
Where the thoughts dwell: the physiology of neuronal-glial "diffuse neural net

THE JOURNAL OF GENE MEDICINE R E S E A R C H A R T I C L EJ Gene Med 2009; 11: 899–912.Published online 28 July 2009 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jgm.1377
Reversible neurochemical changes mediatedby delayed intrastriatal glial cell line-derivedneurotrophic factor gene delivery in a partialParkinson’s disease rat model
Xin Yang1,2
Birgit Mertens3
Enni Lehtonen1,2
Linda Vercammen4
Olivier Bockstael1,2
Abdelwahed Chtarto1,2
Marc Levivier1,5
Jacques Brotchi1
Yvette Michotte3
Veerle Baekelandt4
Sophie Sarre3
Liliane Tenenbaum*1,2
1Laboratory of ExperimentalNeurosurgery, Universite Libre deBruxelles, Brussels, Belgium2Multidisciplinary Research Institute(IRIBHM), Universite Libre deBruxelles, Brussels, Belgium3Department of PharmaceuticalChemistry, Drug Analysis and DrugInformation (Research GroupExperimental Neuropharmacology),Vrije Universiteit Brussel, Brussels,Belgium 4Laboratory forNeurobiology and Gene Therapy,Katholieke Universiteit Leuven,Leuven, Belgium 5Present address:Department of Neurosurgery, CHUV,Lausanne, Switzerland
*Correspondence to:Liliane Tenenbaum, Laboratory ofExperimentalNeurosurgery/IRIBHM, CP602,ULB-Hopital Erasme, Brussels,Belgium. E-mail: [email protected]
Received: 23 March 2009Revised: 9 June 2009Accepted: 23 June 2009
Abstract
Background Efficient protection of dopaminergic neurons against asubsequent 6-hydroxydopamine lesion by glial cell line-derived neurotrophicfactor (GDNF) gene delivery has been demonstrated. By contrast, theneurorestorative effects of GDNF administered several weeks after the toxinhave been less characterized. In particular, whether these were permanent ordependent on the continuous presence of GDNF remains elusive.
Methods A tetracycline-inducible adeno-associated virus (AAV)-1 vectorexpressing human GDNF cDNA was administered unilaterally in therat striatum 5 weeks after 6-hydroxydopamine. Rats were treated withdoxycycline (dox) or untreated from the day of vector injection until sacrifice(4 or 14 weeks). A sub-group was dox-treated for 7 weeks then untreateduntil 14 weeks. The motor behavior was assessed by amphetamine-inducedrotations and spontaneous forelimb asymmetry. The amounts of tyrosinehydroxylase (TH), serine-40-phosphorylated TH (S40-TH) and aromaticamino acid decarboxylase (AADC) proteins were compared by westernblotting and the dopamine levels quantified by high-performance liquidchromatography.
Results Dox-dependent behavioral improvements were demonstrated4 weeks post-vector injection. At later time points, spontaneous partialrecovery was observed in all rats, but no further improvement was foundin dox-treated animals. TH levels were significantly increased in dox-treatedrats at all time points. By contrast, striatal dopamine and S40-TH wereincreased at 4 weeks, but not 14 weeks, and AADC remained unchanged. Doxwithdrawal after 7 weeks, resulted in TH levels comparable to the controls at14 weeks.
Conclusions Delayed GDNF gene delivery only transiently improveddopaminergic function. Over the long term, TH was more abundant, butnot functional, and the increase was lost when GDNF gene expression wasswitched off. Copyright 2009 John Wiley & Sons, Ltd.
Keywords adeno-associated viral vector; glial cell line-derived neurotrophicfactor; 6-hydroxydopamine; Parkinson’s disease; phosphorylation; tetracycline-inducible; tyrosine hydroxylase
Copyright 2009 John Wiley & Sons, Ltd.

900 X. Yang et al.
Introduction
Glial cell line-derived neurotrophic factor (GDNF) [1] andits structural and functional homolog, Neurturin [2] havebeen proposed for gene therapy of Parkinson’s disease onthe basis of their potent neurotrophic activities towardsdopaminergic neurons [3–5].
GDNF is a member of the transforming growth factor-βsuperfamily [6]. Response to GDNF requires the presenceof glycosylphosphatidyl-inositol-linked proteins (GFR-αs)[7], which form a complex with the membrane receptorRet, thereby inducing receptor tyrosine autophosphory-lation [8] and activation of signal transduction, finallyleading to inhibition of apoptosis [9]. Alternatively, GDNFbound GFR-α1 has also been shown to signal throughthe polysialylated neural cell-adhesion molecule trans-membrane receptor [10]. Consistently with Ret signaling,GDNF gene delivery has been shown to prevent dopamin-ergic cell death induced by various insults, such as medialforebrain bundle axotomy [11], 6-hydroxydopamine(6-OHDA) injection [12] or 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) treatment [13]. However,besides promoting dopaminergic neuron survival, GDNFcan also alter dopaminergic functionality at several lev-els, including enhancement of dopamine (DA) synthesisand metabolism [14–17], transcriptional modulation ofenzymes of DA biosynthesis [18,19], increase of DA avail-ability [20,21] and the excitability of DA neurons [22] andstimulation of tyrosine hydroxylase (TH) phosphorylation[23], thereby augmenting TH activity.
Most animal studies were performed in paradigms inwhich the viral vector encoding GDNF was deliveredprior to establishing a dopaminergic lesion. Efficientprotection of nigral dopaminergic neuron cell bodies andstriatal projections as well as behavioral improvementswere demonstrated when vectors were injected in the ratstriatum prior to a 6-OHDA lesion [24–28] or in MPTP-treated macaques [29,30]. By contrast, midbrain vectorinjections generally only provide protection of cell bodiesand no behavioral improvement [12,26].
However, in the clinical situation, enrolled patientswere diagnosed with moderate to severe Parkinson’sdisease and thus presumably had important lesionswhen the treatment started [31]. Therefore, studiesaddressing the neurorestorative effects of GDNF genedelivery evaluated in paradigms of delayed injectionof viral vectors after lesioning are more relevant[32–36]. In these studies, behavioral improvementshave been demonstrated, although whether these werepermanent or dependent on the continuous presenceof GDNF is not clear. For example, Lapchak et al.[32] demonstrated a reduction of apomorphine-inducedrotational behavior from one to 3 weeks post-injectionof an adenoviral vector expressing GDNF (Ad-GDNF)in the 6-OHDA-lesioned substantia nigra, whereasthe spontaneous locomotor activity was significantlyimproved at 1 week post-injection but then decreased.Furthermore, in their study, Ad-GDNF significantly
increased DA levels in the substantia nigra (SN), but notin the striatum. By contrast, two other articles reportedlong-term correction of apomorphine-induced rotationsand spontaneous asymmetrical behavior when adeno-associated virus (AAV) vectors expressing GDNF wereinjected in the striatum 4 weeks after 6-OHDA lesioning[33,34]. One study using recombinant protein has shownthat GDNF withdrawal results in the loss of the GDNFneurorestorative effect [37]. By contrast, another studyusing capsules implanted for 1 week suggests that theGDNF effects are not reversible [38]. These discrepanciescould be a result of the length of the delay and deliveryperiods, the dose of GDNF, the site and method ofdelivery and the endpoints measured. The evaluationof the effects of GDNF treatment interruption in genetherapy paradigms requires the use of inducible viralvectors that can be efficiently switched off.
Tetracycline-regulated AAV [39] and lentiviral [40,41]vectors for GDNF gene delivery that allow modulationof the biological effects of GDNF have been designed. Inparticular, we have previously developed a tetracycline-inducible autoregulated AAV vector that allowed deliveryof GDNF in the healthy striatum at biologically active con-centrations, leading to a previously described undesiredeffect of GDNF (downregulation of TH) [42] in doxycy-cline (dox)-treated rats, but not in uninduced controls[39].
In the present study, using the autoregulatedtetracycline-inducible AAV vector, we show that delayedGDNF delivery 5 weeks after 6-OHDA intrastriatal injec-tion promoted behavioral recovery at 4 weeks post-viral injection. Consistently, the amount of serine-40-phosphorylated (active) TH as well as of DA and itsmetabolite dihydroxyphenylacetic acid (DOPAC) wereincreased at 4 weeks. At 14 weeks post-viral injection,no significant differences in motor behavior and in stri-atal DA content could be found. However, spontaneousrecovery observed in all groups was possibly maskingfurther GDNF-mediated beneficial effects. Surprisingly,total TH levels were approximately increased by ten-foldat 14 weeks. This increase in TH presumably did notreflect an increased neuron survival because: (i) aromaticamino acid decarboxylase (AADC) levels were unchangedand (ii) arrest of GDNF gene expression by dox removalafter 7 weeks resulted in levels of TH similar to those innontreated animals.
These data might be relevant for the interpretationof the absence of significant therapeutical improve-ments in the Phase II AAV-Neurturin clinical trial(http://www.ceregene.com/press 101006.asp).
Materials and methods
Plasmids and viral vectors
The pAC1-M2-WPRE plasmid comprising AAV ITRs brack-eting the bidirectional tetracycline-responsive cassetteexpressing both rtTAM2 and enhanced green fluorescent
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

GDNF gene delivery in a partial Parkinson’s disease 901
protein (eGFP) has been described previously [39].Recombinant AAV serotype 1 viral stocks were pre-pared from pAC1-M2-WPRE-eGFP and pAC1-M2-WPRE-GDNF by the Laboratoire de Therapie Genique (INSERM,Nantes, France). Titers expressed as viral genomes permillilitre were: rAAV2/1-rtTAM2-WPRE-eGFP (hereafterdesignated as ‘AAV-tetON-GFP’), 1.7 × 1011; rAAV2/1-rtTAM2-WPRE-GDNF (hereafter designated as ‘AAV-tetON-GDNF’): 8.8 × 1011.
Surgical procedures
Adult female Wistar rats, weighing 250 g (CharlesRiver, L’Arbresle, France) were used for unilateral 6-OHDA intrastriatal injections. The animals were anes-thetized using a mixture of ketamine [ketalar, 100 mg/kgintraperitoneally (i.p.)] and xylazine (Rompun, BayerHealth Care, Animal Health, Monheim, Germany;10 mg/kg i.p.). Injections were made according to coor-dinates defined by Paxinos and Watson using a Kopfstereotaxic apparatus (David Kopf, Tujunga, CA, USA).Twenty micrograms of 6-OHDA calculated as free base(Sigma, St Louis, MO, USA) dissolved in 4 µl of ascorbate-saline (0.05%) were injected in the right striatum at twodelivery sites (2 µl per site) at the coordinates: anteriopos-terior (AP) = +1.0 mm, mediolateral (ML) = +3.0 mm,dorsoventral (DV) = −5.5; −4.5 mm, relative to thebregma and the dural surface. After 4 weeks, animalswere tested for amphetamine-induced rotations (5 mg/kgadministered i.p.) and only those animals exhibiting fouror more contralateral rotations/min in a 90-min periodwere included in the experiments. Lesioned animals wererandomly assigned to receive AAV-tetON-GDNF, AAV-tetON-GFP or phosphate-buffered saline (PBS) injection.
Viral particles (3.5 × 108) diluted in 2 µl D-PBS(BioWhittaker, Walkersville, MD, USA) were infusedin the right striatum using a motor-driven Hamiltonsyringe (0.2 µl/min) with a 30-gauge needle at thecoordinates: AP = 0.5 mm, ML = +2.5 mm and DV= −5.5 mm. as previously described [43]. After injection,the needle was left in place for 5 min to allow diffusionof the viral suspension in the parenchyma. The needlewas then slowly lifted 1 mm and left in place for5 min, and then slowly removed. When indicated, theanimals were given water containing 3% sucrose and dox(600 µg/ml) continuously from the day of injection untilsacrifice (group GDNF/Dox+) or until 7 weeks post-vectorinjection (group GDNF/Dox+/−).
Animals were maintained, four in each cage, under a12 : 12 h light/dark cycle with free access to rat chowand water. All experimental procedures were conductedin accordance with the Belgium Biosafety AdvisoryCommittee and with the ethical committee of the Facultyof Medicine (CEBEA, ULB).
Animals were killed at 4 or 14 weeks after vectorinjection by an overdose of anesthetic (200 mg/kgketamine and 20 mg/kg xylazine). The animals weredivided into two groups. For immunohistological analysis,
animals were perfused through the ascending aorta firstwith 150–200 ml of saline (NaCl 0.9%) and then with200 ml of 4% paraformaldehyde in 0.1 M PBS, pH 7.4(PF4). After overnight post-fixation in PF4 at 4 ◦C, brainswere transferred to PBS and stored at 4 ◦C.
For western blotting, enzyme-linked immunosorbentassay (ELISA) and high-performance liquid chromatog-raphy (HPLC), animals were decapitated and the brainswere removed, gradually frozen in isopentane/dry ice(−10 ◦C for 10 s and then −20 ◦C for 20 s) and storedat −80 ◦C. Two hundred-micrometer coronal slices werecut using a cryostat and striata and midbrains were dis-sected out and weighed. Tissue (25 mg/ml) was sonicated(Branson Sonifier 250, output 2, 30% duty cycle for 10 s;Branson Ultrasonics, Danbury, CT, USA) in 0.25 M sucrose,pH 7.4, supplemented with protease inhibitors cocktail asindicated by the supplier (catalog no. 04 693 132 001;Roche Diagnostics, Basel, Switzerland). Samples werecentrifuged at 10 000 g and supernatants were harvestedand their protein concentration analyzed using the BCAAssay kit (Pierce, Rockford, IL, USA).
Behavioral tests
Amphetamine-induced rotation testRotational behavior was assessed in automated rotame-ter bowls (Sandown Scientific, Hampton, UK). After i.pinjection of 5 mg/kg D-amphetamine sulphate (Federa,Brussels, Belgium), right and left full body turns wererecorded over 90 min. Mean net rotations are expressed asfull body turns per 90 min ipsilateral minus contralateralto the lesion.
Cylinder testSpontaneous forelimb use was assessed as describedpreviously [44]. The animals were placed in a glasscylinder (approximately 20 cm in diameter and 35 cmin height) and videotaped as they moved freely in thecylinder and explored the environment. Mirrors wereplaced behind the cylinder to allow observation of theanimals when they were turned away from the camera.Five minutes of activity were recorded for each animal.Weight-bearing forelimb contact with the cylinder wallwas scored. Forelimb asymmetry is expressed as thepercentage of touches using the left (impaired) paw outof the total number of touches. An unbiased animal wouldthus receive a score of 50%, whereas lesions exceeding70–80% of the striatal DA innervation usually reducedthe performance of the impaired paw in this test to lessthan 20% of total wall contacts.
Immunohistochemistry
For GDNF staining, 50 µm vibratome (Leica Microsys-tems, Wetzlar, Germany) coronal sections collected at14 weeks post-vector injection were incubated withpolyclonal goat anti-GDNF antibodies (dilution 1 : 200;
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

902 X. Yang et al.
catalog no. AF-212-NA; R&D Systems, Minneapolis, MN,USA) followed by donkey biotinylated anti-goat secondaryantibody (1 : 250; Amersham, Little Chalfont, UK). ForTH staining, the sections were incubated with mono-clonal anti-TH (dilution 1 : 500; Chemicon, Temecula,CA, USA) followed by biotinylated sheep anti-mousesecondary antibody (dilution 1 : 200; Amersham) asdescribed previously [39].
Streptavidin-peroxidase staining was performed usingthe ABC Elite Vectastain kit and diaminobenzidine(Vector; NTL Laboratories, Brussels, Belgium), accordingto the manufacturer’s protocol. Sections were mountedon gelatin-coated slides, dehydrated and mounted usingEntellan mounting fluid (Merck, Darmstadt, Germany).Sections were photographed using a Zeiss Axiophot 2microscope. Images were acquired as jpeg files usingKS300 software (Zeiss, Gottingen, Germany).
The number of TH-positive neurons in the substantianigra was determined by counting the stained cells ata magnification of 40-fold on one representative 50-µmsection at the level (AP = 5.3 mm) at which the substantianigra is clearly separated from the ventral tegmental areaby the medial terminal nucleus (MTN) of the accessoryoptic tract. This approach has been used previously toensure that comparable rostrocaudal levels of the SN aresampled between animals [45,46].
The lesion extent was expressed as a percentage ofTH-positive cell counts in the lesioned (right) side versusthe contralateral unlesioned side.
In vivo GDNF determination by ELISA
GDNF tissue levels were measured by ELISA test (catalogno. CHC2423; Invitrogen, Carlsbad, CA, USA) accordingto the manufacturer’s protocol (BioSource, Nivelles, Bel-gium) and expressed in pg/mg proteins. Recombinanthuman GDNF (provided by the manufacturer) was usedto establish the standard curve. A cross-reaction of 100%was demonstrated with recombinant rat GDNF (Onco-gene Research Products, San Diego, CA, USA) (data notshown).
Western blots
Equivalent amounts of proteins (as specified in the Resultssection) diluted in Laemmli buffer (Bio-Rad, Hercules, CA,USA) and heated at 95 ◦C for 5 min were electrophoresedon 10% Tris-HCl sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) gels and transferred tonitrocellulose membranes (Hybond N; Amersham).
The amounts of TH, serine 40-phosphorylated TH (S40-TH) and AADC were standardized relative to β-actin.
For TH immunodectection, membranes were incubatedsequentially in (i) Odyssey blocking buffer (LI-COR;Westburg, Leusden, The Netherlands) for 1 h at roomtemperature; (ii) monoclonal mouse anti-TH (dilution1 : 5000; Chemicon) and polyclonal rabbit anti-actin
(dilution 1 : 5000; Sigma) primary antibodies overnightat 4 ◦C;
For S40-TH immunodetection, membranes were incu-bated with a phosphorylation-specific rabbit polyclonalprimary antibody (dilution 1 : 200; Zymed LaboratoriesInc., San Francisco, CA, USA) combined with mouse mon-oclonal anti-β-actin (dilution 1 : 5000; Zymed)
For AADC immunodetection, membranes were incu-bated with polyclonal rabbit anti-AADC (dilution 1 : 200;Chemicon) combined with mouse monoclonal anti-β-actin.
Secondary antibodies were: infrared IRDye 680-conjugated goat anti-mouse antibody (dilution 1 : 10000; LI-COR; red fluorescence) and infrared IRDye 800-conjugated goat anti-rabbit antibodies (dilution 1 : 10000; LI-COR; green fluorescence). Blots were scannedwith the LI-COR Odyssey infrared scanner and signalswere quantified with Odyssey, version 2.0, software.
Biochemical determination of thestriatal DA and DOPAC content
To determine the tissue DA and DOPAC concentration inthe striata, brain extracts at a concentration of 25 mg/mlin 0.25M sucrose supplemented with protease inhibitorswere prepared as described above and analyzed byconventional HPLC, as described previously [47] withslight modifications.
The supernatant obtained from the homogenate wasdiluted ten times in acetic acid (0.5 M) and 20 µl of samplewas injected and analyzed directly on a narrowbore (C18column: 15 µm, 150 × 2.1 mm; Alltima, Grace, Lokeren,Belgium) LC system. The mobile phase consisted of0.1 M sodium acetate trihydrate (Merck), 20 mM citricacid monohydrate (Merck), 1 mM 1-octane sulfonic acid(Sigma), 0.1 mM Na2 ethylenediaminetetraacetic acid(Merck) and 1 mM dibutylamine (Sigma), adjusted to pH3.7. Methanol 3% (v/v) was added as organic modifier.The flow rate was set at 0.3 ml/min. The electrochemicaldetection (Antec, Rotterdam, The Netherlands) potentialwas +700 mV versus the reference electrode (Ag/AgCl).Sensitivity was set at 2 nA full scale. The limit ofdetection of the LC system for DA corresponds to aresidual tissue dopamine content of less than 2.5 ng/gwet tissue.
Statistical analysis
Data are expressed as the mean ± SD. Data analysiswas performed using GraphPad software (GraphPadSoftware Inc., San Diego, CA, USA). One-way or two-way (behavioral tests) analysis of variance (ANOVA)followed by the Newman–Keuls post-hoc test, was usedto compare data obtained from independent samples ofsubjects. p < 0.05 was considered statitically significant(∗p < 0.05; ∗∗p < 0.001).
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

GDNF gene delivery in a partial Parkinson’s disease 903
Results
Time course of the study
To characterize the potential regenerative effect of AAV-tetON-GDNF in a Parkinson’s disease model, a partiallesion of the nigrostriatal pathway was first establishedby injection of 6-OHDA in the right striatum of adultfemale rats (time point: −5 weeks; Figure 1). Lesionedrats were selected on the basis of amphetamine-inducedrotational behavior 4 weeks after toxin injection (timepoint: −1 week; Figure 1). Six groups of animals turningmore than four turns per minute, were constitutedfor vector injections: animals injected with AAV-tetON-GDNF continuously treated with dox until theend of the experiment (GDNF/Dox+ group); animalsinjected with AAV-tetON-GDNF not treated with dox(GDNF/Dox− group); animals injected with AAV-tetON-GDNF treated with dox during 7 weeks post-vectorinjection (GDNF/Dox+/− group); and animals injectedwith AAV-tetON-eGFP (GFP group) (time point: 0 weeks;Figure 1). Behavioral tests were performed at 4, 8, 12 and14 weeks post-vector injection. Animals were sacrificed at4 or 14 weeks post-vector injections (Figure 1).
Regulation of GDNF gene expressionby dox in 6-OHDA-lesioned rats
Immunohistochemistry revealed that, after AAV-tetON-GDNF injection and continuous dox treatment (GDNF/Dox+ group), GDNF was localized in the middle part ofthe striatum with minimal diffusion outside the striatum(Figure 2A). The GDNF concentration in the striatumwas approximately ten- and eight-fold higher in theGDNF/Dox+ group than in the control GFP group, respec-tively, at 4 and 14 weeks (Figure 2G). By contrast, theGDNF/Dox− group had a striatal GDNF concentrationsimilar to the basal endogenous level of the 6-OHDAlesioned striatum (GFP group). Interestingly, arrest of dox
treatment after 7 weeks (GDNF/Dox+/− groups) resultedin GDNF levels similar to the background at 14 weeks(Figure 2G).
GDNF was also found in the ipsilateral substantia nigraof the GDNF/Dox+ (Figure 2D), but not the GDNF/Dox−(E) and GFP groups (F). The pattern of the stainingsuggested that GDNF was present in fibers as wellas in the extracellular space in the substantia nigra(data not shown). However, the GDNF concentrationin the substantia nigra of the GDNF/Dox+ groupwas approximately six-fold lower than in the striatum(Figures 2G and 2H).
Improvement of spontaneous andamphetamine-induced asymmetricalbehavior 4 weeks after injection ofAAV-tetON-GDNF
A significant decrease in amphetamine-induced rotationswas observed in the GDNF/Dox+ and GDNF/Dox+/−groups compared to the GDNF/Dox− and GFP groupsat 4 weeks after gene delivery (n = 8 except forGDNF/Dox+/− groups: n = 5, p < 0.05, ANOVA; Figure 3left panel). (Note that the GDNF/Dox+ and GDNF/Dox+/−groups are equivalent at 4 weeks, because dox treatmentwas arrested at 7 weeks in the GDNF/Dox+/− group.) Atlater time points (i.e. 8–14 weeks post-vector injection),there was no significant difference among the four groups(Figure 3 left panel). However, it should be noted thatthere was a significant decrease of rotations in the GFPgroup at 14 weeks compared to zero time (one-wayANOVA, p < 0.05; Figure 3 left panel), suggesting thata spontaneous (GDNF-independent) recovery, possiblymasking the effect of GDNF, had occurred.
Similarly, a significant improvement was observed inthe cylinder test in the GDNF/Dox+ and GDNF/Dox+/−groups at 4 weeks post-vector injection (Figure 3 rightpanel). Indeed, at 4 weeks post-injection of 6-OHDA, theratio of left versus right touches on the cylinder walls wasapproximately 25 : 75% (data not shown). Relative to this
Figure 1. Time course of surgery and brain analysis. 6-OHDA was injected in the right striatum of adult rats. The presence ofa dopaminergic lesion was evaluated 4 weeks later by assessing behavioral asymmetry. Animals, selected on the basis of theirrotational behavior after amphetamine injection (at least four ipsilateral turns per minute), received rAAV2/1-rtTAM2-WPRE-EGFP,3.4 × 108 viral particles; rAAV2/1-rtTAM2-WPRE-GDNF, 3.5 × 108 viral particles, or PBS, 5 weeks after the 6-OHDA lesion. Dox(600 µg/ml in 3% sucrose) was given in drinking water to the GDNF/Dox+ and GDNF/Dox+/− groups starting at the day of vectorinjection. In the GDNF/Dox+/− group, dox treatment was arrested after 7 weeks of treatment. Amphetamine induced rotation testand cylinder test were performed at −5, −1, 4, 8, 12 and 14 weeks. The animals were sacrificed and brains frozen at 4 and 14 weeksfor biochemical analysis [ELISA test, western blot (WB) and HPLC]. Alternatively, animals were perfused at 4 or 14 weeks and theirbrains processed for histochemical analysis (IHC)
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm
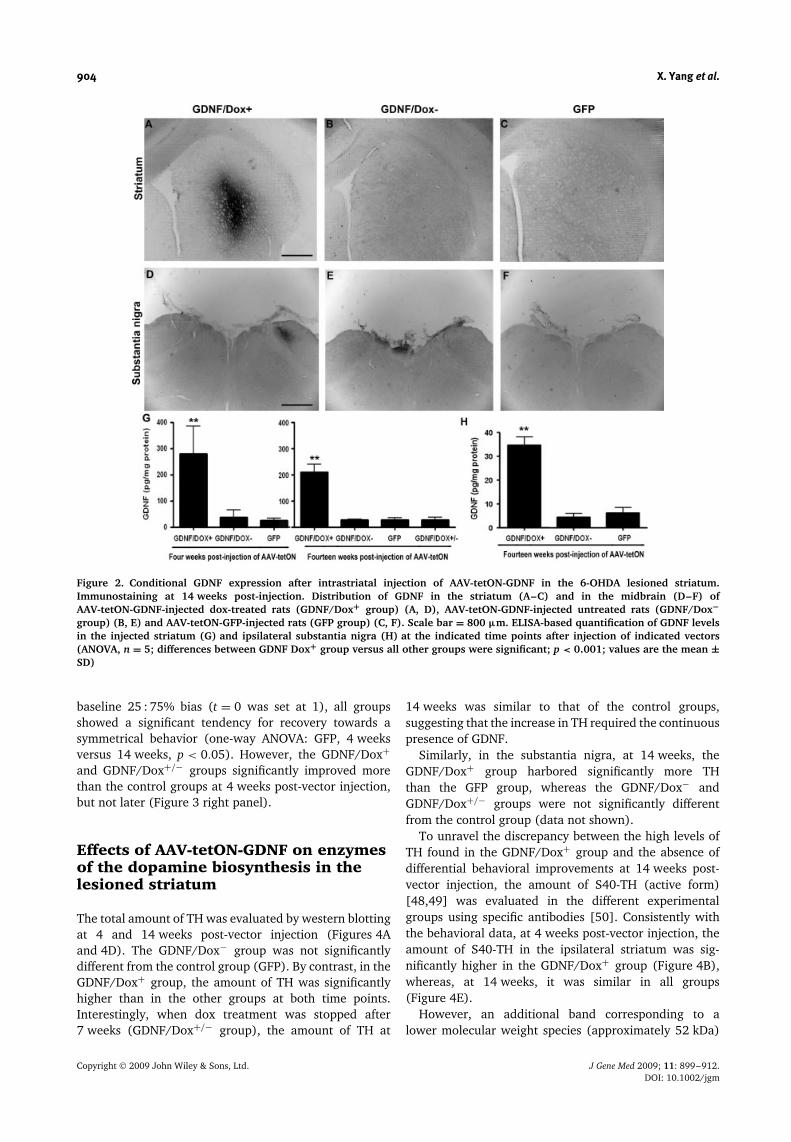
904 X. Yang et al.
Figure 2. Conditional GDNF expression after intrastriatal injection of AAV-tetON-GDNF in the 6-OHDA lesioned striatum.Immunostaining at 14 weeks post-injection. Distribution of GDNF in the striatum (A–C) and in the midbrain (D–F) ofAAV-tetON-GDNF-injected dox-treated rats (GDNF/Dox+ group) (A, D), AAV-tetON-GDNF-injected untreated rats (GDNF/Dox−group) (B, E) and AAV-tetON-GFP-injected rats (GFP group) (C, F). Scale bar = 800 µm. ELISA-based quantification of GDNF levelsin the injected striatum (G) and ipsilateral substantia nigra (H) at the indicated time points after injection of indicated vectors(ANOVA, n = 5; differences between GDNF Dox+ group versus all other groups were significant; p < 0.001; values are the mean ±SD)
baseline 25 : 75% bias (t = 0 was set at 1), all groupsshowed a significant tendency for recovery towards asymmetrical behavior (one-way ANOVA: GFP, 4 weeksversus 14 weeks, p < 0.05). However, the GDNF/Dox+and GDNF/Dox+/− groups significantly improved morethan the control groups at 4 weeks post-vector injection,but not later (Figure 3 right panel).
Effects of AAV-tetON-GDNF on enzymesof the dopamine biosynthesis in thelesioned striatum
The total amount of TH was evaluated by western blottingat 4 and 14 weeks post-vector injection (Figures 4Aand 4D). The GDNF/Dox− group was not significantlydifferent from the control group (GFP). By contrast, in theGDNF/Dox+ group, the amount of TH was significantlyhigher than in the other groups at both time points.Interestingly, when dox treatment was stopped after7 weeks (GDNF/Dox+/− group), the amount of TH at
14 weeks was similar to that of the control groups,suggesting that the increase in TH required the continuouspresence of GDNF.
Similarly, in the substantia nigra, at 14 weeks, theGDNF/Dox+ group harbored significantly more THthan the GFP group, whereas the GDNF/Dox− andGDNF/Dox+/− groups were not significantly differentfrom the control group (data not shown).
To unravel the discrepancy between the high levels ofTH found in the GDNF/Dox+ group and the absence ofdifferential behavioral improvements at 14 weeks post-vector injection, the amount of S40-TH (active form)[48,49] was evaluated in the different experimentalgroups using specific antibodies [50]. Consistently withthe behavioral data, at 4 weeks post-vector injection, theamount of S40-TH in the ipsilateral striatum was sig-nificantly higher in the GDNF/Dox+ group (Figure 4B),whereas, at 14 weeks, it was similar in all groups(Figure 4E).
However, an additional band corresponding to alower molecular weight species (approximately 52 kDa)
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

GDNF gene delivery in a partial Parkinson’s disease 905
Figure 3. Delayed delivery of AAV-tetON-GDNF mediates transient dox-dependent behavioral improvements. (Leftpanel) Amphetamine-induced ipsilateral rotations were first assessed 4 weeks after 6-OHDA. The animals turning more thanfour turns per minute were randomized into the different experimental groups (t = 0; 5 weeks after 6-OHDA injection) and injectedwith AAV-tetON-GDNF (GDNF/Dox+, GDNF/Dox− and GDNF/Dox+/− groups) and AAV-tetON-eGFP (GFP group). The rotationalbehavior was further analyzed at 4, 8, 12 and 14 weeks after vector injection. In the GDNF/Dox+/− group, dox treatment wasarrested at 7 weeks. The values (mean ± SD) indicate the number of ipsilateral turns per 90 min. At 4 weeks, differences betweenGDNF/Dox+ as well as GDNF/Dox+/− groups versus GDNF/Dox− and GFP groups were significant (ANOVA: n = 8 for all groupsexcept GDNF/Dox+/− : n = 5; ∗p < 0.05). (Right panel) Spontaneous motor asymmetry was assessed by the forelimb use asymmetrytest (cylinder test). The bias for left (L) versus right (R) paw touches assessed at 4 weeks after 6-OHDA was found to be 25% : 75%and was set as 1 (baseline value). The animals, randomized on the basis of the amphetamine-induced rotation test, were injectedwith AAV-tetON-GDNF (GDNF/Dox+, GDNF/Dox− and GDNF/Dox+/− groups) or AAV-tetON-eGFP (GFP group). The values of theproportion of left (L) versus total touches (L + R) relative to the baseline are shown at 4, 8, 12 and 14 weeks post-vector injection.At 4 weeks, differences between GDNF/Dox+ as well as GDNF/Dox+/− groups versus GDNF/Dox− and GFP groups were significant(two-way ANOVA: n = 8 for all groups except GDNF/Dox+/−: n = 5; ∗p < 0.05; values are the mean ± SD)
appeared in the GDNF/Dox+ group. This band did notappear in the GDNF/Dox+/− group, suggesting that itwas a consequence of long-term exposure to GDNF orthat it required the continuous presence of GDNF. The52-kDa band could be the result of a proteolytic cleavageof the S40 TH protein, which is known to be less stablethan its unphosphorylated counterpart [51,52].
Complete restoration of functional dopaminergic neu-rons is expected to be accompanied by an increase ofall the enzymes of the dopamine biosynthesis. Therefore,we measured the level of AADC, which converts 3,4-dihydroxy-L-phenylalanine (L-DOPA) to dopamine.
As shown in Figures 4C and 4F, the amount of AADCwas similar in the GDNF/Dox+ group and in the controlgroups.
Levels of dopamine and DOPAC areincreased in AAV-tetON-GDNFdox-treated rats at 4 weeks, but not at14 weeks post-vector injection
Concentrations of DA and DOPAC in striatal extracts weremeasured by HPLC. Consistent with the behavioral andneurochemical data, the concentrations of DA (Figures 5Aand 5B) and DOPAC (Figures 5C and 5D) in the ipsilateral(lesioned) striatum were significantly higher in theGDNF/Dox+ group than in all other groups at 4 weeks,but not at 14 weeks post-vector injection. By contrast,no significant difference between the groups were foundfor the DOPAC : DA ratios at both time points (Figures 5Eand 5F), suggesting that DA metabolism was not modifiedby GDNF.
AAV-tetON-GDNF induces an increasedTH-positive innervation area in thelesioned striatum
Because no functional effect of GDNF was found at14 weeks post-vector injection, we wanted to test whetherthe increased amount of TH in the striatum was the resultof an increased area of TH-positive fibers or increasedamounts of TH in the remaining fibers.
Immunohistochemistry revealed that the 6-OHDAintrastriatal injection resulted in a partial loss of TH-positive fibers in the right striatum more prominent in thelateral and posterior regions in all groups (Figure 6Ato 6C). However, the GDNF/Dox+ group (Figure 6A)harbored a more widespread TH staining compared tothe two other groups.
AAV-tetON-GDNF increases the numberof TH-positive neurons in the lesionedsubstantia nigra
We then aimed to test whether the increased THstaining in the striatum was accompanied by anincreased number of TH-positive neurons in the substantianigra pars compacta. As expected, the number of TH-positive cells in the lesioned (right) substantia nigrawas reduced compared to the contralateral side inall groups (Figures 7A to 7C). However, cell countswere approximately 1.8-fold higher in the GDNF/Dox+(approximately 55% of the contralateral side) than in theGDNF/Dox− and GFP groups (approximately 30% of thecontralateral sides) (Figure 7C).
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

906 X. Yang et al.
Figure 4. Effects of delayed delivery of AAV-tetON-GDNF on enzymes of the dopamine biosynthesis in the lesioned striatum.Animals were sacrificed at 4 weeks (A–C) or 14 weeks (D–F) post-injection of the indicated vector and treated (GDNF/Dox+) or notGDNF/Dox−) with dox during 14 weeks or treated with dox during 7 weeks and then untreated for the next 7 weeks (GDNF Dox+/−).Ten micrograms (A, C, D, F) or 20 µg (B, E) of striatal extracts were loaded on 10% SDS-PAGE gels. After blotting, membranes wereincubated either with (i) monoclonal anti-TH (A, D) combined with polyclonal anti-β-actin antibodies; (ii) polyclonal anti-S40-TH(B, E); or (iii) polyclonal anti-AADC (C, F) combined with monoclonal anti-β-actin antibodies. Secondary antibodies coupled to a redfluorophore (for TH and monoclonal anti-β-actin antibodies) or a green fluorophore (for S40-TH, AADC and polyclonal anti-β-actin)were used to reveal the 60-kDa TH (A, D), 59-kDa S40-TH (B, E), 55-kDa AADC (C, F) and 43-kDa β-actin bands. Amounts of TH,S40-TH and AADC in the striatum were quantified relative to β-actin by dividing the integrated intensities of the red versus thegreen bands (ANOVA: ∗p < 0.05; ∗∗p < 0.001; values are the mean ± SD; 4 weeks: GDNF/Dox+ , n = 4; GDNF/Dox−, n = 6; GFP,n = 6; 14 weeks: GDNF/Dox+, n = 4; GDNF/Dox−, n = 5; GFP, n = 4, Dox+/−, n = 5; ∗, p < 0.05; values are the mean ± SD)
Discussion
GDNF can protect nigrostriatal dopaminergic neuronsagainst further degeneration [12,25,26,28,29], as wellas enhance their function [16,17,19,20,22,23]. In PhaseI open-label clinical trials, GDNF recombinant proteinwas infused into the putamen or into the ventricle ofpatients with Parkinson’s disease. Clinical benefits as well
as increased dopaminergic function, as demonstratedby neuroimaging were reported in the intraputaminalPhase I study [53], but not confirmed in a Phase IIstudy [54]. However, adverse effects were reported inboth human studies [55,56], as well as in rat [57] andmonkey experiments [58]. Therefore, the location, doseand period of GDNF administration should be tightlycontrolled.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

GDNF gene delivery in a partial Parkinson’s disease 907
Figure 5. Delayed delivery of AAV-tetON-GDNF increases striatal DA and its metabolite DOPAC at 4 weeks, but not at 14 weekspost-injection. Determination of DA (A, B) and DOPAC (C, D) levels in striatal extracts by HPLC revealed a dox-dependent increasein the GDNF/Dox+ group at 4 weeks (A, C), but not at 14 weeks (B, D). At 4 weeks, the differences between GDNF/Dox+ groupand the two other groups (GDNF/Dox− and GFP) were significant (ANOVA: 4 weeks: GDNF/Dox+, n = 4; GDNF/Dox−, n = 6; GFP,n = 6; 14 weeks: GDNF/Dox+, n = 4; GDNF/Dox−, n = 5; GFP, n = 4, Dox+/−, n = 5; ∗p < 0.05; values are the mean ± SD). (E, F)Ratio of DOPAC versus DA (DOPAC/DA) at 4 (E) and 14 weeks (F), respectively. No significant difference between the differentgroups was found (ANOVA)
In the present study, an autoregulated tetracycline-inducible AAV serotype 1 vector expressing human GDNFcDNA [39] was delivered to the rat striatum 5 weeksafter intrastriatal injection of 6-OHDA. We have demon-strated that GDNF levels could be tightly controlled bydox administration and removal. Indeed, the amounts ofGDNF were ten-fold higher and similar to the endogenouslevel, respectively, in dox-treated and untreated ani-mals. Furthermore, arrest of dox treatment after 7 weeksresulted in the complete loss of induced GDNF. Accord-ingly, functional (reduction of behavioral asymmetry)and neurochemical effects (increase of amounts of TH) ofGDNF were dox-dependent.
Another important finding of the present study was thatdelayed GDNF gene delivery promoted differential tem-poral effects on the nigro-striatal dopaminergic system.At 4 weeks post-injection of recombinant AAV express-ing GDNF, the total amounts of TH as well as S40-TH
were significantly increased and, consistently, DA lev-els were increased. By contrast, at 14 weeks post-vectorinjection, GDNF significantly increased TH levels, but notS40-TH and DA. By contrast to other studies showing anacceleration of DA metabolism reflected by an increasedDOPAC : DA ratio in the presence of GDNF [15,16], in thepresent study, the DOPAC : DA ratios were unchanged.This discrepancy could be explained by the acute versuscontinuous GDNF delivery method, as well as by the timeof DOPAC : DA ratio measurement after onset of GDNFtreatment.
As a result of the use of the tetracycline-induciblevector, we could show that, when GDNF transgene wasadministered 5 weeks after a partial lesion, interruptionof GDNF expression after 7 weeks resulted in the lossof the GDNF-dependent increase in TH. To our knowl-edge, these data constitute the first demonstration thatdelayed GDNF delivery promotes a reversible effect on
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

908 X. Yang et al.
Figure 6. Effect of AAV-tetON-GDNF on striatal tyrosine hydroxylase fiber staining. TH immunostaining of representative 50-µmstriatal sections taken at three different anteroposterior levels at the indicated coordinates (according to Paxinos and Watson) at14 weeks after vector injection. (A) GDNF/Dox+ group; (B) GDNF/Dox− group; (C) GFP group. Scale bar = 800 µm
TH. Furthermore, it suggests that, under our experimen-tal conditions, delayed GDNF gene delivery did not allowthe rescue of dopaminergic neurons that have not yetdied, but rather that the GDNF effects were mediatedthrough stimulation of remaining neurons and that thiseffect of GDNF was reversible.
The observed increase in TH resulted in statisticallysignificant behavioral improvements in two tests at4 weeks post-vector injection. Spontaneous recovery, aphenomenon already reported previously with single-site intrastriatal 6-OHDA injections [46], was observedin the control groups at 14 weeks (19 weeks after 6-OHDA) compared to time zero (5 weeks after 6-OHDA),masking a potential GDNF-mediated recovery. Similarly,in a previous study using a single intrastriatal 6-OHDA
injection [59], a partial but significant recovery in terms ofdopamine tissue levels as well as a tendency for behavioralimprovements were demonstrated at 8 weeks comparedto 5 weeks post-lesioning. It remains to be determinedwhether, in a more stable lesion model, significantAAV-GDNF-mediated behavioral improvements can bedemonstrated.
An increase in TH transcription and post-transcriptionalstability have been described in neuroblastoma cells [19].Thus, it is possible that the observed increase in TH did notreflect rescue of dying cells, but rather TH transcriptionstimulation in spared dopaminergic neurons. Intrastriatal6-OHDA injections induce a partial and progressive lossof nigro-striatal dopaminergic neurons [60]. It could bethat, at 4 weeks post-vector injection (corresponding to
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

GDNF gene delivery in a partial Parkinson’s disease 909
Figure 7. Effect of AAV-tetON-GDNF on the number of tyrosine hydroxylase-expressing cells in the substantia nigra.Photomicrographs of 50-µm coronal sections through the ventral mesencephalon of a representative GDNF/Dox+ (A), GDNF/Dox−(B) and GFP (C) animal sacrified at 14 weeks post-vector injection and processed for TH immunohistochemistry. Section showsthe levels from the SN proper [only sections in which the medial and lateral parts of the SN–ventral tegmental area were clearlyseparated by the MTN of the accessory optic tract level (AP = 5.3 mm); Paxinos and Watson] were selected for analysis ofTH-immunoreactive (TH-IR) cell numbers (D). This approach ensures that comparable rostrocaudal levels of the SN are sampledbetween animals [45]. Note the relative profound cell loss at levels of SN in the GDNF/Dox− (B) and GFP (C) groups (ANOVA:n = 5; ∗p < 0.05; values are the mean ± SD). Arrows indicate the lesioned side. Scale bar = 800 µm
9 weeks post-toxin injection), some of the degeneratingdopaminergic neurons were still able to be stimulatedto express TH, and contained enough protein kinaseA (PKA) to phosphorylate this additional amount ofTH. At 14 weeks post-vector injection (correspondingto 23 weeks post-toxin injection), it might be the casethat the degenerating neurons had died and that anyadditional TH reflects the transcriptional stimulation ofspared neurons. However, in these spared neurons, theamount of PKA may be limiting and thus not sufficient tophosphorylate more than the normal intracellular level ofTH. It should be noted that, indeed, only less than 5% ofTH is phosphorylated in normal striatum [48].
By contrast to TH, AADC levels were unchanged inGDNF-treated animals at both time points. These datacould be explained by the previously suggested presenceof an additional cell type in the striatum responsiblefor L-DOPA decarboxylation [61,62] in the absence ofdopaminergic neurons. Alternatively, it could be thatGDNF selectively increased TH levels without affectingneuron survival. The reversibility of the increase in
TH after dox withdrawal argues in favor of the latterhypothesis.
Conflicting data exist in the literature concerning theability of delayed intrastriatal delivery of GDNF to rescue6-OHDA-induced partial lesions [33–35,37,38,63,64].These discrepancies could be related to differences in thetime of delivery, the amount and localization of GDNF andthe severity of the lesion. For example, in a study by Kiriket al. [64], behavioral improvements were not maintainedafter the arrest of GDNF protein administration. Bycontrast, in another study by Sajadi et al. [38], behavioralimprovements were maintained after withdrawal ofencapsulated cells secreting GDNF. However, in theformer study, the GDNF administration was started2 weeks after lesioning, whereas, in the latter, it started1 week after lesioning. On the other hand, continuousGDNF secretion by encapsulated genetically modified cellscould be more biologically active than repeated GDNFrecombinant protein produced in Escherichia coli.
By contrast, Wang et al. [33] and Zheng et al. [34]demonstrated both stable behavioral improvements and
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

910 X. Yang et al.
an increase in TH after delivery of AAV-GDNF and Ad-GDNF, respectively, 4 weeks after 6-OHDA. Differentamounts of GDNF could be responsible for the discrepancywith our data. Indeed, the study by Wang et al. [33] useda high-titer of AAV-GDNF: 1.6 × 1013 viral genomes/ml,whereas, in our study, the titer was 1.5 × 1011 viralgenomes/ml. However, to compare these two studies,differences in the AAV serotype and promoter strengthshould also be taken into account.
Adverse effects of GDNF were suggested to be related toinadequate localization. For example, GDNF and/or viralvector transport to the substantia nigra might result inaberrant TH-positive sprouting [57]. In a previous study[65], we demonstrated that, using the AAV1-tetON-GFPvector, the amount of transgene product in the midbrainwas low. This was a result of: (i) inefficient retrogradetransport of viral particles to the substantia nigra parscompacta and (ii) inefficient eGFP expression in thestriatal neurons projecting to the substantia nigra parsreticulata compared to an equivalent vector using thecytomegalovirus promoter. The present data confirm thata relatively low amount of GDNF was transported to thesubstantia nigra after intrastriatal injection of AAV-tetON-GDNF. Indeed, the tissue concentration was six-fold lowerthan in the striatum.
In conclusion, AAV-tetON-GDNF allows the conditionalexpression of GDNF in the striatum of 6-OHDAlesioned rats, resulting in conditional behavioral andneurochemical effects. However, because of a progressivespontaneous recovery of the animals, our experimentaldesign (single intrastriatal 6-OHDA lesion) only allowedassessment of the behavioral effects of GDNF at 4 weekspost-virus injection but not later. Nevertheless, the factthat the main neurochemical effect (i.e. an increase in TH)was reversible suggests that delayed GDNF gene delivery,at least at the dose used (approximately 300 pg/mgprotein corresponding to 50–75 pg/mg tissue) does notallow degenerating dopaminergic neurons to be fullyrescued. Retrograde labeling of nigro-striatal neurons[37] could allow the determination of whether, under ourconditions, GDNF had an effect on atrophic neurons. Thetime delay for the expression of GDNF by the AAV-tetONvector should also be taken into account. Indeed, Sauerand Oertel [60] showed that the degeneration of nigro-striatal neurons was progressive until at least 16 weekspost intrastriatal 6-OHDA delivery.
These data are important in view of the clinical trialsusing AAV2-Neurturin (a structural and functional GDNFhomolog [66]; http://www.ceregene.com/press 101006.asp). Indeed, because patients have extensive DA losswhen they are enrolled, the paradigm of delayedgene delivery is likely to be more relevant to theclinical situation than the neuroprotection paradigm.Furthermore, the reversibility of the neurochemical effectsof GDNF described in the present study could possiblyprovide an explanation for the rapid loss of improvementsexperienced by patients after GDNF capsule removal [67].
Acknowledgements
We thank the vector core of the University Hospital ofNantes supported by the Association Francaise contre lesMyopathies (AFM) for preparing AAV2/1 vectors. PlasmidpAC1: US patent no. 6780 639. We are grateful to Dr TomasGonzalez-Hernandez for critically reading this manuscript.X.Y. was the recipient of a predoctoral fellowship fromAssociation Francaise contre les Myopathies (AFM). B.M. wasthe recipient of a predoctoral fellowship from FWO (Fonds voorWetenschappelijke Onderzoek, Belgium). O.B. was the recipientof a predoctoral fellowship from Fondation Van Buuren. O.B. andE.L. were recipients of a predoctoral fellowship from the FrenchBelgian national research foundation (FNRS-Televie). L.T. wasthe recipient of a ‘Credit aux chercheurs’ (no. 1.5.118.07) fromFNRS. This work was also supported by grants from the FrenchBelgian national research foundation FRSM (no. 3.4510.06), bya grant from the ‘Region Bruxelles-Capitale’ to J.B. and fromAssociation Francaise contre les Myopathies.
References
1. Lin LF, Doherty DH, Lile JD, Bektesh S, Collins F. GDNF: a glialcell line-derived neurotrophic factor for midbrain dopaminergicneurons. Science 1993; 260: 1130–1132.
2. Kotzbauer PT, Lampe PA, Heuckeroth RO, et al. Neurturin, arelative of glial-cell-line-derived neurotrophic factor. Nature1996; 384: 467–470.
3. Bjorklund A, Lindvall O. Parkinson disease gene therapy movestoward the clinic. Nat Med 2000; 6: 1207–1208.
4. Brundin P. GDNF treatment in Parkinson’s disease: time forcontrolled clinical trials? Brain 2002; 125: 2149–2151.
5. Marks WJ Jr, Ostrem JL, Verhagen L, et al. Safety andtolerability of intraputaminal delivery of CERE-120 (adeno-associated virus serotype 2-neurturin) to patients with idiopathicParkinson’s disease: an open-label, phase I trial. Lancet Neurol2008; 7: 400–408.
6. Lin LF, Zhang TJ, Collins F, Armes LG. Purification and initialcharacterization of rat B49 glial cell line-derived neurotrophicfactor. J Neurochem 1994; 63: 758–768.
7. Jing S, Wen D, Yu Y, et al. GDNF-induced activation of the retprotein tyrosine kinase is mediated by GDNFR-alpha, a novelreceptor for GDNF. Cell 1996; 85: 1113–1124.
8. Trupp M, Arenas E, Fainzilber M, et al. Functional receptor forGDNF encoded by the c-ret proto-oncogene. Nature 1996; 381:785–789.
9. Trupp M, Scott R, Whittemore SR, Ibanez CF. Ret-dependentand -independent mechanisms of glial cell line-derivedneurotrophic factor signaling in neuronal cells. J Biol Chem1999; 274: 20885–20894.
10. Paratcha G, Ledda F, Ibanez CF. The neural cell adhesionmolecule NCAM is an alternative signaling receptor for GDNFfamily ligands. Cell 2003; 113: 867–879.
11. Tseng JL, Baetge EE, Zurn AD, Aebischer P. GDNF reduces drug-induced rotational behavior after medial forebrain bundletransection by a mechanism not involving striatal dopamine.J Neurosci 1997; 17: 325–333.
12. Mandel RJ, Spratt SK, Snyder RO, Leff SE. Midbrain injectionof recombinant adeno-associated virus encoding rat glial cellline-derived neurotrophic factor protects nigral neurons in aprogressive 6-hydroxydopamine-induced degeneration model ofParkinson’s disease in rats. Proc Natl Acad Sci USA 1997; 94:14083–14088.
13. Palfi S, Leventhal L, Chu Y, et al. Lentivirally delivered glialcell line-derived neurotrophic factor increases the number ofstriatal dopaminergic neurons in primate models of nigrostriataldegeneration. J Neurosci 2002; 22: 4942–4954.
14. Beck KD, Irwin I, Valverde J, et al. GDNF induces a dystonia-likestate in neonatal rats and stimulates dopamine and serotoninsynthesis. Neuron 1996; 16: 665–673.
15. Hebert MA, Van Horne CG, Hoffer BJ, Gerhardt G A. Functionaleffects of GDNF in normal rat striatum: presynaptic studies using
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

GDNF gene delivery in a partial Parkinson’s disease 911
in vivo electrochemistry and microdialysis. J Pharmacol Exp Ther1996; 279: 1181–1190.
16. Hudson J, Granholm AC, Gerhardt GA, et al. Glial cell line-derived neurotrophic factor augments midbrain dopaminergiccircuits in vivo. Brain Res Bull 1995; 36: 425–432.
17. Martin D, Miller G, Cullen T, et al. Intranigral or intrastriatalinjections of GDNF: effects on monoamine levels and behaviorin rats. Eur J Pharmacol 1996; 317: 247–256.
18. Georgievska B, Kirik D, Rosenblad C, Lundberg C, Bjorklund A.Neuroprotection in the rat Parkinson model by intrastriatalGDNF gene transfer using a lentiviral vector. Neuroreport 2002;13: 75–82.
19. Xiao H, Hirata Y, Isobe K, Kiuchi K. Glial cell line-derivedneurotrophic factor up-regulates the expression of tyrosinehydroxylase gene in human neuroblastoma cell lines.J Neurochem 2002; 82: 801–808.
20. Grondin R, Cass WA, Zhang Z, et al. Glial cell line-derivedneurotrophic factor increases stimulus-evoked dopamine releaseand motor speed in aged rhesus monkeys. J Neurosci 2003; 23:1974–1980.
21. Smith AD, Kozlowski DA, Bohn MC, Zigmond MJ. Effect ofAdGDNF on dopaminergic neurotransmission in the striatumof 6-OHDA-treated rats. Exp Neurol 2005; 193: 420–426.
22. Xu K, Dluzen DE. The effect of GDNF on nigrostriataldopaminergic function in response to a two-pulse K(+)stimulation. Exp Neurol 2000; 166: 450–457.
23. Salvatore MF, Zhang JL, Large DM, et al. Striatal GDNFadministration increases tyrosine hydroxylase phosphorylationin the rat striatum and substantia nigra. J Neurochem 2004; 90:245–254.
24. Bjorklund A, Kirik D, Rosenblad C, et al. Towards aneuroprotective gene therapy for Parkinson’s disease: use ofadenovirus, AAV and lentivirus vectors for gene transfer ofGDNF to the nigrostriatal system in the rat Parkinson model.Brain Res 2000; 886: 82–98.
25. Kirik D, Rosenblad C, Bjorklund A, Mandel RJ. Long-term rAAV-mediated gene transfer of GDNF in the rat Parkinson’s model:intrastriatal but not intranigral transduction promotes functionalregeneration in the lesioned nigrostriatal system. J Neurosci2000; 20: 4686–4700.
26. Choi-Lundberg DL, Lin Q, Chang YN, et al. Dopaminergicneurons protected from degeneration by GDNF gene therapy.Science 1997; 275: 838–841.
27. Choi-Lundberg DL, Lin Q, Schallert T, et al. Behavioral andcellular protection of rat dopaminergic neurons by an adenoviralvector encoding glial cell line-derived neurotrophic factor. ExpNeurol 1998; 154: 261–275.
28. Bilang-Bleuel A, Revah F, Colin P, et al. Intrastriatal injectionof an adenoviral vector expressing glial-cell-line-derivedneurotrophic factor prevents dopaminergic neuron degenerationand behavioral impairment in a rat model of Parkinson disease.Proc Natl Acad Sci USA 1997; 94: 8818–8823.
29. Kordower JH, Emborg ME, Bloch J, et al. Neurodegenerationprevented by lentiviral vector delivery of GDNF in primatemodels of Parkinson’s disease. Science 2000; 290: 767–773.
30. Eslamboli A, Cummings RM, Ridley RM, et al. Recombinantadeno-associated viral vector (rAAV) delivery of GDNF providesprotection against 6-OHDA lesion in the common marmosetmonkey (Callithrix jacchus). Exp Neurol 2003; 184: 536–548.
31. Marks WJ Jr, Ostrem JL, Verhagen L, et al. Safety andtolerability of intraputaminal delivery of CERE-120 (adeno-associated virus serotype 2-neurturin) to patients with idiopathicParkinson’s disease: an open-label, phase I trial. Lancet Neurol2008; 7: 400–408.
32. Lapchak PA, Araujo DM, Hilt DC, Sheng J, Jiao S. Adenoviralvector-mediated GDNF gene therapy in a rodent lesion model oflate stage Parkinson’s disease. Brain Res 1997; 777: 153–160.
33. Wang L, Muramatsu S, Lu Y, et al. Delayed delivery of AAV-GDNF prevents nigral neurodegeneration and promotesfunctional recovery in a rat model of Parkinson’s disease. GeneTher 2002; 9: 381–389.
34. Zheng JS, Tang LL, Zheng SS, et al. Delayed gene therapy ofglial cell line-derived neurotrophic factor is efficacious in a ratmodel of Parkinson’s disease. Brain Res Mol Brain Res 2005;134: 155–161.
35. Kozlowski DA, Connor B, Tillerson JL, Schallert T, Bohn MC.Delivery of a GDNF gene into the substantia nigra after aprogressive 6-OHDA lesion maintains functional nigrostriatalconnections. Exp Neurol 2000; 166: 1–15.
36. McGrath J, Lintz E, Hoffer BJ, et al. Adeno-associated viraldelivery of GDNF promotes recovery of dopaminergic phenotypefollowing a unilateral 6-hydroxydopamine lesion. Cell Transplant2002; 11: 215–227.
37. Lu X, Hagg T. Glial cell line-derived neurotrophic factor preventsdeath, but not reductions in tyrosine hydroxylase, of injurednigrostriatal neurons in adult rats. J Comp Neurol 1997; 388:484–494.
38. Sajadi A, Bensadoun JC, Schneider BL, Lo BC, Aebischer P.Transient striatal delivery of GDNF via encapsulated cells leadsto sustained behavioral improvement in a bilateral model ofParkinson disease. Neurobiol Dis 2006; 22: 119–129.
39. Chtarto A, Yang X, Bockstael O, et al. Controlled delivery of glialcell line-derived neurotrophic factor by a single tetracycline-inducible AAV vector. Exp Neurol 2007; 204: 387–399.
40. Georgievska B, Jakobsson J, Persson E, et al. Regulated deliveryof glial cell line-derived neurotrophic factor into rat striatum,using a tetracycline-dependent lentiviral vector. Hum Gene Ther2004; 15: 934–944.
41. Szulc J, Wiznerowicz M, Sauvain MO, Trono D, Aebischer P. Aversatile tool for conditional gene expression and knockdown.Nat Methods 2006; 3: 109–116.
42. Georgievska B, Kirik D, Bjorklund A. Overexpression of glialcell line-derived neurotrophic factor using a lentiviral vectorinduces time- and dose-dependent downregulation of tyrosinehydroxylase in the intact nigrostriatal dopamine system.J Neurosci 2004; 24: 6437–6445.
43. Tenenbaum L, Jurysta F, Stathopoulos A, et al. Tropism of AAV-2 vectors for neurons of the globus pallidus. Neuroreport 2000;11: 2277–2283.
44. Vercammen L, Van der PA, Vaudano E, et al. Parkin protectsagainst neurotoxicity in the 6-hydroxydopamine rat model forParkinson’s disease. Mol Ther 2006; 14: 716–723.
45. Lee CS, Sauer H, Bjorklund A. Dopaminergic neuronaldegeneration and motor impairments following axon terminallesion by instrastriatal 6-hydroxydopamine in the rat.Neuroscience 1996; 72: 641–653.
46. Kirik D, Rosenblad C, Bjorklund A. Characterization ofbehavioral and neurodegenerative changes following partiallesions of the nigrostriatal dopamine system induced byintrastriatal 6-hydroxydopamine in the rat. Exp Neurol 1998;152: 259–277.
47. Izurieta-Sanchez P, Sarre S, Ebinger G, Michotte Y. Effect oftrihexyphenidyl, a non-selective antimuscarinic drug, ondecarboxylation of L-dopa in hemi-Parkinson rats. Eur JPharmacol 1998; 353: 33–42.
48. Dunkley PR, Bobrovskaya L, Graham ME, von Nagy-Felsobuki EI, Dickson PW. Tyrosine hydroxylase phosphoryla-tion: regulation and consequences. J Neurochem 2004; 91:1025–1043.
49. Bobrovskaya L, Gilligan C, Bolster EK, et al. Sustainedphosphorylation of tyrosine hydroxylase at serine 40: anovel mechanism for maintenance of catecholamine synthesis.J Neurochem 2007; 100: 479–489.
50. Goldstein M, Lee KY, Lew JY, et al. Antibodies to a segment oftyrosine hydroxylase phosphorylated at serine 40. J Neurochem1995; 64: 2281–2287.
51. Martinez A, Haavik J, Flatmark T, Arrondo JL, Muga A.Conformational properties and stability of tyrosine hydroxylasestudied by infrared spectroscopy. Effect of iron/catecholaminebinding and phosphorylation. J Biol Chem 1996; 271:19737–19742.
52. McCulloch RI, Fitzpatrick PF. Limited proteolysis of tyrosinehydroxylase identifies residues 33–50 as conformationallysensitive to phosphorylation state and dopamine binding. ArchBiochem Biophys 1999; 367: 143–145.
53. Gill SS, Patel NK, Hotton GR, et al. Direct brain infusion of glialcell line-derived neurotrophic factor in Parkinson disease. NatMed 2003; 9: 589–595.
54. Lang AE, Gill S, Patel NK, et al. Randomized controlled trialof intraputamenal glial cell line-derived neurotrophic factorinfusion in Parkinson disease. Ann Neurol 2006; 59: 459–466.
55. Nutt JG, Burchiel KJ, Comella CL, et al. Randomized, double-blind trial of glial cell line-derived neurotrophic factor (GDNF)in PD. Neurology 2003; 60: 69–73.
56. Tatarewicz SM, Wei X, Gupta S, et al. Development of amaturing t-cell-mediated immune response in patients withidiopathic Parkinson’s disease receiving r-metHuGDNF via
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm

912 X. Yang et al.
continuous intraputaminal infusion. J Clin Immunol 2007; 27:620–627.
57. Georgievska B, Kirik D, Bjorklund A. Aberrant sprouting anddownregulation of tyrosine hydroxylase in lesioned nigrostriataldopamine neurons induced by long-lasting overexpression ofglial cell line derived neurotrophic factor in the striatum bylentiviral gene transfer. Exp Neurol 2002; 177: 461–474.
58. Hovland DN, Boyd RB, Butt MT, et al. Six-month continuousintraputamenal infusion toxicity study of recombinantmethionyl human glial cell line-derived neurotrophic factor(r-metHuGDNF) in rhesus monkeys. Toxicol Pathol 2007; 35:1013–1029.
59. Yuan H, Sarre S, Ebinger G, Michotte Y. Histological,behavioural and neurochemical evaluation of medial forebrainbundle and striatal 6-OHDA lesions as rat models of Parkinson’sdisease. J Neurosci Methods 2005; 144: 35–45.
60. Sauer H, Oertel WH. Progressive degeneration of nigrostriataldopamine neurons following intrastriatal terminal lesionswith 6-hydroxydopamine: a combined retrograde tracing andimmunocytochemical study in the rat. Neuroscience 1994; 59:401–415.
61. Hefti F, Melamed E, Wurtman RJ. The decarboxylation of DOPAin the parkinsonian brain: in vivo studies on an animal model.J Neural Transm Suppl 1980; 16: 95–101.
62. Sarre S, De KN, Herregodts P, Ebinger G, Michotte Y.Biotransformation of locally applied L-dopa in the corpus
striatum of the hemi-parkinsonian rat studied with microdialysis.Naunyn Schmiedebergs Arch Pharmacol 1994; 350: 15–21.
63. Rosenblad C, Kirik D, Devaux B, et al. Protection andregeneration of nigral dopaminergic neurons by neurturin orGDNF in a partial lesion model of Parkinson’s disease afteradministration into the striatum or the lateral ventricle. Eur JNeurosci 1999; 11: 1554–1566.
64. Kirik D, Georgievska B, Rosenblad C, Bjorklund A. Delayedinfusion of GDNF promotes recovery of motor function in thepartial lesion model of Parkinson’s disease. Eur J Neurosci 2001;13: 1589–1599.
65. Bockstael O, Chtarto A, Wakkinen J, et al. Differential transgeneexpression profiles from rAAV2/1 vectors using the tetON andCMV promoters in the rat brain. Hum Gene Ther 2008; 11:1293–1306.
66. Marks WJ, Jr., Ostrem JL, Verhagen L, et al. Safety andtolerability of intraputaminal delivery of CERE-120 (adeno-associated virus serotype 2-neurturin) to patients with idiopathicParkinson’s disease: an open-label, phase I trial. Lancet Neurol2008; 7: 400–408.
67. Slevin JT, Gash DM, Smith CD, et al. Unilateral intraputamenalglial cell line-derived neurotrophic factor in patients withParkinson disease: response to 1 year of treatment and 1 year ofwithdrawal. J Neurosurg 2007; 106: 614–620.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 899–912.DOI: 10.1002/jgm
Copyright © 2022 FDOKUMEN




















