand and Loubens (1970) showed for Alestes baremoze, where the condition factor (K) fell from
1.30 in April to 1.00 in September. Of course care has to be taken in the interpretation of changes in
weight and condition factor, as the development of gonadial tissue and discharge of eggs or milt is
also reflected in these parameters. However, changes in K are more often slow over the whole dry
period, than abrupt at the time of breeding, as would be the case if the discharge of reproductive
products were the sole cause.
Three main reasons have been advanced for the arrest of growth during several months of the year.
(i) temperature
(ii) effects associated with drawdown; and
(iii) reproduction
In the temperate zone, temperature is clearly the dominant feature and ring formation is correlated
with winter but the growth rate arrest coincides with a drop in temperature. In several tropical river
systems too, for instance, in the Lake Chad basin, winter temperatures drop at least 8°C below the
summer maxima (Durand and Loubens, 1969) and coincide with the minimum growth rate. Similarly
in the Senegal, Reizer (1974) considered the slower growth from December to February to be
correlated with lower temperatures, and in both the lower La Plata system and on the Kafue flats, the

183
minimum rate of growth occurs at the same time as similar drops in temperatures. In most of these
rivers, however, low water coincides with the winter and it is difficult to distinguish the effects of the
two factors. In the southern Okavango swamps the floods arrive during the colder part of the year
(Fox, 1976) which leads to very low growth rates of the fish living there. It is not yet clear when the
growth arrest occurs in these waters, although current work may shed some light on this.
In equatorial rivers growth checks occur regularly where there are only minimal changes in
temperature. In such circumstances Lowe-McConnell (1964) has suggested that crowding and
lessened availability of food, brought about by drawdown conditions, are responsible. That this too
cannot be the whole answer is shown by the Lake Chad fishes which stop growing in the lake even
though there is abundant food. In the Niger river (Daget, 1957) reported that growth arrests can be
distinguished on the scales of carnivores, herbivores, limnivores, insectivores and plankton-eating
species, despite the fact that the predators and limnivores at least have sufficient food. Furthermore,
some fish such as the young of Oreochromis and Tilapia in the Kafue, resume feeding before the
onset of the floods, at a time when conditions are at their most cramped (Dudley, 1974).
The effects of population density on growth rate are somewhat problematic. In the Danube,
Chitravadivelu (1974) was unable to detect changes in the growth rate of Alburnus alburnus and
Rutilus rutilus, despite great differences in biomass and population density from one year to another.
However, Frank (1959) did record increases in growth in Rutilus rutilus and Abramis brama when the
population decreased from 69 124 i/ha to 19 394 i/ha in an Elbe oxbow. This he traced to the greater
availability of planktonic food following the decline in competition for this food source. Clear
relationships between population density and the growth rate and the total production of bullhead
(Cottus gobio) have also been demonstrated by Edwards and Brooker (1982), (Fig. 6.5) from
tributaries of the river Wye. Similar experiments with brown trout show a pronounced drop in
individual growth rate as the number of fish present per unit area increases (Backiel and Le Cren,
1978). Such conflicting results indicate a need for further investigation into the question of the
relationship of food supply to population density, as this influences the amount of fish available to the
fishery.

184
Figure 6.5 Relationships between density and (A) growth, and (B) production in bullhead (Cottus
gobio) in tributaries of the Upper Wye. (From Edwards and Brooker, 1982)
The elaboration of gonadial products during the fasting period probably accelerates the depletion of
fat reserves and exaggerates the low physical condition which is reflected as an annulus. It is,
however, doubtful whether this is the prime reason for growth checks, as maturation is frequently
preceded by a long period of negligible feeding in many species. Furthermore, rings are laid down by
immature fish as well as adults. That there may be deep physiological rhythms which dictate the
seasonal cessation of growth is suggested by an isolated experiment quoted by Johnels (1954). Here
some Barbus gambiensis, which had been transported to Sweden and were being maintained in the
even conditions of an aquarium, still stopped growing and laid down scale rings at precisely those
times when their congeners did so in the Gambia river.
MODELS OF GROWTH
Annuli on scales and other hard parts have been used to calculate growth in many species.
Supplementary information and independent estimates of growth have also been made from the
analysis of length frequency distributions for the progression of individual age groups and also from
the growth of tagged fish. Comparisons between the various methods of age determination show that
they give good agreement at least in sane species (Rao and Rao, 1972; Gupta and Jhingran, 1973).
Several workers have used the Von Bertalanffy model of growth:

185
in which length at time t + 1(Lt+1) is a function of length at time t (Lt) according to the Ford-Walford
equation:
where K is the coefficient of growth and Lo the theoretical asymptotic length achievable by the
species if it grows for an infinite period of time.
Most species seem to conform well to this model in respect of growth in length, or when subject to an
appropriate conversion factor, in respect of growth in weight, as is shown by examples in Table 6.4.
Table 6.4
Representative Von Bertalanffy relationships for growth in length of fish from some tropical river
systems
Species Sex Growth equation Author Alestes baremoze ♂ Lt=237.8[1-e-0.8163(t-0.57)] Durand and Loubens, 1969
♀ Lt=267[1-e)-0.7172(t-0.52)] Durand and Loubens, 1969
Catla catla ♂ + ♀ Lt=1275[1-e)-0.28(t-0.11)] Natarajan and Jhingran,1975
Labeo rohita ♂ + ♀ Lt=1015[1-e)-0.276(t-0.333)] Khan and Jhingran, 1975
Labeo calhasu (Ganga river) ♂ + ♀ Lt=1028[1-e)-0.15(t-0.19)] Gupta and Jhingran, 1973
(Godavari river) ♂ + ♀ Lt=944[1-e)-0.14(t+0.86)] Rao and Rao, 1972
Parapimelodus valenciennesi ♂ + ♀ Lt=333[1-e)-0.14(t-2.4)] Candia et al., 1973
Pimelodus maculatus ♂ Lt=45.4[1-e)-0.2104(t-0.61)] Fenerich et al., 1975
♀ Lt=56.5[1-e)-0.1938(t+0.36)] Fenerich et al., 1975
Prochilodus reticulatus ♂ + ♀ Lt=41.0[1-e-0.20(t-0.35)] Espinosa and Gimenez, 1974
From the table it may be seen that male and female fish of the same species frequently have different
rates of growth and also maximum sizes as indicated by L∞. Growth curves of some representative
species from an African river (Niger) and the Latin American La Plata system shows part of the range
of interspecific variation (Fig. 6.6). Most species grow very rapidly in their first season, a feature
which Dowe-McConnell (1967) regarded as adaptive. Predation is intense in floodplain rivers, so
rapid growth to get to a size too large to be swallowed before the shelter of the floating vegetation on
the floodplain disappears, is a great advantage. Fish probably also need to attain an adequate size to
migrate by the time the floods recede. There seems to be little difference in the size attained by year 1
in fish having a wide range of maximum sizes as shown by Merona (1983) in his examination of over
100 African species.

186
Figure 6.6 Growth in length of representative fishes from rivers: (A) Niger: (i) Heterotis niloticus; (ii)
Lates niloticus; (iii) Mormyrops deliciosus; (iv) Citharinus citharus; (v) Eutropius niloticus (B)
La Plata: (i) Prochilodus platensis (after Cordiviola de Yuan, 1971); (ii) Pimelodus maculatus
(after Fenerich et al., 1975); (iii) Parapimelodus valenciennesi (after Cabrera et al., 1973)
Whereas the Von Bertalanffy growth curve adequately describes year to year progression in length,
growth within any one year does not conform to the model. The long period in which growth either
ceases completely or is considerably restricted, means that most of the year's increase in length occurs
during a comparatively short period. Dudley (1972), for instance, recorded that 75 percent of the

187
expected first year’s growth of Oreochromis andersoni and O. macrochir took place within six weeks
of peak floods in the Kafue river. Growth in weight is even more subject to seasonal variation, often
with temporary losses occurring during the dry season.
Because the within-the-year growth pattern has important implications for the estimation of biological
production, Daget and Ecoutin (1976) have produced a modified growth model applicable to species
with prolonged annual growth arrests. This requires the introduction of two new parameters into the
growth equation. These are q, which represents the duration of the annual growth arrest in months,
and t, which is the duration of the first period of growth also in months. The parameter t is necessary
where reproduction does not coincide with the end of the parent's period of arrested growth. When the
growing period is 12 - q months, the normal Von Bertalanffy curve is expressed as:
Lt = L∞ {1 exp [-g' (t-to)]}
where t and tO are expressed in months and g' = g/12-q; tO is obtained from the equation
The arc of the growth curve thus obtained is thereby compressed into 12-q months and is followed by
a horizontal line q months long. Daget and Ecoutin applied this model to Polypterus senegalus from
the Middle Niger obtaining the mean growth curve shown in Fig. 6.7. A similar modification of Von
Bertalanffy's model was proposed by Cloern and Nichols (1978) to take into account seasonal
variations in growth rate in temperate waters. This model:
where L is length at time (t), L max is maximum body size, L min is body size at time of recruitment
obtains growth predictions with a much smoother transition from growth to arrest phases.

188
Figure 6.7 Mean linear growth of Polypterus senegalus for the first five years of life assuming an
annual growth arrest of six months and a first year's growth period of seven months (T0 = -3).
(After Daget and Ecoutin, 1976)
These models describe situations where growth stops completely during the dry season and have the
advantage of being based on current growth models, but are somewhat inflexible when applied to
situations where growth varies from year to year depending on favourable or unfavourable conditions.
Such conditions require a growth trajectory that is less pre-determined, and Welcomme and Hagborg
(1977) had to adopt a different model for growth within the year to allow for this. Their formula: has
the characteristics of fast initial increase in length followed by a period of slower growth (but not a
complete halt). Values of Lt for successive years can conform to the Von Bertalanffy relationship,
although the form of the curve within one year calculated for successive weeks does not. The
advantage of this relationship in modelling the growth of fish living on floodplains is that the terminal
value of the year’s growth can change according to the intensity of flooding by the operation of an
appropriate coefficient on G.
Lt+t1 = Lt + G exp(t1)
In his studies on fish production in the Kafue river, Kapetsky (1974 and 1974a) was presented with a
similar problem of modelling within year growth patterns. From his own observations as well as those
of Dudley (1972) it was obvious that growth in weight of Sarotherodon and Oreochromis spp. on the
Kafue flats does not stop completely in the dry season, although it is considerably slowed. Kapetsky,
therefore, proposed to rotate the relationship Wt = WO exp (Gt) on its diagonal and then reverse it.
This gives an equation of the form:
Wt = WO + W1 [1 - exp(-gt)]
where W1 = W0 exp G(12). Here W1 = the weight at the end of a year's growth, whereas individual
segments of t and the growth coefficient g are in months.

189
YEAR-TO-YEAR VARIATIONS IN GROWTH
There appear to be few studies on interannual variation in growth rate of fish from the rhithron and
from low order streams of the temperate zone. It may be assumed that, given the relatively stable
conditions of such environments between years, similar rates of growth are obtained. However, there
are indications that density dependant factors may influence the rate of growth in some species,
although the origin of the fluctuations is not clear.
From the studies of growth of fish species inhabiting the potamon it has become obvious that there are
considerable year-to-year variations in growth within the same species. The most detailed
examination of the possible causes of such variations has been carried out for some cichlid species in
the Kafue River. Here, Dudley (1972 and 1974) and Kapetsky (1974) found significant correlations
between some physical variables and the main growth increment. The intensity and duration of
flooding particularly could have accounted for much of the year-to-year variation in the growth of
year class I and II, Tilapia rendalli, Oreochromis andersoni and O. macrochir. Low temperature in
the dry season also appeared to influence some year classes and gave good partial correlations when
entered into the equation after some measure of flood intensity. Typical relationships are shown in
Table 6.5 where TI is an index of temperature, FI is an index of flooding drawn from the area under
the flood curve (Dudley 1972), and HI 2 and HI 3 are indices summarizing the degree of drawdown in
the dry season (Kapetsky, 1974).
Kapetsky's regression equations were successfully used to predict growth increments for certain year
classes, but the consistency of the results is not uniform, possibly due to the short time series upon
which the calculations were based. They are sufficient, however, to indicate the importance of
external physical factors in determining the growth of fish in such systems. This work on the Kafue is
not isolated. As early as 1934 Wimpenny (quoted in Holden, 1963) found that the yield of fish from
the Nile delta, Lake Manzala, was correlated with flood level, high floods being followed by better
than average yields which were due in part to higher growth rates of first year fish. Similarly,
conditions for feeding, and hence growth, of non-anadromous fishes in the Amur river are
considerably improved in years when there is plenty of water (Krykhtin, 1972, quoted by Krykhtin,
1975). The exceptionally poor flood years during the 1968–74 Sahelian drought provided an
opportunity to assess the effects of this on the growth of fish species in the Senegal, Niger and Logone
rivers. In the Senegal, Reizer (1974), discerned great differences in growth of Citharinus citharus
between 1968, a year of particularly poor flood, and other years (Fig. 6.8). The first year class was
missing totally for that year. The second year growth increment for the 1967 year class in the 1968
flood was 3.91 cm, whereas the 1966 year class grew 7.99 cm during the 1967 flood. The third year's
growth showed similar differences; an increment of 2.32 cm for the 1966 year class (1968 flood) and
8.37 cm for the 1967 year class (1969 flood). Differences in growth were also noted from the Niger
where the floods of 1971 and 1972 were particularly bad. Here Dansoko (1975) and Dansoko et al.
(1976) studied two species of Hydrocynus, H. brevis and H. forskahlii, and found that growth,
particularly of the young of the year in both species, was poor during these two years. Hydrocynus
forskahlii, which only inhabits the river, showed this effect less than H. brevis which depends much
on the floodplain for feeding, but nonetheless the differences were still marked. It is also of interest
that year classes with poor first year growth appear to continue to grow badly despite better conditions
in later years. Likewise year classes with good initial growth do not suffer so badly in poor years. In
the Logone Benech and Quensiere (1984) were also able to demonstrate improved growth, as
represented by the mean weight of fish leaving the Yaeres floodplain through the El Beid, as a
correlate of the Logone flood in several species including Hyperopisus bebe, Brachysynodontis
batensoda, Marcusenius cyprinoides, Oreochromis aureus and O. niloticus.
Table 6.5
Parameter estimates for simple and multiple linear regressions of first and second year

190
growth increments on temperature and hydrological indices
Species Year of growth Sex Model r
Oreochromis andersoni 1 M Growth (cm) = 0.02FI+12.87 a 0.92
1 M TL (mm) = 146.51–0.11(HI2) b 0.94
1 F Growth (cm) = 0.014FI+13.4 a 0.78
2 M TL (mm) = -29.47+1.98 (TI) b 0.90
2 F TL (mm) = 38.24–0.30(HI3)+0.83(TI) b 0.93
Oreochromis macrochir 1 M Growth (cm) = 0.2FI+11.02 a 0.9
1 M TL (mm) = 130.39–0.13(HI2) b 0.92
1 F TL (mm) = 130.13–0.32(HI2) b 0.85
2 M TL (mm) = 74.72.0.10(HI3) b 0.58
2 F TL (mm) = 14.69–0.18(HI3) b 0.95
Tilapia rendalli 1 M Growth (cm) = 0.029FI+12.8 a 0.80
a Dudley, 1972 b Kapetsky, 1974
Figure 6.8 Growth of Citharinus citharus in the Senegal River: 1966, 1967, 1969 year classes.
Numbers in parentheses = total length at growth arrest in centimetres. (After Reizer, 1974)

191
REPRODUCTION
SPAWNING SITES AND REPRODUCTIVE ADAPTATIONS
Fish inhabiting rivers show a diversity of reproductive habit which adapts them to the varying
conditions encountered along the length of the river and to the particular difficulties inherent in
breeding in systems with rapidly fluctuating water levels and often extreme conditions of flow or
oxygen deficiency. It seems that physical and behavioural specializations for reproduction are more
varied than those for feeding in these ecosystems. The range of adaptations is indicated by the fact
that nearly all of Balon's reproductive guilds (Balon 1975 and 1981) are represented in the various
rivers of the world. These guilds which form a useful ecological classification of breeding behaviour,
localities and substrates are listed in Table 6.6 together with some representative taxa. Several broad
reproductive strategies have evolved within these guilds which are summarized in Table 6.7.
Table 6.6
Classification of reproductive guilds of fishes with representative taxa from river systems (from
Balon, 1981)
Ethological section - A. Nonguarders
A.1 Open substratum spawners
(Selected key features of early ontogeny)
Pelagic spawners (pelagophils)
Numerous buoyant eggs, none or poorly developed embryonic respiratory organs, little pigment, no
photophobia. Ctenopoma muriei, Lates niloticus.
Rock and gravel spawners with pelagic larvae (lithopelagophils)
Adhesive chorion at first, some eggs soon buoyant, after hatching free embryos pelagic by positive
buoyancy or active movement, no photophobia, limited embryonic respiratory structures.
Prochilodus spp.
Rock and gravel spawners with benthic larvae (lithophils)
Early hatched embryo photophobic, hide under stones, moderately developed embryonic respiratory
structures, pigment appears late. Many cyprinid and characin spp., Barbus and Labeo.
Nonobligatory plant spawners (phytolithophils)
Adhesive eggs on submerged items, late hatching, cement glands in free embryos, photophobic,
moderately developed respiratory structures. Many cyprinids and Rutilus rutilus.
Obligatory plant spawners (phytophils)
Adhesive egg envelope sticks to submerged live or dead plants, late hatching, cement glands, not
photophobic, extremely well developed embryonic respiratory structures. Many cyprinid, characin
and siluroid spp., Puntius gonionotus.
Sand spawners (psammophils)

192
Adhesive eggs in running water on sand or fine roots over sand, free embryos without cement
glands, phototropic, feebly developed respiratory structures, large pectorals, large neuromast rods
(cupulae). Many migration cyprinid and characin spp.
Terrestrial spawners (aerophils)
Small adhesive eggs scattered out of water in damp sod, not photophobic, moderately developed
respiratory structures. Brycon petrosus.
A.2 Brood hiders
Beach spawners (aeropsammophils)
Spawning above the waterline of high tides, zygotes in damp sand hatch upon vibration of waves,
pelagic afterwards. Not represented.
Annual fishes (xerophils)
In cleavage phase blastomeres disperse and rest in first facultative diapause, two more resting
intervals obligate - eggs and embryos capable of survival for many months in dry mud.
Nothobranchius.
Rock and gravel spawners (lithophils)
Zygotes buried in gravel despressions called redds or in rock interstices, large and dense yolk,
extensive respiratory plexuses for exogenous and carotenoids for endogenous respiration, early
hatched free embryos photophobic, large emerging alevins. Many salmonid species.
Cave spawners (speleophils)
A few large adhesive eggs, must hide in crevices, extensive embryonic respiratory structures, large
emerging larvae.
Spawners in live invertebrates (ostracophils)
Zygotes deposited via female's ovipositor in body cavities of mussels, crabs, ascidians or sponges,
large dense yolk, lobes or spines and photophobia to prevent expulsion of free embryos, large
embryonic respiratory plexuses and carotenoids, probable biochemical mechanism for
immunosuppression. Rhodeus sericeus.
Ethological section - B. Guarders
B.1 Substrate choosers
Pelagic spawners (pelagophils)
Nonadhesive, positively buoyant eggs, guarded at the surface of hypoxic waters, extensive
embryonic respiratory structures. Some Ophicephalus and Anabas spp.
Above water spawners (aerophils)
Adhesive eggs, embryos with cement glands, male in water splashes the clutch periodically.
Copeina arnoldi.

193
Rock spawners (lithophils)
Strongly adhesive eggs, oval or cylindrical, attached at one pole by fibres in clusters, most have
pelagic free embryos and larvae. Loricaria parva, L. macrops and some small cichlids.
Plant spawners (phytophils)
Adhesive eggs attach to variety of. aquatic plants, free embryos without coment glands swim
instantly after prolonged embryonic period. Polypterus spp.
B.2 Nest spawners
Froth nesters (aphrophils)
Eggs deposited in a cluster of mucous bubbles, embryos with cement glands and well developed
respiratory structures. Hepsetus odoe, Hoplosternum, some anabantids.
Miscellaneous substrate and material nesters (polyphils)
Adhesive eggs attached singly or in clusters on any available substratum, dense yolk with high
carotenoid contents, embryonic respiratory structures well developed feeding of young on parental
mucus common. Notopterus chitala, Hoplias malabaricus.
Rock and gravel nesters (lithophils)
Eggs in spherical or elliptical envelopes always adhesive, free embryos photophobic or with cement
glands swing tail-up in respiratory motions, moderate to well developed embryonic respiratory
structures, many young feed first on the mucus of parents. Aequidens and other cichlids, some
characins, e.g., Leporinus.
Gluemaking nesters(ariadnophils)
Male guards intensively eggs deposited in next bind together by a viscid thread spinned from a
kidney secretion, eggs and embryos ventilated by male in spite of well developed respiratory
structures. Gasterosteus aculeatus
Plant material nesters (phytophils)
Adhesive eggs attached to plants, free embryos hang on plants by cement glands, respiratory
structure well developed in embryos assisted by fanning parents. Clarias batrachus.
Sand nesters (psammophils)
Thick adhesive chorion with sand grains gradually washed off or bouncing buoyant eggs, free
embryo leans on large pectorals, embryonic respiratory structures feebly developed. Tilapia spp.
Hole nesters (speleophils)
At least two modes prevail in this guild: cavity roof top nesters have moderately developed
embryonic respiratory structures, while bottom burrow nesters have such structures developed
strongly. Several cichlids.
Anemone nesters (actiniariophils)

194
Adhesive eggs in cluster guarded at the base of sea anemone, parent coats the eggs with mucus
against nematocysts, free embryo phototropic, planktonic, early juveniles select host anemone. Not
represented.
Ethological section - C. Bearers
C.1 External bearers
Transfer brooders
Eggs carried for some time before deposition; in cupped pelvic fins, in a cluster hanging from
genital pore, inside the body cavity (earlier ovi ovoviviparous), after deposition most similar to
nonguarding phytophils. Callichthys, Corydoras.
Auxiliary brooders
Adhesive eggs carried in clusters or balls on the spongy skin of ventrum, back, under pectoral fins
or on a hook in the superoccipital region, or encircled within coils of female's body, embryonic
respiratory circulation and pigments well developed. Loricaria spp.
Mouth brooders
Eggs incubated in buccal cavity after internal, external synchronous or asynchronous, or buccal
fertilization assisted by egg dummies, large spherical or oval eggs with dense yolk are rotated
(churning) in the cavity or densely packed when well developed embryonic respiratory structures
had to be assisted by endogenous oxydative metabolism of carotenoids, large young released. Many
cichlids, Osteoglossum.
Gill-chamber brooders
Eggs of North American cave fishes are incubated in gill cavities.
Pouch brooders
Eggs incubated in an external marsupium: an enlarged and everted lower lip, fin pouch, or
membraneous or boy plate covered ventral pouch, well developed embryonic respiratory structures
and pigments, low number of zygotes. Loricaria vetula and L. anus.
C.2 Internal bearers
Facultative internal bearers
Eggs are sometimes fertilized internally by accident via close apposition of gonopores in normally
oviparous fishes, and may be retained within the female's reproductive system to complete some of
the early stages of embryonic development, rarely beyond the cleavage phase; weight decreases
during embryonic development. Rivulus marmoratus, Oryzias latipes, Pantodon buchholzi.
Obligate lecithotrophic livebearers
Eggs fertilized internally, incubate in the reproductive system of female until the end of embryonic
phase or beyond, no maternal-embryonic nutrient transfer; as in oviparous fishes yolk is the sole
source of nourishment and most of the respiratory needs; some specialization for intrauterine
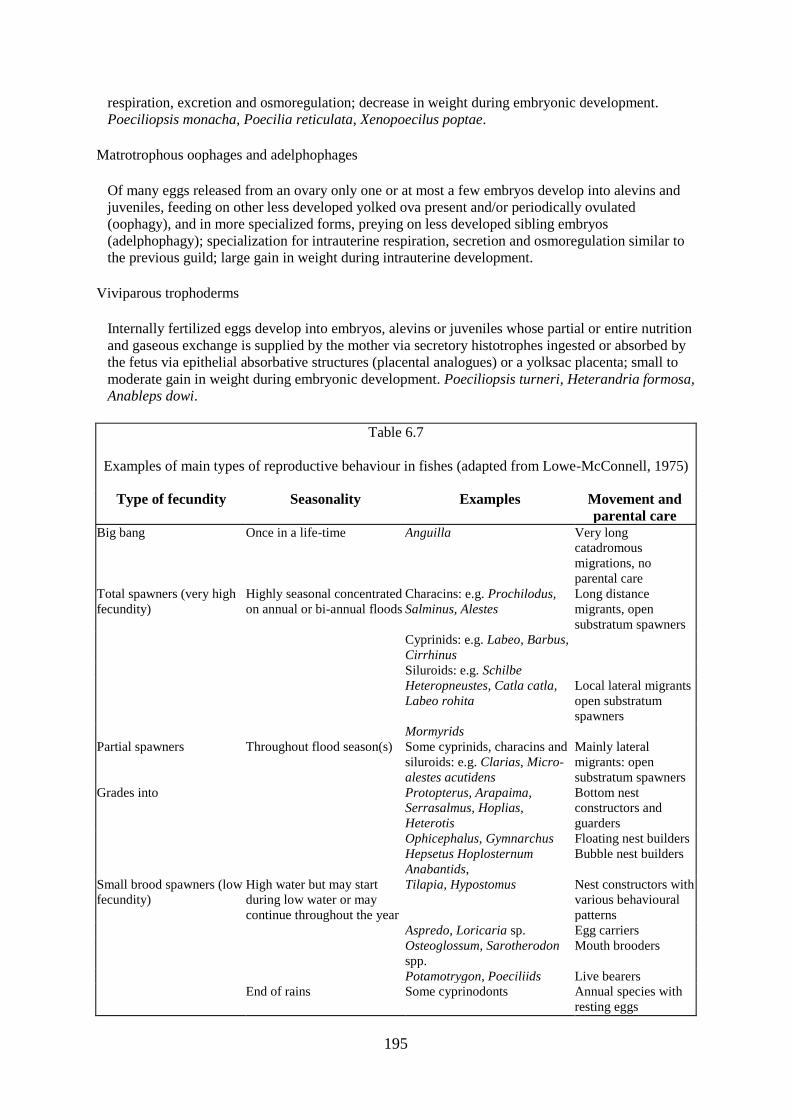
195
respiration, excretion and osmoregulation; decrease in weight during embryonic development.
Poeciliopsis monacha, Poecilia reticulata, Xenopoecilus poptae.
Matrotrophous oophages and adelphophages
Of many eggs released from an ovary only one or at most a few embryos develop into alevins and
juveniles, feeding on other less developed yolked ova present and/or periodically ovulated
(oophagy), and in more specialized forms, preying on less developed sibling embryos
(adelphophagy); specialization for intrauterine respiration, secretion and osmoregulation similar to
the previous guild; large gain in weight during intrauterine development.
Viviparous trophoderms
Internally fertilized eggs develop into embryos, alevins or juveniles whose partial or entire nutrition
and gaseous exchange is supplied by the mother via secretory histotrophes ingested or absorbed by
the fetus via epithelial absorbative structures (placental analogues) or a yolksac placenta; small to
moderate gain in weight during embryonic development. Poeciliopsis turneri, Heterandria formosa,
Anableps dowi.
Table 6.7
Examples of main types of reproductive behaviour in fishes (adapted from Lowe-McConnell, 1975)
Type of fecundity Seasonality Examples Movement and
parental care Big bang Once in a life-time Anguilla Very long
catadromous
migrations, no
parental care
Total spawners (very high
fecundity)
Highly seasonal concentrated
on annual or bi-annual floods
Characins: e.g. Prochilodus,
Salminus, Alestes
Long distance
migrants, open
substratum spawners
Cyprinids: e.g. Labeo, Barbus,
Cirrhinus
Siluroids: e.g. Schilbe
Heteropneustes, Catla catla,
Labeo rohita
Local lateral migrants
open substratum
spawners
Mormyrids
Partial spawners Throughout flood season(s) Some cyprinids, characins and
siluroids: e.g. Clarias, Micro-
alestes acutidens
Mainly lateral
migrants: open
substratum spawners
Grades into Protopterus, Arapaima,
Serrasalmus, Hoplias,
Heterotis
Bottom nest
constructors and
guarders
Ophicephalus, Gymnarchus Floating nest builders
Hepsetus Hoplosternum
Anabantids,
Bubble nest builders
Small brood spawners (low
fecundity)
High water but may start
during low water or may
continue throughout the year
Tilapia, Hypostomus Nest constructors with
various behavioural
patterns
Aspredo, Loricaria sp. Egg carriers
Osteoglossum, Sarotherodon
spp.
Mouth brooders
Potamotrygon, Poeciliids Live bearers
End of rains Some cyprinodonts Annual species with
resting eggs

196
Fish spawning within rhithronic reaches of the main channel usually have to contend with high flow
and turbulence. Consequently they tend either to have adhesive eggs which stick to rocks and plants
(lithophils or phytophils) or to place the eggs in crevices of the rocky substrates of the riffles
(lithophils). Some salmonids for instance cut “redds” into fine gravel which are designed both to
protect and aerate the eggs. Other species spawning in small streams may lay their eggs in very
shallow water or in damp terrestrial environments (aerophils). For example, the group spawning
Brycon petrosus is described by Kramer (1978) as lodging its eggs in the splash zone at the margins
of Panamanian streams. Several other species including Fundulus similis, F. heteroclitus and
Hypomesus pretiosus also have similar behaviour. Other fish from the Amazon, Madeira, Orinoco,
Danube and Amur as well as presumably many other rivers which spawn in the main channel have
semi-buoyant eggs (lithopelagophils) which drift downstream with the current until finding suitable
feeding grounds usually on floodplains or backwaters.
Many species migrate considerable distances upstream in order to spawn in the rhithronic lower order
streams where presumably the good aeration, rich food supply of the riffles and relative freedom from
predation all favour the development of the fry. To compensate for the high risks inherent in these
environments however nearly all species spawning in such regions produce large numbers of eggs and
are total spawners. That upstream migrants do not all seek the same type of headwater is shown by the
observations made by INDERENA (1973) on the Magdalena river. Here Brycon moorei moved into
small short side arms off the main channel, whereas Prochilodus reticulatus stayed within the main
channel, Salminus affinis swam up side streams to areas of high flow and Brycon henni ascended to
the highest accessible reaches of the main river.
In Europe breeding success of some species appears mainly dependent on the presence of suitable
substrates. Even under considerably modified hydrological regimes species such as Rutilus rutilus or
Abramis brama have shown themselves capable of changing from the type of migratory behaviour
they show in the Volga or Lower Danube to their more static behaviour in Western European
modified streams provided the required substrates are available. Other species, such as Chandrostoma
nasus, Barbus barbus or Leuciscus idus have shown themselves much less able to so adapt.
Migratory species may alter their reproductive strategy to adapt to different climatic conditions. For
instance Alosa sapidissima, an anadromous species which enters rivers on the East coast of the United
States to spawn, breeds more frequently, but releases few eggs per breeding in the cooler northern
rivers than in the warmer southern rivers (Legget and Carscadden, 1978). This means that in northern
rivers, more of the available energy is allotted to migration in order to place eggs and fry in the most
favourable conditions. Legget and Carscadden add that available literature suggests that many other
species are equally capable of fine tuning their reproductive strategies to accord with local
environmental circumstances. Such behaviour would mean that populations specific to certain rivers
would develop and that homing capabilities would be needed to ensure that the behavioural patterns
would match the climatic conditions of the spawning environment.
The most diverse breeding strategies are found in those fish spawning on the floodplains, and many
species migrate considerable distances up or downstream to breed so as to place their fry in these
productive habitats. Utilization of the floodplain for spawning is in fact recorded from most tropical
river systems and is also common in such temperate rivers as still have plains. Thus the various
authors writing on larval fish distribution in the Mississippi-Missouri system (e.g. Kallemyn and
Novotny, 1977) state that the floodable marsh areas of the backwaters are vital for the spawning of
many species. Similar observations have also been made in the Danube (e.g., Holcik and Bastl, 1976)
and the Volga (Koblitskaya, 1985).
Migratory blackfishes show a wide range of behaviour. Most are open stratum phytolithophils or
phytophils which attach or scatter their eggs among vegetation or over open bottoms of the floodplain.
Often the localities chosen for egg deposition are in the channels conducting water on to the plain as
in the Alestes species of the Logone river which lay their eggs in the channels feeding the Yaeres

197
floodplain while the adults themselves never penetrate the plain. Alternatively species migrating on to
the floodplains of the Ayuthaya province of Thailand (Chao Phrya river system) illustrate the
selection of substrates for spawning (Tongsanga and Kessunchai, 1966). Crossocheilus reba spawns
in the inundated rice fields as does Wallago attu. Another catfish, Pangasius sutchi scatters its eggs
among submerged weeds and bushes in the shallow inundated margins of the Klongs or canals. The
spring eel Macrognathus aculeatus spawns both in flooded rice fields and in the channels on clear
bottoms or among weeds. Some species, such as Puntius goniotus drop their eggs in the middle of
channels and presumably rely on currents to wash them into suitable nursery sites.
Closely related species and genera also show contrasting behaviour. For example, Hydrocynus brevis
breed on the floodplain of the Niger during the floods whereas the morphologically similar H.
forskahlii breeds in the main river channel during low water. Jackson and Coetzee (1982) have
remarked on the habits of S. African Labeo umbratus which move on to flooded grasslands to lay
their sticky eggs among the temporarily submerged vegetation as contrasted to the local Barbus
johnstoni and B. trapilepidotus which both spawn in gravel areas of swift flowing rivers. Their
conclusion that these behaviour patterns can be enlarged to make a fundamental distinction between
the two genera does not, however, stand up to closer inspection as some Labeo species, e.g. Labeo
parvus of the Niger River which inhabit rapids breed in the riffle zones of the main channel and some
small Barbus equally breed in the flooded grass of the floodplain.
Most blackfish species which at the most are only local migrants have some form of parental care and
in some genera a variety of behavioural patterns are found. This is typified by the genus Loricaria,
where L. parva and L. macrops sit on eggs which are attached to cleared areas of rock. The eggs are
cleaned and fanned by the parents from time to time. Female L. piracicalae develop a special spongy
skin on their ventral surface. The fish roll on the fertilized eggs which adhere to this region. L. vetula
and L. anus have a pouch formed by the enlargement of the lower lip of the males in to which the
eggs are lodged, and in Loricaria sp. (near microdon: Lowe-McConnell, 1964) the lower lip is
extended into a special elongation from which the young are suspended. Other loricariids such as the
Panamanian L. uracantha use hollows and cavities in pieces of wood as nesting sites (Moodie and
Power, 1982).
Nest building is very common among floodplain species. Arapaima gigas scoops out hollows in the
bottom of the flooded savannah and both parents .guard the eggs that are deposited (Sanchez, 1961).
Various species of cichlid make nests. Some of these are simply cleared areas of rock, others holes
under rocks or vegetation, and in yet others, excavated pits sometimes of complex design. Heterotis
niloticus, construct nests in the middle of masses of vegetation at the shallow margins of the plain,
whereas Gymnarchus niloticus make floating flask or raft shape masses of aquatic weeds in which
they lay their eggs (Svensson, 1933). The different architecture of the nests of Protopterus has been
described by Johnels and Svensson (1954) for P. annectens and Greenwood (1958) for P. aethiopicus.
These range from simple sunken areas in a sandy substrate to complex domed structures of papyrus
roots. The characin Hoplias malabaricus constructs a simple nest on the bottom with whatever
material is readily available and some species of Serrasalmus are also reputed to guard egg masses
laid on tree roots, and aquarium observations have shown them to excavate nests in plant masses
(Braker, 1963).
Floating nests are found in some species. Primitive types are made by some ophicephalids.
Ophicephalus micropeltes allows its pelagic eggs to float to the surface in a cluster, and then
surrounds them with a ring of bits bitten off surrounding vegetation (Tongsanga and Kessunchai,
1966). The floating mass is guarded by the male. Hepsetus odoe is unequal amongst the characins in
constructing a nest of foam which is lodged between the stems of weeds or grasses at the margins of
the plain. Froth or foam nests made of mucus secretions from one or both parents are common in
other families. Among the siluroids Callichthys callichthys and Hoplosternum littorale both make a
raft constructed of bubbles and aquatic plants. Floating nests are made by many anabantids including
Ctenopoma damasi in Africa (Berns and Peters, 1969) and Colisa, Betta and Trichogaster in Asia.

198
The building of nests enables the eggs and newly hatched fry to be concentrated in a protected locality
that is easily defended by one or both parents. Floating nests also bring the eggs and young fish into
contact with the better oxygenated upper layers of the water column, a very necessary feature as most
species making this type of nest inhabit highly deoxygenated waters. Juveniles of species which build
their nests on the bottom often have gill-filaments to improve their oxygen uptake, and the parent fish
fan the nest to ensure a flow of aerated water. Furthermore, nests are usually placed at the limits of the
advancing water where the dissolved oxygen levels are still moderately high. Such constructions are,
however, vulnerable to sudden changes in water level which can either leave the nest stranded or
submerge it in too great a depth of anoxic water. To avoid this happening some cichlids move their
eggs up and down by mouth to follow the advancing or receding flood.
Parental care reaches its most extreme among those species which bear their young throughout their
development. Many of the African and some South American cichlids and other species such as
Osteoglossum bicirrhosum incubate their eggs in their mouths, and continue to shelter their fry until
they become independent. The mouth brooding habit enables spawning to take place before the
floods. The young can be conserved in a well oxygenated environment (the parent's mouth)
throughout their development, and they can be deposited on the nursery grounds at the fringes of the
plain which are far distant from the breeding sites. Most mouth brooding species also construct nests
which serve both as territorial markers for their breeding rituals and as clean places upon which the
eggs are deposited prior to being picked up by one or other of the parent. Most cyprinodonts and the
glandulocaudine characins have progressed even further and have developed internal fertilization
which in many cyprinodonts is coupled with live bearing. Nelson (1964) in his study of the
glanducaudines considered that internal fertilization is an adaptation to the floodplain environment
permitting mating to occur when the fish are concentrated in the dry season habitats. Egg laying, or in
the case of the poeciliid cyprinodonts the birth of the young, can thus be delayed until the female can
move into the flooded shallows at the margin of the floodplain.
FECUNDITY AND SPAWNING PATTERNS
The fecundity and spawning pattern of fishes is correlated with the types of breeding behaviour
described above. Two main categories exist: total spawners, in which all the eggs ripen and are shed
within a very short time, and multiple spawners, in which repeated breeding occurs in any one season
with only a small proportion of the total eggs stock becoming ripe at any one spawning.
Long distance migrant species of whitefish, which are usually open substratum spawners showing no
parental care, belong to the first category. Their eggs are usually small and are produced in very large
quantities to compensate for the wastage inherent in this type of spawning. Fish such as Prochilodus
platensis produce between 360 000 and 750 000 eggs for individuals between 40 cm and 65 cm
respectively (Vidal, 1967). Egg counts for P. scrofa (Ihering, 1930) and P. argenteus (Fontenele,
1953) also fall within this range. Salminus maxillosus of between 52 and 100 cm were reported by
Ringuelet et al. (1967) to produce between 1 152 900 – 2 619 000 eggs. The regression for the
fecundity of Hilsa ilisha; F = 0.9550 exp 0.5396, formulated by Pillay and Rosa (1963) predicts
between 250 000 to 1 600 000 eggs for individuals within the normal size range. The fecundity of
Alestes baremoze ranges between 32 000 eggs for a female 24.4 cm long (177 gm) and 111 000 eggs
for a female 31.4 cm long (394 mm) and is described by the relationship: F = 0.345 Wt of female in
gms - 25 (Durand and Loubens, 1970). Really large species such as Lates niloticus can produce
extraordinary quantities of eggs, over 11 million having been recorded from some individuals by
Okedi (1971). Although the above examples indicate the numbers of eggs produced by total spawners,
the listing is far from complete and more details are given by Lowe-McConnell (1975).
Partial spawning is usually associated with some degree of parental care. The eggs are larger as is
reflected in the lesser numbers per gm wet weight of ovary. A total spawner such as Eutropius
niloticus has 2 950 eggs/gm, whereas Oreochronis sp. have about 135 eggs/gm (Willoughby and
Tweddle, 1977). The number of eggs released at any one spawning is much lower than in total

199
spawners. However, partial spawners may breed several times during any one season and it is,
therefore, difficult to estimate the total fecundity of the fish. This is further complicated by the fact
that the number of young reared is more often a function of the parent's capacity to care for a brood,
than of the ovarian egg production. A typical example of this is the mouth brooding cichlid
Oreochromis leucostictus whose ovarian egg production increased approximately as a square of the
standard length (F = 1.091 L2.156), but whose brooding capacity only increased linearly (B = 17.6 L -
78.2) (Welcomme, 1967). The partial spawning habit is adapted to areas with erratic water levels as it
allows for the loss of one or more broods through the unpredictability of the environment. A further
mechanism to cope with this is found in species such as Arapaima gigas where only one third of the
available eggs ripen at any one time, the rest being available should earlier broods be lost due to
sudden changes in water level.
TIMING
Reproduction of fish in rivers tends to be highly seasonal throughout the world. This seasonality
appears correlated primarily with two factors, temperature and flow, which in the temperature zones
are more or less synchronous in that increases in flow result directly from the snow melt and increased
precipitation associated with rising temperatures in spring. Approaching the tropics the influence of
temperatures seemingly diminishes and the flood regime becomes increasingly important as the major
regulator of breeding. Thus throughout the world the onset of reproduction of the majority of fish
species tends to coincide with the earlier parts of the flood. Spawning may occur at low water, rising
water or peak flood but only very rarely during falling floods.
Some species are however known to breed outside the most favoured season. The salmonids with
their low water winter season spawning mode are a noted example in the temperate zone. In the
tropics several of the black fish species with elaborate parental care may also continue breeding well
outside the flood. Species of Oreochromis are well known for round the year breeding in lakes and
apparently continue this behaviour in some rivers, for instance in the Shire River (Willoughby and
Tweddle, 1977). Brooding cichlids generally have been seen to spawn at low water in river elsewhere
(personal observation) although the frequency of breeding is very much reduced at this time. Many
cyprinodont fishes spawn towards the end of the flood, behaviour which is correlated with their
annual habit whereby one year's fish hatch at the beginning of the rains and develop throughout the
life of the temporary waters which they populate. As pools desiccate in the dry season the fish lay
small batches of their dormant eggs in the bottom.
Species of other families have been reported as breeding during the dry season in the Congo river
(Matthes, 1964); Roman (1966) noted that Micralestes acutidens too breeds on the floodplain during
the floods but continues to breed at all times of the year in the main channel of the Volta river, the
only one of the dwarf forms studied by him to do so. Dormitator latifrons also shows a continuation
of breeding into the dry season albeit at a lower rate thus the peak spawning during rising floods in the
Chone River, Equador (Chang and Navas, 1984). Kramer (1978), who investigated the breeding
habits of small characins in a Panamanian stream, found that there were great differences in timing of
breeding. Here Bryconamericus emperador and Piabucina panamensis spawned in temporary
tributaries in the rainy season, Brycon petrosus and Hyphessobrycon panamensis spawned in the dry
season and Gephyrocharax atricaudatus and Roeboides guatamalensis spawned throughout the year.
Kramer concluded that in tropical environments where conditions are only mildly seasonal
ostariophysan fishes may show a greater range of timing in breeding than they do in areas with
marked seasonality. Similarly, observations from the Magdalena River on the number of species with
ripe gonads and the number of juveniles present each month indicate that breeding patterns in this
river may be rather diffuse (Kapetsky et al., 1977). On the basis of 18 species studied, Kapetsky was
able to distinguish three main groups of fish from their spawning behaviour, those species which
apparently breed only once per year, those which have two or more breeding periods and these species
which reproduce almost continuously. More than half the species studied breed during February to
April bridging the period from low water and in the first stages of the rising flood. There was a second

200
peak of breeding activity during the second phase of rising water (October) but very few species bred
during falling water (December - January). This spread in the reproductive season is probably
explained by the sustained period of high water in the Magdalena system.
The considerable flexibility shown by a single species in timing of spawning in different river basins
within a geographic region is illustrated by Brycinus macrolepidotus (Paugy, 1982). This species
spawns once during the peak floods in the Niger and Senegal rivers or only during low water in Lake
Chad. In the Bandama, however, the species breeds throughout the year. These differences appear to
be correlated with different hydrological regimes, as the Niger and Senegal are both flood rivers,
whereas the Bandama is a reservoir river lacking lateral floodplains. Lake Chad is of course
lacustrine.
Despite such exceptions, the observations of most authors indicate the similarity in breeding
periodicity in tropical rivers from Asia, Africa, South America and Northern Australia (Bishop, 1983).
Typical of such are the remarks of Schwassmann (1978), who produced evidence that those fishes
which live in Central Amazonian floodplain lakes and which are total, one-shot spawners, begin
migration to spawn during the rising waters, probably to shallow recently flooded areas. Colossoma
bidens, Prochilodus sp. Anodus spp., Brycon spp. and Potamorhina pristigaster all had fully
developed ovaries in January to February and migrated out of the lakes into feeder channels on the
rising flood in the Janauaca Varzea lakes of the lower Solimoes. The synchronization of the
reproductive cycle with the flood is sometimes so good that, in systems where the flood wave takes a
considerable time to progress down-valley, the breeding of downstream populations is delayed, often
for up to a month relative to the fish upstream. This has been noted from the Mekong by Sao-Leang
and Dom Saveum (1955) and the Parana by Bonetto et al. (1971). It is also evident from the
comparison of Daget's (1954) data for the Middle Niger where fish breed mainly in July-August with
that of FAO/UN (1970) for the Niger in Nigeria, where breeding occurred from August to October.
In some equatorial rivers, such as parts of the Amazon and Congo systems there are two rainy seasons
which produce bi-modal floods. In these areas many species have more than one breeding season per
year (Roberts, 1973). In the Congo most species breed at the start of the September-October floods.
During the April to June high water, there is also a period of reproduction, although Matthes (1964)
considered this to be less important. Biennial breeding takes place in the rivers leading to Lake
Victoria where species ascend the streams during the equinoctial floods to reach the flooded swamps
near the headwaters (Welcomme, 1969). The twin breeding peaks of the Magdalena River have also
been described earlier in this section.
In temperate zones fish species tend to spawn within very well defined ranges of temperature. Whilst
these ranges occur mainly in spring or early summer when the water levels are also rising summer
spawners also breed during falling water in some systems such as the Mississippi (Fig. 6.9). The range
of temperatures can be illustrated by some North American species (Data taken from Scott and
Crossman (1973)) (Table 6.8).
Table 6.8
Characteristic spawning temperature ranges for some North American
species
Family Species Temperature
range°C
Salmonidae Salmo trutta 6.7–8.9
Esocidae Esox lucius 5–12
Cyprinidae Hybognathus nuchalis 13–20
Notropsis cornutus 15–18
Hybopsis storeriana 21

201
Nocomis biguttatus 23.9
Cyprinus carpio 17–26
Catastomidae Catastomus catastomus 5
Catastomus macrocheilus 7.8–8.9
Catastomus platyrhynchus 10.5–18.5
Ictiobus cyprinellus 15.5–18.3
Ictaluridae Ictalurus punctatus 24–30

202
Figure 6.9 Number of fish species spawning in different months of the year compared with air
temperature and river discharge in (A) the Danube River and (B) the Mississippi river

203
Where the highest temperatures of the year occur at the same time as low water some conflict between
the two stimuli may occur. In the Okavango Delta spawning of most species takes place in the warmer
months of the year, (September-March). This period coincides with the flood in the North basin of the
swamp, but in most of the delta the flood occurs in the coldest part of the year and the fish breed
during low water (Fox et al., 1976).
The factors which initiate maturation and stimulate breeding of floodplain fishes are elsewhere less
clear and a number of factors have been implicated including changes in individual physical
parameters such as temperature, conductivity or flow, as well as the assemblage of conditions that
mark the beginning of the flood. It is probable that each species is affected by the various factors in
different ways and that such external releasers are only effective when superimposed on an internal
physiological rhythm of the fish. An illustration of the specificity of those conditions that induce
spawning is Pimelodus maculatus of the Jaguari River. Basile-Martins et al. (1975) considered that
maturation of this species started when temperatures reached 22°C and that a temperature of 25°C
was needed for breeding to occur. However, temperature was not the only regulatory mechanism as a
minimum increase in water level of 1 m over the low water level also seemed necessary to trigger
spawning (Fig. 6.10). Changes in gonadosomatic index (GSI) where:
were also used to define breeding of Alestes baremoze in the Chari delta which coincides both with
rising flood and increase in rainfall (Durand and Loubens, 1970). In this species Paugy (1978)
concluded that the high temperature attained during the dry season was necessary for maturation
whereas the arrival of the flood triggered actual spawning. The relationship between endocrine
activity, spawning, rainfall, temperature and day length have been presented for the Indian catfish
Heteropneustes fossilis (Viswanathan and Sundararaj, 1974). These three examples, together with
other observations show the underlying similarity of timing of maturity and spawning. It is well
known that many of the migratory characins and cyprinids will not spawn unless the flood
materializes. As Bonetto (1975) has noted, individuals of Prochilodus platensis which are isolated in
the floodplain lagoons do not mature when fish in the river channel do. Most Indian investigators
agree that the major carps will not breed if the flooding associated with the monsoon fails
(Parameswaran et al., 1970). Under these conditions, suitable inundated spawning grounds also have
to be available for reproduction to be successfully concluded, although one cyprinid, Cirrhinus reba
differs from this in that it can breed even at low water in the Cauvery and Bhavani Rivers provided
sexual readiness is induced by a short initial spate (Rao et al., 1972). According to Sidthimunka
(1972) Probarbus jullieni shows similar behaviour in the Mekong system and Krykhtin (1975)
reported that, while many food fishes from the river Amur bred only on the floodplain with rises in
water level, some species (e.g., Elopichthys bambusa, Erythroculter sp. also reproduced when the
water level did not rise. Indeed many phytophylic species in European rivers appear to have adapted
to the modified conditions following channelization and flood control by spawning within the main
channel rather than on the fringing flooded areas. However, in these species the main external factor
regulating spawning seems to be temperature rather than flow.
Despite such exceptions it may be concluded that in the majority of total spawning species breeding is
so timed that if floods are delayed or inadequate to trigger migration reproduction may fail in that
year. Partial spawning species appear to be capable of breeding whenever suitable conditions are
present and as has already been noted some such species breed throughout the year. Many others have
longer and more diffuse breeding seasons, starting to breed in the river channel before the floods
arrive and continuing with subsequent broods well into the flood season on the plain.
Maturation tends to be relatively rapid in river species. Most small tropical species are ready to spawn
by the onset of the rainy season following their birth and have very short life cycles (Lowe-

204
McConnell, 1967). Equivalent sized species in the temperate zone mature somewhat later, either
during the first or more usually the second year of life. Larger species in both tropics and temperate
zones often delay maturation until their third or fourth years and extreme examples such as Aspius
aspius or Catastomus catastomus may not begin spawning until their fifth year. Species in unstable
systems may respond to unfavourable circumstances by stunting and accelerating maturation within
the first year of life. Such behaviour is typical of species of the genus Oreochromis (Dudley, 1976).
Hatching times of eggs are in most cases closely related to water temperatures and they differ
considerably in any one species. For instance, eggs of Esox lucius which hatch in 12–14 days at 7–
9°C hatch in 4–5 days at 17–20°C, and in brown trout incubation times of 148 days at 2°C are reduced
to 27 days at 12°C (Table 6.9).
Figure 6.10 Gonadosomatic index (GSI) of Pimelodus maculatus in the River Jaguari compared with
temperature (T°C), water level in river (m) and rainfall (mm³) for 1971–73. (After Basile
Martins et al., 1975)

205
Table 6.9
Average times between fertilization and hatching in brown
trout (Salmo trutta) at different temperatures (from Forst and
Brown, 1967)
Water temperature °C Incubation time days 2.0 148
3.6 118
4.7 97
6.0 77
7.8 60
10.0 41
11.0 35
12.0 27
Because of the cooler and less predictable thermal conditions temperate species have larger and more
variable hatching times usually ranging from 5–10 days. In the tropics, with higher and more constant
temperatures, hatching is rapid and in some fishes may occur within 16–24 hours of the eggs being
laid. Such fast development has been noted particularly from open substratum spawners such as Hilsa
ilisha (Pillay and Rosa, 1963), Labeo rohita (Kahn and Jhingran, 1975) and Labeo victorianus (Fryer
and Whitehead, 1959). The eggs of some nest building species develop equally fast, although in
mouth brooders the eggs take longer to hatch (4–5 days) and the fry often stay with the parent for up
to 2 weeks.
Exceptionally long hatching times are also common in some species. Many of the autumn or winter
spawners of temperate rivers, such as Salmo trutta have development times of up to three months
meaning that hatching usually occurs in early spring. Similarly the drought resistant eggs of
Nothobranchius spp. may arrest development for over a year until favourable conditions prevail.
THE INFLUENCE OF HYDROLOGICAL REGIME ON SPAWNING SUCCESS
Several aspects of the flood regime can have an impact on breeding success or on the survival of fry,
and thereby influence recruitment to the fish stock. That there are considerable differences in the
numbers of young produced in various years is well known. Holden (1963) noted that some species
were only represented by one year class in the Sokoto River. There were also great variations in the
relative abundance of species, indicating year to year differences in breeding success. Non-migratory
species were most abundant in one year (1955) due to a strong 1954 year class correlated with a good
early rise of the river. The same factor obviously did not favour the migratory species. Holden also
mentions the observations of Wimpenny (1934) on the Nile delta lake where heavy floods were
followed by good recruitment giving a high density of fish. LoweMcConnell (1964 and 1967) noted
large fluctuations in abundance of different species in different years, which she attributed to
differential breeding success, short life cycles and rapid maturation. Here strong year classes of
Serrasalmus nattereri and Prochilodus were found in different years.
In the Amur system, the yield of phytophilous species, which breed on the floodplain of the river, is
also directly related to the flood regime. Year classes are usually weak in years of light flooding and
strong when floods are heavy (Nikolsky, 1956). That a similar dependency between recruitment and
flood regime exists in the Danube is indicated by the fluctuations in fish yield as a correlate of water
level analysed by Holcik and Bastl (1977).
From fluctuations in the number of Oreochromis andersoni, O. macrochir, and Tilapia rendalli,
Dudley (1972) deduced that there were seasonal changes in abundance. Furthermore, from differences
in the abundance of Oreochromis juveniles he concluded that breeding success and recruitment to the

206
stock was much better in those years having good floods. Later work on O. andersoni and O.
macrochir indicated that spawning success might have been depressed by high dry season water
levels. The reason advanced for this is that Oreochromis normally stunt under the severer conditions
where drawdown is greater. The stunted fish mature and breed earlier than they would in years when
the high volume of water retained allows fish to grow to their normal maturation size.
The extreme hydrological conditions of the Sahelian rivers during the 1970–1974 period have
provided examples of the effects of failed floods on the recruitment of several fish species. In the
Central Delta of the Niger, Dansoko (1975) and Dansoko et al. (1976) compared the biology of
Hydrocynus brevis and H. forskahlii. The floodplain spawning H. brevis and the main channel
spawning H. forskahlii give a useful contrast in that the effect of variability in the extent of flooding
should be reflected more in one species that the other. This was confirmed by poor recruitment in H.
brevis during the years when the flood failed. A similar succession of events happened in the Senegal
River where recruitment of all freshwater fish was lacking in the poor flood years of 1968 and also in
1971 as exemplified by Citharinus citharus in Fig. 6.8. In the Chari River several species have
disappeared following the sustained Sahelian drought which hindered breeding in the Yaeres
floodplains as well as producing unfavourable conditions in the river channel and in Lake Chad.
Larger species, Distichodus, Citharinus and Labeo and the migratory Alestes dentex and A. baremoze,
declined drastically in abundance and were replaced by other species including Hemisynodontis
membranaceus and Brachysynodontis batensoda. Oreochromis niloticus, because of its multiple
spawning habit could rapidly recolonize the floodplains once more favourable conditions appeared,
whereas in species such as Alestes, which are total spawners where the majority of the new recruits
are produced by the second and third year classes, a failure of two floods in succession so weakened
the stock that it appeared unable to recover especially in the face of continued heavy fishery
(Carmouze et al., 1983).
MORTALITY
CAUSES OF MORTALITY
Any consideration of mortality among river fishes is liable to be complex because of the interplay
among the many causes of death. Two main groups of natural factors may be identified: (i) Density
dependant factors whose rate is a function of the numbers of individuals of the species or of other
species within a given space, and (ii) Density independent factors which usually are non-biotic and
which originate in the changing physical and chemical characteristics of the river system. The two
groups of factors are interrelated especially in rivers with fluctuating water regimes as changes in the
volume of the living space constantly influence the density of the fish themselves. Equally the various
density dependant factors can affect the rate at which others proceed; for instance having mortalities
due to predation at one phase of the flood cycle can so reduce populations of prey species that their
subsequent mortality through competition among themselves is considerably reduced. An indication
of the linkages between some of these factors is shown in Fig. 6.11 for the fry and parr stages of
stream dwelling salmonids.

207
Figure 6.11 Summary of factors which effect changes in numbers of stream dwelling salmonids
during the fry and parr stages (from McFadden, 1969)
To the natural causes of mortality should be added fishing mortality. Attempts at separating natural
and fishing mortalities in river fisheries are rare. In any case, because most techniques employed in
artisanal river fisheries are themselves density dependant, the effects of removal of fish by fishing
may significantly alter the rate at which they die from other causes.
Death due to stranding or isolation of fish in headwater streams, main channel pools or temporary
water bodies on the floodplain is probably one of the major elements of natural mortality. Bonetto et
al. (1969) estimated that some 40 000 t of fish are lost in this way from the Parana system, basing
their estimate on the area of standing water on the plain (2 × 106 ha²) and the standing stocks of 20
kg/ha as estimated from a small sample of them. This figure was greatly superior to the 10 000 t
which were reported to have been caught by the fishery in 1967. There is no doubt that enormous
quantities of fish are lost in this manner in most systems, and this apparent wastefulness has been
widely remarked upon in the literature. Mortality through stranding is obviously closely related to the
flood regime. It is not particularly prominent during the rising flood although temporarily recessions
can leave fish isolated in pools and can destroy young fish in nests at the flood margin. More
extensive flooding can raise the total number or biomass of fish in the system through improved
reproductive success and growth making the whole community more sensitive to later contractions in
the volume of the aquatic environment. The form of the falling flood would appear to be critical, as a
more rapid draining of the plain would seem to give the fish less time to abandon their flood habitats
and would increase the chances of their being isolated in unsuitable places. On the other hand there is
a possibility that there might also be some favourable effects because of the stronger pattern of
currents which may make the fish more responsive to such stimuli. However, the behaviour of fish
during the return movement to the main channel has not been studied in any great detail and needs
considerable work in the future. The duration of the dry season equally affects the proportion of the
standing water that will disappear through evaporation or be rendered unsuitable for habitation by
certain species: Presumably the longer and more severe the dry period, the greater the mortality due to
this cause. The impact of stranding mortality on the dynamics of the community as a whole is difficult
to assess. If all of the fish produced during one wet season were to enter the permanent standing water
at the end of the flood, densities far surpassing the carrying capacity of the water might well result.
This could give rise to a corresponding increase in density dependent mortality with a final survival
not far different from that resulting when a proportion of the individuals are lost by stranding.
Deaths due to unfavourable hydrological conditions during the floods have been described from
several systems. In the Kafue flats (Tait, 1967) and the Parana, these have been traced to the

208
deoxygenation of the whole water column in lagoons when the lowering of the temperature and high
winds cause the breakdown of stratification and sudden overturn. Similar cold spells have been the
source of recorded mortalities in the Nile and in the Amazon, where they are known as “friagems”,
Brinkmann and Santos, 1973). In the Amur River, winter kill conditions arise in years of poor low
water flow when the main channel separates into a number of pools. These become deoxygenated
causing heavy mortalities of adult fishes (Krykhtin, 1975). Heavy fish kills in pools within the
channels of small stream or even major rivers produced by high temperature or deoxygenated
conditions are also common especially in the tropics or in temperate zones where eutrophicated of
polluted conditions prevail. Fish kills due to such causes are often spectacular in that large numbers of
dead fish appear on the surface, but they generally seem to be highly localized in space and time.
Reduced survival of stream living species and of the young of upstream spawners can be anticipated
in years of exceptionally heavy flooding. In such an eventuality the semi-pelagic eggs, juveniles and
even the fish themselves could be swept downstream past the most favourable sites for their
development. Similarly, fish may be lost to the community by the downstream drift of sudd islands in
which they have taken refuge. These islands may eventually reach saline or other unsuitable waters
where the fish die. In estuarine systems too, mortalities of freshwater fish have been observed when
they have been carried out to sea by excessive currents.
Many of the species of fish inhabiting rivers are of small size and have life spans of one or at the most
two years. Death in such fishes is possibly associated with stresses arising from spawning or from
senescence. The mechanisms defining the life span of any species are, however, not well understood
and many fish live for considerably longer periods in captivity than they would seem to do in their
natural habitats.
Intraspecific or interspecific competition is obviously one of the main potential causes of density
dependant mortality among species. Competition may be for food or, perhaps more commonly, for
some other limiting resource such as shade, refuge sites or breeding sites. In some species,
particularly redd cutting species such as salmonids, the phenomenon of overcutting, where one fish
destroys the eggs of a previous spawner, increases at high population densities. Also demonstrated for
salmonids is the exclusion of juveniles from feeding territories by incumbent fish at densities much
above 5 fry/m² (Le Cren, 1965) resulting in a plateaux in population at about this level. Le Cren was
able to obtain experimentally a linear relationship between the coefficient of mortality (%) and the
logarithm of the numbers of Salmo trutta present in a stream. Such relationships indicate the extent to
which direct intraspecific competition may affect populations as density increases.
Interspecific competition may also produce local differences in mortality rates, but in view of the
continual co-survival of multi-speciate fish communities over long periods of time it obviously does
not represent a major cause of mortality. Perhaps the most widespread interspecific contribution to
mortality is that of predation. As has already been noted, the role of piscivorous predators in the
community increases with stream order and predators form a particularly large proportion of the
biomass in the potamon. Here mortality of small species and of the juveniles of larger species due to
predation by fish is probably maximal during the draining of the floodplain after the flood peak has
passed. Evidence presented in the section on feeding indicates that in many tropical rivers, predators
do not feed extensively during the dry season. There is unfortunately no data on the intensity of
predation during the rising floods. At this time cover for prey is maximal and the fish are probably
dispersed widely over the plain. Nevertheless, the condition of the predators improves throughout this
period indicating that they are feeding to a certain extent. The juveniles of most predatory species use
alternative food resources during the earlier stages of the flood, but are already turning to a
piscivorous habit as the flood nears its end. During the run-off phase the stomachs of most predators
are full, and personal observations on the Ouémé and Niger showed them to be typically packed with
the juveniles of a wide range of species. Prey may include the young of the predators themselves
whose non-specific feeding habits may lead them to cannibalism. Selection by length of prey is,

209
however, much more widespread, and there would appear to be a relatively narrow size range at
which different species are vulnerable to predation.
Predation by animals other than fish also appears to be at its highest during the period of the draining
of the plain. Williams (1971) and Lowe-McConnell (1964), together with several other workers, have
remarked particularly on the number of water birds preying on the fish stranded in floodplain
depression or leaving the area by way of the channels.
Another possible contributor to total natural mortality is disease. This may become extremely
important during the phase of high fish concentrations at the end of the dry season. A range of
diseases infect salmonid and cyprinid species in temperate rivers. These are mostly associated with
the high densities of fish attained in intensive fish culture but easily spread to the wild. There is little
knowledge of the epidemiology of tropical fishes in their natural habitats, although Khalil (1971)
recorded 215 genera of helminths, and Awachie et al. (1977) listed a range of bacteria, protozoa and
Crustacea, all of which are parasitic on fish in African inland waters. According to Awachie et al.
(1977) the lotic conditions in rivers are not favourable to heavy infestations with such parasites.
However, large mortalities have been recorded from the various eleotrid species from the Sepik River
in Papua New Guinea. These were traced to the bacterium Aeromonas hydrophila, the causative agent
of dermal necrosis, and occurred not only among species inhabiting the floodplain but also those of
the headwater streams (Coates, 1984). A similar disease has been reported as spread throughout S.E.
Asian rivers in the early 1980s. That reservoirs of infection are present among natural populations in
rivers is also indicated by the rapidity with which diseases appear and spread through populations of
ornamental fish after their capture. Much would appear to depend on the concentration of fish in the
habitat and the swiftness of the current. In sheltered lentic environments the abundance of parasitic
organisms increases. Such conditions are present in floodplain pools or isolated main stream pools in
the dry season where the lack of current, warm temperatures and high population densities of fish
favour the spread of infection. Likewise, parasitic infestations are readily discernible on nursery
beaches where young Oreochromis and Tilapia congregate. Under crowded conditions too, there is a
build up of toxic excretory products, mainly NH3 and N02 in the water. These lead to gill or skin
damage, which in conjunction with heavy parasite infestations can cause death.
SEASONALITY OF MORTALITY
The various factors contributing to total mortality tend to follow a similar pattern in fluctuating
systems. Apart from deaths due to unfavourable conditions during rising floods, and the sweeping
away of juvenile or small fish also during this time, most causes of death intensify as the flood cycle
passes from wet to dry phase. Due to the fluctuating volume of the aquatic components of the river
there is a range of ichthyomass that can be supported by it. Available evidence suggests that the
maximum amount of fish is present during the flood and the minimum just before the onset of the
next flood. This means that there is an overproduction of ichthyomass during the flood which persists
throughout the period of falling water and which must be lost through total mortality during the dry
season. It also implies that the amount of water remaining in the system during the dry season relative
to the amount of water present during the wet is one of the major factors determining mortality.
Because there is an overproduction of ichthyomass during the wet season the biotic system can
respond to differences in the amount of water during the dry season so as to assure maximum survival
into the next year. What proportion of total mortality can be taken as fishing mortality without
affecting this flexibility of response is one of the key questions in the dynamics of river fish
communities and in the management of their stocks for fisheries.
ESTIMATES OF MORTALITY RATES
Total mortality rates (Z) are derived from the formula

210
and represents loss from a habitat or component of the ecosystem by death from all sources including
fishing, and the difference between immigration into emigration from the area under study.
Obviously, the larger the area sampled to establish Nt and Nt0 relative to the size of the whole system,
the more errors arising from immigration and emigration are liable to be reduced and the more Z
tends to represent death rate. Only very few estimates have been made for species of fish inhabiting
tropical rivers and these are given in Table 6.10. Values of Z are generally quite high in the early
years of life (Z 4) but tend to drop during the later years (Z 1). This may be because the fish become
less susceptible to predation as their size increases, and large species such as Colossoma have low
mortality rates even when young although heavy fishing pressure continues on most species
throughout their lives. In the extreme case of fishes whose life spans are very short, for instance the
one to two years of many species of Barbus, cyprinodont or small mormyrid, values of Z, can be very
high.
Comparison with mortality rates of fish from temperate rivers shows that, although survival is better
initially in the temperate species (low mortality rates), their mortality rates tend to increase with age.
An accurate knowledge of mortality rates is essential to the calculation of production an animal
communities, the lack of precise knowledge of this parameter for tropical riverine fish is a severe
handicap in understanding the dynamics of such populations, however indications of mortality rate
may be taken from Pauly's (1980) demonstration of the inverse relationship between mortality and
asymptotic length in fishes.
MODELS OF MORTALITY
The simple exponential model of mortality Nt = Noe-zt has been widely used to describe year to year
mortality of fishes. If used to describe mortality patterns during one year it assumes stable
environmental conditions such as these in reservoir rivers or canals and predicts maximum loss of
numbers early in the period under consideration. Such a model, however, does not adequately
describe mortalities within any one year in rivers with fluctuating hydrological regimes. To more
accurately represent mortality patterns under such regimes Kapetsky (1974) suggested that the form of
the mortality curve might be represented by rotating the curve for exponential decrease in number
around its diagonal axis. This gives a curve of the form:
Nt = N0 - N0 exp - zT[(exp zt)-l]
where z is a weekly mortality coefficient, to is equivalent to the time of recruitment, t = time in weeks
and T = 52 weeks. Such a curve gives a mortality rate which increases steadily throughout the year
(Fig. 6.12). While not wholly satisfactory this model is a useful generalization for use in broader
productivity models. The seasonal patterns of mortality described above indicate very little mortality
during the wet season and increased mortality as the year progresses. The majority of causes of death:
predation, fishing, disease, hostile environments, etc., would seem to be density linked, and for this
reason Welcomme and Hagborg (1977) reduced the number of fish in a simulated floodplain
population by inserting a density dependent mortality parameter in the simple exponential model:
Nt+1 = Nt exp - (exp zM)
Table 6.10
Annual mortality coefficients (Z) for certain fish species from some tropical rivers as compared with

211
fish species from temperate rivers
Species River Year Author 1 2 3 4 5 6 7 8 9
TROPICAL
Polypterus senegalus Chari 0.54 0.53 Daget and Ecoutin, 1976
Brycinus leuciscus Niger 1.20 Daget and Ecoutin, 1976
Hydrocynus brevis Niger 3.08 0.98 Dansoko, 1975
Hydrocynus forskahlii Niger 2.49 2.67 Dansoki, 1975
Oreochromis andersoni Kafue 2.47 0.65 0.65 0.65 0.65 1.70 0.58 0.58 Kapetsky,1974a
Oreochromis macrochir Kafue 3.98 0.70 0.70 0.70 0.70 0.70 Kapetsky, 1974a
Tilapia rendalli Kafue 4.61 1.40 1.40 1.40 1.40 Kapetsky, 1974a
Serranochromis
angusticeps Kafue 3.12 2.20 Kapetsky, 1974a
Hepsetus odoe Kafue 2.74 1.84 1.84 1.84 Kapetsky, 1974a
Total community* Bandama 2.67 Daget et al., 1973
Colossoma macropomum Amazon 0.45 0.45 Petrere, 1983
TEMPERATE
Salmo trutta Bere stream 2.10 1.34 1.81 Mann, 1971
Cottus gobio Bere stream 1.66 1.33 Mann, 1971
Rutilus rutilus Thames 0.42 0.42 0.68 0.70 0.92 1.00 1.26 1.86 Mann, 1971
Rutilus rutilus Danube 0.69 0.98 0.65 0.70 0.56 0.61 0.68 0.75 Chitravadivelu, 1972
Alburnus alburnus Thames 1.29 1.29 1.29 3.90 4.80 4.10 Mann, 1971
Danube 0.88 0.86 0.82 0.84 0.73 0.63 Chitravadivelu, 1972
Leuciscus leuciscus Thames 0.60 0.60 0.60 0.60 0.56 1.15 1.28 1.46 Mann, 1971
Perca fluviatilis Thames 0.98 0.98 0.98 1.61 1.49 1.39 1.45 1.47 Mann, 1971
Gobio gobio Thames 0.88 0.88 0.88 0.88 2.52 4.24 Mann, 1971
* Dominant species Labeo coubie and Alestes rutilus

212
Figure 6.12 Changes in number of fish in one age group over 52 weeks as predicted by a simple
exponential model (—) and Kapetsky's (1974) floodplain model (-----): z = 1.5
In this simulation z was obtained from a mortality coefficient characteristic of selected floodplain fish
species; M was calculated from a formula of the form M = aBx where B is the weight per unit volume
of the fish in the system. M was adjusted to unity at a preselected biomass typical of a floodplain
system, producing slower mortality rates when densities fell below the biomass and increased
mortality rates at greater densities. The results of such a simulation are shown in Fig. 6.13.

213
Figure 6.13 Changes in the number of fish in one age group over 52 weeks assuming recruitment at
the start of week 1 when generated by the simulation of Welcomme and Hagborg (1977). Also
shown are the water levels used in the simulation.
STANDING STOCK AND PRODUCTION
STANDING STOCK
Estimates of ichthyomass or standing stock are available from many rivers and sufficient information
exists to permit a preliminary comparison between temperate and tropical waters. Standing stock
estimates are usually made by one or more of three basic techniques: multiple fishing to the
exhaustion of the stock, mark and recapture experiments, and poisoning a sample area with piscicides.
The assessment of the total fish population by these methods is not wholly reliable due to sampling
errors of various kinds, but the combination of several methods and the taking of samples over large
areas improves the reliability of the estimate.
The density of the population present in rivers with fluctuating water regimes depends much on the
amount of water present at the time of sampling. Population densities (ichthyomass per unit area) are
generally higher during low water when the fish are concentrated together than at high water when
they are dispersed, even though the total weight of fish in the system may be larger during the floods.
Furthermore, the degree of exploitation affects the standing stock with probable reduction in mean

214
ichthyomass as fishing pressure increases. As many water courses are very heavily fished, particularly
in the tropics, the values of ichthyomass might be expected to be lower than they would be in a virgin
or unexploited stock. Comparison between different systems is therefore difficult and can only
legitimately be made on the basis of samples taken during the same phase of the flood cycle and
taking into account the state of exploitation of the stock.
Main River Channel
Differences in standing stock in the main channel may be correlated with ecological parameters such
as slope. In general there is a progressive increase in ichthyomass as one moves downstream, as for
instance in the Luanza River, Congo where Malaisse (1969 and 1976) found 1.3 kg/km, 26.1 kg/km
and 31.7 kg/km in successive downstream reaches (see also Table 6.11). Much of this increase may be
traced to the widening of the river channel downstream; hence standing stock (Ichthyomass/unit area)
may not increase at such a rate. However, where the fall line of the river does increase lower
ichthomasses are present, as in the case of the Kaloma River where a reach with greater slope had an
ichthyomass of 7 kg/km and was intermediate between an upstream reach with 21 kg/km and a
downstream reach with 91 kg/km (Balon and Coche, 1974). This implies that there may be a trend to
increase in overall productivity along the river channel with higher standing stocks, downstream
although much more work is needed to clarify the ecology of small tropical streams.
In most rivers, flows are too swift for accurate samples to be taken during the floods in the main
channel. Consequently most estimates have to be made during low water. It was assumed by the
University of Michigan et al. (1971) that during the floods the ichthyomass in the Kafue river would
not differ significantly from that of the open water areas of the floodplain where, on the basis of
repeated fishing with seine nets and on rotenone samples in the open waters, an ichthyomass of 337
kg/ha was established. Samples taken in the channel at low water showed the standing stock to be
less, 204 kg/ha. In the light of work by Kapetsky (1974) this dry season estimate may be somewhat
low as he found the standing stock of the five main commercial species to be very different for three
separate reaches of the Kafue river, at 106.5 +29.21; 576.7 ± 129.2 and 386.6 ± 63.9 kg/ha
respectively. The differences in standing stocks were attributed to differences in fishing intensity in
the various reaches. The discrepancy between the mean 348.2 ± 59.5 kg/ha and the 204 kg/ha found
by the University of Michigan could be explained in part by the reduction in water volume by a factor
of 0.7 between the two sets of samples, but because the University estimate is based on all fish
species, the actual difference between the two estimates is most probably greater than it appears.
In the Chari river, Loubens (1969) carried out an estimate of standing stock in a 360 m² pool left in
the main channel at low water obtaining 15 138 fish/ha weighing 861 kg/ha. More comprehensive
results were quoted by Daget et al. (1973), whose rotenone sampling at two sites in the Bandama
River enabled the evaluation of changes occurring in the dry season. These were reduced into the
formulae: for the reduction in number (dj) with time in days (j). At one site, numbers were reduced
from 3 417 fish/ha on 31 January to 1 411 fish/ha on 30 May, and biomass fell from 125 kg/ha to 50
kg/ha in the same period. At the other site, 2 271 fish/ha on 31 January decreased to 996 fish/ha on 2
August, a loss of biomass of 144 kg/ha from 257 kg/ha to 113 kg/ha. The lower estimates of standing
stock obtained from the Mekong by Sidthimunka (1970) of 6.0 and 7.4 kg/ha were attributed to
difficulties in sampling and one sample from the Mong tributary of the river gave the much higher
value of 137.4 km/ha. Other small rivers in Thailand have been recorded with very similar levels of
standing stock, 118, 186 and 81 kg/ha for the Bori Pat, Lami Pi and Muak Lek rivers respectively
(Geisler et al., 1979). Bishop (1973) also calculated an ichthyomass of 179 kg/ha in the small black
water Gombak river in Malaysia although that estimate was arrived at by assuming only 20%
efficiency with electro fishing gear applied to catches equivalent to about 38 kg/ha.
Log dj = 1.63292 - 0.003202j and Log dj = 1.67286 - 0.003166j

215
Unpublished fishery studies relating to the lower Mekong basin, prepared under the auspices of the
Mekong Committee present values of 60 kg/ha for the upstream Mun river tributary to the Mekong,
and 91.9 ± 51.7 kg/ha for 11 other tributaries. The value of 60 kg/ha for the Mun river is a mean of
several estimates which fall from 120 kg/ha at low water to 5 kg/ha during the rising flood. The
progressive drop of ichthyomass per unit area is attributed to the dispersal of the stock with increasing
volume of water. Downstream in the main inundation zone the riverine standing crop was estimated at
135 kg/ha, but as this and the other values quoted are derived from capture fisheries the authors
consider them to be low and they probably exclude the first year fish which, as has been shown,
contribute the major part of the ichthyomass. The basic productivity of the water may also affect the
standing stock, especially in exploited populations. In black water rivers relatively low values of
ichthyomass may be anticipated, such as the 88.7 ± 75.7 kg/ha from the Baram River (Watson and
Balon, 1983), although no correlation between nutrient status and standing stock was found by Geisler
et al. (1979) in Amazonian rivers and analysis of standing stock against conductivity yields no
significant results. Considerably more data is needed from black waters in order to more fully
understand their ecology.
Much work has been done on the fish populations of temperate rivers to determine their ichthyomass
(Table 6.11). For example, Mann (1965) found a total ichthyomass of 659 kg/ha in the river Thames,
a value which he considered high for temperate rivers, although Backiel (1971) estimated that total
ichthyomass in the Vistula river may be between 200 and 1 100 kg/ha. In Belgian rivers of various
sizes, Timmermans (1961) and Huet and Timmermans (1963) found between 130 and 300 kg of
fish/ha by electrofishing. Philippart (1978) found that the total ichthyomass in the river Ourthe
corresponded to 315 kg/ha, or approximately 900 kg/km. Within this mean biomass fluctuated
following various factors such as temperature or depth of water but the main correlate was with
channel width according to the formula:
Ichthyomass = 6.9569 x 1.8456 (r = 0.919)
Detailed analysis of 40 Belgian rivers shows the main biomass to be 197.8 ± 137.8 kg/ha although
potamon reaches such as the Ourthe or Semois Rivers range between 250–350 kg/ha. An inverse
relationship was established by Timmermans and by Cuinat (1971) for French rivers:
posit that small rivers with high slope have a larger biomass per unit area, at least in Europe.
Philippart (1978a) proposed a modified model:
B = -295 + 0.19A + 5.72T+17.58S +16.8W
where A = alkalinity; T = mean temperature; S = mean slope and W = mean width based on the set of
Belgian rivers. Similar values have been obtained from temperate rivers other than those of Europe.
For example, the Horokiwi stream in New Zealand supported up to 311 kg/ha of fish (Allen, 1951)
and ichthyomass in trout streams in North America may reach 471 kg/ha (McFadden and Cooper,
1962). Swingle (1954) recorded 143 kg/ha from deep parts of the Coosa river and 154 kg/ha for
shallow areas. Ichthyomass in deep parts of other large Alabama rivers varied from 51.2 to 1 730
kg/ha, this latter being attained in the Tenson river which had a particularly wide floodplain. Further
to the North, Mahon et al. (1979) found 11 126-74 765 individuals/ha, equivalent to 32.4 to 190 kg/ha
for the Speed river Ontario.
This brief survey of standing stocks from streams all over the world indicates a wide range of values
at all latitudes. It is perhaps surprising, however, to find that there is little basis for any supposed

216
higher carrying capacity in the tropics. In fact values of between 100 and 600 kg/ha are common in all
continental areas.
Backwaters
Secondary channels and blind river arms, which have many of the characteristics of floodplain
standing waters but remain connected to the main channel for most of the year, are richer habitats than
the main river.
Perhaps the most detailed studies of fish populations of river backwaters come from the Danube. Bastl
et al. (1969), Holcik (1972), Holcik and Bastl (1973 and 1976) and Chitravadivelu (1974) have
investigated the Biskupicke, Zofin and Vojka branches using mark-recapture and repeated catching
methods. Their work led to the generalization that ichthyomass varies between 300 and 500 kg/ha.
About 20 species were present although species number varied with the area of the arm. Population
densities were high in the summer when fish took refuge in the side arm from the current in the main
channel. In the autumn the ichthyomass dropped as larger fish disappeared from the shallower waters.
Very low values, often below 100 kg/ha were associated with unusually strong flooding which
dispersed the population, or with pollution which was especially common in same side arms of the
Danube. A similar ichthyomass has been recorded from the Poltruba arm of the Elbe, where Oliva
(1960) found a population density of Rutilus rutilus and Alburnus alburnus of 222.3 kg/ha using
rotenone. As this was obtained from only two species the total ichthyomass was presumably
somewhat higher. As many as 22 350 fish/ha were found in a secondary arm of the Chari by Loubens
(1969) although 98 percent of these were small fish of less than 10 cm length. By contrast, 96 percent
of the 2 150 kg/ha were contributed by the few fish larger than this length. The same backwater had 5
616 kg of fish/ha at the end of one flood which had fallen to 1 600 kg/ha two months later. A fourth
sample from the same area a year later, only yielded 369 kg/ha, indicating the year to year variability
that can be expected in the same body of water. Another backwater of the Chari gave an estimate of 2
166 kg/ha from an area of 6 000 m². In the Bandama, Daget et al. (1973) showed that population
densities can increase in such backwaters. Contrary to the trends shown in the main channel, the
number of fish in a minor river arm increased from 1 408 individuals/ha to 3 311 individuals/ha in
June. This corresponded to an increase in weight from 149 kg/ha in March to 350 kg/ha in June.
These figures, coupled with those for the main river, indicated that there is some movement during the
dry season from the open pools of the main river into the more shaded habitats of the backwaters.
Sidthimunka's (1970) figure of 219.8 kg/ha for a backwater to the main stream of the Mekong was
also considerably higher than the estimates from the main river itself, although the Mekong studies
figure of 125 kg/ha for a non-flowing backwater is similar to that in the main river. In the large
Alabama Tombigbee river the ichthyomass in a backwater sampled by Swingle (1954) was much
higher at 2 084 kg/ha, than either deep or shallow areas of the main stream with 457 and 570 kg/ha
respectively.
Standing Waters of Floodplains
Populations of fish in the floodplain lakes of the Danube are of similar densities to those of the
backwaters. Balon (1967) in his sample of 13 water bodies, found between 335 to 318 632 fish/ha.
The composition of the population varied considerably from one sample to another. The ichthyomass
also had a wide range of variability; 230.4± 277.6 kg/ha. It was lowest during high water (136 ± 127
kg/ha in July) and highest during low water (480 ± 334 kg/ha). Other observations from seasonal
water bodies of the Danube floodplain by Holcik et al. (1981) gave a mean of 366.7 ± 293.6 kg/ha. In
the Russian Nyamunas River the floodplain is inundated by the spring floods which bring large
quantities of fish into the oxbows and artificial channels. These are isolated during the summer and
Gaygalas and Blatneve (1971) estimated that about 200 kg/ha of fish remained in the autumn. As
about twice this number were caught by anglers the total spring biomass was probably closer to 500
kg/ha. The floodplain pools of the Canadian Irvine Creek System described by Halek and Balon
(1983) supported standing stocks of 132.6 ± 57.7 kg/ha. Much of the variation between pools was

217
attributed to the duration of the connection between the pools and the main river. Pools in which this
connection persisted for some time had greater density of young of the year fish and hence higher
mean biomass.
In tropical rivers standing stocks of floodplain pools tend to be somewhat higher although on some
floodplains, such as the Magdalena and Mekong, there is a considerable movement out of the
depressions toward the main river channel which may account for the low values obtained from such
system. Table 6.11 summarizes the dry season ichthyomass from the standing waters of a sample of
tropical rivers. Ichthyomass apparently varies according to a number of factors. As with other parts of
the aquatic system, population densities are normally greater during the dry season. Thus when
Kapetsky et al. (1976) estimated a minimum biomass of between 0.23 and 251 kg/ha (mean 55.7
kg/ha) as being present in the open water and between 20 and 232 kg/ha (mean 79.8 kg/ha) in the bay
areas of the cienagas of the Magdalena floodplain he also found a good negative correlation between
the population density and the water level in the cienagas. Strangely enough, whilst the University of
Michigan et al. (1971) also found population densities to be higher in the open waters of the lagoons
of the Kafue river during the dry season (426 kg/ha) than during the floods (337 kg/ha), vegetated
areas showed the opposite tendency and very high concentrations of fish (up to 2 682 kg/ha) were
present in such areas at high water. High densities of fish under floating vegetation seem to be
comparably rare, however, and permanent swamps, particularly those under papyrus, are notorious for
the poverty of their fish communities. This situation may change temporarily during the flood when
currents can oxygenate the water column under the plants. The University of Michigan study also
indicated the loss of ichthyomass during the dry season when in one pool an initial standing stock of 2
693 kg/ha diminished by 75% to 684 kg/ha in three months. In a second pool a similar decline from 3
306 to 501 kg/ha occurred in 10 weeks, although in this case the pool was connected to the river for
part of the time and emigration might have occurred. Bonetto (1980a) studied a population of
Prochilodus platensis which colonized the Laguna Gonzalez when it was flooded by the Riachuelo
River in 1971/72. The lagoon subsequently remained isolated from the river and he was thus able to
trace a single year class through a number of years by extrapolation from mark-recapture experiments.
The isolation of the population from other year classes of Prochilodus undoubtedly gives a rather
distorted picture of what happens in more normal lagoons. At the time of separation from the river the
fish were probably about 1 year old and, perhaps because of the limitation or food availability, the fish
grew to a relatively small terminal length of 36 cm, as opposed to 60 cm in lagoons connected to the
river. The standing stock increased from 232 kg/ha in the first year to 351 kg/ha in the second and
decreased thereafter to 199 kg/ha in 1976 when observations ceased.
The form of the lagoon and the nature of the bottom may also influence ichthyomass. In the Senegal
river, long narrow pools formed from isolated drainage channels supported a much higher standing
stock than round depression pools, 205 ± 155 kg/ha as against 13 ± 6 kg/ha (Reizer, 1974). In the
Sokoto, Holden's (1963) analysis shows that a greater proportion of fish preferred intermediate
sand/mud bottoms (1 012 kg/ha) as opposed to sand (785 kg/ha) or mud (233 kg/ha).
Differences in standing stock have been related to the degree of organic fertilization of the water body
by Fox (1976). His estimates for small unenriched pools in the southern Okavango delta showed
between 100–200 kg/ha to be usual, whereas a highly enriched peripheral lagoon had the highest
estimate of 700 kg/ha. While this evidence is far from conclusive due to the small sample size it
would appear reasonable that eutrophicated waters should support higher densities of fish relative to
those less rich in nutrients. Nutrient poor black waters on the other hand, appear to support lower
standing stocks. For example, eight floodplain pools from the Rio Pacaya and Rio Saniria tributaries
of the Peruvian Amazon gave 89.6 ± 37.1 kg/ha (Montreuil pers.com.). However, there is still
insufficient data available to attempt drawing some relationship between nutrient content and standing
stock of fish in rivers.
Table 6.11

218
Estimates of dry season ichthyomass in pools and lagoons of the floodplains of
some tropical rivers
River Size of
sample Estimated
ichthyomass kg/ha Author
Apure 1 982 Mago-Leccia, 1970
Candaba - 500–700 Delmendo, 1969
Chari 2 701–2 166 Loubens, 1969
Kafue 8
444 University of Michigan et al.,
1971
Magdalena 28 122 Kapetsky et al., 1977
Mekong 1 63 Sidthimunka, 1970
390
Mekong Fish. Studies
(pers.comm.)
Mogi Guassu 1 313 Gomez and Montiero, 1955
Nile Sudd 2 306–433 Mefit-Babtie, 1983
Okavango 2 200–700 Fox, 1976
Oueme 68 1 835±825 FAO/UN, 1971
Parana
Temporary
lakes
18 959±1 512 Bonetto, 1976
Permanent
lakes
4 766±348
Sabaki 1 786 Whitehead, 1960
Senegal 8 110±144 Reizer, 1974
Sokoto 25 661±557 Holden, 1963
Standing stocks may also be influenced by the preceding flood history of the river, a topic treated in
more detail under “Catch”. An example of such variability comes from the Atchafalaya distributary of
the lower Mississippi where Bryan and Sabins (1979) recorded a lowering of standing crop of 795 kg
ha in 1977 from 904 kg ha in 1975–76. The higher estimate comes from years preceded by good
spring floods (1973–75) whereas the lower estimate comes from years preceded by poor floods
(1976–77).
Total System
Studies analysing the contribution of the various components of a riverine system to the total standing
stock are rare. However, one such study was carried out by the University of Michigan et al., (1971)
who extrapolated rather widely from a limited number of samples of four different habitats of the
Kafue River and floodplain. Their results are shown in Table 6.12.
A similar analysis was carried out by Holcik et al. (1981) for the Middle reaches of the Danube River
with results as shown in Table 6.13.
Table 6.12
Summary of high water and low water estimates of ichthyomass from the Kafue
River and floodplain system
Area (ha²) Ichthyomass
(kg/ha)
Total
ichthyomass
(t)
High water
Open water lagoon 126 000 337 42 462
Vegetated lagoon 16 000 2 682 42 912

219
Grass marsh 136 000 64 8 704
River channel 5 300 337 1 786
Total 283 300 338.4 95 864
Low water
River channel 4 800 204 959
Open water lagoon 113 000 426 48 138
Vegetated lagoon 14 000 592 8 288
Total 131 800 435.6 57 405
Table 6.13
Present status of ichthyomass in the Danube between the mouth of the Morava
and Ipel rivers (from Holcik et al., 1981)
Area (ha²) Ichthyomass (kg/ha) Total ichthyomass (t)
Main channel 7 936.9 35.0 277.8
Backwaters
Czechoslovak bank 1 777.9 350.0 622.3
Hungarian bank 1 336.0 400.0 534.4
Both banks 3 113.9 371.5 1 156.5
Total 11 050.8 129.76 1 434.3
A general estimate of ichthyomass has also been made for Atchafalaya River Louisiana from samples
both in the main channel and some floodplain lakes. Here a mean standing crop equivalent to 860
kg/ha was calculated for the lower part of the basin and 555 kg/ha for the upper part (Bryan and
Sabins, 1979).
PRODUCTION
ESTIMATES OF PRODUCTION
Production can be defined as the total elaboration of fish tissue during any specified period, including
gonadial products and material formed by individuals that do not survive to the end of the period
(Ivlev, 1966). Standing stock and production estimates for a number of rivers are given in Table 6.14.
Estimates of production range from a maximum of 2 000 kg/ha/yr in the Thames to as little as 16.2
kg/ha/yr in the Speed River but have a mean of 218.6±335.6 kg/ha/yr. Several factors may influence
production among which the latitude, the basic productivity of the waters and the age structure of the
population are perhaps the most important.
Table 6.14
A selection of data on total biomass and production of fish in rivers
River Country Biomass
kg/ha Production
kg/ha/yr P:B Authority
Amazon Manaus Brazil 1600.0 2800.0 1.75 Bayley (1983)
Thames
(0+fish) UK 1315.0 2000.0 1.52 Mann (1972)
(1+fish) UK 659.0 426.0 0.65 Mann (1965)
Hinaki N.
Zealand
880.8 735.4 0.82
Hopkins (1971)
Kafue Zambia 520.0 618.0 1.19 Kapetsky (1974)

220
Ellis Canada 376.0 375.4 1.0 Mahon (1981)
Warkosc Poland 307.5 161.4 0.5 Mahon (1981)
Carrol Canada 274.8 280.3 1.0 Mahon (1981)
Hopewell Canada 228.4 263.5 1.0 Mahon (1981)
Hinau II N.
Zealand
217.7 241.7 1.11
Hopkins (1971)
Lemki USA 212.0 136.0 0.64
Goodnight and Bjornn
(1971)
Jaruma Spain 178–221 221–583 1.2–2.6 Lobon-Cervia and Penczak
(1984)
Tarrant UK 198.0 596.0 3.01 Mann (1971)
Kejin Borneo 173.1 261.5 1.50 Watson and Balon (1984)
Bere UK 161.7 270.0 1.69 Mann (1971)
Irvine Canada 149.9 226.4 1.50 Mahon (1981)
Hall UK 129.0 52.0 0.40 LeCren (1969)
Swann Canada 124.4 174.5 1.40 Mahon (1981)
Struga Poland 111.2 110.3 1.0 Mahon (1981)
Deer USA 84.7 160.0 1.89 Chapman (1965)
Utrata Poland 84.5 289.3 3.4 Penczak (1981)
Big Springs USA 84.2 118.0 1.4
Goodnight and Bjornn
(1971)
Mesta Bulgaria 80+82 80.0 1.0 Penczak et al. (1984)
Dockens UK 75.0 140.0 1.87 Mann (1971)
Clemons Fork
(3rd 0) USA 71.5 77.2 1.08 Lotrich (1973)
Kejin Borneo 71.0 98.3 1.4 Watson and Balon (1984)
Clemons Fork
(2nd 0) USA 63.6 105.5 1.66 Lotrich (1973)
Appletreeworth UK 62.0 30.0 0.48 LeCren (1969)
Black Brows UK 59.0 100.0 1.69 LeCren (1969)
Clemons Fork
(1st 0) USA 54.9 83.5 1.52 Lotrich (1973)
Bobrza Poland 50.2 83.4 1.7 Mahon (1981)
Needle Branch USA 45.9 90.0 1.96 Chapman (1965)
Zalewka Poland 43.9 54.0 1.23 Penczak (1981)
Kaka Borneo 38.5 47.5 1.2 Watson and Balon (1984)
Wolborka Poland 37.4 35.3 0.94 Penczak et al. (1982)
Speed (3rd Order) Canada 32.3 54.0 1.67 Mahon et al. (1979)
Lava Borneo 30.5 39.3 1.3 Watson and Balon (1984)
Payau Borneo 27.1 31.2 1.2 Watson and Balon (1984)
Lawa Borneo 21.3 33.3 1.6 Watson and Balon (1984)
Bulu Borneo 21.5 26.0 1.2 Watson and Balon (1984)
Hinau I N.
Zealand
19.6 42.8 2.18
Hopkins (1971)
Speed (4th Order) Canada 7.0 16.2 2.31 Mahon et al. (1979)
It is commonly held that tropical waters are more productive than their temperate counterparts, and in
lakes this undoubtedly appears to be the case. Unfortunately few observations exist on the
productivity of tropical waters but those that do exist give little support to this contention. In fact the
range, mean and standard deviation of standing stocks of 17 tropical rivers, 21.4 (154.4±198.7) 861
kg/ha/yr falls entirely within those of standing stocks for 26 temperate rivers, 7.0 (205.0±300.3) 1315
kg/ha/yr.

221
There is as yet equally little evidence for the lower productivity of rivers with lower ionic content,
partly because so many other factors appear to influence the characteristics of the stock. However, in
the tropics Watson and Balon's (1984) figure for black water streams from Borneo (conductivity >50
μmhos) is decidedly lower at 179.9 kg/ha/yr than Kapetsky's (1974) estimate of 678 kg/ha/yr for the
richer Kafue (conductivity 130 – 320 μ mhos).
The influence of the proportion of juveniles in the stock is much clearer and Table 6.15 summarizes
the percentage of total productivity contributed by various year classes in some fish species.
Table 6.15
Percentage contribution to total production by year class in some species of river
fish
Species Percentage products by year
class Author
0+ I II III IV+
Cottus cognatus 65.1 20.0 12.2 2.5 0.2 Petrovsky and Waters, 1975
Salvelinus fontinalis 40.8 41.0 14.7 3.1 0.4 Dept. of Natural Resources,
1974
Alburnus alburnus 46.2 10.1 20.7 17.0 5.9 Matthews, 1971
Cottus gobio 32.0 18.0 27.0 20.0 3.0 Matthews, 1971
Oreochromis
andersoni 61.2 22.8 12.2 2.2 1.6 Kapetsky, 1974
Oreochromis
macrochir 46.4 26.3 12.7 7.6 7.0 Kapetsky, 1974
Theoretically there should be a smooth relationship such as that shown by S. fontinalis or C. cognatus
where data was averaged over several years, but differences in year class strength due to differential
success in recruitment, such as is shown by A. alburnus or R. rutilus may cause variations in overall
productivity from year to year. Because of the great significance of juvenile fish to the total
productivity, the efficiency of sampling for the younger year classes is of great importance and
differences in population estimates may arise according to the degree to which the smaller fish are
included in the sample. That year to year variations do occur in some systems is a matter of record but
others show a remarkable constancy in productivity despite considerable changes in biomass. For
example, the production of Salvelinus fontinalis varied by less than 20% around a mean of 117 kg/ha
over 11 years of sampling in Lawrence Creek, Wisconsin. During the same period the biomass
underwent almost threefold fluctuations from 13.5 to 99 kg/ha. This means that the efficiency of
production was in this case strongly density dependent and a regression of P/B against biomass gave a
significantly high correlation coefficient (Dept. of Natural Resources, 1974). From Table 6.14 it may
be seen that, whereas standing stocks tend to be lower in lower order streams than in those of higher
order, the productivity tends to increase resulting in higher P/B ratios. This high productivity of the
rhithron, especially in the riffles, may be accounted for in part by the larger numbers of juvenile fish
in such areas (Schlosser, 1982). Production/Biomass ratios are generally useful indicators of the
rapidity of turnover of biomass and, while productivity is strongly correlated to standing stock:
variations do occur for a variety of reasons. As mentioned above the population density may influence
the rate of production as also might the proportion of juvenile fish in the stock and a further factor that
has been implicated is species composition. This latter may operate in two ways, firstly by the
longevity of the species and secondly by its trophic status. The contention that short lived species
have greater productivities (Leveque et al., 1977) cannot be verified from the present data set,
although it is to be anticipated that communities composed of small short lived species will have
greater productions than communities of larger species inhabiting the same body of water. For similar
reasons it may be supposed that predatory species, partly because of their generally large size and
partly because of their position in the food chain, would have relatively lower productivities. In this

222
case the mean value for P/B for 13 observations on 7 predatory species was much lower at 0.54±0.16
than the value of 1.2±0.5 for 95 observations on 37 species of all trophic categories and from both
tropical and temperate waters. The difficulties inherent in this type of analysis are illustrated by the
very wide range of P/B values obtained for some species; S. fontinalis, for instance, have been
recorded as having ratios as low as 0.4 (LeCren, 1969) and as high as 3.0 (Mann, 1971).
P = 3.3 + 1.2 B (r² = 0.85 for the data listed in Table 6.14)
MODELS OF STANDING STOCK AND FISH PRODUCTION IN RIVERS
Several approaches have been used to study and estimate standing stocks in rivers. These vary
according to the type of river in question, mainly because of difference in the ease with which various
kinds of waters can be sampled. In general the difficulties of sampling large streams, usually defined
as any water deeper than wading depth, and the large numbers of watercourses dictate the use of some
type of predictive model, usually an index. Obviously such models have to be based on a certain
number of estimates from the field and have to be verified from time to time in actual rivers. A second
approach is through simulations based on theoretical concepts of the ecology of the fish community.
Simple models based upon environmental characteristics
Several attempts have been made at relating some characteristic of the fish community to the
environment in which it lives. Usually such models or indices have been derived for practical
purposes of fisheries management and it is not always clear exactly what is being predicted. For
example, models will often claim to predict production when it is in fact catch that is meant and there
is therefore some overlap between models dealt with in this section and those described in the section
on appraisal of catch.
During their studies on the fishery resources of the rivers of Belgium and Northern France Leger, and
later Huet, were able to formulate some generalizations as to the zonification of the running waters of
temperate Europe. Arising from this Huet (1949 and 1964) proposed a simple model:
P = BLK
to assess the approximate ichthyomass available for exploitation in temperate rivers. In this model P =
annual productivity (or harvest) of the water in kg/km of river, L = average width of the river, B =
'biogenic capacity' and K = coefficient of productivity. Values between 1 and 10 are assigned to B
according to the amount of fish food (benthos) available. The coefficient K is the sum of K1, K2 and
K3 where the value of K1 is derived from mean annual temperature, K2 depends on the acidity of the
water and K3 summarizes the type of fish population.
This original method was modified by Lassleben (1977) for the assessment of fish catch from the
German stretch of the Danube (Kolbing, 1978). In this case fish catch (C) for a section of river is
determined as:
C = BK 10 kg/ha/yr
where B and the various components of K are as above except for K3 whose value may be derived
from a simple equation establishing the proportion of each type of species present. This method has
given good results in north temperate streams and were extended to larger rivers such as the Danube
by Holcik (1979) using the simple percentage of rheophilic and limnophilic species. The biogenic
capacity of the river may be assessed using the biomass of benthic invertebrates instead of the
quantity of aquatic vegetation as used in the other formulae. According to Albrecht (1953 and 1959)
streams with a biomass of less than 60 kg/ha can be considered as poor, those with a benthic biomass

223
of 60 to 300 kg/ha as medium and streams with 300 to 700 kg/ha good. Binns and Eisermann (1979)
followed a similar approach in developing a Habitat Quality Index to predict trout standing crops in
streams in Wyoming by deriving two models which were moderately easy to estimate. Their most
successful model was the expression:
logY+1 = [(-0.903) + 0.807 log(X1+1) + 0.877 log(X2+1) + 1.233 log(X3+1) + 0.631 log(F+1) + 0.182
log(S+1)] [1.12085]
where
Y = predicted trout standing crop
X1 = late summer stream flow
X2 = annual stream flow variation
X3 = maximum summer stream temperature
F = food index consisting of the product of;
X4 = nitrate nitrogen
X9 = substrate type
X10 = water velocity
S = shelter index consisting of;
X7 = cover
X8 = eroding stream banks
X11 = stream width
The various parameters were rated on a scale of 1 to 5 according to a simple table. This model, which
works well for the trout streams for which it was developed, illustrates the complexity of models of
this type. It also indicates the number of factors which may influence standing stocks and
productivity, as well as the difficulties of forming reliable estimates on the basis of any one factor.
This approach has been further elaborated by other workers in the United States using Habitat
Suitability Indices based upon a wide range of parameters. These are usually species specific (Rabern,
1984) and are based on a thorough understanding of the ecological requirements of the species
concerned as well as detailed information on a large number of parameters.
Models of dynamics of fish populations in floodplain rivers
Models of population dynamics and production derived for lakes may well be applied to rivers which
have stable hydrological regimes, such as reservoir rivers or those whose flow has been modified by
man. The situation is, however, very different in rivers having a seasonal fluctuation in water level
where the generally accepted simple exponential models of mortality and growth are generally
inconsistent with what is known of the biology of floodplain fishes. They are thus inadequate to
derive estimates of within the year changes in biomass and production. The catch that can be expected
from flood rivers is most probably linked to the excess of ichthyomass produced in the flood over that
which can be supported in the dry season. A more detailed knowledge of within the year changes in
these parameters is necessary for the management of both the fish stock and the hydrological regime.
Alternative models based on the assumptions of rapid growth and low mortality during the flood, and
low growth and high mortality during the dry season, have been proposed. Kapetsky (1974)
contrasted the production derived from standard models of growth and mortality with linear and
floodplain models of these parameters, and found that production estimates for age III fish onwards
gave very different results according to which model was used. The floodplain model combines
number:
Nt = No - Noexp - zT[(exp Zt) - 1]
and mean weight
(Wt = W0 + W1 exp gT [(-1-(exp gt)]

224
Bt = NtWte.
It gives estimates for production of up to 4.4 times that of the simple exponential models and twice
that of the linear model. The reason for this is shown in the differences between the theoretical
biomass-time curves for age group 0 fish which are plotted in Fig. 6.14 from the simple exponential
(a) and floodplain (b) models for the two values of z and a growth rate (G) = 5.3.
Figure 6.14 Theoretical biomass-time curves for age group 0 fish during one year when calculated
from (a) simple exponential expressions for growth and mortality, and (b) “floodplain”
expressions
Daget and Ecoutin (1976) also derived a model of biomass and production based on their growth
equation for fish which have an annual growth arrest. Biomass is calculated from Bt=NtWt where:
Wt = 0.68 10-5 Lt³
Lt is derived from the formulae discussed in the section on growth and Nt is derived from the simple
exponential mortality equation:
Wt = N0e-zt
The model was applied to Polypterus senegalus, giving good comparability with observed results.
Estimates of production ranged from 528.5 kg/ha/yr (P/B=0.56) to 281.6 kg/ha/yr (P/B=1.12) for
mortality coefficients z=0.04 and z=0.10 respectively. This illustrates the importance of correctly
estimating mortality rate in deriving production and biomass values. The use of a constant mortality
factor in this model is somewhat limiting and the abrupt termination of growth produces a rather
sharply peaked annual biomass curve (Fig. 6.15) which does not appear to reflect the natural situation.
A more detailed simulation of the dynamics of a floodplain fish community was proposed by
Welcome and Hagborg (1977) based on a combination of their growth and mortality models. By
introducing density dependent mortality rates, and growth and recruitment which were dependent on
the intensity of the simulated flood model enabled the exploration of the effects on a theoretical fish

225
community of changes in high and low water components of the hydrological regime. The simulation
gave a series of curves for total ichthyomass derived from hydrological regimes in which either the
flood intensity or the amount of water remaining in the system in the dry season could be varied
independently of one another (Fig. 6.16). It indicated that differences in the flood regimes produce
great differences in within the year ichthyomass whereas the magnitude of the population passing
through to the following year largely depends on the amount of water remaining in the system at low
water. They also indicated that the greater amount of water remaining in the system at low water the
more the differences induced by the high water regime are transmitted to the following years. The
curves in Fig. 6.16 were combined into a three dimensional plot of mean ichthyomass for different
high and low water regimes (Fig. 6.17). Values for fish production per mean flooded area derived
from this simulation agree well with the general findings on flood rivers ranging from 241 kg/ha/yr to
564 kg/ha/yr depending on flood regime with P/B values of between 1.35 and 1.77. Production and
Biomass are both maximal in the higher floods but the ratio between them increases as the maximum
area flooded decreases. This model, appropriately modified to the growth and mortality of local
Oreochromis stocks, has been applied to Lakes Alaotra and Ihotry in Madagascar (Moreau, 1980,
1982). Here the area under water in the dry season was 22 000 ha, whereas in the wet season the water
flooded 57 000 ha. Apart from some differences in the numbers of fish the predictions of the model
agreed closely with results obtained from experimental studies.
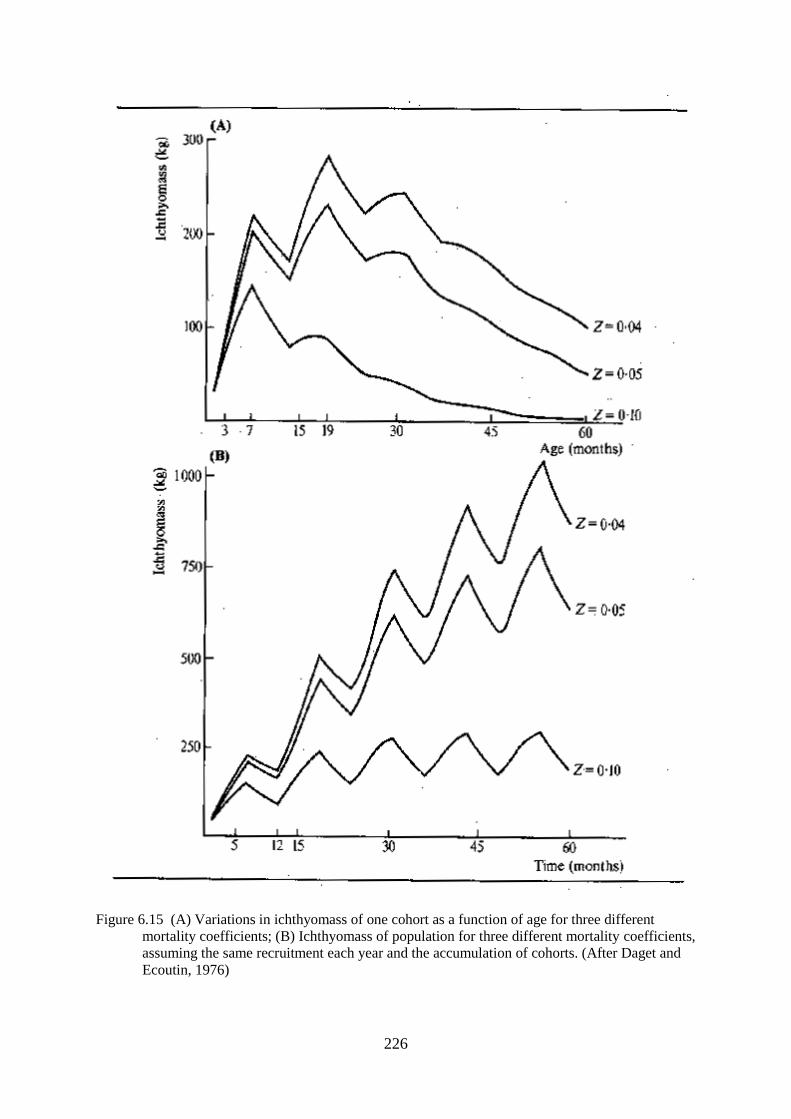
226
Figure 6.15 (A) Variations in ichthyomass of one cohort as a function of age for three different
mortality coefficients; (B) Ichthyomass of population for three different mortality coefficients,
assuming the same recruitment each year and the accumulation of cohorts. (After Daget and
Ecoutin, 1976)

227
Figure 6.16 Computer generated curves showing changes in total ichthyomass with time for different
flood regimes where (A) the low water regime is constant and the high water regime varies;
and (B) the high water regime is constant and the low water regime varies. Also shown is a
typical water regime (----). (After Welcomme and Hagborg, 1977)

228
These three models indicate a similar evolution of biomass throughout the year with a convex
ichthyomass-time curve. According to this there is an initial rapid increase in ichthyomass which
attains a maximum at about bankfull on the declining flood and which would seem to represent the
natural state fairly accurately. The models equally emphasise the great importance of the 0+ age group
which contribute up to 80% of the total ichthyomass depending on the mortality rate. Both the shape
of the biomass-time curve and the great preponderance of juvenile fish have important implications
for the management of the fish community for fisheries. The variations in production and ichthyomass
corresponding to differences in flood regimes predicted by the simulation suggest that equivalent year
to -year variations occur in the fish populations themselves. From records in many systems it is
known that the catch does indeed fluctuate in this manner, and so presumably do production and
ichthyomass.

229
Figure 6.17 Computer generated curves showing changes in population density (kg/ha) with time for
different flood regimes where: (A) the low water regime is constant and the high water regime
varies; and (B) the high water regime is constant and the low water regime varies. Also shown
is a typical water regime (----). (After Welcomme and Hagborg, 1977)
Curves plotted from simulation for ichthyomass per unit area indicate that the population is very
dispersed during the flood but concentrates rapidly as the water drains from the floodplain until
bankfull (Fig. 6.17). This phase is followed by a steady attrition which reflects the real situation
observed by Daget et al. (1973) in the Bandama and the University of Michigan et al. (1971) in the

230
Kafue River. Changes in the ichthyomass per unit area are also evident from the seasonality of the
fisheries which are at their most intense during those periods when the fish are concentrated in the late
falling flood and the dry season.
CHAPTER 7
THE FISHERY
THE FISHERMEN
All fisheries are to a great extent shaped by the nature of the environment and the characteristics of
the fish stocks they exploit. The factors influencing fisheries in rivers are very different from those
shaping lacustrine and marine fisheries. Three factors in particular give riverine fisheries their
character, diffuseness in space, seasonality and diversity. Because these are common to the vast
majority of the world's rivers there is a remarkable parallelism in the general form of riverine fisheries
and the communities that exploit them. This is subject, of course, to certain modifications according
to the general state of development of the countries in which fishery is practiced.
Rivers are lineaform and of limited width so the total area of water that can be reached from any point
on the bank is limited by the capacity to move up- or downstream. In most temperate rivers
commercial fisheries are usually motorized and the developed road networks make access relatively
easy. In many of the tropical rivers movement is by hand propelled canoe, although outboard motors
are also used. Whatever the mode of propulsion it is rare for there to be an expanse of water available
to one landing sufficiently large to support a capture and marketing operation of any size. In many
regions the problems posed by the linear form of rivers are aggravated by the swampy and changeable
nature of the terrain, which, together with the periodic submergence of any flanking floodplains,
hinders the installation and maintenance of permanent roads and other forms of communication. In
most areas of the world spatial dispersion and inaccessibility have combined to make fisheries labour
intensive, artisanal operations which are located in a series of small settlements along the river
channels or spread over the floodplain on islands of higher elevation. There are some obvious
exceptions to this general principle in estuarine 10/12/0waters, large floodplains or lakes and
reservoirs associated with rivers where the available area of water is greater.
In flood rivers the seasonal expansion and contraction of the water area can separate settlements from
the main course of the river by many kilometres in the dry season even though they are at the water’s
edge during the flood. Coupled to the hydrological cycle are the migrations of fish within the system
which causes the stock to undergo changes in abundance density and location. In response to the
fluctuations in the ecosystem fishermen either have to alternate with other occupations or they
themselves have to migrate. Because of this it is very difficult to define accurately who is, or is not, a
fisherman. In some systems the situation is fairly clear, as fishing is the task of one particular ethnic
group or tribe, and practically the whole of the active population of such a group may be interpreted
as being involved in the fishery in some way. In such cases the fishing group often does not own land
and coexists with other groups equally specialized in agriculture or pastoralism. In other rivers, the
riparian population remains relatively unspecialized as a whole, although sub-groups may concentrate
on one or more of the specialized activities including fishing. Within the fishing community there is
usually a well defined division of labour. The men fish, construct and maintain the gear and build the

231
boats, whilst the women collect, treat and market the produce. In such populations most able bodied
male, and sometimes female, individuals fish at some time or other. Furthermore, the role of small
pre-adolescent boys in many artisanal fisheries should not be underestimated as they act as fisheries
aids. In this capacity they paddle canoes, bait and control long lines, lift traps and generally keep the
fishery going when the adult members of the community may be occupied elsewhere.
For the purposes of convenience fishermen may be classified into three categories on the basis of the
time they spend fishing (FAO/UN, 1962). These are:
(i) occasional fishermen;
(ii) part-time fishermen;
(iii) full-time fishermen.
Such classification refers mainly to food fisheries in the tropics and sub-tropics and excludes such
special categories as temperate zone recreational fishermen.
OCCASIONAL FISHERMEN
In contrast to lakes, where a boat is usually essential to reach fishing grounds and to operate gear,
many river channels and floodplain waters can be reached on foot during part of the year at least and
fished by simple apparatus from the bank or by wading. This relative ease of access for the inhabitants
of floodplains coupled with a certain amount of free time between the sowing and the harvest of
floodplain crops means that casual fishing for subsistence is popular in floodplain communities.
Furthermore, in many areas of Africa and Asia, certain depression lakes or sectors of the river and
floodplain are traditionally reserved for the inhabitants of particular villages. These are fished during
festivals or fish drives which take on all the aspects of a holiday and in which all members of the
community participate. Most of the fish caught enters directly into the diet of the fishing community.
The individual time spent is low and for the most part the gear used is simple and relatively
unproductive. On the other hand the numbers participating are often very high. It is therefore difficult
to assess the contribution of such efforts to the total catch of any particular system.
PART-TIME FISHERMEN
Many sedentary peoples living on floodplains fish during part of the year. This is an activity that is
co-equal to or inferior to the alternative activities of such populations. The flood cycle, the biological
cycle of the fish, and the seasonal needs of agriculture impose a cyclicity on such communities.
During the floods there is very little activity in either domain, but as the waters drain from the plain,
fishing increases. As the floodplain dries the preparation of the soil and sowing of the seeds take
priority, to be followed with a second burst of fishing at low water. Harvesting of the crops follows
and the cycle repeats itself year by year. It is perhaps not surprising that, while part-time fishermen
use most of the types of gear used by professional fishermen, they also have a tendency to practice
various types of extensive aquaculture techniques. For example, the development of drain-in fish
ponds is associated with a certain agricultural type of land and water tenure in countries as widely
separated as Cambodia (Chevey and Le Poulain, 1940) and Benin (Hurault, 1965). Similarly, the
association of drain in ponds with paddy fields is a feature of many rice growing communities (Tang
Cheng Eng et al., 1973).
PROFESSIONAL FISHERMEN
In most aquatic systems there are groups of individuals that live entirely by fishing. The need for year
round employment and the movements of the fish stocks often force such groups to be nomadic.

232
Migratory fishermen have been noted from many systems and are particularly a feature of river
fisheries. For example, Bhuiyan (1959) described the migrations of the Hilsa fishermen of the Indus
who followed the movements of that species up and down the river. The fishermen of the Kafue
similarly move around depending on the water level and abundance of the fish stocks (Everett, 1974).
In the Magdalena fishery the fishermen move from the cienagas where they fish in the dry season, to
intercept the “Subienda” migration in the river (Bazigos et al., 1977). Some of these migrations by
fishermen can be of very large proportions. The historical upstream movements of the Haoussa in the
Niger River (Daget in FAO, 1962) might reasonably be compared to the operations of a factory
trawler in oceanic fisheries when the size of the investment relative to the per capita income of the
community is taken into account. This movement, which may be taken as fairly typical, involved
whole families of 20–30 persons who moved upstream in September and October to fish the northerly
portions of the Central Delta on the falling flood. The distance moved was in excess of 1 000 km. The
main vessels, which were often up to 15 m long and were equipped to support whole families on the
journey, were accompanied by a flotilla of small craft which were used in the actual fishing. On the
return journey, which was made on the next flood, the main vessels were loaded with many tons of
fish for sale in the Nigerian markets. These northern parts of the Central Delta are far from the main
centres of population, and have been exploited by wandering fishermen of the Haoussa and Bozo
tribes for many centuries. The incidence of nomadism therefore increases from the southern parts of
the basin where only two percent of the population are mobile to 52 percent in the heart of the lake
district (Raimondo, 1975).
Not all nomadic fishermen undertake such long journeys, but most have to leave their native villages
during at least part of the annual cycle. Many fishermen construct temporary 10/12/01fishing camps
on high ground within the floodplains which they occupy during the fishing season and which they
move following changes in the river level. To encourage this, it has been proposed that artificial
islands be constructed on some floodplains, especially the Kafue and the Sudan Sudd, to enable the
fishermen to exploit areas that have so far remained undeveloped for lack of suitable living space. In
some flooded areas, such as the Mesopotamia area of the Tigris-Euphrates system or the Ganges delta
in Bangladesh, the rural community including fishermen and their families already live on islands
made of soil and domestic refuse accumulated over the years. As an alternative many fishing
communities have developed houses on stilts which remain above all but the highest floods. These are
typical of the lowland rivers of the West African coast and of Asia. In Asia, too, some fishermen live,
at least temporarily, on boats or rafts which are associated with traps, stownets or lift nets in the river
channel (see Fig. 7.1).

233
Figure 7.1 Weighted average monthly landings related to mean water level of the Yamuna River at
Agra during 1968–69 (After Wishard, 1978)
Professional fishermen use a vast range of fishing gear, but in recent years have tended to concentrate
on one or two of the fishing methods based on modern materials such as seine nets, gillnets or cast
nets. Such fishermen, too, use powered craft to a great extent, if not for fishing then for the transport
of fish from the fishing grounds to the markets.
In recent years long distance nomadism has been discouraged in many parts of the world as this
practice has involved the movements of the nationals of one country into the waters of another. With
increasing awareness of the potentialities of their inland fisheries most countries have felt the need to
retain these benefits for their own nations and have trained local people accordingly. Many of the
traditional movements, such as that described for the Niger, have therefore broken down and in their
place shorter local migrations have intensified.
Most artisanal fishing communities have developed elaborate traditions, legends and even religious
systems which enable them to integrate culturally with the general ecology of the river on which they
live. Parallel institutions are found, not only among tropical communities, but also in many early
European and North American temperate peoples. Such studies as those of Smith (1981) or the many
papers included in Gunda (1984) indicate the rich folklore associated with such communities, many
elements of which serve genuinely conservationist ends. Such traditional mechanisms for regulating
the fishery include the definition of closed seasons, the delimitation of refuge areas or limitation on
access to the fishery. Such traditional institutions are fragile and disappear under pressure from more
modern political and socio-economic systems. For example, fishing is often seen as a kind of relief
occupation from the chronic unemployment and land hunger of much of the tropical world. The influx
of new and inexperienced fishermen into the traditional fishing communities results in their
disruption. Unfortunately no new institutions arise to replace the traditional management of the
fishery and the resulting anarchy may harm not only the fishing communities but also the fishery
resources on which they depend.

234
FISHING GEAR
SEASONALITY
In stable water bodies, including reservoir rivers and the larger permanent water bodies associated
with river systems, fisheries are pursued around the year. In cold temperate zones fishing may be
intensified in such rivers during the otherwise unfavourable cold season because of the greater activity
of salmonids at such times and in some areas fishing even continues under the ice cover. In flood
rivers, on the other hand, fish are vulnerable to the fishery for only part of the year when their
accessibility depends much on the ichthyomass per unit area as represented in Fig. 6.17 or during
temporary concentrations of migrating fish. During high water fish are dispersed over the floodplain
and concealed by mats of vegetation. The bulk of the population are juveniles, often too small to be
captured by any practical gear. Physical conditions are also unfavourable to the fishery as strong
currents sweep down the main channel and out over the plain, carrying with them logs and floating
masses of vegetation that carry away any fixed engines. As a consequence there is a slackening or
complete absence of fishing during the flood season in most river systems, although some gears such
as the ‘Atalla’ lift net of the lower Niger river, are operated specifically at such times to catch small
species which have taken refuge in vegetation fringing the main channel or juvenile fish which are
concentrated in the shallow margins of the lagoons (Awachie and Walson, 1978). Fishermen may also
continue to be active in some of the larger floodplain lakes where conditions are more stable or in the
flooded forest where specific fisheries for frugivorous fishes take place (Goulding, 1981). In older and
more tradition bound fisheries this cessation of activity is often reinforced by laws or taboos which
close the fishery to some or all gears. Many such traditional bans seem to have arisen from the
recognition of the flood season as a period of breeding essential for the continuation of the stock.
Fish become more available for capture as they congregate in the channels and pools of the floodplain
as the water begins draining off the main channel. At this time there is a very heavy and concentrated
fishery. Intensive fishing continues throughout the dry season both in the standing waters of the
floodplain and in the main river channel. Later as the water begins to rise again fairly specialized
fisheries concentrate on the adult fish migrating to their breeding sites. In most river systems this
sequence gives a pattern of capture whereby the efficiency of capture is inversely proportional to the
strength of the flood or is related to water level by more complex relationships such as those described
by Wishard (1976) for the fisheries of the Yamuna river at Agra (Fig. 7.1).
CAPTURE METHODS
The types of fishing method chosen for use in rivers are conditioned by three factors; first, the nature
of the fish stock, second, the form of the river and third the degree of development of the fishing
community. As has been described the fish communities of tropical and sub-tropical rivers are
particularly diverse containing a large number of species, most of which differ to some degree in their
selection of habitat, diet, migration pattern and ease of capture. They are also represented by an age
structure which is more than usually biased toward juvenile fish. The fishermen have, therefore, to
establish priorities for their fishery with respect to which species and age classes they attempt to
capture. Preference is often imposed by local food taboos or customs, although there are a few
fisheries which have been founded on the export of species which are not locally accepted items of
diet. In temperate climates the diversity of the community is reduced but, even in complex fisheries
such as that of the Mississippi, a few species only are selected as forming the basis for capture. In
some tropical fisheries too, particularly those at an early stage of development, only the larger species
are captured and in such restricted fisheries the range of gear in use is quite limited. In many other
fisheries, however, the fishery exploits all species that are catchable and so a great number of fishing
methods have evolved. Until recently, when cotton and later nylon twine and nets were introduced to
river fisheries, gear was constructed uniquely from local materials. Roots, vines, plant fibres, leaves,
stems etc. have all been, and still are, used for much of the gear encountered in rivers and floodplains.
The similarity in comportment of the different elements of the fish communities and the raw materials

235
of which the gear is made, have led to a considerable parallelism in most methods, even though they
have clearly been developed independently.
There are a number of local variations, and these are described in the many catalogues of fishing
methods in various countries such as Chevey and Le Poulain (1940) for the Mekong, Ahmad (1956)
for the waters of Bangladesh or Blache and Miton (1962) for the Chari/Lake Chad basin. A detailed
description of all gear used in river fisheries is inappropriate here so this chapter will examine the
general principles of the capture methods used in as far as they are relevant to the broad ecology of
these systems. Clearer descriptions of artisanal and other fishing gears, classified by type are available
elsewhere, for instance in Andreev's (1966) “Handbook of fishing gear and its rigging” or in Brandt's
(1984) book on “Fish catching methods of the world”.
While general types of gear used are determined by the above there is also a seasonal selection
depending on the flow regime. In flood rivers this tends to fluctuate during the year leading to the
successive use of a variety of gears. Typical examples of this are the fisheries of the Oueme R. in
Africa (Fig. 7.2) and the Mekong in Asia (Fig. 7.3). Similar examples of differential use of gears
according to flood stage are given by Goulding (1981) and Smith (1981) for rivers of the Amazon
system. In rivers with more constant flows the methods used are more restricted and generally
correspond to the gear types used during the low water phase in flood rivers.
Figure 7.2 Fishing timetables for the Mekong fishery. Thin line = fishing possible; thick line = peak
effort with method. Subsistence methods are not included in the tabulation. (Adapted from Fily
and d'Aubenton, 1966)

236
Figure 7.3 Fish gear utilization in the lower Oueme River during the course of one year
Certain types of fishing gear relying on modern industrially manufactured twines are in widespread
use. These are the seine net, the gillnet and the cast net. In many systems these methods are replacing
much of the traditional gear, and they are especially favoured by professional (full-time) fishermen as
their individual catching power is superior. Their care and maintenance requires an expertise often
lacking amongst the more casual elements of the fishing community. The cast net is in fact one of the
mainstays of these fisheries. Conditions in reservoir rivers and in flood rivers in the floodplain lakes
and the main stream at low water are well suited to its use. The water is fairly shallow, the bottom
unencumbered and the fish are sufficiently concentrated to give a good chance of capture. The mesh
size can be varied according to the species and size of fish sought. Fishermen using cast nets may fish
either individually or in combination with others whose manoeuvres serve to concentrate the fish still
further. The seine net requires relatively large teams to operate it and is very expensive. It is,
therefore, the gear of the professional “par excellence”. Its use is limited by current and especially by
the availability of bottoms which are sufficiently free of obstructions which would otherwise cause
the net to snag. The tenure of suitable seining beaches is often hotly disputed in the fishing
community. As a gear it has several precursors among traditional methods and communal fish drives
often have a seine like approach with lines of fishermen wielding baskets or clap nets. Alternatively a
barrage fence, which is one of the basic items of equipment for the floodplain fisherman, is set in a
large semi-circle and moved inshore in the same way as the seine. The gillnet is also common
although it is sensitive to floating vegetation and will not operate in strong flows. On the other hand, it
does come into its own in the larger floodplain lagoons and lakes where the depth, the large stretches
of open water and lack of current make it one of the most effective gears.
These gears either singly or together dominate certain fisheries. In the Magdalena River the cast net in
the main method, although seines, gillnets, traps and spears are also used to some extent. In recent
years the seine net has become the principal gear in the Senegal River, to the exclusion of many
traditional methods which are still current in the neighbouring Niger River (Reizer, 1974). The Kafue
river fishery also relies almost entirely on the combination of gillnets and seine nets (Everett, 1974).
Catches by these methods vary considerably according to the construction of the gear, the habits of
the fishermen and the time of year.

237
Low Water
During the low water period the majority of fish tend to remain relatively static. Riverine migrant
species which ascend the main course of the river and its tributaries during low water are an exception
to the more general behaviour pattern and form the basis of such fisheries as those of the “Subienda”
and “Piracema” in Latin America, the Alestes fishery of the Chari river, the Khone Falls fishery as
described by Chanthepha (1972), and the “Roak” fishery of the Yamuna river (Wishard, 1976).
Fishing methods tend to be active although some static gear is also used during low water. This is
often baited, for fish are still attracted to baited gear despite the general lowering of feeding intensity.
At this time the use of gillnets, seine nets and cast nets is maximal both in the river and in the lakes
and lagoons, but these gears on their own cannot reach all habitats or catch all species, consequently
many other fishing methods are in use. Hook lines of various types are common in the main river
channel. Longlines, baited with starch paste, offal, small fish, etc., are stretched across the river and
catch mainly the larger predatory species which are often not vulnerable to other gears. Unbaited
snagging and entangling lines are laid in the deeper portions of channels where big fish are
accustomed to rest, and small boys are usually to be seen in most places with a rod or leger line with
which they capture a seemingly endless succession of small fish.
Barrages are common throughout river systems, sometimes taking complex labyrinth-like forms, but
more often simply dividing the main river channel or the smaller drainage canals into sections which
limit fish movement and facilitate their capture by other means. Many types of trap, (Fig. 7.4), are
associated with the barrages where they capture the fish that are milling about trying to pass the
barrier. Traps are also set by themselves amongst vegetation where they attract fish seeking refuge
there. The traps may be unbaited, but they may also be baited to select for certain species. In West
African rivers for instance the same type of cylindric trap catches greater proportions of Clarias when
baited with oil palm fruits, Tilapia/Oreochromis when baited with maize meal and Macrobrachium
when rotting meat is used.
The problem of extracting fish from under the vegetation which fringes lagoons and river channels is
tackled in a number of ways. Special robust hand nets, which may be made out of netting or basket
work (Fig. 7.5), are scraped along the under-surface of the vegetation, or the vegetation mass might be
surrounded by a fish fence. In the latter case the plants are cut out piece by piece and the fence
advanced inwards so as to enclose the fish within a small space from which they can be captured by
hand nets or baskets. In the Oueme River such vegetation masses may be planted deliberately at the
end of the flood, either attached to the bank or recessed into it at the mouth of the channels which
drain the plain. They are left to collect fish for about two months after which they are fished and
replaced to be emptied again toward the end of the dry season. Harvests from such “refuge traps” or
“fish parks” can be quite high and in the Oueme river 15 installations of this type of mean area 440m²
gave a mean harvest equivalent to 1.88 t/ha (of park) per fishing or 3.88 t/ha/yr between 1958 and
1968.
A variety of gears have been developed to fish the main channel, especially the deeper portions where
the larger fish come to rest. Many of these are drawn or propelled by boats. A fairly widespread
device is the frame trawl drawn by one or two boats but the most popular is probably the Vee-shaped
net (Fig. 7.5) which can be mounted in a variety of ways and has great operational flexibility. Armed
with small mesh netting, it can, for instance, skim the surface to catch the small pelagic clupeid,
cyprinid and characin species found there, or be scraped along the bottom to capture many small
bottom-living species. With larger mesh it can also be plunged near the bottom where it drags for
bigger bottom-living species. With larger mesh it can also be plunged near the bottom where it drags
for bigger bottom-living siluroids. Nets similar to this may also be mounted on the bank where they
take migrating fish as they follow the shoreline or move inshore away from the current (Fig 7.6). In
Asian rivers lift nets perform a similar function. They may be mounted on the bank, but are
sometimes operated from rafts on which the fishermen live with their family. Such an apparatus is the
“Sadung” of the middle Mekong, whose operation was described by Fraser (1972).

238
The standing waters of the floodplain are often the property of a particular village or group, which
exploits them communally using clap nets or plunge baskets to virtually empty the water of any living
thing. But many waters are exploited by individuals.
Figure 7.4 Various types of fish trap from tropical rivers: (A) cylindrical drum trap (worldwide); (B)
vertical slit trap (Asia, Bangladesh and Mekong River); (C) folded woven trap (Niger River);
(D) funnel trap (worldwide); (E) spring trap (Africa, Niger, Chari and Congo rivers)
Larger lakes, of course, are fished in much the same manner as the main channel with a full variety of
gear. However, smaller floodplain lakes are usually either poisoned or fished out with fences in much
the same manner as the brush parks. There are many plant toxins which are used to capture fish both
in the lagoons and in pools of the main river channel. Table 7.1 gives some indication of the number
of bio-active plants in the Benue river system alone; although many of the shrubs listed are also found
in other parts of the tropical world, and Gunda (1984) describes an equally large range of plants used
for poisoning fish in European rivers. In recent years, even more powerful poisons have become
available in the form of insecticides such as dieldrin or endrin, which are used with apparent abandon.
Fishing with poisons is one of the main methods used by occasional fishermen and has the
unfortunate side-effect that it selects against the young fish which are particularly susceptible to these
substances. Its effect on the fish stock is of course minimal where temporary pools are fished, for
there the fish would die anyway before they could rejoin the stock but in permanent standing waters
the annihilation of the population is more serious, as here the fish take refuge during the dry season to
form a reconstituted stock in the floods. Poisons are also used to eradicate unwanted species, for
example, the piranhas (Serrasalmus spp.) in Brazil (Braga, 1976).

239
Figure 7.5 (A) Basket dip-net (Ouémé river); (B) “Vee” shaped dip-net (worldwide); (C) “Vee”
shaped net mounted on a canoe (Chari river) (adapted from Blache, Miton and Stauch, 1962)

240
Figure 7.6 (A) Bank mounted lift-net (widespread, example from Niger River); (B) handheld life-net
(widespread, example from Bangladesh); (C) raft mounted lift-net “Sadung” (middle Mekong)
Table 7.1
Ichthyotoxic plants used in fishing in Benue River (after Stauch, 1966)
Plant Active part Effect
Balanites aegyptica Bark which is crushed Kills fish within a few hours
Tephrosia vogelii Leaves and young shoots crushed Fish appear on surface very quickly and die
soon after
Momordica charantia Dried leaves and fruits: usually mixed
with Balanites
Effect very slow

241
Unidentified plant: (local
name: Horesoungsoungko)
Whole plant used after crush- ing in a
mortar. The shallow lagoon to be
fished is stirred up and the poison
mixes with the mud
Especially effective for catch- ing bottom
living mud-eating species such as Clarias
Crinum sp. Bulbs crushed in a mortar and put in a
sack which is drawn through the water
Very effective, kills all fish in a short time
Indigofera pilosa Ripe seeds Useful in waters of little volume but is
more often mixed with other products
Parkia filicoidea Pre-ripe seeds pulped Slow in action and ineffective against
siluroids
Syzygium guineense Bark which is crushed Rapid effect against all species
Euphorbia kamerunica Latex Renders fish inedible in large quantities:
small amounts are used to intensify the
effect of other poisons
Prosopsis africana Seed pods, dried and crushed Very slow (three days to produce death) but
intensifies effect of other poisons
Sarcocephalus esculentis Bark which is rubbed between two
stones
Irritant which fish avoid by taking refuge in
traps which are placed in the pool to be
fished
Adenium obaesum Fresh wood cut in discs and sun dried
for 2 days
Kills fish within three hours
Moringa pterygosperma Bark crushed between two stones Kills fish within three hours
Acacia ataxacantha Flowers dried in the sun Never used alone but acts as an intensifier
to Balanites or Momordica
Ximenia americana Bark which is pounded in water Particularly effective against cyprinids and
characins which appear belly up on the
surface within an hour. Frequently mixed
with Balanites
Ziziphus mucronata Flowers which are poun- ded in a
mortar
Kills all fish within two hours even
siluroids
As has been shown in the section on standing stocks, many lagoons contain a very high ichthyomass,
most of which is removed during intensive fishing of these waters. This fact has apparently been
recognized and, as we have seen in some areas, fishermen attempt to retain the maximum volume of
water in the depressions by damming the outlet channel. In more developed floodplains, the shape of
the lagoons may be regularized and eventually drain-in fish ponds may be dug into the surface of the
plain.
Fishing during the Rising and Receding Floods
As the floods start to rise there is a burst of activity by the fish as the adults move preparatory to
breeding. Locally high concentrations are present giving rise to very heavy but localized fisheries.
When the floods recede from the plain the water becomes confined increasingly into depressions and
channels. The fish follow these flow patterns to reach either the main channel or what will become the
standing waters of the plain itself. Fishing methods take advantage of these movements and are
mainly aimed at either directing the fish into places where they are more easily captured, or to
retaining the fish in floodplain depressions from which they may be more easily removed later. Such
gear may be based on bamboo or palm frond fish fences which are installed across the plain or the
channels through which the water enters or leaves. They can reach a considerable length and can be
arranged in complex forms giving a labyrinth-like effect. Capture is either in trap shaped chambers
(Fig. 7.7) or in special cylindrical traps or nets which are placed in openings in the barrage. Durand
(1970), for instance, has described a cross river barrage from El Beid River which drains the Yaeres
floodplains of the Logone River. Here the fences are arranged in a series of Vee's at the apexes of

242
which are held large hand nets. Both upstream and downstream migrants may be caught in the same
type of trap.
Figure 7.7 Barrage trap from coastal floodplain of Benin; note heart-shaped capture chamber
Thus in the Nzoia river the ‘kek’ type of barrier trap (Fig. 7.8) intercepts breeding fish moving
upstream, as well as adult fish returning from the spawning grounds (Whitehead, 1959a). Such fences
can contribute a large proportion of the total annual catch in some systems. In the Lubuk Lampam
(Indonesia) guide fences accounted for about 50 percent of the fish caught in 1975 (Arifin and Arifin,
1976) and on the African Barotse plain the “maalelo” fishery produced about 25 percent (631 t) of the
total catch in 1969. The “maalelo” of the Barotse floodplain is perhaps typical of the more open plain
type of barrier fishery. Here the fish guides are usually earth bunds some 75 cm high and between 3 to
40 m long, which deflect fish into traps placed at intervals along their length, although reed, wire
mesh or brushwood fences may also be used. Alternatively the bunds may join two areas of high
ground so as to retain a pool behind them. When eventually the dam thus formed is breached, the fish
are caught in traps and baskets at the outflow giving a yield equivalent to about 33.6 kg/ha of area
impounded (Bell-Cross, 1971). A few “maalelo” are fished during rising flood. Weiss (FAO/UN,
1970a), estimated that there were 10 000 of these weirs operating each year on the Barotse plain.

243

244
Figure 7.8 Fishing basket used in Kenya (A) and arrangement in a “kek” barrage (B). Fish enter
through the vertical funnel. (After Whitehead, 1958)
Complete blocking of small channels leading water out of depressions is common in other systems; it
has been noted from the Gambia River by Svensson (1933) and by Chevey and Le Poulain (1940)
from the Mekong. The pool thus created is fished, often considerably later in the dry season, either by
breaching the dam and catching the fish in traps, baskets or nets or by bailing the water out until the
enclosed section is dry. This principle has been suggested as a means for improving the fishery
productivity of floodplains as an extensive form of aquaculture.
In large channels which are not easily blocked with dams, stownets and wing traps form an alternative
to the barrage trap. The stownet is a conical fixed gear which operates rather as a static trawl, the
water passing through the net rather than vice versa. In the River Rhine these are operated using a
single anchored otter board, in the Oueme such nets are slung from poles securely stuck into the
bottom, but in the Tonle Sap, which drains the Grand Lac into the Mekong, they reach complex
proportions in the form of the “day” (Fig. 7.9). Such gears give very high yields during the main
migrations into and out of the major flood depression and in the main channel. Fily and D'Aubenton
(1966) recorded a mean catch of 33 t per unit for two lunar catching periods for 11 installations in
1962–63. Earlier results by Chevey and Le Poulain for 1938-39 gave about twice this figure (64.5 t).
In the rivers and canals of the Chao Phrya and Mekong deltas, stationary wing traps of the type
illustrated in Fig. 7.10, also take large quantities of migrating fish as they moved up and down the
channels during the dry season. According to Tongsanga and Kessunchai (1966) catch rates ranged
between 1.2 and 3.8 t/day in various canals.

245
Figure 7.9 Contribution of a “day” stow-net from the Tonle Sap: (A) cylindrical trap (“day”); (B)
disposition of individual traps in stow-nets mounted in a barrage; (C) method of use (i) with
gear lowered, and (ii) with gear raised (after Chevey and LePoulain, 1940)
Fishing at Peak Flood
Although in most rivers fishing is highly seasonal with only minimum effort being expended at high
water, some fisheries are pursued during the floods particularly in forested rivers such as the Amazon.

246
Such fisheries concentrate on frugivorous species which congregate in areas rich in fruit bearing trees
and shrubs. Here fish are captured mostly by hook lines, although bows and harpoons are also
employed for larger species (Goulding, 1981; Smith, 1981). In savannah rivers fishing continues in
some systems using traps, lines or gill nets set in areas of slack water or in the deeper parts of
floodplain lakes. However, the risk of loss of gear, the low return on effort and the difficulty of
working on a vegetation encumbered floodplain tend to restrict the practice.
Figure 7.10 stationary wing-traps from the Chao-Phrya delta. (After Tongsanga and Kessunchai,
1966)
BOATS
A detailed description of the various kinds of fishing craft is not appropriate here, but the availability
of suitable means of water transport is crucial in a fishery which depends much on the mobility of the
fishermen. Furthermore, in flood rivers the plains are inundated for several months of the year, and
the communities inhabiting them are forced to adopt a semi-aquatic way of life. During the floods
water-born transport is the only means of communication and even the markets are conducted from
canoes. As a simplification, two main types of boats are used in tropical and sub-tropical fisheries.
The first are the fishing craft themselves which are usually dugout canoes or planked craft between 4–
6 m long. Motorization of the smaller fishing boats is comparatively rare, especially among part-time
fishermen, and is in many cases of no great advantage. The second class of boats are longer, often up
to 10 m long and are used for transport of fish from the landing to the market. These are more
frequently motorized. In the main equatorial rivers, the construction of boats presents few problems as
wood is plentiful and dug-outs are easily made. In the savannah rivers, however, the lack of good
wood is limiting and canoes are often scarce and expensive as they have to be imported from
elsewhere. The development of suitable substitutes for traditional types of craft, which are cheap
enough for the fishermen, is one of the main preoccupations of fisheries administrators in such areas.

247
PRESERVATION OF FISH
TYPES OF PRODUCTS
A certain amount of the fish caught in river systems is consumed fresh by the fishermen themselves
and by communities with a limited radius of the fish landings, but in the more important fisheries, a
surplus to local requirements is produced which is sold for transport elsewhere. To improve the
quality of their product, fishermen as far apart as the Mekong River and the Magdalena River, keep
their catch in live chambers and even transport it in special boats with wet holds. Some species
survive and keep better than others in the fresh state after landing, and air-breathing fish particularly
are sought after because of the time they can be kept alive after capture. In India and part of Africa,
murrels, cat fish and some anabantids are transported for considerable distances in baskets lined with
damp weeds or moss. However, because of the dispersion and inaccessibility of fishing sites in the
river-floodplain system, and the rapidity with which fish deteriorate under tropical conditions, most
fish have to be preserved by one means or another for it to arrive in the markets in an acceptable
condition. Furthermore, the seasonal nature of the fishery means that a period of excess fish
production is followed by one of scarcity. Preservation techniques are thus needed to prevent fish that
are in excess of demand being lost and to even the supply of the year. Several types of treatment are
used, depending on local conditions and preferences.
Chilled Fish
The preservation of fish by icing or freezing is a comparative innovation to the river fisheries. Even
now its use is limited to areas where sufficient fish is caught to justify the expense of ice-making
plants, and where communications are sufficiently easy for the fish to be collected, iced and removed
rapidly. This method of keeping fish has grown up on major fisheries located in the main river
channel such as that for Hilsa ilisha in the Indus (Husain, 1973) where the fish are collected at certain
landings for icing prior to transport by rail to the major towns. Icing is especially popular in Latin
America, where the comparatively recent development of the river fisheries has left little time for
traditional methods of preservation to have arisen. On the Magdalena River and the Amazon, for
instance, fish traders come to the fishing sites in motorized boats equipped with ice to preserve the
fish. In recent years, preservation in ice has tended to replace other methods of treatment. In the
Mekong, Fraser (1972) recorded the addition of ice preserved fish to the customary range of products,
and in the Kafue, Williams (1960) commented on the growing proportion of fish which is carried in
this form.
Dried Fish
Sun drying of fish without salting is not practical in many of the world's river systems because of the
high atmospheric humidity. But in desertic or Sahelian savannah rivers the practice is common,
especially for smaller species. In the Senegal basin, for example, simple drying is the usual form of
treating the fish after they have been eviscerated, scaled and, in the case of larger species, cut into
strips. On the Niger, only the smaller species, such as Alestes, are sun-dried and in the Chad basin one
of the traditional fish products, “Salanga”, consists of Alestes dentex and A. baremoze which are split
open ventrally and laid on mats to dry in the sun. In the Mekong, and other Asian rivers, sun-drying is
also common, although this is sometimes combined with salting.
There is a noticeable loss of weight as fish dry. This varies much with the species of fish, for instance,
Clarias with their massive head bones tend to lose less than do characins or cyprinids. It is generally
agreed that the ratio wet fish: dry fish is about 3:1.
Salting

248
Preserving fish with salt is not common in inland waters, largely because of the high cost of the salt,
and secondly because in the more humid areas the deliquescence of the salt shortens the life of the
product rather than increasing it. In the Indus valley, salting is used only when there is a very heavy
catch of Hilsa which exceeds the capacity of the ice plants (Husain, 1973). Certain species are
prepared in this way in the Parana River (Vidal, 1969), where Lycengraulis olidus and more recently
Hoplias malabaricus and Pseudoplatystoma spp. are treated with salt prior to sun-drying.
Smoking
Smoke-drying is perhaps the most widespread way of preserving fish in Africa and is practised in
nearly all river systems. Techniques vary somewhat and the type of oven used changes from place to
place. In Malawi, fish are smoked in a special thatched smoke house, whereas in other regions ovens
constructed of clay are open to the air (Fig. 7.11). Small fish are usually smoked whole. Medium sized
fish are scaled, eviscerated, and sometimes split open or slashed down the sides. The largest fish are
cut into pieces before treatment. As they are smoked fish lose weight in about the same proportion (3
kg fresh fish = 1 kg smoked fish) as when they are dried. One treatment is rarely sufficient, especially
in more humid areas, consequently the fish have to be retreated at intervals of between a week and ten
days to keep them in acceptable condition. The smoke-dried product is often stockpiled over a
considerable period of time, especially in those temporary camps furthest from major centres of
communication.

249
Figure 7.11 Different types of smoking oven: (A) circular oven (Haussa); (B) rectangular oven made
of baked earth; (C) pit oven; (D) sections of rectangular and pit ovens (adapted from Blache,
Miton and Stauch, 1962)
In some areas the lack of wood for smoking is causing problems for the fishery. In the Sahel, for
instance, the availability of domestic firewood is becoming one of the factors limiting human
occupation. Studies carried out in Mali by the Operation Peche, indicate that about 1 kg of wood is
necessary to produce 1 kg of smoked fish, using traditional ovens. An improved oven is being
introduced in the Central Delta of the Niger, which will reduce the demand to 0.5 kg of wood for
every kg of smoked fish produced (Operation Peche, 1976). A similar type of oven is being used in
the development of the Elephant marsh fishery in Malawi where it is also intended to plant copses of
trees to supply them with wood; a copse of about 1 hectare is thought adequate to serve one oven
(Tweddle et al., 1977).
Fish Meal

250
The only river fishery whose catch is used for making fish meal is the Prochilodus fishery of the
Parana. According to Vidal (1967) these “Sabelerias” accounted for nearly 70 percent of the total
inland catch of Argentina, although these have declined considerably from about 1965 onwards.
Fish Oil
Several species of fish are particularly rich in fats and are traditionally used for extracting oil. Oil is,
for instance, a by-product of the “Sabelerias” fish meal factories. Brycinus leuciscus is exploited early
in the fishing season in the Niger for its oil, and several species of the Mekong, principally Cirrhinus
and Dangila are taken for the same purpose. The extraction process involves boiling for a certain time
to release the oil which floats to the surface and may be skimmed off. Dangila spp. can produce up to
15 percent of their weight in oil (Chevey and Le Poulain, 1940).
Fermented Products
Fermentation of the cleaned and gutted fish in water for 12 hours is a common preliminary to salting
in the Mekong, or to sun-drying in the Senegal River. In Asia a number of fermented products
including fish pastes and sauces with a high salt content are also produced.
PROTECTION AGAINST INSECT INFESTATION
One of the major curses of the fishing industry in many parts of the world is the infestation of fish
with insects. Moist fish is susceptible to be damaged by blowflies and their larvae in particular. Blow-
fly larvae consume fish flesh so it is only when the fish is sufficiently dried that it becomes
unattractive to the adult fly for egg laying.
Dried or smoked fish are subject to be attacked by beetles, mainly Dermestes spp. The level of losses
due to Dermestes spp is directly related to the length of storage of the fish.
Prevention measures include the re-heating or re-smoking of the infested fish in a temperature above
50°. The use of insecticides is not advisable as there is no safe insecticide officially recognized yet
that can be used on cured fish.
However, keeping the fish off the ground and improving the hygienic conditions of the processing and
storage areas as well as their surroundings can significantly reduce the problem of insect infestation.
In West Africa early attempts to control Dermestes infestations included scorching, whereby during
the hot smoking process the outer skin of the flesh was blackened and hardened to lessen the
successful penetration of the flesh by the beetle. Other techniques have included packing the fish in
bales wrapped with matting as soon as it is smoked, but such attempts have largely failed. The
Operation Pêche in Mali has claimed a certain success by treating the fish with natural insecticides
(Bioresmetrine) after drying or smoking but improved hygiene and heat treatment using a polythene
tent and solar energy and simply keeping fish off the ground probably provide more satisfactory
alternatives.
SPECIES CAUGHT
Because of the large number of species involved in river fisheries, detailed species lists serve little
purpose. Analysis of catch data from most gears shows them to conform to the canonical distribution
of species abundance already described. Distributions of this type predict that only a few species will
be dominant in the catch of any gear, and knowledge of these is essential, both for the management of
the stocks and for establishing priorities for research.

251
LATIN AMERICA
The river fisheries of Latin America have concentrated mainly on the low water “Piracema” or
“Subienda” migrations. The fisheries are still relatively undeveloped and have selected for species of
large size and consumer appeal. Consequently catches are composed of the larger characins or
siluroids. In the Magdalena River well over half the catch consists of Prochilodus reticulatus
(Granados, 1975) although Pseudoplatystoma fasciatum, Pimelodus clarias, P. grosskopfii, Brycon
moorei, Sorubim lima, Plagioscion surinamensis and Ageneiosus caucanus also contribute
significantly. The preoccupation with the subienda species in Columbia led to a neglect of other
potential food fishes and Bazigos et al. (1977) drew attention to an unexploited stock of
Hemiancistrus and Pterygoplichthys in the cienagas which could substantially increase the yield from
this system. Fish of the genus Prochilodus are the mainstay of other South American fisheries. P.
reticulatus forms the basis of a heavy fishery in the southwestern portion of Lake Maracaibo and the
inflowing floodplain river Catatumbo (Espinosa Giminez, 1974). In the Apure and Upper Orinoco
systems, the fishery concentrates on the larger Pimelodidae: Pseudoplatyptoma fasciatum, P.
tigrinum, Brachyplatystoma filamentosum and B. vaillantii (Canestri, 1972) but a greater diversity of
genera including Prochilodontids are found in the Lower Orinoco and Delta (Novoa, 1982) (Table
7.2).
Table 7.2
Main Commercial Fish Species from the Lower Orinoco (from Novoa,
1982)
Prochilodus mariae Cynoscion spp.
Pseudoplatystoma fasciatum Colossoma brachypomum
Semaprochilodus laticeps Colossoma macropomum
Mylossoma duriventris Brachyplatystoma spp.
Phractocephalus hemiliopterus Brycon spp.
Hydrolicus scombroides Sorubimichthys spp.
Arius parkeri Pellona flavipinnis
Hypophthalmus edentatus Pterodoras spp.
Pinirampus pinirampus Schizodon isognathus
In the Parana, Vidal (1969) listed 18 species as being the principal ones of commercial value (Table
7.3). Further upriver, in the Mogi Guassu, P. scrofa makes up 0–60 percent of the catch which also
contains 16 other species of characin (Godoy, 1975). Although the Amazon basin contains some 2
000 species, only a small proportion of these are captured by the fishery. Meschkat (1975) listed the
major commercial species, based on the catch statistics for Amazonas state (Table 7.4).
Table 7.3
Species of major commercial value in the fishery zone of Rosario, R. Parana (not
listed in order of importance) (from data in Vidal, 1969)
Lycengraulis olidus Colossoma mitrei Pimelodus albicans
Brycon orbygnianus Ageneiosus brevifilis Zungaro zungaro
Salminus maxillosus A. valenciennesi Luciopimelodus pati
Prochilodus platensis Oxydoras kneri Pseudoplatystoma fasciatum
Leporinus spp. Rhinodoras d'orbignyi P. coruscans
Hoplias malabaricus Pimelodus clarias Basilichthys bonariensis

252
The heavy and selective fishing pressure on the largest species A. gigas and C. bidens, had already led
to a sharp decline in the populations of these fishes by 1975 and Bayley (1981) posit that the catch of
all large species cannot be maintained at present fishing pressure. In exchange the share of the catch
contributed by species of such genera as Semaprochilodus, Prochilodus and Triportheus has, and will,
increase until they dominate the fishery.
Table 7.4
Species of commercial value in the Amazon (listed in order of
importance) (from data in Meschkat, 1975)
Arapaima gigas Leporinus and Schizodon spp.
Colossoma bidens Brachyplatystoma flavicans
Prochilodus insignis Brycon hilarii
Plagioscion surinamensis Oxydoras kneri
Plecostomus spp. Osteoglossum bicirrhosum
Brycon nattereri Cichla ocellaris
Colossoma spp. Hypophthalmus edentatus
Rhinosardinia spp. Pseudoplatystoma fasciatum
Prochilodus corimbata Astronotus ocellatus
AFRICA
In Africa, the river fisheries are exploited at a much greater intensity and a very broad spectrum of
species are caught, especially in the basins of the western side of the continent. In the Niger River gill
net and cast net catches can contain over 50 species and it is difficult to identify the major elements of
the catch. However, Raimondo (1975) listed the nine most important species of the Upper Niger
(Table 7.5).
Table 7.5
Predominant species in the catch from the Central Delta of the Niger
from data in Raimondo, 1975)
Alestes dentex Lates niloticus
Brachysynodontis batensoda Bagrus bayad
Hydrocynus forskhalii Mormyrus rume
Oreochromis niloticus Citharinus latus
Labeo senegalensis
Other species commonly represented are Auchenoglanis occidentalis, Clarias anguillaris, and
particularly Brycinus leuciscus which forms the basis of a specialized fishery for fish oil production.
Catches from other West African rivers have a similar combination of species. In the Senegal River,
Reizer (1974) investigated the number of species captured as a function of mesh in gillnets (Fig. 8.7)
and showed that the number of species increased as mesh size decreased. On the basis of these
experimental fishings it appeared that the ten most important species to the fishery were Schilbe
mystus, Lates niloticus, Alestes dentex and A. baremoze, Hydrocynus brevis, Labeo senegalensis,
Eutropius niloticus, Citharinus citharus, Heterotis niloticus and Hepsetus odoe. In some systems,
such is the diversity of gear that it is almost impossible to establish the true weighting of the various
species in the total catch. In these instances, studies of the abundance of fish in the markets adjacent
to the fishery give some idea. For instance, in the Oueme system the order of abundance of the
various species was as shown in Table 7.6 although as many as 40 species figured in the fishery as a

253
whole. This abundance is of course, biased by the food preferences of the fishermen themselves as
some of the species caught rarely reach the market.
Table 7.6
Order of abundance of major species appearing in the markets of the
Oueme valley
Clarias ebriensis Synodontis melanopterus
Clarias lazera Synodontis schall
Parachanna obscurus Schilbe mystus
Heterotis niloticus Distichodus rostratus
Mormyrids Ctenopoma kingsleyae
In the Chari-Logone system the elements of the catch are very difficult to separate from those
produced in the lake. However, Blache and Miton (1962) listed the principal elements of the catch
from a number of fishing methods in the Chari and Logone rivers. From these it appeared that the
migratory Alestes dentex and A. baremoze were by far the most important species to the fishery.
Several larger species were also of major importance including Citharinus citharus and C. latus,
Distichodus rostratus and D. brevipinnis, Labeo senegalensis, Hydrocynus brevis and H. forskahlii,
and Lates niloticus. Some smaller species were also important including Oreochromis galilaeus and
O. niloticus, Schilbe mystus, Brycinus nurse and divers Synodontis spp. In the swamps Clarias lazera
and C. anguillaris were particularly abundant. The drying of the Lake Chad basin induced by the
Sahelian drought has, however, radically changed this composition and the catch is now dominated by
tilapiine cichlids.
In contrast to the specific richness of West African fisheries, the rivers of East Africa produce only a
restricted variety. The Shire river fishery was found to have five species, Clarias gariepinus, Clarias
ngamensis, Oreochromis mossambicus, Marcusenius macrolepidotus and Eutropius depressirostris
which made up about 90 percent of the catch (Willoughby and Tweddle, 1977) despite the fact that
there are 39 species in the system. The Kafue fishery takes a greater number of species (18) but of
these, 6 contributed about 90 percent of the catch (Everett, 1974). These were: Oreochromis
andersoni, O. macrochir, T. rendalli, Serranochromis angusticeps, Schilbe mystus and Clarias
gariepinus.
ASIA
In the rivers of the Indian sub-continent there is one migratory species, Hilsa ilisha, which, in the
Indus, Ganges and Godavari systems, is the subject of specialized fisheries. Apart from this the
fisheries are based largely on a series of large cyprinids known as the major carps, as well as some
siluroids, ophicephalids and notopterids. The major Gangetic carps are Labeo rohita, L. calbasu,
Catla catla and Cirrhinus mrigala, and the principal siluroids Mystus aor, M. seenghala and Wallago
attu. In addition to these, Jhingran (1975) listed a further 12 species which contribute significantly to
the catch. A similar species complex occurred in the Brahmaputra river, with the addition of Labeo
gonius, Puntius sarana and Notopterus notopterus. In the Cauvery river a somewhat different group
of species dominated the fishery: Acrossocheilus hexagonolepis, Tor putitora, Barbus carnaticus,
Labeo kontius, Cirrhinus cirrhosa and Osteocheilus brevidorsalis amongst the cyprinids,
Glyptothorax madraspatanus, Mystus aor, M. seenghala, Pangasius pangasius, Wallago attu and
Silonia silondia amongst the siluroids, together with Channa (= Ophicephalus) marulius a murrel and
Notopterus notopterus.
Husain (1973) listed the 13 principal species of the lower Indus fishery as shown in Table 7.7
although these were drawn from a pool of 66 species. From the list it will be seen that there are many

254
elements in common with the Indian rivers, although O. mossambicus has been introduced from
Africa.
Fishing in the Chao Phrya river is largely done by stationary wing traps which block most of the main
channel (Tongsanga and Kessunchai, 1966). Of the 77 species commonly captured, Crossocheilus
reba made up about 60 percent of the catch. Other important species were Wallago attu,
Macrognathus aculeatus, Ophicephalus micropeltes, Ophicephalus striatus, Puntius gonionotus,
Pangasius sutchi and Cirrhinus microlepis.
Table 7.7
Main food fishes of the River Indus (after Husain, 1973)
Hilsa ilisha Rita rita
Notopterus chitala Mystus spp.
Catla catla Oreochromis mossambicus
Cirrhinus mrigala Channa (=Ophicephalus) marulius
Labeo calbasu Channa (=Ophicephalus) striatus
Labeo rohita Channa (=Ophicephalus) punctatus
Wallago attu
More than 150 of the total 800 species that inhabit the Mekong make up the bulk of the catch in that
system. Of these a few may be singled out as being particularly conspicuous or sought after. These
vary with the region of the river fished, and separate authors have identified different dominant
components. Pangasianodon gigas particularly is distinguished by its size but has diminished in
importance due to overfishing. Adopting the summary of Fily and D'Aubenton (1966) the species
listed in Table 7.8 made up over one percent of the catch in Cambodian waters.
Table 7.8
Species comprising over 1 percent of total catch in the Cambodian
water of the Mekong in order of importance (after Fily and
D'Aubenton, 1966)
Pseudosciaena soldado Ambassis wolffii
Cirrhinus jullieni Puntius orphoides
Cirrhinus auratus Puntius altus
Ophicephalus micropeltes Notopterus notopterus
Thynnichthys thynnoides Hampala macrolepidota
Kryptopterus apogon Puntius bramoides
Macrones nemurus Pangasius larnaudi
Cyclocheilichthys enoplus Wallago attu
Labeo chrysophekadion Clupea thibaudeani
EUROPE AND NORTH AMERICA
Catches in North American and European highland streams are both dominated by salmonid species
but richer faunas are to be found in the lowland reaches. In Europe these are based mainly on
cyprinids and of the 20 species listed from the middle of Danube by Liepolt (1972) and Holcik et al.
(1981) over half belong to this family (Table 7.9). In the lower Danube, as with other Black Sea and
Caspian rivers, migratory species, such as Huso huso, Acipenser ruthensis, A. stellatus, A.
guldenstaedtii and Alosa pontica make up a considerable part of the catch.

255
The Vistula in Poland contains many less species although the fishery does have some of the elements
present in the Danube. According to Backiel (1983) the following species appeared in order of
importance: Abramis brama, Vimba vimba, Anquilla anquilla, Esox lucius, Stizostedion lucioperca,
Chondrostomus nasus, Rutilus rutilus and Blicca bjoerkna. This relative poverty in species is
exaggerated by environmental degradation which has already eliminated some of the previously most
important anadromous fishes from the river.
The range of families appearing in the catches of North American lowland rivers is greater than in
Europe and the diversity of the catch is equally high as is illustrated by the commercial and
recreational catches of the Middle Mississippi (Rasmussen, 1979) (Table 7.10).
Table 7.9
Main commercial fish special fish species from the Middle Danube
Rutilus rutilus Alburnus alburnus
Aspius aspius Leuciscus cephalus
Vimba vimba Chondrostoma nasus
Abramis brama Leuciscus idus
Stizostedion lucioperca Cyprinus carpio
Esox lucius Barbus barbus
Silurus glanis Tinca tinca
Acipencer ruthensis Salmo trutta
Ctenopharyngodon idella Hucho hucho
Anguilla anguilla
Table 7.10
Species composition of Upper Mississippi Fisheries (a) commercial
and (b) recreational
(a)
Cyprinus carpio Ictiobus spp.
Ictalurus spp. Aplidonotus grunniens
Polyodon spathula Catostomid spp.
Acipenser spp. Lepisosteus spp.
Amia calva Hiodon tergisus
Esox lucius Anguilla rostrata
Ctenopharyngodon idella Pomoxis spp.
(b)
Stizostedion canadense Aplidonotus grunniens
Stizostedion vitreum Morone chrysops
Pomoxis spp. Lepomis macrochirus
Ictalurus punctatus Esox lucius
Cyprinus carpio Micropterus salmoides
Perca flavescens Pylodictus olivaris
Ictalurus spp. Micropterus dolomieu
Lepomis cyanellus Catastomid spp.
Ambloplites rupestris Centrarchid spp.
Dorosoma cepedianum Lepidosteus spp.
Amia calva Acipencer fulvescens
Ictalurus furcatus Hiodon tergisus
Polyodon spathula Anguilla rostratus

256
Scaphyrhnchus platorhynchus Lepomis gulosus
Morone mississippiensis
These examples show that even fisheries in rivers from the temperate zone can be based on a large
number of species.
FISHERIES FOR JUVENILE FISH
The very heavy exploitation of juvenile fish, in the form of fish of the year moving to the dry season
habitats at the end of the flood, is a particular feature of floodplain fisheries. In the Oueme, as in many
African and Asian fisheries, small mesh nets of various types are used intensively in the canals
draining the plain. Cross channel dams and barrages, such as those of the El Beid or the Barotse plain
are also designed for the capture of young fish. Durand (1970) estimated that up to 90% of the catch
by number and weight of the El Beid river was made up of juvenile fish moving from the Yaeres
floodplain towards Lake Chad. The “maalelo” fishery of the Barotse removed about 3.7 percent of the
juveniles of the 15 most important species each year (Bell-Cross, 1971). Regression analysis also
indicates that considerable proportions of the catch from African rivers (ranging from 90% in the
Shire and drought affected Niger to 50% in the Niger under more normal regimes) are repeatedly
drawn from 0+ fish. Many millions of fingerlings and fry of the major carps are withdrawn annually
from Indian rivers to stock reservoirs in their basins, and the capture of fry is common throughout
Asia for the stocking of floodplain depressions, rice paddies and culture ponds.
It is common prejudice that the removal of large quantities of juvenile fish will prove harmful to the
stock. The persistence of many of the fisheries indicates that there is little danger so long as the
practice is kept within reasonable limits. Both Reed et al. (1967) in his defence of the “atalla” fishery
for juveniles on the Niger river, and Bell-Cross 1971) in his analysis of the “maalelo” fishery, made
the point that, with the high mortality rates current among river fishes, the loss of a proportion of year
class 0+ fish is hardly liable to affect the final population at all. A theoretical analysis of such a
fishery (Welcomme and Hagborg, 1977) indicated that a high proportion of the juveniles can be
removed during the period of drain-off without damaging the fishery and in simulated fisheries, where
juveniles were exploited at the same time as adults, the combined catch exceeded the maximum catch
of either juvenile or adult fisheries on their own. Careful control of these fisheries is, however,
essential and further studies are needed on actual situations where the juveniles are heavily exploited.
CATCH
ANALYSIS OF CATCH IN DIFFERENT RIVERS
Catch as a Function of River Form
Catch statistics from rivers are often of low quality because of the difficulties inherent in collecting
data from fisheries which operate from many landings dispersed along a system that may traverse
several countries. As in most fields of fisheries, there is a need to improve the quality and quantity of
the catch statistics and the collection of biological and other data as a prerequisite for the proper
management of the riverine stocks. However, because of the lack of concise information on many of
the individual systems, attempts have been made to extrapolate general principles from the small
group of water bodies about which something is known. Some approaches to this have already been
discussed in the section which describes methods used to establish either standing stock, production or
yield in temperate rivers from a wide variety of ecological parameters. The information needed for
such exercises in tropical rivers and an alternative means of analysing catch patterns has been sought.
Despite the inadequacies of the data, an analysis of fish yield patterns from African rivers has given a
fairly coherent picture of the factors involved in determining the catch that can be expected from any
particular system (Welcomme, 1976). In the rivers used in this analysis, see Table 7.11 all of which
were moderately to heavily fished, there was a good correlation between the drainage basin area of the

257
river system in km² (A) and the catch in tons obtained from it (C). Excluding catches from
exceptionally large flooded areas, the sample conformed to the relationship:
C = 0.03A0.97 (r = 0.91)
Because the basin areas and the total length of the longest channel of the river are also simply related.
(Main channel length = 4.95 basin area0.45, in African rivers) this equation transformed into a
relationship for yield in t as a function of the main channel length in km (L):
C = 0.0032L1.98 (r = 0.90)
or approximately one three hundredth of the square of the length of the stream (L²/300).
Table 7.11
Main channel length, basin area and catch from African rivers
River Channel length
(km) Basin area (km²) Catch (t)
Nile 6 669 3 000 000 40 840
Congo 4 700 4 014 500 82 000
Ubangi 1 060 772 800 4 670
Kasai 1 735 342 116 7 750
Niger 4 183 1 125 000 30 000
Benue 1 400 219 964 12 570
Zambezi 2 574 1 300 000 21 000
Senegal 1 641 335 000 16 000
Gambia 1 120 77 000 3 000
Volta B. 650 45 324 1 560
Volta R. 260 6 871 370
Volta W. 255 6 602 70
Pendjari 330 11 226 140
Oueme 700 40 150 646
Mono 360 22 000 533
Tana 600 38 000 500
Bandama 950 97 000 3 408
Sassandra 650 75 000 1 518
Comoe 1 160 78 000 2 142
Rufigi/Ruaha 750 17 700 3 600
The catch of any reach of river of length x km at distance y from its source can be calculated from xCy
= Cy+x - Cy, where values of Cy can be obtained from the preceding equation. In its most extreme
form, where x = 1 km, this yields a theoretical equation for the catch that might be expected for any
kilometre or river at different distances from the source (i.e., catch at kmy = 0.0064y0.95). When catches
from rivers with extensive floodplains (those whose flood area exceeds 2% of the total basin) are also
included in the analysis, a second relationship emerges whereby C = 0.44A0.90. Values predicted by
these relationships are of course averages over a number of systems from which actual values from
individual systems deviate quite widely. Nevertheless these formulae have been used to predict catch
in a number of systems lying outside the original set. The Mekong, Danube and Magdalena rivers,
with floodplain areas of between 3 and 8 percent of their respective basins, are distributed around the

258
extensive floodplain line. Rivers such as the Indus, Mogi Guassu and Pongolo with no extraordinary
development of their floodplains fit the ‘normal’ relationship nearly exactly. Furthermore some
independent estimates of potential, such as that obtained by Paiva (1973) from the Parnaiba river,
Brazil, of 7370 t/yr for a basin area of 362 000 km², agree completely with the predictions of the
formulae. However, caution should be exercised in their application of rivers outside Africa owing to
the different relationships between basin area and channel length for the various continents.
Deviations from the mean regression line may arise for two main natural reasons. First, there is a
possibility that edaphic factors may influence the basic productivity of a system in such a manner as
to determine yield. There is so far no strong evidence for this in the data set available on savannah
rivers, although there is more than a strong suspicion that certain types of poor river, such as the
tropical black waters, are indeed less capable of supporting sustained fishing than is normal. Second,
morphological factors have been shown to determine the yield characteristics of river systems. The
very fact that it has proved necessary to derive two different relationships for yield against basin area
indicates the predominant role of floodplain area in determining the productivity of a system. This
factor alone accounts for 70% of the differences between actual and predicted catch in different
reaches of the same river. From the data in Tables 7.12 and 7.13 mean catch levels of 17 fully
exploited African floodplains were 54.7 ± 36.5 kg/ha/yr. Six Asian floodplains had mean catches of
44.02 ± 17.9 kg/ha/yr and, given the means and standard deviations of the two populations they can
be considered one set which can be merged. When data from the Orinoco and Magdalena rivers,
which also lie within the limits of confidence, are added, the mean yield for the 25 rivers is 51.55 ±
32.1 kg/ha/yr. With the present data it is not possible to detect any differences between the tropical
continents but at 18.7 kg/ha catches from the temperate Danube are slightly lower. However, the
catches from 1481 km of the Upper Mississippi river (Rasmussen, 1979) show that the 171 136 ha of
pooled river, produced 6499 t of fish (mean of 25 yrs) equivalent to 37.8 kg/ha. In this section of the
river there is a succession of 26 pools in which the lower ends have lacustrine characteristics and the
upstream portions have retained their floodplain. Catches obtained from the three blackwater rivers
for which data are available indicate catch levels of between 11 and 19 kg/ha/yr which are
considerably less than the mean for savannah rivers. When catch is plotted against floodplain area for
the 25 tropical floodplains which are exploited at a reasonably intense level, a relationship:
Table 7.12
Maximum inundated area and catch from some floodplains additional to the
information in Table 7.11
River Area (km²) Catch (t) CPUA
(kg/ha)
AFRICA
Niger Benin 242 1 200 49.59
Niger C.D. Mali 20 000 90 000 45.00
Massilli 150 475 31.67
Pongolo 104 400 38.46
Yaeres 7 000 17 500 25.00
Kamulondo (Blackwater) 6 639 7 355 11.08
SOUTH AMERICA
Orinoco 228 1 000 43.80
Amazon (Peru) 9 960 13 700 13.80
ASIA
Mahaweli 121 413 34.13
Bangladesh 93 000 727 000 78.17
Lubuk Lampan 12 29 24.17
Lower Mekong 54 000 220 000 40.74
Ganges 1958–61 296 1 538 51.96

259
Ganges 1962–69 296 1 430 48.31
EUROPE
Danube 26 450 49 400 18.68
C = 4.23A1.005
is obtained. Although the best fit is a power curve the exponent is sufficiently close to 1 as to make
the relationship almost linear. It does, however, indicate that large floodplains may be marginally
more productive per unit area than smaller ones, and values range from 42.8 kg/ha for a 1 hectare
plain to 44.6 kg/ha for a 5 million hectare plain. These figures all confirm the earlier estimates of
between 40 and 60 kg/ha/yr that may be expected from tropical floodplains.
One question that frequently arises is the degree to which the catch of reservoir rivers or modified
rivers which have no annual flood, compare to the levels of production of flood rivers. The only two
fully fished rivers of this type for which data are available, the Mississippi and the Nile, lie within the
same range of values appropriate to their level of exploitation as do flood rivers. More data are
needed, however, to clarify this as the dynamic processes underlying the fisheries of such rivers are
likely to differ considerably from those in rivers having a pronounced seasonal flood.
Catch as a Function of Fishing Intensity
Standard models relating catch and effort, such as those of Schaefer and Fox, have been used
successfully in some fisheries of small rivers or in the analysis of fisheries based on single species, for
example those for Colossoma (Petrere, 1983) or Plagioscion spp. (Annibal, 1983) both of the Amazon
basin. Examination of catch and effort data in large rivers, however, generally illustrates the
difficulties of applying such models to complex fisheries which are pursued with a variety of methods
on the whole fish community in any river or river reach. Because of the scarcity and poor quality of
most catch and effort data from river fisheries any analysis of the factors regulating catch must be
regarded as somewhat speculative. Two approaches can be adopted for the analysis of the data
available. First, data from individual rivers can be studied to identify any relationships arising from
changes in temporal fishing patterns. Second, data from a number of rivers can be grouped for
treatment as one set with a range of differing regimes. Data sets from individual rivers are rare and
only two sets, one from the Mississippi and one from the Nile, have so far been traced. Similar power
curves fit the data for catch per boat for 30 years from the Mississippi:
(CPUE = 29731E-1.24: data from Rasmussen, 1979)
and for catch per boat for 10 years from the Nile:
(CPUE = 11117E-1.03: data from Borhan, 1981) (Fig. 7.12).
Curves of this type have been described previously for individual fisheries, for instance by Beverton
(1959) who used one to describe the response of Tilapia to increased fishing in Lake Victoria. When
22 observations from 16 of the worlds rivers are compared (Fig. 7.13, Table 7.13) the single best fit
regression line is also an inverse power curve of the form CPUE = a E-b although the value of the
relationship is limited by a lack of points in the middle range (5 – 20 fishermen/ha). The inclusion of
data from various continents and climatic zones appears justified at this time as the points for the
Magdalena River, Ganges and Misissippi all lie well within the confidence limits of the African data.
The relationship between effort and catch per unit effort is perhaps better described by a curve within
the log-log plot which may be interpreted as two different regression lines calculated from all points
between 0.05 and 3 fishermen/km2, where CPUE = 2.72E-0.47, and between 3 and 30 fishermen/km2,
where CPUE = 10.2E-1.09. Separate plots of total catch against catch per unit effort extrapolated from
the two regression lines cross over at about 8 fishermen/km2.

260
The form of these relationships may be explained in three ways:
(a) That there is an interference effect between fishermen as densities increase, whereby gears
compete with one another for an increasingly limited resource. This would mean that fishing mortality
is no longer linearly related to nominal effort.
(b) That the measures of effort customarily adopted for ease of recording, i.e., numbers of fishermen
or numbers of canoes is unsatisfactory. This may be particularly significant in artisanal and
subsistence river fisheries where fishermen have alternative occupations. They may then allocate the
time spent on fishing according to its rewards relative to such occupations, in which case fishing
mortality is again no longer linearly dependent on the variable selected to represent effort. Here some
other measure, for instance fisherman/days, might be a more appropriate index. Such measures
demand an increased effort in surveying the fishery which exceeds the financial possibilities of most
developing countries. In many river systems the switch between farming and fishing is in any case
dictated by the exigencies of the agricultural cycle, which may leave unstructured time which can
only be filled by fishing. Further verification of these questions rests upon detailed socio-economic
studies in such communities but the possibility signals the need for caution in adopting such measures
of effort for management.
Table 7.13
Number of fishermen, catch and maximum flooded area of some floodplains
River No.
fishermen Catch (t)
Area
(km²) No. fisherman
/km²
Catch
fisherman
/year (t)
CPUA (kg/ha/
year)
Seasonally inundated
AFRICA
Shire 1970 2 445 9 545 665 3.68 3.90 143.53
Shire 1975 3 324 7 890 665 5.00 2.37 118.65
Kafue 1963 1 112 8 554 4 340 0.26 7.69 19.71
Kafue 1970 670 6 747 4 340 0.15 10.07 15.55
Oueme 1957 29 800 6 500 1 000 29.80 0.22 65.00
Senegal 10 400 30 000 5 490 1.89 2.88 54.65
Niger Mali 1971 54 112 90 000 20 000 2.71 1.66 45.00
Niger Niger 1965 1 314 4 700 630 2.09 3.58 74.60
Niger Niger 1982 3 200 3 200 600 5.33 1.00 53.33
Niger Nigeria 4 600 14 340 4 800 0.96 3.12 29.88
Benue 5 140 9 570 3 100 1.66 1.86 30.87
Pendjari 65 140 40 1.63 2.15 35.00
Barotse 912 3 500 5 120 0.18 3.84 6.84
Rufigi 3 000 3 589 1 450 2.07 1.20 24.75
Kilombero 341 4 536 6 700 0.05 13.30 6.77
Logomatia 70 300 600 0.12 4.29 5.00
SOUTH AMERICA
Magdalena 1978 30 000 65 000 20 000 1.50 2.17 32.50
Orinoco 200 1 000 228 0.88 5.00 43.80
Amazon (Peru) 3 360 13 700 9 960 0.34 4.08 13.80
ASIA
Ganges 1 600 1 480 296 5.41 0.93 50.00
Mahakam 8 000 14 500 7 178 1.11 1.81 20.20
Stabilized
NORTH AMERICA
Mississippi 1950 3 036 4 037 1 711 1.77 1.33 23.59
Mississippi 1960 1 977 5 337 1 711 1.16 2.70 31.19
Mississippi 1970 2 232 5 256 1 711 1.30 2.35 30.72
AFRICA

261
Nile 3 725 8 410 800 4.66 2.26 105.13
(c) There is a real effect at the fish community level, whereby the yield curve concept as applied to
individual species is not applicable. Some of the changes occurring in exploited fish communities
described below indicate that such an effect does occur.
Figure 7.12 Catch per unit effort as a function of effort in (A) the Nile and (B) the Mississippi

262
Figure 7.13 Catch per unit effort as a function of effort for 17 rivers: Solid line = regression for all
points; dashed lines = best fit lines for catches from 0.05–3 fishermen/km² and 3–30
fishermen/km²
The form of these relationships also implies that the relationship between effort and total catch shows
an initial rise followed by a somewhat flat line during which there is little change in catch over a
considerable range of effort (Fig. 7.14). The Nile downstream of Aswan, for instance, produced 8 410
± 542 t for ten years despite a threefold increase in effort. Plainly such a plateau can be extended
indefinitely neither to the right nor the left of the relationship, and it is suggested that curves of catch
and catch per unit effort have three phases as set out in Fig. 7.15.

263
Figure 7.14 Plots of total catch (---•---) against effort for the Nile. Also shown is catch per unit effort
against effort (---o---)

264
Figure 7.15 theoretical diagrams of changes in catch and catch per unit effort as a function of effort,
letters explained in text
(1) Initiation phase: Limitations on catch per unit effort, and hence total catch, are set independently
off the relationship by the physical capacity of the fisherman to handle his catch. Observations from
all over the world on both lacustrine and riverine fisheries indicate that this capacity rarely exceeds 1
– 4 t per year per fishing unit although exceptionally up to 13 t/yr may be handled. The attainment of
maximal levels at low fisherman densities depends much on the state of development of the society in
which the fishery is pursued. Thus, whereas in advanced economies sufficient support exists to permit
the fisherman to land and market large catches, in developing economies individual efficiency may be
poor for lack of efficient gear and the absence of infrastructure for landing, marketing and
transportation of the catch. Fishing therefore tends to remain a subsistence activity. As the numbers of
fishermen increase the creation of markets will give rise to a rapid expansion of the fishery.
(2) Sustained exploitation phase: During this phase, catch levels are maintained in the face of
increasing effort. At the same time changes occur in the composition and abundance of the exploited
fish community which lead to a progressive shift in catch composition from large to small species.
(3) Collapse phase: Few cases have been documented where the fishery has been intensified to the
point of total collapse of the fish community. It is to be supposed that the fishery becomes
uneconomic long before and most reported overfishing has occurred where larger and highly favoured
species have declined while the rest of the fish community remains intact or underexploited.
It is conceivable that real collapse at the community level can occur under exceptionally heavy
exploitation. This may be impelled by socio-economic factors, as in the case of the Oueme, where the
disappearance of nearly all the larger species, the very small size at capture and the loss of catch over
the decade 1957–68 would suggest a community at the limits of its tolerance. The exceptionally high
fisherman densities of 30/km2 in this river result from a populous ethnic group which is confined to a
floodplain area by demographic pressures. Other drastic declines in fisheries have been recorded in
the African Sahel where the failure of the floods over a number of years has resulted in a decrease in
natural productivity in rivers such as the Niger, Senegal and Chari/Logone. The fish stocks, thus
diminished, have been called upon to support an increased fishing pressure as other food sources have
disappeared.
CHANGES IN COMMUNITY STRUCTURE WITH INCREASING FISHING
PRESSURE
The form of the relationship between catch and effort, whereby there is an initial rise followed by a
more or less prolonged plateau where catch remains stable in the face of increasing effort, masks a
series of adjustments in the fish community. Unfortunately systematic monitoring is still not carried
out in most river fisheries. As a consequence theories on the impacts of fisheries in rivers with
complex multi-species stocks must be derived from a number of river and lake fisheries. Experience
has shown that a similar succession of events, which has come to be termed the fishing up process,
occurs as fishing pressure is applied and increased. It is a common observation that there is a
progressive disappearance of the larger species as pressure is applied. Similarly, increases in fishing
intensity and total catch were correlated with a reduction both in mean size and as a proportion of
catch of the large pimelodid catfishes of the Orinoco river (Novoa, 1982), whereas the proportion of
smaller Semaprochilodus in the catch increased to more than compensate for the deficit. In the
Amazon at Manaus larger species such as Arapaima gigas and Colossoma bidens have progressively
disappeared from the catch and have been replaced by increased quantities of smaller prochilodontids
and clupeids as well as new species such as the pimelodid catfishes. Lates niloticus, Heterobranchus
longifilis and Bagrus docmac have completely disappeared from the heavily fished lower reaches of
the Oueme River, and other large species such as Citherinus latus, Distochodus rostratus, Labeo
senegalensis and Heterotis niloticus diminished in abundance to be replaced by small catfishes,

265
characins and mormyrids. Sritingsook and Yoovetwatana (1977) remarked on the decline of the larger
migratory species such as Probarbus jullieni from the Mekong River and its tributaries.
It is useful to examine the history of the Oueme fishery in more detail as one of the best examples of
community overfishing in rivers. The total catch of the Oueme floodplain was estimated at 10 400 t in
1955–57 (CTFT, 1957). By 1968–69 this had fallen to an estimated 6 484 t (FAO/UN, 1971). The fall
was accompanied not only by the shifts in species composition noted above, but by a fall in catch per
unit effort in most gears as summarized in Table 7.14. There was some increase in total effort during
the period of study (about 19%) but this was insufficient to account for the drop in CPUE. Larger
mesh fishing gears were eliminated from the fishery by 1968 and the mean weight of the species
caught decreased in most of the gears that remained in use. For example, the medium-mesh cast net
caught fish of 124.9 g in 1955–57 but the mean weight had dropped to 26.5 g by 1968–69. Similarly
the small-mesh net caught fish of 25.2 g in 1955–57 but only 9.8 g in 1968–69. The only gear in
which catches increased was a trap for Macrobrachium, a detritus eating freshwater prawn that was
previously heavily preyed upon by the fishes.
Table 7.14
Changes in catch per unit effort in gear from the Oueme river between
1955–57 and 1968–99
Catch per unit effort Percentage
Type of gear 1955–57 (g/h) 1968–69 (g/h) changes
Cast nets
large-mesh 1 186.0 249.8 -79
medium-mesh 694.4 372.0 -46
small-mesh 589.5 322.6 -45
Longline 286.0 193.0 -33
(g/d) (g/d)
Traps
Clarias trap 74.0 43.2 -42
Macrobrachium trap 115.0 309.0 +269
Other changes have been noted from a number of systems. In Lake Tanganyika, for instance,
increased fishing led to the elimination of the major predator (Lates) with a considerable increase in
total catch from the small pelagic clupeids and smaller predator (Luciolates) which form the basis of
the fishery (Coulter, 1970). In Lake Victoria the fishery was being pushed from a dominance of large
tilapiine cichlids towards a preponderance of small haplochromine cichlids and even smaller
Rastrineobola before the spread of introduced Nile perch reversed the trend. Further changes occurred
within the haplochromine population under fishing pressure from trawls with the disappearance of
larger piscivorous and molluscivorous species and a progressive reduction in size in the remaining
planktivores and limnivores (Witte and Goudswaard, 1984). The establishment of a trawl fishery in
Lake Malawi also resulted in the successive displacement of the species caught to smaller and smaller
lengths (Turner, 1981). Certain types of environmental pressure would appear to mimic or to reinforce
the effects of increased fishing pressure, which makes it difficult to fully interpret the cause of
observed changes. Thus in the Caspian Sea there has been a long standing tendency toward smaller
species in the fishery in the face of sustained effort and environmental stress although the level of
catch has remained the same (Carre, 1978). Similarly, Regier and Loftus (1972) were able to trace a
succession of alterations in the composition of the fish communities of the N. American Great Lakes
arising from a combination of fishing, ecological and environmental pressures.

266
From this evidence, it may be supposed that individual species within the community pass through a
typical production curve of catch against effort. The succession of such curves agrees with the
observations that exploitation eliminates first the larger individuals of the larger species then the
larger species themselves (a and b in Fig. 7.16). This selective disappearance tends to be due to a
preference, both by fishermen and consumers, for larger individuals, as well as the lower capacity of
the long-lived larger species to support high levels of fishing mortality. In an effort to sustain catch
the fishing methods tend to be replaced by those which capture smaller fish. As the few largest
species are removed from the fishery larger numbers of intermediate-size species (c-j in Fig. 7.16). In
other words, the community is pushed successively from K dominance to r dominance. A feature of r
selected species with short life spans, high fecundity and high productivity is that they fluctuate
considerably in abundance. It is, therefore, reasonable to suppose that as a fishery becomes more and
more dependent on only a few such species it may become destabilized with large fluctuations in
catch with time. Indeed, trajectories of this sort are characteristic of fisheries for pelagic clupeids
throughout the world.
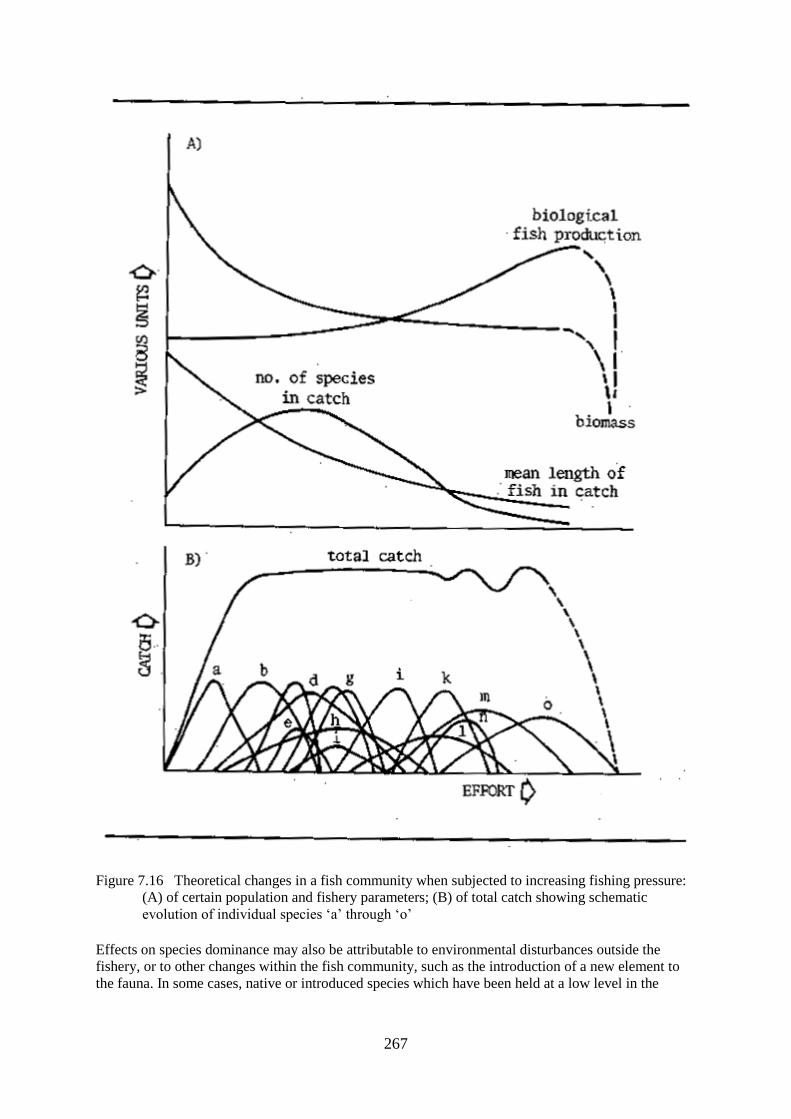
267
Figure 7.16 Theoretical changes in a fish community when subjected to increasing fishing pressure:
(A) of certain population and fishery parameters; (B) of total catch showing schematic
evolution of individual species ‘a’ through ‘o’
Effects on species dominance may also be attributable to environmental disturbances outside the
fishery, or to other changes within the fish community, such as the introduction of a new element to
the fauna. In some cases, native or introduced species which have been held at a low level in the

268
community are able to expand to occupy a dominant position when indigenous species are overfished
or when the habitat is changed in their favour by human action.
In river communities where there is as paucity of species at lower trophic levels, as shown by high
Predator/Forage fish ratios, the capacity of the system to benefit by such changes is limited. Similar
limitations have been observed in nutrient poor systems such as tropical blackwaters where the
amount of nutrients accumulated within the fish community represents a significant proportion of the
total nutrient pool. Here removal of the larger predators, whether fish or crocodiles, has even been
held accountable for the declining long term productivity of the environment (Fittkau, 1973). In such
systems rapid declines in catch may follow the onset of exploitation as happened in the Kamulondo
depression, where catch fell from 9 063 to 4 810 t in the space of four years after the introduction of
an intensive fishery based on nylon gillnets (Poll and Renson, 1948). More recently similar rapid
declines on individual fishing grounds have been observed in the blackwater tributaries of the
Amazon. Eventually even in the most productive of systems the capacity of the fish community to
absorb further increases in exploitation pressure will be exceeded, recruitment will fail and numbers
and biomass will drop sharply.
During the period when yield changes relatively little with increasing exploitation, the ichthyomass
will tend to remain relatively constant or to decrease as the species composition favours those with
higher turnover rates. At the same time biological fish production will increase. The tendency for
ichthyomass to remain constant may be reinforced in flood rivers where only a certain proportion of it
is able to survive through the dry season anyway. The number of species which make up the catch
will initially increase, although in some regions market preferences may be restricted so that only a
few stocks may be fished. When these are reduced, it may take some time before the market can be
reoriented to replacement species. Until this happens catches may fall and fishing be reduced, as
happened in the Parana River with the decline of the ‘sabalo’ (Prochilodus platensis) fisheries and in
temperate rivers where salmonid stocks were reduced by eutrophication. Where the market demand is
highly diverse, as it is in most African and Asian countries, the number of species in the catch will
first rise rapidly and then diminish as the more susceptible forms succumb to fishing and other
pressures.
These changes are most noticeable in fisheries using mesh selective gears, such as gill nets, cast nets
or seine nets, where changes in the size and species composition of the stock are accompanied by the
classic pattern of the successive replacement of large mesh gear by smaller and smaller mesh sizes.
Such changes are more difficult to detect in traditional fisheries which from the outset exploit a broad
spectrum of species and sizes with a variety of gear. The available evidence from rivers such as the
Oueme indicates that the use of gears designed to capture larger species and individuals progressively
declines as the fishery becomes heavily exploited.
FLUCTUATION IN CATCH BETWEEN YEARS
The available evidence indicates that the year-to-year variations in fish stock abundance are directly
linked to the degree of variability of the hydrological regime. Thus in stable systems such as reservoir
rivers or those with flood control the magnitude of the standing stock varies little from year to year,
whereas it is apparent that the catches from flood systems fluctuate in a manner that is in some way
dependent on changes in the flood cycle. Such changes in yield were noted early in the study of river
fisheries when Antipa (1910) concluded that the fisheries production of the Danube delta was directly
proportional to the extent and duration of the floods. This sentiment was echoed by subsequent
workers on that river (Botnariuc, 1968; Holcik and Bastl, 1977). That a similar state of affairs existed
in tropical rivers was suspected by many workers including Wimpenny (1943) in the Nile prior to its
complete control.
Models of production as a function of water regime would suggest that year-to-year variation in catch
arises from two main sources. First, the amount of water in the system at low water can affect catches

269
of the same year by altering the ease with which fish are captured or can influence catches in
succeeding years through dry season mortality. Second, the intensity of the flood determines the
magnitude of the stock in future years through differences in recruitment, survival and growth. In
certain rivers it can also alter the ease of access between the channel and the floodplain water bodies
thus altering catch within the same year. In combination these effects render analysis difficult and
obscure trends over time but sufficient examples of each exist to illustrate these various cases.
Improved catches during exceptionally low waters are well known from most fisheries as the
shallower waters favour the capture of fish. Usually such comments take the form of qualitative
statements such as those of Vidy (1983) when describing the Logomatia fishery of the Yaeres
floodplain. The relationship has been quantified in the Magdalena River as:
Catch y(t) = 171779.36 - 23706.17 Water Level y
for the whole fishery (Arias pers.comm.) and by Annibal (1983) as:
Catch y(kg) = 56937 DDFy - 164206: (r = 0.84)
for the Plagioscion fishery of the Lago do Rei of the Amazon basin, where DDF is an index of the
amount of water remaining in the system during the dry season.
The amount of water remaining in the system during the dry season may also influence the catches in
subsequent years as was shown by the University of Michigan et al. (1971) for the Kafue Flats
fishery. Here the relationship:
Cy = - 6630 + logeDDFn-1: (r = 0.77)
which was originally formulated for the years 1956–71, has continued valid through 1983 (Hayward,
1984) despite the construction of two sets of flow control dams in the intervening period.
Sufficient data are available from the Kafue and Shire floodplains and from the Central Delta of the
Niger for an assessment of the effects of the intensity of the flood on fish catch. Unfortunately none of
these sets of data include reliable estimates of changes in fishing effort, which may themselves be
responsible for some year-to-year variations in catch.
In the Kafue river records are available from 1954 until the present. However, the fishery was not
judged to have reached maximum expansion until 1958 (Muncy, 1978), and the flood regime was
changed following the closure of the Kafue Gorge dam in 1972. This was followed by the
construction of an upstream dam at Itezhitezhi and the two dams, acting together substantially altered
the duration but not the periodicity of flooding. Calculations based on the years 1958–71 gave good
correlations between catch in year y and the flood of the preceding year (y - 1) (r = 0.72) or the flood
two years previously (y - 2) (r = 0.71). Muncy (1973 and 1978) presented a complete analysis of the
various factors also finding good positive correlations between catch in year y and flood intensity in
the preceding years (y - 1 and y - 2). Although detailed analyses were not continued after 1971 the
predictions of these regressions are consistent with the greater catches that have been obtained
because of the increased flooding following the installation of the two dams.
Sets of data for water level and catch are also available for the Shire river (1969–73) and the Central
Delta of the Niger (1966–74) which were analysed by Welcomme (1975). This analysis used a simple
sum of all weekly mean water levels which exceeded the bankfull stage. The results showed a highly
significant correlation to exist between catch in year y and the flood regime in y - 1 in all three
systems. Correlations of catch with flooding in the same year were not so satisfactory. Because the
fisheries of most tropical rivers are based on fish that are one or two years old, it might be expected

270
that the flood regime in the both preceding years might exert an effect on catch of any year. When the
regressions of combined and weighted hydrological indices from both receding years were tested
some improvements in correlation were noted. As a result it was finally concluded that catch in year y
is best explained by a combination of the flood histories of the two preceding years. Thus the best fit
linear regression lines which are plotted in Fig. 7.17 were as follows:
Kafue: Cy = 2962 + 70.54 (0.7 HIy-1 + 0.3 HIy-2)
Shire: Cy = 5857 + 38.11 (0.9 HIy-1 + 0.1 HIy-2)
Niger: Cy = 3239 + 32.10 (0.5 HIy-1 + 0.5 HIy-2)
Differences in Hydrological Index accounted for 57 percent of the variation in catch between years in
the Kafue, 82 percent in the Shire and 92 percent in the Niger. Subsequently the Niger fishery has
continued to show a strong correlation with flood regime and a data set for the 19 years from 1966 to
1984 yields a linear correlation:
Cy = 19.172 + 0.027 (0.7 HIy-1 + 0.3 HIy-2) (R² = 0.84)

271

272
Figure 7.17 Best-fit regression lines for the relationship between catch and flood regime for: (A) the
Kafue Flats; (B) the Shire river; (C) the Central Delta of the Niger at Mopti
which HI is calculated from the mean six monthly flood discharge on the Kaulikoro gauge. A linear
relationship was not at best felt, however, and an improvement of correlation to R² = 0.87 was
obtained with the regression:
Cy = 151.73 log (0.7 HIy-1 + 0.3 HIy-2) - 428.26
The log-normal nature of this regression indicate a slight lessening of the response of fish
communities as flood densities increase over a certain level. Other rivers of the Sahel also showed
decline in catch over its period of the drought (Table 7.15)
Table 7.15
Catches of fish in the Senegal river and the Central delta of the Niger
(data from Senegal, Direction des Eaux, Forets et Chasses, 1976 and
Laboratoire d'hydrobiologie de Mopti, 1981)
Year Senegal Niger Upper Chari
1966 110
1967 30 98
1968 25 105 20–25
1969 20 107
1970 18 107
1971 18 94
1972 15 88
1973 12 73 20–25
1974 21 63
1975 25 87
1976 89
1977 87
1978 77
1979 83
1980 88
1981 75
1982 73 4.0
1983 61 1.8
1984 54 0.8
There is considerable interest in identifying the relative role of the two components of the
hydrological regime in determining productivity although it is sometimes difficult to separate them
statistically. For example, where the intensity of flooding is highly correlated with the severity of
drawdown, as in the Kafue (r = -0.78), it is difficult to determine whether the apparent dependence of
catches on the preceding drawdown is a statistical artefact. Some indication of the relative roles might
be deduced the Shire river, which shows a lesser correlation between drawdown and flood (r = -0.45)
and also shows a lower correlation between drawdown and catch. However, this should not be taken
to imply that the high water phase is the more influential of the two phases in all systems. The
difference in response could equally be due to differences in the amount of water remaining in the
system at low water relative to the amount at peak flood. In the Kafue the dry season area was about
27 percent of that during the floods, whereas in the Shire the equivalent figure was 48 percent. It may
be surmised that the more stringent the drawdown, as reflected by the lessened percentage of residual
water during the dry season, the greater the influence of the low water regime on catch in the next

273
year. This is supported by the predictions of Welcomme and Hagborg's (1977) simulation of a
floodplain and its fishery which predicted that the more water remaining in a system at low water, the
more the differences induced by variations in the high water regime are transmitted to subsequent
years.
Analysis of catch relative to the flood history of a river has been carried out independently by several
workers. Catches in the Orinoco gave two relationships depending on species (Novoa, 1982). Thus the
total catch over 15 years was:
Cy = -634.8 + 0.53 (0.5 HIy + 0.5 HIy-1) : (r=0.62)
and the catch of Prochilodus mariae over 10 years was:
Cy = -389.4 + 0.19 (0.5 HIy + 0.5 HIy-1):(r=0.73)
In this river a second relationship between catch and flooding was noted for Semaprochilodus laticeps
where, because more water bodies were in contact with the river channel in higher floods, a logistic
relationship was found between an index of flood intensity (HI) and catch during the same year:
Catch was also found to be related to flood height in the Madeira river by Goulding (1981). Here
catch at low water was improved when low water flows were less than average. Heavier floods are
reported by the fishermen to have the effect of releasing fish imprisoned within floodplain water
bodies and increasing the recruitment, the fishery thus leading to better catches in the source as
subsequent years.
Flood strength has also been found to influence catches in temperate rivers. Ivanov (quoted by
Chitravadivelu, 1974) also noted similar effects in the Zofin arm of the Danube, but with a lag time of
one year.
Because of the number of factors involved, for instance changes in fishing effort or the reduction in
the age and length of first capture associated with the fishing up process, it is frequently difficult to
interpret these relationships by direct regression analysis, although visual comparison of plots of flood
regime and catch indicate some relationship to exist. Krykhtin (1975), for example, obtained excellent
relationships between flood strength and catch in the Amur River where the best correlation was
obtained with the flood regime three to four years previously. Dunn (1982) provided graphical
evidence for a correlation between an index of catch in year y and of the flood five years previously
for the Hilsa fishery of Bangladesh. Hilsa, which is a marine anadrome, is thought to enter the fishery
in its 4th or 5th years. In analysing these results fluctuations between 1966 and 1976 are best
explained by a combination of y-5 and y-6 hydrological indices (r=0.76) but between 1976 and 1981
there is little clear correlation. Similarly, graphical inspection of Rasmussen's (1979) catch data as a
function of the Upper Mississippi floods shows there to be parallel trends but this data set is less
consistent and did not yield a conclusive regression.
The lag between the year of flooding and the time when its effects are reflected in the catch are
probably dependent on the time taken for fish to grow to the size range captured by the fishery. In
tropical rivers this is very short, often less than a year, because of the small size and rapid growth of
many of the species caught, and also because of the heavy fishing for fish of the year as they leave the
floodplain. An example of this was provided by Benech and Quensiere (1984) in their studies of the
fishery of the El Beid river which drains the Yaeres of Cameroun. Here there was a strong positive

274
correlation between the quantity of juvenile fish moving down river and the floods of the same year.
The same study showed the level of production to be independent of the species composition when a
regular sequence of floods exists. Thus in the El Beid the relation of production to the intensity of
floods compared well between a series of readings from 1968 and another series between 1974 and
1978, although the population structure was very different. By contrast, when the regular series of
floods is disturbed, aberrant patterns may be detected in the following years. Ivanov also remarked on
the high proportion of young year classes in the Danube fishery one year after high floods and a
corresponding drop in catch a year after particularly poor flow. In some rivers where growth is slower
and the larger species are favoured by the fishery, fish may take two or more years before they are
susceptible to the gear. In the Amur river, for instance, Krykhtin proposed that the effect on catch is
only felt after the incorporation of 20–30 percent of the new year class into the target stock.
The fisheries of some reservoirs with highly fluctuating water levels seem to behave like floodplains.
In the Parakrama Sumadra of Sri Lanka a 20 year study by Scheimer (1983) has shown the effects of
fluctuating water levels on fish catch to be described by a relationship:
Cy = 232.2 + 16.2 HIy-3
In the lake fish take from 2–3 years to enter the fishery. This data together with that of Moreau (1980
and 1982) indicate that reservoirs and shallow lakes with highly fluctuating water levels are
ecologically homologous to floodplains.
Cy = 232.2 + 16.2 HIy-3
On the basis of the obvious correlation between the intensity of flooding and catch it is tempting to try
to predict catches in future years from the flood of the year using regression formulae similar to the
ones above. To test this it may be assumed that if one can indeed predict the catch in any one year
from its flood or the flood of preceding years the accuracy of the prediction would improve as an
increasing number of years of data are added to the regression. When the data for the Niger were
treated in this way the accuracy of forecast did improve, from which it was concluded that it is
possible to validly predict catches in river systems from regression analyses of the past performance
of the fishery provided enough terms are available. In the case of the Niger, at least 14 years data were
needed to accurately predict future trends; such predictions assume that there are no major changes to
the resources base through overfishing, environmental change, etc. Likewise, correlations between
water levels in years y and y+1 and catch in the Czechoslovak reaches of the Danube, were used by
Holcik and Bastl (1977) to predict future catches from hydrological flow data. Because of the
importance of such predictions both for the regulation of the fishery and for the establishment of
criteria for mitigation of falling catches due to upstream flood control structures further work on this
topic is desirable.
CHAPTER 8
MANAGEMENT OF RIVER FISHERIES
EFFECTS OF OTHER USES OF RIVERS AND THEIR BASINS ON FISHERIES
Because the river is only one component of a larger system, the basin, fish communities in rivers are
affected not only by events occurring within the channel and its associated waters but are also subject
to a range of external influences. Precipitation falling within the basin eventually finds its way into the

275
river by surface and sub-surface flow carrying with it a variety of materials including the topsoil and
any contaminants it might contain. Changing conditions in the basin can produce differences in water
quality and quantity, as well as in loading with silt and other material which can directly affect
channel form. Many such effects are the result of natural variability, particularly climatic shifts but
more frequently such changes are the result of some human intervention. In river reaches with
floodplains the situation is more complex as the floodplain has a dry season phase during which time
terrestrial use is made of space which is covered with water during the flood. Because of the fertility
of their soils, and their proximity to water courses for irrigation and transport, floodplains are much in
demand for a wide range of activities. They are also regarded as having high development potential
and efforts to ‘reclaim’ them by flood control are widespread. This chapter examines some of these
effects and also describes the observed impacts on river fish communities of uses of the river and its
basin other than fisheries.
CHANGES IN FLOW
Rivers respond to changes in mean annual flow by a period of adjustment to the new regime after
which they stabilize in a form adapted to the altered conditions. Clearly reductions in flow will result
in the progressive restriction of the stream to a smaller bed within the original channel with a
concomitant loss of habitats for fish and other aquatic organisms. Conversely, overall increases in
flow will lead to the enlargement of the river channel by erosion of former banks and other features;
these, in time, should lead to an increase in habitat diversity through extension of the aquatic system.
Biotic sensitivity to changes in water regimes are not limited to alterations in absolute quantity but
also to the distribution of flow in time as food peaks can be moved to different times of the year or
even suppressed altogether.
The living aquatic organisms in rivers are usually adapted to the particular patterns of flow found
there; consequently changes in flow will produce changes in the biotic components of the ecosystem
quite apart from those arising from the contraction or extension of the aquatic habitat. Fish
communities tend to be either limnophilic or rheophilic, depending on the type of water regime
prevalent in the river reach in which they live. Changes in flow will, therefore, tend to favour one or
other of these communities, increasing flows leading to a larger presence of rheophilic species,
decreasing flows encouraging colonization by limnophilic species. Adequate flows are also essential
for the breeding and migration of many fishes. Physiologically fish respond to flood conditions3 by
becoming sexually ripe and by movement to breeding grounds. Conversely, should the appropriate
flow regime not occur, eggs may be resorbed, as in the case of the Rutilus rutilus, Abramis brama and
Cyprinus carpio of the Arakum lakes described by Shikhshabekov (1979). Certain critical levels of
flow are also needed to maintain certain types of breeding substrate in a suitable condition for
spawning. For this reason, there has been much concern over the determination of minimum flow
requirements for fish, particularly in rhithronic areas where high value angling resources coincide
with high demands for water. Examples of such research are discussed by Stalnaker (1980) who has
identified three main approaches to the general problem of evaluating instream flow requirements: (a)
rule of thumb and hydrographic analyses, (b) physical habitat analyses and (c) crop-flow analyses. An
example of the rule of thumb approach is the ‘Montana method’ described by Tennant (1976) (Table
8.1) whereby the effect of the various flow regimes relative to the original regime are described.
Table 8.1
Instream flow regimes for fish and related environmental resources (after Tennant, 1976)
Description Base flow regimes
(% of original average flow in periods)
October-March April-September
Flushing or maximum 200 200
Optimum range 60–100 60–100

276
Outstanding 40 60
Excellent 30 50
Good 20 40
Fair or degrading 10 30
Poor or minimum 10 10
Severe degradation 10–0 10–0
The above figures have been found to describe the effects of flow reductions on a wide range of North
American rivers. Similar series have possible applications elsewhere in the world but, unfortunately,
there is insufficient data in the tropics upon which to base such indices.
In Potamon reaches the problem of maintenance of flow is especially critical as the yield of systems
with floodplains is closely linked to the extent of flooding. Thus, if at any time the floodplain fails to
be inundated recruitment fails in that year for all those species which spawn on the plain.
Furthermore, the fertility of the plain during its dry phase is conditional on its being flooded for at
least part of the year.
Even if average flows within the system remain unaltered changes in the timing or form of the flood
may have grave consequences. In many fish species breeding success depends on a coincidence of
characters of which flow is but one. Consequently races or strains of species have adapted to a
particular timing in their breeding and displacement of the floods to a different time of year may not
permit them to reproduce. There are, however, signs that in some systems fish may adapt to the
altered flow regimes following dam construction. In the Volga River, for example, changes in the
timing of floods and in temperature of the water led to shifts in the spawning time and to an increase
in the duration of the reproductive period in many cyprinid and percid species, even to the extent that
a twin-peaked spawning period has appeared following reservoir construction (Kutznetzov and
Fedeyev, 1979). Equally, abrupt changes in flow characteristics can influence breeding success
adversely. Overly rapid rises and falls in water level can leave nests or spawning grounds dry at
critical periods or can result in eggs or fry being washed away. The precipitous decline of the flood
can result in fish being trapped in temporary water bodies for lack of time to find passage to the main
channel of the river.
CHANGES IN SILT LOAD
Increases in silt load resulting from changes in land or water use accelerate the natural evolutionary
processes of the river system, but in doing so cause a number of problems. In the rhithron deposition
of fine particles of silt on what is normally a coarse substrate suffocates the rheophilic organisms that
normally inhabit such reaches, cutting down on the availability of food. Such choking of the substrate
may also render it unsuitable for spawning by those species requiring swift, well aerated flows and
clear pebble or gravel bottoms. The silt provides an anchorage for vegetation, blocking low order
streams and even diverting them into new courses. Further downstream deposition of silt on levees
and on the river bottom may lead to progressive elevation of the whole channel until it stands above
the level of the surrounding plain. An extreme flood in a channel so encumbered may cause the river
to jump its bed changing its course by some kilometres. Excessive silting of floodplains chokes the
standing waters, which disappear faster than new ones can be generated by erosion. Similarly,
channels and dead arms are filled and new channels are cut to such an extent that the whole delta of a
river may shift along the coastline. At the same time coastal deltaic floodplains grow rapidly,
especially at their seaward end, where new land continuously appears.
Heavy silt loads also directly affect living organisms. The poor light penetration into silt laden waters
reduces the depth to which phytoplankton can develop and shade out submersed vegetation. Choking
of bottom substrates has also been implicated in the disappearance of benthic organisms in the
potamon as well as being identified as the cause of mortalities of fish scattering eggs in such areas.

277
CHANGES IN WATER QUALITY
The succession of physical and chemical conditions in rivers from the headwaters to the mouth may
be regarded as a natural eutrophication process. Most downstream reaches are normally enriched, and
further loading with nitrogen, phosphorus and organic compounds from agricultural, industrial and
urban sources appears acceptable up to a point, and may even be beneficial in initially impoverished
systems. If the capacity of the ecosystem to satisfy the BOD is exceeded, however, conditions can
deteriorate rapidly. Fish communities inhabiting potamon reaches of rivers are usually adapted to
eutrophicated conditions and can support a measure of deoxygenation, although some of the more
active species can disappear, Godoy (1975) for instance, traced the disappearance of Triurobrycon
lundii from the Mogi Guassu river to deoxygenated conditions produced by such eutrophication.
General pollution with toxic substances has not so far been widely reported from tropical systems,
although the situation in the rivers of India described by Patil (1977) leaves little room for
complacency. Here several rivers are suffering from severe contamination with industrial effluents
which have adversely affected their fish stocks. Severe local pollution can occur, however, and is
commonly associated with mining where seepage and direct discharge of toxic wastes can create
fishless zones in the rivers affected.
In temperate rivers the situation has been more serious and the widespread degradation of the quality
of water in many European and North American rivers has been widely described. Even large rivers
such as the Rhine or the Vistula have deteriorated to the point where many elements of their fish
fauna have been lost. Liebman and Reichenbach Klinke (1967) also recorded bad conditions in certain
reaches of the Danube caused by domestic and industrial pollution, but the self-purifying capacity of
this large and swift flowing river has been sufficient to keep the main stream at an acceptable quality.
Studies on the floodplain, however, indicated that conditions in some standing waters can deteriorate
to a point where they damage the fish stock, although in others moderate eutrophication by sewage
can raise the productivity.
The effects of pollution on the aquatic life of the system may be summarized as:
(i) lethal toxicity which kills fish at some stage of its life history. In the case of floodplain rivers this
may be indirectly in that the reduction of dissolved oxygen in standing waters of the floodplain and
river channel may make them unsuitable for fish that normally live there;
(ii) sub-lethal effects which are usually difficult to detect or prove but which alter the fish's
behaviour in such a manner as to prevent it completing its normal life cycle, or simply to reduce its
growth or increase its susceptibility to disease;
(iii) cumulative effects which can render fish either unsafe or unpalatable for consumption. Most
pollution effects tend to be very broad affecting many different species. Whatever their immediate
effects, the response at the community level is a reduction in diversity and a shift in species
composition towards relatively smaller, shorter-lived forms. In other words they tend to mimic the
changes expected from heavy fishing and are therefore apt to reduce the amount of fish available to
the fishery. Moderate enrichment with organic substances, on the other hand, can increase the amount
of fish (ichthyomass) supported by the system.
INTERACTION WITH OTHER USES
Terrestrial Animals
Wildlife

278
Many species of game animals move on to the floodplain during the dry season in search of the rich
grazing to be found there. Certain species are more specialized to this habit than are others, and some
are virtually limited to the floodplains for their distribution. Such species as the Lechwe (Kobus leche
kafuensis) migrate from the centre of the plain toward its periphery as the flood rises, and return in the
wake of the falling water (Fig. 8.1). Their distribution is, therefore, a mirror image of that of the fish
and their dynamics are somewhat similar with a maximum in lambing as the water leaves the plain
exposing new pasture (Sayer and Van Lavieran, 1975). Before the construction of the new dams on
the Kafue river there was a stable lechwe population of about 94 000 individuals, but changes in the
regime following the closing of the Kafue Gorge dam has reduced the amount of grazing available
and most probably will cause a drop in the number of individuals supported by this plain. According
to Gonzalez-Jimenez (1977) the capybara (Hydrochoerus hyrochaeris) exists in huge numbers on the
floodplains of Latin America where it feeds on grasses, and also eats aquatic plants. Unlike the
lechwe it swims well and inhabits permanently swampy areas.
Figure 8.1 Distribution of the Kafue lechwe: (A) at maximum flood, April 1971; (B) at low water, 2–
14 September 1971. (After Sayer and Van Lavieren, 1975)

279
Floodplain wildlife is apt to be affected adversely by alterations to their environment in the same way
as the fish. As a further example of this, Attwell (1970) noted some effects of the Kariba dam. Here
the Mana floodplain is of great importance for the conservation of wildlife in the valley. Since the
closure of the dam a lesser area of the plain has been flooded and the remaining portion is under
severe pressure of utilization by the larger species of mammals which has produced changes in the
vegetation. The impact of overpopulation is reinforced by changes in the type of grasses favoured by
the new flood regime, which are tougher. Growth of plants is also less lush as the rich alluvium which
used to fertilize the plain is now removed from the Zambezi water before it reaches the plain. By
stabilizing the flow the dam has reduced the ecological dynamism of the river whose discharges are
wrongly timed, disrupting reproductive patterns of the mammals. A similar theory to this appears in
many systems where mammals and waterfowl have suffered by the drying up of the wetlands (Smart,
1976) and will also emerge when we examine the consequences of dam building to the fish.
The presence of wild ungulates and hippopotami on the floodplain has been considered by Kapetsky
(1974) to be beneficial to the fish and we have already seen the importance attached by Fittkau (1973)
to the crocodile populations of Amazonian rivers for the maintenance of balanced communities. In
fact, the amount of nutrients recycled by the terrestrial components of the system is probably very
large and must have some effect on its overall productivity. It has been suggested that the tendency
for wildlife to disappear in favour of cattle early in the development of the plains, lowers this
productivity as the cattle deposits less of their wastes directly into the aquatic system.
Cattle
Most unmodified floodplains are used as ranges for cattle during the dry season. In certain areas
seasonal migrations of the cattle herding peoples dominate the demography. Rzoska (1974 and 1976)
has described the way in which the pastoral Nuer and Dinka of the Nile Sudd migrate away from their
permanent villages on the higher ground of the swamps following the receding water. As they
progress they burn the dead and drying aquatic vegetation to obtain the fresh shoots upon which the
cattle feed. At the beginning of the flood they return to their island villages after migrating distances
of up to 80 km. Similar movements have been noted from the Central Delta of the Niger (Gallais,
1967; Fig. 8.2) and the Okavango swamps (Stannard, pers. comm.), and are a prominent feature of all
African wetlands. Patterns do differ in some plains, for instance, the Oueme, where the characteristic
“lagunair” cattle are confined to the levees which are ditched or fenced off from the back swamp
depressions. During the flood the cattle are corralled on artificial islands and fed with aquatic
vegetation cut from the floating mats on the plain. Extensive cattle ranching is also a feature of Latin
America, and at present the vast “llanos” of Venezuela and Colombia, which are drained by the
Apure, Arauca and Meta rivers, are used mainly for this purpose. Because the llanos are either
submerged by sheet flooding or extremely arid, the Venezuelan ranchers are erecting large crescent
shaped dykes called “modulos” across the plain. These trap and retain the water as it retreats towards
the main river channels and by evaporation and filtration they slowly dry out leaving a well-watered
fringe of vegetation available for the cattle. The “modulos” also tend to trap and retain fish although
their potential as fish collecting and rearing devices needs to be explored.
Large numbers of cattle are present on the plains, Rzoska (1974) recorded some 625 000 head as
being present in the Sudd. On the Barotse plain there were 310 000 head (FAO/UN, 1969), on the
Kafue 250 000 cattle grazed the plain from May to October (FAO/UN, 1968a), on the Shire
floodplains 148 000 head are present in the Elephant and Ndinde marshes, and in West Africa on the
Central Delta of the Niger, Gallais (1967) estimated some 200 000 head, whereas on the Gambia
floodplain there were 300 000 head. In general cattle and fisheries are compatible, even
complementary utilizations of the plain. The dung dropped by the cattle, estimated at about 500
kg/ha/yr (Shepherd, 1976) converts much of the dry season primary production into readily dissolved
organic and mineral nutrients which have an important impact on the chemistry of the flood waters.
Several workers have noted the enrichment of some standing waters of the floodplain by cattle that
use the lakes for drinking. The exceptional fertility of the Bangula lagoon on the Shire has been

280
attributed to this cause, and according to Gilmore (1976) lagoons of the Okavango frequented by
cattle had more than three times the standing crop of more normal pools, 700 kg/ha as against 200
kg/ha. Some adverse effects of cattle have also been noted. Extensive use of portions of the Oueme
floodplain for pasture have resulted in trampling and breaking down of the banks of the drain-in
ponds found there, with their later abandonments and filling through siltation (Hurault, 1965). On the
flooded banks of small channels and streams, clearance of vegetation for intensive grazing may result
in a lessening of cover with a consequent reduction of fish population. Gunderson (1968) cited the
case of a Montana stream where ungrazed reaches had 76 percent more cover, and the brown trout
living there were more numerous and had about 44 percent more ichthyomass than those then in the
reaches where grazing was common. The literature on the interactions between grazing in riparian
environments, particularly those associated with smaller water courses, and the quality and quantity of
water in United States streams has been surveyed by Platts (1981). It is clear that improper grazing
practices, involving high densities of animals over prolonged periods degrades bottom land and
associated water courses. Changes include loss of vegetation cover, instability and erosion of stream
banks, increase in silt load and consequently in siltation of channels and a decrease in water quantity.
Because of these changes, fish populations, in these cases consisting of salmonids, are much higher
(by 200–400%) in ungrazed than in grazed reaches of lower order streams (order 6 or less).

281
Figure 8.2 The seasonal migrations of the pastoral peoples of the Central Delta of the Niger prior to
the Sahelian drought. The numbers refer to separate ethnic groupings. (Adapted from Gallais,
1967)
Forestry
Because forests tend to conserve water, topsoil and nutrients, they exert a conservative effect on the
aquatic systems draining forested landscapes. The removal of the trees has several readily observable
effects, notably on the water yields in streams, the timing and nature of runoff and the production of
silt. Other effects are less immediately discernible but may nevertheless be important and include
increased concentrations of nutrients, changes in pH and water temperature and more recently
introduction of pollutants into the system.
Deforestation of the catchment area of rivers leads to changes in the flood characteristics whereby
flood peaks tend to become higher and shorter as run-off is decanted straight into the channels. In
forested slopes much water is retained by the vegetation and also in the top soil. As the top soil
disappears there is nothing to delay the water in its move down slope. The faster rise and fall, the
more unpredictable spiky flood regimes and the lower dry season flows are detrimental to many
species of fish which require a smoother transition from one water phase to another. The lack of top
soil and the exposure of the bedrock also tend to lower the amount of nutrients entering solution. The
conductivity of the water drops leading eventually to impoverished conditions in the river. Such
changes are summarized in Table 8.2.
Table 8.2
Changes in stream flow resulting from various land management practices
Treatment Mean
precip.
(mm)
Annual
flow (*) Annual water yield response (stream flow)
Conversion of
Forest to grassland 1 850 840 Grass cover yielded equal stream-flow as former
forest when vigorous but declining grass gave 27
mm more water than forest
Brush to grass 480 30 Increased 127 mm
Abandoned
farmland/ 1150 640 Decreased 25 mm
grassland to forest
Hardwood to conifer 1950 686 Decreased 200 mm after 15 years
Sclerophyll scrub - 490 Increased 142 mm after 8–12 years
to conifer
Natural event
Forest destroyed 580 150 Increased 89 mm
by fire
Forest management
Clearcut regrowth 1220 710 Increases of 284 mm first 3 years
prevented by
herbicides for 3
years
Clearcut 1524 534 Increased 130 mm first year
Clearcut 760 280 Increased 112 mm first 5 years
Selection cut 2030 1290 Increased 55 mm
Strip cut 1220 710 Increased 32 mm after first two- year phase and 114
mm after second two-year phase

282
Regrowth after
clearcut
1221 737 Decrease 52 mm during 7 years of regrowth
* Rise in level in mm caused by increased flow of 1 1/m²
The process of logging itself contributes to the degradation of stream quality. With bad logging
techniques excessive amounts of waste timber and soil enter the stream causing increases in stream
bed load, suspended sediments and dissolved solid concentrations (Graynoth, 1979).
Clearing of the forests allows sunlight to fall on the aquatic system. Thus in rivers in the North
Western United States total salmonid standing crops were significantly greater (some 1.5x) in
deforested sites than in forested areas, although there were also associated charges in year class
strength and species composition of the community. Similarly the more rapid recycling of nutrients
through the repeated growth and death of floodplain grasses on floodplains free of trees would suggest
higher levels of productivity on savannah plains.
It is now apparent that many of the great open savannah floodplains of the present day were once
lined with gallery forests and the plains themselves were covered with scrub forests of the bush
savannah type. Clearance of trees for agriculture, grazing and firewood has denuded these and this
process is still continuing in some of the Latin American plains. As the process of denudation is
historically slow it is difficult to assess the impact of these changes on the fish populations although
these have undoubtedly occurred. It seems probably that the rivers have become less stable with more
frequent changes of channel. The flood regimes would also have changed as extensively forested
plains tend to retain the flood waters longer. There would be a reduction too in the amount of
allochthonous food available to the fish. Nevertheless, most studies seem to indicate that most
savannah plains are indeed more productive than those that are forested possibly because they are at a
younger stage of their succession. However, deforestation may lead to lowered productivity in some
cases such as that which took place in the Grand Lac of the Mekong. Here the surrounding forest was
cleared for agriculture and for firewood, and was accompanied by a decline of the fish catch by about
half over twenty five years. Sao Leang and Dom Saveun (1955) attributed this to erosion and siltation
in the basin and the lessened availability of allochthonous food arising from the reduced area of
forests. The silting led to increased turbidity in the lake through re-suspension of particles by wave
action in the increasingly shallow water body. This in turn led to a drop in primary productivity which
was reflected in a lowering of the fish populations. According to the information now available this
process was halted and reversed during the late 1970s when political troubles brought about the
collapse of population in the Khmer republic. The area around the Grand Lac became re-afforested
and the fish populations increased again.
Much of the increased silting noted in tropical rivers in recent years has been traced to the denudation
of land in the upper reaches of their basins. Here the steeper slope of the land encourages erosion and
the top soil is rapidly lost especially where marginal agriculture is practised (Eckholm, 1976).
Reforestation of hill slopes has been proposed for the control of soil erosion, the protection of
rhithronic streams and for the eventual diminution of siltation in the lower courses of rivers. Locally,
however, reforestation programmes may not be universally beneficial, as experience in Scotland have
shown. Here plantations of Sitka spruce (Picea sitchensis) decreased the pH of streams flowing over
quarzite, schist or slate by collection of acid contaminants from rainfall (pH c. 4.3 – 4.5). In such
streams salmon eggs die within a few weeks and trout are also absent. In adjacent unforested streams
good trout populations exist (Harriman and Morrison, 1982).
Agriculture
The new deposit of alluvial material during the floods each year, a valuable input of organic nitrogen
from blue-green algae and a mobilization of phosphorous and potash through the alternation of

283
aerobic and anaerobic conditions (Bramer, 1980) make flood-plains some of the richest of agricultural
lands. For this reason, they have attracted man's attention from early in his history as a farmer and
some of the earliest civilizations apparently arose in response to the need for communal control of the
flooding of the Nile, the Mesopotamean rivers or the rivers of the great plains of China. Where human
population density is low, simple culture is common whereby small plots are cleared of the moribund
aquatic vegetation as the floods subside. Cereal crops such as maize, sorghum or millet are usually
sown and, aided by the high water table, grow to be harvested before the next flood. At this level of
exploitation little modification of the plain is needed and the cultivated areas are dispersed among
wooded plains. As pressure on the land increases, the floodplain woodland is progressively cleared
and the environment further modified by drainage or irrigation. The needs of intensive dry season
agriculture lead to the filling in of many floodplain depressions and reclamation of permanent
swamps. To assist in this drainage canals are dug which dry out permanently wet areas, but these also
hasten the run-off from the plain so as to lengthen the growing season. Irrigation systems often have
to be installed in floodplain areas deficient in water or to compensate for water lost through improved
drainage. There is thus an increasing tendency to control the flooding of the plain either by poldering,
which keeps the water out of some areas, or artificial levee construction which keeps the river channel
within its banks and stops the annual flood. To complete the control, upstream flood control dams and
reservoirs are also built. Levels of cropping at different intensities of exploitation of bottom lands
have been summarized by Thompson (1983) as:
- Static subsistence 300–600 kg/ha/yr
- Slash and burn 1500 kg/ha/yr
- Intensive cropping 6000 kg/ha/yr
In its simple state agriculture does not conflict with fisheries and on such rivers as the Oueme in
Africa a pattern of intensive use has evolved where both activities are pursued together. On the
floodplain the maize fields are interspersed with drain-in ponds, the banks of which are used for
various types of market culture, tomatoes, peppers or green vegetables. In addition, the higher levees
which follow the river are used for grazing cattle, giving a balanced economy which supports a dense
population. Fig. 8.10 illustrates the typical arrangement of these activities according to Hurault (1965)
who described the land use patterns of this area in some detail.
Unfortunately as agriculture is intensified beyond this level and pursued as a sole objective, the
effects on the aquatic system of the flood control works that become necessary are far reaching. One
result, for instance, is that when the floods are controlled depositions of alluvial silt no longer fertilize
the soil which can quickly become exhausted. One finds a decrease in productions such as that
experienced downstream of the Kainji dam, where within ten years of its closure 50 percent of the
land was no longer suitable for agriculture (Adeniji, 1975). To compensate for this applications of
fertilizer are needed, putting up costs, and raising the risk of eutrophication and eventually pollution
of the water courses. Furthermore, in regions where there is relatively little water and where there are
high evaporation rates, salinization of the soil arises either through repeated flooding with salt water
or though the drawing to the surface of ground waters whose evaporation leaves the salts in the
surface layers of the soil. Should there be insufficient flood water to flush the accumulated salts into
the river, the ground rapidly becomes unsuitable for crops and eventually totally barren. As yet little is
known of the salinity tolerance of floodplain biota, but the salinization of lagoons definitely inhibits
most freshwater forms and, although in regions near the coast colonization with halophytic species of
plants may occur, the ecosystem is in some measure simplified.
The hydrological regime of a floodplain environment is well-suited to rice culture. This may be at the
primitive level of floating rice such as is practiced in the Central Delta of the Niger, but in Asia, the
adaptation of the lowlands of the river systems to intensive rice growing is one of the major features
of the landscape. Intensive rice culture requires a complete control over the hydrological regime.
Flood control dams, polders and networks of irrigation and drainage ditches are therefore installed for
this purpose.

284
Heckman (1979) has examined in considerable detail the ecology of rice fields in Northern Thailand
which become naturally colonized by numerous fish species which contribute to the crops of the area.
In fact the capture of fish in rice paddies in Thailand amounts to some 65-80 kg/ha/yr (figures similar
to those estimated for natural flood-plains), which means that for the 8 million ha of paddies, the total
production could be as much as 580 000 t/yr, most of which is for home consumption. His findings on
general ecology of rice fields (Fig. 8.3) may be taken as representative not only of cultivated semi-
aquatic ecosystems but also, to a certain degree, of natural floodplains as well. The nutrient flow
within the system emphasizes the enormous proliferation in the biomass of rooted aquatic plants
relative to other elements of the community. Evidently in the natural system the offtake toward man
and domestic animals would be absent and this block of nutrients would be returned to the floodplain
by the natural processes of decay.

285
Figure 8.3 General overview of ecosystem nutrient flow in rice culture: (A) early rainy season (1
May); (B) late rainy season; (C) cool season (16 October); (D) dry monsoon (5 December).
(After Heckman, 1979)

286
Because rice growing and fisheries use the same phase of the hydrological cycle there are sometimes
considerable conflicts between them. Fish are frequently accused of destroying rice and Matthes
(1977) investigated the ways in which the fishes of the Niger river interact with this type of culture.
He found that many species were present in the rice fields most of which were juveniles. Certain
species were, however, present as adults and these could be classed in three groups:
(i) fish feeding predominantly on rice, of which four emerged as the most destructive, Alestes
dentex, A. baremoze, Distichodus brevipinnis and Tilapia zillii of which the last two nibble through
the stalks at mid-height thus cutting down the whole plant;
(ii) fish feeding occasionally on rice, particularly Brycinus nurse which bites at the leaves of young
plants, and Oreochromis niloticus which nibbles epiphytic algae from the stems of the rice plants but
also tears away at the stalk;
(iii) species causing occasional damage through other activities such as Heterotis and Gymnarchus
which construct nests of the stems, or Clarias, Heterobranchus and Protopterus which uproot small
plants when probing the mud in search of their benthic food. Some species probably benefit the rice
by seeking out and eating the stem borer and other insect pests or by cleaning the stems of epiphytic
vegetation.
In the Central Delta of the Niger fishermen construct low dykes, 50 cm high, which protect the young
rice during the early part of the rising flood when it is most vulnerable to attack and these work
reasonably well, as by the time fish can penetrate the field the rice is past its most tender stage. For
complete protection, permanent dams with screens are needed to exclude fish from the field entirely,
but these all too often succeed only in sealing fish in rather than out.
Rice culture may affect fisheries adversely in two ways. First, the need to control insect pests has
encouraged the use of insecticides leading to possible pollution of the water in the rice field and
downstream of it. Second, the modifications of the environment associated with intensive rice culture
are detrimental to the fish. The fact that rice and fish are not necessarily mutually exclusive is shown
by the widespread practice of fish culture in rice fields.
Urbanization
In most parts of the world early human communities were established along the courses of rivers and
streams and about 5 000 years b.p. the systematic colonization of the flood-plains of the Nile, Indus,
Yangtse, Tigris and Euphrates rivers lead to the growth of man's earliest civilizations. With
heightened demand for land brought about by population growth, the pressures for physical
occupation of the lower part of water courses has increased despite the risks inherent in the season
flooding of such areas. This breeds the need for flood control which in turn encourages further
occupancy of the plain to the point where a considerable proportion of the population is located on the
100 year flood-plain. For example, about 16 percent of the urban areas of the United States lie within
such areas, and of this over 50 percent is developed for industry or human habitation (Sabol, 1974).
Small wonder, therefore, at a certain preoccupation with flooding in these areas as flood losses mount
annually despite the best efforts to contain them. An analysis of the world occupancy of floodplains
(UN, 1969) summed up as follows: In Western Europe new flood losses are mounting slowly as a
result of the intensified use of areas in major cities which are subject to large but infrequent
overflows. In Eastern Europe there is also an increase in flood losses though somewhat slower than in
North America. In South and Southeast Asia the intense use of long settled land and the development
of new floodplains have more than offset the effects of major river control works. In South America,
there is widespread encroachment of urban growth on floodplains. The situation in Africa is similar
for, although the older settlements were sited away from the main flood zones, new cities are
expanding into these areas. The result of this gradual invasion is a growing tendency to control floods.

287
The construction of cities tends to produce local disturbances in discharge as the existence of large
areas of impermeable surface accelerate run-off in the vicinity of the city, but such effects are still
slight in the basin as a whole. Some changes to flood-plain morphology also follow from the
communication systems which support the city and its surrounds. Navigation by commercial craft on
the river often requires some measure of regularization of flow including the installation of weirs and
locks, and the canalization of exceptionally tortuous stretches to make passage easier. Wash from
boats erodes banks and accelerates siltation and the destruction of marginal habitats. Roads and
railways cut across the floodplains, usually on raised embankments which act as polders or dams to
seal off large areas of the plain restricting movements of fish and, even more important, containing
the flood waters within a smaller area. Urbanization has two additional effects which also bear on the
aquatic environment. One of these, the growing need for power, has similar results to the flood control
measures in that it is associated with upstream dam and reservoir construction. The second, loading of
the waters with organic and inorganic substances, produces pollution and eutrophication in the waters
at the level of the city and downstream of it.
Hydraulic Engineering
Control of the flood in rivers is thought necessary for many of the foregoing uses, and is increasingly
pursued as a major objective in the development of river basins. Three main types of structure are
used either separately or together:
(i) dams and impoundments;
(ii) levees;
(iii) canals;
These all have impacts on the riverine and associated ecosystems upstream and downstream of the
structures which have caused considerable concern in a number of fora and have found expression in
such publications as Ward and Stanford (1979) or Petts (1984). The main effects on fisheries of such
structures are briefly reviewed in Table 8.3 and in the following sections.
Dams and impoundments
The storing of water behind a dam so that it may be released more slowly throughout the year is
probably the most popular of flood control devices. It has the additional advantage that the water can
do work for the generation of power and that it can be used for irrigation or cattle watering.
Furthermore, fish can be grown in the sometimes extensive water bodies that are retained. In size, the
reservoir can range from vast lakes such as that backed up behind the Akosombo dam on the Volta
river, which has a total flooded area of 8 500 km², to small dams which desiccate completely in the
dry season. Some major flood rivers such as the Volga, the Missouri and the Columbia have already
been converted into a chain of reservoirs, and others such as the Danube, the Indus or the Mekong
seem destined for that fate in the near future. The durability of such barrages is questionable as silting
is proceeding at a considerable rate in many rivers and the smaller dams have a predicted useful life of
only a few years before their ability to control floods diminishes progressively. Eventually the
reservoir will fail in its purpose, whether it be water storage or flood control. Larger reservoirs, of
course, may take much longer to silt up, and may last for over a century but the process is progressing
faster than predicted in many areas. As major sites are something of a non-renewable resource the
success of a policy of hydraulic control through the use of dams seems somewhat dubious in the long-
term. Meanwhile, dams produce their changes on the floodplain and the fish. These are brought about
by alternations in the flood regime and by changes in silt loading which in turn alter the dynamics
determining the channel shape. This in turn affects the distribution and persistence of vegetation in the
downstream stretches of river. For example, below the Volta dam the more stable hydrological
conditions and lack of scouring during the flood favoured the rapid development of extensive stands
of submersed vegetation such as Potomogeton octandrus and Vallisneria aethiopica (Hall and Pople,
1968).

288
The flood regime may be suppressed completely, or at least altered in magnitude so that it does not
flood such extensive areas. Very large amounts of floodplain have been lost in this manner as a few
documented examples will show. In the Missouri, the amount of floodable wetlands over a 145 km
sample reach of river dropped from 15 167 ha in 1879 to 7 414 ha in 1967, a loss of 67 percent of the
area (Whitley, 1974). The Illinois river, too, lost about half of its floodplain as 80 939 ha of the
original 161 874 ha were drained between 1903 and 1920. Subsequently, 3 238 ha have been restored
as lakes. Following the closure of a dam on the Peace river, the 2 560 km² of the Peace-Athabaska
delta were transformed from a thriving floodplain environment into a series of isolated mud flats
(Blench, 1972). Although drainage is not so advanced in other parts of the world the intentions are
there. Liepolt (1972) stated that all of the 26 450 km² flood-plain of the Danube will eventually be
drained for irrigated agriculture and the original floodable area of the river is already much restricted.
In the Mekong the intention is to eliminate flooding from the 49 560 km² delta area and 1 480 km²
have already been lost below the Pa Mong dam site. Similarly, in the Senegal river most of the 5
000km² valley floodplain will become dry after the dams which are at present being planned in the
head-waters are built. This presumably is going to be the fate of most of the world's great rivers, at
least temporarily. Many smaller water courses are being modified in a similar manner, and although
this is happening with less attention being drawn to individual cases, the effects of the accumulation
of environmental modifications brought about by many small dams may well be impressive. A typical
example of such an intervention is the Strydom dam on the Pongolo river, South Africa, which has
resulted in the disappearance of 100 km² floodplain and its associated lakes, although in this case
there has been an attempt to keep the wetlands intact through controlled discharges (Coke, 1970). In
the case of the Mogi Guassu-Rio Grande system in Brazil described by Godoy (1975) there has been a
progressive loss of feeding areas on the floodplains of the Rio Grande, by the construction of a series
of dams for industrial use.
Table 8.3
Summary of the effects of hydraulic works on river fish communities
Changes in flow
Temporal changes
Disruption of spawning patterns through
inappropriate stimuli or unnatural short-term
flows
Changes in community structure away from seasonal spawners to
species with more flexible spawning
Shift from pulse regulated to stable system
dynamics
Diminished productivity at community level
Changes in velocity
Increases in flow rate (usually due to
channelization)
Young fish in drift swept past appropriate sites for colonization
Local shifts in species composition in tail race with accumulation
of rheophilic predators
Decreased flow rate Shifts from rheophilic to lentic communities in reservoir
upstream and in controlled reaches downstream Changes in
flushing rate resulting in accumulation or low dilution of toxic
wastes or anoxic conditions leading to fish mortalities
Loss of habitat
Prevention of flooding by dams and levees Loss of floodplain area available for spawning growth; loss of
habitat diversity; change in species composition with loss of
obligate floodplain spawners
General diminution in productivity of whole system
Drowning of spawning substrates upstream of
dams or in channelized reaches
Variable effects usually involving decline of lithophils or
psammophils although new wave washed shore or rock rip-rap
may simulate rhithronic habitats

289
Blocking of channel
Interruption of migratory pathways by dam
walls or by the creation of unsuitable
conditions for passage
Elimination of diadromous or obligate migrants by preventing
movement to upstream breeding sites by adults and slowing
down- stream movements of juveniles
Changes in silt loading
Changes in channel form (due to
channelization or to changes in
deposition/erosion process)
Reduction of habitat and community diversity: loss of species
Increased rate of silt deposition (usually
upstream of dams but also in newly cut off
portions of channel or channelized reaches
downstream)
Choking of substrates for reproduction leading to failure to
reproduce in lithophils/ psammophils
Changes in density of vegetation usually in favour of phytophilis
Changes in quantity and type of food available and in the benthos
leading to restructuring of the fish community toward
iliophagous
Decrease in suspended silt load Changes in fish community reduction in number of non-visual
predators and omnivores
Lack of sediment (downstream of dams) Changes in nutrient cycle and in the nature of the benthos leading
to loss of iliophagous and increase in benthic limnivores
Changes in plankton abundance
Increases in phytoplankton in reservoir or
downstream due to slower flow and higher
water transparency
Increase in abundance of planktonivorous fish
Changes in temperature
Changes in mean temperature caused by low
flow regimes
Increasing temperature variation can cause shifts in success of
spawning due to adverse temperatures either for cold or warm
water spawners
Stratification in reservoirs Difficulties of passage for migrant species
Elimination of fish in deoxygenated hypolimnion
Mortalities downstream of dams due to emission of anoxic
waters and H2S
Uptake of water
Induction of water into power stations or
through pumps or irrigation canals
Entrainment of fish into currents diverting them; impingement of
fish on turbines and pumps resulting in loss of fish particularly
juveniles
Water transfers between river systems Transfer of species and disease organisms from one system to
another
The loss of floodplain area for feeding and breeding has serious effects on the fish populations, and
the reaction of the fish communities of the Chari, Niger and Senegal rivers to flood failures provoked
by natural climatic variations such as has occurred in the Sahel also confirm the highly detrimental
effects of suppressing the flood. The significance of floodable plains and backwaters as breeding and
nursery areas in North American rivers is documented by Guillory (1979) and was early appreciated
by Richardson (1921) who found a mean fish yield of 199 kg/ha where 90 percent of the water areas
of the Illinois river was in backwaters, 146 kg/ha where 83 percent was backwater and 78 kg/ha where
63 percent was backwater.
In the Missouri river, Whitley (1974) traced the steady decline of catch from 680 t in 1894 to 122 t in
1963 mainly to the loss of fish habitats following the construction of the reservoirs. On this river, too,
there have been modifications of the main channel by dykes on side channels and backwaters have

290
been blocked. The river has been trained by reveted banks into a series of bends. Prior to 1900
Ictalurus punctatus and Ictiobus spp. made up the major part of the catch, but since then the
introduced common carp Cyprinus carpio has become dominant.
Widespread changes occurred in the Colorado River following its conversion into a cascade system
(Holden and Stalnaker, 1975; Holden, 1979). A combination of hindrance to migration, reduction in
river flow and lowered mean water temperatures seriously reduced populations of endemic cyprinid
species such as the Colorado squawfish (Ptychocheilus lucius) hump back chub (Gila cypha), bonytail
chub (Gila elegans) and the razorback sucker (Xyrauchen tecarus) all of which are obligate riverine
species. These have to a large measure been replaced by species exotic to the Colorado basin such as
Micropterus salmoides, Lepomis cyanellus and Notropsis lutrensis which are more tolerant of lentic
conditions. A similar replacement of endemic fish species by introduced species which are better
adapted to the changed hydrological conditions has also been described from Soviet rivers. In the
Volga Chikova (1974) has noted changes in species composition below the V.I Lenin Volga
Hydroelectric station (Kuibyshev reservoir) where the phytophilous fishes, Abramis brama, A.
ballerus and Rutilus rutilus decreased in number, whereas Stizostedion (=Lucioperca) lucioperca and
“Sichel” (Pelecus cultratus ?) increased. Eliseev and Chikova (1968) concluded that the decline of the
phytophilous species is due to the failure and unpredictability of the flood, which when it does
inundate the plain, does so in a sporadic manner which arbitrarily strands young fish and spawn in
isolated pools. Main stream spawners such as the various sturgeons, Stizostedion and “Sichel” are not
so affected. Furthermore the flood peak of the new regime is now somewhat retarded favouring the
later spawning species which generally belong to the second group. Long distance migrant species
also declined in the Volga river following its conversion into a cascade reservoir system. Such species
as Huso huso, Caspialosa kessleri or Stenodus leucicithys disappeared from all but the lowest reaches
of the system whereas the regulated flows and reservoirs favoured such lacustrine species as
Clupeonella delicatula which populated large areas (Poddubnyi, 1979). In the Volga delta the
reduction of duration of flooding of the floodplain water bodies reduced the time available for their
occupation by young fish from 50–70 days before the closure of the dams (1950s) to only 10–15 days
afterwards (1960s–70s). This meant that the downstream migration of the young has been advanced
with less time for growth and survival on the plain. As a result there has been a great reduction in year
class strength (Koblitsbeya, 1985). At least 40 days of regular annual flood are judged necessary for
the maintenance of an adequate fish community in the lower reaches of this river. According to Lelek
and El Zarka (1973) and Adeniji (1975) the changes in the fish fauna of the Niger river below the
Kainji dam are also traceable to the unpredictable nature of the flood. Catch in the reach between
Jebba and Lokoja fell by about 50 percent in three years from about 4 400 t/yr (1967–69) (Otobo,
1978). This was accompanied by changes in species composition whereby the Characidae,
Mormyridae and Clariidae declined but predatory species such as Lates and Bagridae increased in
abundance (Sagua, 1978). Changes were also felt further downstream where the Lower Anambra
basin fisheries declined by about 60 percent (ie. 4 000 t) because of desiccation of much of the
floodplain following the closure of the dam (Awachie, 1978; 1979).
A localized increase in the abundance of fish, particularly of predatory species, immediately below
dams has been noted from many rivers. In the Nile below the Owen falls dam populations of Barbus
altianalis and the recently introduced Lates niloticus were particularly abundant. In the Niger, as we
have seen, Lates also appeared in quantity especially in the reach immediately below the Kainji dam
(Kainji to Jebba); although more recently it declined in abundance (Sagua, 1978) in favour of various
species of Mochokidae, Characidae and Cyprinidae. Chikova also commented on the maximum
accumulation of fish in the Volga as occurring in the same regions downstream of the dams. Whitley
(1974) explained this locally increased production in terms of the enriched water from the reservoir
which, in passing through the sluices, carries with it zooplankton, insects and fish. This enrichment
does not persist for any great distance, however, and in the Missouri was only detectable for about 2
km downstream. Such concentrations are probably associated primarily with feeding, but fish also
accumulate below dams where they are interrupted in their migrations and form the basis of rich
tailrace fisheries such as that described by Otobo (1978) for the Kainji dam.

291
Fish may be entrained by the flow out of dams either through the sluices or through the associated
power stations. This usually results in their death by gas disease caused by the explosive
decompression from high pressure upstream to low pressure downstream of the dam.
A history of diminishing catches in the Columbia river from a peak production of 22 440 t in 1911 to
only 6 800 t at present, is mainly due to the blocking of passage to migrating fish and changes in the
flow characteristics of the river, rather than to the loss of floodplain area (Trefethen, 1972). In the
Murray river, Australia, the 800 km migrations of the golden perch (Plectroplites ambiguus) have also
been stopped by a combination of water management practices, including flood control weirs, which
have left few of the original characteristics of the uncontrolled river system unchanged (Butcher,
1967). The Sakkur dam of the Indus did not affect populations of the migratory Hilsa ilisha, but the
construction of the Gulam Mahommed dam further downstream of it deprived the fish of 60 percent
of their previous spawning areas. Further upstream in the same river the Tarbela dam poses a threat to
Tor putitora, a large cyprinid which migrates from the floodplain to the foothills of the Himalayas to
breed. Hilsa stocks have become depleted in several other rivers of the Indian sub-continent. In the
Godavari river a combination of upstream dams and silting through erosion are restricting the species
increasingly to the estuarine delta (Nagaraja Rao and Rajalakshmi, 1976); in the Cauvery H. ilisha has
ceased to ascend the river following the regulation of flow by the Stanley reservoir (Sreenivasan,
1977). Following the construction of the reservoir, Puntius dubius has disappeared from the Cauvery
and Tor khudree is now much more restricted in its distribution. Species have also declined in
abundance in Brazilian rivers where dam construction has been intensive in recent years. Many of the
“Piracema” species have largely disappeared from the Rio Grande and its affluents in the Sao Paulo
state, and Pseudoplaytstoma coruscans was eliminated from the Tiete river soon after the closure of a
series of barrages there. In Venezuela the construction of the Guri dam on the Caroni River impeded
the upstream migration of many species of Pimelodid catfishes as well as the prochilodontid,
Prochilodus mariae and Semaprochilodus spp.
The importance of changes in silt and nutrient discharge from rivers on larger aquatic systems
following damming has been noted from several areas. Ryder (1978) traces the decline of the East
Mediterranean pelagic fishery to the withholding of nutrients in the Nile within Lake Nasser which in
turn has led to some increase in the yield of the lake (Fig. 8.4). Increases also occurred in Lake
Manyala in the Nile Delta where the reduction in flow caused by the Aswan High Dam led to the
silting of the channels connecting the lake to the sea. The freshwater environment then created with
the eutrophicating conditions originating from organic pollution has in recent years (1962–1973)
supported a greater fish population than previously (1908–1935) (Shaheen and Youssef, 1979). A
disastrous decline has been noted in fish production in the Black Sea and Sea of Azov since the main
inflowing rivers, the Danube, Dnieper and Dniester have been subject to increasing control in the
form of cascade systems (Tolmazin, 1979). This has led to a reduction in the inflow of nutrients which
used to support large stocks of fish and shellfish in the estuarine seas. It has also prevented the
discharge of freshwater which formed a well oxygenated layer over the deeper, saline and largely
anoxic waters. A similar decline in water quality and fisheries of the Caspian followed the
construction of the dams of the Volga River. Berdicevsky (1975) (in Carre, 1978) showed that while
the overall catch remained the same a change occurred in community structure away from higher
valued species to small pelagics of less commercial interest.

292
Figure 8.4 Regression of the annual fish catch from Lake Nasser on the annual yield of the Eastern
Mediterranean fishery four years earlier. (After Ryder and Henderson, 1975)
The importance of river inputs into larger aquatic systems is also demonstrated by the influence of
river flow on pelagic fish catches in Lake Kariba. Here highly significant relationships between the
wet season flow of the nutrient rich tributaries (Gwaai and Sanyati rivers) and the catch in the
subsequent year were established by Marshall (1982). Variations in the flow of the larger but nutrient
poor Zambezi did not affect catches to the same extent. Although these relationships originated in a
reservoir, the great flow to volume ratio makes nutrient conditions more closely resemble a river.
Thus by extension fluctuations in discharge of nutrient rich tributaries are also liable to influence the
productivity of larger but nutrient poor major rivers. This indicates the possible dangers of even
comparatively small tributary dams for lowering the overall productivity of river, lake or even marine
ecosystems.
Levees
The man-made levee is a type of linear dam which heightens the natural levee upwards to prevent
water spreading laterally on to the plain. The main impact of this is of course to deny fish access to
the feeding and breeding grounds that are necessary for their survival. Catch and specific diversity
drop in much the same way as they do for dams. As the water can no longer spread over such large
areas, sediment and pollution are concentrated into the river and such standing waters as remain,
much as described for the Illinois river by Starret (1972) (Fig. 8.5). Here the lack of spawning
grounds caused species such as pike (Esox lucius), large-mouth bass (Micropterus salmoides) and
yellow perch (Perca flavescens) to diminish in abundance compared to new species such as Cyprinus
carpio which were introduced into the system. The commercial catch declined from 2 613 t in 1950 to
182 in 1973, and angling success showed a similar reduction; effects which were attributed directly to
loss of habitat and siltation following the leveeing of the river and the draining of the bottom lands
(Sparks and Starrett, 1975).

293
Figure 8.5 Schematic drawing demonstrating the impact man has had during the past century on the
ecology of the Illinois river and two of its adjoining bottom-land lakes near Havana. (After
Starret, 1972)
The flow during the floods is also increased and as a result fish can be swept out of protected
positions into unfavourable environments. The construction of levees in low lying areas of the Danube
delta has increased the rate of descent of the larvae of many of the migratory fishes. Instead of
spreading out over the floodplain, to grow in the standing waters, these are ejected into the sea. As a
result catches of such species as Alosa pontica, Aspius aspius, Tinca tinca, Blicca bjoerkna and
Abramis brama, are much reduced relative to other arms of the river where no levees have yet been
constructed (Zambriborsch and Nguen Tan Chin, 1973). In the Middle Danube the Yugoslavian
fishery yields have declined from 20 000 t/yr to 1 200 t/yr following the enclosure of the flood-plain
zones by levees. A similar loss in fish production from Rumanian tributaries of the Danube has also
been noted by Bacalbasa following the reclamation of floodable bottom land for agriculture. Here not
only has the catch declined but the remaining population has come to be dominated by Carassius
auratus gibelio which is useless to the fishery (Bacalbasa and Popa, 1978). Similar losses are
described by Butcher (1967) from the Murray river, and pose a potential source of concern when
levees are to be constructed on floodplains supporting “Subienda” or “Piracema” types of migrants.
Effects comparable to those produced by levees can be anticipated from polders which essentially
enclose areas of floodplain within dykes to prevent them from uncontrolled flooding. In this way
many potential breeding areas may be removed from the ecosystem especially when the poldered area
is of any great extent.

294
Standing stocks of fish in the poldered Northern portion of the Atchafalaya basin, a distributory of the
Mississippi, has a lower standing stock at 555 kg/hr than the less controlled southern basin with 860
kg/hr. Land clearance and agricultural runoff leading to eutrophication are also implicated in the
lowered yield from the North whereas, in the South, the original allochthonous detrital based food
chains in the annually inundated habitats have been maintained (Bryan and Sabins, 1979).
Gaygalas and Blatneve (1971), traced the poor recruitment of bream, Abramis brama to the poor
spawning conditions in the 23 000 ha² of summer polders, which were constructed to reclaim the
floodplain depressions of the Nyamunas river delta. Summer polders are normally inundated during
the winter floods but are emptied during the spring. Overly rapid drainage does not allow the fish to
breed successfully, whereas with correct management, a slight prolongation of the flooded phase
improved recruitment in other years, and large fish stocks survived in the canals within the enclosed
area. Polders can also be flooded in a controlled manner for rice culture, forms of irrigated agriculture
or, as in Venezuela, for grazing reserves for cattle. Such practices can be combined with extensive
aquaculture. Where polders occupy a large proportion of the plain, the reduction in area available for
the expansion of the floodwaters can result in the acceleration of flow which can alter conditions on
the remaining flooded areas in a way unfavourable to fish.
Channelization
The channel is frequently used as a system for simplifying the natural complexity of a river. The
banks are smoothed and straightened in the interest of the more rapid and contracted evacuation of
water. Whilst channelized reaches of river may protect a particular area their net effect is to increase
the energy content of the water making it potentially more troublesome downstream. Flow patterns
are concentrated in time, producing shorter, higher spates. The rivers themselves may be shortened to
some degree by the evening out of meander bends; about 120 km have been subtracted from the
Missouri river between 1890 and 1947 in this way and the surface area of water has been reduced
from 49 250 ha in 1879 to 24 645 ha in 1972. Trees have largely been eliminated and the flood-plain
forest coverage has declined from 76% (1826) to 13% (1972). By contrast the percentage of land
under cultivation has increased from 18 to 83 over the same period (Burke and Robinson, 1979).
Channelization has much the same effects on species composition and abundance of fish and other
organisms as other flood control measures. In the United States workers such as Congdon (1973) and
Adkins and Bowman (1976) have found that the channelization of streams considerably reduces both
species diversity and biomass when compared with unchannelized stretches of the same river, and
Gorman and Karr (1978) also found that the general reduction in habitat diversity following
disturbances to the ecosystem such as channelization produced a corresponding reduction in both
diversity and stability of the fish community. As examples of this effect Groen and Schmulbach
(1978) reported that fish catch per mile was 2 to 2.5 times greater in the unchannelized reaches than in
channelized reaches of the Missouri. In channelized reaches many of the larger species such as lake
sturgeon (Acipenser fulvescens) paddlefish, (Polyodon spathula) and blue catfish (Ictalurus furcatus)
have almost been eliminated (Burke and Robinson, 1979) and catch has fallen by 80% between 1897
and 1963. Tarplee et al. (1971) found an average ichthyomass of 174.0 kg/ha in unchannelized and
55.3 kg/ha in channelized streams in N. Carolina. Jahn and Trefethen (1973) found 632.8 kg/ha of fish
in unchannelized, 502.9 kg/ha in lightly channelized and 146.7 kg/ha in heavily channelized reaches
of the Blackwater river, Missouri where the mean size of the fish was also much reduced in
channelized reaches. Many organisms other than fish are affected by channelization and Bulkley et al.
(1976) and Griswold et al. (1978) among others, have shown the drift organisms, which constitute the
bulk of fish food, are also much reduced by the siltation of the bottom in channelized reaches. Further
references describing the reduction of aquatic fauna in U.S. rivers due to this type of management
were listed by Schneberger and Funk (1971) and Schoof (1980). Schoof found that in addition to the
effects on the biotic elements of the system channelization can affect the physical environment by
draining wetlands, cutting off oxbow lakes and meanders, reducing ground water levels and the
recharge of ground waters from stream flow and by increasing sedimentation rates. Channels can also
contribute to the diminution of flooded area by deflecting the flow from one river to another, as in the

295
case of water transfers, or by bypassing major floodplain areas. The greatest projected work of this
nature is probably the Jonglei Canal, which if completed will lead much of the Nile water past the
Sudd to decant it back into the main river downstream of the swamp at Malakal. This will make much
more water available for irrigation in the arid Nile basin, but the estimated effects of the altered
regime within the Sudd will result in a reduction of maximum flooded area of between 26 and 36%
depending on the water regime adopted (Mefit-Babtie, 1984).
An early phase in the channelization process is the clearance of streams of snags, logs, boulders and
other large debris. This effectively reduces habitat diversity and has been responsible for a great loss
of fish. Even in torrential streams from the North-West United States floodplain habitats resulting
from such debris represented 6 – 29% of the summer rearing habitat and 55 – 70% of the standing
crop of coho salmon juveniles. As a result large log-jammed side channels had eight times the density
of coho than side channels lacking such features (Sedell, Yuska and Speaker, 1983).
Not all changes associated with channelization are detrimental to fish stocks. For example, Hesse et
al. (1982) noted that the rock rip-rap, wing dykes and stone revetments used to contain the flow
enormously increase the area available for periphyton. This offsets the lack of benthic food organisms
present in the accelerated flow of the river.
DEVELOPMENT AND MANAGEMENT OF RIVER SYSTEMS FOR FISHERIES
OBJECTIVES AND STRATEGY
Development and management are here considered as those intentional activities which are directed at
the manipulation of living aquatic resources, the environment in which they live and the human
communities associated with them. The nature of these activities depends on a variety of factors
among which are the purpose for which the resource is to be utilised, the objectives of management,
the type of fish community and the developmental context within which management is pursued.
Strategies for effective implementation of management plans also have to be conceived at different
levels. At the level of the basin, priorities must be set among a number of competing uses of which
fisheries is but one. At the level of the fishery the resource may have to be allocated among several
interested groups and direct management techniques applied to control the type and amount of fish
being caught. At the level of the fish community certain management measures can be employed to
improve the productivity of the fishery. Less directly fisheries may be managed by inducing social
changes or by the development of new types of fishery.
Objectives
Objectives, as opposed to uses, are here considered as those socio-economic criteria which underlie
the planning process. The first level of decision is the assignment of the resource among several
possible alternative uses. Such decisions usually arise from social or economic pressures. For
example, whereas production of fish for food may have high priority throughout the developing
world, the earning of foreign currency through export of luxury items may be judged more important.
Also in industrialised countries the commercial fisheries of the past have largely been supplanted by
sporting interests. Questions of access dominate recreational fisheries, and whether the resource is
open to all-comers or restricted to a certain group is a matter of deep concern to the recreational
fishery manager. Several main objectives consistently recur in development and management plans
for food fisheries; first the production of the maximum quantity of food at an appropriate price,
second the maintenance of the quality of the fish caught, third the provision of employment and fourth
the improvement of the standard of living of the fishing community. Such objectives are frequently in
conflict and the attainment of one in its entirety will impede the realization of the others. For example,
production of the maximum amount of cheap fish protein can only be at the cost of the quality of the
fish caught and usually of the well being of the fishing community. Alternatively, regulating access to
the fishery reduces employment opportunities but increases economic returns from the fishery.

296
Decision makers should, therefore, give same weighting to the various objectives to allow for a more
realistic orientation of management plans.
One of the commonest concepts for the regulation of fisheries is that of maximum sustainable yield
(MSY), which predicts that any fish stock has a constant surplus production which may be removed
by a fishery each year. The concept is applicable mainly to single stocks whose abundance is
relatively unaffected by changes in the environment. Few river fisheries are typical of this situation,
and because the concept introduces severe economic weaknesses in that to reach MSY costs incurred
exceed the revenues added by increasing effort. MSY has been also somewhat discredited in recent
years from the population dynamics standpoint. The hunt for a successor concept has proved
unrewarding and MSY is still cited in a loose way as representing the general productive capacity of a
fishery. The concept is particularly inappropriate to those river fisheries which are based on multi-
species communities and here the type of model described is more applicable. As an illustration of the
interaction between the objectives selected and planning, three points may be considered as critical on
the yield curve in this figure.
A. Represents a strategy based upon the management of the fishery for a few high valued species
only. It implies some loss in total potential catch and a great limitation in employment opportunities
but is often coincident with the maximum value to individual fishermen who maintain an elevated
catch per unit effort of a highly priced product. The fishery must be highly regulated to remain in this
state for an extended period.
B. Represents a compromise at which catch is maximal while conserving acceptable quality.
Individual welfare is high but access to the fishery must be restrained. In practise a fishery generally
fails to remain long at this point as social pressures tend to encourage drift towards the right hand end
of the curve.
C. Represents the other extreme. It provides maximum employment and a near maximum yield of
protein but a poor quality product and low level of individual welfare. Given the nature of the catch
per unit effort curve, however, where decreases in catch per unit effort decline and effort intensifies,
the welfare of the individual fisherman may not fall excessively when compared with point D for
instance. This strategy accepts the risks inherent in pushing the fish community near to its limits of
tolerance whereby any further pressure, environmental disturbance or climatic change can readily
destroy the fragile equilibrium and bring about the collapse of the community.
Because it is apparently more difficult for the fishery to move from the right-hand side of the model to
the left for socio-economic and biological reasons, the early setting of objectives is highly desirable to
avoid arriving inadvertently at a position where options are limited. Furthermore, because impacts of
human induced changes in the ecosystem tend to accelerate the fishing-up process, anticipated
changes in water quality or quantity should also be taken into account when setting objectives.
Uses of Fish Resources
The various species of fish that are found in rivers can be used for a variety of purposes.
(a) Food: This is by far the most widespread and important use of fish resources, especially in
tropical and subtropical areas far removed from the coast where freshwater fish often contribute a
significant proportion of the total animal protein in the diet. Fish for food may be either captured as
adults for direct consumption, or may be caught while still in the juvenile or fry stages for stocking
into other bodies of water or for rearing in ponds or cages through a variety of aquaculture techniques.
Most of the world's rivers contain stocks of fish that are used to a greater or lesser extent for food but
such fisheries are usually diffuse with relatively low yields at any one point. However, most of the
great lateral expansion areas, where extensive floodplains exist, support intensive fisheries which are
among the most productive in the world. The location of most of these are indicated in Figs. 1.13–

297
1.19. It is difficult to separate the production obtained from rivers from that of lakes and intensive
aquaculture in ponds in any given geographical region. It seems fair, however, to assume that about
half of the world's inland fisheries production of 8.6 million tons (1983 estimate) comes from running
waters, their floodplains and from aquaculture associated with lands inundated for rice culture or other
purposes. Certainly in Africa just under half of the 1.4 million tons is attributed to these sources
(Welcomme, 1979) and in S. America most of the 323 000 t comes from rivers as there are very few
lakes there. By far the greatest proportion of inland fish is derived from Asia, and in particular China,
where various types of extensive aquaculture are common, but river fisheries are also pursued. Fish
rearing in floodplain rice paddies is widespread throughout Southeast Asia and the production from
major rivers such as the Ganges, Brahmaputra, Mekong and Indus alone combine to give some 1.25
million tons.
(b) Sport: Recreational fishing is particularly common in the industrialised countries of the temperate
zones although this type of utilization is now spreading to other areas of the world where there is
heavy industrialization. It is often difficult to define the limits between recreational and food fisheries
as most fish caught for sport is also eaten, thus providing a valuable addition to diet. Furthermore
there is sometimes a recreational element in subsistence fisheries even in non-industrial economies.
Sport and commercial fisheries often compete for the same resource and, because the value of the
same fish is considerably greater to the sport fishery than to the commercial fishery, there is a trend to
favour the recreational interest with subsequent declines in the commercial catch.
(c) Ornament: The beauty and the interesting behaviour of many of the species of riverine fishes,
particularly those of the tropics, together with their small size have favoured their use for ornament or
hobby. Several hundred species are regularly captured or cultured for export providing a small but
significant income to some river communities. The magnitude of the world trade in aquarium fish has
been described by Conroy (1975) and I.T.C. (1979). From these it emerges that large numbers are
caught to compensate for wastage through disease and transport mortalities. Colombia, for instance,
exported some 10 million fish in 1974, Peru 15 million, Brazil 3.5 million, Venezuela 10 million - all
from the Amazon and Orinoco basins. As mortalities between capture and export range between 50
and 70 percent, this represents an offtake of at least 70 million fish, most of which are withdrawn
from such restricted environments as floodplain pools or small feeder streams. Local disappearances
of species due to overexploitation have already been reported from some countries. To compensate for
this, and to reduce the costs of capture and collection from the wild, many of these species are now
cultured in suitable warm waters. The transfer and spread of species by escape from such farms has
resulted in several of the commoner forms becoming pan-tropical in distribution.
(d) Other uses: There are a number of minor uses for riverine fish species including disease vector
control, where fish such as Gambusia or Lebistes have been introduced to control mosquito larvae.
Other species notably Ctenopharyngodon idella have been transferred widely for the control of
undesirable aquatic vegetation.
Developmental Stage
Four main stages can be identified in the modification of river systems and their fisheries which call
for different approaches to research and management (Table 8.4) and three cutting points seem
significant between one developmental stage and the next. The first of these occurs with the
introduction of systematic agriculture to the riparian area which marks the end of the unmodified
stage. Agriculture demands at least some attempt at drainage and regulation of floodplains and also
may involve some abstraction or diversion of water courses. As the support capacity of the basin rises
population increases and fishing pressure rises to the point where it produces a noticeable effect on
the fish stock. The second cutting point seems to arrive with the installation of flood control structures
which begin to modify the flood, impound water in lower order streams and restrict the flood area
available to the fishery. At this stage exploitation of the wild stock is very intensive and its effects,
added to the stresses induced by environmental modifications, can bring about rapid changes in

298
species composition and abundance. The last cutting point is reached with the installation of one or
more barrages along the course of the river. These change the nature of the downstream reaches from
flood rivers to reservoir rivers with the consequent loss of much of the previously flooded area. The
four stages listed do not necessarily represent an evolutionary sequence although most rivers in
Europe and North America have passed successively through these steps to reach their presently
highly controlled state. Similar fates are projected for many tropical rivers, but other rivers will
inevitably stay at a slightly or extensively modified stage without ever becoming completely
controlled. Nor does the developmental process necessarily proceed at an even pace along the whole
length of the river. Indeed in many large river systems, such as the Danube or the Mekong, different
reaches and tributaries may be at different developmental stages.
Table 8.4
Stages in the modification of rivers
Stage Basin use Fisheries Research and
management problems Unmodified
Channel and floodplain
show most
characteristic features,
flood regime
unhindered by direct
human interventions,
but indirect effects of
activities elsewhere in
the river basin may be
apparent, e.g., Sepik,
Niger, Sudd
In wild state often forested,
supports game and later used
for grazing cattle. Vegetation
modified by burning. Seasonal
occupation by nomadic
fishermen, hunters and
pastoralists. Slash and burn
agriculture practised in basin.
Fish stocks largely in
original condition of
diversity but size structure
may be modified by fishing
in both river channels and
standing waters. Whole
channel and plain available
for fisheries. Accidental
introductions could result
in the presence of several
exotic fish species.
Exploratory fishing for
description of composition
of fish stock. Identification
of major resources. Studies
of biology of individual
species and their
geographical and seasonal
distribution. Studies on local
fishing methods and
introduction of appropriate
additional techniques.
Establishment of simple
regulatory measures for
protection of major stocks.
Improve access and
marketing network.
Slightly modified
Some drainage
channels for more
rapid and efficient
removal of flood
waters from floodplain.
Smaller depressions
filled or regularized.
Flood still largely
unaltered in timing and
duration. Some small
dams on lower order
streams, e.g., Senegal,
Oueme.
Floodplain largely cleared of
forest, extensive drawdown
agriculture some floating rice in
suitable depressions. Some
areas reserved for grazing and
zonation of flood-plain for
different uses often highly
developed. Settlement on levées
and higher ground, or on
artificial islands and stilt
villages.
Fish stock largely unaltered
although larger species may
be becoming rarer and size
structure heavily biased
toward smaller individuals.
Some depressions may be
dammed as holding ponds,
or for extensive
aquaculture, or fish holes
may be excavated. Whole
floodplain available for
fisheries.
Population dynamics of
major elements of
community to give refined
estimates of potential yields.
Continue studies on biology
to identify possible sub-
populations and to describe
ecological interactions
between species.
Monitoring of fishery to
detect potential over-fishing
of major stocks coupled
with intensification of
regulatory measures to
protect fish stock.
Investigation of simple
forms of extensive
aquaculture. Improvement
and concentration of fish
landings and preservation
techniques.
Extensively modified
Lower order streams Flood agriculture(usually rice) Some modification to fish Examination of general

299
largely dammed for
flood control or
irrigation. Drainage
and irrigation common,
some flood control
through dams and
levées which contain
main channel.
Depressions usually
filled or regularized.
Flood often modified
in timing and duration,
e.g., Chao Phrya,
Mekong.
and intensive dry season
agriculture. Moderately
extensive occupation of the
dryer areas of the plain for
habitation - beginnings of
urbanization. Much of plain still
subject to flooding. Degradation
of lower order streams through
degradation of environment
produced by deforestation and
intensive agriculture, mining,
industrial pollutants and urban
sewage entering river
frequently without treatment.
Pesticides and herbicides from
large-scale monoculture
treatment also enter the river.
stock with disappearance of
larger species. Wild
fisheries often very intense
in main river channels, with
some new fisheries in
reservoirs. Disappearance
of most long-distance
migrant species. Rice fish
culture in suitable areas.
Drain-in ponds and some
intensive fish culture in
regularized depressions.
River area available for
fisheries restricted.
dynamics of fish community
to judge reaction to various
sources of loading.
Intensification of
monitoring of fisheries with
increased control of
catching methods by
licensing and legislation.
Examine impacts of other
activities in the river basin
on the fishery and
endeavour to ensure that
suitable conditions are
maintained. Investigation of
intensive aquaculture
methods. Consider
development of reservoir
fisheries and seek
alternative employment to
reduce fishing pressure on
main river.
Completely modified
Flood control by large
upstream dams and by
levées. Main channel
sometimes
channelized. Flood-
plain largely dry
although still subject to
occasional catastrophic
floods. River often
reduced to a chain of
reservoirs, e.g.,
Mississippi.
Urbanization, intensive use of
river basin for agriculture,
industry, habitation. Mining,
industrial and urban pollution to
some degree controlled,
eutrophication usual. Pesticides
and herbicides input.
Fish stock changed by loss
of some species through
pollution and
channelization and
sometimes by introduction
of exotic species. Some
sport fisheries in main
channels or in few lakes
that have been retained on
flood-plain. Some intensive
aquaculture in specially
constructed ponds. River
area available for fisheries
very small, but intensive
fisheries may be developed
in the reservoirs.
Investigation of
eutrophication and pollution
and other management
impacts to establish criteria
for maintenance of fish
stock. Regulation of
discharge and effluent
according to these criteria.
Contemplate introduction of
new elements to fish
community or stocking to
support threatened species.
Study access problems to
fishery to resolve
conflicting demands of sport
and commercial fishermen.
Intensify development of
aquaculture and reservoir
fisheries.
The four stages proposed correspond approximately to those presented by Yon and Tendron (1981)
(Figure 3.7) for European rivers. A similar series of categories has been proposed by Scudder and
Conelly (1984) on the basis of socio-economic criteria, which to a certain extent parallel the above
classification. Their four categories are:
(a) Primarily subsistence; in which fishing methods are limited to traditional, locally manufactured
gears and both men and women participate in the fishery. The catch is consumed entirely within the
boundaries of the fishing community.
(b) Incipient commercialisation; where fishing methods are still diversified but more modern twines
and gear are used. Division of labour is more apparent with the men fishing and the women carrying
out support activities. The fishery usually produces sufficient surplus for part of the catch to be sold
outside the fishing community.

300
(c) Primarily commercial; gill nets and seines are now the most common gear and the traditional
methods have largely disappeared. The fishery is pursued almost entirely for markets external to the
fishing region.
(d) Increasing marginalization of traditional communities; a period marked by a decrease in total
catch and decline in the fishery corresponding to a depletion in the resource.
MANAGEMENT OF THE RIVERINE ENVIRONMENT
There is an increasing trend at the present time to attempt to master rivers and to control their floods
so as to make their basins more productive for agriculture and safer for human occupation. Whatever
the long-term benefit and success of these attempts they do modify the aquatic environment in such a
manner that the fish stocks are adversely affected. If rivers are to continue to produce fish, it must be
within an entirely different framework from the capture fishery as practised in unmodified rivers.
PRESERVATION OF THE NATURAL SYSTEM
The biology and ecology of the majority of fishes inhabiting rivers show them to be extremely
sensitive to modifications of the flood cycle and to the environmental changes that occur in modified
rivers. In order to retain something of the natural diversity of the stocks of fish certain areas have to
be maintained in their wild state and it is hoped that the setting aside of land or some river systems for
suitable reserves be considered in the planning of land use within a river basin. As has been discussed
various types of agricultural engineering and construction works also interfere with the natural pattern
of flooding. Thus management policies involve planning of land use within the river basin as a whole.
Levees, polders and embankments for transport systems prevent the water reaching floodplain
depressions, and the remaining water is restricted within main channels in such a way as to modify its
flow. An essential part of planning for fisheries is, therefore, to make sure that any lateral expansion
zones which are designated as reserve areas should be kept free of such structures. A second
important feature is the provision of sufficient water to produce a flood which will have the
characteristics needed for the reproduction of fish species. This is both a question of quantity and
timing which have to be based on an adequate knowledge of the biology of the fish species concerned.
Controlled releases of water have been tried in some systems, and on the Pongola river, a series of
experimental floods have successfully filled the lagoons and induced breeding in the species
inhabiting them (Coke and Pott, 1970; Phelines, Coke and Nicol, 1973). Similarly, sufficient water
has been released from the Shire river dam to fill the lagoons of the Elephant and Ndinde marshes,
with satisfactory results for the fisheries. Kenmuir (1976) showed that the juveniles of several species
of fish, including Clarias gariepinus, Brycinus imberi, B. macrolepidotus, Labeo altivelis, L.
cylindricus, Eutropius depressirostris, Synodontis zambezensis, Oreochromis mortimeri and Tilapia
rendalli appeared in lagoons of the Mana floodplain of the Zambezi river following flood releases
from the Kariba reservoir. Furthermore sexually active adults of these species were observed moving
on to the plain during flood releases that corresponded to the time when the Zambezi would normally
flood. He therefore concluded that artificial releases of water (simulated floods) can stimulate
breeding in riverine fishes provided that they occur during the previously normal flood period.
The correlations between drawdown factors and catch detected in some floodplain rivers, as well as
the theoretical predictions of Welcomme and Hagborg's (1977) simulation indicate that standing stock
and yield do not depend solely on the flooded area, but also vary according to the amount of water
retained in the system during the dry season. Because of this, the shortfall in catch following a
reduction in flooded area can to a certain extent be compensated for by increasing the area of residual
water, provided the reduced floods still occur at the appropriate time. Where water management
actions are being planned which will result in the loss of areas of floodplain the provision of large
permanent bodies of water may be considered at an early stage. In all probability fish community of
systems managed in this way will undergo changes in species composition probably in favour of the
blackfishes much as in ordinary reservoirs.

301
The management of the natural system should not be restricted to the floodplain, but should be
extended upstream and downstream as part of the manipulation of the whole basin. As we have seen,
activities in the headwaters of the basin can alter the siltation, runoff and flow patterns of the potamon
reach, often with far reaching consequences to the fishery. Equally, pollution or damming
downstream can prevent species from moving up to the floodplains to feed or breed.
INSTREAM IMPROVEMENT STRUCTURES
A variety of structures can be used to restore more natural patterns where they have been destroyed or
to stabilize river channels whilst maintaining the diversity of habitats therein. Although such
structures tend to diminish the effectiveness of channels modified for drainage, irrigation, flood
control, etc. they can be used in a controlled fashion to improve the biological productivity of the
systems concerned.
According to Swales and O'Hara (1980) three main categories exist:
(a) structures which impound or modify flow;
(b) structures which provide direct cover;
(c) improvement of spawning areas.
Structures which impound or modify flow
Traditionally groynes or wing deflectors have been used for many years to improve salmonid streams
and have been proved effective in introducing artificial sinuosity into a straightened channel. They
also create slacks in the flow shadow immediately downstream and by scouring and deposition help
re-establish pool riffle sequences along the channel. As a pool riffle sequence characteristically recurs
at intervals of five to seven channel widths such devices should be installed with this spacing between
them. Low dams and nets are even more effective in the restructuring of the bed of the river to create
pool-riffle sequences. Both these types of device should be accompanied by bank stabilization to
prevent selective erosion by the deflected current. The use of structure of this type is not limited to the
rhithronic reaches of rivers. Burke and Robinson (1979) and Schick et al. (1982) describe a number of
structures which have been used to restore habitat diversity to channelized potamonic sections of the
Missouri and Mississippi rivers. These include (i) notched dykes, where portions of transverse barriers
are lower than the rest of the structure. These allow water to flow through the barrier so as to develop
or preserve side channels or to prevent further land accretion; (ii) Low elevation structures, which are
below the water surface for much of the time and which prevent island formation that can support
vegetation. They also lead to diversified depths particularly in that a deep pool develops downstream
of the structure; (iii) rootless structures are transverse dykes which do not abut on to the back, a
variant, vane dykes, are angled relative to the current. Such structures encourage multiple sub-channel
formation and create more bank area; (iv) chute closure structures are placed in such a way that side
arms, fringing lakes etc are maintained in contact with the main river channel. They also combat
permanent land accretion and the loss of spawning areas. On the whole such structures appear
successful in increasing habitat diversity and although there is no immediate evidence for increases in
biomass which would in any eventuality be difficult to detect in such a large river, various species do
appear to be showing distinct preferences within the new richer ecosystem.
Structures which provide direct cover
At the most primitive, such structures consist of fixed or floating objects which provide shade in
simulation of missing vegetation cover in larger river channels. Submerged objects may also serve a
similar purpose. Thus logs, boulders, branches etc, may be placed adjacent to the bank, a principle
elaborated by many tropical fisheries into brush parks or artificially created or anchored masses of
vegetation.

302
Because snags and other large organic debris are demonstrably important in maintaining habitat
diversity in small to moderate size streams (order 1 to 4) Sedell et al. (1982) proposed that, where
channels had been cleared, such structures be either artificially reintroduced or be allowed to
accumulate in order to increase the productivity of such waters.
Spawning area improvements
Several systems have been attempted for the improvement of bottom texture for fish which spawn
over gravel, particularly where siltation has resulted in the loss of the normal substrate. These
methods often consist in mechanical or hydraulic disturbance of the gravel causing deposited
sediments to be washed out, although the more costly and arduous physical replacement of the
substrate has also been tried. Letichevsky (1981) for instance, quotes the success of the recreation of
sandy spawning substrate of Stenodus leucichithys in the lower course of the Volga, which has now
supplanted a more expensive intensive fry rearing and stocking programme.
Fish passes
One of the remedies commonly proposed for blockages to migrations caused by dams is the
construction of fish ladders or passes similar to those which work so satisfactorily for the salmons in
North Temperate rivers. In some cases fish passes have been successfully installed in the tropics. One
such, at Cachoeira de Emas on the Mogi Guassu river, has been functioning since 1936 with little
apparent detrimental effect on the stock. Here many characin species leap up a series of shallow steps.
The siluroids, which do not jump, migrate past the barrage through special tunnels. The several
fishways at dams on the Tigris and Euphrates rivers have also been used successfully by many of the
migratory cyprinid species in these rivers (FAO/UN, 1956). Furthermore fish passes in dams blocking
the Himalaya tributaries of the Ganges also appear to function adequately for Tor spp. and Indian
major carps to the extent that the upper chambers of the ladders are frequently used as fish traps.
Most attempts at using fish ladders with tropical species have been less successful. Bonetto et al.
(1971) claimed that the efficiency of the fish ladders which bypass a series of dams on the Caracana
tributary of the Parana was very low and that Salminus maxillosus, one of the major migratory species
of the basin, was unable to negotiate them. Bonetto (1980) repeated the pessimistic view in describing
the anticipated performance of fish ladders to be installed in the Salto Grande and Yacyreta dams,
although subsequent experience with the Borland lifts installed in the Salto Grande dam showed that
certain species including: Salminus maxillosus, Pseudoplatystoma coruscans, Colossoma mitrei,
Doras spp., Oxydoras spp. and Prochilodus platensis did in fact use them to pass upstream. The fish
ladders installed at the Markala barrage on the Niger also did not fulfil their intended function
satisfactorily. Here enormous quantities of fish were blocked during their upstream migration at low
water although some fish did get through. The fisheries above the dam declined considerably after it
was closed as these reaches depended on a replenishment of their stocks from fish moving out of the
Central Delta. The failure of the ladder was attributed by Daget (1960) to insufficient capacity in view
of the very large numbers of fish wanting to pass over it. Furthermore, the migration of the species
concerned was a simple dispersal movement rather than a breeding migration and the stimulus to
surmount obstacles was possibly relatively low as a consequence. The failure of conventional
salmonid ladders in many tropical rivers is hardly surprising in view of the complex behavioural
factors which contribute to the functioning of such structures. For example, it has been shown that the
form of the standing wave downstream of weirs is essential for salmon to be able to generate enough
speed to clear the crest of the weir. Presumably other species have very different requirements which
need to be studied before the installation of effective fish passes can be contemplated. In any case the
provision of a fish pass or ladder for upstream movement of fish would only be justified where the
migration is absolutely essential for the maintenance of the fish stocks.

303
MANAGEMENT OF THE FISH STOCK
In principle a fishery exploits a community which in its unexploited state exists in some kind of
equilibrium with itself and with the environment. Under normal circumstances such a community may
be assumed to tend to maximize its biological productivity and to continue to do so when subjected to
a reasonable level of exploitation. In such a situation manipulation of the community in all probability
adds little or nothing to the yield of the fishery. There are two circumstances, however, under which
such interventions may be desirable. First, where the fish community is lacking some element to
exploit a food resource or habitat (i.e. a ‘vacant niche’) it may be considered advisable to introduce
one or more new species into the waterway. Second, where the fish community as a whole, or some
preferred element of it, is overfished a policy of stocking may be adopted.
INTRODUCTION OF NEW SPECIES
In their unmodified condition most rivers support fish communities that are sufficiently diverse to fill
most of the available trophic and spatial niches. Nevertheless some systems do exist which, for
zoogeographic reasons, have poor or incomplete faunas. Furthermore as systems are modified there is
a tendency for certain indigenous species to disappear and for others to take their place. This is most
noticeable in impoundments where many of the migratory riverine species are lost soon after the
stabilization of flow. In many rivers species which are adequately adapted to the new conditions are
absent from the original fauna and to overcome this additional species may have to be introduced
from other river basins. In some cases introductions have been performed uncritically, and such
species as the carp (Cyprinus carpio) or Oreochromis mossambicus have themselves been blamed for
degradation of the environment or fish community structure. These and others, for instance Salmo
gairdneri, have been implicated in the elimination of sometimes delicate native resources, such as the
unique faunal associations of Northeast California (Cooper, 1983) or the endemic galaxiids and
Protroctes oxyrhynchus from New Zealand. Such cases often arise when there has been a
contemporary disturbance of the environment which may have led to the elimination of the species in
any case. The presence of introduced species may thus be considered an advantage as they are able to
colonise the disturbed habitats and thereby maintain or increase species diversity (Leidy, 1983).
Fishes such as these, which are among the main elements for intensive aquaculture, find their way
into natural waters by accidental release from farm ponds. Any introduction of a new species into the
fish fauna of a river should, therefore, be preceded by a very careful analysis of its anticipated
impacts. The procedures for such an analysis have been defined by EIFAC/CECPI (1984).
While widespread or impetuous transfer of new species is to be decried, the great success of these
same species under other circumstances points to the role of properly considered introductions. Thus
most of modern aquaculture is founded on a comparatively small number of introduced species, the
tilapiine cichlids have been invaluable in increasing the yield from Asian reservoirs (Fernando, 1976)
and the common carp supports fisheries in many highly eutrophicated streams. Typical of such
successful transfers is that of the chinese carps into the Amu Darya river. A combination of the
macrophyte eating Ctenopharyngodon idella with the planktonophage Mylopharyngodon piceus,
Hypophthalmichthys molitrix, Aristichthys nobilis and Parabramis pekingensis colonised the system
including the adjoining Kara Kum canal thereby laying the basis for a rich fishery.
STOCKING
Three main motives for stocking exist:
(a) Stocking to maintain production in the face of intensive exploitation
(b) Stocking to mitigate or compensate for adverse effects of some activity within the river basin

304
(c) Stocking to increase production from individual components of the aquatic system, usually
through extensive aquaculture or ranching;
With the exception of trout streams and some sport fisheries in the temperate zones systematic
stocking of rivers to maintain or improve their fish stocks is not yet a widespread practice. In fact the
tendency has been more for rivers to act as a source of supply of juvenile fish seed for stocking into
reservoirs and fish culture installations. The requirements for successful stocking of salmonids have
been extensively studied and rules of thumb for numbers of fish and age of fish to be stocked have
been derived for various waters (EIFAC, 1982). The requirements and indeed the rationale for
stocking with other species is less well understood and in many cases current practice appears
arbitrary and counterproductive. There is considerable room for the expansion of stocking as a tool for
the management of river systems.
MANAGEMENT OF THE FISHERY
Traditionally a number of practices exist for the control of the fishery. These usually involve direct or
indirect control on the fishermen by limiting access to the fishery, or on the practise of fishing through
control of season, location or type of gear (Fig. 8.6). Usually attempts at direct control through
legislation result in costly and sometimes oppressive enforcement programmes, whereas less direct
manipulation of the economic forces regulating the fishery may be more effective. Whichever the
approach adopted, if controls do not correspond to biological realities regulating the fish community
attempts at regulating the fishery will be counterproductive.
REGULATION OF ACCESS
As shown in Fig. 7.13 the total catch is related to the number of fishermen operating on the river.
Furthermore the individual artisanal fisherman seems to have a limited fishing power in that he is
physically capable of removing only a certain quantity of fish from the system in any one year.
Because of this the solution that seems most appealing in this type of fishery is a simple restriction on
the number of fishermen operating in a certain region. Such control of access through licensing
remains one of the most important managerial tools, particularly in commercial or recreational
fisheries. An allied technique, the fixing of catch quotas for individual fishermen poses enforcement
problems in artisanal fisheries but may be practical where more intensive industrialized practices are
adopted. Many traditional systems also practice forms of access control by allowing only certain
groups within the community to fish the waters. In many rivers, especially in Asia, such control is
achieved by dividing the river and the plain into lots, for which individuals or groups of fishermen
compete at auction each year. The job of policing the lot rented then devolves upon the fishermen
groups. The renting of stretches of river to fishermen for restricted periods has been criticized on the
grounds that any group with only temporary tenure of a fishery will attempt to gain the maximum
profit within the time available to it. This leads to ruinous fishing practices where the stock is
exploited to the point of exhaustion. On the other hand, where areas are held by tradition or contract
for a number of years as in many parts of Africa, the fishermen are free to husband the resources and
develop the fishery. A final form of access control is usually less direct by manipulating the economic
conditions of the fishery so as to dissuade fishermen from entering the fishery. This may take the form
of taxation, price control on fish landed or regulating the supply of fishing gear all of which are aimed
at lowering net revenues to the fishermen.
Unfortunately the regulation of access to the fishery by traditional or by planned management tends to
break down under the pressure of unstable socio-economic conditions. This occurs particularly where
there is a high rate of rural unemployment and landlessness and fishing is viewed as a relief
occupation producing an influx of numerous new and uneducated persons into the fishery.

305
Figure 8.6 Diagram of the relationships between the main techniques for fishery management
INCREASING THE CATCH CAPACITY OF FISHERMEN
Improvements in the catch capacity of the individual fisherman can be brought about in several ways.
Introduction of new types of gear or the amelioration of existing ones enables the individual to
capture larger numbers of fish of a greater range of species. Better materials mean that the gear is less
liable to damage; consequently less time has to be spent in its maintenance and more on the fishing
grounds. Proper boat design and motorization of craft can get the fishermen to and from the fishing
grounds faster. Historically these processes have already occurred in many rivers, first with the
introduction of perfected gill nets, seines and cast nets, second by the introduction of nylon twine and
netting, and third by the adoption of the outboard motor. In some fisheries too, traditional methods,
especially those centred on cross river barrier traps which capture migrating fish, are as efficient as
the more modern techniques that seek to replace them. The effectiveness of the fishery may also be
conditioned by geographical accessibility or by the lack of adequate facilities for treatment which
limit the amount of fish that can be handled. Because of this some fisheries do operate below their full
capacity. However, the efficiency of much of the traditional and improved modern gear is such that
individual stocks have collapsed in some river systems and other whole communities are near
maximum exploitation, if not actually overfished. Improving the efficiency of the individual
fisherman does not, therefore, appear to be appropriate in many cases and should be regarded more as
a means of fine tuning the fishery as development of other sectors occurs. In other words, in areas
where there is a great demand for rural employment it is probably advisable to restrain the
introduction of more efficient methods and to concentrate on better marketing and other support
facilities. In such circumstances, where better gear is introduced, the improved performance is either
dissipated in a diminished catch per unit effort as the stock declines, or the less prosperous and
efficient fishermen, who cannot use or adapt to the new gear, are forced out of the fishery. As the
general economy of a country increases on the other hand, there is a tendency for labour to be
withdrawn from the rural sector including fishing. In such cases, fisheries have declined due to

306
fishermen leaving their trade, which in any case becomes less and less profitable. Here a rapid
increase in the individual efficiency of a few fishermen can to a certain extent halt the decline and
preserve a reasonable way of life for those who choose to remain in their previous profession.
Closed Seasons
By the nature of their biology, most river fisheries have built-in closed seasons. In the floodplain this
lasts from just after bankfull on the rising flood to peak floods when the fish population is too
dispersed and individual fish are too small in size for them to be readily available to the majority of
methods of capture. In the torrential upper reaches flows are frequently too high for effective fishing
during some of the year. This generally discourages fishing throughout the period as yields are low,
effort needed to capture fish high, and there are additional dangers of loss of gear in the currents and
floating vegetation masses. This effective closed season makes biological sense in that it allows the
fish to reproduce relatively undisturbed and for the young to grow to a reasonable size before they are
exposed to the fishery. Closed seasons outside this time are of limited value, although restrictions on
the overly heavy fishing of migrating adult fish to the spawning grounds may be necessary in some
places, as experience with Labeo in Africa have shown.
Reserved Waters
As fishing of the floodplain and river channel in the dry season becomes more intensive there is a risk
of local over-exploitation of the stock. For this reason, traditional fisheries have long been based on
the designation of certain floodplain depression lakes and reaches of the river as reserves which
remained unfished. In larger systems, there are usually inaccessible areas which form reserves as they
are infrequently exploited and it is probably from such areas that the Niger and Senegal rivers were
re-colonized during temporary reversals of the arid conditions of the Sahelian drought. As a
management measure the conservation of certain areas is probably a wise move and gains force when
other pressures are being applied to the system.
Mesh Regulations
The use of mesh selective gear almost always entails a consideration of the mesh sizes to be adopted
which can only be viewed relative to the characteristics of the stock to be exploited. As has been
described, the fishing-up process almost always involves a drift downwards in mesh size which needs
considerable enforcement of legislation to stop. Any lower limit on mesh size, therefore, has to be
imposed in the face of a natural trend to disregard it. It also poses the classic dilemma of how to
manage multi-species stocks consisting of species with a range of sizes. If the objective of the fishery
is to exploit only the large species of the community, the imposition of mesh limits which protect the
immature fish is probably the only way to do it effectively. However, one almost certainly neglects a
considerable proportion of the potential ichthyomass in this type of fishery as is shown by the number
of species caught by progressively lower mesh sizes (see Fig. 8.7). If the mesh size is lowered to take
advantage of the smaller species, then almost automatically the larger ones will disappear. One
possible solution to the dilemma is the limitation of mesh size in major gears such as seine nets, gill
nets, etc., coupled with a use of a variety of minor gears which are aimed at particular smaller
elements of the stock. In other words, in a fish community which is highly diverse it is as well to
maintain an equally diversified fishery.
Banning of Certain Gears
The restriction or complete outlawing of more destructive fishing practices is most important.
However, even such methods may be appropriate in some circumstances. Poisoning of water courses
is of course liable to damage the stocks of fish when carried out in the main channel of the river,
whereas its use for removing fish from temporary floodplain pools or for eradicating undesirable

307
species may be quite permissible. Unfortunately were the use of poisons allowed in one habitat it
would rapidly extend to others. Gear is often prohibited for reasons other than those bearing directly
on the fish stock. Long lines, for instance, are regarded with disfavour by users of cast nets which may
become entangled in the hooks. Barriers which completely block the river channel, thus stopping fish
migrations, are as likely to be removed for reasons of navigation as for fisheries.
Figure 8.7 The number of species captured by gill nets of different mesh sizes (after Reizer, 1974)
Accessibility
One of the main factors determining the degree to which the fisheries of the world's rivers are
exploited is their accessibility and proximity to major centres of population. Many of the tributaries of
major rivers such as the Ganges or the Amazon are still relatively unexploited due to the absence of
access roads, and some of the greatest floodplains are still relatively little utilized by reason of their
distance from suitable markets and lack of living space for the fishermen. The Okavango delta, the
Gran Pantanal of the Paraguay river or the Central flood regions of the Congo river are typical
examples of areas where development is conditional on the provision of suitable access routes and
centres for the marketing of catch, as well as artificial mounds or other appropriate locations for the
fishermen to settle near the fishing grounds during the flood.
DEVELOPMENT OF NEW OR ALTERNATIVE FISHERIES
Aquaculture

308
Natural production of rivers can be exceeded by various forms of husbandry, intensive or extensive
aquaculture. Such methods are widespread particularly on floodplains, and various techniques are
available for the different developmental stages of rivers.
Blocking of natural floodplain depressions
Complete blocking of small channels draining water from depressions is common in many systems.
Fish are conserved in the pools thus created and are fished later in the seasonal cycle at times when
fish from other sources are becoming scarce. This method is often the first systematic attempt at
husbandry or extensive aquaculture. Experiments with more permanent dams have been carried out by
Reed (FAO/UN, 1969a) in the Niger and by Reizer (1974) in the Senegal. Reed's work showed that
the area of standing water on the plain was increased by such installations (Fig. 8.8), and Reizer
showed how the level and area of such pools differ before and after damming (Fig. 8.9). Harvest from
such pools may be quite respectable - about 185 kg/ha/yr in the case of otherwise unmanaged pools in
the Niger and up to 500 kg/ha/yr in depressions which are stocked with fry and fed.
Figure 8.8 Tracing from aerial photograph of the Niger floodwater retention dams, showing the
original water area and the area flooded after the placing of the dams. (After FAO/UN, 1969a)
Drain-in ponds
In more advanced culture systems on floodplains depressions are regularized and depended and
eventually their number may be increased by digging new ponds into the surface of the plain. Three
river systems particularly have benefitted from this form of management. In the Oueme about 3
percent of the 1000 km2 surface area is occupied by drain-in ponds some of which are several
kilometres long (Fig. 8.10). Similar constructions on the Mekong floodplain ranged from 20 to 100 m
in length and 2–3 m in depth. In Bangladesh some 30 000 ponds have been formed from the borrow-
pits which were excavated during the construction of the artificial islands upon which much of the
rural population lives. Drain-in ponds are usually fished by blocking a portion with a bamboo barrier,

309
successively removing the accumulated vegetation and advancing the barrier until the fish are
confined in a small space from which they are easily removed. Drain-in ponds are also used in
conjunction with rice fields where, as the water is drained prior to harvesting, the fish may retreat into
ponds or ditches constructed for this purpose. Yields from drain-in ponds may be high. In 1955–58,
34 ponds from the Oueme produced a mean of 2.1 t/ha and even though the catch dropped the same
installations were still producing over 1.5 t/ha ten years later. In the Mekong this type of pond was
suppressed following the recommendations of Chevey and Le Poulain (1940) who felt that the
stagnant waters retained at the end of the dry season damaged the crops as they spread out over the
plain at the beginning of the next flood. A similar system is used in Equator where the “Chameras”
fisheries for Dormitator latifrons of the Chone river involve the construction of ponds on the
floodplain, either by dyking or by excavation. The young of D. latifrons stocked into these are grown-
on through the dry season.
Husbandry and fish culture in backwaters
Backwaters and other residual waterbodies separated from the main channel as a result of such river
control works as reverting and channelization may be used for rearing fish. In Hungary, for example,
such waterbodies are being developed for aquaculture (Pinter, 1983) and compare very favourably
with the more traditional rearing of fish in ponds. Yearling fish are stocked into such waterbodies
which are particularly suited to the raising of herbivorous species. Little additional feed is required
over and above the natural production, although this is encouraged with a limited application of
fertilizers. The success of this method is indicated by comparison with pond farming (Table 8.5).

310
Figure 8.9 Difference in changes in (A) depth and (B) area of a floodplain pool before and after
damming of the main access channel. Encircled letters refer to individual illustrations in Figure
1.8
Refuge traps
In the Oueme river vegetation masses may be planted deliberately at the end of the flood, either
attached to the bank (Fig. 8.10) or recessed into it at the mouth of the channels which drain the plain.
They are left to collect fish for about two months after which they are fished and replaced to be
emptied again towards the end of the dry season. Harvest from such “refuge traps” or “fish parks” can
be quite high and in the Oueme river 15 installations of this type of mean area 440 m² gave a mean
harvest equivalent to 1.88 t/ha (of park) per fishing or 3.88 t/ha/yr between 1958 and 1968.
The practice of deliberately planting vegetation or branches on the water to attract fish is in fact very
widespread in rivers as well as in coastal lagoons having been recorded by Chevey and Le Poulain
(1940) from the Mekong, Stauch (1966) and Reed et al. (1967) from the Niger and Benue systems and
Meschkat (1972) for Equador. Welcomme, (1972a) carried out an evaluation of this method of fishing
as it is practised in the lagoons associated with the delta of the Oueme river. The refuge traps in this
river and its associated lakes and lagoons are of two types in addition to the masses of floating

311
vegetation described above. There are small circular types about 22 m² in area which give up to 2.8
t/ha for each harvest. As they are harvested up to ten times during the 7 month dry season an annual
yield as high as 28 t/ha can be obtained without apparently affecting the catch by other fishing
methods in the area. Larger rectangular parks or “acadjas” are harvested less frequently, but also
achieve very high yields. The yield of brush parks is directly related to the frequency of fishing and to
the density of planting with greater yields of fish coming from parks with the greater density of
branches. In the freshwater zone, a variety of species are attracted to the Oueme fish parks from which
up to 32 species were recorded. By contrast, in the brackish water zone, only two species,
Sarotherodon melanotheron and Chrysichthys nigrodigitatus made up to 95 percent of the individuals
present. These breed actively in the park throughout the year and as a consequence the population
builds up rapidly (Fig. 8.11) thus accounting for the very high yields of up to 8 t/ha/yr in 1957–59.

312

313
Figure 8.10 Portions of the Oueme floodplain showing the distribution of different activities: (A)
river channel with brush parks and drain-in ponds; (B) a dense aggregation of drain-in ponds.
In both illustrations the plain is divided into fields. (By courtesy of IGN, Paris)
Table 8.5
Comparison between production parameters in Hungarian fish ponds
and in backwaters used for aquaculture (From Pinter, 1983)
Fish ponds Backwaters
Stocking rates Kg/ha 422 488
Cyprinus carpio 314 223
Herbivorous fish 98 257
Others 10 8
Yield Kg/ha 1305 1301
Cyprinus carpio 946 476
Herbivorous fish 334 749
Others 25 76
Total yield 883 813
Of which: fed
ponds 588 (66.6%) 316 (38.9%)
fertilised
ponds 295 (33.4%) 497 (61.1%)
Amount of feed in
starch value
(Kg)/1 kg
total yield) 2.3 1.3
More recent investigations have shown that the fish stock increased in a similar manner in 1969–70
although yields were somewhat lower as a result of changes in the lagoon environment brought about
by the construction of a port. The only other such installations that have been systematically
investigated are the ‘Samra’ parks of the Grand Lac which according to Chevey and Le Poulain
(1940) would appear to have given comparable yields to the Benin type of installation. Brush park
fisheries are regarded by many as a somewhat mixed blessing. There is little doubt that they put up the
overall productivity of the body of water in which they are used, but they also may shorten its life
through accelerated silting.
Rice-fish culture
The widespread practice of fish culture in rice fields may be considered an almost ideal method of
land use which produces both a carbohydrate and protein crops from the same piece of land. It has
reached very high standards in Asia and Madagascar but is still in the experimental stage in Africa
and Latin America. Coche (1967) has dealt comprehensively with the situation up to 15 years ago.
The combined culture of fish and rice is widespread throughout Southeast Asia and represents a
logical, integrated approach to the management of these types of wetlands. Ruddle (1980) classifies
the various ways in which rice and fish can be managed as crops from the same piece of land.

314
Figure 8.11 Relation between yield and length of time of installation of fish parks in the Oueme
Delta: (•) means for 1957–59; (x) means for 1968–70
Ricefield fisheries capture systems
culture systems concurrent cultivation
of fish and rice
rotational cultivation
of fish and rice
alternate intermediate
cropping cropping
He estimates that capture yields are about 135 kg/ha in the Malaysian Peninsula.
Two main types of exploitation can be distinguished:
(i) capture systems where the fish that gain access through natural channels may be trapped.
(ii) culture systems where the field is deliberately stocked with fry.
The fish may be taken as a single annual crop with a single rice crop, an intermediate crop between
one rice harvest and the next planting or during the growing season of the rice. Frequently fish are
trapped casually during the growing season, and are then drained into specially constructed sump
ponds or canals as the fields are dried (see Fig. 8.12). Yields from trapping during the growing period
have been measured at about 132 km/ha by Tang Cheng Eng et al. (1973). If the rice is cropped only
once per year, the sump ponds yield about 162.8 to 262.9 kg/ha (mean 200 kg/ha) but if the field is
double cropped fish yields drop to 68.2–143.0 (mean 95.7) kg/ha. At the same time between 2 277
and 2 975 kg/ha of rice were produced. Fish represents between 12–50 percent of the total income in
single cropping and 6–27 percent in double cropping systems. Similar yields have been recorded in

315
Thailand in the Mekong basin where 433 hm² of paddy fields were sampled giving fish harvests
ranging between 50 and 1 710 kg/ha (mean 433 kg/ha) (Thailand, Department of Fisheries, 1974). In
Sri Lanka rice-fish culture has not been practised because of the short (3–4 weeks) duration of the
flood phase.
There has been a long term decline in yield from ricefield fish production. The main reasons for this
are: First the increasing use of pesticides, weedicides and fungicides associated with the modern
advanced agrarian practices needed to grow the high yielding varieties of paddy. The effects of
pesticide treatment of rice for the control of stem borer insects, has caused concern and such workers
as Kok (1972) or Saanin (1969) have attempted to define the toxic and sub-lethal effects of the
common pesticides used (which include Endrin, Methyl Parathion, Dieldrin, Diazinon and -BHC).
While no definite conclusions can yet be drawn as to the precise effects of such environmental
contaminants, it is evident that the use of less toxic products can result in improved yields of rice
whilst safeguarding the supply of fish.
Figure 8.12 Adaptations of the paddy field for concurrent rice-fish cultivation (after Ruddle, 1980)
Second the concept of multiple cropping in rice cultivation leaves little scope for fish culture because
of the short duration of the flood over the rice field (Kassim, et al. 1979). The production of a plant
and a protein crop from the same area of floodable land is undoubtedly of great value in maintaining a
nutritionally balanced yield pattern, although one that is not necessarily maximal in terms of absolute
tonnage consequently projects have been suggested for the extension of this type of culture. For
instance, the management of the 32 000 ha Candaba swamp. This plain is flooded by the Pampanga
and Angat rivers and is one of a similar series of floodplains in the Philippines. The plain has been
enclosed in a series of dykes which include a gate for the harvesting of fish. The fields are at present
used for water melon or rice growing during the dry season when the fish remain in the canals. Fish

316
yields based solely on natural productivity range from 300–500 kg/ha in 7 months. The melon crop
produces 5 000 kg/hm. Delmendo suggested ways in which this area could be improved with the
inclusion of fish canals at the foot of the main dykes and an intensification of fish culture by stocking
and manuring the ponds.
Intensive aquaculture in ponds
The intensive culture of salmonids in ponds associated with the valleys of rivers is very common in
temperate countries, but the use of the potamon in the tropics for the purpose is more recent.
Once the flood is completely controlled the former floodplain is converted to irrigated agriculture.
Aquaculture farms, with ponds or pools specially adapted to the rearing of fish, can be associated with
the irrigation net-works. Such farms can either rear fish to a size where they can be sold directly for
consumption, or can raise fry for stocking into such permanent lakes as remain in the plain or into
fields where fish are reared as a secondary or alternative crop. Developments of this type are perhaps
best seen in Thailand, although they are also common in many other parts of Southeast Asia. In the
Chao Phrya river basin for instance, the control of the water regime over much of the floodplain has
decreased natural fish production. To compensate for this loss many state and private fish farmers rear
various species of cyprinid and siluroids as well as freshwater prawns (Macrobrachium rosenbergii).
The impervious nature of the alluvial soils, the flat terrain and the ready access to water supplies
through irrigation canals makes this type of modified floodplain particularly suitable for aquaculture
in ponds.
Cage culture in rivers
Once flow is stabilized within the main river channels, or even before such stabilization occurs, fish
may be cultured in floating cages. This type of culture usually applies to species of high commercial
value, such as Pangasius sutchii which is cultured in this manner in Thailand. The fish are usually
caught as fry and reared to marketable size in such structures.
Reservoir Fisheries
Reservoirs support fish populations and fisheries where yields are comparable to or higher than the
rivers they submerge. The net gain is usually far higher in rapids reaches where the relatively low
productivity of the main channel is easily exceeded by the new lake, but in any system where a
reservoir replaced a floodplain either by submerging it or by stopping its flooding the benefit or loss
to the fishery as a whole has to be carefully evaluated. In the case of the Upper Mekong, many of the
impounded tributary rivers have little floodplain either in the submerged reaches or downstream of the
dams (Sidthimunka, 1972). However, according to the Mekong studies the mainstream Pa Mong
reservoir will eliminate flooding for some 700 km downstream of the dam, resulting in a loss of catch
of about 2 150 t from the river, whereas the reservoir is expected to produce only about the same
amount of fish. Similarly, at least 6 000 tons of fish have been lost below the Kainji dam in Africa
which must be deducted from the 5 400 t produced by the reservoir behind the dam.
In many other reservoirs the amount of fish lost downstream of the dam surpasses the potential of the
dam itself, although such considerations are usually secondary where power generation or irrigated
agriculture are also taken into account.
The fish community that establishes itself in the reservoir is based entirely on that pre-existing in the
river basin, unless exotic species are introduced. There are usually considerable modifications in
community structure as many species are unable to adapt to the new environment. Earliest to
disappear from the body of the reservoir are the migratory whitefishes although they might survive in
the upper portion of the lake from which they can readily move upstream to breed. Blackfishes tend to
adapt better, and the main body of the lake is often colonized by species that were relatively

317
insignificant elements of the pre-impoundment fauna. In some rivers, for example, those of the Indian
subcontinent, Fernando (1976) suggested that few suitable blackfish occur. Consequently, lacustrine
fish communities are not readily established with the species native to the system, and, failing the
introduction of exotics, populations have to be maintained by continual stocking.
ACKNOWLEDGEMENTS
Although published under one name this book is the result of the work of many people. The
knowledge has been acquired not only by scientists working on the topic as an academic problem but
also by the fishermen who depend on their wisdom for the food they eat. During the course of travel
to and on many of the worlds rivers I have benefitted from the friendship and cooperation of many
who work directly with the fisheries described here and to them I owe an enormous debt. The
scientists and fishery administrators of many countries have contributed considerably to this work
through technical meetings of various types including the working parties of the FAO Regional
Fishery Bodies dealing with inland waters. Much of the stimulus to collect and interpret this
information comes from my colleagues in the FAO Department of Fisheries without whose help and
encouragement it would be difficult to proceed. In this respect the many hours spent in the physical
preparation of the paper by a number of secretaries should be particularly acknowledged.
REFERENCES
Academia Republicii Socialiste Romania, 1967. Limnologia sectorilui Romanesa al Dunarii.
Bucharest, Bucaresti Studiu Monografic Acadamiei Republicii Socialiste Romania, (Comisia de
Hydrobiolgie), 651 p.
Adams, J.C., 1964. Contribution a l'étude de la vegetation du lac de Guiers (Sénégal).
Bull.Inst.Fondam.Afr.Noire (A.Sci.Nat.), 26:1–72
Adebisi, A.A., 1981. The physico-chemical hydrology of a tropical seasonal river-upper Ogun River.
Hydrobiologia, 79:157–65
Adeniji, E.O., 1975. Observations on agriculture and fisheries downstream of the Kainji dam. In The
ecology of Lake Kainji, the transition from river to lake, edited by A.M.A. Imevbore and O.S.
Adegoke. Ile-Ife, Nigeria, University of Ife Press, pp. 181–6
Adis, J., K. Furch and U. Urmler, 1979. Litter production of a Central amazonian black water
inundation forest. Trop.Ecol., 20(1):236–45
Adkins, G. and P. Bowman, 1976. A study of fauna in dredged canals of coastal Louisiana.
Tech.Bull.La.Wildl.Fish.Comm., (18):72 p.

318
Ahmad, N., 1956. Fishing gear of East Pakistan. Dacca, Directorate of Fisheries, 33 p.
Al-Hamed, M.I., 1966. Limnological studies on the inland waters of Iraq. Bull.Iraq Nat.Hist.Mus.,
3(5):1–22
Al-brecht, M.L., 1953. Ergebmissa quantitativer untersuchungen an flissender. Gewass.
Ber.Limnol.Flusstn.Freudental, 4:10–1
Al-brecht, M.L., 1959. Die quantative untersuchung der Bodenfauna fliessender. Gewass.Z. Fisch.,
8:481–550
Alexander, G.R., 1979. Predators of fish in coldwater streams. In Predator prey systems in fisheries
management. Washington, D.C., Sport Fishing Institute, pp. 153–70
Allen, K.R., 1951. The Horokiwi stream: A study of a trout population, Fish.Bull., N.Z., (10):231 p.
Andreev, N.M., 1966. Handbook of fishing gear and its rigging. Jerusalem, Israel Program for
Scientific Translation, IPST Cat. No. (1654):454 p.
Angermeier, P.L., 1982. Resource seasonality and fish diets in an Illinois stream. Environ.Biol.Fish.,
7(3):251–64
Angermeier, P.L. and J.R. Karr, 1983. Fish communities along environmental gradients in a system
on tropical streams. Environ.Biol.Fish., 9(2):117–35
Annibal, S.R. 1983. Avaliacao bio-ecologico e pesqueira das “pescadas” (Plagioscion
squamosissimus Heckel, 1840 e Plagioscion montei Soares, 1978) no “sistema Lago do Rei” - Il ha
do Careiro - AM - Brasil. Manaus, INPA (Instituto Nacional de Pesquisas da Amazonia) MSc
Dissertation., pag. var.
Antipa, G., 1910. Regiunea inundabila a Dunarii. Starea ei actuala si mijloacele de a o pue in valoare.
Bucharest, 318 p.
Ardizzone, G.D. and R. Ziesler, 1977. Lista prelimanar de las aguas continentales de America Latina.
Rome, FAO/COPESCAL, 144 p.
Arias, P.A., 1975. Contribucion al conocimiento limnologico de la cienaga de Guarinocito y su
relacion con el rio Magdalena. Tesis, Fundacion Universidad de Bogota, Colombia, pag. var.
Arias, P.A., 1977. Evaluacion limnologica de las planicies inundables de la cuenca norte del Rio
Magdalena. Proyecto para el desarollo de la pesca continental en Colombia. Cartagena,
INDERENA-FAO, pag. var.
Arifin, O. and Z. Arifin, 1976. Fisheries on the floodplain area of south Sumatra, a case study of the
Lubuk Lampam in 1973. Paper presented at IPFC Symposium on the development and utilisation
of inland fishery resources, Colombo, 27 October-5 November 1976. IPFC/76/SYM/10 (mimeo)
Attwell, R.I., 1970. Some effects of Lake Kariba on the ecology of a floodplain of the Mid-Zambezi
Valley of Rhodesia. Biol.Conserv., 2(3):189–96
Awachie, J.B.E. and E.C. Walson, 1978. Atalla fishery of the lower Niger, Nigeria. CIFA
Tech.Pap./Doc.Tech.CPCA, (5):296–311

319
Awachie, J.B.E., P.C.O. Ilozumba and W.I. Azugo, 1978. Fish parasites in the ecology, management
and productivity of river and floodplain fisheries in Africa. CIFA Tech.Pap./Doc.Tech.CPCA,
(5):253–65
Bacal basa, N. and N. Popta, 1978. Consideratii ecologice si teknologice asupra limnologiei aplicate
la conditule jedetului Galati. Ocrot.Nat.Med.Inconj., 22(2):119–26
Backiel, T., 1971. Production and food consumption of predatory fish in the Vistula River. J.Fish
Biol., 3:369–405
Backiel, T., 1983. Rybactwo i ryby w Wisle. In Ekologicne podstawy zagospodarowania i jej
dorzecza, edited by Z. Kajak. Warsawa, PWN, pp. 511–42
Backiel, T. and E.D. LeCren, 1978. Some density relationships for fish population parameters. In
Ecology of freshwater fish, edited by S. Gerking. Oxford, Blackwell Scientific Publications, pp.
279–302
Bain, M.B., 1984. Fish community structure in rivers with natural and modified daily flow regimes.
Paper presented at the 114th Annual General Meeting of the American Fisheries Society
Bakare, O., 1970. Bottom deposits as food for inland freshwater fish. In Kainji Lake studies. Vol. 1.
Ecology, edited by S.A. Visser. Ibadan, Ibadan University Press, pp. 65–85
Baker, A.L. and K. Kromerbaker, 1979. Effects of temperature and current discharge on the
concentrations and photosynthetic activity of the phytoplankton of the upper Mississippi River.
Freshwat.Biol., 9:191–8
Balogun, R.A., 1970. Occurence of free amino acids and sugars in Lake Kainji. In Kainji Lake
studies, Vol. 1, Ecology. Ibadan, Ibadan University Press, pp. 103–6
Balon, E.K., 1967. Vyvos ichtyofauny Duneja jej sucasny stav a pokus o prosnozu dalksich zmien po
vystavbe vodnych diel. Biol.Pr., 8(1):5–121
Balon, E.K., 1975. Reproductive guilds of fishes: a proposal and a definition. J.Fish. Res.Board Can.,
32(6):821–64
Balon, E.K., 1981. Additions and ammendments to the classification of reproductive styles in fishes.
Environ.Biol.Fish., 6(3–4):377–89
Balon, E.K. and A.G. Coche, 1974. Lake Kariba: A man-made tropical ecosystem in Central Africa.
The Hague, Junk, Monogr.Biol., 24:767 p.
Balon, E.K. and D.J. Stewart, 1983. Fish assemblages in a river with unusual gradient (Luongo,
Africa, Congo system), reflections on river zonation, and description of another species.
Environ.Biol.Fish., 9(3/4):225–52
Bardach, J., 1959. Report on fisheries in Cambodia. Phnom Penh, Cambodia, USOM, 80 p. Issued
also in French
Basile-Martins, M.A., et al., 1975. Influencia de factores abioticos sobre a maturacao dos ovarios de
Pimelodus maculatus Lac. 1803 (Pisces: Siluroidea), Bol.Inst. Pesca Santos, 4(1):1–28

320
Bastl, I., J. Holcik and I. Krupka, 1969. Abundance and ichthyomass of fish populations in the
Biskupicke branch of the Danube River. Pr.Lab.Rybar., 1969(2):253–68
Bayley, P.B., 1973. Studies on the migratory characin, Prochilodus platensis Holmberg, 1889,
(Pisces: Characoidea) in the R. Pilcomayo, South America. J.Fish Biol., 5:25–40
Bayley, P.B, 1981. Fish yield from the Amazon in Brazil: comparison with African river yields and
management possibilities. Trans.Am.Fish.Soc., 110:351–9
Bayley, P.B, 1983. Central Amazon Fish populations. Biomass, Production and some dynamic
characteristics. Dabhousie University, Dissertation. 330 p.
Bazigos, G.P., et al., 1977. The present state of the fishery of the Magdalena River Basin, Colombia.
Rome, FAO, FI:DP/COL/72/552:30 p.
Beadle, L.C., 1974. The inland waters of tropical Africa: An introduction to tropical limnology.
London, Longman. 365 p.
Beauchamp, R.S.A., 1956. The electrical conductivity of the head-waters of the White Nile. Nature,
Lond., 198(4534):616–9
Bell-Cross, G., 1971. Weir fishing on the central Barotse flood plain in Zambia. Fish.Res.Bull.,
Zambia, 5:331–40
Belyy, N.D., 1972. Downstream migration of the pike perch Lucioperca lucioperca (L.) and its food
in the early development stages in the lower reaches of the Dneiper. J.Ichthyol., 12:465–72
Benech, V. and J. Quensiere, 1982. Migrations de poissons vers le Lac Tchad a la decrue de la plaine
inondée du Nord-Cameroun. 1 Note - Methodologie d'echantillonage et resultats generaux.
Rev.Hydrobiol.Trop., 15(3):191–282
Benech, V., 1983. Migrations de poissons vers le Lac Tchad a la decrue de la plaine inondée du Nord-
Cameroun. 2 Note - Comportement et rythmes d' activité des principales éspèces.
Rev.Hydrobiol.Trop., 16(1):79–101
Benech, V., 1984. Migrations de poissons vers le Lac Tchad a la decrue de la plaine inondée du Nord-
Cameroun. 3 Note - Influence de l'hydrologie sur les variations interannuelles.
Rev.Hydrobiol.Trop., 16(3):287–316
Bernacsek, G.M. and S. Lopes, 1984. Investigations into the fisheries and limnology of Cahora Bassa
reservoir seven years after dam closure. Rome, FAO, FAO/GCP/MOZ/006/SWE., Field doc. 9,
pag. var.
Berner, L.M., 1951. Limnology of the Lower Missouri River. Ecology, 32(1):1–12
Berns, S. and H.M. Peters, 1969. On the reproductive behaviour of Ctenopoma muriei and Ctenopoma
damasi. Ann.Rep.EAFFRO, (1968):44–9
Best, R., 1981. Foods and feeding habits of wild and captive Sirenia. Mammal.Res., 11(1):3–29
Beverton, R.J.H., 1959. Report on the state of the Lake Victoria fisheries. Lowestoft, Fisheries
Laboratory, (mimeo)

321
Bhatnagar, G.K. and B.J. Karamchandani, 1970. Food and feeding habits of Labeo fimbriatus (Bloch)
in River Narbada near Hoshangabad (MP), J.Inland Fish.Soc. India, 2:30–40
Bhuiyan, N.I., 1959. Hilsa fishing in River Indus. Agric.Pak., 11(4):511–9
Binns, N.A. and F.M. Eiserman, 1979. Quantification of fluvial trout habitat in Wyoming.
Trans.Am.Fish.Soc., 108(3):215–28
Bishop, J.E., 1973. Limnology of a small Malayan river, Sungai Gombak. The Hague, W. Junk, 485
p.
Bishop, K.A., 1983. Fish movement as a possible regional dispersal mechanism. In Environmental
protection in the Alligator river region. Jabiru, 17–20 May 1983. Jabiru, Office of Supervising
Scientist, vol. 2, Pap.72:18 p.
Bishop, K.A. and M. Forbes, The freshwater fishes of Northern Australia. In Monsoonal Australia
edited by C. Haynes, M. Ridpath and M. Williams. Rotterdam, A.A. Balkema (in press)
Bisson, P.A. and J.R. Sedell, Salmonid populations in logged and unlogged stream sections of western
Washington. In Proceedings of a Symposium on fish and wildlife relationships in old-growth
forests, April 12–13 1982, Juneau, Alaska, edited by W.R. Mechan, T. Hanley and T.R. Merrill.
American Institute of Fishery Research Biologists (in press)
Bisson, R.A., et al., 1982. A system of naming habitat types in small streams, with examples of
habitat utilisation by salmonids during low streamflow. In Aquisition and utilisation of aquatic
habitat inventory information, edited by N.B. Armantrout. Proceedings of a Symposium held 28–
30 October 1981, Portland, Oregon. Western Division, American Fisheries Society, pp. 62–73
Blache, J. and J. Goosens, 1954. Monographie piscicole d'une zone de peche au Cambodge. Cybium,
8:1–49
Blache, J., F. Miton and A. Stauch, 1962. Premiére contribution à la connaissance de la pêche dans le
bassin hydrographique Logone-Chari Lac Tchad. Mem.ORSTOM, 4(1):143 p.
Blache, J., et al., 1964. Les poissons du bassin du Tchad et du bassin adjacent du Mayo Kebbi.
Mem.ORSTOM, 4(2):483 p.
Blackburn, W.M. and T. Petr, 1979. Forest litter decomposition and benthos in a mountain stream in
Victoria, Australia. Arch.Hydrobiol., 86(4):453–98
Blanc, M., J. Daget and F. D'Aubenton, 1955. Recherches hydrobiologiques dans le bassin du Moyen-
Niger. Bull.Inst.Fr.Afr.Noire (A.Sci.Nat.), 17:619–746
Blazka, P., 1958. The anaerobic metabolism of fish. Physiol.Zool., 31:117–28
Blench, T., 1972. Morphometric changes. In River ecology and man, edited by R.T. Oglesby, C.A.
Carlson and J.A. McCann. New York, Academic Press, pp. 287–308
Bonetto, A.A., 1975. Hydraulic regime of the Parana River and its influence on ecosystems.
Ecol.Stud., 10:175–97

322
Bonetto, A.A., 1976. Calidad de las aguas del Rio Parana - introduccion a su estudio ecologico.
Corrientes, Argentina, Instituto Nacional de Construcciones Portuarias y Vias Navigables -
Instituto Nacional de Ciencias y Tecnica Hidrica (INCYTH), 202 p.
Bonetto, A.A. and A.M. de Ferrato, 1966. Introduccion al estudio del zooplankton en las cuencas
islenas del Parana Medio. Physis, B.Aires, 26(72):385–96
Bonetto, A.A. and C. Pignalberi, 1964. Nuevos aportes al conocimiento de la migración de los peces
en los rios mesopotamicos de la Republica Argentina. Commun.Inst. Nac.Limnol., Argent., 1:1–14
Bonetto, A.A. and I.D. Ezcurra, 1964. La fauna bentonica de algunas aguas rapidas del Parana Medio.
Physis, B.Aires, 24:311–6
Bonetto, A.A., W. Dioni and C. Pignalberi, 1969. Limnological investigations on biotic communities
in the middle Parana river valley. Verh.Int.Ver.Theor.Angew. Limnol., 17:1035–50
Bonetto, A.A., et al., 1971. Informaciones complementarias sobre migraciones de peces en la cuenca
de la Plata. Physis, B.Aires, 30:305–20
Bonetto, A.A., 1978. Estudios limnologicos en la cuenca del Riachuelo (Corrientes, Argentina).
Ecosur, 5(9):57–84
Bonetto, A.A., 1978a. Estudios limnologicos en la cuenca del Riachuelo (Corrientes, Argentina), 2.
Lagunas Totoras y Gonzales. Ecosur, 5(9):17–55
Bonetto, A.A., 1978b. Estudios limnologicos en la cuenca del Riachuelo (Corrientes, Argentina), 3.
Laguna La Brava. Ecosur, 5(3):57–84
Bonetto, C.A., 1980. Densidad de poblacion, crecimiento y production del sabalo (Prochilodus
platensis) en la Laguna Gonzales (Corrientes, Republica Argentina). Hist.Nat., 1(18):121–36
Bonetto, C.A., 1980a. Estimación de la transperancia del agua en fonción de las variaciones de altura
hidrometrica en la alta Parana. Hist.Nat., 1(14):93–100
Bonetto, C.A., 1982. Estimación de la producción primaria de fitoplankton en fonción de algunas
variables limnologicas del Rio Parana (Corrientes, Argentina). Hist.Nat., 2(15):125–36
Bonetto, C.A., et al., 1979. Producción primaria del fitoplankton del Rio Parana en el area de su
confluencia con el Rio Paraguay. Ecosur, 6(12):207–27
Bonetto, C.A., 1981. Contribución al conocimiento limnologico del Rio Paraguay en su tramo
inferior. Ecosur, 8(16):55–88
Borhan, M.A., 1981. River Nile fish and fisheries 1980. Progr.Rep.Inst.Oceanogr.Fish. Inland Waters
Fish Cult.Branch, Alexandria, (5):pag. var.
Botnariuc, N., 1967. Some characteristic features of the floodplain ecosystems of the Danube.
Hydrobiologia, 8:39–49
Botnariuc, N., 1968. La conception de Gr. Antipa concernant la production biologique de la zone
inondable du Danube, du point de vue des recherches actuelles. Trav.Hist.Nat.Grigoire Antipa,
8:60–8

323
Bowen, S.H., 1979. A nutritional constraint in detrivory by fishes: the stunted population of
Sarotherodon mossambicus in Lake Sibaya, South Africa. Ecol.Monogr., 49(1):17–31
Bowen, S.H., 1979a. Determinants of the chemical composition of periphytic detrital aggregate in a
tropical lake (Lake Valencia), Venezuela. Arch.Hydrobiol., 87:166–77
Bowen, S.H., 1980. Detrital non-protein amino acids are the key to rapid growth of Tilapia in Lake
Valencia, Venezuela. Science Wash., 204:1216–8
Bowen, S.H., 1981. Digestional assimilation of periphytic detrital aggregate by Oreochromis
mossambicus. Trans.Am.Fish.Soc., 110:239–45
Bowmaker, A.P., 1963. Cormorant predation on two central African lakes. Ostrich, 34(1):2–26
Braga Adhemar, R., 1976. Ecologia e etologia de piranhas no nordeste do Brasil (Pisces-Sarrasalmus
Lacepede 1803). Tese, Instituto de Biociencias de Universidade de Sao Paulo. Fortaleza, Ceara,
DNOCS, 268 p.
Braker, W.P., 1963. Black piranhas spawned in the Shedd aquarium. Aquarium, Philad., 32(10):12–4
Bramer, H., 1980. Some innovations don't wait for experts: a report on applied research by
Bangladeshi peasants. Ceres, 13(2):24–8
Brandt, A. von, 1984. Fish catching methods of the world. 3rd ed. Farnham, Surrey, Fishing News
Books Ltd., 418 p.
Braum, E. and W.F. Junk, 1982. Morphological adaptations of two Amazonian characoids (Pisces) for
surviving in oxygen deficient waters. Int.Rev.Ges.Hydrobiol., 67(6):869–86
Braun, R., 1952. Limnologische untersuchungen as einegen seen in Amazonsgebeit.
Schweitz.Z.Hydrol., 14(1):128 p.
Brinkman, W.L. and U de M. Santos, 1973. Heavy fish-kill in unpolluted floodplain lake of central
Amazonia, Brazil. Biol.Conserv., 5:147–9
Brook, A.J. and J. Rzoska, 1954. The influence of the Gebel Aulia Dam on the development of the
Nile plankton. J.Anim.Ecol., 23:101–15
Brown, M.E., 1957. The physiology of fishes. New York, Academic Press, 2 vols., 447 p.
Bruns, D.A., et al., 1984. Tributaries as modifiers of the river continuum concept: analysis by polar
ordination and regression models. Arch.Hydrobiol., 99(2):208–20
Bruton, M.N., 1979. The survival of habitat desiccation by air breathing clariid catfishes.
Environ.Biol.Fish., 4(3):273–80
Bryan, C.F. and D.S. Sabins, 1979. Management implications in water quality and fish standing stock
information in the Atchafalaya river basin, Louisiana. In Proceedings of the Third coastal marsh
and estuary management symposium, edited by J.W. Day, et al., Baton Rouge, Louisiana,
Louisiana State University, pp. 293–316
Bryan, C.F., et al., 1975. Annual report - A limnological survey of the Atchafalaya basin. Baton
Rouge, Louisiana State University, Louisiana Cooperative Fishery Research Unit, 203 p.

324
Bryan, C.F., 1976. Annual Report - A limnological survey of the Atchafalaya basin. Loiusiana
Cooperative Fishery Research Unit. 285 p.
Bryson, R.A., 1974. A prospective on climatic change. Science, Wash., 184:753–60
Bulkley, R.V., et al., 1976. Warmwater stream alteration in Iowa: extent, effects on habitat, fish, and
fish food, and evaluation of stream improvement structures (Summary report). Iowa Cooperative
Fishery Research Unit, 39 p.
Burke, T.D. and J.W. Robinson, 1979. River structure modifications to provide habitat diversity. The
mitigation symposium. Gen.Tech.Rep.U.S.Dep.Agric., Fort Collins, (RM-65):556–61
Butcher, A.D., 1967. A changing aquatic fauna in a changing environment. IUCN Publ. (New Ser.),
9:197–219
Cabrera, S.E., 1962. Crecimiento de pejerrey del Rio de la Plata y algunos datos ecologicos sobre la
especie Atherinidae: Basilichthys bonariensis (Cuv. et Val.). Buenos Aires, Secretaria de Estado de
Agricultura y Ganaderia de la Nacion, Direccion General de Pesca y Consevacion de la Fauna,
Departamento de Investgaciones Pesqueras, 53 p.
Cabrera, S.E. and C. Candia, 1964. Contribución al conocimiento de la biologia de sabalo
(Prochilodus platensis Holmberg) del Rio de la Plata. Rev.Invest.Agropec. B.Aires, 1 (4):57–83
Cabrera, S.E., et al., 1973. Algunos aspectos biologicos de las especies di ictiofauna de la zona de
Punto Lara (Rio de la Plata) la parte, Alimentación natural del pejerrey (Basilichthys bonariensis).
Publ.Serv.Hidrogr.Nav., Argent., (H1028):29 p.
Cabrera, S.E., 1973a. Algunos aspectos biologicos de las especies di ictiofauna de la zona de Punto
Lara (Rio de la Plata) 2a parte, Alimentacion natural de bagre porteno (Parapimelodus
valenciennesi), Publ.Serv.Hidrogr.Nav., Argent., (H1029):47 p.
Cadwalladr, D.A., 1965. Notes on the breeding biology of Labeo victorianus Boulenger
(Pisces:Cyprinidae) of Lake Victoria. Rev.Zool.Bot.Afr., 72(1–2):109–34
Candia, C.R., M. de Lourdes Baiz and S.E. Cabrera, 1973. Algunos apectos biologicos de las especies
de ictiofauna de la zona de Punta Lara (rio de la Plata). 3a parte. Estudio de la edad y crecimiento
del bagre porteno (Parapimelodus valenciennesi) con algunos datos sobre su reproduccion.
Publ.Serv.Hidrogr. Nav., Argent., (H1030):33 p.
Canestri, V., 1972. El recurso icticola continental, Regionalisacion ictiofaunistica. Caracas, Ministerio
de Agricultura y Cria, Oficina Nacional de Pesca, 72 p.
Carey, T.B., 1971. Hydrological survey of the Kafue floodplain. Fish.Res.Bull., Zambia, (5):245–95
Carey, T.G., 1967. Some observations on distribution and abundance of the invertebrate fauna.
Fish.Res.Bull., Zambia, (3):22–4
Carey, T.G., 1967a. Kafue river and floodplain research. Fish populations in lagoon and riverine
environments. Fish.Res.Bull., Zambia, (3):9–12
Carey, T.G., 1971. Hydrological survey of the Kafue floodplain. Fish.Res.Bull., Zambia, (3):9–12

325
Carmouze, J.P., J-R. Durand and C. Leveque, 1983. Lake Chad, ecology and productivity of a shallow
tropical ecosystem. The Hague, Junk, 575 p.
Caro, P.M., C.C. Bonetto and Y. Zalocar, 1979. Produccion primaria de fitoplankton de lagunas del
noroeste de la Provincia de Corrientes. Ecosur, 6(11):83–100
Carre, F., 1978. La peche en Mer Caspienne. Ann.Geogr., (479):1–39
Carter, G.S., 1953. The papyrus swamps of Uganda. Cambridge, Heffer, 25 p.
Carter, G.S. and L.C. Beadle, 1931. The fauna of the Paraguayan Chaco in relation to its environment.
1. Physico-chemical nature of the environment. J.Linn.Soc., Lond.(Zool.), 37:205–58
Castella, E., et al., 1984. Macroinvertebrates as ‘describers’ of morphological and Hydrological types
of aquatic ecosystems abandoned by the Rhone River. Hydrobiologia, 119:219–25
CECOAL, 1977. Estudios ecologicos en el area de Yacyreta: informe de avance, 2. Argentina,
Corrientes, Centro de Ecologia Aplicada del Litoral, 204 p.
CECOAL, 1979. Estudios ecologicos en el area de Yacyreta: informe de avance, 3. Argentina,
Corrientes, Centro de Ecologia Aplicada del Litoral, 153 p.
Chang, B.D. and W. Navas, 1984. Seasonal variations in growth, codition and gonads of Dormitator
latifrons in the Chone river basin, Equador. J.Fish Biol., 24(6):607–48
Chanthepa, S., 1972. Nam Ngum fishery programme and fishery in Khone Falls (Mekong River).
Paper presented at SEADAG Seminar, Santa Barbara, 23 p. (mimeo)
Chapman, D.W., 1967. Production in freshwaters. In The biological basis of freshwater fish
production, edited by S.D. Gerking. Oxford, Blackwell Scientific Publications, pp. 3–29
Chevey, P. and F. Le Poulain, 1940. La peche dans les eaux douces du Cambodge. Mem.
Inst.Oceanogr.Indochine, (5):193 p.
Chikova, V.M., 1974. Species and age composition of fishes in the lower reach (downstream) of the
V.I. Lenin Volga Hydroelectric power station. In Biological and hydrological factors of local
movements of fish in reservoirs, edited by B.S. Kuzin. New Delhi, Amerind Publishing Company
Co., pp. 185–92
Chitravadivelu, K., 1974. Growth, age composition, population density, mortality, production and
yield of Alburnus alburnus (Linnaeus, 1758) and Rutilus rutilus (Linnaeus, 1758) in the inundation
region of the Danube-Zofin. Acta Univ. Carol.(Biol.), 1972:1–76
Clifford, H.F., 1972. Downstream movements of white sucker Catastomus commersoni fry in a brown
water stream of Alberta. J.Fish.Res.Board Canada, 29(7):1091–3
Cloern, J.E. and F.H. Nichols, 1978. A von Bertalanffy growth model with a seasonally varying
coefficient. J.Fish.Res.Board Can., 35(11):1479–82
Coates, D., 1984. An ulcer disease outbreak amongest the freshwater fish populations of the Sepik
River system, with notes on some freshwater parasites. Rep.Dep. Primary
Ind.Fish.Res.Surv.Branch, Port Moresby, (84–02)

326
Coche, A.G., 1967. Fish culture in rice fields, a world-wide synthesis. Hydrobiologia, 30(1):1–44
Coke, M., 1970. The water requirements of Pongolo floodplain pans. Paper presented to the Republic
of South Africa Water Year 1970 Convention: Water for the future, 6 p. (mimeo)
Coke, M. and R. Pott, 1970. The Pongolo floodplain pans: a plan for conservation. Pietermaritzberg,
Natal Parks Board, 34 p.
Congdon, J.C., 1973. Fish populations of channelized and unchannelized sections of the Chariton
River, Missouri. Spec.Publ.N.C.Div.Amer.Fish.Soc., (2):52–62
Conroy, D.A., 1975. An evaluation of the present state of the world trade in aquarium fish. FAO
Fish.Tech.Pap., (146):128 p.
Cooper, J.S., 1983. Distributional ecology of native and introduced fishes of the Pit River system,
northeast California, with notes on the Modoc sucker. Calif. Fish Game, 69(1):39–53
Cordiviola de Yuan, E., 1971. Crecimiento de peces del Parana medio. 1. ‘Sabalo’ (Prochilodus
platensis Holmberg). Physis, B.Aires, 30(81):483–504
Cordiviola de Yuan, E. and C. Pignalberi, 1981. Fish poulations in the Parana River, 2. Santa Fe and
Corrientes area. Hydrobiologia, 77:261–72
Cott, H.B., 1961. Scientific results of an enquiry into the ecology and economic status of the Nile
crocodile (Crocodilus niloticus) in Uganda and Northern Rhodesia. Trans.Zool.Soc.Lond.,
29(4):211–356
Coulter, G.W., 1970. Population changes within a group of fish species in L. Tanganyika following
their exploitation. J.Fish Biol., 2:329–53
CTFT, 1957. Notes et documents sur la pêche et pisciculture (Dahomey, Vallée inférieure de
l'Oueme). Nogent-sur-Marne, Centre Technique Forestier Tropical, Ser.D.G.2
Cuinat, R., 1971. Pricipaux caracteres demographique observés sur 50 rivieres à truite francaises.
Influence de la pente et du calcium. Ann.Hydrobiol., 2(2):187–207
Culp, J.M. and R.W. Davies, 1982. Analysis of longitudinal zonation and the river continuum concept
in the Oldman-South Saskatchewan river system. Can.J.Fish. Aquat.Sci., 39(9):1258–66
Daget, J., 1952. Mémoires sur la biologiedes poissons du Niger. 1. Biologie et croissance des éspèces
du genre Alestes. Bull.Inst.Fr.Afr.Noire, 14(1):191–225
Daget, J., 1954. Les poissons du Niger Superieur. Mem.Inst.Fr.Afr.Noire, (36):391 p.
Daget, J., 1956. Mémoires sur la biologie des poissons du Niger moyen. 2. Recherches sur Tilapia
zillii (Gerv.). Bull.Inst.Fr.Afr.Noire, 36:391 p.
Daget, J., 1957. Mémoires sur la biologie des poissons du Niger moyen. 3. Reproduction et croissance
d'Heterotis niloticus Ehrenberg. Bull.Inst.Fr.Afr.Noire, 19:295–323
Daget, J., 1957a. Donées recentes sur la biologie de poissons dans le Delta Central du Niger.
Hydrobiologia, 9:321–47

327
Daget, J., 1960. Les migrations des poissons dans les eaux douces tropicales africaines. Proc.IPFC,
8(3):79–82
Daget, J., 1966. Abondance relative des poissons dans les plaines inondées par la Bénoué à hauteur de
Garoua (Cameroun). Bull.Inst.Fondam.Afr.Noire(A.Sci.Nat.), 28:247–58
Daget, J. and P.S. Economidis, 1975. Richesse spécifique de l'ichtiofaune de Macedoine orientale et
de Thrace occidentale (Grèce). Bull.Mus.Nat.Hist.,Paris (3° ser.) (Ecol.Gen.7)), 346:81–4
Daget, J. and J.-M. Ecoutin, 1976. Modèles mathematiques de production applicables aux poissons
subssant un arret annuel prolongé de croissance. Cah.ORSTOM (Hydrobiol.), 10(2): 59–70
Daget, J., N. Planquette and P. Planquette, 1973. Premieres données sur la dynamique des
peuplements de poissons du Bandama (Cote d'Ivoire). Bull.Mus.Natl.Hist.Nat., Paris, (3°
Ser.)(Ecol.Gen.7), 151:129–43
Dahl, G., 1971. Los peces del Norte de Colombia. Bogota, D.E., Ministerio de Agricultura, Instituto
de Desarollo de los Recursos Naturales Renovables, INDERENA, 319
Dansoko, F.D., 1975. Contribution a l'etude de la biologie des Hydrocyon dans le delta central du
Niger. Thesis, Mali, Bamako, Ministere de l'Education Nationale, 105 p.
Dansoko, F.D., H. Breman and J. Daget, 1976. Influence de la secheresse sur les populations
d'Hydrocynus dans le delta central du Niger. Cah.ORSTOM (Hydrobiol.), 10(2):71–6
D'Aubenton, F., 1963. Rapport sur le foncionnement d'un barrage mobile sur le Tonle-Sap. Paris,
Museum d'Histoire Naturelle. 38 p. (mimeo)
Day, F., 1958. The fishes of India; being a natural history of the fishes known to inhabit the seas and
freshwaters of India, Burma and Ceylon. London, W. Dawson, 2 vols., vol. 1:778 p., vol. 2:pag.
var.
Delmendo, L.M., 1969. Lowland fishponds in the Philippines. Curr.Aff.Bull.IPFC, (54/55):1–11
Desai, V.R., 1970. Studies on the fishery and biology of Tor tor (Hamilton) from River Narbada. 1.
Food and feeding habits. J.Inland Fish.Soc. India, 2:101–12
Dickie, L.M., 1975. Predation, yield and ecological efficiency in aquatic food chains.
J.Fish.Res.Board Can., 33(2):313–6
Donnelly, B.G., 1978. Evidence of fish survival during habitat desiccation in Rhodesia.
J.Limnol.Soc.South.Afr., 4(1):75–6
Drury, D.H., 1969. Hydraulic geometry. In Introduction to fluvial processes, edited by R.J. Chorley.
New York, Barnes and Noble, pp. 146–56
Ducharme, A., 1975. Informe tecnico de biologia pesquera (Limnologia). Publ.Proy. Desarr.Pesca
Cont.INDERENA/FAO Colombia, (4):42 p.
Dudgeon, D., 1982. Spatial and seasonal variations in the standing crop of periphyton and
allochthonous detritus in a forest stream in Hong Kong, with notes on the magnitude and fate of
riparian leaf fall. Arch.Hydrobiol., Suppl.64(2):189–220

328
Dudley, R.G., 1972. Biology of Tilapia on the Kafue floodplain, Zambia: predicted effects of the
Kafue Gorge Dam. PhD. Dissertation, University of Idaho, Moscow, U.S.A., 50 p.
Dudley, R.G., 1974. Growth of Tilapia of the Kafue floodplain, Zambia: predicted effects of the
Kafue Gorge Dam. Trans.Am.Fish.Soc., 103:281–91
Dudley, R.G., 1976. Status of major fishes of the Kafue floodplain, Zambia, five years after the
completion of the Kafue Gorge Dam. Final report submitted to the National Science Foundation
Scientists and Engineers in Development. Athens, Georgia, Georgia University Grant no. OIP75–
09239: 71 p.
Dunn, I.G., 1982. The Hilsa fishery of Bangladesh, 1982: an investigation of its present status with an
evaluation of current data. FAO, Rome, FI:DP/BGD/81/034, 71 p.
Dunn, I.G. and G. Othe, 1982. Fisheries development in the Middle Mahakam Area. East Kalemiarta
Tranomigration Area Development Project. TAD Materialian 13. Eschborn, GTZ, pag.var.
Durand, J.R., 1970. Les peuplements ichtyologiques de l'El Beid. Prémiere note. Presentation du
milieu et resultats généraux. Cah.ORSTOM (Hydrobiol.), 4(1):3–36
Durand, J.R., 1971. Les peuplements ichtyologiques de l'El Beid. 2e. Note. Variations inter et
intraspécifiques. Cah.ORSTOM (Hydrobiol.), 5(2):147–59
Durand, J.R. and G. Loubens, 1969. Croissance en longueur d'Alestes baremoze (Joannis, 1835)
(Poissons; Characidae) dans le bas Chari et la Lac Tchad. Cah.ORSTOM (Hydrobiol.), 3(1):59–105
Durand, J.R., 1970. Variations du coefficient de condition chez les Alestes baremoze (Pisc. Charac.)
du bas Chari et du lac Tchad. Cah.ORSTOM (Hydrobiol.), 4(1):27–44
Durand, J.R., 1970a. Observation sur la sexualite et la reproduction des Alestes baremoze du bas
Chari et du lac Tchad. Cah.ORSTOM (Hydrobiol.), 4(2):61–81
Eckholm, E.P., 1976. Losing ground: environmental stress and world food prospects. New York,
W.W. Norton, 223 p.
Edwards A.M. and J.B. Thorne, 1970. Observations on the dissolved solids of the Casiquiare and
Upper Orinoco, April-June 1968. Amazoniana, 2(3):245–56
Edwards, R.W. and M.P. Brooker, 1982. The ecology of the Wye. The Hague, W. Junk
Egborge, A.B.M., 1971. The chemical hydrology of the River Oshun, Western State, Nigeria.
Freshwat.Biol., 1(3):257–72
Egborge, A.B.M., 1974. The seasonal variation and distribution of phytoplankton in the River Oshun,
Western State, Nigeria. Freshwat.Biol., 4(2):177–91
EIFAC, 1982. Report of the Symposium on stock enhancement in the management of freshwater
fishes, Held in Budapest, Hungary, 31 May - 2 June, 1982 in conjunction with the Twelfth session
of EIFAC. EIFAC Tech.Pap., 42:43 p. Suppl.Vol.1:281 p., Vol.2:283–554. Issued also in French
EIFAC/CECPI, 1984. Report of the EIFAC Working Party on stock enhancement. Hamburg, Federal
Republic of Germany, 16–19 May 1983. Rapport du Groupe de travail de la CECPI sur

329
l'amelioration des stocks. Hambourg, Republique fédérale d'Allemagne, 16–19 mai 1983. EIFAC
Tech.Pap./Doc.Tech.CECPI. (44):22 p.
Eliseev, A.I. and V.M. Chikova, 1974. Conditions of fish reproduction in the lower reach
(downstream) of the V.I. Lenin Volga Hydroelectric Station. In Biological and hydrological factors
of local movements of fish in reservoirs, edited by B.S. Kuzin. New Delhi, Amerind Publishing
Corporation, pp. 193–200
Elouard, J.M. and C. Leveque, 1977. Rythme nycthemeral de derive des insectes et des poissons dans
les rivieres de Cote d'Ivoire. Cah.ORSTOM (Hydrobiol.), 11(2):179–83
Enaceanu, V., 1957. Contributiuni la determinarea productivitatii apelor piscicole.
Bul.Inst.Cercet.Piscic., 16:83–5
Enaceanu, V., 1964. Das Donauplankton auf rumanischen Gebeit (Km 488 bis km 345).
Arch.Hydrobiol., (Suppl.27):442–56
Ertl, M., 1966. Zooplankton and chemistry of two backwaters of the Danube River. Hydrobiol.Stud.,
1:267–95
Ertl, M., S. Juris and J. Tomajka, 1972. Vorlaufige angaben iche jahrezeitliche Veranderungen und
die vertikale verteilung des periphytons in mitteleren abshnitt der Donau. Arch.Hydrobiol., Suppl.
44 (1):34–48
Espinosa, V. de and C.B. Gimenez, 1974. Estudio sobre la biologia y pesca del bocachico Prochilodus
reticulatus (Valenciennes) en el Lago de Maracaibo. Inf.Tec.Of. Nac.Pesca, Caracas, (63):32 p.
Everett, G.V., 1974. An analysis of the 1970 commercial fish catch in three areas of the Kafue
floodplain. Afr.J.Trop.Hydrobiol.Fish., 3(2):148–59
FAO/UN, 1954. Report to the Government of Iraq on the Development of inland fisheries, based on
the work of A. van den Eilaart. Rep.FAO/UNDP(TA), (270):38 p.
FAO/UN, 1956. Report to the Government of Iraq on fishways at dams on the Tigris and Euphrates
Rivers. Rep.FAO/UNDP(TA), (526):20 p.
FAO/UN, 1962. Rapport au Gouvernement de la Republique du Niger sur la situation et evolution de
la peche au Niger. Base sur le travail de Mr. J. Daget. Rapp.FAO/PEAT, (1525):27 p.
FAO/UN, 1968. Report to the Government of Zambia on fishery development in the Central Barotse
plain. Based on the work of D.W. Kelley, Inland Fishery Biologist. Rep.FAO/UNDP(TA),
(2554):83 p.
FAO/UN, 1968a. Multipurpose survey of the Kafue River Basin, Zambia. Vol. 4. The ecology of the
Kafue flats. Part 1. ‘Ecology and Development’, prepared by H.J. van Rensburg. Rome,
FAO/SF:35/ZAM, 138 p.
FAO/UN, 1968b. Report to the Government of Zambia on fishery development in the Central Barotse
Plain. Based on the work of D.W. Kelley, Inland Fishery Biologist. Rep.FAO/UNDP(TA),
(2554):83 p.
FAO/UN, 1969. Report to the Government of Zambia on fishery development in the Central Barotse
floodplain. Second phase. Based on the work of D. Duerre. Rep.FAO/UNDP(TA), (2638):80 p.

330
FAO/UN, 1969a. Report to the Government of Nigeria on fishing technology relating to river and
swamp fisheries of Northern Nigeria. Based on the work of W. Reed, FAO/TA Fishery
Technologist. Rep.FAO/UNDP(TA), (2711):90 p.
FAO/UN, 1970. Report to the Government of Nigeria on fishery investigations on the Niger and
Benue rivers in the Northern Region and development of a programme of riverine fishery
management and training. Based on the work of M.P. Motwani. Rep.FAO/UNDP(TA), (2771):196
p.
FAO/UN, 1970a. Report to the Government of Zambia on the fishery development of the Central
Barotse floodplain. Based on the work of G.F. Weiss. Rep.FAO/UNDP (TA), (2816):19 p.
FAO/UN, 1971. Rapport au Gouvernment du Dahomey sur l'evolution de la pêche interieur, son eetat
actuel et ses possibilites, etabli sur la base des travaux de R.L. Welcomme, Specialiste de la peche.
Rapp.FAO/PNUD(AT), (2938):97 p.
FAO/UN, 1973. Kainji Lake Research Project, Nigeria. Pelagic primary production in Kainji Lake.
Report prepared for the Government of Nigeria by the FAO of the UN acting as executing agency
for the UNDP. Based on the work of S.G. Karlman, Rome, FAO/UNDP FI:SF/NIR 24. Tech.Rep.,
(3):59 p.
Fenerich, N.A., M.Y. Narahara and H.M. Godinho, 1975. Curva de crecimiento e primeira maturacao
sexual do mandi, Pimelodus maculatus Lac. 1803 (Pisces: Siluroidei). Bol.Inst.Pesca Santos,
4(1):15–28
Fernando, C.H., 1977. Reservoir fisheries in South East Asia; past, present and future. Proc.IPFC,
17(3):475–89
Ferreira da Silva, V.M., 1983. Ecologia alimentar dos golfinhos da Amazonia. Dissertation, Manaus,
Instituto Nacional de Pesquisas da Amazonia, pag. var.
Fily, M. and F. d'Aubenton, 1966. Cambodge: Grand Lac-Tonle Sap. Technologie des peches 1962–
63, Paris, Ministere des Affaires Etrangeres, Service de Cooperation Technique, 373 p.
Fittkau, E.J., 1967. On the ecology of Amazonian rain-forest streams. In Atas de Simposio sobre a
Biotica Amazonica. Rio de Janiero, Conselho Nacional Pesquisas Vol. 3, Limnologia, pp. 97–108
Fittkau, E.J., 1970. Role of caimans in the nutrient regime of mouth-lakea of Amazon affluents (a
hypothesis). Biotropica, 2(2):138–42
Fittkau, E.J., 1973. Crocodiles and the nutrient metabolism of Amazonian waters. Amazoniana,
4(1):103–33
Fittkau, E.J. and H. Klinge, 1973. On biomass and trophic structures of the Central Amazonian rain
forest ecosystem. Biotropica, 5(1):2–14
Fittkau, E.J., et al., 1975. Productivity, biomass and population dynamics in Amazonian water bodies.
Ecol.Stud., 11:289–311
Floyd, K.B., R.D.Hoyt and S. Timbrook, 1984. Chronology of appearance and habitat partitioning by
stream larval fishes. Trans.Am.Fish Soc., 113:217–33

331
Fontenele, O., 1953. Contribucao para o conhecimiento da biologaa da curimata pacu Prochilodus
argentaeus (Spix). Rev.Bras.Biol., 13:87–102
Fox, P.J., et al., 1976. Preliminary observations on fish communities of the Okavango Delta. In
Proceedings of the Symposium on the Okavango Delta and its future utilization. 30 August-2
September 1976. Gaberones, National Museum, Botswana Society, pp. 125–30
Franceschi, E.A. and J.P. Lewis, 1979. Notas sobre la vegetacion de valle Santafesina del Rio Parana
(Republica Argentina). Ecosur, 6(11):55–82
Frank, S., 1959. Zavislost rustu nechterych druhu ryb na potravnich podminkach v polabske tuni
poltruba. Vestn.Cesk.Spol.Zool., 23:247–53
Fraser, T.M., 1972. Fishermen of the middle Mekong. Paper presented to the SEADAG Mekong
development seminar, Santa Barbara, Calif., 1972 (mimeo)
Frost, W.E. and M.E. Brown, 1967. The trout. London, Collins, 286 p.
Fryer, G. and P.J.P. Whitehead, 1959. The breeding habits, embryology and larval development of
Labeo victorianus Boulenger (Pisces: Cyprinidae). Rev.Zool.Bot. Afr., 59(1–2):33–49
Gagliano, S.M. and J.L. van Beek, 1975. Environmental base and management study. Atchafalaya
Basin, Louisiana, U.S. Environment Protection Agency Report
Gallais, J., 1967. Le Delta Interieur du Niger; etude du geographie regionale. Mem.
Inst.Fondam.Afr.Noire., (79)2 vols.: 619 p.
Gawrishova, N.A., A.I. Iwanow and L.E. Kostikowa, 1982. Bacterio- und Phytoplankton der Donau-
und Dnjestr-Mundungsgebeit. In XX Internationale Tagung, Arbeitsgemeinschaft Donauforschung,
Kiev, Naukowa Dumka, pp. 146–50
Gaygalas, K.S. and D.P. Blatneve, 1971. Growth characteristics and the structure and abundance of
the bream (Abramis brama (L.)) stock in the watercourses of the summer polders of the Nyumunas
River Delta. J.Ichthyol., 11(5):682–92
Geisler, R., 1969. Untersuchungen uber der biochemischen sauerst offbedarf und der
sauerstoffvenbranch von Fischer in einem tropischen schwartzwasser (rio Negro, Amazonien,
Brazilien). Arch.Hydrobiol., 66:307–25
Geisler, R., H.A. Knoppel and H. Sioli, 1973. The ecology of freshwater fishess in Amazonia; present
status and future tasks for research. In Applied sciences and development, Vol. 2. Tubingen,
Institute for Scientific Cooperation, pp. 144–62
Geisler, R., G.W. Schmidt and S. Sookvibul, 1979. Diversity and biomass of fishes in three typical
streams of Thailand. Int.Rev.Ges.Hydrobiol., 64(5):673–97
Gessner, F., 1960. Limnologische untersuchungen am zusammerflus des rio Negro und des Amazonas
(Solimoes). Int.Rev.Ges.Hydrobiol., 45:55–79
Gibbs, R.J. 1970. Mechanisms controlling world water chemistry. Science, Wash., 170:1088–90

332
Gilmore, K.S., 1976. Development potential and constraints of a fishing industry in Okavango Delta.
In Proceedings of the Symposium on the Okavango Delta and its future utilisation. National
Museum, Gaberone, Botswana. Gaberone, Botswana Society, pp. 175–8
Godoy, M.P., 1959. Age, growth, sexual maturity, behaviour, migration, tagging and transplantation
of the curimbata, Prochilodus scrofa, Steindachner, 1881, of the Mogi Guassu River, Sao Paulo
State, Brazil. Ann.Acad.Bras.Cienc., 31:447–77
Godoy, M.P., 1975. Peixes do Brasil Suborden Characoidei Bacia do Rio Mogi Guassu. Piracicaba,
Editora Franciscana, 4 vols., pag. var.
Gomez, A.L. and F.P. Montiero, 1955. Estudo da populacao total de peixes da represa da Estacao
Experimental de Biologia e piscicultura em Pirassununga Sao Paulo. Rev.Biol.Mar.Valp., 6:82–154
Gonzales-Jimenez, 1977. The Capybara. World Anim.Rev., 21:24–30
Goodnight, W.H. and T.C. Bjornn, 1971. Fish production in two Idaho streams. Trans. Am.Fish Soc.,
4:769–80
Gorman, O.T. and J.R. Karr, 1978. Habitat structure and stream fish communities. Ecology,
59(3):507–15
Gosse, J.P., 1963. Le milieu aquatique et l'ecologie des poissons dans la region de Yangambi.
Ann.Mus.R.Afr.Cent., 116:113–271
Goulding, M., 1980. The fishes and the forest. Explorations in Amazonian natural history. Berkley,
Los Angeles, Universitu of California Press, 280 p.
Goulding, M., 1981. Man and fisheries on an Amazonian frontier. The Hague, Dr. W. Junk,, 137 p.
Goulding, M. and M.L. Carvalho, 1982. Life history and management of the tambaqui (Colossoma
macropomum; Characidae): an important Amazonian food fish. Rev. Bras.Zool., 1:107–33
Granados, J.F., 1975. Estimaciones de la captura, esfuerzo y poblacion pesquera en los rios
Magdalena, Cauca y San Jorge 1974–75. Bogota, INDERENA, 48 p.
Graynoth, E., 1979. Effects of logging on stream environments and faunas in Nelson.
N.Z.J.Mar.Freshwat.Res., 13(1):79–109
Green, E., 1963. Zooplankton of the River Sokoto, the rhizopod Testacea. Proc.Zool.Soc. Lond.,
141:497–514
Green, E., 1970. Freshwater ecology of the Mato Grosso, Central Brazil. 1. The conductivity of some
natural waters. J.Nat.Hist.Lond., 4:289–99
Green, E., 1970a. Freshwater ecology of the Mato Grosso, Central Brazil. 3. Associations of rotifera
in meandering lakes of the Rio Suia Missu. J.Nat.Hist. Lond., 6:229–41
Green, E., 1972. Freshwater ecology of the Mato Grosso, Central Brazil. 2. Associations of Cladocera
in meandering lakes of the Rio Suia Missu. J.Nat.Hist.Lond., 6:215–27
Greenway, P.J. and D.F. Vesey-Fitzgerald, 1969. The vegetation of Lake Manyara National Park.
J.Ecol., 57:129–49

333
Greenwood, P.H., 1958. Reproduction of the East African lung fish Protopterus aethiopicus Heckel.
Proc.Zool.Soc.,Lond., 130:547–67
Gregory, K.J. and D.E. Walling, 1973. Drainage basin form and process. New York, J. Wiley and
Sons, 458 p.
Griswold, B.L., et al., 1978. Some effects of stream channelization on fish populations, macro-
invertebrates, and fishing in Ohio and Indiana. Columbia, Missouri, U.S. Fish and Wildlife
Service, 64 p.
Groen, C.L. and J.C. Schmulbach, 1978. The sport fishery of the unchannelized and channelized
middle Missouri river. Trans.Am.Fish.Soc., 107(3):412–8
Grossman, G.D., P.B. Moyle and J.O. Whitaker, Jr., 1982. Stochasticity in structural and functional
characteristics of an Indiana stream fish assemblage: a test of community theory. Am.Nat.,
120(4):423–54
Guillory, V., 1979. Utilization of an inundated floodplain by Mississippi river fishes. Fl.Sci.,
42(4):222–8
Gunda, B., 1984. The fishing culture of the world: studies in ethology, cultural ecology and folklore.
Budapest, Akademiai Kiado, 2 Vols. 1253 p.
Gunderson, D.R., 1968. Floodplain use related to stream morphology and fish populations.
J.Wildl.Manage., 32:507–14
Gupta, S.D. and A.G. Jhingran, 1973. Ageing Labeo calbasu (Hamilton) through its scales. J.Inland
Fish.Soc.India, 5:126–8
Halek, L.C. and E.K. Balon, 1983. Structure and ecological production of the fish taxocene of a small
floodplain system. Can.J.Zool., 61:2446–64
Hall, J.B. and W. Pople, 1968. Recent vegetational changes in the lower Volta River. Afr.J.Sci., 8(1–
2):24–9
Hall, J.B., I.M. Valente and B.R. Davies, 1977. The Zambezi river in Mocambique: the physico-
chemical status of the Middle and Lower Zambezi prior to the closure of the Cabora Bassa Dam.
Freshwat.Biol., 7:187–206
Hammerton, D., 1972. The Nile River - a case history. In River ecology and man, edited by T.
Oglesby, C.A. Carlson and J.A. McCann. New York, Academic Press, pp. 171–213
Hardlo, W.L. and G.J. Williams, 1984. Development of the Kafue Flats: the last five years. Lusaka,
University of Zambia
Harrel, R.C., B.J. Davis and T.C. Dorris, 1967. Stream order and species diversity of fishes in an
intermittent Oklahoma stream. Am.Midl.Nat., 78:428–36
Harriman, R., and B.R.S. Morrison, 1982. Ecology of streams draining forested and non-forested
catchments in an area of Central Scotland subject to acid precipitation. Hydrobiologia, 88:251–63
Harrison, A.D., 1965. River zonation in Southern Africa. Arch.Hydrobiol., 61(3):380–86

334
Harrison, A.D., 1966. Recolonization of a Rhodesian stream after frought. Arch. Hydrobiol., 62:405–
21
Harrow, L.G. and A.B. Schlesinger, 1980. The larval fish recruitment study. Environ.
Ser.Bull.Omaha, Nebraska, (5):92 p.
Hastings, R.E., 1972. Interim report 1970–1972. Fisheries Research Unit, Lower Shire. Zomba,
Malawi, Fisheries Department, July. Unpag.
Hawkes, H.A., 1975. River zonation and classification. In River ecology, edited by R.B. Whitton.
London, Blackwell, pp. 312–74
Hawkins, C.P. and J.R. Sedell, 1981. Longitudinal and seasonal changes in functional organization of
macroinvertebrate communities in four Oregon streams. Ecology, 62(2):389–97
Hayward, P.B. 1984. Status report on the Kafue fishery. In Development on the Kafue Flats: the last
five years. Kafue Basin Research Project, Lusaka, University of Zambia, pp. 58–73.
Heckman, C.W., 1979. Rice field ecology in Northeastern Thailand. The Hague, Junk, 228 p.
Heeg, J. and C.M. Breen, 1982. Man and the Pongolo floodplain. S.Afr.Nat.Sci.Prog.Rep., (56):117 p.
Hesse, L.W., et al., 1982. The middle Missouri River. Norfolk, Missouri, The Missouri Study Group,
301 p.
Holcik, J., 1972. Abundance, ichthyomass and production of fish populations in three types of water-
bodies in Czechoslovakia. In Productivity problems of freshwaters, edited by Z. Kajak and A.
Hillbricht-Ilkowska. Warsaw, Polish Scientific Publishers, pp. 834–55
Holcik, J., 1979. Assessment of fish stock and prediction of catch in large rivers. FAO
Fish.Tech.Pap., (194):26–35
Holicik, J. and I. Bastl, 1973. Ichtyocenozy dvoch Dunajskych ramien so zertelom na zmeny v ich
druhovom ziozeni a hustote vo vztaha ku kolisaniu hladiny v hlavnom toku. Biol.Pr., 19(1):106 p.
Holcik, J., 1976. Ecological effects of water level fluctuations upon the fish populations of in the
Danube River floodplain in Czechoslovakia. Acta Sci. Nat.Brno, 10(9):1–46
Holcik, J., 1977. Predicting fish yield in the czechoslovakian section of the Danube River based on
the hydrological regime. Int.Rev.Ges.Hydrobiol., 62(4):523–32
Holcik, J., et al., 1981. Hydrobiology and ichthyology of the Czechoslovak Danube in relation to
predicted changes after the construction of the Gabcikovo-Nagymaros river barrage system.
Pr.Lab.Rybar.Hydrobiol., 3:19–158
Holden, M.J., 1963. The populations of fish in dry season pools of the River Sokoto.
Fish.Publ.Colon.Off.,Lond., (19):58 p.
Holden, M.J. and J. Green, 1960. The hydrology and plankton of the River Sokoto. J.Anim.Ecol.,
29:65–84

335
Holden, P.B., 1979. Ecology of riverine fishes in regulated stream systems with emphasis on the
Colorado River. In The ecology of regulated streams, edited by J.V. Ward and J.A. Stanford. New
York, Plenum Publishing Corporation, pp. 57–74
Holden, P.B. and C.B. Stalnaker, 1975. Distribution and abundance of mainstream fishes in the
middle and upper Colorado River basins, 1967–1973. Trans.Am.Fish.Soc., 104:217–31
Holland, H.D. 1978. The chemistry of the atmosphere and oceans. Chapter IV. Physical and chemical
transport in river systems. New York, John Wiley and Sons, pp. 81–152
Honda, E.M.S., 1974. Contribuçao as conhecimientos da biologia de peixes do Amazonas. 2.
Alimentaçao de tambaqui, Colossoma bidens (Spix). Acta Amazonica, 4:47–53
Hopkins, C.L., 1971. Production of fish in two small streams in the North Island of New Zealand.
N.Z.J.Mar.Freshwat.Res., 5(2):280–90
Horton, R.E., 1945. Erosional development of streams and their drainage basins: hydrophysical
approach to quantitative morphology. Bull.Geol.Soc.Am., (56):275–370
Horwitz, R.J., 1978. Temporal variability patterns and the distributional patterns of stream fishes.
Ecol.Monogr., (48):307–21
Howard-Williams, C. and W.J. Junk, 1976. The decomposition of aquatic macrophytes in the floating
meadows of a central Amazonian varzia lake. Biogeographica, 7:115–23
Howard-Williams, C. 1977. The chemical composition of central Amazonian aquatic macrophytes
with special reference to their role in the ecosystem. Arch.Hydrobiol., 79(4):446–64
Howard-Williams C.M. and G.M. Lenton, 1975. The role of the littoral zone in the functioning of a
shallow tropical lake ecosystem. Freshwat.Biol., 5:445–59
Huet, M. 1947. Appreciation de la valeur piscicole des eaux douces. Trav.Stn.Rech.Eaux
For.Groenendaal-Hoeilaart (D), (10):1–15
Huet, M, 1949. Apercu des relations entre la pente et la population des eaux courantes.
Schweitz.Z.Hydrol., 11:333–51
Huet, M, 1964. The evaluation of the fish productivity in fresh waters. The coefficient of productivity.
Verh.Int.Ver.Theor.Angew.Limnol., 15:524–8
Huet, M. and J.A. Timmermans, 1963. La population piscicole de la Semois inferieur, grosse riviere
Belge du type superieur de la zona a barbeau. Trav.Stn.Rech. Eaux For.Groenendaal-Hoeilaart
(D), (36):31 p.
Hughes, J.M. (ed.), 1976. Respiration of amphibious vertebrates. London, Academic Press, 402 p.
Hurault, J., 1965. Les principaux types de peuplements du sud-est Dahomey, et leur representation
cartographique. Etud.Photo-Interpret.Inst.Geogr.Natl., Paris, (2):79
Husain, Z., 1973. Fish and fisheries of the lower Indus basin (1966–67). Agric.Pak., 24(3–4):297–322
Husar, S.L., 1975. A review of the literature on the dugong (Dugong dugon).
Wildl.Res.Rep.U.S.Fish.Wildl.Serv., (4):30 p.

336
Hynes, H.B.N., 1970. The ecology of running waters. Liverpool, Liverpool University Press, 555 p.
Hynes, J.D., 1975. Annual cycles of macro-invertebrates of a river in Southern Ghana.
Freshwat.Biol., (5):71–83
Ihering, R. von, 1930. La piracema ou montee du poisson. C.R.Seanc.Soc.Biol., Paris, 103:1336–8
Illies, J. and L. Botosaneanu, 1963. Problemes et methodes de la classification de la zonation
ecologique des eaux courantes, considerees surtout du point de vue faunistique.
Mitt.Int.Verein.Theor.Angew.Limnol., 12:1–57
Iltis, A., 1982. Peuplements algaux des rivieres de Cote d'Ivoire. II. Variations saisonnieres des
biovolumes, de la composition et de la diversite specifique. Rev.Hydrobiol.Trop., 15(3):241–52
Imevbore, A.M.A. and O. Bakare, 1974. A preimpoundment study of the swamps of the Kainji lake
basin. Afr.J.Trop.Hydrobiol.Fish., 3(1):79–94
INDERENA, 1973. Operación Subienda 1973. Investigacion pesquera. Bogota, Colombia, Instituto
de Desarollo de los Recursos Naturales Renovables, 133 p.
Inger, R.F. and P.K. Chin, 1962. The Freshwater fishes of North Borneo. Fieldiana (Zoology), 45:1–
268
Inyang, N.M., 1984. On the biology of Macrobrachium felicinum Holthius in Lower Niger River of
South-eastern Nigeria. Rev.Zool.Afr., 98(2):440–9
Islam, B.N. and G.B. Talbot, 1968. Fluvial migration, spawning and fecundity of Indus River Hilsa,
Hilsa ilisha. Trans.Am.Fish.Soc., 97:350–5
ITC (International Trade Centre), 1979. International trade in tropical aquarium fish. Geneva,
UNCTAD/GATT, 137 p.
Ivlev, V.S., 1966. The biological production of rivers. J.Fish.Res.Board Can., 23(11):1727–59
Izvoranu, 1982. Einege aspkte der phytophylen Fauna des Porcu- und Rosu-Sees des Donaudelta,
1978. In Internationalen Arbeitsgemeinschaft Donauforschung, pp. 145–6
Jackson, P.B.N. 1961. The fishes of Northern Rhodesia. Lusaka, Zambia, Government Printer, 140 p.
Jackson, P.B.N. 1963. Ecological factors affecting the distribution of freshwater fishes in tropical
Africa. Ann.Cape Prov.Mus., 2:223–8
Jackson, P.B.N. and P.W. Coetzee, 1982. Spawning behaviour of L. umbratus (Smith)
(Pisces:Cyprinidae). S.Afr.J.Sci., 78:293–5
Jahn, L.R. and J.B. Trefethen, 1973. Placing channel modifications in perspective. In Proceedings of
the National Symposium on waterbodies in transition. Urbana, Illinois, American Water Resources
Association, pp. 15–21
Jhingran, V.G., 1968. Synopsis of biological data on Catla catla (Hamilton 1833). FAO Fish.Synop.,
(32)Rev.1:pag. var.

337
Jhingran, V.G., 1975. Fish and fisheries of India. Delhi, India, Hindustan Publishing Corporation, 954
p.
Johnels, A.G., 1954. On the fishes from the Gambia River. Ark.Zool., 6(17):131–44
Johnels, A.G. and G.S.O. Svensson, 1954. On the biology of Protopterus annectens. Ark. Zool.,
7:131–44
Johnson, C.R., 1976. Diel variation in the thermal tolerance of Gambusia affinis (Pisces:Poeciliidae).
Comp.Biochem.Physiol.(A.Comp.Physiol.), 55:337–40
Johnson, D.S., 1967. On the chemistry of freshwaters of southern Malaya and Singapore.
Arch.Hydrobiol., 63:477–96
Johnson, D.S., 1968. Malayan blackwaters. In Proceedings of the Symposium on recent advances in
tropical ecology. Part 1, edited by R. Misra and B. Gopal. Varanasi, India, International Society for
Tropical Ecology, pp. 303–10
Junk, W.J, 1970. Investigations on the ecology and production biology of the floating meadows
(Paspalo echinochloetum) on the middle Amazon. Part 1. The floating vegetation and its ecology.
Amazoniana, 2(4):449–95
Junk, W.J, 1971. Primeiros resultados das investigacoes acerca a povoacao animal em substrato
fluctuante identico em differentes tipos de aguas de Amazonas. In IIe Simposio y Foro de biologia
tropical Amazonica, Florencia (Caqueta) y Leticia (Amazonas). Bogota, Asosiacion pro Biologia
Tropical, pp. 81–5
Junk, W.J., 1973. Investigations on the ecology and production biology of the floating meadows
(Paspalo echinochloetum) on the middle Amazon. Part 2. The aquatic fauna of the root zone of
floating vegetation. Amazoniana, 4:9–102
Junk, W.J, 1976. Faunal ecological studies in inundated areas and the definition of habitats and
ecological needs. Anim.Res.Dev., 4:47–54
Junk, W.J, 1983. Ecology of swamps in the middle Amazon. In Mires, swamp, bog, fen, moor. E.
Regional studies, edited by A.J.P. Gore. Amsterdam, Elsevier, pp. 269–96
Junk, W.J, Temporary fat storage, an adaptation of some fish species to the waterlevel fluctuations
and related environmental changes of the Amazon River. From Cooperation between Masc-Plank-
Institute for Limnology, Working Group “Tropical Ecology” Plon, West Germany and INPA,
Manaus, Brazil (in press)
Junk, W.J., G.M. Soares and F.M. Carvalho, 1983. Distribution of fish species in a lake of the
Amazon river floodplain near Manaus (Lago Cameleao), with special reference to extreme oxygen
conditions. Amazoniana, 7(4):397–431
Juris, S., 1969. Die entwicklung der alginkomponente des periphytons. In Tchechoslowakischen
bereich der Donau in jahr 1966: Limnologische Donauforchungun. Kiev, Berichte der II
Internationalen Konferenz zur limnologie der Donau, pp. 235–57
Juris, S., 1973. Quantitative studien der algenkomponente des periphytons de Tchechoslowakischen
teiles der Donau. Acta Rer.Nat.Mus.Natl.Slov., Bratislava, 19:103–11

338
Juris, S., 1975. K poznaniu fytoplanktonu Bakaskeha ramena Duneja. Acta Rer.Nat. Mus.Natl.Slov.,
Bratislava, 21:55–69
Kallemeyn, L.W. and J.F. Novotny, 1977. Fish and fish food organisms in various, habitats of the
Missouri River in South Dakota, Nebraska and Iowa. Biological Services Program. Washington,
D.C., U.S. Fish abd Wildlife Service, FWS/OBS/77/25:100 p.
Kapetsky, J.M., 1974. Growth, mortality and production in five fish species of the Kafue River
floodplain, Zambia. PhD. Dissertation, University of Michigan, 194 p.
Kapetsky, J.M., 1974a. The Kafue river floodplain: an example of preimpoundment potential for fish
production. In Lake Kariba: a man-made tropical ecosystem in Central Africa, edited by E.K.
Balon and A.G. Coche. The Hague, W. Junk, pp. 497–523
Kapetsky, J.M., Management of fisheries on large African reservoirs - an overview. Proceedings of
the National Symposium on managing reservoir fishery resources (in press)
Kapetsky, J.M., et al., 1976. Fish populations in the floodplain lakes of the Magdalena River. Second
report. Bogota, INDERENA-FAO, 30 p. (mimeo)
Kapetsky, J.M., 1977. Some ecological aspects of the shallow lakes of the Magdalena floodplain,
Colombia. Paper presented to the Intertropical Symposium on stability and diversity in tropical
communities, Panama, March 1977
Kaushik, N.K. and H.B.N. Hynes, 1971. The fate of the dead leaves that fall into streams.
Arch.Hydrobiol., 68(4):465–515
Kenmuir, D.H.S., 1976. Fish spawning under artificial flood conditions on the Mana Floodplain,
Zambezi River. Kariba Stud., 6:86–97
Kensley, B. and I. Walker, 1982. Palaeomonid shrimps from the Amazon basin, Brazil
(Crustacea:Decapoda:Natantia). Smithson.Contrib.Zool., (362):28 p.
Keulder, P.P., 1970. Orange River Research. Rep.South Afr. IBP/PF Working Group, 4:7–9
Khalil, K.F., 1971. Check list of helminth parasites of African freshwater fish.
Tech.Commun.Commonw.Inst.Helminthol., G.B., (42):80 p.
Khan, H.A. and V.G. Jhingran, 1975. Synopsis of biological data on rohu, Labeo rohita (Hamilton,
1822). FAO Fish.Synop., (111):100 p.
Kimpe, P. de, 1964. Contribution à l'étude hydrobiologique du Luapula-Moero. Ann.Mus.R.
Afr.Cent.(Ser.8 Sci.Zool.), 128:1–238
Knoppel, H.A., 1970. Food of central Amazonian fishes. Contribution to the nutrient ecology of
Amazonian rain forest streams. Amazoniana, 2(3):257–352
Koblitskaya, A.F., 1985. The significance of bayou-type spawning grounds in the Volga Delta.
J.Ichthyol., 24(4):108–18
Kofoid, C.A., 1908. The plankton of the Illinois River 1894–1899 with introductory notes upon the
hydrography of the Illinois River and its basin. Pt. 2. Constituent organisms and their spatiasl
distribution. Bull.Ill.State.Lab.Nat.Hist., (8):1–361

339
Kok, L.T., 1972. Toxicity of insecticides used for Asiatic rice borer control to tropical fish in rice
paddies. In The careless technology: ecology and international development, edited by M.T. Farvar
and J.P. Milton. Garden City, New York, National History Press, pp. 489–98
Kolbing, A., 1978. The European method of fish harvest prediction in fluvial systems.
Environ.Biol.Fish., 5(3):249–51
Kramer, D.L., 1978. Reproductive seasonality in the fishes of a tropical stream. Ecology, 59(5):976–
85
Kramer, D.L., 1981. Aquatic surface respiration in the fishes of Panama: distribution in relation to
risk of hypoxia. Environ.Biol.Fish., 8(1):49–54
Kramer, D.L. and J.B. Graham, 1976. Synchronous airbreathing, a social component of respiration in
fishes. Copeia, 1976 (4):689–97
Kramer, D.L. and M. McClure, 1982. Aquatic surface respiration, a widespread adaptation to hypoxia
in tropical freshwater fishes. Environ.Biol.Fish., 7(1):47–55
Kramer, D.L., D. Manley and R. Bourgeois, 1982. The effect of respiratory mode and oxygen
concentration on the risk of aerial predation in fishes. Can.J.Zool., 61:653–65
Kramer, D.L., et al., 1978. The fishes and the aquatic environment of the Central Amazon basin with
particular reference to repiratory patterns. Can.J.Zool., 5:717–29
Krykhtin, K.L., 1975. Causes of periodic fluctuations in the abundance of the non-anadromous fishes
of the Amur River. J.Ichthyol., 15(5):826–9
Krykhtin M.I. and E.I. Gorbach, 1981. Reproductive ecology of the grass carp, Ctenopharyngodon
idella and the silver carp, Hypophthalmichthys molitrix in the Amur basin. J.Ichthyol., 21(2):109–
23
Kuehne, R.A., 1962. A classification of streams illustrated by fish distribution in an east Kentucky
creek. Ecology, 43:608–14
Kushlan, J.A., 1976. Environmental stability and fish community diversity. Ecology, 57:821–5
Kutznetsov, V.A., and N.I. Fedeyev, 1979. Some features of the reproduction of fishes in parts of the
Volga before and after flow regulation. J. Ichthyol., 19(1):81–90
Laboratoire d'Hydrobiologie de Mopti, 1981. Recherches ichthyologiques dans le Delta Central du
fleuve Niger. Projet NOVIB - Rapport final, Mopti, 113 p.
Lagler, K.E., et al., 1977. Ichthyology. New York, J. Wiley and Sons, 506 p. 2nd ed.
Lassleben, P., 1977. Las Schatzverfakren fur Fischwasser nach Leger und Huet. Osterr.Fisch., 28:53–
64
Le Van Dang, 1970. Contribution to a biological study of the Lower Mekong. In Proceedings of the
IBP regional meeting of inland water biologists in Southeast Asia at Kuala Lumpur and Malacca
(Malaysia), 5–11 May 1969. Djakarta, Unesco Field Science Office for Southeast Asia, pp. 59–64

340
LeCren, E.D., 1965. Some factors regulating the size of populations of freshwater fish.
Mitt.Int.Ver.Theor.Angew.Limnol., 13:88–105
LeCren, E.D., 1969. Estimates of fish populations and production in small streams in England. In
Symposium on salmon and trout in streams edited by T.G. Northcote. Vancouver, Institute of
Fisheries, pp. 269–80
Leggett, W.C. and J.E. Carscadden, 1978. Latitudinal variation in reproduction characteristics of
American shad (Alosa sapidissima): evidence for population specific life history strategies in fish.
J.Fish.Res.Board Can., 35(11):1469–78
Leibman, M. and H. Reichenbach-Klinke, 1967. Eingriffe des menschens und deren biologische
auswirkung. In Limnologie der Donau, edited by R. Liepolt. Stuttgart, E. Schweitzerbart'sche
Verlagsbuchhandlung, pp. 1–25
Leidy, R.A., 1983. Distribution of fishes in streams of the Walnut creek basin, California. Calif.Fish
Game, 69(1):29–32
Lelek, A., 1968. The vertical distribution of fishes in the Ebo stream and notes to the fish occurence in
the L. Bosumtwi, Ashanti, Ghana. Zool.Listy., 17(3):245–52
Lelek, A. and S. El Zarka, 1973. Ecological comparison of the preimpoundment and
postimpoundment fish faunas of the River Niger and Kainji Lake, Nigeria. Geophys.Monogr.,
17:655–60
Lellack, J., 1966. Influence of the removal of the fish population on the bottom animals of five Elbe
backwaters. Hydrobiol.Stud., 1:323–80
Leopold L.B. and T. Maddock Jr., 1953. The hydraulic geometry of stream channels and some
physiographic implications. Prof.Pap.U.S.Geol.Surv., (252)
Leopold, L.B., M. B. Wolman and J. P. Miller 1964. Fluvial processes in geomorphology. San
Francisco, W. H. Freeman, 522 p.
Letichevskiy, M.A., 1981. Prospects of increasing the artificial and natural reproduction of the
inconnu, Stenodus leucichthys, in the lower course and delta of the Volga. J.Ichthyol., 5:103–7
Leveque, C., J-R. Durand and J-M. Ecoutin, 1977. Relation entre le rapport P/B et al lngevite des
organismes. Cah.ORSTOM (Hydrobiol.), 11(1):17–31
Levine, J.S. and E.F. MacNichol Jr., 1982. Color vision in fishes. Sci.Am., 246(2):140–9
Levine, J.S., et al., 1980. Visual communication in fishes. In Environmental physiology of fishes,
edited by M.A. Ali. New York, Plenum Press, pp. 443–75
Lewis, W.M., 1970. Morphological adaptations in cyprinodonts for inhabiting oxygen deficient
waters. Copeia, 1970(2):319–26
Liepolt, R., 1972. Uses of the Danube River. In River ecology and man, edited by R.T. Oglesby, C.A.
Carlson and J.A. McCann. New York, Academic Press, pp. 233–49
Liepolt, R. (ed.), 1967. Limnologie der Donau. Stuttgart, E. Schweitzerbart'sche
Verlagsbuchhandlung, Pag. var.

341
Livingstone, D.A., 1963. Chemical composition of rivers and lakes. In Data of Geochemistry, edited
by M. Fleischer. Washington D.C., Government Printing Office, Ch. 4.
Lobon-Cervia, J. and T. Penczak, 1984. Fish production in the Jaruma River, Central Spain. Holarctic
Ecol., 7:128–37
Loftus, K.H., 1976. Science for Canada's fisheries rehabilitation needs. J.Fish.Res.Board Can.,
33(8):1822–57
Lotrich, V.A., 1973. Growth, production and community composition of fishes inhabiting a first-,
second-, and third order stream of Eastern Kentucky. Ecol.Monogr., 43(3):377–97
Loubens, G., 1969. Etude de certains peuplements ichtyologiques par des peches au poison (lre note),
Cah.ORSTOM (Hydrobiol.), 3(2):45–73
Loubens, G., 1970. Etude de certains peuplements ichtyologiques par des peches au poison (2e note).
Cah.ORSTOM (Hydrobiol.), 4(1):45–61
Lowe-McConnell, R.H., 1964. The fishes of the Rupununi savanna district of British Guiana, South
America. Part 1. Ecological groupings of fish species and effects of the seasonal cycle on the fish.
J.Linn.Soc.(Zool.), 45:103–44
Lowe-McConnell, R.H., 1967. Some factors affecting fish populations in Amazonian waters. In Atas
do Simposio sobre a biota Amazonia, Conselho Nacional Pesquisas. Rio de Janiero, Consevacao de
Natureza e Recursos Naturais, pp. 117–86
Lowe-McConnell, R.H., 1975. Fish communities in tropical freshwaters. London, Longman, 337 p.
Lowe-McConnell, R.H., 1977. Ecology of fishes in tropical waters. Inst.Biol.Stud.Biol., (76):64 p.
Mago-Leccia, F., 1970. Estudios preliminares sobre la ecologia de los peces de los llanos de
Venzuela. Acta.Biol.Venez., 7(1):71–102
Mahon, R., 1981. Patterns of fish taxocenes of small streams in Poland and Ontario: a tale of two river
basins with a test for accuracy of quantitative sampling. PhD. Thesis, University of Guelph,
Guelph, Ontario, pag. var.
Mahon, R., 1984. Divergent structure in fish taxocenes of north temperate streams.
Can.J.Fish.Aquat.Sci., 41(2):330–50
Mahon, R., E.K. Balon and D.L.G. Noakes, 1979. Distribution, community structure and production
in the Upper Speed River, Ontario, A preimpoundment study. Env.Biol.Fish., 4(3):219–44
Malaisse, F., 1969. Les faciès d'un cours d'eau tropical: La Luanza (Haut-Katanga, Rép. Dém.
Congo). Verh.Int.Ver.Theor.Angew.Limnol., 17:956–1040
Malaisse, F., 1976. Ecologie de la riviere Luanza, edited by J.J. Symoens. Bruxelles, Cercle
Hydrobiologique de Bruxelles, 151 p.
Mann, K.H., 1965. Energy transformations by fish in the River Thames. J.Anim.Ecol., 40:155–90
Mann, K.H., 1972. Case history: the River Thames. In River Ecology and man, edited by R.T.
Oglesby, C.A. Carlson and J.A. McCann. New York, Academic Press, pp. 215–32

342
Mann, R.H.K., 1971. The population, growth and production of fish in four small streams in Southern
England. J.Anim.Ecol., 40:155–90
Marlier, G., 1951. Recherches hydrobiologiques dans les rivieres du Congo oriental. Composition des
eaux. La conductibilite electrique. Hydrobiologia, 3:217–27
Marlier, G., 1967. Ecological studies of some lakes of the Amazon valley. Amazoniana, 1:91–115
Marlier, G. 1973. Limnology of the Congo and Amazon rivers. In Tropical forest ecosystems in
Africa and South America: A comparative review, edited by B.J. Meggers, S.A. Ayensu and W.D.
Duckworth. Washington, D.C., Smithsonian Institution Press, pp. 223–38
Marlier, G., 1967. Ecological studies on some lakes of the Amazon valley. Amazoniana, 1:91–115.
Marshall, B.E., 1982. The influence of river flow on pelagic sardine catches in L. Kariba. J.Fish Biol.,
20:465–69
Martin, J.M. and M. Meybeck, 1976. The content of major elements in the dissolved and particulate
load of rivers. In Proceedings of the Unesco/SCOR Workshop 29 Nov-Dec 1976. Paris, Unesco,
pp. 96–110
Martin, J.M. and M., 1979. Elemental mass balance of material carried by major world rivers.
Mar.Chem., 7:173–206
Marxsen, J. von, 1980. Investigations on the ecology of suspended bacteria in small streams.
Arch.Hydrobiol., Suppl.58(1/8):26–55
Mathews, C.P., 1971. Contribution of young fish to total production of fish in the River Thames near
Reading. J.Fish Biol., 3:157–80
Matthes, H., 1964. Les poissons du lac Tumba et de la region d'Ikela. Etude systématique et
écologique. Ann.Mus.R.Afr.Cent.(Ser.8 Sci.Zool.), (126):204 p.
Matthes, H., 1977. The problems of rice eating fish in the Central Delta, Mali. CIFA
Tech.Pap./Doc.Tech.CPCA, (5):225–52
McArthur, R.H. and E.O. Wilson, 1967. The theory of island biogeography. Princeton, N.J., Prnceton
University Press, 203 p.
McFadden, J.T., 1969. Dynamics and regulation of salmonid populations in streams. In Symposium
on salmon and trout in streams. H.R. MacMillan lectures in Fisheries. Vancouver, University of
British Colombia, pp. 313–29
McFadden J.T. and E.L. Cooper, 1962. An ecological comparison of six populations of brown trout
(Salmo trutta). Trans.Am.Fish.Soc., 91:53–62
McIntosh, R., 1978. Distribution and food of the cormorant on the lower reaches of the River Tweed.
Fish.Manage., 9(4):107–13
Mefit-Babtie, 1983. Development studies in the Jonglei canal area; Final Report to Government of the
Democratic Republic of the Sudan. Glasgow, Mefit–Babtie, 4 Vols:107 p., vol.2:273 p., vol.3:265
p., vol.4:87 p.

343
Menezes, N.A., H.J. Wagner and M.A. Ali, 1981. Retinal adaptations in fishes from a floodplain
environment in the central Amazon basin. Rev.Can.Biol., 40(1):111–32
Merona, B. de, 1981. Zonation ichtyologique du bassin du Bandama (Cote d'Ivoire). Rev.
Hydrobiol.Trop., 14(1):63–75
Merona, B., 1983. Modele d'estimation rapide de la croissance des poissons. Application aux poissons
d'eau douce d'Afrique. Rev.Hydrobiol.Trop., 16(1):103–13
Meschkat, A., 1972. Nuevas tecnicas de pesca fluvial recomendadas para el Equador. Quito, Servicio
Florestal y Piscicultura, 34 p.
Meschkat, A., 1975. Aquacultura e pesca em aguas interiores no Brasil. Doc.Tec.SUDEPE, (9):35 p.
Meybeck, M., 1976. Total mineral dissolved transport by world major rivers.
Hydrol.Sci.Bull.Sci.Hydrol., (21):265–84
Meybeck, M., 1979. Concentrations des eaux fluviales en éléments majeurs et apport en solution aux
oceans. Rev.Geol.Dyn.Geogr.Phys., Paris, 21(3):215–46
Micha, J.C., 1973. Etude des populations piscicoles de l'Oubangui et tentatives de selection et
d'adaptation de quelques especes a l'etang de pisciculture. Nogent -sur-Marne, Centre Technique
Forestier Tropical
Mikkola, H. and P.A. Arias, 1976. Evaluacion preliminar de la limnologia y de las poblaciones de
peces en el sistema del canal del Dique. Parte 1. Limnologia. Bogota, Proyecto Pesca Continental,
INDERENA/FAO, 65 p.
Mitchell, D.S., T. Petr and A.B. Viner, 1980. The water fern Salvinia molesta in the Sepik River,
Papua New Guinea. Environ.Conserv., 7(2):115–28
Mizuno, T. and S. Mori, 1970. Preliminary hydrobiological survey of some Southeast Asian inland
waters. Biol.J.Linn.Soc., 2:77–117
Moghraby, A.I. el., 1977. A study on diapause of zooplankton in a tropical river - the Blue Nile.
Freshwat.Biol., 7:77–117
Monakov, A.V., 1969. The zooplankton and the zoobenthos of the White Nile and adjoining waters in
the Republic of the Sudan. Hydrobiologia, 33:161–85
Moodie, G.E. and M. Power, 1982. The reproductive biology of an armoured catfish, Loricaria
uracantha, from Central America. Environ.Biol.Fish., 7(2):143–8
Mordukhai-Boltovskoi, P.D., 1979. The River Volga and its life. The Hague, W. Junk, Monogr.Biol.,
33:473 p.
Moreau, J., 1980. Essai d'application au Lac Alaotra (Madagascar) d'un modele d'etude de pecheries
pour les plaines d'inondation intertropicales. Cah.ORSTOM (Hydrobiol.), 13(1–2):83–91
Moreau, J., 1982. Le Lac Ihotry, lac plat hypersalé (Madagascar); ecologie et peuplement piscicole.
Rev.Hydrobiol.Trop., 15(1):1–82

344
Morris, L.A., et al., 1968. Effects of main stem impoundments and channelization upon the limnology
of the Missouri River, Nebraska. Trans.Am.Fish.Soc., 91:380–8
Moyle, P.B. and H.W. Li, 1979. Community ecology and predator-prey relations in warmwater
streams. In Predator-prey systems in fisheries management, edited by H. Clepper. Washington,
D.C., Sport Fisheries Institute, pp. 171–80
Mucha, V., 1967. Die microbiologie der Donau. In Limnologie der Donau, edited by R. Liepolt.
Stuttgart, E. Schweitzerbart'sche Verlagsbuchhandlung. Pag. var.
Mulholland, P.J., 1981. Deposition of riverborne organic carbon in floodplain wetlands and deltas. In
Flux of organic carbon by rivers to the oceans, U.S. Department of Energy. Springfield, National
Technical Information Service, CONF-8009140 UC-11, pp. 142–72
Muncy, R.J., 1973. A survey of the major fisheries of the Republic of Zambia. Rome, FAO, FI:DP
9/10 ZAM 511/3, 69 p.
Muncy, R.J., 1978. An evaluation of the Zambian Kafue River floodplain fishery. CIFA
Tech.Pap./Doc.Tech.CPCA, (5):153–64
Muth, R.T. and J.C. Schmulbach, 1984. Downstream transport of fish larvae in a shallow prairie river.
Trans.Am.Fish Soc., 113:224–30
Nagaraja Rao, S. and T. Rajalakshmi, 1977. Investigations on the fisheries of River Godavari, Andhra
Pradesh: India. Proc.IPFC, 17(3):162–6
Natarajan, A.V. and A.G. Jhingran, 1963. On the biology of Catla catla (Ham.) from the River
Yamuna. Proc.Natl.Inst.Sci.India (B), 29(3):326–55
Nelson, K., 1964. Behaviour and morphology in the glandulocaudine fishes (Ostariophysi,
Characidae). Univ.Cal.Publ.Zool., 75(2):59–152
Newbold, J.D., et al., 1981. Nutrient spiralling in streams: the concept and its field measurement.
Can.J.Fish.Aquat.Sci., 38(7):860–3
Nezdoliy, V.K., 1984. Downstream migration of young fishes during the initial period of flow
regulation of the Ili River. J.Ichthyol., 24(2):34–46
Nicolau, A., 1952. Vegetatia acuatica factor important in determinarea conditiilor hydrobiologice din
baltile regiunii inundbile a Dunarii. Bull.Inst.Cerct. Piscic., 11(4):53–69
Nieff, J.J., 1975. Fluctuaciones annuales en la composicion fitocenotico y biomasa de la hidrofita en
lagunas islenas del Parana medio. Ecosur, 2(4):153–83
Nikolsky, G.V., 1937. On the distribution of fishes according to the nature of their food in the rivers
flowing from the mountains of Middle Asia. Verh.Int.Ver. Theor.Angew.Limnol., 8:170–6
Nikolsky, G.V., 1956. Ry by basseyna Amura (The fishes of the Amur basin), Moscow, Akadamiia
Nauk SSR, 551 p.
Nishiwaki, M., et al., 1982. Recent survey on the distribution of the African manatee. Sci.Rep.Whales
Res.Inst., Tokyo, (34):137–47

345
Norman, J.R., 1975. A history of fishes. London, Ernest Benn, 467 p. 3rd ed.
NOVIB, 1981.
Novoa, D. (ed.), 1982. Los recursos pesqueros del Rio Orinoco y su explotacion. Caracas,
Corporacion Venezolana de Guayana, Division de desarollo agricola, 386 p.
Novotna, M. and V. Korinek, 1966. Effect of the fish-stock on the quantity and species composition
of the plankton of two backwaters. Hydrobiol.Stud., 1:297–322
Ohya, M., 1966. Comparative study on the geomorphology and flooding in the plains of the Cho-
Shui-Chi, Chao-Phrya, Irrawaddy and Ganges. In Scientific problems of the humid tropical zone
deltas and their implications. Proceedings of the Dacca Symposium, 1964. Paris, Unesco, pp. 23–8
Okedi, J., 1969. Observations on the breeding and growth of certain mormyrid fishes of the L.
Victoria basin. Rev.Zool.Bot.Afr., 79(1–2):34–6
Okedi, J., 1971. Further observations on the ecology of the Nile perch (Lates niloticus) in Lake
Victoria and Lake Kyoga. Rep.E.Afr.Freshwat.Fish.Res.Org., (1970):42–54
Oliva, O., 1960. Further contributions to the fish population composition in middle Bohemia.
Vestn.Cesk.Spol.Zool., 24:42–9
OMVS (Organisation pour la mise en valeur du fleuve Senegal), Evaluation des effects sur
L'environment d'amenagements prevues dans le bassin du fleuve Senegal. Senegal, Dakar, OMVS.
Operation Pêche, 1977. Rapport annuel 1976. Mopti, Mali, Operation Pêche, 44 p.
Osmera, S., 1973. Annual cycle of zooplankton in backwaters of the flood area of the Dyje.
Hydrobiol.Stud., 3:219–56
Otobo, F.O., 1978. Commercial fishery in the middle Niger, Nigeria. CIFA Tech.Pap./Doc.
Tech.CPCA, 5:185–208
Paiva, M.P., 1973. Recursos pesqueiros e a pesca ,na bacia do rio Parnaiba (Brasil).
Bol.Ceará.Agron., (14):49–82
Paiva, M.P., 1976. Estimativa do potencial da producao de pescado em grandes represas brasileiras.
Rio de Janeiro, Electrobras
Paiva, M.P. and S.A. Bastos, 1982. Marcacao de peixes nas regioes do alto e medio Sao Francisco
(Brasil). Cien.Cult., 34:1362–5
Pantulu, V.R., 1970. Some biological cosiderations related to the Lower Mekong development. In
Proceedings of the Regional Meeting of Inland Water Biologists in Southeast Asia, at Kuala
Lumpur and Malacca (Malaysia) 5–11 May 1969. Djakarta, UNESCO Field Science Office for
Southeast Asia, pp. 113–9
Parameswaran, S., C. Selvaraj and S. Radhakrishnan, 1970. Observations on the maturation and
breeding cycle of carps in Assam. J.Inland Fish.Soc., India, 2:16–29

346
Pardo, G., 1976. Inventario y zonificacion de la cuenca para fines hidroagricolas, in Conferencias del
Foro. In Foro aprovechamientos de proposito multiple proteccion contra las inundaciones. Bogota,
Proyecto Cuenca de la Magdalena, Cauca, Convenio Colombo Holandes, D3–1 to D3–7
Patil, M.R., 1977. Pollutional effects of industrial wastes on riverine fisheries of India. Proc.IPFC,
17(30:268–71
Paugy, D., 1978. Ecologie et biologie des Alestes baremoze (Pisces, Characidae) des rivieres de Cote
d'Ivoire. Cah.ORSTOM (Hydrobiol.), 12(3–4):245–75
Paugy, D., 1982. Synonomie d'Alestes rutilus Blgr., 1916, avec A. macrolepidotus (Val., 1849).
Biologie et variabilite morphologique. Rev.Zool.Afr., 96(2):286–315
Paugy, D., 1980. On the interrelationships between natural mortality, growth parameters and mean
environmental temperature in 175 fish stocks. J.Cons. CIESM, 39:175–92
Pavlov, D.S.,et al., 1977. Some features of the downstream migrations of juvenile fishes in the Volga
and Kuban Rivers. J.Ichthyol., 19(3):363–74
Penczak, T., 1981. Ecological fish production in two small lowland rivers in Poland. Oecologia,
48:107–11
Penczak, T., E. Suszycka and M. Molinsky, 1982. Production, consumption and energy
transformations by fish populations in a small lowland river. Ekol.Pol., 30(1–2):111–37
Penczak, T., et al., Fish production in the Mesta River, Rila Mountains, Samkov, Bulgaria (in press)
Petr, T., 1968. Population changes in aquatic invertebrates living in two water plants in a tropical
man-made lake. Hydrobiologia, 32:449–85
Petr, T., 1974. A pre-impoundment study limnological study with special reference to the fishes of the
Great Ruaha river (Tanzania) and some of its tributaries (River Yovi and Little Ruaha) in and
around the impoundment area. Uppsala, Institute of Systematic Biology, Cyclostyled report, 104 p.
Petr, T., 1976. Limnology of the Great Ruaha river above Kidetu. In Ecological Studies of the Mtera
basin, edited by D. Johansson. Stockholm, SWECO, pp. 87–99
Petrere, M., 1983. Yield per recruit of the tambaqui Colossoma macropomum Cuvier in the Amazonas
State, Brazil. J.Fish Biol., 22:133–44
Petrovsky C.E. and T.F. Waters, 1975. Annual production by the slimy sculpin population in a small
Minnesota trout stream. Trans.Am.Fish.Soc., 1042:237–44
Petts, G.E., 1984. Impounded rivers: perspectives for ecological management. Chichester, England,
John Wiley and Sons, 326 p.
Phelines, R.F., M. Coke and S.M. Nichol, 1973. Some biological consequencea of the damming of the
Pongolo River. In Proceedings lleme Congres des Grands Barrages. Commission Internationale des
Grands Barrages, pp. 175–90
Philippart, J.C., 1978. Evaluation des ressources piscicoles et halieutiques dans les rivieres du bassin
de la Meuse. Definition d'une methode d'etude. Paper presented at the Colloque international
problematique et gestion des eaux interieueres, Liege, 16–19 May 1978, 11 p. (mimeo)

347
Philippart, J.C., 1978a. Etude des bases ecologique de l'amenagement piscicole de l'Ourthe, Grosse
riviere de la zone a barbeau. Rep. 8. Commission provinciale de Liege du Fonds Piscicole. Liege,
Université de Liege
Pierce, R.S. and H.M. Keller, 1978. Forest land-use impacts on upstream water resources. In Program
on man and the Biosphere (MAB) Project 5 workshop. Land-use impacts on lake and reservoir
ecosystems, May 26–June 2 1978 Warsaw, Poland. Proceedings edited by N. Duncan and J.
Rzoska, Vienna, Facultas
Pillay, S.R. and H. Rosa Jr., 1963. Synopsis of biological data on hilsa, Hilsa ilisha (Hamilton, 1822).
FAO Fish.Biol.Synop., (25):pag. var.
Pinter, K., 1983. Hungary. In World fish farming, cultivation and economics, edited by E.E. Brown,
Westport, Connecticut, AVI Publishing Company, Inc., pp. 297–4
Platts, W.S., 1979. Relationships among stream order, fish populations and aquatic geomorphology in
an Idaho river drainage. Fisheries, 4(2):5–9
Platts, W.S., 1981. Influence of forest and rangeland management on anadromous fish habitat in
western North America. 7. Effects of livestock grazing. Gen.Tech. Rep.U.S.Dep.Agric.For.Serv.,
(PNW-124):124 p.
Poddubnyi, A.G. 1979. The Ichthyofauna of the Volga. In The River Volga and its life, edited by P.D.
Mordukhai-Boltovskoi. The Hague, Dr. W. Junk, pp. 304–39
Poi de Nieff, A., 1981. Mesofauna relacionada a la vegetacion acuatica en una laguna del valle, Alto
Parana, Argentina. Ecosur, 8(16):41–53
Poi de Neiff, A. and J.J. Nieff, 1980. Los camelotes de Eichhornia crassipes en aguas loticas del
Parana y su fauna asociada. Ecosur, 7(16):185–93
Poll, M., 1959. Recherches sur la faune ichtyologique de la region du Stanley Pool. Ann.Mus.R.Congo
Belge (Ser.8 Sci.Zool.), 71:75–174
Poll, M., 1959a. Aspets nouveau de la faune ichtyologique du Congo Belge. Bull.Soc. Zool.Fr.,
84:259–71
Poll, M. and H. Renson, 1948. Les poissons, leur milieu et leur peche au bief superieur du Lualaba.
Bull.Agric.Congo Belge, 39(2):427–46
Power, M.E., 1984. Habitat quality and the distribution of algae-grazing catfish in a Panamanian
stream. J.Anim.Ecol., 53(2):357–74
Powers, D.A., et al., 1979. A comparative study of the oxygen equilibrium of blood from 40 genera of
Amazonian fishes. Comp.Biochem.Physiol.(A Comp.Physiol.), 62:67–85
Preston, F.W., 1962. The canonical distribution of commonness and rarity. Part 1. Ecology,
43(2):185–215
Preston, F.W., 1962a. The canonical distribution of commonness and rarity. Part 2. Ecology,
43(3):410–32
Preston, F.W., 1969. Diversity and stability in the biological world. Brookhaven Symp.Biol., 22:1–12

348
Prowse, G.A. and J.F. Talling, 1958. The seasonal growth and succession of plankton algae in the
White Nile. Limnol.Oceanogr., 3:222–38
Quensiere, J., 1976. Influence de la secheresse sur les pecheries du delta du Chari (1971–1973).
Cah.ORSTOM (Hydrobiol.), 10(1):3–18
Quensiere, J., 1981. De l'interet que présente l'approche synécologique dans les études d'aménagement
halieutique (étude de renouvellement des stocks lacustres tchadiens pr la plaine d'inondation du
nord Cameroun). CIFA Tech.Pap./Doc. Tech.CPCA, (8):173–99
Quiros, R., B.L. Prenski and C. Baigun, 1981. Relacion entre resultados de ensayos de captura y
factores ambientales en al embalse de Salto Grande. Paper presented to the COPESCAL
Symposium on the Development and management of artificial lakes. Santo Domingo, 30 Nov.–1
Dec. 1981 (mimeo)
Rabern, D.A., 1984. Development of habitat based models for predicting standing crops of nine
species of riverine fishes in Georgia. MSc. Thesis. University of Georgia. Athens, Georgia, 126 p.
Rai, H., 1979. Microbiology of central Amazonian lakes. Amazoniana, 6(4):583–99
Rai, H. and G. Hill, 1981. Physical and chemical studies on Lago Tupe, a central Amazonian
blackwater ria lake. Int.Rev. Ges.Hydrobiol., 66(1):37–82
Raimondo, P., 1975. Monograph on operation fisheries, Mopti. In Report of the Consultation on
fisheries problems of the Sahelian zone. Bamako, Mali, 13–20 November 1974. CIFA Occas.Pap.,
(4):294–311. Issued also in French
Rajyalakshmi, T. and T.V. Premswarup, 1975. Primary productivity in River Godavari. Indian
J.Fish., 22(102):205–14
Rao, N.G. and L.H. Rao, 1972. On the biology of Labeo calbasu (Ham. Buch.) from the River
Godavari. J.Inland Fish.Soc.India, 4:26–37
Rao, N.G.S., P. Ray and K. Gopinath, 1972. Observations on the spawning of Cirrhinus reba
(Hamilton) in the Cauvery and Bhavani rivers. J.Inland Fish.Soc.India, 4:69–73
Rasmussen, J.L. (ed.), 1979. A compendium of fishery information on the Upper Mississippi River.
Rock Island, Illinois, Upper Mississippi River Conservation Committee, 259 p.
Reed, W., et al., 1967. Fish and fisheries in Northern Nigeria. Zaria, Ministry of Agriculture, 226 p.
Regier, H.A. and H.F. Henderson, 1973. Towards a broad ecological model of fish communities and
fisheries. Trans.Am.Fish.Soc., 102(1):56–72
Regier, H.A. and K.H. Loftus, 1972. Effects of fisheries exploitation on salmonid communities in
oligotrophic lakes. J.Fish.Res.Board Can., 29(6):959–68
Reiss, F., 1977. Qualitative and quantitative investigations on the macrobenthic fauna of Central
Amazon lakes. I. Lago Tupé, a blackwater lake on the lower Rio Negro. Amazoniana, 6(2):203–35
Reizer, C., 1971. Contribution a l'etude hydrobiologique du Bas-Senegal. Premieres recommandations
d'amenagement halieutique. Nogent-sur-Marne, Centre Technique Forestier Tropical, 142 p.

349
Reizer, C., 1974. Definition d'une politique d'amenagement des resources halieutiques d'un
ecosysteme aquatique complex per l'etude de son environnement abiotique, biotique et
anthropique. Le fleuve Senegal moyen et inferieur. Doctorat en Sciences de l'environnement,
Dissertation, Arlon, Fondat
Reynolds, L.F., 1983. Migration patterns of five fish species in the Murray-Darling River system.
Aust.J.Mar.Freshwat.Res., 34:857–71
Ribeiro, M.C.L., 1983. Ecologia das migracoes das jaraquis no Rio Negro. In X Congrreso Brasiliera
do Zoologia, Resumos, Belo Horizonte, pp. 287–8
Ribeiro, M.C.L., 1983a. As migracoes dos jaraquis (Pisces: Prochilodontidae) no Rio Negro,
Amazonas, Brasil, Dissertation, INPA, Manaus, 192 p.
Richardson, R.E., 1921. The small bottom and shore fauna of the middle and lower Illinois River and
its connecting lakes Chillicote to Grafton: its valuation; its source of food supply; and its relation to
the fishery. Bull.State Ill. Nat.Hist.Surv., 13:363–522
Richey, J.E., 1981. Fluxes of organic matter in rivers relative to the global carbon cycle. In Flux of
organic carbon by rivers to the oceans. United States Department of Energy, Springfield, Virginia,
National Technical Information Services CONF-8009140 UC 11, pp. 270–93
Ringuelet, R.A., R.H. Aramburu and A.A. de Arumburu, 1967. Los peces Argentinos de agua dulce.
Buenos Aires, Librart, 602 p.
Roberts, T.R., 1973. Ecology of fishes in the Amazon and Congo basins. In Tropical forest
ecosystems in Africa and South America: a comparative review, edited by B.J. Meggers and E.S.
Ayers. Washington, D.C., Smithsonian Institution Press, pp. 239–54
Robertson, C.H., 1983. Aspects of the biology of various Macrobrachium spp. found in the Sepik
River. Rep.Dep.Primary Ind.Port Moresby, (83–05):61 p.
Roman, B., 1966. Aportación al estudio de la fauna ictiologica dulceacuicola de Rio Volta. Doctoral
Thesis, Barcelona, Spain
Ruddle, K., 1980. A preliminary survey of fish cultivation with special reference to West Java,
Indonesia. Bull.Natl.Mus.Ethno.,Osaka, 5(3):801–82
Russev, B., 1967. Das zoobenthos der Donau, in Limnologie der Donau, edited by R. Liepolt,
Stuttgart, E. Schweitzerbart'sche Verlagsbuchhandlung, pp. 242–71
Russev, B., 1981. Gegenwartiger stand der untersuchungen uber das zoobenthos im Donau-becken.
Schweitz.Z.Hydrol., 33(1):303–13
Russev, B., 1982. Der einfluss der Stauanlagen auf die entwicklung des zoobnthos im
Osterreichischen Donauabshnitt. In Internationalen Arbeitsgemeinschaft Donauforschung, pp.
139–41
Ryder, R.A., 1978. Fish yield assessment of large lakes and reservoirs-a prelude to management. In
Ecology of freshwater fish production, edited by S.A. Gerking. London, Blackwell Scientific
Publications, pp. 403–23
Rzoska, J., 1974. The Upper Nile swamps, a tropical wetland study. Freshwat.Biol., 4:1–30

350
Rzoska, J. (ed.), 1976. The Nile: biology of an ancient river. The Hague, Junk, Monogr.Biol., (20):417
p.
Rzoska, J. and J.F. Talling, 1966. Plankton development in relation to hydrology and reservoir regime
in the Blue Nile. Verh.Int.Ver.Theor.Angew.Limnol., 16:716–8
Saanin, H., 1953. Notes on the fishery of the cyprinid fish Thynnichthys vaillanti in Indonesia.
Proc.IPFC, 4(2):192–207
Saanin, H., 1969. Toxicity of insecticides and herbicides to some Indonesian cultivatable fish species.
Proceedings of the Fourth Pan Indian Ocean Science Congress, Biological Sciences, pp. 237–303
Sabol, K.J. (ed.), 1974. National Conference on floodplaoin management, 1974. League City, Texas,
National Association of Conservation Districts, 261 p.
Sagua, N.O., 1978. The effect of Kainji Dam, Nigeria, upon the fish production in the River Niger
below the dam at Faku. CIFA Tech.Pap./Doc.Tech.CPCA, (5):209–24
Sanchez Romero, J., 1961. El Paiche. Aspectos de su historia natural, ecologia y aprovechiamiento.
Lima, Servicio de Pesqueria, 48 p.
Sandon, H. and A. Tayib, 1953. The food of some common Nile fish. Sudan Notes Rec., (34):205–23
Sao-Leang and Dom Saveun, 1955. Apercu général sur la migration et la reproduction des poissons
d'eau douce du Cambodge. Proc.IPFC, 5(2–3):138–62
Sayer, J.A. and L.P. van Levieren, 1975. The ecology of the Kafue lechwe population of Zambia
before the operation of hydro-electric dams on the Kafue River. E.Afr.Wildl.J., 13:9–37
Scheimer, F., 1983. Limnology of the Parakrama Samudra - Sri Lanka. A case study of an ancient
man made lake in the tropics. The Hague, Junk, Dev.Hydrobiol., (12):236 p.
Schick, R.A., et al., 1982. Mitigation enhancement techniques for the Upper Mississippi River system
and other larger river systems. U.S. Fish Wildl.Ser.Res.Pub., 149:714 p.
Schlosser, I.J., 1982. Fish communiy structure and function along two habitat gradients in a headwater
stream. Ecol.Monogr., 52(4):395–414
Schmid, M., 1961. Incidences possibles sur la végetation en bordure du Tonle Sap de la construction
d'un barrage concu pour regulariser le regime de Bas-Mekong. Report submitted to Committee for
Investigations of the Lower Mekong Basin, UN/ECAF, 15 p. (mimeo)
Schmidt, G.W., 1969. Vertical distribution of bacteria and algae in a tropical lake.
Int.Rev.Ges.Hydrobiol., 54:791–7
Schmidt, G.W., 1970. Number of bacteria and algae and their interrelation in some Amazonian
waters. Amazoniana, 2:393–400
Schmidt, G.W., 1972. Amounts of suspended solids and dissolved substances in the middle reaches of
the Amazon in the course of one year (August 1969–July 1970). Amazoniana, 3:208–23
Schmidt, G.W., 1972a. Seasonal changes in water chemistry of a tropical lake (Lago do Castanho,
Amazonia, South America). Verh.Int.Ver.Theor.Angew.Limnol., 18:613–21

351
Schmidt, G.W., 1973. Primary production of phytoplankton in a three types of Amazonian waters. 2.
The limnology of a tropical floodplain lake in Central Amazonia, Lago do Castanho, Amazonas,
Brazil. Amazoniana, 4(2):139–203
Schmidt, G.W., 1973a. Primary productivity of phytoplankon in a tropical floodplain lake of central
Amazonia, Lago do Castanho, Amazonas, Brazil. Amazoniana, 4:379–404
Schmidt, G.W., 1976. Primary production of phytoplankton in a three types of Amazonian waters. 4.
The primary productivity of phytoplankton in a bay on the lower rio Negro (Amazonas, Brazil).
Amazoniana, 5(4):517–28
Schneberger, E. and J.L. Funk, 1971. Stream channelization; a symposium. Spec.Publ.N.
Cent.Div.Am.Fish.Soc., (2):83 p.
Schoof, R., 1980. Environmental impact of channel modification. Water Resour.Bull., 16(4):697–701
Schramm, H.L. Jr., et al., 1974. Study of the importance of backwater chutes to a riverine fishery.
Vicksburg, Mississippi, U.S. Army Engineer Waterways Experimental Station, (AD–786544):144
p.
Schwassman, H.O., 1978. Times of annual spawning and reproductive strategies in Amazonian fishes.
In Rhythmic activities of fishes, edited by J.E. Thorpe. London, Academic Press, pp. 187–200
Scott, W.B. and E.J. Crossman, 1973. Freshwater fishes of Canada. Bull.Fish.Res.Board Can.,
(184):966 p.
Scudder, T. and T. Conelly 1984. Management systems for riverine fisheries. California Institute of
Technology, Institute for Development Authropology, 89 p.
Sedell, J.R. and K.J. Luchessa, 1981. Using the historical record as an aid to salmonid habitat
enhancement. Paper presented at the Symposium of Aquatic Inventory Information, Portland,
Oregon (mimeo)
Sedell, J.R. J.E. Yuska and R.W. Speaker, 1983. Study of westside fisheries in Olympic National
Park, Washington. Washington, D.C., USDI, National Park Service, (CX–9000–0–E–081):55 p.
Sedell, J.R., et al., 1974. Sources and fate of organic inputs in coniferous forest streams. In Integrated
research; the coniferous forest Biome, edited by R.H. Waring and R.L. Edmonds. Proceedings of
the AIBS Symposium on coniferous forest ecosystems. Conf.For.Biome Bull., (5):57–69
Sedell, J.R., 1982. Fish habitat and streamside management: past and present. Proc.Soc. Am.For.,
1982
Senegal, Direction des Eaux, Forets et Chasses, 1976. La peche continentale: preparation du Ve plan
du developpement economique et social. Dakar, Direction des Eaux, Fôrets et Chasses, 10 p.
(mimeo)
Shaheen, A.H. and S.F. Youssef, 1979. The effect of the cessation of the Nile flood on the
hydrographic features of Lake Manzalla, Egypt. Arch.Hydrobiol., 85(2):166–91
Shepherd, C.J. (ed.), 1976. Investigation into fish productivity in a shallow freshwater lagoon in
Malawi 1975/76. London, Ministry of Overseas Development, 90 p.

352
Shikhshabekov, 1979. Resorption of the gonads in some semi-diadromous fishes of the Arakum lakes
(Dagestan ASSR) as a result of the regulation of discharge. J.Ichthyol., 11(3):427–30
Shiraishi, Y., 1970. The migration of fishes in the Mekong River. In Proceedings of the IBP Regional
Meeting of Inland Water Biologists in Southeast Asia, Kuala Lumpur and Malacca (Malaysia) 5–
11 May 1969. Sponsored by Unesco. Djakarta, Field Science Office for Southeast Asia, pp. 135–
40
Sidthimunka, A., 1970. A report on the fisheries survey of the Mekong River in the vicinity of the Pa
Mong dam site. Tech.Pap.Dep.Fish.Thail., (8):75 p.
Sidthimunka, A., 1972. Fisheries in relation to impoundment in the Mekong basin experience in
Thailand. In Proceedings of the SEADAG Seminar on the Impact of Development on the Fisheries
of the Lower Mekong River, 3–5 Feb 1972
Silva, S.S. de, and K. Kortmulder, 1977. Some aspects of the biology of three species of Puntius (=
Barbus) (Pisces: Cyprinidae), endemic to Sri Lanka. Neth.J.Zool., 27(27):182–94
Silva, S.S. de, K. Kortmulder and M.J.S. Wijeyratne, 1977. A comparative study of the food and
feeding habits of Puntius bimaculatus and P. titteya (Pisces: Cyprinidae). Neth.J.Zool., 27(3):253–
63
Silva, S.S. de, P.R.T. Cumaranatunga and C.D. de Silva, 1980. Food, feeding ecology and
morphological features associated with feeding of four co-occurring cyprinids (Pisces:
Cyprinidae). Neth.J.Zool., 30(1):54–73
Simpson, B.R.C., 1979. The phenology of annual killifishes. Symp.Zool.Soc.Lond., 44:243–61
Sioli, H., 1964. General features of the Limnology of Amazonia. Verh.Int.Ver.Theor. Angew.Limnol.,
15:1053–8
Sioli, H., 1968. Principal biotypes of primary production in the waters of waters of Amazonia. In
Proceedings of the Symposium on recent advances tropical ecology, edited by R. Misra and B.
Gopal. Varanasi, India, International Society for Tropical Ecology, pp. 591–600
Sioli, H., 1975. Amazon tributaries and drainage basins. Ecol.Stud., 10:199–213
Smart, M., (ed.), 1976. Proceedings of the International Conference on the conservation of wetlands
and waterfowl. Slimbridge, Gloucestershire, U.K., International Waterfowl Research Bureau, 492
p.
Smith, N.J.H., 1981. Man, fishes and the Amazon. New York, Colombia University Press, 180 p.
Smith, P.A., 1976. An outline of the vegetation of the Okavango drainage system. In Proceedings of
the Symposium on the Okavango Delta and its future utilisation. National Museum, Gabarone,
Botswana, 30 Aug–2 Sept. 1976. Gabarone, Botswana Society, pp. 93–112
Soulsby, J.J., 1959. Status of the lake Mweru fishery. Rep.Jt.Fish.Res.Org., 8(1958):309–8
Sparks, R.E. and W.C. Starret, 1975. An electrofishing survey of the Illinois River 1959–1974.
Ill.Nat.Hist.Bull., 31(8):317–80

353
Sreenivasan, A., 1977. Fisheries in the Stanley Reservoir (Mettur Dam) and three other reservoirs in
Tamilnadu, India - a case history. Proceedings of the IPFC Symposium on the development and
utilisation of inland fishery resources, Colombo, 27–29 Oct. 1976. Proc.IPFC, 17:292–302
Sreenivasan, A., 1978. Fish production and fish population changes in some South Indian reservoirs.
Indian J.Fish., 23(1–2):134–52
Sritingsook, C. and T. Yoovetwatana. 1977. Induced spawning of pla yee-sok, Probarbus jullieni
Sauvage. Proc.IPFC, 17(3):404–7
Stalnaker, C.B., 1982. Effects on fisheries of abstractions and purturbations in streamflow. In
Allocation of fishery resources. Proceedings of the Technical Consultation on allocation of fishery
resources, Vichy, France, 20–23 April 1980, edited by J.H. Grover. Published by arrangement with
FAO and the American Fisheries Society, pp. 366–83
Starret, W.C., 1972. Man and the Illinios River. In River ecology and man, edited by R.T. Oglesby,
C.A. Carlson and J.A. McCann. New York, Academic Press, pp. 131–69
Stauch, A., 1966. Le bassin Camerounais de la Benoue et sa peche. Mem.ORSTOM, Paris, (15):152 p.
Stott, B., 1967. The movements and population densities of roach (Rutilus rutilus (L.)) and gudgeon
(Gobio gobio (L.)) in the River Mole. J.Anim.Ecol., 36:407–23
Strahler, A.N., 1957. Quantitative analysis of watershed geomorphology. Trans.Am. Geophys.Union,
38:913–20
Svensson, G.S.O., 1933. Freshwater fishes from the Gambia River (British West Africa). Results of
the Swedish expedition, 1931. K.Sven.Vetenskapsakad Handl., 12:1–102
Swales, S. and K. O'Hara, 1980. Instream habitat improvement devices and their use in freshwater
fisheries management. J.Environ.Manage., 10:167–79
Swanson, F.J., et al., 1982. Land-water interactions: the riparian zone. In Analysis of coniferous forest
ecosystems in the Western United States. US/IBP Synth. Ser., (14)
Swingle, H.S., 1954. Fish populations in Alabama rivers and impoundments. Trans.Am. Fish.Soc.,
83(1953):47–57
Sydenham, D.J.H., 1978. The qualitative composition and logitudinal zonation of the fish fauna of the
River Ogun. Rev.Zool.Afr., 91(4):974–96
Szemes, G., 1967. Das Phytoplankton der Donau. In Limnologie der Donau, edited by R. Liepolt.
Stuttgart, E. Schweitzerbart'sche Verlagsbuchhandlung, pp. 158–79. (English summary)
Tait, C.C., 1967. Hydrological data. Fish.Res.Bull.Zambia, (3):26–8
Tait, C.C., 1967a. Mass fish mortalities. Fish.Res.Bull.Zambia, (3):28–30
Talling, J., 1957. The longitudinal succession of water characteristics in the White Nile.
Hydrobiologia, 9(1):73–89
Talling, J.F. and J. Rzoska, 1967. The development of plankton in relation to hydrological regime in
the Blue Nile. J.Ecol., 55:637–62

354
Talling, J.F. and I.B. Talling, 1965. The chemical composition of african lake waters.
Int.Rev.Ges.Hydrobiol., 50(3):421–63
Tan, E.S.P., 1980. Ecological aspects of some Malaysian riverine cyprinids in relation to their
aquaculture potential. In Tropical ecology and development. Proceedings of the fifth International
Symposium of tropical ecology, 16–21 April 1979, Kuala Lumpur, Malaysia, edited by J.I.
Furtado. Kuala Lumpur, International Society of Tropical Ecology, pp. 757–62
Tang Cheng Eng, et al., 1973. A report on paddy and paddy field fish production in Krian Perak.
Malaysia, Ministry of Agriculture and Fisheries, 57 p.
Tarplee, W.H.Jr., D.E. Louder and A.J. Weber, 1971. Evaluation of the effects of channelization on
fish populations in North Carolina coastal plains streams. North Carolina Wildlife Resources
Commission
Tennant, D.L., 1976. Instream flow regimes for fish, wildlife, recreation and related environmental
resources. Fisheries, 1(4):6–10
Thailand, Department of Fisheries, 1974. Fish farm and pond production for 1973. Bangkok, Ministry
of Agriculture, 64 p.
Thompson, K., 1976. The primary productivity of African wetlands, with particular reference to the
Okavango Delta. In Proceedings of the Symposium on the Okavango Delta and its future
utilisation. Gaberone, Botswana, 30 August2 September 1976. Gaberone, Botswana Society, pp.
67–79
Thompson, K., 1983. The rise and fall of the floodplain. Commonw.Hum.Ecol.Counc.J., 6:13–6
Timmermans, J.A., 1961. La population piscicole de l'Eau Blanche. Petite riviere du type superieur de
la zone a barbeau. Trav.Stn.Rech.Eaux For.Groenendaal-Hoeilaart (D), (30):16 p.
Tolmazin, D., 1979. Black Sea - Dead Sea. New Sci., 84(1184):767–9
Tongsanga, S. and P. Kessunchai, 1966. Fish populations and ecological studies of flood water
fisheries in Pra Nakorn Sri-Ayuthaya Province. Occas.Pap.IPFC, (6):38 p.
Trefethen, P., 1972. Man's impact on the Columbia River. In River ecology and man, edited by R.T.
Oglesby, C.A. Carlson and J.A. McCann. New York, Academic Press, pp. 77–98
Turner, J.L., 1981. Changes in multispecies fisheries when many species are caught at the same time.
CIFA Tech.Pap./Doc.Tech.CPCA, (8):201–11
Tweddle, D., R.E. Hastings and T. Jones, 1978. The development of a floodplain fishery, the Elephant
Marsh, Malawi. CIFA Tech.Pap./Doc.Tech.CPCA, (5):341–56
UN, 1969. Report of the United Nations Interregional Seminar on Flood Damage Prevention
Measures and Management. New York, United Nations, ST/FAO/SER.C/144, 117 p.
Ungemach, H., 1971. Chemical rain water studies in the Amazon region. In II Simposio y foro de
biologia tropical Amazonica, Florencia (Caqueta) y Leticia (Amazonas) Colombia, Enero 1969.
Bogota, Asociacion pro biologia tropical, pp. 354–8

355
Ungemach, H., 1972. Die ionenfracht das rei Negro, Staat Amazonas, Brasilien nach Untersuchungen.
Amazoniana, 3:175–85
University of Idaho, et al., 1971. Ecology of fishes in the Kafue River. Report prepared for FAO/UN
acting as executing agency for UNDP. Moscow, Idaho, University of Idaho. FI:SF/ZAM 11:Tech.
Rep. 2:66 p.
University of Michigan, et al., 1971. The fisheries of the Kafue River Flats, Zambia, in relation to the
Kafue Gorge Dam. Report prepared for the FAO/UN acting as executing agency for the UNDP.
Ann Arbor, Michigan, University of Michigan Press, FI:SF/ZAM 11: Tech. Rep. 1:161 p.
Vaas, K.F., 1953. Fisheries in the lake district along the River Kapuas in west Borneo. Proc.IPFC,
4(2):192–207
Van Leynseele, P., 1979. Ecological stability and intensive fish production: the case of the Libinza
people of the Middle Ngiri (Congo). In Social and ecological systems, edited by P.C. Burnham and
R.F. Ellen. London, Academic Press, Adv.Sci.Assoc.Monogr., 18:167–84
Vannote, R.L., et al., 1980. The river continuum concept. Can.J.Fish.Aquat.Sci., 37(1):130–7
Varela, M.E., et al., 1978. Estudios limnologicos en la cuenca del Riachuelo, V. Biota acuatica de la
alguna la Brava y caracteres del habitat. Ecosur, 5(9):97–118
Vidal, J.C., 1964. Un caso de mortandad de peces en el rio Parana. Buenos Aires, Argentina,
Dirección General de Pesca. 26 p.
Vidal, J.C., 1967. Contribución al estudio biologico del sabalo de los Rios Parana y Uruguay.
(Prochilodus platensis Holmberg). Buenos Aires, Argentina, Direccion General de Pesca y
Conservacion de la Fauna, 51 p.
Vidal, J.C., 1969. Actividades pesqueras en Rosario. Rosario, Argentina, Estación Hidrobiologica de
Rosario, 41 p.
Vidy, G., 1983. Peche traditionelle en bordure du Grand Yaeres nord-camerounias: Le Logomatia.
Rev.Hydrobiol.Trop., 16(4):353–72
Visser, S.A., 1970. The distribution of organic compounds in freshwater lakes and rivers. In Kainji
Lake studies Vol. 1, Ecology, edited by S.A. Visser. Ibadan, Ibadan University Press, pp. 107–25
Visser, S.A., 1974. Composition of waters of lakes and rivers in East and West Africa.
Afr.J.Trop.Hydrobiol.Fish., 3:42–60
Visser, S.A., 1974a. Seasonal changes in the hypophthalmo-hypophyseal-ovarion system in the
catfish, Heteropneustes fossilis (Bloch). J.Fish Biol., 6:331–40
Volker, A., 1966. Surface hydrology of deltaic areas. In Scientific problems of the humid tropical
zone deltas and their implications. Proceedings of the Dacca Symposium, 1964. Paris, Unesco, pp.
153–9
Vranovsky, M., 1974. Zooplankton baciaskeho systemu Ramien pred vyustenim do Llavneho toku a
Jeho Vyznam preformovanie zooplanktonu v Dunaji. Biol.Pr.Slov.Akad. Vied., 7(20):77 p.

356
Vranovsky, M., 1974a. Zur kenntnis der verteilung, biomasse und drift des zoolplanktons im
tchechoslowakisch-ungarischen Donau-abschnitt. Arch.Hydrobiol., Suppl.44 (3):360–3
Ward, J.V. and J.A. Stanford, 1979. The ecology of regulated rivers. New York, Plenum Press, 398 p.
Wassink, E.C., 1975. Photosynthesis and productivity in different environments - conclusions. In
Photosynthesis and productivity in different environments, edited by J.P. Cooper. London,
Cambridge University Press, International Biological Programme 3:675–87
Watson D.J., 1982. Subsistance fish exploitation and implications for the management in the Baram
River system, Sarawak, Malaysia. Fish.Res., 1:299–310
Watson, D.J. and E.K. Balon, 1984. Ecomorphological analysis of fish taxocenes in rainforest streams
of northern Borneo. J.Fish Biol., 25:371–84
Watson, D.J., 1984a. Structure and production of fish communities in tropical rain forest streams of
northern Borneo. Can.J.Zool., 62:627–40
Watson, L., 1981. Sea guide to whales of the world. London, Hutchinson, 302 p.
Weiss, H.W. and D.C. Midgley, 1975. Mathematical floodplain modelling. Vol. 1.
Rep.Univ.Witwatersrand Hydrobiol.Res.Unit, (7/75):pag. var.
Weiss, H.W., 1976. Mathematical floodplain modelling. Vol. 2. Rep.Univ.Witwatersrand
Hydrobiol.Res.Unit, (3/76):pag. var.
Welcomme, R.L., 1964. The habitats and habitat preferences of the young of the Lake Victoria tilapia
(Pisces: Cichlidae). Rev.Zool.Bot.Afr., 40(1–2):1–28
Welcomme, R.L., 1967. The relation between fecundity and fertility in the mouth-brooding cichlid
fish (Tilapia leucosticta). J.Zool.Lond., 151:453–68
Welcomme, R.L., 1969. The biology and ecology of the fishes of a small tropical stream.
J.Zool.Lond., 158:485–529
Welcomme, R.L., 1972. The inland waters of Africa. Les eaux interieures d'Afrique. CIFA
Tech.Pap./Doc.Tech.CPCA, (1):117 p.
Welcomme, R.L., 1975. The fisheries ecology of African floodplains. CIFA Tech.Pap., (3):51 p.
Issued also in French
Welcomme, R.L., 1976. Some general and theoretical considerations on the fish yield of African
rivers. J.Fish Biol., 8:351–64
Welcomme, R.L., 1979. The fisheries ecology of floodplain rivers. Londong, Longman, 317 p.
Welcomme, R.L. and D. Hagborg, 1977. Towards a model of a floodplain fish population and its
fishery. Environ.Biol.Fish., 2(1):7–22
Werner, E.E. and D.J. Hall, 1979. Foraging efficiency and habitat switching in competing sunfishes.
Ecology, 60(2):256–64
Westlake, D.F., 1963. Comparison of plant productivity. Biol.Rev.Camb.Philos.Soc., 38(3):385–425

357
Westlake, D.F., 1975. Primary production of freshwater macrophytes. In Photosynthesis and
productivity in different environments, edited by J.P. Cooper. London, Cambridge University
Press, International Biological Programme 3:186–206
Whitehead, P.J.P., 1958. Indigenous river fishing methods in Kenya. East.Afr.Agric.J., 24:111–6
Whitehead, P.J.P., 1959. The anadromous fishes of Lake Victoria. Rev.Zool.Bot.Afr., 59(3–4):329–63
Whitehead, P.J.P., 1959a. The river fisheries of Kenya. Part 1: Nyanza Province. East.Afr.
Agric.For.J., 24:214
Whitehead, P.J.P., 1960. The river fisheries of Kenya. Part 2: The lower Athi (Sabaki) River.
East.Afr.Agric.For.J., 25(4):259–65
Whiteside, B.G. and R.M. McNatt, 1972. Fish species diversity in relation to stream order and
physicochemical conditions in the Plum Creek drainage basin. Am.Midl.Nat., 88:90–101
Whitley, J.R., 1974. Some aspects of water quality and biology and biology of the Missouri River.
Trans.Miss.Acad.Sci., (7–8):60–72
Williams, N.V., 1960. A review of the Kafue River fishery. Rhodesia Agric.J., 57:86–92
Williams, R., 1971. Fish ecology of the Kafue River and floodplain environment. Fish.Res.Bull.,
Zambia, (5):305–30
Willoughby, N.C. and D. Tweddle, 1977. The ecology of the commercially important species in the
Shire valley fishery, Southern Malawi. CIFA Tech.Pap./Doc.Tech.CPCA, (5):137–52
Wimpenny, R.S., 1934. The fisheries of Egypt. Sci.Prog.Twentieth Centur., Lond., 29(114):210–27
Winstanley, D., 1975. Rainfall and river discharges in the sub-Sahara zone (10°–20°N) of Africa.
CIFA Occas.Pap., (4):97–114 Issued also in French
Wisconsin Department of Natural Resources, 1974. Annual production by brook trout in Lawrence
Creek during eleven successive years. Tech.Bull.Dep.Nat.Resour. Wisc., (82):28 p.
Wishard, S.K., 1976. Roak fishing and its probable effects on the capture fishery of River Yamuna in
Agra district. Indian J.Fish., 23(1–2):213–31
Witte, F. and P.C. Goudswaard 1984. Prospects of the haplochromine fishery in southern Lake
Victoria. Leiden, The Netherlands, Leiden University HEST Report (33):pag. Var.
Wolverton, B. and R.C. McDonald, 1976. Don't waste waterweeds. New Sci., 71:318–20
Wourms, J.P., 1972. The developmental biology of annual fishes. 3. Pre-embryonic diapause of
variable duration in the eggs of annual fishes. J.Exp.Zool., 182:389–414
Wynne-Edwards, V.C., 1962. Animal dispersion in relation to social behaviour. Edinburgh, Oliver
and Boyd, 653 p.
Yon, D. and G. Tendron, 1981. Alluvial forests of Europe. Counc.Eur.Nat.Environ.Ser., (22):65 p.

358
Zambriborsch, F.S. and Nguen Tan Chin, 1973. The descent of fish larvae into the sea through the
Kiliya arm of the Danube. J.Ichthyol., 13(1):90–5
Zarate, M. and A. Cubides. 1977. Estudio ecologico de la orilla de las cienagas del plono inundable
del rio Magdalena. Tesis, Fundacion Universidad de Bogota, Colombia, pag. var.
Zaret, T.M. and A.S. Rand, 1971. Competition in tropical stream fishes: support for the competitive
exclusion principle. Ecology, 52(2):336–42
Zhadin, V.I. and S.V. Gerd, 1970. Fauna and flora of the rivers, lakes and reservoirs of the USSR.
Jerusalem, Israel Program for Scientific Translations, IPST Cat.No.1000:626 p.
View publication statsView publication stats




















