
Feasibility of bilateral crossing c7 intralaminar screws: a cadaveric study
Upload
independentCategory
view
0download
0
h%-uroscience Vol. 42, No. 1, pp. 73-102, 1991 Printed in Great Britain
0306-4522/91 $3.00 + 0.00 Pergamon Press plc
0 1991 IBRO
RESTRICTED CORTICAL TERMINATION FIELDS OF THE
MIDLINE AND INTRALAMINAR THALAMIC NUCLEI IN THE RAT
H. W. BERENDS? and H. J. GROEMWRGEN
Department of Anatomy and Embryology, Vrije Universiteit, Van der Boecborststraat 7, 1081 BT Amsterdam, The Netherlands
Abstract-The projections from the midline and intralaminar thalamic nuclei to the cerebral cortex were studied in the rat by means of anterograde tracing with Pheolus vulgar&leucoagglutinin.
The midline and intralaminar nuclear complex taken as a whole projects to widespread, predominantly frontal, cortical areas. Each of the constituent thalamic nuclei has a restricted cortical projection field that overlaps only slightly with the projection fields of adjacent midline and intralaminar nuclei. The projections of the intralaminar nuclei cover a larger cortical area than those of the midline nuclei. The laminar distributions of fibres from individual midline and intralaminar thalamic nuclei are different and include both deep and superficial cortical layers.
The parataenial, paraventricular and intermediodorsal midline nuclei each project to circumscribed parts of the prefrontal cortex and the hippocampal and parahippocampal regions. In the prefrontal cortex, the projections are restricted to the medial orbital, infralimbic, ventral prelimbic and agranular insular fields, and the rostral part of the ventral anterior cingular cortex. In contrast to the other midline nuclei, the rhomboid nucleus projects to widespread cortical areas. The rostra1 intral~inar nuclei innervate dorsal parts of the prefrontal cortex, i.e. the dorsal parts of the prelimbic, anterior cingular and dorsal agranular insular cortical fields, the lateral and ventrolateral orbital areas, and the caudal part of the ventral anterior cingular cortex. Additional projections are aimed at the agranular fields of the motor cortex and the caudal part of the parietal cortex. The lateral part of the parafascicular nucleus sends fibres predominantly to the lateral agranular field of the motor cortex and the rostral part of the parietal cortex. The medial part of the parafascicular nucleus projects rather sparsely to the dorsal part of the prelimbic cortex, the anterior cingular cortex and the medial agranular field of the motor cortex.
Individual midline and intralaminar thalamic nuclei are thus in a position to directly influence circumscribed areas of the cerebral cortex. In combination with previously reported data on the organization of the midline and intralaminar thalamostriatal projections and the prefrontal corticostriatal projections the present results suggest a high degree of differentiation in the convergence of thalamic and cortical afferent fibres in the striatum. Each of the recently described parallel basal ganglia-thalamocortical circuits can thus be expanded to include projections at both the cortical and striatal levels from a specific part of the midline and in~~aminar nuclear complex. The distinctive laminar dist~butions of the fibres originating from the different nuclei emphasize the specificity of the midline and intralaminar thalamocortical projections.
The midline and intralaminar thalamic nuclei con- stitute a conspicuous nuclear complex in the dorsal thalamus that receives afferents from a wide variety of sources25~26,8’ (for review see Ref. 88). The main targets of this complex include the cerebral cortex, the striatum, the reticular thalamic nucleus, the amygdala and the subthalamic nucteusU L273&7VW ~1,1w27.141 (for
review see Ref. 77). The ~lationship of the midline and intralaminar nuclei with the cerebral cortex is often characterized as non-specific, because of the widespread distribution of their projections over the cortex.69,80,‘27 Furthermore, the mode of termination in the cortex of the fibres originating from these nuclei is reminiscent of the laminar dis~bution of the “unspecific fibres” of Lorente de No.67*69,86 These anatomical data concur with the physiological
*To whom correspondence should be addressed. Abbreviations: CAI, field CA1 of Ammon’s horn; PHA-L,
Phaseolus u~lgaris-leucoagglutinin.
concept of a non-specific corticopetal system, parallel to the cortical projections from the specific or relay nuclei of the thalamus.6473*74,98 The concept is based on the results of experiments in which cortical recruit- ing and arousal reactions were elicited by electrical stimulation of several thalamic areas, including the midline and intralaminar nuclei.3’,73~74*97 The morph- ology of the neurons within these nuclei, observed in studies using the Golgi method,13’ has strengthened the above-mentioned assumption of non-specificity. However, an increasing body of evidence indicates a higher degree of specificity in the afferent and efferent connections of the midline and intralaminar nuclear complex than was assumed previously. The results of tracing studies have revealed that, in addition to their shared afferents, individual midline and intralaminar nuclei receive specific sets of inputs2S,26 (see references in Refs 11 and 88). Retrograde tracing studies have further shown that the midline and intralaminar projections to separate cortical areas come from
73

14 H. W. BERENDSE and H. J. GROENEWEGEN
different populations of neurons that are either inter- mingled or spatially segregated.9~‘5~78~89’90’1” Finally,
physiological and behavioural studies have provided evidence for the involvement of various nuclei in specific processes. 4,16,30,70,93,108.113,133135
The projections of the midline and intralaminar nuclei to the striatum constitute one of the two major striatal inputs8~‘1~75 and may consist of collaterals given off by the thalamocortical fibres. 22.23.76.'%124,139 The ot,,er
major input is derived from the entire cortical mantle and is arranged in such a way that functionally different cortical areas project to separate striatal regions.82~94,‘3*~‘5’,‘57 This suggests that these striatal sectors are functionally different, a notion substan- tiated by the results of physiological and behavioural studies, 18.32.39~42,103,109~110,121,122 In the monkey, Alexander
et al.’ have distinguished several parallel circuits lead- ing through the basal ganglia, each including distinct parts of the cerebral cortex, the striatum, the pallidum and the thalamus. In the rat, similarly organized basal ganglia-thalamocortical circuits may exist,5s but the precise organization of these circuits awaits further
elucidation.
prefrontal corticostriatal connections.12 In addition, the laminar distribution of the midline and intra-
laminar thalamic afferent fibres in the cerebral cortex will be described and, for the prefrontal cortex, related to the laminar origin of the patch- versus matrix- directed corticostriatal projections.47 To these aims, the anterograde tracer Phase&r vulgaris-leucoagglutinin (PHA-L), first introduced by Gerfen and Sawchenko,49 was used since it allows the placement of very small deposits of tracer and, furthermore, permits a differentiation between passing and terminating fibres.
EXPERIMENTAL PROCEDURES
Anierograde tracing
In line with the parallel arrangement of forebrain circuits involving cortex, thalamus and basal ganglia, we have demonstrated previously that inputs from the midline and intralaminar nuclei converge with functionally related cortical inputs in three distinct striatal sectors.” One of the objectives of the present study was to determine the degree of specificity in the projections of these thalamic nuclei to the cerebral cortex. For the prefrontal cortex, the organization
of these projections is compared with the parallel arrangement of the thalamostriatal” and that of the
Injections of 2.5% PHA-L49 (Vector Laboratories, U.S.A.) were placed stereotaxically in the thalamus of 38 female Wistar-rats (180-220 g), foilowing anaesthesia with a mixture (4: 3) of ketamine and 2-(2.6-xvlidine)5.6-dihvdro- 1,3-thiazine (dompun), in a dosage of -1 ml/kg. The tracer was applied iontophoretically through glass micropipettes (external tip diameter 15525 pm) using a positive-pulsed 7.5pA current (7 s on/7 s 0% CCS-3 current source, Midgard, U.S.A.) for 15-30min. The coordinates for the injections were derived from the atlas of Paxinos and Watson.‘o7 Survival times of the animals ranged between 6 and 20 days, after which the animals were reanaesthetized with pentobarbital and perfused transcardially with 100 ml saline, followed by 750ml of a fixative containing 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) either with or without 0.05% glutaraldehyde. Following a postfix- ation in the same fixative for 1-2 h, the brains were stored overnight at 4°C in 30% sucrose in 0. I M phosphate buffer (pH 7.4) or in a mixture of 20% glycerol and 2% dimethyl- sulphoxide in distilled water. Subsequently, sections were cut at 40 nm on a freezing microtome and collected in 0.1 M phosphate buffer (pH 7.4).
Abbreviations used in the figures
MO mt oc Par I Par II
ACd ACv AGm AGI AId AIP AIv AM cc cg CL CM CPU Ent FL fr G hbc Hif HL IL IMD LHb LO LV MD MHb
anterior cingular cortex, dorsal part anterior cingular cortex, ventral part agranular field of motor cortex, medial part agranular field of motor cortex, lateral part agranular insular cortex, dorsal part agranular insular cortex, posterior part agranular insular cortex, ventral part anteromedial thalamic nucleus corpus callosum cingulum central lateral thalamic nucleus central medial thalamic nucleus caudate-putamen entorhinal cortex forelimb area fasciculus retroflexus gelatinosus thalamic nucleus habenular commissure hippocampal fissure hindlimb area infralimbic cortex intermediodorsal thalamic nucleus lateral habenular nucleus lateral orbital cortex lateral ventricle mediodorsal thalamic nucleus medial habenular nucleus
PC PF Pir PL PrC PRh PT PV PVA PVP Re Rh RSA RSG S
;r Tu VLO vo 3v
medial orbital cortex mammillothalamic tract occipital cortex parietal area I parietal area II paracentral thalamic nucleus parafascicular thalamic nucleus piriform cortex prelimbic cortex precommissural nucleus perirhinal cortex parataenial thalamic nucleus paraventricular thalamic nucleus paraventricular thalamic nucleus, anterior part paraventricular thalamic nucleus, posterior part nucleus reuniens thalami rhomboid thalamic nucleus agranular retrosplenial cortex granular retrosplenial cortex subiculum stria medullaris temporal cortex olfactory tubercle ventrolateral orbital cortex ventral orbital cortex third ventricle
Thalamocortical projections 15
Every third section was incubated in rabbit anti-PHA-L antiserum (1:2000; E-Y Labs, U.S.A. or Dakopatts, Denmark) for 18 hat 4”C, following three pre-rinses in 0.05 M Tris-buffered saline (pH 7.6) with 0.5% Triton X-100. Thereafter, sections were incubated in swine anti-rabbit whole serum (1: SO; Nordic Immunology, The Netherlands) and in rabbit ~ro~da~-ant~~roxidase (1: 800, Dakopatts, Denmark), successively, each for 1 h at 20°C. All incubations and intervening rinses were in 0.05 M T&-buffered saline (pH 7.6) with 65% Triton X-100. Finally, the sections were treated with diaminobenxidine (50 mg in 100 ml Tris-HCl. pH 7.6), preceded and followed ‘by th&e rinses in Tris-HCi (pH 7.6), and mounted from 0.2% gelatin in Tris-HCI (pH 7.6). Half the PHA-L-stained sections were counterstained with Cresyl Violet before coverslipping; the other sections
were coverslipped unstained. The distribution of PHA-L- labelled fibres and terminals was charted with the aid of a drawing tube on a standard Zeiss microscope.
RESULTS
For the dehneation of the various midline and intraiaminar thalamic nuclei, we refer to a previous study in which the striatal projections of these nuclei were described.” In the present study we adhere to the same nomenclature, adopted from Paxinos and Watson.“’
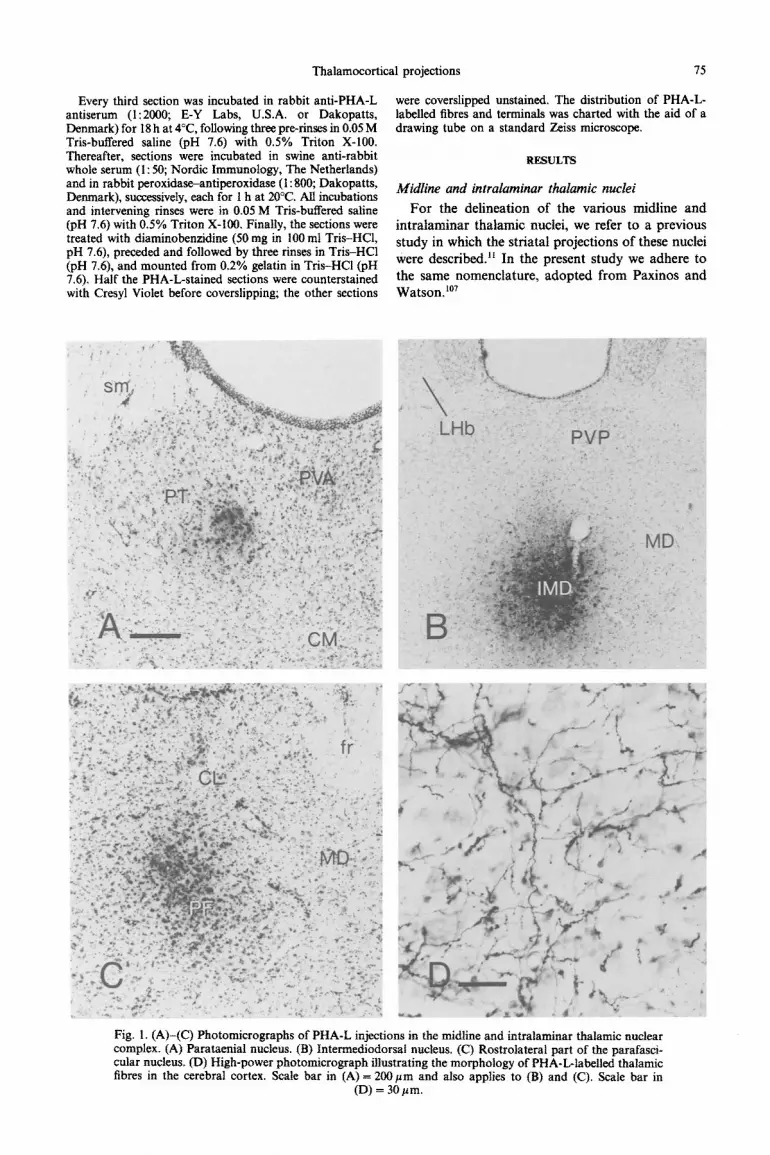
Fig. 1. (A)-(C) Photomicrographs of PHA-L injections in the midline and intralaminar thalamic nuclear complex. (A) Parataenial nucleus. (B) Inte~ediodorsal nucleus. (C) Rostrolateral part of the parafasci- cular nucleus. (D) High-power photomicro~aph illus~ating the mo~hology of PHA-L-la~ll~ thalamic fibres in the cerebral cortex. Scale bar in (A) = 2OOpm and also applies to (B) and (C). Scale bar in
(D) = 30 pm.

76 H.W. BEXENDSE and H.J. GROENEWEQEN
Prefrontal and other cortical areas
The parcellation of the prefrontal cortex largely follows the accounts of Krettek and Pricee4 and Van Eden and Uylings145 (see also Refs 55 and 114). According to these authors, the prefrontal cortex can be subdivided into medial and lateral portions. The medial prefrontal cortex includes the medial orbital, infralimbic, prelimbic, anterior cingular and medial precentral co&ices. The lateral prefrontal cortex encompasses the remaining orbital cortical fields and the agranular insular cortices. For the frontoparietal, retrosplenial and perirhinal cortical regions, the descriptions of Donoghue and Wise,37 Zilles and Wreersg and Reep et al. ‘I6 have been adopted. In these accounts, medial and lateral agranular fields are distinguished in the frontal cortex that correspond cytoarchitectonically to the medial and lateral pre- central cortices of Krettek and Price,@ Van Eden and Uylings145 and Groenewegen. s5 In the present report, we will employ the terms medial and lateral agranular cortex. The first and secondary somatic sensory cortices are indicated as parietal area I and parietal area II, respectively; forelimb and hindlimb areas are identified as separate fields.37~“6,‘59 The temporal and occipital cortices will not be divided into subfields, since anterograde label was sparse in these areas following PHA-L injections in the midline and intra- laminar nuclei.
Midline t~a~arno~ort~cal projections
PI-IA-L was injected in the paravent~cular, parataenial, inte~~iodo~al and rhom~id nuclei, Except for the injections in the parataenial nucleus, all PHA-L deposits involve the nuclei bilaterally. Examples of the injections are given in Fig. 1A and B. The diameter of the injection sites varies from 150 to 600 pm. With the exception of the fibres originating in the rhomboid nucleus (see corresponding paragraph), PHA-L-labelled fibres from all other nuclei terminate bilaterally in circumscribed areas of the prefrontal cortex and the hippocampal and/or parahippocampal cortices. The labelling is most dense ipsilaterally. The contralateral projection of the parataenial nucleus is negligible.
Most labelled fibres reach their cortical target via the inferior thalamic peduncle, the substantia innominata, the pallidum and the striatum. In the striatum, the thalamic fibres follow the fibre bundles of the internal capsule. A minority of fibres enter the striatum via the stria terminalis. It could not be determined whether the thalamocortical fibres give off collateral branches that terminate in the striatum. The majority of fibres destined for the posterior parts of the insular cortex, the perirhinal cortex and the entorhinal cortex follow a ventral route through the substantia innominata and the ~ygdaloid complex.
In the various cortical areas, the fibres o~~nating from the midline thalamic nuclei are not restricted to a single cortical layer, but rather distribute over two
or more layers. More importantly, the laminar distri- butions of the fibres from individual midline thalamic nuclei are different. Passing fibres, that can easily be distinguished from terminating fibres on the basis of their smooth appearance, are present in large numbers in the deeper cortical layers, coursing towards neigh- bouring cortical areas or more superficial cortical layers. In the layers that are most densely labelled, the fibres have a high density of varicosities, branch frequently and are oriented more or less parallel to the pial surface (Fig. ID). The fibres that terminate in sparsely labelled layers bear relatively few varicosities and follow a rather straight course (see for example Fig. 4B).
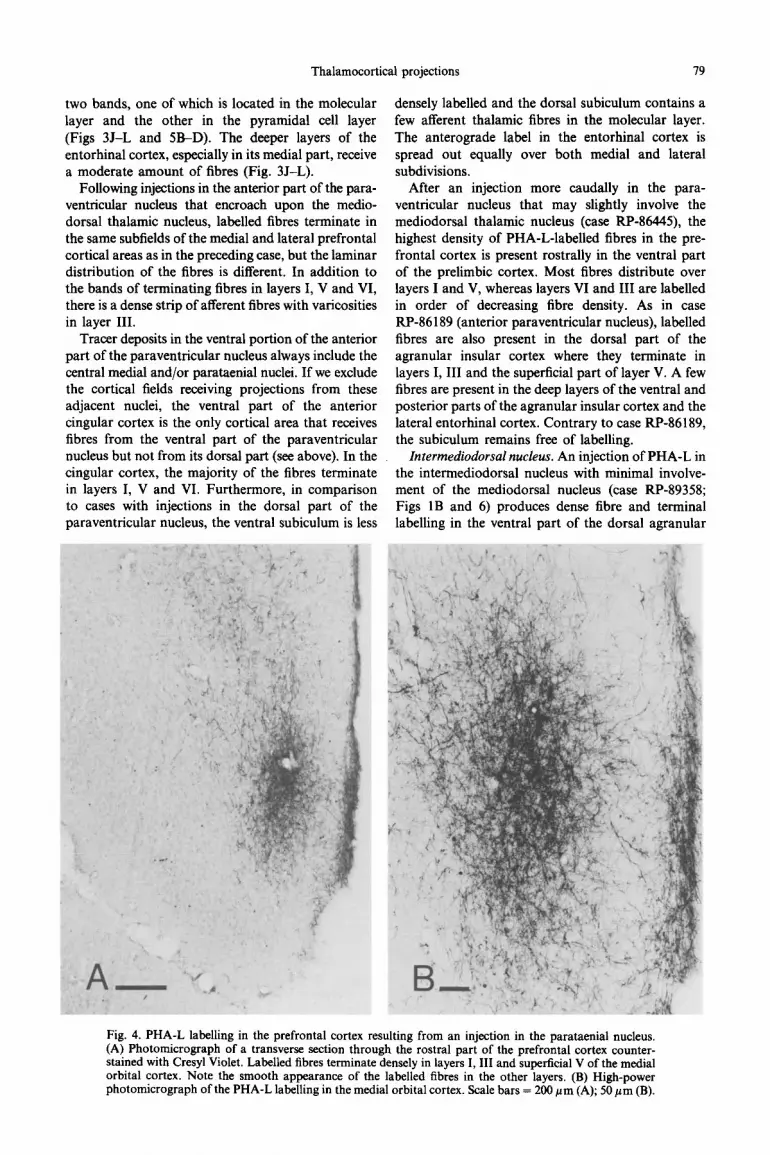
Parataenial nucleus. In case RP-88050, a very small injection of PHA-L was placed in the parataenial thalamic nucleus (Figs IA and 2). The resulting anterograde labelling in the medial prefrontal cortex pr~ominantly involves the medial orbital and infra- limbic cortical areas (Figs 2A-C and 4). Sparse fibres terminate in the ventral part of the prelimbic cortex (Fig. 2B, C). The terminal plexus in the medial orbital cortex is concentrated in its dorsal part and involves layers I and III and, to a lesser extent, superficial layer V (Figs 2A and 4). The plexus is continuous with that in the rostra1 part of the infralimbic cortex which equally involves layers I and III and, though sparsely, layer V (Fig. 28, C). At caudal levels of the infralimbic cortex, the labelling is less dense. A few frbres from the parataenial nucleus terminate in the ventral agranular insular (Fig. 2C, D) and the perirhinal cortical fields (Fig. 21, J).
A sparse projection from the parataenial nucleus is directed to the entorhinal cortex and the ventral subiculum (Fig. 2J-L). The fibres in the entorhinal cortex are located predominantly in layers V and VI of the lateral subdivision; those in the ventral subiculum are confined to the molecular layer.
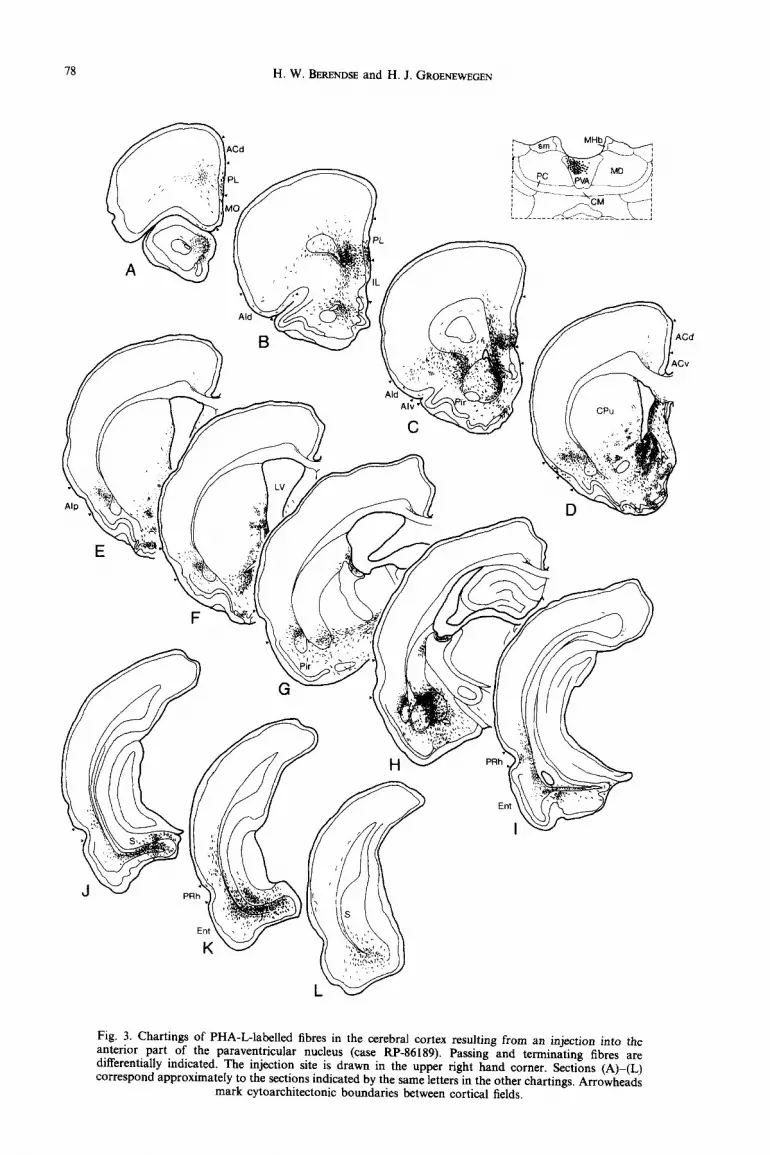
Paraoentricular nucleus. PHA-L was injected at various rostrocaudal levels of the paraventricular nucleus. The deposit in case RP-86189 is located in the anterior part of the paraventricular nucleus and involves the dorsal portion of the nucleus only (Fig. 3). In the ventral part of the prelimbic cortex, antero- gradely labelled fibres form a terminal plexus that is most dense caudally. The fibres terminate predomi- nantly in layers I, V and VI (Figs 3A-C and SA). At the most caudal levels of the prelimbic cortex, the label in layer VI extends into the dorsal part of this cortical field (Fig. 3C). The agranular insular cortex receives paraventricular fibres as well. Rostrally, the fibres distribute over layers I, V and VI of the dorsal and, to a lesser extent, the ventral subfield (Fig. 3B-D). In the posterior agranular insular cortex, sparse thalamic flbres terminate predominantly in layer VI (Fig. 3E, F). The perirhinal cortex contains a moderate amount of anterogradely labelled fibres in layers V and VI and sparse fibres in layer III (Fig. 31-K). More dense anterograde label is present in the ventral subiculum, where the labelled fibres are organized in

Thalamocortical projections
Fig. 2. Chartings of PHA-L-labelled fibres in the cerebral cortex following an injection into the parataenial nucleus (case RP-88050). Passing and terminating fibres are differentially indicated, The injection site is drawn in the upper right hand comer and is shown photographically in Fig. 1A. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings. Arrowheads
mark cytoarchitectonic boundaries between cortical fields.
78 H. W. BERENDSE and H. J. GROENEWEGEN
Fig. 3. Chartings of PHA-L-labelled fibres in the cerebral cortex resulting from an injection into the anterior part of the paraventricular nucleus (case RP-86189). Passing and terminating fibres are differentially indicated. The injection site is drawn in the upper right hand corner. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings. Arrowheads
mark cytoarchitectonic boundaries between cortical fields.
Thalamocortical projections 79
two bands, one of which is located in the molecular densely labelled and the dorsal subiculum contains a layer and the other in the pyramidal cell layer few afferent thalamic fibres in the molecular layer. (Figs 3J-L and 5B-D). The deeper layers of the The anterograde label in the entorhinal cortex is entorhinal cortex, especially in its medial part, receive spread out equally over both medial and lateral a moderate amount of fibres (Fig. 3J-L). subdivisions.
Following injections in the anterior part of the para- ventricular nucleus that encroach upon the medio- dorsal thalamic nucleus, labelled fibres terminate in the same subfields of the medial and lateral prefrontal cortical areas as in the preceding case, but the laminar distribution of the fibres is different. In addition to the bands of terminating fibres in layers I, V and VI, there is a dense strip of afferent fibres with varicosities in layer III.
Tracer deposits in the ventral portion of the anterior part of the paraventricular nucleus always include the central medial and/or parataenial nuclei. If we exclude the cortical fields receiving projections from these adjacent nuclei, the ventral part of the anterior cingular cortex is the only cortical area that receives fibres from the ventral part of the paraventricular nucleus but not from its dorsal part (see above). In the cingular cortex, the majority of the fibres terminate in layers I, V and VI. Furthermore, in comparison to cases with injections in the dorsal part of the paraventricular nucleus, the ventral subiculum is less
After an injection more caudally in the para- ventricular nucleus that may slightly involve the mediodorsal thalamic nucleus (case. RP-86445) the highest density of PHA-L-labelled fibres in the pre- frontal cortex is present rostrally in the ventral part of the prelimbic cortex. Most fibres distribute over layers I and V, whereas layers VI and III are labelled in order of decreasing fibre density. As in case RP-86 189 (anterior paraventricular nucleus), labelled fibres are also present in the dorsal part of the agranular insular cortex where they terminate in layers I, III and the superficial part of layer V. A few fibres are present in the deep layers of the ventral and posterior parts of the agranular insular cortex and the lateral entorhinal cortex. Contrary to case RP-86189, the subiculum remains free of labelling.
Intermediodorsal nucleus. An injection of PHA-L in the intermediodorsal nucleus with minimal involve- ment of the mediodorsal nucleus (case RP-89358; Figs 1B and 6) produces dense fibre and terminal labelling in the ventral part of the dorsal agranular
Fig. 4. PHA-L labelling in the prefrontal cortex resulting from an injection in the parataenial nucleus. (A) Photomicrograph of a transverse section through the rostra1 part of the prefrontal cortex counter- stained with Cresyl Violet. Labelled fibres terminate denselv in lavers I. III and sunerticial V of the medial orbital cortex. Note the smooth appearance of the labelied fibres in the other layers. (B) High-power photomicrograph of the PHA-L labelling in the medial orbital cortex. Scale bars = 200 pm (A); 50 pm (B).

80 H. W. BERENDSE and H. J. GROENR~EGEN
Fig. 5. Photomicrographs of PHA-L-labelied frbres in the prefrontal cortex (A) and the subiculum (ED) following an injection in the anterior part of the paraventrkadar nucleus. (A> LabaIled fibres in the ventral part of the prelimbic cortex. The tibres terminate in layers I, V and VI. Note that the labelling in layer I is confined to the deep Part. (B)-(D) In the ventral subiculum, PHA-L-labelled fibres are arranged in two bands (B). The inset in (B) is shown at a higher magnification in (D). A comparison with an adjacent section that was counterstained with Cresyl Violet (C) demonstrates that one of the two bands of labelled fibres is located deeply in the molecular layer, whereas the other involves the deep part of the pyramidal cell layer. Asterisks in (C) and (D) indicate corresponding blood vessels. Scale bar in (A) = 200 pm and
also applies to (B). Scale bar in (C) = 50pm and also applies to (D).

Thalamocortical projections
Fig. 6. Chartings of the anterograde labelling in the cerebral cortex resulting from a PHA-L injection in the intermediodorsal nucleus (case RP-89358). Passing and terminating fibres are differentially indicated. The tracer deposit is illustrated in the upper right hand corner of this figure and in Pig. 1B. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings.
Arrowheads mark cytoarchitectonic boundaries between cortical fields.

82 H. W. BERENDSE and H. J. GROENEWEGEN
Fig. 7. Photomicrographs of two transverse sections through the dorsal agranular insular cortex illustrating the distribution of PHA-L-labelled fibres resulting from an injection in the intermediodorsal nucleus. The section in (A) was counterstained with Cresyl Violet. The fibres terminate in layers I, III and superficial V (A). Note in (B) the smooth appearance of the fibres in deep layer V. Scale bar in
(A) = 200 pm and also applies to (B).
insular cortex, throughout its rostrocaudal extent. The laminae with the highest density of labelling are layers I, III and the superficial part of layer V (Figs 6A-D and 7). The labelling is continuous with that in the rostra1 part of the posterior agranular insular cortex (Fig. 6E). At more caudal levels of the latter cortical field, label is sparse and restricted to the deep cortical layers (Fig. 6F-H). In the medial prefrontal cortex, labelled fibres are limited to the rostroventral part of the prelimbic cortex involving layers I, III and superficial V (Fig. 6A-C). Dense labelling is found throughout layer V of the lateral part of the entorhinal cortex. A few labelled fibres terminate in layers I and V of the ventral part of the perirhinal cortex (Fig. 61-K).
Rhomboid nucleus. Injections in the rhomboid nucleus (e.g. case RP-87157; Fig. 8) result in a quite different distribution of PHA-L-labelled fibres in the cerebral cortex than in the preceding cases. In case RP-87157, many fibres are present in layer I of the most ventral part of the infralimbic cortex which lies immediately adjacent to the taenia tecta and is some- times referred to as the dorsal peduncular cortex lo7 (Fig. 8C). Other concentrations of label involve layers I and V of the perirhinal cortex and the molecular layers of the dorsal parts of CA1 and the subiculum (Fig. 8G-L). In addition to these areas of dense labelling, there is diffuse and widespread labelling of virtually all cortical areas. The labelled fibres terminate in layers I and/or superficial V; in some areas the label extends from layer V into layer IV (Fig. 8). The fibres in layer VI are predominantly of the passing
type.
Intralaminar thalamocortical projections
Deposits of PHA-L were placed in the rostra1 and caudal intralaminar nuclei (see for example Fig. 1C). The range in size of the injections is similar to that of the midline injections. In all cases, the labelling in the cerebral cortex is restricted to the ipsilateral side of the brain, except for the central medial cases in which the injection sites extend across the midline. Most fibres reach the cerebral cortex along similar routes as the midline thalamic fibres. Following injections dorsolaterally in the intralaminar nuclei, the fibres enter the internal capsule directly from the reticular thalamic nucleus.
The intralaminar nuclei project to restricted areas of the prefrontal cortex and the neocortex. A medio- lateral gradient in the thalamus corresponds to a mediolateral gradient in the cerebral cortex. Further- more, rostra1 injections produce a terminal plexus in the cortex characterized by an extensive rostrocaudal distribution, with relative sparing of extreme rostra1 levels, whereas caudal injections result in labelling that is largely confined to rostra1 cortical areas. In general, the projections to the caudolateral parts of the cerebral cortex are less dense and cover larger areas. The laminar distribution of the fibres in the cortex is dependent upon the thalamic nucleus injected. The morphology of the fibres that arise from the intralaminar nuclei is similar to that of the fibres originating in the midline nuclei (see above).
Central medial nucleus. In case RP-86108, the PHA-L injection was placed in the central medial nucleus (Fig. 9). The resulting label in the cerebral

Thalam~orti~l projections
. AGm
m
Fig. 8. Chartings of PHA-L-lazily fibres in the cerebral cortex foilowi~g an injection in the rhomboid nucleus (case RP-87157). Passing and terminating fibres are differentially indicated. The injection site is drawn in the upper right hand comer. Note that the labelled fibres are present in widespread cortical areas. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other
chartings. Arrowheads mark cytoarchitectonic boundaries between cortical fields.
H. W. BEREND~E and H. 1. GROENEWEGEN
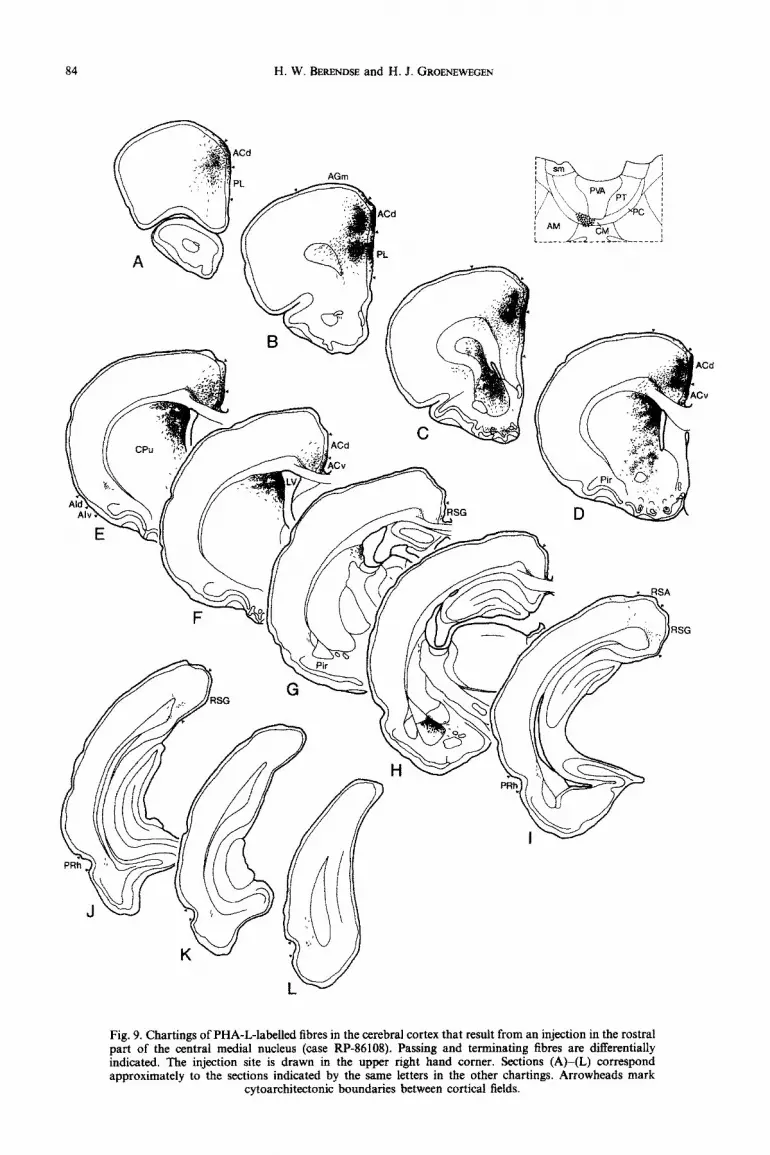
Fig. 9. Chartings of PHA-L-labelled fibres in the cerebral cortex that result from an injection in the rostra1 part of the central medial nucleus (case RP-86108). Passing and terminating fibres are differentially indicated. The injection site is drawn in the upper right hand corner. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings. Arrowheads mark
cytoarchitectonic boundaries between cortical fields.
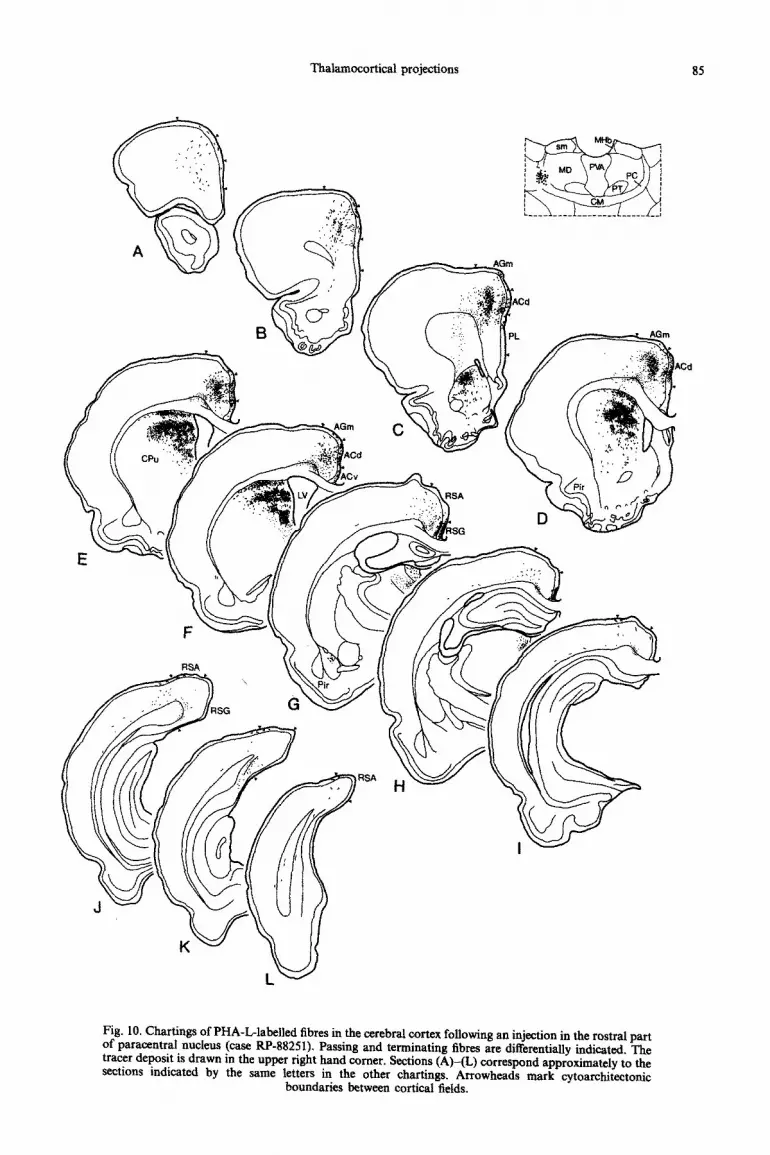
Thalamocortical projections 85
r SnJ
j&” pIATPc
:= \ j
CM L____ ____ ___-..-.._ ____- __I
d
Fig. 10. Chartings of PHA-L-labelled fibres in the cerebral cortex following an injection in the rostra1 part of paracentral nucleus (case RP-88251). Passing and terminating fibres are differentially indicated. The tracer deposit is drawn in the upper right hand comer. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings. Arrowheads mark cytoarchitectonic
boundaries between cortical fields.

86 H. W. BERENDSE and H. J. GROENEWEGEN
cortex is largely confined to prefrontal areas. A dense plexus of te~inating tibres covers the rostrocaudal extent of the dorsal part of the prelimbic cortex and the rostra1 two-thirds of the dorsal anterior cingular cortex (Figs 9A-E and llA, B). At the level of the forceps minor of the corpus callosum, the field of terminations breaks up into two separate areas, with a narrow intervening zone that is unlabelled and coincides with the border zone between the prelimbic and the anterior cingular cortical fields (Fig. 9B). In both cortical fields, the labelled fibres terminate most densely in layers I and V. The superficial part of layer V is more densely labelled than its deep part. Layer III is only slightly involved (Fig. 11 A, B). At caudal levels of the anterior cingular cortex, the terminal plexus shifts from the dorsal into the ventral subfield (Figs 9E, F and llC, D). A few scattered fibres terminate in the deep layers of the perirhinal cortex (Fig. 91, J). In other cases with similarly placed but somewhat larger injections, sparse labelling can also be observed in the deep layers of the occipital cortex.
Paracentral nucleus. The PHA-L injection in case RP-88251 is centred in the rostra1 part of the para- central nucleus. At this thalamic level the central lateral nucleus is not present.“*ro7 As a consequence of the small size of the rostra1 part of the paracentral nucleus, the PHA-L deposit involves the adjacent anteroventral and mediodorsal thalamic nuclei (Fig. 10). The injection produces dense fibre and terminal labelling in the dorsal anterior cingular cortex that extends into the medial part of the medial agranular cortex (Fig. IOB-F). The labelling is strongest at intermediate rostrocaudal levels and is concentrated in layers I and superficial V. A few fibres terminate in layer III. An additional plexus of labelled fibres in layers I and III of the retrosplenial area (Fig. IOG, H) probably results from the involve- ment of the anteroventral nucleus in the injection site. A few smooth fibres pass through the deep layers of the occipital cortex (Fig. IOJ).
In case RP-86041, the PHA-L deposit involves the caudal part of the paracentral nucleus and the adjacent ventral part of the mediodorsal nucleus. Three areas of termination are present in the cerebral cortex. The first and most dense area occupies the rostra1 part of the dorsal agranular insular cortex, where the majority
of labelled fibres terminate in layers I, III and V. Layer VI contains pr~ominantly passing ftbres. At the rostra1 tip of the hemisphere, the labelling extends from the dorsal agranular insular cortex into the lateral orbital cortex (Fig. 11F). The second plexus of terminating fibres is situated rostrally in the lateral agranular and the lateral part of the medial agranular cortical fields. The labelled fibres terminate in layer I and at the border between layers IV and V (Fig. 1lE). The last and least densely labelled area involves the posterior agranular insular and the perirhinal cortices,
Central lateral nucleus. The PHA-L deposit in case RP-88277, placed at a more caudal thalamic level compared to that in case RP-88251, is centred in the rostra1 part of the central lateral nucleus (Fig. 12) and may slightly involve the adjacent mediodorsal and ventrolateral nuclei. Subsequent to this injection labelled fibres are present in the frontal and parietal cortices. In the frontal cortex, the labelling is re- stricted to the medial agranular field and the adjacent medial part of the lateral agranular field. The fibres in the frontal cortex are organized in three bands (Figs 12B-F and 14A). One of these bands is only weak and consists of a few fibres in layer I that course parallel to the pial surface. Other fibres terminate at the border of layers III and IV and are oriented perpendicular to the pial surface. The third band of labelling has the highest density of fibres and resides in the deep part of layer V. The other cortical layers predominantly contain passing fibres (Fig. 14A). At the caudal end of the prefrontal cortex, the label extends into the rostra1 part of the retrosplenial cortex where it tapers off rapidly in a caudal direction (Fig. 12G, H). The labelling of the parietal cortex is restricted to the caudal part of parietal area I (Fig. 12F-I). A small number of fibres terminate in layer I, whereas one band of more extensive labelling involves layer IV, and another deep layer V and super- ficial layer VI (Fig. 14B). The sparse fibres observed rostrally in the occipital cortex bear only very few varicosities (Fig. 12H, I). A few fibres terminate in the deep layers of the ventrolateral orbital cortex (Fig. 12A, B).
Following an injection of PHA-L in the caudal part of the central lateral nucleus, that involves the lateral part of the mediodorsal nucleus (case RP-87219) the
Fig. 1 I. (A)-(D) PHA-L-la~ll~ fibres in frontal sections through the prefrontal cortex following an injection in the central medial nucleus. In both the dorsal part of the prelimbic (A) and the dorsal anterior cingular cortex (B) labelled fibres terminate densely in layers I and superficial V. Smooth fibres of passage are present in the other layers. The labelling in the ventral anterior cingular cortex is sparse (C). A high-power photomicrograph of a section counterstained with Cresyl Violet (D) demonstrates that the fibres terminate in layers I and superficial V. (E),(F) Photomicrographs of the PHA-L labelling in the cerebral cortex that results from an injection in the caudal part of the paracentral nucleus. In the lateral agranular field of the motor cortex, labelled fibres terminate predominantly in layer I and at the border between layers IV and V (E). The extremely dense labelling of the lateral orbital and dorsal agranular insular cortical fields is concentrated in layers III and V. A few fibres terminate in layer I. Scale bar in
(A) = 2OOpm and also applies to (B), (C), (E) and (F). Scale bar in (D) = 50 pm.
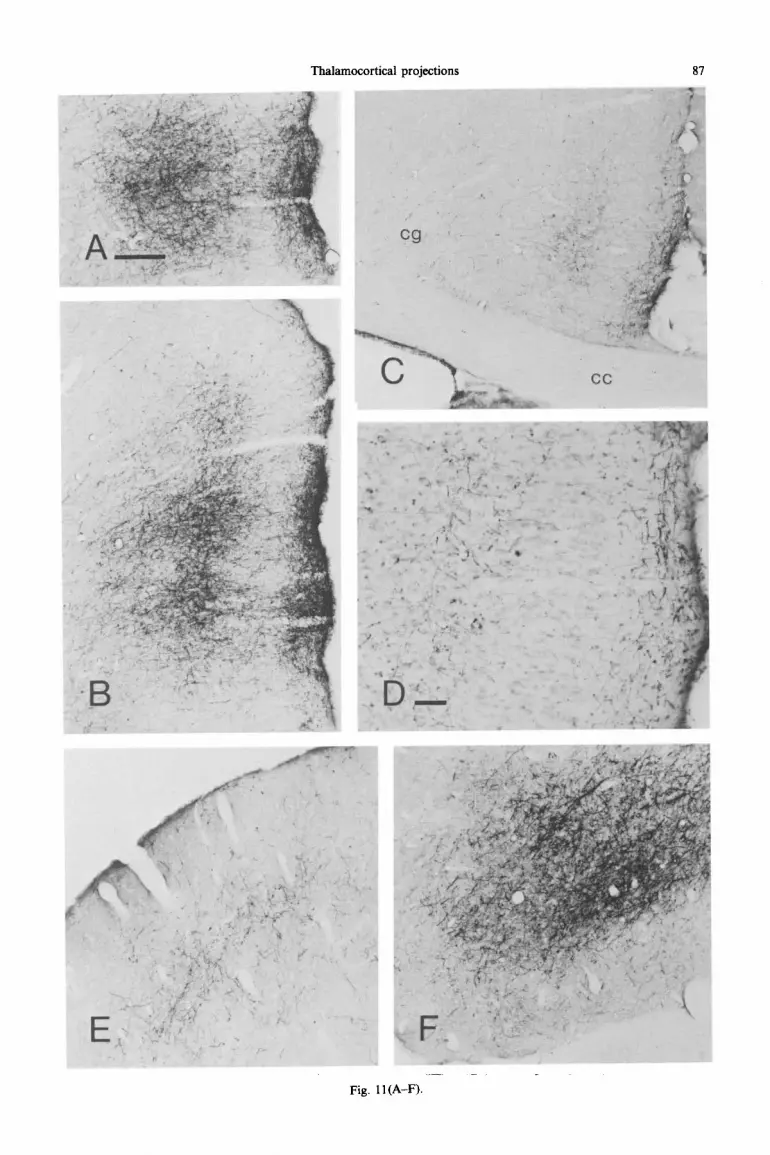
Thalamocortical projections 87
Fig. 11 (A-F).

H. W. BERENDSE and H. J. GROENEWEGEN
Fig. 12. Chartings of the anterograde labelling in the cerebral cortex following a PHA-L injection into the rostra1 part of the central lateral nucleus (case RP-88277). Passing and terminating fibres are differentially indicated. The injection site is illustrated in the upper right hand corner. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings. Arrowheads
mark cytoarchitectonic boundaries between cortical fields.

Thalamocortical projections 89
highest density of labelling is present rostrally in layer III of the dorsal anterior cingular and agranular cortical fields. Very few fibres terminate in layer I. In the agranular fields, some fibres terminate in&tperIicial layer V. Contrary to case RP-88277, no labelled fibres are found in the parietal cortex in case RP-87219, but the labelling in the ventrolateral orbital cortex, which includes layers I, III and V, is more dense than that in case RP-88277.
Parafascicular nucleus, lateral part. Following injections of PHA-L in the parafascicular nucleus, labelled fibres distribute over the lateral agranular field of the frontal cortex and the parietal cortex. The majority of the fibres terminate rostrally in these cortical areas and are concentrated in four bands situated in layers I, IV, deep V and deep VI (Fig. 14C, D). Layers V and VI are most densely labelled. In the agranular field of the frontal cortex the strip of label in layer IV is uninterrupted, whereas in the dorsolateral part of the parietal cortex it is more sparse and discontinuous.
The tracer deposit in case RP-88049 is located in the centre of the lateral part of the parafascicular nucleus (Fig. 13). In the cerebral cortex, the PHA-L- labelled fibres occupy the lateral agranular cortex, the most lateral part of the medial agranular cortex and the dorsolateral part of parietal area I (Figs 13AG and 14C, D). A minority of fibres terminate in the forelimb and hindlimb areas. The labelling in the caudal part of the parietal cortex is weak and restricted to the deep cortical layers. A few fibres are present in the rostra1 part of the occipital cortex (Fig. 13H, I). In case RP-88090, the PHA-L deposit is smaller than that in case RP-88049 and involves only the most rostro- lateral part of the parafascicular nucleus (Fig. 1C). In the cerebral cortex, the labelled fibres have a more medial position than in case RP-88049 and thus occupy the rostrolateral part of the lateral agranular cortex and the rostra1 part of the parietal cortex. The most caudally located fibres in the parietal cortex are concentrated in parietal area II. The laminar distri- bution in the latter cortical area is somewhat different from that described above. Two bands of terminating fibres are present, the first in layer I and the other at the border between layers IV and V. The label in layer VI consists of smooth passing fibres.
Parafascicular nucleus, medial part. The cortical projections of the medial part of the parafascicular nucleus are rather sparse. The highest density of labelled fibres is present in the dorsal part of the pre- limbic cortex, the anterior cingular cortex, and the caudal part of the medial agranular cortex. In the latter areas, the label is restricted to layers I, V and VI. It is interesting to note that the projections to these areas overlap to a large extent with those of the central medial nucleus (cf. case RP-86108). A few fibres are further observed in the deep layers of the agranular insular cortex, the occipital, perirhinal and entorhinal cortices, the subiculum, and the parasubiculum.
DISCUSSION
The most striking finding of the present study of the midline and intralaminar thalamocortical projections in the rat is that, contrary to the results of previous anterograde tracing studies in rat@‘@ and cats,80,‘27 the individual nuclei of the midline and intralaminar nuclear complex project to specific parts of the cerebral cortex. Thus far, a topographically organized pro- jection to the cerebral cortex has been reported only for the centromedian-parafascicular complex in the cat.‘26 Of equal importance is the present observation that the laminar distributions in the cerebral cortex of the midline and intralaminar fibres are not the same for all nuclei. Both findings support the notion that there is a high degree of specificity in the connections of the midline and intralaminar nuclear complex, li,25,26@ A combination of the present results with those of studies on the thalamostriatal” and the prefrontal corticostriatal projections’* reveals that particular parts of the midline and intralaminar thala- mic nuclear complex project to specific cortical areas as well as to the striatal targets of these areas. The implication of this arrangement is that each of the recently proposed basal ganglia-thalamocortical circuits3@ can be influenced both at the cortical and the striatal level by the same part of the midline and intralaminar nuclear complex.
Area1 organization of midline thalamocortical projections
Previous studies have demonstrated that the cortical projections of the midline thalamic nuclei are predominantly ipsilateral; only a small number of neurons in the midline of the thalamus project contralaterally.96~“2~‘42 In agreement with this, the present PHA-L injections in the midline nuclei gave rise to bilateral labelling. Following injections in the parataenial nucleus, well outside the midline, labelhng was virtually absent from the contralateral hemisphere. The major cortical targets of the midline thalamic nuclei, not including the rhomboid and reuniens nuclei, are the ventral part of the medial prefrontal cortex, several subfields of the lateral prefrontal cortex, and the hippocampal and parahippocampal regions. A sparse innervation of the perirhinal cortex was found to originate from the paraventricular and intermediodorsal nuclei (cf. Refs 29 and 63). The projections to the prefrontal cortex are virtually non- overlapping, which is at variance with the results of a recent retrograde tracing study with fluorescent tracers in the rat by Conde et af.24 In the present anterograde tracing study, it could be demonstrated that many passing fibres course through the ventral parts of the medial prefrontal cortex on their way to more dorsal cortical areas. The uptake of fluorescent tracer by these fibres is a likely explanation for the more widespread and overlapping projections from the midline and intralaminar thalamic nuclei to the medial prefrontal cortex, as found by CondC et a1.24
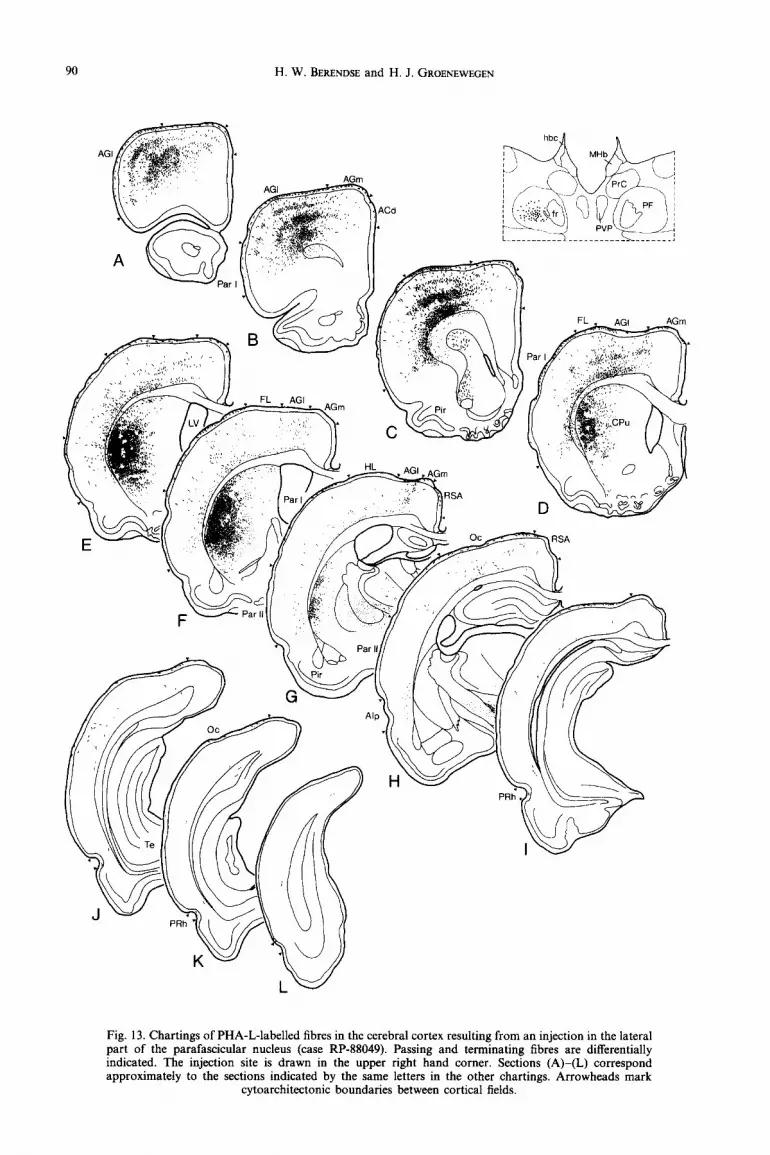
H. W. BERENDSE and H. J. GROENEWEGEN
Fig. 13. Chartings of PHA-L-labelled fibres in the cerebral cortex resulting from an injection in the lateral part of the parafascicular nucleus (case RP-88049). Passing and terminating fibres are differentially indicated. The injection site is drawn in the upper right hand corner. Sections (A)-(L) correspond approximately to the sections indicated by the same letters in the other chartings. Arrowheads mark
cytoarchitectonic boundaries between cortical fields.

Fig. 14. Photomicrographs of the PHA-L labelling in the cerebral cortex following injections in the rostra1 part of the central lateral (A, B) and in the lateral part of the parafascicular nucleus (C, D). (A) Transverse section through the agranular fields of the motor cortex. Labelled fibres terminate in layer I, at the border between layers III and IV, and in deep layer V. Note that the highest density of terminating iibres is in layer V. (B) High-power photomicrograph of PHA-L-labelled fibres in layer V of the parietal cortex. Countemtained with Cresyl Violet. (C) In the lateral agranular field of the motor cortex and in the parietal cortex, PHA-L-la~ll~ fibres from the parafascicular nucleus distribute over layers I, IV, deep V and deep VI. Many smooth passing fibres intermingle with the te~inating fibres in layer VI. Note that the fibres terminating in layer IV are oriented perpendicular to the pial surface. (D) High-power photomicrograph of PHA-L-labelled hbres from the parafascicular nucleus in deep layer V of the parietal cortex. Counterstained with Cresyl Violet. Scale bar in (A) = 200 pm
and also applies to (C). Scale bar in (B) = 50 ym and also applies to (D).

92 H. W. BERENDSE and H. J. GROENEWEGEN
For the parataenial nucleus we found a projection to the medial orbital and infralimbic subfields of the medial prefrontal cortex. Previous retrograde tracing studies have shown a projection from the parataenial nucleus to the infralimbic cortex,33,78,‘46 whereas a projection to the medial orbital cortex was inferred from the results of several large injections of tritiated proline in the mediodorsal and adjacent thalamic nuclein4 Interestingly, the parataenial nucleus has been related to the basal parts of the medial surface of the hemisphere as early as 1951 by Droogleever- Fortuyn,” who noted degenerative changes in this nucleus following lesions of the cerebral cortex in the rabbit. The absence of a projection from the medio- dorsal nucleus to the medial orbital and infralimbic cortices55,84 suggests that the relationship of the parataenial nucleus with these cortical areas may be partly comparable to the relationship of the medio- dorsal nucleus with the other prefrontal cortical areas (see Laminar organization of midline and intralaminar thalamocortical projections). The presence of sparse labelling in the ventral subiculum and the entorhinal cortex in our experiments is in accord with the results of retrograde tracing studies on the thalamic afferents of the hippocampal and parahippocampal regions.7~71~119~'"
In the cat, retrograde labelling of neurons in the paraventricular nucleus has been reported following tracer injections in the cingular and medial prefrontal cortices.‘OO~‘O’ In the rat, projections from the para- ventricular nucleus to the cerebral cortex have either been denied’* or were considered to be sparse and widespread.69 The present anterograde tracing study is the first to demonstrate that the projections from the paraventricular nucleus are substantial but, neverthe- less, restricted to a few cortical areas. These areas include the ventral part of the prelimbic cortex and, though to a lesser extent, the agranular insular cortex and the ventral part of the anterior cingular cortex. Furthermore, our results indicate that the projection to the latter area is probably derived from the ventral part of the paraventricular nucleus. In agreement with the results of previous retrograde tracing studies 7~71~“9~‘37~‘40~‘54~‘55 we found projections from the anterior part of the paraventricular nucleus to the subiculum and the entorhinal cortex. The posterior part of the nucleus does not seem to provide any significant contribution to either of these projections. The paraventricular nucleus projects sparsely to the perirhinal cortex (present results).
Contrary to the parataenial and paraventricular nuclei, the intermediodorsal nucleus projects most densely to the lateral part of the prefrontal cortex. In our experiments, labelled fibres were observed in layers I, III and superficial V of the posterior agran- ular insular cortex and the ventral part of the dorsal agranular insular cortex. A comparison with retro- grade tracing studies is difficult, since in these studies the intermediodorsal nucleus was not identified as a separate entity but was instead included in the caudo-
medial part of the mediodorsal nucleus48s55 or in the central medial nucleus.“4 Some retrogradely labelled neurons illustrated in these studies appear to lie quite close to or even in that part of the thalamus which we identify as intermediodorsal nucleus. Moreover, a fluorescent tracer injection, which is located super- ficially in the dorsal agranular insular cortex, results in extensive retrograde labelling of both the caudo- medial part of the mediodorsal nucleus and the inter- mediodorsal nucleus (Groenewegen, unpublished observations). The projection to layer V of the agranular insular cortex that we presently observed most probably originates from the intermediodorsal nucleus. The labelling in layers I and III may derive from the caudomedial part of the mediodorsal nucleus for which a projection to these layers has been reported.55,84 As the projection to layer I from the caudomedial part of the mediodorsal nucleus in the experiments of Krettek and Pric@“ and Groenewegen55 is quite sparse compared to that demonstrated in the present experiments, this layer presumably receives projections from both the mediodorsal and inter- mediodorsal nuclei.
The presence of a sparse projection from the intermediodorsal nucleus to the ventral part of the prelimbic cortex is in agreement with the results of retrograde tracing studies.24*55’46 As we discussed for the agranular insular cortex, the involvement in the injection site of the caudomedial part of the medio- dorsal nucleus probably accounts partly or entirely for the labelling in layers I and III (see preceding paragraph). The projection to the lateral entorhinal area, which we observed in our material, has not been reported in retrograde tracing studies in rats or cats.7,‘54,‘55 In the monkey, many retrograde neurons have been described in the centralis complex of the thalamus following tracer injections in lateral parts of the entorhinal cortex.” According to Jones,” part of this complex in the monkey is comparable to the intermediodorsal nucleus of the rat.
The pattern of cortical projections of the rhomboid and reuniens nuclei is quite different from that described above which emphasizes that the group of midline nuclei is heterogeneous as far as their connec- tions are concerned (see also Refs 11, 140 and 153). The rhomboid nucleus projects densely to only a few cortical areas, including the most ventral part of the infralimbic cortex, the perirhinal cortex, and the dorsal parts of the subiculum and CAI. Additionally, there is a diffuse projection to widespread areas of the cerebral cortex, most strongly to the frontopari- eta1 areas (present results; cf. Ref. 104). Consistent with these results, retrograde tracing studies have demonstrated the labelling of neurons in the rhom- boid nucleus subsequent to injections in various parts of the medial and lateral prefrontal cortical areas.7*~‘oo~‘o’~“4~120~‘46 Like the rhomboid nucleus, the nucleus reuniens projects densely to a few cortical areas and more diffusely to a larger cortical area. The dense projections involve the ventral part of the infra-

Thalamocorticsl projections 93
limbic cortex, the ventral anterior cingular cortex, the subiculum, and CAl.66*‘04,‘53
Area1 organization of intralaminar thalamocortical projections
The rostra1 and caudal intralaminar nuclei project pr~omin~tly to the ipsilateral cerebral cortex (present results and Refs 67, 69, 78, 80, 89, 90, 126, 127 and 144). In the central medial nucleus, ipsi- and contralaterally projecting neurons intermingle in the midline.~“2~‘42 Except for the centromedian- parafascicular complex of the cat, which was found to project topographically to specific regions of the rostra1 cerebral cortex,‘26 anterograde tracing exper- iments have so far failed to demonstrate any sub- stantial differences in the cortical projections of the intralaminar nuclei.67~69~80~‘27 A rough topographical organization in the intralaminar thalamocortical projections was already demonstrated with the retro- grade degeneration method in 1966 by Murray.* Furthermore, the results of retrograde tracing studies in several mammalian species suggest that the intra- laminar nuclei have preferential zones of termination in the cerebral cortex72~78**3**g~‘4 (for further references see Ref. 88). This is substantiated by the finding in anatomical and physiological studies that branching of single intralaminar neurons to separate cortical areas is either minimal or entirely absent.9~1J~80~90~139 The present study is the first to substantiate the results of the retrograde tracing experiments by means of anterograde tract tracing. As a group, the intralaminar nuclei project predominantly to rostra1 parts of the cerebra1 cortex, including the frontal and parietal areas. The projections to the prefrontal cor- tex are restricted to the dorsal parts of the prelimbic and the agranular insular cortical fields, the caudal part of the ventral anterior cingular cortex, and the lateral and ventrolateral orbital areas, and are thus complementary to those of the midline nuclei (see above).
The central medial nucleus densely innervates the dorsal part of the prelimbic cortex and the rostra1 two-thirds of the dorsal anterior cingular cortex. A less dense projection is aimed at the caudal part of the ventral anterior cingular cortex. These data are in agreement with the results of retrograde tracing studies in rats and cats,‘s89*‘oo.‘43~‘” Since our injections involve only the rostra1 part of the central medial nucleus, we could not con&m the existence of thalamic inputs from this nucleus to the agranular insular cortex, the temporal cortex and the face representa- tion of the motor cortex, which appear to originate exclusively from the caudal part of central medial nucleusz~1’4~13’~‘44 (see also references in Ref. 88). In the present study, the perirhinal cortex is the only temporal cortical area that receives a sparse pro- jection from the central medial nucleus. The presence of sparse labelling in the occipital cortex in our experiments with large injections in the central nucleus substantiates the results of previous studies.@J2’ Using
anterograde tracing of tritiated amino acids from the rostra1 part of the central medial nucleus in the rat, Herkenham67,6g reported that this nucleus projects to virtually all cortical areas. The discrepancy with the results of our PHA-L injections can be explained by the fact that Herkenham@ includes the ventrally adjacent rhomboid nucleus in his central medial nucfeus. The involvement of the rhomboid nucleus may account for the observed widespread projections (present results). In the cat, the cortical projections of the central medial nucleus heavily involve the orbitofrontal and cingular fields.127 The additional projections to other cortical areas may be the result of the involvement in the injection site of the ventral part of the central medial nucleus which corresponds to the rhomboid nucleus of the rat.“*77*‘07
The cortical projection field of the paracentral nucleus comprises the dorsal anterior cingular cortex, the agranular fields of the frontal cortex, the agranular insular cortex, the perirhinal cortex and the lateral orbital cortex. Involvement of the mediodorsal nucleus in the injection site probably accounts for the fibres in layer III of the dorsal anterior cingular and agranular insular cortical fields, and may contribute to the pro- jections to layer Iss@ (see also Laminar organization of midline and intralaminar thalamocortical projections). Rostra1 parts of the paracentral nucleus project to intermediate rostrocaudal levels of the dorsal anterior cingular cortex and the medial part of the medial agranular field. The presence of PHA-L-labelled fibres in layers I and III of the retrosplenial cortex in case RP-8825 I probably results from the involvement of the adjacent anteroventral thalamic nucleus in the injection site.?’ Fibres from the caudal parts of the paracentral nucleus terminate rostrally in the lateral orbital cortex, the dorsal agranular insular cortex, the lateral agranular field and the lateral part of the medial agranular field. Minor projections were found to the posterior agranular insular and perirhinal cortices. These findings are largely con- sistent with observations in retrograde tracing stud- ies 2~35~‘8~8~~‘~~‘20~130~‘~3,161 however, he projections to
visual, auditory and temporal cortical areas that have been reported in the cat’@‘” (see also references in Ref. 88) could not be co&rned in the rat. The projections described in the present study are more restricted than those reported in previous anterograde tracing studies.67@~80*‘27 This may be explained by the fact that with PHA-L, in contrast to tritiated amino acids, very small deposits of tracer can be made and that terminating fibres can be distinguished from passing fibres. Alternatively, the fact that the small PHA-L deposits in the present study do not involve the total rostrocaudal extent of the paracentral nucleus may have caused us to underestimate the total projection of this nucleus. However, this could only have been the case if specific subgroups of neurons with projections different from the ones described in the present study were unevenly distributed over the rostrocaudal axis, as our injections were placed at

94 H. W. BERENDSE and H. J. GROENEWEGEN
several rostrocaudal levels. The discrepancies with the studies in the cat80~‘27 could be explained by species differences. However, the high degree of differen- tiation in the projections to the striatum from the same nuclei in both rat and cat8*n,75 render the latter explanation quite unlikely.
The central lateral nucleus is considered to be a source of input to the frontal and parietal cortical gelds ~6,21.35.3~~,65,72~,89,105.115,128,132.144 me present data
confirm the presence of projections to the agranular fields of the frontal cortex and to caudal parts of the parietal cortex and give some indication that a ros- tral-to-caudal gradient in the central lateral nucleus corresponds to a caudal-to-rostra1 gradient in the cerebral cortex. The projection to the parietal cortex originates exclusively from the rostra1 part of the central lateral nucleus. Since the lateral part of the mediodorsal nucleus projects to layers I and III of the anterior cingular and medial agranular cortical fields, the involvement of this part of the mediodorsal nucleus in the injection sites can explain the projections to layers I and III of these cortical areas.ss,84 Projections to widespread frontal and parietal cortical areas have been reported for the paralaminar part of the ventro- lateral nucleus,69 which lies immediately adjacent to the central lateral nucleus and may have been involved slightly in our rostra1 central lateral injection. How- ever, retrograde tracing studies have demonstrated that the populations of neurons in the ventrolateral nucleus projecting to the lateral agranular field of the motor cortex and the first somatosensory area are not adjacent to the rostra1 part of the central lateral nucleus.2’s36 Neurons projecting to the medial agran- ular field of the motor cortex are located both in the rostra1 part of the central lateral nucleus and in the adjacent part of the ventrolateral nucleus.“’ There- fore, the involvement of the ventrolateral nucleus in our injection in the rostra1 part of the central lateral nucleus may have contributed to the projection to the medial agranular field of the motor cortex. The present observation of a projection to the agranular part of the retrosplenial cortex is consistent with the results of previous retrograde tracing studies.‘aC,‘20,‘43,is6 Finally, a sparse projection from the central lateral nucleus to the ventrolateral orbital cortex was present in our material. In the cat and the monkey, a more fine- grained topography of the cortical projections of the central lateral nucleus has been demonstrated with retrograde tracing methods.72~132
In the present study, we have demonstrated that in the rat the lateral part of the parafas~icular nucleus, corresponding to the centromedian nucleus of the cat,77 projects more densely to the cerebral cortex than the medial part, which is comparable to the parafascicular nucleus of the cat.77 The projections from the lateral part of the parafascicular nucleus predominantly involve the rostra1 portions of the lateral agranular and parietal cortical areas. The forelimb and hindlimb areas and the occipital cortex are only sparsely innervated. The above-mentioned
projections thus appear to converge with those of the adjacent posterior nucleus, for which projections to both motor and parietal cortical areas have been demonstrated in the anterograde tracing study of Herkenham.69 Retrograde tracing studies confirm that both the parafascicular and the posterior nucleus project to the parietal cortex and the lateral agranular field of the motor cortex.s,2’,36 The cortical projections of the medial part of the parafasci~ular nucleus are to some extent complementary to those of the lateral part and include the prelimbic cortex, the anterior cingular cortex and the medial agranular cortex. Minor projec- tions reach the agranular insular, occipital, perirhinal and entorhinal cortices, and the hippocampus. The above-mentioned findings are consistent with the results of the anterograde tracing study in the cat by Royce and Mourey.iz6 In their study, the motor and premotor cortical fields constitute the major target of the centromedian-parafascicular complex, but non- motor areas are involved as welln6 In particular, the parafascicular nucleus has been mentioned as a source of afferents to the medial prefrontal cortex in cats and monkeys. ‘01,‘46 In the rat, projections from the parafascicular nucleus to the motor cortex have been reported using both anterograde and retrograde trac- ing techniques (Refs 2, 34, 67 and present results). Jones and Leavitt” related the parafascicular nucleus to the ventrolateral surface of the hemisphere, near the frontal pole, whereas Ullbn’” reported widespread cortical projections with a frontal predominant. In studies of the afferent connections of the agranular insular cortex in rodents, retrograde labelling of neurons in the medial part of the parafascicular nucleus has been reported.48.“4
Relationship with the parallel arrangement of basal ganglia-thalamocortical circuits
The midline and intralaminar nuclei each project to a specific part of the cerebral cortex, although there is some overlap with the projections from neighbour- ing nuclei. The cortical targets of the midline nuclei are located in the ventral parts of the cerebral cortex, whereas the intralaminar nuclei predominantly target more dorsally located cortical areas. Furthermore, rostra1 parts of the midline and intralaminar complex are related to the medial and dorsomedial surface of the hemisphere, whereas caudal parts innervate the ventrolateral, lateral and dorsolateral surfaces of the cerebral cortex. Lastly, a rostral-to-caudal gradient in the thalamus corresponds to a caudal-to-rostra1 gradient in the cerebral cortex.
In a previous report, we have described the topo- graphical organization of the projections from the midline and intralaminar nuclei to the striatum in the rat” (see also Refs 8, 75 and 129). On the basis of the available literature on the afferent connections of these nuclei, we distinguished three nuclear groups, of which the limbic-afferented group of midline nuclei projects to medial parts of the striatum. The motor- related lateral parafascicular nucleus issues flbres
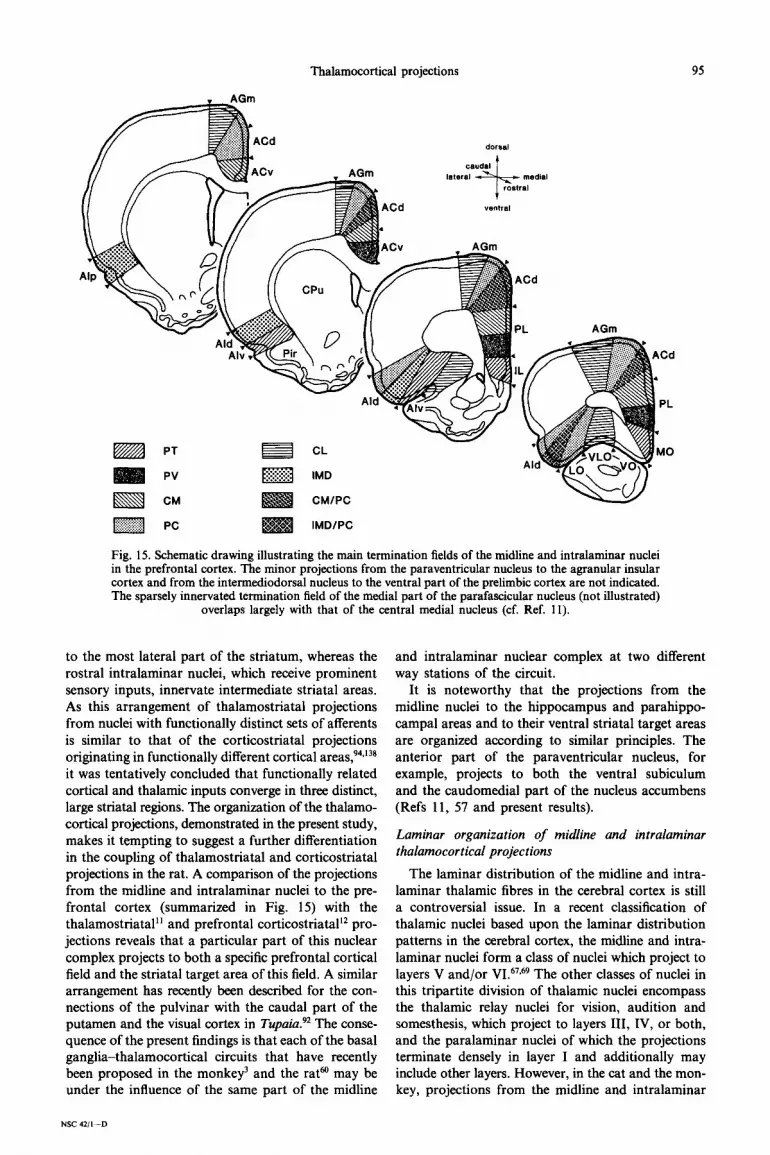
Thalamocortical projections
PV
lss CM m pc
m ,MD
CMlPC
MD/PC
Fig. 15. Schematic drawing illustrating the main termination fields of the midline and intralaminar nuclei in the prefrontal cortex. The minor projections from the paraventricular nucleus to the agranular insular cortex and from the intermediodorsal nucleus to the ventral part of the prelimbic cortex are not indicated. The sparsely innervated termination field of the medial part of the parafascicular nucleus (not illustrated)
overlaps largely with that of the central medial nucleus (cf. Ref. 11).
to the most lateral part of the striatum, whereas the rostra1 intralaminar nuclei, which receive prominent sensory inputs, innervate intermediate striatal areas. As this arrangement of thalamostriatal projections from nuclei with functionally distinct sets of afferents is similar to that of the corticostriatal projections originating in functionally different cortical areas,94,138 it was tentatively concluded that functionally related cortical and thalamic inputs converge in three distinct, large striatal regions. The organization of the thalamo- cortical projections, demonstrated in the present study, makes it tempting to suggest a further differentiation in the coupling of thalamostriatal and corticostriatal projections in the rat. A comparison of the projections from the midline and intralaminar nuclei to the pre- frontal cortex (summarized in Fig. 15) with the thalamostriatal” and prefrontal corticostriatal’* pro- jections reveals that a particular part of this nuclear complex projects to both a specific prefrontal cortical field and the striatal target area of this field. A similar arrangement has recently been described for the con- nections of the pulvinar with the caudal part of the putamen and the visual cortex in Tupuia?* The conse- quence of the present findings is that each of the basal ganglia-thalamocortical circuits that have recently heen proposed in the monkey’ and the rata may be under the influence of the same part of the midline
NSC 4211-D
and intralaminar nuclear complex at two different way stations of the circuit.
It is noteworthy that the projections from the midline nuclei to the hippocampus and parahippo- campal areas and to their ventral striatal target areas are organized according to similar principles. The anterior part of the paraventricular nucleus, for example, projects to both the ventral subiculum and the caudomedial part of the nucleus accumbens (Refs 11, 57 and present results).
Laminar organization of midline and intralaminar thalamocortical projections
The laminar distribution of the midline and intra- laminar thalamic fibres in the cerebral cortex is still a controversial issue. In a recent classification of thalamic nuclei based upon the laminar distribution patterns in the cerebral cortex, the midline and intra- laminar nuclei form a class of nuclei which project to layers V and/or VI.67,69 The other classes of nuclei in this tripartite division of thalamic nuclei encompass the thalamic relay nuclei for vision, audition and somesthesis, which project to layers III, IV, or both, and the paralaminar nuclei of which the projections terminate densely in layer I and additionally may include other layers. However, in the cat and the mon- key, projections from the midline and intralaminar

96 H. W. BERENDSE and H. J. GROENEWEGEN
nuclei to superficial cortical layers, including layers I and III, have been reported repeatedly using the anterograde transport of both tritiated amino acids43s0~‘26~‘27 and PHA-L.‘25 Although these results may reflect species differences or spread of tracer in the adjacent paralaminar thalamic nuclei, superficial applications of horseradish peroxidase to the parietal cortex lead to labelling of intralaminar neurons in both cats and rat.“’ The relatively sparse labelling of the intralaminar nuclei in the latter study is quite compatible with the sparse layer I projections to the parietal cortex observed in the present study. Contra- dictory results have been obtained with superficial horseradish peroxidase applications to the visual cortex.‘9,“8 The present findings using the sensitive PHA-L technique establish that the midline and intralaminar nuclei in the rat project to deep as well as to superficial cortical layers. Physiological data are consistent with a superficial termination of afferents from the central lateral thalamic nucleus in the rat and the cat.10s~‘28
In the prefrontal cortex, the fibres from all midline and intralaminar nuclei, except for those from the paraventricular thalamic nucleus, terminate predomi- nantly in layers I and superficial V. Many passing fibres are present in layer VI and deep layer V. The additional plexus of terminating fibres in layer III, which is found in some cases, probably results from involvement of the mediodorsal nucleus in the injec- tion site.55@ In these cases, the mediodorsal nucleus may also contribute to the layer I projections.55T84 The parataenial nucleus projects to layers I, III and V of the medial orbital and infralimbic cortical fields. Interestingly, until now a projection of the medio- dorsal nucleus to the projection field of the parataenial nucleus has not been demonstrated.55~84 It may there- fore be concluded that the parataenial nucleus has characteristics of both the mediodorsal nucleus and the midline and intralaminar nuclear complex. The projections from the paraventricular nucleus have a different laminar organization than those from the other nuclei, as the fibres from this nucleus terminate not only in layers I and superficial V, but also in deep layer V and in layer VI. The projections from the midline nuclei to the entorhinal and perirhinal cortices are largely confined to layers V and VI. The wide- spread projections from the rhomboid nucleus to the prefrontal cortex predominantly involve layer I. The fibres from the rhomboid nucleus in other parts of the cerebral cortex terminate in layers I and V; in some cortical areas the fibres from the rhomboid nucleus terminate in deep layer IV (cf. Ref. 104).
The laminar distribution of the intralaminar thalamic fibres in the agranular fields of the frontal cortex and the parietal cortex as well as the relative density of labelhng in each of the layers involved are dependent upon the nuclear origin of the fibres. All nuclei project relatively sparsely to layer I. A second plexus of terminating fibres is present in layer IV and may extend into layer III. The fibres in deep layer V
may extend into the superficial part of layer VI. A projection to deep layer VI is found only for the parafascicular nucleus.
The most striking difference between the rat and the cat with respect to the laminar organization of the projections of the intralaminar nuclei is the absence of a projection to layer V in the latter species. Instead, a dense innervation of layer III has been described.80~‘26’27 Taking into consideration the intimate relationship of the midline and intralaminar nuclei with the striatum,8*“~75 the observed discrep- ancy may be related to the fact that most cortico- striatal projection neurons in the cerebral cortex of the cat reside in layer III,‘23 whereas in the rat these projections originate predominantly from neurons in layer V.94
Relationship with the compartmental organization of the striatum
A characteristic feature of the striatum is its compartmental organization, which is reflected in the heterogeneous distribution of virtually all histo- chemical markers, receptors, uptake sites, and alTerent and efferent systems. ‘4.45,46,53,54,58,59,68,147 Recently, it has
been demonstrated that in the rat the cortical afferents of the patch versus the matrix compartments in the dorsal striatum, as defined by immunohistochemistry for calbindin D,,,,, are derived from different cortical layers.47 Corticostriatal neurons in deep layer V and layer VI project principally to the patches, whereas superficial layer V and layer III and II corticostriatal neurons show a preference for the matrix. The laminar organization of the cortical projections from the mid- line and intralaminar thalamic nuclei could well be in register with this arrangement in the corticostriatal system. The striatal projections from the thalamic nuclei that are related to layers I and superficial V of the prefrontal cortex avoid areas of high immuno- reactivity for enkephalin in the ventromedial part of the caudate-putamen and the adjacent part of the nucleus accumbens.“*13 These areas coincide with areas of low immunoreactivity for calbindin D28,,.‘47 The paraventricular nucleus, which projects to the areas of high immunoreactivity for enkephalin in the aforementioned striatal regions, is the sole representa- tive of the midline and intralaminar nuclei that issues fibres to deep layer V and layer VI of the prefrontal cortical areas projecting to these striatal areas. Although these data are by no means conclusive without information on the microcircuitry at the corti- cal level, it is tempting to suggest that a particular midline or intralaminar thalamic nucleus, projecting to a distinct compartment in the striatum,‘1,‘3 can most directly inIluence those prefrontal corticostriatal neurons that project to the same striatal compart- ment. More detailed knowledge of the prefrontal corticostriatal projections and the cortical micro- circuitry is necessary to substantiate this assumption and to generalize it for the entire ventral striatum.

Thaiamocortical projections 97
Functional considerations
Over the past decade, the ideas about the midline and intralaminar nuclear complex of the thalamus have changed considerably. The original concept of a non-specific corticopetal system that relays the activity of the reticular formation to widespread areas of the cerebral cortexS2,M,73,74,98*139 has been replaced by a more differentiated view of a system of distinguishable nuclei, which each receive specific sets of afferents, project to specific parts of the striatum and the cerebral cortex, and are involved in quite specific processes. 4.8,10,11,16,25,26,30,70,87,88,93,108,113.131-135
The concept of a highly differentiated group of midline and intralaminar nuclei does not necessarily imply that these nuclei process the same type of info~ation as the relay nuclei. For example, extero- ceptive information is conveyed via separate popula- tions of neurons in the spinal cord to the midline and intralaminar nuclei, and to the relay nuclei.28~50~91~‘02~IJ2 More importantly, the spinal cord neurons projecting to the latter nuclei have small receptive fields, whereas those projecting to the midline and intralaminar nuclei have large, often bilateral, receptive fields, are not somatotopically organized, and may be polymoda11~“*5’~‘08 (cf. Ref. 17). Therefore, the relay nuclei probably subserve discriminative aspects of the ascending information, contrary to the midline and intralaminar nuclei which may be committed to non- di~~minative or affective aspects of the informa- tion.‘*20*95*‘58 These features may be required to alert
the organism of the occurrence of new and potentially dangerous situations. 61~Lo8 Since the relay nuclei of the thalamus project only sparsely to the striatum,B~*‘~7s di~~minative aspects of the ascending info~ation are relayed mainly to the cerebral cortex. The non- discriminative aspects of the information are conveyed to both the striatum and the cerebral cortex (Refs 8, 11,67, 69, 75, 80, 126, 127 and present results), either by the same or by closely adjacent neurons.Z23~7690~124,139 At the level of the cerebral cortex, both aspects of the info~ation can be integrated and an adequate response planned. Depending on the affective value, the non-discriminative aspects of the ascending infor- mation that reach the striatum by way of the midline and intralaminar nuclei can either prepare the striatum for an imminent cortical input or induce a timely first behaviouraf response to be modified by the integrated cortical input. The importance of the midline and intralaminar nuclear complex for spontaneous and reactive behaviour is stressed by the reduction in responsiveness to external stimuli (neglect) and the lack of spontaneous behaviour (hypokinesia, mutism, brad~hrenia) following damage to these nuclei both in human patients62,85,1~,i48,*~ and in animals.‘W*‘49
Acknowledgemenrs-We would like to thank Prof. Dr A. H. M. Lohman for his critical review of this manuscript, Mmes Anita Hartog-te Kortschot and Yvon Galis-de Graaf for technical assistance, Mr Dirk de Jong for his invaluable photo~phi~al skills and MS Joan Hage for secretarial support.
1.
2. 3.
4.
5.
6.
7.
8. 9.
10.
11.
12.
13.
14.
IS.
REFERENCES
Albe-Fessard D., Berkley K. J., Kruger L., Ralston H. J. III and Willis W. D. Jr (1985) Dien~phali~ mechanisms of pain sensation. Brain Res. Rev. 9, 217-296. Aldes L. D. (1988) Thalamic connectivity of rat somatic motor cortex. Brain Res. Bull. 20, 333-348. Alexander G. E., DeLong M. R. and Strick P. L. (1986) Parallel organization of functionally segregated circuits linking basal ganglia and cortex. A. Rev. Neurosci. 9, 357-381. Andersen E. and Dafny N. (1983) An ascending serotonergic pain modulation pathway from the dorsal raphe nucleus to the parafascicular nucleus of the thalamus. Brain Res. 269, 57-67. Avendano C., Rausell E. and Reinoso-Suarez F. (1985) Thalamic projections to areas Sa and 5b of the parietal cortex in the cat: a retrograde horseradish peroxidase study. J. Neurosci. 6, 1446-1470. Beckstead R, M. (1976) Convergent thalamic and mesencephalic projections to the anterior medial cortex in the rat. J. camp. Neural. 166, 403-416. Beckstead R. M. (1978) Aflerent connections of the entorhinal area in the rat as demonstrated by retrograde cell-labeling with horseradish peroxidase. Brain Res. 152, 249-264. Beckstead R. M. (1984) The thalamostriatal projection in the cat. J. camp. Nearol. 223, 313-346. Bentivoglio M., Macchi G. and Albanese A. (198 1) Tbe cortical projections of the thalamic intralaminar nuclei, as studied in cat and rat with the multiple fluoresamt retrograde tracing technique. Neauosci Left. 26, S-10. Bentivoglio M., Minciacchi D., Molinari M., Granato A., Spreafico R. and Mac&i R. (1988) The intrinsic and extrinsic organization of the thalamic intralaminar nuclei. In Cellular Thalamic Mechanisms (eds Bentivoglio M. and Spreafico R.), pp. 221-237. Elsevier, Amsterdam. Berendse H. W. and Groenewegen H. J. (1990) Organization of the thalamostriatal projections in the rat, with special emphasis on the ventral &at&. J. camp. Neuro?. 299, 187-228. Berendse H. W. and Groenewerren H. J. (1990) Parallel arrangement of forebrain circuits in the rat. i. Prefrontal cortex. midline and intralaminar tha&nic nuclei, aid ventral striafum. Sot. Neurosci. Abstr. 16, 426. Berendse H. W., Voom P., te Kortschot A. and Groenewegen H. J. (1988) Nuclear origin of thalamic afferents of the ventral striatum determines their relation to patch/matrix configurations in enkephalin-immunoreactivity in the rat. J. &em. Neuroanat. 1, 3-10. Bolam J. P., Izzo P. N. and Graybiel A. M. (1988) Cellular substrate of the histochemically defined striosome/matrix system of the caudate nucleus: a combined Golgi and immunocytochemical study in cat and ferret. Neuroscience 24, 853-875. Brysch I., Brysch W., Creutzfeldt O., Hayes N. L. and Schlingensiepen K.-H. (1984) The second, intralaminar thalamo-cortical projection system. Anat. Embryoi. 169, 11 l-l 18.

98 H. W. BERENDSE and H. J. GROENEWEGEN
16. Buchanan S. L., Thompson R. H. and Powell D. A. (1989) Midline thalamic lesions enhance conditioned bradycardia and the cardiac orienting reflex in rabbits. Psychobiology 17, 300-306.
17. Bushnell M. C. and Duncan G. H. (1989) Sensory and affective aspects of pain perception: is medial thalamus restricted
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
to emotional issues? Expl Brain Res. 78, 415-418. Caan W., Perret D. I. and Rolls E. T. (1984) Responses of striatal neurons in the behaving monkey. II. Visual processing in the caudal neostriatum. Brain Res. 290, 53-65. Carey R. G., Fitzpatrick D. and Diamond I. T. (1979) Layer I of striate cortex of Tupaia gIis and Gafago senegdensis: projections from thalamus and claustrum revealed by retrograde transport of horseradish peroxidase. J. camp. Neural. 186, 393-438. Carr K. D. and Bak T. H. (1988) Medial thalamic injection of opioid agonists: p-agonist increases while K-agonist decreases stimulus thresholds for pain and reward. Brain Rex 441, 173.-184. Carve11 G. E. and Simons D. J. (1987) Thalamic and corticocortical connections of the second somatic sensory area of the mouse. J. camp. Neural. 265, 409427. Cesaro P., Nguyen-Legros J., Berger B., Alvarez C. and Albe-Fessard D. (1979) Double labelling of branched neurons in the central nervous system of the rat by retrograde axonal transport of horseradish peroxidase and iron dextran complex. Neurosci. L&t. 15, l-7. Cesaro P., Nguyen-Legros J., Pollin B. and Laplante S. (1985) Single intralaminar thalamic neurons project to cerebral cortex, striatnm and nucleus reticularis thalami. A retrograde anatomical tracing study in the rat. Brain Res. 325, 29-37. Conde F., Audinat E., Maire-Lepoivre E. and Crepe1 F. (1990) Afferent connections of the medial frontal cortex of the rat. A study using retrograde transport of fluorescent dyes. 1. Thalamic afferents. Brain Res. Bull. 24, 341-354. Cornwall J. and Phillipson 0. T. (1988) Afferent projections to the parafascicular thalamic nucleus of the rat, as shown by the retrograde transport of wheat germ agglutinin. Brain Res. Bull. 20, 139-150. Cornwall J. and Phillipson 0. T. (1988) Afferent projections to the dorsal thalamus of the rat as shown by retrograde lectin transport. II. The midline nuclei. Brain Res. Bull. 21, 147-161. Cowan W. M. and Powell T. P. S. (1955) The projection of the midline and intralaminar nuclei of the thalamus of the rabbit. J. Neural. N~ro~rg. Psychiat. 18, 266-279. Craig A. D. Jr, Linington A. J. and KnilIki K.-D. (1989) Cells of origin of spinothalamic tract projections to the medial and lateral thalamus in the cat. J. camp. Neural. x89, 568-585. Deacon T. W., Eichenbaum H., Rosenberg P. and Eckmann K. W. (1983) Afferent connections of the perirhinal cortex in the rat. J. camp. Neurol. 220, 168-190. Delacour J. (1971) Effects of medial thalamic lesions in the rat. A review and an interpretation. Neuropsychologia 9, 157-174. Dempsey E. W. and Morison R. S. (1942) The production of rhy~mically recurrent cortical potentials after localized thalamic stimulation. Am. J. Physiol. 135, 293-300. Divac I., Markowitsch H. J. and Pritzel M. (1978) Behavioral and anatomical consequences of small intrastriatal injections of kainic acid in the rat. Brain Res. 151, 523-532. Divac I., Kosmal A., Bjljrklund A. and Lindvall 0. (1978) Subcortical projections to the prefrontal cortex in the rat as revealed by the horseradish peroxidase technique. Neuroscience 3, 785-796. Dong W. K., Ryu H. and Wagman 1. H. (1978) Nociceptive responses of neurons in medial thalamus and their relationship to spinothalamic pathways. J. Neurophysiol. 41, 1592-1613. Donoghue J. P., Kerman K. L. and Ebner F. F. (1979) Evidence for two organizational plans within the somatic sensory-motor cortex of the rat. .I. camp. Neural. 183, 647-664. Donoghue J. P. and Parham C. (1983) Afferent connections of the lateral agranular field of the rat motor cortex. J. camp. Neural. 217, 390-404. Donoghue J. P. and Wise S. P. (1982) The motor cortex of the rat: cytoarchitecture and microstimulation mapping. J. camp. Neural. 212, 76688. Droogleever-Fortuyn J. (1950) On the configuration and the connections of the medioventral area and tlte midline cells in the thalamus of the rabbit. Folia psychiat. neuroi. neurochir. need. 53, 213-254. Dunnett S. B. and Iversen S. D. (1980) Regulatory impai~ents following selective kainic acid lesions of the neostriatum. Behav. Brain Res. I, 497-506. Dunnett S. B. and Iversen S. D. (1981) Learning impairments following selective kainic acid-induced lesions within the neostriatum of rats. Behav. Brain Res. 2, 189-209. Dunnett S. B. and Iversen S. D. (1982) Neurotoxic lesions of ventrolateral but not anteromedial neostriatum impair differential reinforcement of low rates (DRL) performance. Behav. Bruin Res. 6, 213-226. Dunnett S. B. and Iversen S. D. (1982) Sensorimotor impairments following localized kainic acid and 6-hydroxy- dopamine lesions of the neostriatnm. Brain Res. 248, 121-127. Friedman R. P., Bachevalier J., Ungerleider L. G. and Mishkin M. (1987) Widespread projections to layer I of primate cortex. Sot. Neurosci. Abstr. 13, 251. Gerebtzoff M. A. (1937) Systdmatisation des connexions thalamo-corticales, des aires frontales, centrales et parietales. La Cellule 46, l-53. Gerfen C. R. (1984) The neostriatal mosaic: compartmentalization of corticostriatal input and striatonigral output systems. Nature 311, 461-464. Gerfen C. R. (1985) The neostriatal mosaic. I. Compartmental organization of projections from the striatum to the substantia nigra in the rat. J. camp. Neurof. 236, 454-476. Gerfen C. R. (1989) The neostriatal mosaic: striatal patch-matrix organization is related to cortical lamination. Science 246, 385-388. Gerfen C. R. and Clavier R. M. (1979) Neural inputs to the prefrontal agranular insular cortex in the rat: horseradish peroxidase study. Brain Res. Bull. 4, 347-353. Gerfen C. R. and Sawchenko P. E. (1984) An anterograde neuroanatomical tracing method that shows the detailed morphology of neurons, their axons and terminals: immunohistochemical localization of an axonally transported plant l&in, Phaseolus v~lgar~s-leucoa~lutinin (PHA-L). Brain Res. 290, 219-238.

Thalamocortical projections 99
50. Giesler G. J. Jr, Men6trey D. and Basbaum A. I. (1979) Differential origins of spinothalamic tract projections to medial and lateral thalamus in the rat. J. camp. Neural. 184, 107-126.
51. Giesler G. J. Jr, Yezierski R. P., Gerhart K. D. and Willis W. D. (1981) Spinothalamic tract neurons that project to medial and/or lateral thalamic nuclei: evidence for a physiolo~c~ly novel population of spinal cord neurons. J. NeurophysioZ. 46, 1285-1308.
52. Glenn L. L. and Steriade M. (1982) Discharge rate and excitability of cortically projecting intralaminar thalamic neurons during waking and sleep states. J. Neurosci. 2, 1387-1404.
53. Graybiel A. M. (1984) Neurochemically specified subsystems in the basal ganglia. In Functions of the Basal GangZiu (eds Evered D. and O’Connor M.), Ciba Foundation Symposium 107, pp. 114-149. Pitman, London.
54. Graybiel A. M. (1990) Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci. 13, 244-254. 55. Groenewegen H. J. (1988) Organization of the tierent connections of the mediodorsal thalamic nucleus in the rat,
related to the medi~or~-prefrontal topo~aphy. Neuroscience 24 379-431. 56. Groenewegen H. J. and Berendse H. W. (1990) Connections of the sub~al~c nucleus with ventral st~atop~li~l
parts of the basal ganglia in the rat. J. camp. Neural. 294, 607-622. 57. Groenewegen H. J., Vermeulen-Van der Zee EL, te Kortschot A. and Witter M. P. (1987) Organization of the
projections from the subiculum to the ventral striatum in the rat. A study using anterograde transport of Phaseolus vulgaris-leucoagglutinin. Neuroscience 23, 103-120.
58. Groenewegen H. J., Meredith G. E., Berendse H. W., Voorn P. and Wolters J. G. (1989) The compartmental organization of the ventral striatum in the rat. In Neural Mechanisms in Disorders of Movement (eds Crossman A. R. and Sambrook M. A.), Current Problems in Neurology, Vol. 9, pp. 45-54. John Libbey, London.
59. Groenewegen H. J., Berendse H. W., Meredith G. E., Haber S. N,, Voorn P., Walters J. G. and Lohman A. H. M. (1991) Functional anatomy of the ventral, limbic system-innervated striatum. In The Mesolimbic Dopamine System: from Motivation to Action (eds Willner P. and Scheel-Kriiger J.), pp. 19.-59. John Wiley, Chichester.
60. Groenewegen H. J., Berendse H. W., Wolters J. G. and Lohman A. H. M. (1990) The anatomical relationship of the prefrontal cortex with the striatopallidal system, the thalamus and the amygdala: evidence for a parallel organization. In The Prefrontai Cortex: Its Structure, Function, and Pathology (eds Uylings H. B. M., Van Eden C. G., De Bruin J. P. C., Corner M. A. and Feenstra M. G. P.), Prog. Brain Res., Vol. 85, pp. 95-118, Elsevier, Amsterdam.
61. Grunwerg B. S. and Kmu~~er G. M. (1990) Vibris~-r~~nsive neurons of the su~~or,co~ic~us that project to the intralaminar thalamus of the rat. Neurosci. I&t. 111, 23-27.
62. Guberman A. and Stuss D. (1983) The syndrome of bilateral paramedian thalamic infarction. Neurology 33,540-546. 63. Guldin W. 0. and Markowitsch H. J. (1983) Cortical and thalamic afferent connections of the insular and adjacent
cortex of the rat. J. camp. Neural. 215, 135-153. 64. Hanbery J., Ajmone-Marsan C. and Dilworth M. (1954) Pathways of non-specific thalamo-cortical projection system.
Electroenceph. c&n. Neurophysiol. 6, 103-I 18. 65. Hendry S. H. C., Jones E. G. and Graham J. (1979) Thalamic relay nuclei for cerebellar and certain related fiber
systems in the cat. J. camp. Neural. 185, 679-714. 66. Herkenham M. (1978) The connections of the nucleus reuniens thalami: evidence for a direct thalamo-hippocampal
pathway in the rat. J. camp. Neural. 177, 589-610. 67. Herkenham M. (1980) Laminar organization of the thalamic projections to the rat neocortex. Science 207, 532-535. 68. Herkenham M., Moon Edley S. and Stuart J. (1984) Cell clusters in the nucleus accumbens of the rat, and the mosaic
relationship of opiate receptors, acetylcholinesterase and subcortical afferent terminations. Neuroscience 11, 561-593. 69. Herkenham M. (1986) New perspectives on the organization and evolution of nonspecific thalamocortical projections.
In Cerebral Cortex Vol. 5, Sensory-motor Areas and Aspects of Cortical connectivity feds Jones E. G. and Peters A.), pp. 403-445. Plenum Press, New York.
70. Hunsperger R. W. and Roman D. (1976) The integrative role of the intralaminar system of the thalamus in visual orientation and perception in the cat. Expr Brain Res. 25, 231-246.
71. Insausti R., Amaral D. G. and Cowan W. M. (1987) The entorhinal cortex of the monkev. III. Subcortical afferents. J. camp. Neural. 264, 396-408.
72. Itoh K. and Mizuno N. (1977) Topographical arrangement of thalamocortical neurons in the centrolateral nucleus (CL) of the cat, with special reference to a spine-th~amo-motor cortical path through the CL. Expr Brain Res. 30, 471~480.
73. Jasper H. H. (1949) Diffuse projection systems: the integrative action of the thalamic reticular system. Efectroenceph. clin. Neurophysiol. 1, 405-420.
74. Jasper H. H. (1960) Unspecific thalamocortical relations. In Handbook of Physiology, Section I: Neurophysiology (eds Field J., Magoun H. W. and Hall V. E.), Vol. II, pp. 1307-1321. American Physiological Society, Washington, DC.
75. Jayaraman A. (1985) Organization of thalamic projections in the nucleus accumbens and the caudate nucleus in cats and its relation with hip~m~l and other subcortical afferents. J. camp. Neurof. 231, 396-420.
76. Jinnai K. and Matsuda Y. (1981) ~alam~audate projection neurons with a branching axon to the cerebral motor cortex. Neurosci. Lett. 26, 95-99.
77. Jones E. G. (1985) The Thalamus. Plenum Press, New York. 78. Jones E. G. and Leavitt R. Y. (1974) Retrograde axonal transport and the demonstration of non-specific projections
to the cerebral cortex and striatum from thalamic intralaminar nuclei in the rat, cat and monkey. J. camp. Neural. 154, 349-378.
79. Jones E. G., Wise S. P. and Coulter J. D. (1979) Differential thalamic relationships of sensory-motor and parietal cortical fields in monkeys. J. camp. Neural. 183, 833-882.
80. Kaufman E. F. S. and Rosenquist A. C. (1985) Efferent projections of the thalamic intralaminar nuclei in the cat. Brain Res. 335, 257-279.
81. Kaufman E. F. S. and Rosenquist A. C. (1985) Afferent connections of the thalamic intralaminar nuclei in the cat. Brain Res. 335, 281-296.
82. Kemp J. M. and Powell T. P. S. (1970) The cortico-striate projection in the monkey. Brain 93, 525-546. 83. Kievit J. and Kuypers H. G. J. M. (1975) Subcortical afferents to the frontal lobe in the rhesus monkey studied by
means of retrograde horseradish peroxidase transport. Bruin Res. 85, 261-266.

100 H. W. BERENDSE and H. J. GRO~~WEGEN
84. Krettek J. E. and Price J. L. (1977) The cortical projections of the mediodorsal nucleus and adjacent thalamic nuclei in the rat. J. camp. Neurol. 171, 157-192.
85. Laplane D., Escourolle R., Degos J. D., Sauron B. and Massiou H. (1982) La n&ligence motrice d’origine thalamique. Rev. Neural., Paris 138, 201-211.
86. Lorente de No R. (1938) The cerebral cortex: architecture, intracortical connections and motor projections. In Physiology of the Nervous System (ed. Fulton J. F.), pp. 291-325. Oxford University Press, New York.
87. Macchi G. and Bentivoglio M. (1982) The organization of the efferent projections of the thalamic intralaminar nuclei: past, present and future of the anatomical approach. Ital. J. neurol. Sci. 2, 83-96.
88. Macchi G. and Bentivoglio M. (1986) The thalamic intralaminar nuclei and the cerebral cortex. In Cerebral Cortex. Vol. 5, Sensory-motor Areas and Aspects of Cortical Connectivity (eds Jones E. G. and Peters A.), pp. 355-401. Plenum Press, New York.
89. Macchi G., Bentivoglio M., D’Atena C., Rossini P. and Tempesta E. (1977) The cortical projections of the thalamic intralaminar nuclei restudied by means of the HRP retrograde axonal transport. Neurosci. Left. 4, 121-126.
90. Macchi G., Bentivoglio M., Molinari M. and Minciacchi D. (1984) The thalamo-caudate versus thalamo-cortical projections as studied in the cat with fluorescent retrograde double labeling. ExpZ Bruin Res. Ss, 225-239.
91. Mantyh P. W. (1983) The spinothal~c tract in the primate: a reexamination using wheatgerm agglutinin conjugated to horseradish peroxidase. ~euros~jen~e 9, %7-862.
92. Matelli M., Luppino G., Fitzpatrick D. and Diamond I. T. (1988) The pulvinar nucleus of Tupaiu: comparative study of its connections with the superior colliculus, the neocortex and the corpus striatum. In Cellular Thalamic Mechanisms (eds Bentivoglio M. and Spreafico R.), pp. 2077220. Elsevier, Amsterdam.
93. McClung R. E. and Dafny N. (1980) The parafascicular nucleus of thalamus exhibits convergence input from the dorsal raphe and the spinal tract of the trigeminal nerve. Bruin Res. 197, 525-531.
94. McGeorge A. J. and Faull R. L. M. (1989) The organization of the projection from the cerebral cortex to the striatum in the rat. Neuroscience 29, 503-537.
95. Melzack R. and Casey K. L. (1968) Sensory, motivational, and central control determinants of pain. In The Skin Senses (ed. Kenshalo D.), pp. 423-439. Thomas, Springfield, IL.
96. Minciacchi D. and Granato A, (1989) Development of the thalamocortical system: transient-crossed projections to the frontal cortex in neonatal rats. J. camp. Neuroi. 281, l-12.
97. Morison R. S. and Dempsey E. W. (1942) A study of thalamo-corticai relations. Am. J. Physioi. 135, 281-292. 98. Moruzzi G. and Magoun H. W. (1949) Brain stem reticular formation and activation of the EEG. Electroenceph. clin.
N~rophys~o~. 1, 455-473. 99. Murray M. (1966) Degeneration of some in~alaminar thalamic nuclei after cortical removals in the cat. J. camp.
NeuroL 127, 341-368. 100. Musil S. Y. and Olson C. R. (1988) Organization of cortical and subcortical projections to anterior cingulate cortex
in the cat. J. camp. Neural. 272, 203-218. 101. Musil S. Y. and Olson C. R. (1988) Organization of cortical and subcortical projections to medial prefrontal cortex
in the cat. .I. camp. Neural. 272, 219-241. 102. Nahin R. L. (1988) Immunocytochemical identification of long ascending, peptidergic lumbar spinal neurons
terminating in either the medial or lateral thalamus in the rat. Brain Res. 442, 345-349. 103. Oberg R. G. E. and Divac I. (1979) “Cognitive” functions of the neostriatum. In The Neostriarum (eds Divac I. and
Oberg R. G. E.), pp. 291-313. Pergamon Press, Oxford. 104. Ohtake T. and Yamada H. (1989) Efferent connections of the nucleus reuniens and the rhomboid nucleus in the rat:
an anterograde PHA-L tracing study. Neurosci. Res. 6, 556-568. 105. Olausson B., Shyu B. C. and Rydenhag B. (1989) Projection from the thalamic intralaminar nuclei on the isocortex
of the rat: a surface potential study. Expl Brain. Res. 75, 543-5.54. 106. Orem J., Schlag-Rey M. and Schlag J. (1973) Unilateral visual neglect and thalamic intralaminar lesions in the cat.
Expi Neural. 40, 784-797. 107, Paxinos G. and Watson C. (1986) The Rat Brain in Siereoraxic Coordinates, 2nd edn. Academic Press, Sydney. 108. Peschanski M., Guilbaud G. and Gautfon M. (1981) Posterior intralaminar region in rat: neuronal responses to
noxious and non-noxious cutaneous stimuli. Expi Neural. 72, 226-238. 109. Pisa M. (1988) Motor somatotopy in the striatum of rat: manipulation, biting and gait. Behav. Brain Res. 27, l-35. 110. Pisa M. and Cyr J. (1990) Regionally selective roles of the rat’s striatum in modality-specific discrimination learning
and forelimb reaching. Behav. Brain Res. 37, 281-292. 111. Powell T. P. S. and Cowan W. M. (1954) The connexions of the midline and intralaminar nuclei of the thalamus of
the rat. J. Anal. 88, 307-319. 112. Preuss T. M. and Goldman-Rakic P. S. (1987) Crossed corticothalamic and thalamocortical connections of macaque
prefrontal cortex. J. camp. Neural. 257, 269-281. 113. Prieto-Gomez B., Dafny N. and Reyes-Vgzquez C. (1989) Dorsal raphe stimulation, 5-HT and morphine micro-
iontophoresis effects on noxious and non-noxious identified neurons in the medial thalamus of the rat. Brnin Res. Bull. 22, 937-943.
114. Reep R. L. and Winans S. S. (1982) Affefent connections of dorsal and ventral agranular insular cortex in the hamster ~eso~r~ce~us mater. Neuroscience 7, 12651288.
IIS. Reep R. L., Corwin J. V., Hashimoto A. and Watson R. T. (1984) Afferent connections of medial precentral cortex in the rat. Neurosci. Left. 44, 247-252.
116. Reep R. L., Goodwin G. S. and Corwin J. V. (1990) Topographic organization in the corticocortical connections of medial agranular cortex in rats. J. camp. Neuroi. 294, 262-280.
117. Rieck R. W. and Carey R. G. (1982) Ventral thalamic projections to layer I of parietal cortex. Anal. Rec. 202, 157-158. 118. Rieck R. W. and Carey R. G. (1985) Organization of the rostra1 thalamus in the rat: evidence for connections to layer
I of visual cortex. J. Eomp. Neural: 234; 137-154. 119. Rilev J. N. and Moore R. Y. (1981) Diencenhalic and brainstem afferents to the hippocamoal formation of the rat.
Brain Res. Bull. 6, 437-444. . , _ __ _
120. Robertson R. T. and Kaitz S. S. (1981) Thalamic connections with hmbic cortex. I. Thalamocortical projections. J. camp. Neural. 195, 501-525.

Thalamocortical projections 101
121. Rolls E. T. and Williams G. V. (1987) Sensory and movement-related neuronal activity in different regions of the primate striatum. In Basal Ganglia and Behavior: Sensory Aspects of Motor Functioning (eds Schneider J. S. and Lidsky T. I.), pp. 37-59. Hans Huber, Toronto.
122. Rolls E. T., Thorpe S. J. and Maddison S. P. (1983) Responses of striatal neurons in the behaving monkey. I. Head of the caudate nucleus. Behuu. Brain Res. 7, 179-210.
123. Royce G. J. (1982) Laminar origin of cortical neurons which project upon the caudate nucleus: a horseradish peroxidase investigation in the cat. J. camp. Neurol. UM, 8-29.
124. Royce G. J. (1983) Single thalamic neurons which project to both the rostra1 cortex and caudate nucleus studied with the fluorescent double labeling method. Expl Neural. 79, 773-784.
125. Royce G. J. (1988) PHA-L study of thalamocortical fibres of the central lateral nucleus. Sot. Neurosci. Abstr. 14, 1028. 126. Royce G. J. and Mourey R. J. (1985) Efferent connections of the centromedian and parafascicular thalamic nuclei:
an autoradiographic investigation in the cat. J. camp. Neural. 235, 277-300. 127. Royce G. J., Bromley S., Gracco C. and Beckstead R. M. (1989) Thalamocortical connections of the rostra1
intralaminar nuclei: an autoradiographic analysis in the cat. J. camp. Neural. 288, 555-582. 128. Rydenhag B., Olausson B., Shyu B. C. and Andersson S. (1986) Localized responses in the midsuprasylvian gyrus
of the cat following stimulation of the central lateral nucleus in thalamus. Expl Brain Res. 62, 11-24. 129. Sadikot A. F., Parent A. and Francois C. (1990) The centre median and parafascicular thalamic nuclei project
respectively to the sensorimotor and associative-limbic striatal territories in the squirrel monkey. Brain Res. 510, 161-165.
130. Saper C. B. (1982) Convergence of autonomic and limbic connections in the insular cortex of the rat. J. camp. Neural. 210, 163-173.
131. Scheibel M. E. and Scheibel A. B. (1967) Structural organization of nonspecific thalamic nuclei and their projection toward cortex. Brain Res. 6, 60-94.
132. Schell G. R. and Strick P. L. (1984) The origin of thalamic inputs to the arcuate premotor and supplementary motor areas. J. Neurosci. 4, 539-560.
133. Schlag J. and Schlag-Rey M. (1984) Visuomotor functions of central thalamus in monkey. II. Unit activity related to visual events, targeting, and fixation. J. Neurophysiol. 51, 1175-l 195.
134. Schlag J., S&lag-Rey M., Peck C. K. and Joseph J.-P,. (1980) Visual responses of thalamic neurons depending on the direction of gaze and the position of targets in spa&. Expl Brain Res. 40, 170-184.
135. Schlag-Rey M. and S&lag J. (1984) Visuomotor functions of central thalamus in monkey. I. Unit activity related to spontaneous eye movements. J. Neurophysiol. 51, 1149-l 174.
136. Schott B., Mauguiere F., Laurent B., Serclerat 0. and Fischer C. (1980) L’Amnesie thalamique. Rev. Neural., Paris 136, 117-132.
137. Segal M. (1977) Afferents to the entorhinal cortex of the rat studied by the method of retrograde transport of horseradish peroxidase. Expl Neural. 57, 750-765.
138. Selemon L. D. and Goldman-Rakic P. S. (1985) Longitudinal topography and interdigitation of corticostriatal projections in the rhesus monkey. J. Neurosci. 5, 776-794.
139. Steriade M. and Glenn L. L. (1982) Neocortical and caudate projections of intralaminar thalamic neurons and their synaptic excitation from midbrain reticular core. J. Neurophysiol. 48, 352-371.
140. Su H.-S. and Bentivoglio M. (1990) Thalamic midline cell populations projecting to the nucleus accumbens, amygdala, and hippocampus in the rat. J. camp. Neural. 297, 582-593.
141. Sugimoto T., Hattori T., Mizuno N., Itoh K. and Sato M. (1983) Direct projections from the centre median- parafascicular complex to the subthalamic nucleus in the cat and rat. J. camp. Neural. 214, 209-216.
142. Takada M., Fishell G., Li Z. K., Van der Kooy D. and Hattori T. (1987) The development of laterality in the forebrain projections of midline thalamic cell groups in the rat. Deul Brain Res. 35, 275-282.
143. Thompson S. M. and Robertson T. (1987) Organization of subcortical pathways for sensory projections to the limbic cortex. I. Subcortical projections to the medial limbic cortex in the rat. J. camp. Neural. 265, 175-188.
144. Ullln J. (1985) Cortical topography of thalamic intralaminar nuclei. Brain Res. 328, 333-340. 145. Van Eden C. G. and Uylings H. B. M. (1985) Cytoarchitectonic development of the prefrontal cortex in the rat.
J. camp. Neural. 2.41, 253-267. 146. Vogt B. A., Pandya D. N. and Rosene D. G. (1987) Cingulate cortex of the rhesus monkey: I. Cytoarchitecture and
thalamic afferents. J. camp. Neural. 262, 256-270. 147. Voorn P., Gerfen C. R. and Groenewegen H. J. (1989) Compartmental organization of the ventral striatum in the
rat: immunohistochemical distribution of enkephalin, substance P, dopamine, and calcium-binding protein. J. camp. Neurol. 289, 1899201.
148. Watson R. T. and Heilman K. M. (1979) Thalamic neglect. Neurology 29, 690-694. 149. Watson R. T., Miller B. and Heilman K. M. (1978) Nonsensory neglect. Ann. Neurol. 3, 505-508. 150. Watson R. T., Valenstein E. and Heilman K. M. (1981) Thalamic neglect. Possible role of the medial thalamus and
nucleus reticularis in behavior. Archs Neural. 38, 501-506. 151. Webster K. E. (1961) Cortico-striate interrelations in the albino rat. J. Anaf. 95, 532-544. 152. Willis W. D., Kenshalo D. R. Jr and Leonard R. B. (1979) The cells of origin of the primate spinothalamic tract.
J. camp. Neural. 188, 543-574. 153. Wouterlood F. G., Saldana E. and Witter M. P. (1990) Projection from the nucleus reuniens thalami to the
hippocampal region: light and electron microscopic tracing study in the rat with the anterograde tracer Phaseolus vulgaris-leucoagglutinin. J. camp. Neural. 296, 179-203.
154. Wyss J. M., Swanson L. W. and Cowan W. M. (1979) A study of subcortical afferents to the hippocampal formation in the rat. Neuroscience 4, 463-476.
155. Yanagihara M., Niimi K. and Ono K. (1987) Thalamic projections to the hippocampal and entorhinal areas in the cat. J. camp. Neurol. 266, 122-141.
156. Yasui Y., Itoh K., Kamiya H., Ino T. and Mizuno N. (1988) Cingulate gyrus of the cat receives projection fibers from the thalamic region ventral to the ventral border of the ventrobasal complex. J. camp. Neural. 274, 91-100.
157. Yeterian E. H. and Van Hoesen G. W. (1978) Cortico-striate projections in the rhesus monkey: the organization of certain cortico-caudate connections. Brain Res. 139, 43-63.

102 H. W. BERENDSE and H. J. GROENEWEGEN
158. Yeung J. C., Yaksh T. L. and Rudy T. A. (1978) Effect on the nociceptive threshold and EEG activity in the rat of morphine injected into the medial thalamus and the periaqueductal gray. Neuropharmacology 17, 525-532.
159. Zilles K. and Wree A. (1985) Cortex: area1 and laminar structure. In The Rut Nervous System Vol. I, Forebrain and Midbrain (ed. Paxinos G.), pp. 375-415. Academic Press, Sydney.
(Accepted 3 1 October 1990)
Copyright © 2022 FDOKUMEN




















