
Mapping resting-state functional connectivity using perfusion MRI

NeuroImage 56 (2011) 1546ndash1555
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage wwwe lsev ie rcom locate yn img
Resting-state networks in the macaque at 7 T
R Matthew Hutchison abd L Stan Leung ab Seyed M Mirsattari acd Joseph S Gati dRavi S Menon ad Stefan Everling abda Graduate Program in Neuroscience University of Western Ontario London Ontario Canadab Department of Physiology and Pharmacology University of Western Ontario London Ontario Canadac Department of Clinical Neurological Sciences University of Western Ontario London Ontario Canadad Robarts Research Institute London Ontario Canada
Grant sponsor Canadian Institutes of Health Resea Corresponding author at The Centre for Brain andM
100 Perth Drive London Ontario Canada N6A 5K8 FaxE-mail address severlinuwoca (S Everling)
1053-8119$ ndash see front matter copy 2011 Elsevier Inc Aldoi101016jneuroimage201102063
a b s t r a c t
a r t i c l e i n f o
Article historyReceived 28 October 2010Revised 18 February 2011Accepted 21 February 2011Available online 26 February 2011
KeywordsFunctional connectivityMacaqueResting-stateIndependent component analysis (ICA)Spontaneous activityFunctional MRI (fMRI)
Assessment of brain connectivity has revealed that the structure and dynamics of large-scale networkorganization are altered in multiple disease states suggesting their use as diagnostic or prognostic indicatorsFurther investigation into the underlying mechanisms organization and alteration of large-scale brainnetworks requires a homologous animal model that would allow neurophysiological recordings andexperimental manipulations The current study presents a comprehensive assessment of macaque resting-state networks based on evaluation of intrinsic low-frequency fluctuations of the blood oxygen-level-dependent signal using group independent component analysis Networks were found underlying multiplelevels of sensory motor and cognitive processing The results demonstrate that macaques share remarkablehomologous network organization with humans thereby providing strong support for their use as an animalmodel in the study of normal and abnormal brain connectivity as well as aiding the interpretation ofelectrophysiological recordings within the context of large-scale brain networks
rch (CIHR)ind Robarts Research Institute +1 519 931 5233
l rights reserved
copy 2011 Elsevier Inc All rights reserved
Introduction
The mammalian brain is composed of functional networksoperating at different spatial and temporal scales mdash characterized bypatterns of interconnections linking sensory motor and cognitivesystems (Felleman and Van Essen 1991 Young 1993 Friston 2002)Neuroimaging has afforded unparalleled access in the exploration ofthe topology of these systems and has revealed that neural processingrelies on the dynamic integration of cortical and subcortical areaswithin large-scale and distributed brain networks (Sporns et al 2005Guye et al 2008) Network activity is typically assessed usingfunctional connectivity measures This is an examination of temporalcorrelations that exist between distinct brain areas (Friston 1994)connected directly or indirectly by long-range cortical and subcorticalpolysynaptic pathways (Hagmann et al 2008) Using this definitionfunctional connectivity has been derived using spontaneous bloodoxygenation-level-dependent (BOLD) fluctuations measured by func-tional magnetic resonance imaging (fMRI Biswal et al 1995)Correlation of low frequency fluctuations (LFFs 001ndash01 Hz) of theBOLD signal acquired in the absence of a task has been shown to
reflect anatomical connectivity (Vincent et al 2007 Honey et al2009 Greicius et al 2009) and presumed to be a hemodynamicmanifestation of functional connectivity between slow fluctuations inneuronal activity (Fox and Raichle 2007 Shmuel and Leopold 2008)
Investigations of functional connectivity through the evaluation ofLFF synchrony during rest have demonstrated that the human brain isspatially organized into coherent patterns characterized as networksThese robust and reproducible resting-state networks (RSNs) havebeen reported for visual motor auditory language memoryexecutive and attention systems as well as the default-modenetwork (Cordes et al 2000 Raichle et al 2001 Hampson et al2002 Beckmann et al 2005 Damoiseaux et al 2006) RSNs parallelpreviously identified task-based networks and spatio-temporalnetwork synchronization is preserved during sedation and anesthesiain humans monkeys and rats (Kiviniemi et al 2005 Vincent et al2007 Hutchison et al 2010) RSNs can be identified through a seed-region analysis in which spatial functional connectivity maps areinferred by a cross-correlation analysis of the voxel-wise fMRIrecordings against a reference time-course The shortcoming of thistechnique is that it tests a specific hypothesis and is limited to thoseareas that are selected as seed-regions (Cole et al 2010) Owing to theconstraints of seed-region analysis exploratory techniques such asindependent component analysis (ICA) are now frequently applied tohuman functional data sets where they have revealed RSNs that hadnot been previously shown with seed-region techniques (Beckmannet al 2005 Smith et al 2009)
1547RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Although their meaning is not fully understood changes infunctional RSNs have been recently reported in several psychiatricand developmental disorders including depression attention deficit-hyperactivity disorder schizophrenia Alzheimers disease epilepsyand multiple sclerosis (Auer 2008 Greicius 2008) Given theextraordinary potential for RSNs as possible diagnostic or prognosticmarkers it is crucial to understand the physiological mechanisms offluctuation regulation and entrainment of LFFs and the RSNs that arerevealed through their synchronization
Nonhuman primates and in particular macaque monkeys havebeen used as surrogates for the study of human brain function forseveral decades and might therefore represent an ideal animal modelfor the study of RSNs Although macaque and human brains share ahigh degree of similarity in terms of cytoarchitecture (Petrides andPandya 1999 2002a Onguumlr et al 2003) functional organization(Rees et al 2000 Koyama et al 2004 Petrides et al 2005 Nakaharaet al 2007) and anatomical connections (Croxson et al 2005 Kellyet al 2010) there also exist structural (Preuss 2000 Rilling 2006)morphological (Buxhoeveden et al 2001) and functional (Orbanet al 2004 Preuss 2004) differences between the brains of these twoprimate species (Passingham 2009)
While a few recent studies have revealed homologous RSNsbetween human and nonhuman primates these studies eitherutilized seed-region analysis (Vincent et al 2007 2010 Margulieset al 2009) or ICA in only two animals (Moeller et al 2009) Here wepresent a comprehensive evaluation of macaque RSNs at 7 T usinggroup ICA and an analysis methodology that is very similar to whathas been used in humans The results show striking similarity ofmacaque RSNs to previously described human RSNs
Material and methods
Animal preparation
Data was obtained from 6macaque monkeys (Macaca fascicularis 2male 4 female)whoseweights ranged from36 kg to 53 kg (mean+minusSD=458+minus14 kg) Surgical and experimental procedures werecarried out in accordance with the Canadian Council of Animal Carepolicy on the use of laboratory animals and approved by the Animal UseSubcommittee of the University of Western Ontario Council on AnimalCare
Animals were initially prepared for imaging by undergoing asurgical procedure to place an MRI-compatible custom-built acrylichead post that served to restrain the head during image acquisitionThe post was anchored to the skull with 6 mm ceramic bone screws(Thomas Recording Giessen Germany) and dental acrylic
On the day of scanning anesthesia was first induced by intramus-cular injections of atropine (04 mgkg) ipratropium (0025 mgkg)and ketamine hydrochloride (75 mgkg) Animals were then adminis-tered3 mlof propofol (10 mgml) intravenouslyvia the saphenousveinFollowing oral intubation with an endotracheal tube anesthesia wasmaintained using 15 isoflurane mixed with oxygen Animals werespontaneously breathing throughout the duration of the experimentThe animal was placed in a custom-built primate chair containingfixation for head immobilization and an integrated custom RF coil andinserted into the bore for image acquisition The isoflurane level wasthen lowered to 1 for imaging experiments Rectal temperature via afiber-optic temperature probe (FISO Quebec City QC) respiration viabellows (Siemens Corp Union NJ) and end-tidal CO2 via capnometer(Covidien-Nellcor Boulder CO) were continuously monitored Physio-logical parameters were in the normal range (temperature 365 degCbreathing 25ndash30 breathsmin end-tidal CO2 24ndash28 mmHg) through-out the duration of the experiment Warmth was maintained using aheatingdisk (Snugglesafe LittlehamptonWest SussexUK) and thermalinsulation
Anesthesia was utilized in this study to eliminate motion effectsphysiological stress and training requirements Although isofluranehas been shown to have vasodilator properties (Farber et al 1997)altering cerebrovascular activity in a dose-dependent manner(Vincent et al 2007) synchronous spontaneous BOLD fluctuationshave been previously reported using an isoflurane regime in bothmonkeys (Vincent et al 2007) and rats (Hutchison et al 2010)However it is still likely that anesthesia can affect RSNs
Data acquisition
All data were acquired on an actively shielded 7 Tesla 68 cmhorizontal borehumanhead scannerwith aDirectDrive console (VarianYarnton UK Walnut Creek CA) and a Siemens AC84 gradient sub-system (Erlangen Germany) operating at a slew rate of 350mTms Anin-house designed and manufactured conformal 5 channel transceiveprimate head RF coil was used for all experiments Magnetic fieldoptimization (B0 shimming)wasperformedusing anautomated three-dimensional mapping procedure over the specific imaging volume ofinterest For each monkey 2 runs of 300 continuous EPI functionalvolumes (TR=2000 ms TE=16 ms flip angle=70deg slices=30matrix=72times72 FOV=96times96 mm acquisition voxel size=13times13times15 mm) were acquired The total acquisition time of each scanwas 10 min EPI images were acquired with GRAPPA at an accelerationfactor of 2 Every image was corrected for physiological fluctuationsusing navigator-echo-correction A high-resolution T2-weighted ana-tomical reference volume was acquired along the same orientation asthe functional images using a fast spin echo acquisition scheme(TR=5000 ms TE=386 ms echo train length=5 effectiveecho=3 slices=30 matrix=256times250 FOV=96times96 mm acquisi-tion voxel size=375 μmtimes384 μmtimes15 mm) T2 imagingusing fast spinecho as compared to gradient echo based imaging such as T1-weightedMP-RAGE reduced the amount of image distortion caused by skullimplants such as the ceramic bone screws and plastic head post
Image preprocessing
All preprocessing was implemented using the FSL softwarepackage (httpwwwfmriboxacuk) and included motion correc-tion (six parameter affine transformation) brain extraction spatialsmoothing (FWHM=3 mm) high-pass temporal filtering (Gaussian-weighted least-squares straight line fitting with sigma=100 s) low-pass temporal filtering (HWHM=28 s Gaussian filter) and normal-ization (12 DOF linear affine transformation implemented in FLIRT) tothe F99 atlas template (Van Essen 2004 see httpsumsdbwustledusumsmacaquemoredo)
Group independent component analysis
Group ICA unlike single-subject ICA allows inferences to be madeat the group level When examining subjects individually it is difficultto compare components since they are not ordered and differentcomponents may be revealed in each subject Entering all subjectsinto an ICA analysis and estimating one set of components has theadvantage of ordering the components of different subjects in thesame way This produces a single set of ldquogrouprdquo components that canthen be interpreted Additionally weak sources with differentcharacteristics across subjects (ie noise) will be suppressed allowinga more accurate reflection of population dynamics
Group ICA was conducted using the GIFT software package(Calhoun et al 2001 httpicatbsourceforgenet) Data from bothruns of all animals were concatenated and the temporal dimension ofthis aggregated data set was reduced by means of principalcomponent analysis (PCA) This was followed by spatial componentestimation using the Infomax algorithm Component time-coursesand spatial maps for each animal were then back-reconstructed using
1548 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
the aggregated components and the results from the data reductionstep (Jafri et al 2008 Calhoun et al 2001) Because ICA is a stochasticestimation process the final component maps can vary depending onthe initial algorithm conditions To quantify the reliability of thedecomposition ICA was reiterated 20 times using the ICASSO toolbox(Himberg et al 2004) ICASSO represents each of the estimatedcomponents for each iteration as a point in the signal space It thenreturns a stability index of the estimate cluster computed as thedifference between the average intra- and inter-cluster similarities Inthe ideal case the repeated estimates are concentrated in compactand close-to-orthogonal clusters resulting in an index of all estimate-clusters that approaches one
There are currently no well-established criteria to guide theselection of an optimal number of independent components (ICs) for agiven data set (Cole et al 2010) The model order or dimensionestimate defines the number of components that the algorithm willextract and in that regard represents a prediction about the number ofunderlying sources in the data If the model order is increased agreater number of networks will be found accounting for the data in amore detailed way albeit causing networks to branch into smallersub-networks (Smith et al 2009 Abou-Elseoud et al 2010) Oneproposed method uses the minimum description length criterion fordimension estimation (Jafri et al 2008 Li et al 2007) howeverwhenapplied to our data set this technique resulted in an estimation of 253ICs Instead 20 components were chosen as this gave a manageablenumber of components and approximated model orders commonlyused in human studies large-scale brain networks (Smith et al 2009Abou-Elseoud et al 2010 Calhoun et al 2001 Demirci et al 2009)
Themean group ICswere then scaled to empirically derived z-scoresby dividing by the standard deviation of the original time-course Thez-scores approximate the temporal correlation between each voxeland the associated component where the magnitude of the z-scorespecifies the strength of the linear relationship A negative z-scoreindicates voxels in which the BOLD signals are modulated opposite tothat of the IC time-course A z-score value of +minus1 was used as thelower limit threshold of functional connectivity
RSN identification and visualization
The ICA derived components were visually inspected and labeledbased on the activation patterns in reference to known anatomical andfunctional locations Classification of the components in terms of RSNswas performed by comparison with known macaque functionalnetworks and previously identified RSNs from ICA studies in humans(Beckmann et al 2005 Damoiseaux et al 2006 Smith et al 2009) Inaddition to identifying spatially independent networks ICA is able toparse out noise due to physiological fluctuations motion and hardwareartifacts from the original source data (Thomas et al 2002 Liao et al2006 Perlbarg et al 2007) Components with high spatial correspon-dence to cerebrospinal spinal fluid (see Supplementary Fig 1) or withlow correlation to gray matter were discarded
Group data were projected from volume data to the F99 corticalsurface using the CARET (httpwwwnitrcorgprojectscaret)enclosed-voxel method
Single-subject ICA
Single-subject ICA is better suited to reveal individual features ofresting-state connectivity as subtle differences between subjects mayget lost in an ICA of a group data set To examine individual subjectmaps and allow comparison with a previously published report(Moeller et al 2009) single subject-ICA was conducted using thesame parameters as the group-ICA The unthresholded group-ICAderived networks were then used as templates to order thecomponent maps from the individual ICA by calculating the spatialcorrelation coefficient of the unthresholded individual maps
Functional network connectivity
Spatial ICA maximizes the statistical independence in the spatialdomain though the components are not temporally constrained andICs can exhibit considerable temporal correlations (Calhoun et al2003) The temporal dependencies are significant albeit not as high asthose between regions within an IC (Jafri et al 2008) Functionalnetwork connectivity (FNC) analysis examines these temporaldependencies among ICs to establish the functional connectivityamong the large scale networks and has been shown to provideadditional information regarding macroscopic brain organization(Jafri et al 2008 Demirci et al 2009) FNC analysis was conductedusing the FNC software package (httpmialabmrnorgsoftware)using analysis steps described previously (Jafri et al 2008) One-sample Students t-tests were used to test statistically significantmaximal lagged correlation (+minus3 s) combinations from the 55possible combinations (pb005 corrected for multiple comparisons)
Results
Resting-state networks
Group-ICA successfully decomposed the resting-state functionaldata of 6monkeys into 20 independent components ICASSO returned astability index for each estimate-cluster that approached 1 (mean+minusSD=0980+minus0006) indicating that the components are concen-trated in compact and close-to-orthogonal clusters and highly consis-tent across multiple ICA runs Eleven components were found to have ahigh correspondence with gray matter as well as showing networkcharacteristics (ie spatial maps containing more than one functionalarea) and were thereby deemed to be physiologically relevant Thespatial maps of the RSNs obtained with ICA analysis are illustrated inFig 1 (for RSNmaps displayed on a flattened cortex see SupplementaryFig 2) The components accounted for 5981 of the datas variance Thedegree of cortical coverage is illustrated in Supplementary Fig 3 Despiteinclusion of the entire brain components were primarily restricted tocortical areas Therefore we focus on cortical connectivity in this studyOn the basis of our classification results the 11 RSNs can be described asfollows
bull RSN A (precentralndashtemporal) The RSN included extended bilateralareas of the ventral precentral gyrus corresponding to ventralpremotor areas F4 and F5 activation in the inferior ramus of thearcuate sulcus the precentral opercular cortex primary andsecondary somatosensory cortex and insular cortex In additionconnectivity was observed with the auditory cortex The networkincluded areas that correspond to the ventral motor system thoughtto play an important role in understanding action and visuomotorcontrol of grasping and also included areas underlying the control ofmouth and tongue (Nelissen et al 2005 Joly et al 2009 Phillipsand Porter 1977)
bull RSN B (fronto-parietal) Bilateral connectivity was found in theanterior bank fundus and posterior bank of the inferior arm of thearcuate sulcus The anterior extent of this region corresponds to thelocation of the frontal eye fields (Bruce and Goldberg 1985)Connectivity was found in the ventral prefrontal cortex and dorsalpremotor cortex Further network connectivity was found in area 3Bin the dorsal central sulcus fundus and in both banks of theintraparietal sulcus In addition the right hemisphere showedpositive correlation of area 6 corresponding to the supplementarymotor area or possibly the supplementary eye fields (Schlag andSchlag-Rey 1987) The network includes brain areas associatedwiththe oculomotor system (also referred to as the dorsal attentionnetwork) responsible for goal-oriented saccadic eye movements(Johnston and Everling 2008) and mediating goal-directed top-down processing (Noudoost et al 2010) The functional network
Fig 1 Cortical representation of eleven resting-state networks (RSNs) identified by independent component analysis of fMRI data acquired from isoflurane-anesthetized macaquemonkeys (N=6) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 see httpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of both hemispheres and theright images depict the lateral and medial views of right hemisphere Potential functional roles of each network are discussed in the text
1549RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
has been previously reported in eye movement tasks (Baker et al2006 Ford et al 2009) and resting-state analysis (Vincent et al2007) of the macaque
bull RSN C (posterior-parietal) The network encompassed the posteriorcingulateprecuneus cortex bilateral areas PG and PE of the parietal
cortex and visual areas V2 and V3 In addition this networkcontained connectivity patterns in the dorsolateral prefrontal cortexand in the dorsal premotor cortex in the left hemisphere The brainareas include areas found in the default-mode network (DMN) areasthat are deactivated during attention-demanding cognitive tasks
Table 1Spatial correlations of single subject ICA derived networks to group-ICA derivednetworks
Network Mean STD ERR Range
A 0691 0068 0383ndash0841B 0560 0082 0176ndash0750C 0463 0062 0227ndash0649D 0501 0078 0215ndash0743E 0373 0046 0275ndash0584F 0459 0073 0184ndash0623G 0372 0060 0200ndash0586H 0559 0059 0272ndash0661I 0501 0065 0199ndash0655J 0375 0045 0207ndash0546K 0264 0017 0186ndash0302
Note Mean correlation values are significantly different from 0 (one sample t-testpb001)
1550 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
and in humans has been suggested to support higher-order mentalfaculties (Raichle et al 2001)
bull RSN D (occipito-temporal) Network areas included bilateral area TOV4 TEO and the arcuate sulcus Bilateral area 29 and Brodmann area10 (left hemisphere) were also functionally connected Previousstudies have shown these temporal lobe areas are critical for higher-order visual processing (Tsao et al 2003)
bull RSN E (frontal) This RSN encompassed bilateral regions in severalprefrontal areas including the anterior bank of the arcuate sulcuscorresponding to the frontal eye fields the upper ramus of thearcuate sulcus the posterior portion of the principal sulcus as wellas the dorsal bank of the principal sulcus In addition LFFsynchronization was also found in area 9 the premotor cortex andthe anterior cingulate cortex These frontalprefrontal areas havebeen shown to be components of the executive system suggested toprovide bias signals to other areas of the brain in order to implementcognitive control (Miller and Cohen 2001)
bull RSN F (superior-temporal) A network encompassing the auditory beltparabelt and bilateral area 22 on the superior temporal gyrus Anti-correlated areas were found in the left principal sulcus and in the leftarcuate sulcus The network pattern resembles the mean functionalactivity resulting from the presentation of multiple sounds categoriesin a previous task-based fMRI study of the awake monkey (Petkovet al 2008) and is likely responsible for acoustic processing andinterpretation (Rauschecker and Scott 2009)
bull RSN G (cingulo-insular) A network of areas including bilateral regionsin the insular cortex the anterior cingulate cortex area 24ab and theorbitofrontal cortex area 14 These areas have been associated withthe reward system involved in the regulation and control of behavior(Kringelbach 2005)
bull RSN H (paracentral) A network reflective of the dorsal motorsystem involved in the control of limb movements (Dum and Strick2002) Network connectivity included bilateral primary motorcortex dorsally and also in the central sulcus area F2 and thedorsal bank of the superior ramus of the arcuate sulcus In additionwidespread connectivity was found in themedial wall including thecingulate motor area supplementary motor area and medialparietal cortex The network pattern closely resembles the ldquosoma-tomotorrdquo network previously described in the anesthetized ma-caque using a seed-region based approach (Vincent et al 2007)
bull RSN I (parieto-occipital) A network consisting of regions involved invisual processing including bilateral areas V1 V2 V2 V3 area POand area MTMST Connectivity was also observed unilaterally inarea 8A (left hemisphere) and 46d (right hemisphere) Similarnetwork patterns have been observed in the macaque in bothresting-state (Vincent et al 2007) and awake fMRI studies(Stefanacci et al 1998)
bull RSN J (postcentral) A network including the postcentral andprecentral gyrus areas dedicated to somatosensory processing(Kaas 1993) Opposite modulation was found to occur in area POand in the anterior cingulate cortex area 24 ab
bull RSN K (hippocampal) A medial temporal network corresponding toareas associated with the macaque declarative memory system(Squire and Zola-Morgan 1991) The RSN bilaterally encompassedthe hippocampus entorhinal perirhinal and parahippocampalcortical areas Anti-correlated bilateral connectivity of anteriorcingulate cortex area 25 was also observed The network althoughpartially explored in several previous electrophysiological studies(eg Rolls et al 1993 Wirth et al 2003) has not been shown withmonkey fMRI possibly due to the difficult task demands required ofthe animals in order to elicit activations in these areas
Single subject ICA
To compare single subject ICA with group ICA components wespatially correlated unthresholded single subject IC maps with the
unthresholded group ICmaps The correlation coefficients for all mapsand all animals was significant (pb10minus20 p value corrected formultiple comparisons using Bonferroni correction) Table 1 shows themean correlation coefficients between the single-subject ICA compo-nents and the group-ICA networks Mean correlation values werefound to be significantly different from 0 (one sample t-test pb001)A representative network (RSN B mdash fronto-parietal) is shown for allmonkeys in Fig 2 Networks for all animals are shown in Supple-mentary Figs 4ndash9 In some animals the same single subjectcomponent was best correlated to two or more of the group ICAnetworks (for example Supplementary Fig 5 componentscorresponding to networks D and I are the same)
Functional network connectivity
Fig 3 shows a FNC diagram for the 11 identified RSNs RSNs arerepresented by circular nodes and significantly correlated RSNs arerepresented by connecting lines For example a line connectsnetworks I and J representing significant functional connectivitybetween those two networks RSN C (posterior-parietal) and RSN I(parieto-occipital) represent the most connected nodes RSN D(occipito-temporal) and RSN E (frontal) also show a high degree ofFNC The other sensory networks show little or no connectivity withother RSNs
Discussion
Alterations in functional connectivity recorded using spontaneousBOLD fluctuations have been suggested as the origin or product ofmultiple disease states (Auer 2008 Greicius 2008) Assessing theirelectrophysiological correlate(s) and establishing the relationshipbetween large-scale functional network connectivity and diseaserequire a suitable animal model Here to the best of our knowledgewe report the first comprehensive application of group independentcomponent analysis (ICA) to monkey fMRI data and the first resting-state examination of the macaque at 7 T ICA successfully identified 11prominent macaque RSNs representing multiple levels of neuralprocessing Networks encompassing sensory and motor areasincluding the visual auditory motor and somato-sensory regionscan be considered to be lower-order in a cognitive processinghierarchy The RSNs comprising areas known to be involved inexecutive control attention reward evaluation and default-modeactivity may represent higher-order processing with temporal net-works putatively responsible for visual processing and memoryproviding intermediate processing In cases where the system hasbeen explored with fMRI the patterns of intrinsic functionalconnectivity are consistent with stimulus-evoked patterns found intask-based studies (see Results) The functional RSNs reported in the
Fig 2 Resting-state networks (RSN) of all monkeys (M1ndashM6) following single-subject independent component analysis (ICA) that were most spatially correlated to group-ICAidentified RSN B (fronto-parietal) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 seehttpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of bothhemispheres and the right images depict the lateral and medial views of right hemisphere
Fig 3 Average functional network connectivity of macaque resting-state networks (RSNs) Lines and numerical values indicate functional connectivity between two RSNs in whichthere was a significant temporal correlation of their respective time-courses (one-sample Student t-test pb005 corrected for multiple comparisons between 55 pairs with a timeleadlag of +minus3 s) Network letters refer to spatial representations shown in Fig 1
1551RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

1547RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Although their meaning is not fully understood changes infunctional RSNs have been recently reported in several psychiatricand developmental disorders including depression attention deficit-hyperactivity disorder schizophrenia Alzheimers disease epilepsyand multiple sclerosis (Auer 2008 Greicius 2008) Given theextraordinary potential for RSNs as possible diagnostic or prognosticmarkers it is crucial to understand the physiological mechanisms offluctuation regulation and entrainment of LFFs and the RSNs that arerevealed through their synchronization
Nonhuman primates and in particular macaque monkeys havebeen used as surrogates for the study of human brain function forseveral decades and might therefore represent an ideal animal modelfor the study of RSNs Although macaque and human brains share ahigh degree of similarity in terms of cytoarchitecture (Petrides andPandya 1999 2002a Onguumlr et al 2003) functional organization(Rees et al 2000 Koyama et al 2004 Petrides et al 2005 Nakaharaet al 2007) and anatomical connections (Croxson et al 2005 Kellyet al 2010) there also exist structural (Preuss 2000 Rilling 2006)morphological (Buxhoeveden et al 2001) and functional (Orbanet al 2004 Preuss 2004) differences between the brains of these twoprimate species (Passingham 2009)
While a few recent studies have revealed homologous RSNsbetween human and nonhuman primates these studies eitherutilized seed-region analysis (Vincent et al 2007 2010 Margulieset al 2009) or ICA in only two animals (Moeller et al 2009) Here wepresent a comprehensive evaluation of macaque RSNs at 7 T usinggroup ICA and an analysis methodology that is very similar to whathas been used in humans The results show striking similarity ofmacaque RSNs to previously described human RSNs
Material and methods
Animal preparation
Data was obtained from 6macaque monkeys (Macaca fascicularis 2male 4 female)whoseweights ranged from36 kg to 53 kg (mean+minusSD=458+minus14 kg) Surgical and experimental procedures werecarried out in accordance with the Canadian Council of Animal Carepolicy on the use of laboratory animals and approved by the Animal UseSubcommittee of the University of Western Ontario Council on AnimalCare
Animals were initially prepared for imaging by undergoing asurgical procedure to place an MRI-compatible custom-built acrylichead post that served to restrain the head during image acquisitionThe post was anchored to the skull with 6 mm ceramic bone screws(Thomas Recording Giessen Germany) and dental acrylic
On the day of scanning anesthesia was first induced by intramus-cular injections of atropine (04 mgkg) ipratropium (0025 mgkg)and ketamine hydrochloride (75 mgkg) Animals were then adminis-tered3 mlof propofol (10 mgml) intravenouslyvia the saphenousveinFollowing oral intubation with an endotracheal tube anesthesia wasmaintained using 15 isoflurane mixed with oxygen Animals werespontaneously breathing throughout the duration of the experimentThe animal was placed in a custom-built primate chair containingfixation for head immobilization and an integrated custom RF coil andinserted into the bore for image acquisition The isoflurane level wasthen lowered to 1 for imaging experiments Rectal temperature via afiber-optic temperature probe (FISO Quebec City QC) respiration viabellows (Siemens Corp Union NJ) and end-tidal CO2 via capnometer(Covidien-Nellcor Boulder CO) were continuously monitored Physio-logical parameters were in the normal range (temperature 365 degCbreathing 25ndash30 breathsmin end-tidal CO2 24ndash28 mmHg) through-out the duration of the experiment Warmth was maintained using aheatingdisk (Snugglesafe LittlehamptonWest SussexUK) and thermalinsulation
Anesthesia was utilized in this study to eliminate motion effectsphysiological stress and training requirements Although isofluranehas been shown to have vasodilator properties (Farber et al 1997)altering cerebrovascular activity in a dose-dependent manner(Vincent et al 2007) synchronous spontaneous BOLD fluctuationshave been previously reported using an isoflurane regime in bothmonkeys (Vincent et al 2007) and rats (Hutchison et al 2010)However it is still likely that anesthesia can affect RSNs
Data acquisition
All data were acquired on an actively shielded 7 Tesla 68 cmhorizontal borehumanhead scannerwith aDirectDrive console (VarianYarnton UK Walnut Creek CA) and a Siemens AC84 gradient sub-system (Erlangen Germany) operating at a slew rate of 350mTms Anin-house designed and manufactured conformal 5 channel transceiveprimate head RF coil was used for all experiments Magnetic fieldoptimization (B0 shimming)wasperformedusing anautomated three-dimensional mapping procedure over the specific imaging volume ofinterest For each monkey 2 runs of 300 continuous EPI functionalvolumes (TR=2000 ms TE=16 ms flip angle=70deg slices=30matrix=72times72 FOV=96times96 mm acquisition voxel size=13times13times15 mm) were acquired The total acquisition time of each scanwas 10 min EPI images were acquired with GRAPPA at an accelerationfactor of 2 Every image was corrected for physiological fluctuationsusing navigator-echo-correction A high-resolution T2-weighted ana-tomical reference volume was acquired along the same orientation asthe functional images using a fast spin echo acquisition scheme(TR=5000 ms TE=386 ms echo train length=5 effectiveecho=3 slices=30 matrix=256times250 FOV=96times96 mm acquisi-tion voxel size=375 μmtimes384 μmtimes15 mm) T2 imagingusing fast spinecho as compared to gradient echo based imaging such as T1-weightedMP-RAGE reduced the amount of image distortion caused by skullimplants such as the ceramic bone screws and plastic head post
Image preprocessing
All preprocessing was implemented using the FSL softwarepackage (httpwwwfmriboxacuk) and included motion correc-tion (six parameter affine transformation) brain extraction spatialsmoothing (FWHM=3 mm) high-pass temporal filtering (Gaussian-weighted least-squares straight line fitting with sigma=100 s) low-pass temporal filtering (HWHM=28 s Gaussian filter) and normal-ization (12 DOF linear affine transformation implemented in FLIRT) tothe F99 atlas template (Van Essen 2004 see httpsumsdbwustledusumsmacaquemoredo)
Group independent component analysis
Group ICA unlike single-subject ICA allows inferences to be madeat the group level When examining subjects individually it is difficultto compare components since they are not ordered and differentcomponents may be revealed in each subject Entering all subjectsinto an ICA analysis and estimating one set of components has theadvantage of ordering the components of different subjects in thesame way This produces a single set of ldquogrouprdquo components that canthen be interpreted Additionally weak sources with differentcharacteristics across subjects (ie noise) will be suppressed allowinga more accurate reflection of population dynamics
Group ICA was conducted using the GIFT software package(Calhoun et al 2001 httpicatbsourceforgenet) Data from bothruns of all animals were concatenated and the temporal dimension ofthis aggregated data set was reduced by means of principalcomponent analysis (PCA) This was followed by spatial componentestimation using the Infomax algorithm Component time-coursesand spatial maps for each animal were then back-reconstructed using
1548 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
the aggregated components and the results from the data reductionstep (Jafri et al 2008 Calhoun et al 2001) Because ICA is a stochasticestimation process the final component maps can vary depending onthe initial algorithm conditions To quantify the reliability of thedecomposition ICA was reiterated 20 times using the ICASSO toolbox(Himberg et al 2004) ICASSO represents each of the estimatedcomponents for each iteration as a point in the signal space It thenreturns a stability index of the estimate cluster computed as thedifference between the average intra- and inter-cluster similarities Inthe ideal case the repeated estimates are concentrated in compactand close-to-orthogonal clusters resulting in an index of all estimate-clusters that approaches one
There are currently no well-established criteria to guide theselection of an optimal number of independent components (ICs) for agiven data set (Cole et al 2010) The model order or dimensionestimate defines the number of components that the algorithm willextract and in that regard represents a prediction about the number ofunderlying sources in the data If the model order is increased agreater number of networks will be found accounting for the data in amore detailed way albeit causing networks to branch into smallersub-networks (Smith et al 2009 Abou-Elseoud et al 2010) Oneproposed method uses the minimum description length criterion fordimension estimation (Jafri et al 2008 Li et al 2007) howeverwhenapplied to our data set this technique resulted in an estimation of 253ICs Instead 20 components were chosen as this gave a manageablenumber of components and approximated model orders commonlyused in human studies large-scale brain networks (Smith et al 2009Abou-Elseoud et al 2010 Calhoun et al 2001 Demirci et al 2009)
Themean group ICswere then scaled to empirically derived z-scoresby dividing by the standard deviation of the original time-course Thez-scores approximate the temporal correlation between each voxeland the associated component where the magnitude of the z-scorespecifies the strength of the linear relationship A negative z-scoreindicates voxels in which the BOLD signals are modulated opposite tothat of the IC time-course A z-score value of +minus1 was used as thelower limit threshold of functional connectivity
RSN identification and visualization
The ICA derived components were visually inspected and labeledbased on the activation patterns in reference to known anatomical andfunctional locations Classification of the components in terms of RSNswas performed by comparison with known macaque functionalnetworks and previously identified RSNs from ICA studies in humans(Beckmann et al 2005 Damoiseaux et al 2006 Smith et al 2009) Inaddition to identifying spatially independent networks ICA is able toparse out noise due to physiological fluctuations motion and hardwareartifacts from the original source data (Thomas et al 2002 Liao et al2006 Perlbarg et al 2007) Components with high spatial correspon-dence to cerebrospinal spinal fluid (see Supplementary Fig 1) or withlow correlation to gray matter were discarded
Group data were projected from volume data to the F99 corticalsurface using the CARET (httpwwwnitrcorgprojectscaret)enclosed-voxel method
Single-subject ICA
Single-subject ICA is better suited to reveal individual features ofresting-state connectivity as subtle differences between subjects mayget lost in an ICA of a group data set To examine individual subjectmaps and allow comparison with a previously published report(Moeller et al 2009) single subject-ICA was conducted using thesame parameters as the group-ICA The unthresholded group-ICAderived networks were then used as templates to order thecomponent maps from the individual ICA by calculating the spatialcorrelation coefficient of the unthresholded individual maps
Functional network connectivity
Spatial ICA maximizes the statistical independence in the spatialdomain though the components are not temporally constrained andICs can exhibit considerable temporal correlations (Calhoun et al2003) The temporal dependencies are significant albeit not as high asthose between regions within an IC (Jafri et al 2008) Functionalnetwork connectivity (FNC) analysis examines these temporaldependencies among ICs to establish the functional connectivityamong the large scale networks and has been shown to provideadditional information regarding macroscopic brain organization(Jafri et al 2008 Demirci et al 2009) FNC analysis was conductedusing the FNC software package (httpmialabmrnorgsoftware)using analysis steps described previously (Jafri et al 2008) One-sample Students t-tests were used to test statistically significantmaximal lagged correlation (+minus3 s) combinations from the 55possible combinations (pb005 corrected for multiple comparisons)
Results
Resting-state networks
Group-ICA successfully decomposed the resting-state functionaldata of 6monkeys into 20 independent components ICASSO returned astability index for each estimate-cluster that approached 1 (mean+minusSD=0980+minus0006) indicating that the components are concen-trated in compact and close-to-orthogonal clusters and highly consis-tent across multiple ICA runs Eleven components were found to have ahigh correspondence with gray matter as well as showing networkcharacteristics (ie spatial maps containing more than one functionalarea) and were thereby deemed to be physiologically relevant Thespatial maps of the RSNs obtained with ICA analysis are illustrated inFig 1 (for RSNmaps displayed on a flattened cortex see SupplementaryFig 2) The components accounted for 5981 of the datas variance Thedegree of cortical coverage is illustrated in Supplementary Fig 3 Despiteinclusion of the entire brain components were primarily restricted tocortical areas Therefore we focus on cortical connectivity in this studyOn the basis of our classification results the 11 RSNs can be described asfollows
bull RSN A (precentralndashtemporal) The RSN included extended bilateralareas of the ventral precentral gyrus corresponding to ventralpremotor areas F4 and F5 activation in the inferior ramus of thearcuate sulcus the precentral opercular cortex primary andsecondary somatosensory cortex and insular cortex In additionconnectivity was observed with the auditory cortex The networkincluded areas that correspond to the ventral motor system thoughtto play an important role in understanding action and visuomotorcontrol of grasping and also included areas underlying the control ofmouth and tongue (Nelissen et al 2005 Joly et al 2009 Phillipsand Porter 1977)
bull RSN B (fronto-parietal) Bilateral connectivity was found in theanterior bank fundus and posterior bank of the inferior arm of thearcuate sulcus The anterior extent of this region corresponds to thelocation of the frontal eye fields (Bruce and Goldberg 1985)Connectivity was found in the ventral prefrontal cortex and dorsalpremotor cortex Further network connectivity was found in area 3Bin the dorsal central sulcus fundus and in both banks of theintraparietal sulcus In addition the right hemisphere showedpositive correlation of area 6 corresponding to the supplementarymotor area or possibly the supplementary eye fields (Schlag andSchlag-Rey 1987) The network includes brain areas associatedwiththe oculomotor system (also referred to as the dorsal attentionnetwork) responsible for goal-oriented saccadic eye movements(Johnston and Everling 2008) and mediating goal-directed top-down processing (Noudoost et al 2010) The functional network
Fig 1 Cortical representation of eleven resting-state networks (RSNs) identified by independent component analysis of fMRI data acquired from isoflurane-anesthetized macaquemonkeys (N=6) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 see httpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of both hemispheres and theright images depict the lateral and medial views of right hemisphere Potential functional roles of each network are discussed in the text
1549RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
has been previously reported in eye movement tasks (Baker et al2006 Ford et al 2009) and resting-state analysis (Vincent et al2007) of the macaque
bull RSN C (posterior-parietal) The network encompassed the posteriorcingulateprecuneus cortex bilateral areas PG and PE of the parietal
cortex and visual areas V2 and V3 In addition this networkcontained connectivity patterns in the dorsolateral prefrontal cortexand in the dorsal premotor cortex in the left hemisphere The brainareas include areas found in the default-mode network (DMN) areasthat are deactivated during attention-demanding cognitive tasks
Table 1Spatial correlations of single subject ICA derived networks to group-ICA derivednetworks
Network Mean STD ERR Range
A 0691 0068 0383ndash0841B 0560 0082 0176ndash0750C 0463 0062 0227ndash0649D 0501 0078 0215ndash0743E 0373 0046 0275ndash0584F 0459 0073 0184ndash0623G 0372 0060 0200ndash0586H 0559 0059 0272ndash0661I 0501 0065 0199ndash0655J 0375 0045 0207ndash0546K 0264 0017 0186ndash0302
Note Mean correlation values are significantly different from 0 (one sample t-testpb001)
1550 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
and in humans has been suggested to support higher-order mentalfaculties (Raichle et al 2001)
bull RSN D (occipito-temporal) Network areas included bilateral area TOV4 TEO and the arcuate sulcus Bilateral area 29 and Brodmann area10 (left hemisphere) were also functionally connected Previousstudies have shown these temporal lobe areas are critical for higher-order visual processing (Tsao et al 2003)
bull RSN E (frontal) This RSN encompassed bilateral regions in severalprefrontal areas including the anterior bank of the arcuate sulcuscorresponding to the frontal eye fields the upper ramus of thearcuate sulcus the posterior portion of the principal sulcus as wellas the dorsal bank of the principal sulcus In addition LFFsynchronization was also found in area 9 the premotor cortex andthe anterior cingulate cortex These frontalprefrontal areas havebeen shown to be components of the executive system suggested toprovide bias signals to other areas of the brain in order to implementcognitive control (Miller and Cohen 2001)
bull RSN F (superior-temporal) A network encompassing the auditory beltparabelt and bilateral area 22 on the superior temporal gyrus Anti-correlated areas were found in the left principal sulcus and in the leftarcuate sulcus The network pattern resembles the mean functionalactivity resulting from the presentation of multiple sounds categoriesin a previous task-based fMRI study of the awake monkey (Petkovet al 2008) and is likely responsible for acoustic processing andinterpretation (Rauschecker and Scott 2009)
bull RSN G (cingulo-insular) A network of areas including bilateral regionsin the insular cortex the anterior cingulate cortex area 24ab and theorbitofrontal cortex area 14 These areas have been associated withthe reward system involved in the regulation and control of behavior(Kringelbach 2005)
bull RSN H (paracentral) A network reflective of the dorsal motorsystem involved in the control of limb movements (Dum and Strick2002) Network connectivity included bilateral primary motorcortex dorsally and also in the central sulcus area F2 and thedorsal bank of the superior ramus of the arcuate sulcus In additionwidespread connectivity was found in themedial wall including thecingulate motor area supplementary motor area and medialparietal cortex The network pattern closely resembles the ldquosoma-tomotorrdquo network previously described in the anesthetized ma-caque using a seed-region based approach (Vincent et al 2007)
bull RSN I (parieto-occipital) A network consisting of regions involved invisual processing including bilateral areas V1 V2 V2 V3 area POand area MTMST Connectivity was also observed unilaterally inarea 8A (left hemisphere) and 46d (right hemisphere) Similarnetwork patterns have been observed in the macaque in bothresting-state (Vincent et al 2007) and awake fMRI studies(Stefanacci et al 1998)
bull RSN J (postcentral) A network including the postcentral andprecentral gyrus areas dedicated to somatosensory processing(Kaas 1993) Opposite modulation was found to occur in area POand in the anterior cingulate cortex area 24 ab
bull RSN K (hippocampal) A medial temporal network corresponding toareas associated with the macaque declarative memory system(Squire and Zola-Morgan 1991) The RSN bilaterally encompassedthe hippocampus entorhinal perirhinal and parahippocampalcortical areas Anti-correlated bilateral connectivity of anteriorcingulate cortex area 25 was also observed The network althoughpartially explored in several previous electrophysiological studies(eg Rolls et al 1993 Wirth et al 2003) has not been shown withmonkey fMRI possibly due to the difficult task demands required ofthe animals in order to elicit activations in these areas
Single subject ICA
To compare single subject ICA with group ICA components wespatially correlated unthresholded single subject IC maps with the
unthresholded group ICmaps The correlation coefficients for all mapsand all animals was significant (pb10minus20 p value corrected formultiple comparisons using Bonferroni correction) Table 1 shows themean correlation coefficients between the single-subject ICA compo-nents and the group-ICA networks Mean correlation values werefound to be significantly different from 0 (one sample t-test pb001)A representative network (RSN B mdash fronto-parietal) is shown for allmonkeys in Fig 2 Networks for all animals are shown in Supple-mentary Figs 4ndash9 In some animals the same single subjectcomponent was best correlated to two or more of the group ICAnetworks (for example Supplementary Fig 5 componentscorresponding to networks D and I are the same)
Functional network connectivity
Fig 3 shows a FNC diagram for the 11 identified RSNs RSNs arerepresented by circular nodes and significantly correlated RSNs arerepresented by connecting lines For example a line connectsnetworks I and J representing significant functional connectivitybetween those two networks RSN C (posterior-parietal) and RSN I(parieto-occipital) represent the most connected nodes RSN D(occipito-temporal) and RSN E (frontal) also show a high degree ofFNC The other sensory networks show little or no connectivity withother RSNs
Discussion
Alterations in functional connectivity recorded using spontaneousBOLD fluctuations have been suggested as the origin or product ofmultiple disease states (Auer 2008 Greicius 2008) Assessing theirelectrophysiological correlate(s) and establishing the relationshipbetween large-scale functional network connectivity and diseaserequire a suitable animal model Here to the best of our knowledgewe report the first comprehensive application of group independentcomponent analysis (ICA) to monkey fMRI data and the first resting-state examination of the macaque at 7 T ICA successfully identified 11prominent macaque RSNs representing multiple levels of neuralprocessing Networks encompassing sensory and motor areasincluding the visual auditory motor and somato-sensory regionscan be considered to be lower-order in a cognitive processinghierarchy The RSNs comprising areas known to be involved inexecutive control attention reward evaluation and default-modeactivity may represent higher-order processing with temporal net-works putatively responsible for visual processing and memoryproviding intermediate processing In cases where the system hasbeen explored with fMRI the patterns of intrinsic functionalconnectivity are consistent with stimulus-evoked patterns found intask-based studies (see Results) The functional RSNs reported in the
Fig 2 Resting-state networks (RSN) of all monkeys (M1ndashM6) following single-subject independent component analysis (ICA) that were most spatially correlated to group-ICAidentified RSN B (fronto-parietal) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 seehttpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of bothhemispheres and the right images depict the lateral and medial views of right hemisphere
Fig 3 Average functional network connectivity of macaque resting-state networks (RSNs) Lines and numerical values indicate functional connectivity between two RSNs in whichthere was a significant temporal correlation of their respective time-courses (one-sample Student t-test pb005 corrected for multiple comparisons between 55 pairs with a timeleadlag of +minus3 s) Network letters refer to spatial representations shown in Fig 1
1551RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

1548 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
the aggregated components and the results from the data reductionstep (Jafri et al 2008 Calhoun et al 2001) Because ICA is a stochasticestimation process the final component maps can vary depending onthe initial algorithm conditions To quantify the reliability of thedecomposition ICA was reiterated 20 times using the ICASSO toolbox(Himberg et al 2004) ICASSO represents each of the estimatedcomponents for each iteration as a point in the signal space It thenreturns a stability index of the estimate cluster computed as thedifference between the average intra- and inter-cluster similarities Inthe ideal case the repeated estimates are concentrated in compactand close-to-orthogonal clusters resulting in an index of all estimate-clusters that approaches one
There are currently no well-established criteria to guide theselection of an optimal number of independent components (ICs) for agiven data set (Cole et al 2010) The model order or dimensionestimate defines the number of components that the algorithm willextract and in that regard represents a prediction about the number ofunderlying sources in the data If the model order is increased agreater number of networks will be found accounting for the data in amore detailed way albeit causing networks to branch into smallersub-networks (Smith et al 2009 Abou-Elseoud et al 2010) Oneproposed method uses the minimum description length criterion fordimension estimation (Jafri et al 2008 Li et al 2007) howeverwhenapplied to our data set this technique resulted in an estimation of 253ICs Instead 20 components were chosen as this gave a manageablenumber of components and approximated model orders commonlyused in human studies large-scale brain networks (Smith et al 2009Abou-Elseoud et al 2010 Calhoun et al 2001 Demirci et al 2009)
Themean group ICswere then scaled to empirically derived z-scoresby dividing by the standard deviation of the original time-course Thez-scores approximate the temporal correlation between each voxeland the associated component where the magnitude of the z-scorespecifies the strength of the linear relationship A negative z-scoreindicates voxels in which the BOLD signals are modulated opposite tothat of the IC time-course A z-score value of +minus1 was used as thelower limit threshold of functional connectivity
RSN identification and visualization
The ICA derived components were visually inspected and labeledbased on the activation patterns in reference to known anatomical andfunctional locations Classification of the components in terms of RSNswas performed by comparison with known macaque functionalnetworks and previously identified RSNs from ICA studies in humans(Beckmann et al 2005 Damoiseaux et al 2006 Smith et al 2009) Inaddition to identifying spatially independent networks ICA is able toparse out noise due to physiological fluctuations motion and hardwareartifacts from the original source data (Thomas et al 2002 Liao et al2006 Perlbarg et al 2007) Components with high spatial correspon-dence to cerebrospinal spinal fluid (see Supplementary Fig 1) or withlow correlation to gray matter were discarded
Group data were projected from volume data to the F99 corticalsurface using the CARET (httpwwwnitrcorgprojectscaret)enclosed-voxel method
Single-subject ICA
Single-subject ICA is better suited to reveal individual features ofresting-state connectivity as subtle differences between subjects mayget lost in an ICA of a group data set To examine individual subjectmaps and allow comparison with a previously published report(Moeller et al 2009) single subject-ICA was conducted using thesame parameters as the group-ICA The unthresholded group-ICAderived networks were then used as templates to order thecomponent maps from the individual ICA by calculating the spatialcorrelation coefficient of the unthresholded individual maps
Functional network connectivity
Spatial ICA maximizes the statistical independence in the spatialdomain though the components are not temporally constrained andICs can exhibit considerable temporal correlations (Calhoun et al2003) The temporal dependencies are significant albeit not as high asthose between regions within an IC (Jafri et al 2008) Functionalnetwork connectivity (FNC) analysis examines these temporaldependencies among ICs to establish the functional connectivityamong the large scale networks and has been shown to provideadditional information regarding macroscopic brain organization(Jafri et al 2008 Demirci et al 2009) FNC analysis was conductedusing the FNC software package (httpmialabmrnorgsoftware)using analysis steps described previously (Jafri et al 2008) One-sample Students t-tests were used to test statistically significantmaximal lagged correlation (+minus3 s) combinations from the 55possible combinations (pb005 corrected for multiple comparisons)
Results
Resting-state networks
Group-ICA successfully decomposed the resting-state functionaldata of 6monkeys into 20 independent components ICASSO returned astability index for each estimate-cluster that approached 1 (mean+minusSD=0980+minus0006) indicating that the components are concen-trated in compact and close-to-orthogonal clusters and highly consis-tent across multiple ICA runs Eleven components were found to have ahigh correspondence with gray matter as well as showing networkcharacteristics (ie spatial maps containing more than one functionalarea) and were thereby deemed to be physiologically relevant Thespatial maps of the RSNs obtained with ICA analysis are illustrated inFig 1 (for RSNmaps displayed on a flattened cortex see SupplementaryFig 2) The components accounted for 5981 of the datas variance Thedegree of cortical coverage is illustrated in Supplementary Fig 3 Despiteinclusion of the entire brain components were primarily restricted tocortical areas Therefore we focus on cortical connectivity in this studyOn the basis of our classification results the 11 RSNs can be described asfollows
bull RSN A (precentralndashtemporal) The RSN included extended bilateralareas of the ventral precentral gyrus corresponding to ventralpremotor areas F4 and F5 activation in the inferior ramus of thearcuate sulcus the precentral opercular cortex primary andsecondary somatosensory cortex and insular cortex In additionconnectivity was observed with the auditory cortex The networkincluded areas that correspond to the ventral motor system thoughtto play an important role in understanding action and visuomotorcontrol of grasping and also included areas underlying the control ofmouth and tongue (Nelissen et al 2005 Joly et al 2009 Phillipsand Porter 1977)
bull RSN B (fronto-parietal) Bilateral connectivity was found in theanterior bank fundus and posterior bank of the inferior arm of thearcuate sulcus The anterior extent of this region corresponds to thelocation of the frontal eye fields (Bruce and Goldberg 1985)Connectivity was found in the ventral prefrontal cortex and dorsalpremotor cortex Further network connectivity was found in area 3Bin the dorsal central sulcus fundus and in both banks of theintraparietal sulcus In addition the right hemisphere showedpositive correlation of area 6 corresponding to the supplementarymotor area or possibly the supplementary eye fields (Schlag andSchlag-Rey 1987) The network includes brain areas associatedwiththe oculomotor system (also referred to as the dorsal attentionnetwork) responsible for goal-oriented saccadic eye movements(Johnston and Everling 2008) and mediating goal-directed top-down processing (Noudoost et al 2010) The functional network
Fig 1 Cortical representation of eleven resting-state networks (RSNs) identified by independent component analysis of fMRI data acquired from isoflurane-anesthetized macaquemonkeys (N=6) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 see httpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of both hemispheres and theright images depict the lateral and medial views of right hemisphere Potential functional roles of each network are discussed in the text
1549RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
has been previously reported in eye movement tasks (Baker et al2006 Ford et al 2009) and resting-state analysis (Vincent et al2007) of the macaque
bull RSN C (posterior-parietal) The network encompassed the posteriorcingulateprecuneus cortex bilateral areas PG and PE of the parietal
cortex and visual areas V2 and V3 In addition this networkcontained connectivity patterns in the dorsolateral prefrontal cortexand in the dorsal premotor cortex in the left hemisphere The brainareas include areas found in the default-mode network (DMN) areasthat are deactivated during attention-demanding cognitive tasks
Table 1Spatial correlations of single subject ICA derived networks to group-ICA derivednetworks
Network Mean STD ERR Range
A 0691 0068 0383ndash0841B 0560 0082 0176ndash0750C 0463 0062 0227ndash0649D 0501 0078 0215ndash0743E 0373 0046 0275ndash0584F 0459 0073 0184ndash0623G 0372 0060 0200ndash0586H 0559 0059 0272ndash0661I 0501 0065 0199ndash0655J 0375 0045 0207ndash0546K 0264 0017 0186ndash0302
Note Mean correlation values are significantly different from 0 (one sample t-testpb001)
1550 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
and in humans has been suggested to support higher-order mentalfaculties (Raichle et al 2001)
bull RSN D (occipito-temporal) Network areas included bilateral area TOV4 TEO and the arcuate sulcus Bilateral area 29 and Brodmann area10 (left hemisphere) were also functionally connected Previousstudies have shown these temporal lobe areas are critical for higher-order visual processing (Tsao et al 2003)
bull RSN E (frontal) This RSN encompassed bilateral regions in severalprefrontal areas including the anterior bank of the arcuate sulcuscorresponding to the frontal eye fields the upper ramus of thearcuate sulcus the posterior portion of the principal sulcus as wellas the dorsal bank of the principal sulcus In addition LFFsynchronization was also found in area 9 the premotor cortex andthe anterior cingulate cortex These frontalprefrontal areas havebeen shown to be components of the executive system suggested toprovide bias signals to other areas of the brain in order to implementcognitive control (Miller and Cohen 2001)
bull RSN F (superior-temporal) A network encompassing the auditory beltparabelt and bilateral area 22 on the superior temporal gyrus Anti-correlated areas were found in the left principal sulcus and in the leftarcuate sulcus The network pattern resembles the mean functionalactivity resulting from the presentation of multiple sounds categoriesin a previous task-based fMRI study of the awake monkey (Petkovet al 2008) and is likely responsible for acoustic processing andinterpretation (Rauschecker and Scott 2009)
bull RSN G (cingulo-insular) A network of areas including bilateral regionsin the insular cortex the anterior cingulate cortex area 24ab and theorbitofrontal cortex area 14 These areas have been associated withthe reward system involved in the regulation and control of behavior(Kringelbach 2005)
bull RSN H (paracentral) A network reflective of the dorsal motorsystem involved in the control of limb movements (Dum and Strick2002) Network connectivity included bilateral primary motorcortex dorsally and also in the central sulcus area F2 and thedorsal bank of the superior ramus of the arcuate sulcus In additionwidespread connectivity was found in themedial wall including thecingulate motor area supplementary motor area and medialparietal cortex The network pattern closely resembles the ldquosoma-tomotorrdquo network previously described in the anesthetized ma-caque using a seed-region based approach (Vincent et al 2007)
bull RSN I (parieto-occipital) A network consisting of regions involved invisual processing including bilateral areas V1 V2 V2 V3 area POand area MTMST Connectivity was also observed unilaterally inarea 8A (left hemisphere) and 46d (right hemisphere) Similarnetwork patterns have been observed in the macaque in bothresting-state (Vincent et al 2007) and awake fMRI studies(Stefanacci et al 1998)
bull RSN J (postcentral) A network including the postcentral andprecentral gyrus areas dedicated to somatosensory processing(Kaas 1993) Opposite modulation was found to occur in area POand in the anterior cingulate cortex area 24 ab
bull RSN K (hippocampal) A medial temporal network corresponding toareas associated with the macaque declarative memory system(Squire and Zola-Morgan 1991) The RSN bilaterally encompassedthe hippocampus entorhinal perirhinal and parahippocampalcortical areas Anti-correlated bilateral connectivity of anteriorcingulate cortex area 25 was also observed The network althoughpartially explored in several previous electrophysiological studies(eg Rolls et al 1993 Wirth et al 2003) has not been shown withmonkey fMRI possibly due to the difficult task demands required ofthe animals in order to elicit activations in these areas
Single subject ICA
To compare single subject ICA with group ICA components wespatially correlated unthresholded single subject IC maps with the
unthresholded group ICmaps The correlation coefficients for all mapsand all animals was significant (pb10minus20 p value corrected formultiple comparisons using Bonferroni correction) Table 1 shows themean correlation coefficients between the single-subject ICA compo-nents and the group-ICA networks Mean correlation values werefound to be significantly different from 0 (one sample t-test pb001)A representative network (RSN B mdash fronto-parietal) is shown for allmonkeys in Fig 2 Networks for all animals are shown in Supple-mentary Figs 4ndash9 In some animals the same single subjectcomponent was best correlated to two or more of the group ICAnetworks (for example Supplementary Fig 5 componentscorresponding to networks D and I are the same)
Functional network connectivity
Fig 3 shows a FNC diagram for the 11 identified RSNs RSNs arerepresented by circular nodes and significantly correlated RSNs arerepresented by connecting lines For example a line connectsnetworks I and J representing significant functional connectivitybetween those two networks RSN C (posterior-parietal) and RSN I(parieto-occipital) represent the most connected nodes RSN D(occipito-temporal) and RSN E (frontal) also show a high degree ofFNC The other sensory networks show little or no connectivity withother RSNs
Discussion
Alterations in functional connectivity recorded using spontaneousBOLD fluctuations have been suggested as the origin or product ofmultiple disease states (Auer 2008 Greicius 2008) Assessing theirelectrophysiological correlate(s) and establishing the relationshipbetween large-scale functional network connectivity and diseaserequire a suitable animal model Here to the best of our knowledgewe report the first comprehensive application of group independentcomponent analysis (ICA) to monkey fMRI data and the first resting-state examination of the macaque at 7 T ICA successfully identified 11prominent macaque RSNs representing multiple levels of neuralprocessing Networks encompassing sensory and motor areasincluding the visual auditory motor and somato-sensory regionscan be considered to be lower-order in a cognitive processinghierarchy The RSNs comprising areas known to be involved inexecutive control attention reward evaluation and default-modeactivity may represent higher-order processing with temporal net-works putatively responsible for visual processing and memoryproviding intermediate processing In cases where the system hasbeen explored with fMRI the patterns of intrinsic functionalconnectivity are consistent with stimulus-evoked patterns found intask-based studies (see Results) The functional RSNs reported in the
Fig 2 Resting-state networks (RSN) of all monkeys (M1ndashM6) following single-subject independent component analysis (ICA) that were most spatially correlated to group-ICAidentified RSN B (fronto-parietal) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 seehttpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of bothhemispheres and the right images depict the lateral and medial views of right hemisphere
Fig 3 Average functional network connectivity of macaque resting-state networks (RSNs) Lines and numerical values indicate functional connectivity between two RSNs in whichthere was a significant temporal correlation of their respective time-courses (one-sample Student t-test pb005 corrected for multiple comparisons between 55 pairs with a timeleadlag of +minus3 s) Network letters refer to spatial representations shown in Fig 1
1551RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

Fig 1 Cortical representation of eleven resting-state networks (RSNs) identified by independent component analysis of fMRI data acquired from isoflurane-anesthetized macaquemonkeys (N=6) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 see httpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of both hemispheres and theright images depict the lateral and medial views of right hemisphere Potential functional roles of each network are discussed in the text
1549RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
has been previously reported in eye movement tasks (Baker et al2006 Ford et al 2009) and resting-state analysis (Vincent et al2007) of the macaque
bull RSN C (posterior-parietal) The network encompassed the posteriorcingulateprecuneus cortex bilateral areas PG and PE of the parietal
cortex and visual areas V2 and V3 In addition this networkcontained connectivity patterns in the dorsolateral prefrontal cortexand in the dorsal premotor cortex in the left hemisphere The brainareas include areas found in the default-mode network (DMN) areasthat are deactivated during attention-demanding cognitive tasks
Table 1Spatial correlations of single subject ICA derived networks to group-ICA derivednetworks
Network Mean STD ERR Range
A 0691 0068 0383ndash0841B 0560 0082 0176ndash0750C 0463 0062 0227ndash0649D 0501 0078 0215ndash0743E 0373 0046 0275ndash0584F 0459 0073 0184ndash0623G 0372 0060 0200ndash0586H 0559 0059 0272ndash0661I 0501 0065 0199ndash0655J 0375 0045 0207ndash0546K 0264 0017 0186ndash0302
Note Mean correlation values are significantly different from 0 (one sample t-testpb001)
1550 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
and in humans has been suggested to support higher-order mentalfaculties (Raichle et al 2001)
bull RSN D (occipito-temporal) Network areas included bilateral area TOV4 TEO and the arcuate sulcus Bilateral area 29 and Brodmann area10 (left hemisphere) were also functionally connected Previousstudies have shown these temporal lobe areas are critical for higher-order visual processing (Tsao et al 2003)
bull RSN E (frontal) This RSN encompassed bilateral regions in severalprefrontal areas including the anterior bank of the arcuate sulcuscorresponding to the frontal eye fields the upper ramus of thearcuate sulcus the posterior portion of the principal sulcus as wellas the dorsal bank of the principal sulcus In addition LFFsynchronization was also found in area 9 the premotor cortex andthe anterior cingulate cortex These frontalprefrontal areas havebeen shown to be components of the executive system suggested toprovide bias signals to other areas of the brain in order to implementcognitive control (Miller and Cohen 2001)
bull RSN F (superior-temporal) A network encompassing the auditory beltparabelt and bilateral area 22 on the superior temporal gyrus Anti-correlated areas were found in the left principal sulcus and in the leftarcuate sulcus The network pattern resembles the mean functionalactivity resulting from the presentation of multiple sounds categoriesin a previous task-based fMRI study of the awake monkey (Petkovet al 2008) and is likely responsible for acoustic processing andinterpretation (Rauschecker and Scott 2009)
bull RSN G (cingulo-insular) A network of areas including bilateral regionsin the insular cortex the anterior cingulate cortex area 24ab and theorbitofrontal cortex area 14 These areas have been associated withthe reward system involved in the regulation and control of behavior(Kringelbach 2005)
bull RSN H (paracentral) A network reflective of the dorsal motorsystem involved in the control of limb movements (Dum and Strick2002) Network connectivity included bilateral primary motorcortex dorsally and also in the central sulcus area F2 and thedorsal bank of the superior ramus of the arcuate sulcus In additionwidespread connectivity was found in themedial wall including thecingulate motor area supplementary motor area and medialparietal cortex The network pattern closely resembles the ldquosoma-tomotorrdquo network previously described in the anesthetized ma-caque using a seed-region based approach (Vincent et al 2007)
bull RSN I (parieto-occipital) A network consisting of regions involved invisual processing including bilateral areas V1 V2 V2 V3 area POand area MTMST Connectivity was also observed unilaterally inarea 8A (left hemisphere) and 46d (right hemisphere) Similarnetwork patterns have been observed in the macaque in bothresting-state (Vincent et al 2007) and awake fMRI studies(Stefanacci et al 1998)
bull RSN J (postcentral) A network including the postcentral andprecentral gyrus areas dedicated to somatosensory processing(Kaas 1993) Opposite modulation was found to occur in area POand in the anterior cingulate cortex area 24 ab
bull RSN K (hippocampal) A medial temporal network corresponding toareas associated with the macaque declarative memory system(Squire and Zola-Morgan 1991) The RSN bilaterally encompassedthe hippocampus entorhinal perirhinal and parahippocampalcortical areas Anti-correlated bilateral connectivity of anteriorcingulate cortex area 25 was also observed The network althoughpartially explored in several previous electrophysiological studies(eg Rolls et al 1993 Wirth et al 2003) has not been shown withmonkey fMRI possibly due to the difficult task demands required ofthe animals in order to elicit activations in these areas
Single subject ICA
To compare single subject ICA with group ICA components wespatially correlated unthresholded single subject IC maps with the
unthresholded group ICmaps The correlation coefficients for all mapsand all animals was significant (pb10minus20 p value corrected formultiple comparisons using Bonferroni correction) Table 1 shows themean correlation coefficients between the single-subject ICA compo-nents and the group-ICA networks Mean correlation values werefound to be significantly different from 0 (one sample t-test pb001)A representative network (RSN B mdash fronto-parietal) is shown for allmonkeys in Fig 2 Networks for all animals are shown in Supple-mentary Figs 4ndash9 In some animals the same single subjectcomponent was best correlated to two or more of the group ICAnetworks (for example Supplementary Fig 5 componentscorresponding to networks D and I are the same)
Functional network connectivity
Fig 3 shows a FNC diagram for the 11 identified RSNs RSNs arerepresented by circular nodes and significantly correlated RSNs arerepresented by connecting lines For example a line connectsnetworks I and J representing significant functional connectivitybetween those two networks RSN C (posterior-parietal) and RSN I(parieto-occipital) represent the most connected nodes RSN D(occipito-temporal) and RSN E (frontal) also show a high degree ofFNC The other sensory networks show little or no connectivity withother RSNs
Discussion
Alterations in functional connectivity recorded using spontaneousBOLD fluctuations have been suggested as the origin or product ofmultiple disease states (Auer 2008 Greicius 2008) Assessing theirelectrophysiological correlate(s) and establishing the relationshipbetween large-scale functional network connectivity and diseaserequire a suitable animal model Here to the best of our knowledgewe report the first comprehensive application of group independentcomponent analysis (ICA) to monkey fMRI data and the first resting-state examination of the macaque at 7 T ICA successfully identified 11prominent macaque RSNs representing multiple levels of neuralprocessing Networks encompassing sensory and motor areasincluding the visual auditory motor and somato-sensory regionscan be considered to be lower-order in a cognitive processinghierarchy The RSNs comprising areas known to be involved inexecutive control attention reward evaluation and default-modeactivity may represent higher-order processing with temporal net-works putatively responsible for visual processing and memoryproviding intermediate processing In cases where the system hasbeen explored with fMRI the patterns of intrinsic functionalconnectivity are consistent with stimulus-evoked patterns found intask-based studies (see Results) The functional RSNs reported in the
Fig 2 Resting-state networks (RSN) of all monkeys (M1ndashM6) following single-subject independent component analysis (ICA) that were most spatially correlated to group-ICAidentified RSN B (fronto-parietal) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 seehttpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of bothhemispheres and the right images depict the lateral and medial views of right hemisphere
Fig 3 Average functional network connectivity of macaque resting-state networks (RSNs) Lines and numerical values indicate functional connectivity between two RSNs in whichthere was a significant temporal correlation of their respective time-courses (one-sample Student t-test pb005 corrected for multiple comparisons between 55 pairs with a timeleadlag of +minus3 s) Network letters refer to spatial representations shown in Fig 1
1551RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

Table 1Spatial correlations of single subject ICA derived networks to group-ICA derivednetworks
Network Mean STD ERR Range
A 0691 0068 0383ndash0841B 0560 0082 0176ndash0750C 0463 0062 0227ndash0649D 0501 0078 0215ndash0743E 0373 0046 0275ndash0584F 0459 0073 0184ndash0623G 0372 0060 0200ndash0586H 0559 0059 0272ndash0661I 0501 0065 0199ndash0655J 0375 0045 0207ndash0546K 0264 0017 0186ndash0302
Note Mean correlation values are significantly different from 0 (one sample t-testpb001)
1550 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
and in humans has been suggested to support higher-order mentalfaculties (Raichle et al 2001)
bull RSN D (occipito-temporal) Network areas included bilateral area TOV4 TEO and the arcuate sulcus Bilateral area 29 and Brodmann area10 (left hemisphere) were also functionally connected Previousstudies have shown these temporal lobe areas are critical for higher-order visual processing (Tsao et al 2003)
bull RSN E (frontal) This RSN encompassed bilateral regions in severalprefrontal areas including the anterior bank of the arcuate sulcuscorresponding to the frontal eye fields the upper ramus of thearcuate sulcus the posterior portion of the principal sulcus as wellas the dorsal bank of the principal sulcus In addition LFFsynchronization was also found in area 9 the premotor cortex andthe anterior cingulate cortex These frontalprefrontal areas havebeen shown to be components of the executive system suggested toprovide bias signals to other areas of the brain in order to implementcognitive control (Miller and Cohen 2001)
bull RSN F (superior-temporal) A network encompassing the auditory beltparabelt and bilateral area 22 on the superior temporal gyrus Anti-correlated areas were found in the left principal sulcus and in the leftarcuate sulcus The network pattern resembles the mean functionalactivity resulting from the presentation of multiple sounds categoriesin a previous task-based fMRI study of the awake monkey (Petkovet al 2008) and is likely responsible for acoustic processing andinterpretation (Rauschecker and Scott 2009)
bull RSN G (cingulo-insular) A network of areas including bilateral regionsin the insular cortex the anterior cingulate cortex area 24ab and theorbitofrontal cortex area 14 These areas have been associated withthe reward system involved in the regulation and control of behavior(Kringelbach 2005)
bull RSN H (paracentral) A network reflective of the dorsal motorsystem involved in the control of limb movements (Dum and Strick2002) Network connectivity included bilateral primary motorcortex dorsally and also in the central sulcus area F2 and thedorsal bank of the superior ramus of the arcuate sulcus In additionwidespread connectivity was found in themedial wall including thecingulate motor area supplementary motor area and medialparietal cortex The network pattern closely resembles the ldquosoma-tomotorrdquo network previously described in the anesthetized ma-caque using a seed-region based approach (Vincent et al 2007)
bull RSN I (parieto-occipital) A network consisting of regions involved invisual processing including bilateral areas V1 V2 V2 V3 area POand area MTMST Connectivity was also observed unilaterally inarea 8A (left hemisphere) and 46d (right hemisphere) Similarnetwork patterns have been observed in the macaque in bothresting-state (Vincent et al 2007) and awake fMRI studies(Stefanacci et al 1998)
bull RSN J (postcentral) A network including the postcentral andprecentral gyrus areas dedicated to somatosensory processing(Kaas 1993) Opposite modulation was found to occur in area POand in the anterior cingulate cortex area 24 ab
bull RSN K (hippocampal) A medial temporal network corresponding toareas associated with the macaque declarative memory system(Squire and Zola-Morgan 1991) The RSN bilaterally encompassedthe hippocampus entorhinal perirhinal and parahippocampalcortical areas Anti-correlated bilateral connectivity of anteriorcingulate cortex area 25 was also observed The network althoughpartially explored in several previous electrophysiological studies(eg Rolls et al 1993 Wirth et al 2003) has not been shown withmonkey fMRI possibly due to the difficult task demands required ofthe animals in order to elicit activations in these areas
Single subject ICA
To compare single subject ICA with group ICA components wespatially correlated unthresholded single subject IC maps with the
unthresholded group ICmaps The correlation coefficients for all mapsand all animals was significant (pb10minus20 p value corrected formultiple comparisons using Bonferroni correction) Table 1 shows themean correlation coefficients between the single-subject ICA compo-nents and the group-ICA networks Mean correlation values werefound to be significantly different from 0 (one sample t-test pb001)A representative network (RSN B mdash fronto-parietal) is shown for allmonkeys in Fig 2 Networks for all animals are shown in Supple-mentary Figs 4ndash9 In some animals the same single subjectcomponent was best correlated to two or more of the group ICAnetworks (for example Supplementary Fig 5 componentscorresponding to networks D and I are the same)
Functional network connectivity
Fig 3 shows a FNC diagram for the 11 identified RSNs RSNs arerepresented by circular nodes and significantly correlated RSNs arerepresented by connecting lines For example a line connectsnetworks I and J representing significant functional connectivitybetween those two networks RSN C (posterior-parietal) and RSN I(parieto-occipital) represent the most connected nodes RSN D(occipito-temporal) and RSN E (frontal) also show a high degree ofFNC The other sensory networks show little or no connectivity withother RSNs
Discussion
Alterations in functional connectivity recorded using spontaneousBOLD fluctuations have been suggested as the origin or product ofmultiple disease states (Auer 2008 Greicius 2008) Assessing theirelectrophysiological correlate(s) and establishing the relationshipbetween large-scale functional network connectivity and diseaserequire a suitable animal model Here to the best of our knowledgewe report the first comprehensive application of group independentcomponent analysis (ICA) to monkey fMRI data and the first resting-state examination of the macaque at 7 T ICA successfully identified 11prominent macaque RSNs representing multiple levels of neuralprocessing Networks encompassing sensory and motor areasincluding the visual auditory motor and somato-sensory regionscan be considered to be lower-order in a cognitive processinghierarchy The RSNs comprising areas known to be involved inexecutive control attention reward evaluation and default-modeactivity may represent higher-order processing with temporal net-works putatively responsible for visual processing and memoryproviding intermediate processing In cases where the system hasbeen explored with fMRI the patterns of intrinsic functionalconnectivity are consistent with stimulus-evoked patterns found intask-based studies (see Results) The functional RSNs reported in the
Fig 2 Resting-state networks (RSN) of all monkeys (M1ndashM6) following single-subject independent component analysis (ICA) that were most spatially correlated to group-ICAidentified RSN B (fronto-parietal) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 seehttpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of bothhemispheres and the right images depict the lateral and medial views of right hemisphere
Fig 3 Average functional network connectivity of macaque resting-state networks (RSNs) Lines and numerical values indicate functional connectivity between two RSNs in whichthere was a significant temporal correlation of their respective time-courses (one-sample Student t-test pb005 corrected for multiple comparisons between 55 pairs with a timeleadlag of +minus3 s) Network letters refer to spatial representations shown in Fig 1
1551RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18
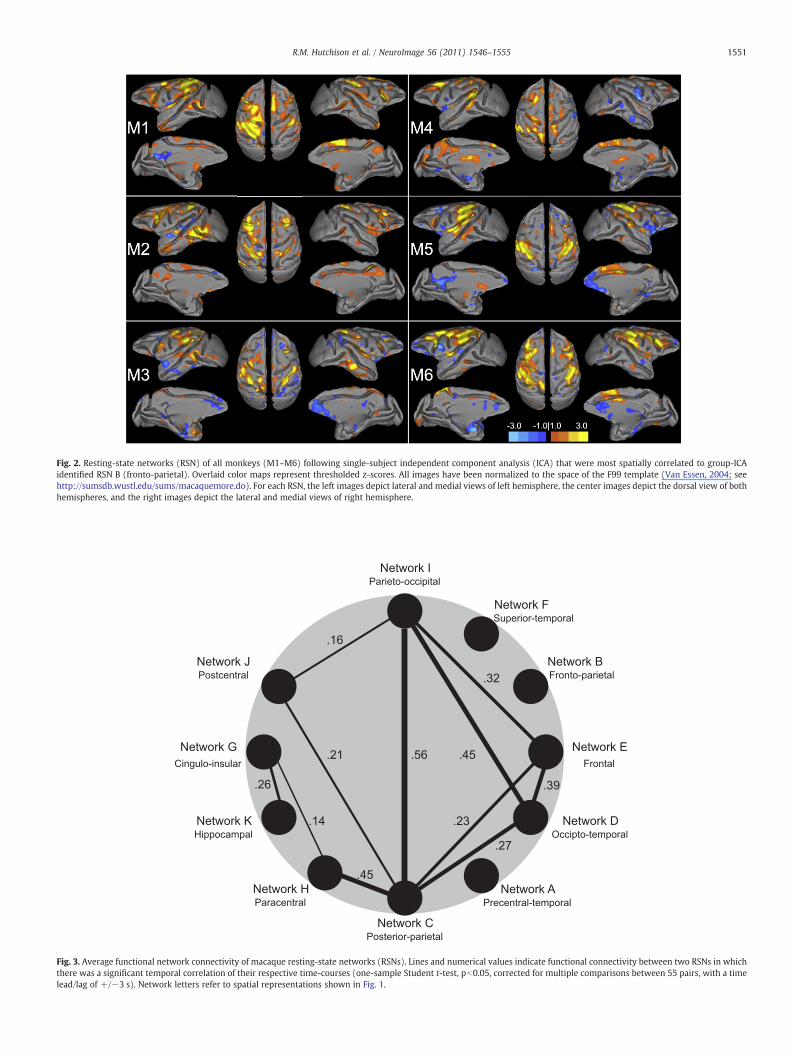
Fig 2 Resting-state networks (RSN) of all monkeys (M1ndashM6) following single-subject independent component analysis (ICA) that were most spatially correlated to group-ICAidentified RSN B (fronto-parietal) Overlaid color maps represent thresholded z-scores All images have been normalized to the space of the F99 template (Van Essen 2004 seehttpsumsdbwustledusumsmacaquemoredo) For each RSN the left images depict lateral and medial views of left hemisphere the center images depict the dorsal view of bothhemispheres and the right images depict the lateral and medial views of right hemisphere
Fig 3 Average functional network connectivity of macaque resting-state networks (RSNs) Lines and numerical values indicate functional connectivity between two RSNs in whichthere was a significant temporal correlation of their respective time-courses (one-sample Student t-test pb005 corrected for multiple comparisons between 55 pairs with a timeleadlag of +minus3 s) Network letters refer to spatial representations shown in Fig 1
1551RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

1552 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
current study revealed highly similar possibly homologous macro-scopic brain organization between macaques and humans RSNs B(fronto-parietal) C (posterior-parietal) D (occipito-temporal) E(frontal) F (superior-temporal) H (paracentral) and I (parieto-occipital) have been commonly reported following ICA of humanresting-state data (Beckmann et al 2005 Jafri et al 2008 Smithet al 2009) These have been labeled oculomotordorsal attention(see Figs 6g h of Beckmann et al 2005) default-mode (see Fig 6e ofBeckmann et al 2005) higher-order visual (see Fig 6b of Beckmannet al 2005) executive (see Fig 6f of Beckmann et al 2005) auditory(see Fig 6c of Beckmann et al 2005) somatomotor (see Fig 6d ofBeckmann et al 2005) and primary visual (see Fig 6a of Beckmannet al 2005) networks respectively Further RSNs G (cingulo-insular)J (postcentral) and K (hippocampal) though not consistently reportedin studies utilizing ICA have homologous networks that can be foundwhen comparing task-based or seed-region analysis of human fMRIdata (Kringelbach 2005 Blatow et al 2007 Burton et al 2009Vincent et al 2006)
Two recent studies have examined resting-state connectivity withhypotheses derived from experimental anterograde tracer studies ofthe macaque monkey (Margulies et al 2009 Kelly et al 2010) Kellyet al (2010) examined the connectivity of ventrolateral frontal areaswith parietal and temporal cortex in the human cortex They foundthat the human brain maintains the same basic patterns observed innonhuman primates (Petrides and Pandya 2009) In the same wayMargulies et al (2009) found functional subdivisions of the precuneusto be preserved between both species and also consistent with tracerstudies (Pandya and Seltzer 1982) These findings suggest thatresting-state functional connectivity reflects the underlying structuralanatomy (discussed below) and taken together with our currentresults support the role of the macaque as a suitable animal model inthe study of human brain organization and cross-species comparisonsof functional neuroanatomy
Despite the strong similarities in many of the macaque RSNs tohuman RSNs our study also revealed a number of notable differencesin RSNs between the two species There was an absence of twocommonly reported lateralized fronto-parietal RSNs implicated incognitive attentional processes as well as memory and languagefunctions (Beckmann et al 2005 Jafri et al 2008 Smith et al 2009)The homologous macaque network RSN B (frontoparietal) thoughencompassing the same brain regions as the human networks wasrelatively symmetrical and did not suggest lateralization of functionGiven that the macaque brain appears to be less lateralized than thehuman brain (Kagan et al 2010) it is possible that the networkrepresents the evolutionary predecessor to the lateralized humannetworks Another functional connectivity difference was the lack ofthe dorsal medial prefrontal cortical (dmPFC) component of the DMN(Raichle et al 2001) A relatively weak network connection has beenpreviously reported (Vincent et al 2010) though it was absent in twoother studies (Vincent et al 2007 Teichert et al 2010) It is difficultto determine whether the dmPFC represents a less connectedweaklysynchronized area of the DMN in the macaque a brain area morevulnerable to BOLD fMRI artifactual sources (possibly due to theproximity to the eyes) an area highly sensitive to anesthesia level orphysiological variability in connectivity between animals Finally RSNK (hippocampal) shares a similar network pattern to the humanmemory RSN however there is an absence of parietal connectivitywithin the network that is found in the human brain (Vincent et al2006) RSN A (precentralndashtemporal) has not been reported as aseparate network in resting-state or task-based studies of humansthough the brain areas are implicated in more diffuse networkpatterns (Peeters et al 2009) The ventral motor areas are critical forgoal directed movements ndash particularly of the hand ndash in both species(Joly et al 2009 Callaert et al in press) In humans however there isa lateralization of motor function in which there is increasedinvolvement of left motor areas resulting in functional asymmetries
(Callaert et al in press) Monkeys show a weaker motor dominancethan humans (Leca et al 2010) and the interhemispheric connectivityof the ventral motor system is supported by strong callosalconnections between the homotopic functional areas These factorscould explain why a bilateral ventral motor RSN may not be found inthe human at lower ICA model orders (20ndash40 ICs) as the synchroni-zation between the systems might not be as tightly coupled
RSNs are inferred from endogenous neural activity and theirorganization is likely shaped by structural connections (Sporns2010) There is increasing evidence to suggest that patterns ofsynchronous LFFs track underlying anatomical connectivity (Vincentet al 2007 Hagmann et al 2008 Skudlarski et al 2008 Honey et al2009 Margulies et al 2009 Kelly et al 2010) Anatomicalconnectivity may underlie some of the RSNs that we identified inthe present study In some cases the observed RSNs are in goodagreement with the known major bundles of fibers that connectsensory association areas of posterior cortex to frontal cortex RSN B(fronto-parietal) contains areas in frontal and parietal cortex that areconnected by subcomponents II and III of the superior longitudinalfasciculus and RSN C (posterior-parietal) is comprised of areas thatare connected by the fronto-occipital fasciculus (Petrides and Pandya2002b) Structural connectivity via the uncinate fasciculus mayunderlie RSN G (cingulo-insular) (Petrides and Pandya 2002b) Atleast some of the areas in RSN D (occipito-temporal) are anatomicallyconnected though the inferior longitudinal fasciculus (Petrides andPandya 2002b) Some of the other RSNs may reflect knownconnectivity by intrinsic connections (RSN E (Barbas and Pandya1989) RSN F (Pandya 1995) RSN H (Vogt and Pandya 1978))
We also observed the opposite pattern in which little to nofunctional connectivity between hippocampal and parietal areas wasobserved whereas tracer methodology has revealed substantialconnectivity between parietal and parahippocampal regions (Rock-land and Van Hoesen 1999) Similarly RSN F (auditory) did notcontain any prefrontal areas despite the known connections of thisarea with ventral and dorsal areas through the extreme capsule(Petrides and Pandya 2002b) It is important to note however thelimitations of the data analysis when discussing the possibleconclusions that are being drawn in terms of lateralization connec-tivity and absent homologous networks Despite the link to structuralorganization resting-state connectivity is not anatomical connectiv-ity The patterns of functional networks have been shown to capturepolysynaptic connections (Vincent et al 2007 OReilly et al 2009)and functional connections have been shown where no directstructural connections exist (Uddin et al 2008 Vincent et al 2008Honey et al 2009) The discrepancies further highlight the need foran animal model as a method to constrain and interpret the falsepresence and absence of known connectivity in the human brainfound using diffusion tensor imaging and resting-state methods Onlytracer studies can address direct connections and these are nottypically feasible in human investigations
A previous report examining changes in functional connectivity ofthe macaque during various visual stimulus contexts also examinedtwo monkeys at rest under ketamine anesthesia using single-subjectICA (Moeller et al 2009) Similar to the present study networksencompassing primary sensory areas including auditory and visualsystems were identified These and other networks were typicallybilateral only comprising hemispheric functional homologous aproperty also shown in rats (Hutchison et al 2010) Our group datamore closely resemble RSN organization of humans in which multiplesubdivisions of a system are functionally connected Functionallyconnected regions can split into separate components at highermodelorders (Abou-Elseoud et al 2010) a property that reflects thehierarchical functional organization of the brain (Cole et al 2010)Moeller and colleagues used a model order of 300ndash1000 ICs whereasin the current study we used a model order of 20 The relatively largeestimate could overestimate the number of networks and the use of
1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

1553RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
an automated sorting algorithm specifying bilateralism could bias theresults towards those reported in the study
Todelineate the effects of single-subject ICA andmodel order single-subject ICA was conducted on our same data set using a model order of20 The individual componentmaps varied in their spatial correlation tothe group RSNs within and between animals The differences couldreflect individual differences in morphology structural connectivity orfunctional connectivity as well as an exacerbation of weaker spatialdependencies causing the ICA to decompose the data differentlyVariability could also arise from noise sources unique to each animalthat were not extracted as a unique component using group-ICA Theresults also highlighted intersubject variability in regards to networkdecomposition as often single subject networks comprised areasencompassed by two (or more) group RSNs Overall the individualnetworks had more diffuse functionally connectivity patterns thangroup-ICA and considerably more connectivity than the small-scalenetworks previously identified (Moeller et al 2009) Therefore thedifferences between our study and thework of Moeller et al (2009) aremore likely the result of a lower model order than group-level analysisIt is important to note however that given the methodologicaldifferences including model order and anesthetic regime a directcomparison between these studies is not possible
Various network analysis strategies examining human and non-human primate brain connectivity have revealed that the cortexcontains a small number of nodes having a disproportionately highnumber of connections (Sporns et al 2007 Hagmann et al 2008Buckner et al 2009) These highly connected nodes are referred to ashubs and serve to integrate diverse informational sources enablingglobally efficient information flow (Sporns et al 2007) Hubs alsofacilitate small-world network organization minimizing wiring andmetabolic costs by providing long-distance connections that integratelocal networks (Bassett and Bullmore 2006) The locations of highfunctional centrality have a close correspondencewith structural hubs(Honey et al 2007 Hagmann et al 2008 Buckner et al 2009) Thepresent study revealed substantial inter-network functional connec-tivity with RSN C (posterior-parietal) and RSN I (parieto-occipital)The precuneousposterior cingulate cortex areas encompassed by RSNC have been previously shown to possess both structural andfunctional hub properties in the human brain (Hagmann et al2008 Buckner et al 2009) and could play a substantial role inintegrating or regulating activity of other RSNs particularly at rest(Greicius et al 2003) A detailed analysis of the structural connec-tivity of the macaque cortex based on tracer studies suggested severalstructural hubs including frontal area 46 and visual area V4 (Honeyet al 2007 Sporns et al 2007) mdash areas encompassed by RSN I Theseareas have been classified as association or integrative areas againreflecting their hub like properties Thus the results of the functionalnetwork connectivity analysis fit the current framework of knownfunctional organization and further support the use of resting-statedata in the evaluation of large-scale network dynamics and the use ofFNC measures It is important to note that like other functionalconnections hubs have been shown to engage and disengage acrosstime mdash dynamically altering their centrality (Honey et al 2007)Evaluation of resting-state functional connectivity could provide anappropriatemethod to characterize the process bywhich the topologyof functional networks changes over time
Conclusions
In summary our results demonstrate that ICA can identify RSNs inmacaque monkeys that are likely homologous to those found inhumans thereby strongly supporting the use of monkeys as an idealanimal model for human brain function (Passingham 2009) whilealso reinforcing the use of resting-state functional connectivity indelineating complex neural circuits in vivo
Supplementarymaterials related to this article can be found onlineat doi101016jneuroimage201102063
Acknowledgments
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to SE and a scholarship from theNatural ScienceandEngineeringResearchCouncil (NSERC) to RMHWe thank S Hughesand B Soper for technical assistance
References
Abou-Elseoud A Starck T Remes J Nikkinen J Tervonen O Kiviniemi V 2010 Theeffect of model order selection in group PICA Hum Brain Mapp 31 1207ndash1216
Auer DP 2008 Spontaneous low-frequency blood oxygenation level-dependentfluctuations and functional connectivity analysis of the lsquorestingrsquo brain Magn ResonImaging 26 1055ndash1064
Baker JT Patel GH Corbetta M Snyder LH 2006 Distribution of activity across themonkey cerebral cortical surface thalamus and midbrain during rapid visuallyguided saccades Cereb Cortex 16 447ndash459
Barbas H Pandya DN 1989 Architecture and intrinsic connections of the prefrontalcortex in the rhesus monkey J Comp Neurol 286 353ndash375
Bassett DS Bullmore E 2006 Small-world brain networks Neuroscientist 12512ndash523
Beckmann CF DeLuca M Devlin JT Smith SM 2005 Investigations into resting-state connectivity using independent component analysis Philos Trans R SocLond B Biol Sci 360 1001ndash1013
Biswal B Yetkin FZ Haughton VM Hyde JS 1995 Functional connectivity in themotor cortex of resting human brain using echo-planar MRI Magn Reson Med 34537ndash541
Blatow M Nennig E Durst A Sartor K Stippich C 2007 fMRI reflects functionalconnectivity of human somatosensory cortex Neuroimage 37 927ndash936
Bruce CJ Goldberg ME 1985 Primate frontal eye fields I Single neurons dischargingbefore saccades J Neurophysiol 53 603ndash635
Buckner RL Sepulcre J Talukdar T Krienen FM Liu H Hedden T Andrews-Hanna JR Sperling RA Johnson KA 2009 Cortical hubs revealed by intrinsicfunctional connectivity mapping assessment of stability and relation toAlzheimers disease J Neurosci 29 1860ndash1873
Burton H Dixit S Litkowski P Wingert JR 2009 Functional connectivity forsomatosensory and motor cortex in spastic diplegia Somatosens Mot Res 2690ndash104
Buxhoeveden DP Switala AE Roy E Litaker M Casanova MF 2001 Morpholog-ical differences between minicolumns in human and nonhuman primate cortexAm J Phys Anthropol 115 361ndash371
Calhoun VD Adali T Pearlson GD Pekar JJ 2001 A method for making groupinferences from functional MRI data using independent component analysis HumBrain Mapp 14 140ndash151
Calhoun VD Adali T Pekar JJ Pearlson GD 2003 Latency (in) sensitive ICA groupindependent component analysis of fMRI data in the temporal frequency domainNeuroimage 20 1661ndash1669
Callaert DV Vercauteren K Peeters R Tam F Graham S Swinnen SP Sunaert SWenderoth N in press Hemispheric asymmetries of motor versus nonmotorprocesses during (visuo)motor control Hum Brain Mapp
Cole DM Smith SM Beckmann CF 2010 Advances and pitfalls in the analysis andinterpretation of resting-state FMRI data Front Syst Neurosci 4 8
Cordes D Haughton VM Arfanakis K Wendt GJ Turski PA Moritz CH QuigleyMA Meyerand ME 2000 Mapping functionally related regions of brain withfunctional connectivity MR imaging Am J Neuroradiol 21 1636ndash1644
Croxson PL Johansen-Berg H Behrens TE Robson MD Pinsk MA Gross CGRichter W Richter MC Kastner S Rushworth MF 2005 Quantitativeinvestigation of connections of the prefrontal cortex in the human and macaqueusing probabilistic diffusion tractography J Neurosci 25 8854ndash8866
Damoiseaux JS Rombouts SA Barkhof F Scheltens P Stam CJ Smith SMBeckmann CF 2006 Consistent resting-state networks across healthy subjectsProc Natl Acad Sci USA 103 13848ndash13853
Demirci O Stevens MC Andreasen NC Michael A Liu J White T Pearlson GDClark VP Calhoun VD 2009 Investigation of relationships between fMRI brainnetworks in the spectral domain using ICA and Granger causality reveals distinctdifferences between schizophrenia patients and healthy controls Neuroimage 46419ndash431
Dum RP Strick PL 2002 Motor areas in the frontal lobe of the primate PhysiolBehav 77 677ndash682
Farber NE Harkin CP Niedfeldt J Hudetz AG Kampine JP Schmeling WT 1997Region-specific and agent-specific dilation of intracerebral microvessels by volatileanesthetics in rat brain slices Anesthesiology 87 1191ndash1198
Felleman DJ Van Essen DC 1991 Distributed hierarchical processing in the primatecerebral cortex Cereb Cortex 1 1ndash47
Ford KA Gati JS Menon RS Everling S 2009 BOLD fMRI activation for anti-saccades in nonhuman primates Neuroimage 45 470ndash476
Fox MD Raichle ME 2007 Spontaneous fluctuations in brain activity observed withfunctional magnetic resonance imaging Nat Rev Neurosci 8 700ndash711
Friston KJ 1994 Functional and effective connectivity in neuroimaging a synthesisHum Brain Mapp 2 56ndash78
1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

1554 RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Friston KJ 2002 Beyond phrenology what can neuroimaging tell us about distributedcircuitry Annu Rev Neurosci 25 221ndash250
Greicius M 2008 Resting-state functional connectivity in neuropsychiatric disordersCurr Opin Neurol 21 424ndash430
Greicius MD Krasnow B Reiss AL Menon V 2003 Functional connectivity in theresting brain a network analysis of the default mode hypothesis Proc Natl AcadSci USA 100 253ndash258
Greicius MD Supekar K Menon V Dougherty RF 2009 Resting-state functionalconnectivity reflects structural connectivity in the default mode network CerebCortex 19 72ndash78
Guye M Bartolomei F Ranjeva JP 2008 Imaging structural and functionalconnectivity towards a unified definition of human brain organization CurrOpin Neurol 21 393ndash403
Hagmann P Cammoun L Gigandet X Meuli R Honey CJ Wedeen VJ SpornsO 2008 Mapping the structural core of human cerebral cortex PLoS Biol 6e159
Hampson M Peterson BS Skudlarski P Gatenby JC Gore JC 2002 Detection offunctional connectivity using temporal correlations in MR images Hum BrainMapp 15 247ndash262
Himberg J Hyvarinen A Esposito F 2004 Validating the independent components ofneuroimaging time series via clustering and visualization Neuroimage 221214ndash1222
Honey CJ Koumltter R Breakspear M Sporns O 2007 Network structure of cerebralcortex shapes functional connectivity on multiple time scales Proc Natl Acad SciUSA 104 10240ndash10245
Honey CJ Sporns O Cammoun L Gigandet X Thiran JP Meuli R Hagmann P2009 Predicting human resting-state functional connectivity from structuralconnectivity Proc Natl Acad Sci USA 106 2035ndash2040
Hutchison RM Mirsattari SM Jones CK Gati JS Leung LS 2010 Functionalnetworks in the anesthetized rat brain revealed by independent componentanalysis of resting-state fMRI J Neurophysiol 103 3398ndash3406
Jafri MJ Pearlson GD Stevens M Calhoun VD 2008 A method for functionalnetwork connectivity among spatially independent resting-state components inschizophrenia Neuroimage 39 1666ndash1681
Johnston K Everling S 2008 Neurophysiology and neuroanatomy of reflexive andvoluntary saccades in non-human primates Brain Cogn 68 271ndash283
Joly O Vanduffel W Orban GA 2009 The monkey ventral premotor cortexprocesses 3D shape from disparity Neuroimage 47 262ndash272
Kaas JH 1993 The functional organization of somatosensory cortex in primates AnnAnat 175 509ndash518
Kagan I Iyer A Lindner A Andersen RA 2010 Space representation for eyemovements is more contralateral in monkeys than in humans Proc Natl Acad SciUSA 107 7933ndash7938
Kelly C Uddin LQ Shehzad Z Margulies DS Castellanos FX Milham MPPetrides M 2010 Brocas region linking human brain functional connectivitydata and non-human primate tracing anatomy studies Eur J Neurosci 32383ndash398
Kiviniemi VJ Haanpaumlauml H Kantola JH Jauhiainen J Vainionpaumlauml V Alahuhta STervonen O 2005 Midazolam sedation increases fluctuation and synchrony of theresting brain BOLD signal Magn Reson Imaging 23 531ndash537
Koyama M Hasegawa I Osada T Adachi Y Nakahara K Miyashita Y 2004Functional magnetic resonance imaging of macaque monkeys performing visuallyguided saccade tasks comparison of cortical eye fields with humans Neuron 41795ndash807
Kringelbach ML 2005 The human orbitofrontal cortex linking reward to hedonicexperience Nat Rev Neurosci 6 691ndash702
Leca JB Gunst N Huffman MA 2010 Principles and levels of laterality in unimanualand bimanual stone handling patterns by Japanese macaques J Hum Evol 58155ndash165
Li YO Adali T Calhoun VD 2007 Estimating the number of independentcomponents for functional magnetic resonance imaging data Hum Brain Mapp28 1251ndash1266
Liao R McKeown MJ Krolik JL 2006 Isolation and minimization of head motion-induced signal variations in fMRI data using independent component analysisMagn Reson Med 55 1396ndash1413
Margulies DS Vincent JL Kelly C Lohmann G Uddin LQ Biswal BB VillringerA Castellanos FX Milham MP Petrides M 2009 Precuneus shares intrinsicfunctional architecture in humans and monkeys Proc Natl Acad Sci USA 10620069ndash20074
Miller EK Cohen JD 2001 An integrative theory of prefrontal cortex function AnnuRev Neurosci 24 167ndash202
Moeller S Nallasamy N Tsao DY Freiwald WA 2009 Functional connectivityof the macaque brain across stimulus and arousal states J Neurosci 295897ndash5909
Nakahara K Adachi Y Osada T Miyashita Y 2007 Exploring the neural basis ofcognition multi-modal links between human fMRI and macaque neurophysiologyTrends Cogn Sci 11 84ndash92
Nelissen K Luppino G Vanduffel W Rizzolatti G Orban GA 2005 Observingothers multiple action representation in the frontal lobe Science 310 332ndash336
Noudoost B Chang MH Steinmetz NA Moore T 2010 Top-down control of visualattention Curr Opin Neurobiol 20 183ndash190
OReilly JX Beckmann CF Tomassini V Ramnani N Johansen-Berg H 2009Distinct and overlapping functional zones in the cerebellum defined by restingstate functional connectivity Cereb Cortex 20 953ndash965
Onguumlr D Ferry AT Price JL 2003 Architectonic subdivision of the human orbitaland medial prefrontal cortex J Comp Neurol 460 425ndash449
Orban GA Van Essen D Vanduffel W 2004 Comparative mapping of higher visualareas in monkeys and humans Trends Cogn Sci 8 315ndash324
Pandya DN 1995 Anatomy of the auditory cortex Rev Neurol (Paris) 151486ndash494
Pandya DN Seltzer B 1982 Intrinsic connections and architectonics of posteriorparietal cortex in the rhesus monkey J Comp Neurol 204 196ndash210
Passingham R 2009 How good is the macaque monkey model of the human brainCurr Opin Neurobiol 19 6ndash11
Peeters R Simone L Nelissen K Fabbri-Destro M Vanduffel W Rizzolatti GOrban GA 2009 The representation of tool use in humans andmonkeys commonand uniquely human features J Neurosci 29 11523ndash11539
Perlbarg V Bellec P Anton JL Peacuteleacutegrini-Issac M Doyon J Benali H 2007CORSICA correction of structured noise in fMRI by automatic identification of ICAcomponents Magn Reson Imaging 25 35ndash46
Petkov CI Kayser C Steudel T Whittingstall K Augath M Logothetis NK 2008 Avoice region in the monkey brain Nat Neurosci 11 367ndash374
Petrides M Pandya DN 1999 Dorsolateral prefrontal cortex comparative cytoarch-itectonic analysis in the human and the macaque brain and corticocorticalconnection patterns Eur J Neurosci 11 1011ndash1036
Petrides M Pandya DN 2002a Comparative cytoarchitectonic analysis of the humanand the macaque ventrolateral prefrontal cortex and corticocortical connectionpatterns in the monkey Eur J Neurosci 16 291ndash310
Petrides M Pandya DN 2002b Association pathways of the prefrontal cortex andfunctional observations In Stuss DT Knight RT (Eds) Principles of Frontal LobeFunction Oxford University Press New York NY pp 31ndash84
Petrides M Pandya DN 2009 Distinct parietal and temporal pathways to thehomologues of Brocas area in the monkey PLoS Biol 7 e1000170
Petrides M Cadoret G Mackey S 2005 Orofacial somatomotor responses in themacaque monkey homologue of Brocas area Nature 435 1235ndash1238
Phillips CG Porter R 1977 Corticospinal Neurons Academic Press LondonPreuss TM 2000 Taking the measure of diversity comparative alternatives to the
model-animal paradigm in cortical neuroscience Brain Behav Evol 55287ndash329
Preuss TM 2004 Specializations of the human visual system the monkey modelmeets human reality In Kaas JH Collins CE (Eds) The Primate Visual SystemCRC Press pp 231ndash259
Raichle ME MacLeod AM Snyder AZ Powers WJ Gusnard DA Shulman GL2001 A default mode of brain function Proc Natl Acad Sci USA 98 676ndash682
Rauschecker JP Scott SK 2009 Maps and streams in the auditory cortex nonhumanprimates illustrate human speech processing Nat Neurosci 12 718ndash724
Rees G Friston K Koch C 2000 A direct quantitative relationship between thefunctional properties of human and macaque V5 Nat Neurosci 3 716ndash723
Rilling J 2006 Human and nonhuman primate brains are they allometrically scaledversions of the same design Evol Anthropol 15 65ndash77
Rockland KS Van Hoesen GW 1999 Some temporal and parietal cortical connectionsconverge in CA1 of the primate hippocampus Cereb Cortex 9 232ndash237
Rolls ET Cahusac PM Feigenbaum JD Miyashita Y 1993 Responses of singleneurons in the hippocampus of the macaque related to recognition memory ExpBrain Res 93 299ndash306
Schlag J Schlag-Rey M 1987 Evidence for a supplementary eye field J Neurophysiol57 179ndash200
Shmuel A Leopold DA 2008 Neuronal correlates of spontaneous fluctuations in fMRIsignals in monkey visual cortex implications for functional connectivity at restHum Brain Mapp 29 751ndash761
Skudlarski P Jagannathan K Calhoun VD Hampson M Skudlarska BA PearlsonG 2008 Measuring brain connectivity diffusion tensor imaging validates restingstate temporal correlations Neuroimage 43 554ndash561
Smith SM Fox PT Miller KL Glahn DC Fox PM Mackay CE Filippini NWatkins KE Toro R Laird AR Beckmann CF 2009 Correspondence of thebrains functional architecture during activation and rest Proc Natl Acad Sci USA106 13040ndash13045
Sporns O 2010 Networks of the Brain MIT Press CambridgeSporns O Tononi G Koumltter R 2005 The human connectome a structural description
of the human brain PLoS Comput Biol 1 245ndash251Sporns O Honey CJ Koumltter R 2007 Identification and classification of hubs in brain
networks PLoS ONE 2 e1049Squire LR Zola-Morgan S 1991 The medial temporal lobe memory system Science
253 1380ndash1386Stefanacci L Reber P Constanza J Wong E Buxton R Zola S Squire L Albright T
1998 fMRI of monkey visual cortex Neuron 20 1051ndash1057Teichert T Grinband J Hirsch J Ferrera VP 2010 Effects of heartbeat and
respiration on macaque fMRI implications for functional connectivity Neuropsy-chologia 48 1886ndash1894
Thomas CG Harshman RA Menon RS 2002 Noise reduction in BOLD-based fMRIusing component analysis Neuroimage 17 1521ndash1537
Tsao DY Freiwald WA Knutsen TA Mandeville JB Tootell RB 2003 Faces andobjects in macaque cerebral cortex Nat Neurosci 6 989ndash995
Uddin LQ Mooshagian E Zaidel E Scheres A Margulies DS Kelly AM ShehzadZ Adelstein JS Castellanos FX Biswal BB Milham MP 2008 Residualfunctional connectivity in the split-brain revealed with resting-state functionalMRI NeuroReport 19 703ndash709
Van Essen DC 2004 Surface-based approaches to spatial localization and registrationin primate cerebral cortex Neuroimage 23 (Suppl 1) S97ndashS107
Vincent JL Snyder AZ Fox MD Shannon BJ Andrews JR Raichle ME BucknerRL 2006 Coherent spontaneous activity identifies a hippocampalndashparietalmemory network J Neurophysiol 96 3517ndash3531
1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18

1555RM Hutchison et al NeuroImage 56 (2011) 1546ndash1555
Vincent JL Patel GH Fox MD Snyder AZ Baker JT Van Essen DC Zempel JMSnyder LH Corbetta M Raichle ME 2007 Intrinsic functional architecture inthe anaesthetized monkey brain Nature 447 83ndash86
Vincent JL Kahn I Snyder AZ Raichle ME Buckner RL 2008 Evidence for afrontoparietal control system revealed by intrinsic functional connectivity J Neuro-physiol 100 3328ndash3342
Vincent JL Kahn I Van Essen DC Buckner RL 2010 Functional connectivity of themacaque posterior parahippocampal cortex J Neurophysiol 103 793ndash800
Vogt BA Pandya DN 1978 Cortico-cortical connections of somatic sensorycortex (areas 3 1 and 2) in the rhesus money J Comp Neurol 177179ndash192
Wirth S Yanike M Frank LM Smith AC Brown EN Suzuki WA 2003 Singleneurons in the monkey hippocampus and learning of new associations Science300 1578ndash1581
Young MP 1993 The organization of neural systems in the primate cerebral cortexProc R Soc Lond B 252 13ndash18
Copyright © 2022 FDOKUMEN




















