
Responses of four Indo-West Pacific seagrass species to shading
13
Responses of four Indo-West Pacific seagrass species to shading Catherine J. Collier ⇑ , Michelle Waycott, Ana Giraldo Ospina School of Marine and Tropical Biology, James Cook University, Townsville, Queensland 4811, Australia article info Keywords: Shading Water quality Great Barrier Reef Monitoring Submerged aquatic vegetation (SAV) abstract Seagrasses of the Great Barrier Reef predominantly occur in coastal regions where terrestrial inputs modify water quality and photosynthetic light is highly variable. Responses to shading were tested for Cymodocea serrulata, Halodule uninervis, Thalassia hemprichii and Zostera muelleri. In aquaria, four light treatments – high (66% surface light), moderate (31%), low (14%) and very low light (1%) treatments – were applied for 102 d. Stress responses in the low and very low light treatments occurred in the follow- ing sequence: metabolic and physiological changes (reduced growth, increased pigment concentrations and photosynthetic efficiency); shedding (leaf loss, shoot loss) and production of new, altered tissue (leaves with reduced length, width and thickness). Complete shoot loss was projected after 76 (Z. muel- leri) to 130 d (T. hemprichii). Responses were slower in the low than in the very low treatment, therefore, efforts to minimize water quality degradation will be rewarded with delayed impacts to seagrasses. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction The productivity, extent and ecological value of seagrass mead- ows have become threatened globally due to increasing anthropo- genic pressures, with reduced light penetration resulting from poor water quality being amongst the most significant of impacts (Waycott et al., 2009). The extreme nature of the impact in marine and aquatic environments is unique, as the episodes of light reduc- tion can be very severe (down to 0% light) and very prolonged (many months, Campbell and McKenzie, 2004; Onuf, 1996; Preen et al., 1995). In addition, chronic declines in water quality have led to broad-scale reductions in photosynthetic light available to benthic primary producers around many coastlines (Fabricius, 2011). The combination of these acute (localized) and chronic (broad-scale) reductions in light have led to world-wide loss of seagrass meadows (Rasheed and Unsworth, 2011; Waycott et al., 2009). In the Great Barrier Reef, water quality in inshore areas has declined since European settlement, but the region of greatest impact is south of Cairns and north of the Burdekin River, which experiences the greatest frequency of exposure to water quality pollutants that reduce photosynthetic light to benthic primary producers (Devlin et al., 2012). In some circumstances, such as dredging, controls can be put in place to minimize the impact and manage the consequences for adjacent seagrass meadows (Erftemeijer and Robin Lewis Iii, 2006; Sofonia and Unsworth, 2009). However, for most seagrass species, the thresholds of light reduction are not known, hampering the ability of coastal resource managers to identify impacts and apply appropriate mitigation procedures (Ralph et al., 2007). Seagrasses have adapted somewhat to their highly variable light environment and they respond in ways that provide tolerance to short-term periods of light reduction. This is partially achieved by balancing carbon supply and respiratory requirements (Ralph et al., 2007). There are a number of possible changes that can help to achieve this balance: both photosynthetic light capture and pho- tosynthetic efficiency can be increased (Abal et al., 1994; Durako et al., 2003; Enríquez, 2005); carbohydrate reserves can be re-allo- cated from storage organs, such as the rhizome, to respiration and growth; and respiratory requirements can be reduced by slowing growth rates and reducing the amount of respiratory-demanding biomass through morphological and meadow-scale changes (Collier et al., 2009; Fourqurean and Zieman, 1991). At sub-lethal light levels, these changes may be permanent enabling long-term survival, but at lethal light levels (below minimum light require- ments) die-off is inevitable. The sequence of change is outlined in Fig. 1. This broad strategy for tolerating low light is adopted to some extent in all species studied thus far; however, the threshold levels of light reduction (both duration and intensity of impact) leading to these responses, as well as the specific nature and mag- nitude of the adaptations are not known for many seagrass species including the tropical species of the Indo-West Pacific (Ralph et al., 2007; Walker et al., 1999; Waycott et al., 2005). There are ecological consequences of the plants adaptating to local conditions. For example, as productive carbon fixers, seagrass meadows support numerous foraging animals including the obli- gate seagrass feeders, dugongs; loss of productivity will reduce capacity to support these foragers. Seagrass productivity also drives nutrient cycling and nutrient regeneration: these place the 0025-326X/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2011.06.017 ⇑ Corresponding author. E-mail address: [email protected] (C.J. Collier). Marine Pollution Bulletin 65 (2012) 342–354 Contents lists available at ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul
Transcript of Responses of four Indo-West Pacific seagrass species to shading

Marine Pollution Bulletin 65 (2012) 342–354
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Responses of four Indo-West Pacific seagrass species to shading
Catherine J. Collier ⇑, Michelle Waycott, Ana Giraldo OspinaSchool of Marine and Tropical Biology, James Cook University, Townsville, Queensland 4811, Australia
a r t i c l e i n f o a b s t r a c t
Keywords:ShadingWater qualityGreat Barrier ReefMonitoringSubmerged aquatic vegetation (SAV)
0025-326X/$ - see front matter � 2011 Elsevier Ltd.doi:10.1016/j.marpolbul.2011.06.017
⇑ Corresponding author.E-mail address: [email protected] (C.J. C
Seagrasses of the Great Barrier Reef predominantly occur in coastal regions where terrestrial inputsmodify water quality and photosynthetic light is highly variable. Responses to shading were tested forCymodocea serrulata, Halodule uninervis, Thalassia hemprichii and Zostera muelleri. In aquaria, four lighttreatments – high (66% surface light), moderate (31%), low (14%) and very low light (1%) treatments –were applied for 102 d. Stress responses in the low and very low light treatments occurred in the follow-ing sequence: metabolic and physiological changes (reduced growth, increased pigment concentrationsand photosynthetic efficiency); shedding (leaf loss, shoot loss) and production of new, altered tissue(leaves with reduced length, width and thickness). Complete shoot loss was projected after 76 (Z. muel-leri) to 130 d (T. hemprichii). Responses were slower in the low than in the very low treatment, therefore,efforts to minimize water quality degradation will be rewarded with delayed impacts to seagrasses.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
The productivity, extent and ecological value of seagrass mead-ows have become threatened globally due to increasing anthropo-genic pressures, with reduced light penetration resulting from poorwater quality being amongst the most significant of impacts(Waycott et al., 2009). The extreme nature of the impact in marineand aquatic environments is unique, as the episodes of light reduc-tion can be very severe (down to 0% light) and very prolonged(many months, Campbell and McKenzie, 2004; Onuf, 1996; Preenet al., 1995). In addition, chronic declines in water quality haveled to broad-scale reductions in photosynthetic light available tobenthic primary producers around many coastlines (Fabricius,2011). The combination of these acute (localized) and chronic(broad-scale) reductions in light have led to world-wide loss ofseagrass meadows (Rasheed and Unsworth, 2011; Waycott et al.,2009). In the Great Barrier Reef, water quality in inshore areashas declined since European settlement, but the region of greatestimpact is south of Cairns and north of the Burdekin River, whichexperiences the greatest frequency of exposure to water qualitypollutants that reduce photosynthetic light to benthic primaryproducers (Devlin et al., 2012). In some circumstances, such asdredging, controls can be put in place to minimize the impactand manage the consequences for adjacent seagrass meadows(Erftemeijer and Robin Lewis Iii, 2006; Sofonia and Unsworth,2009). However, for most seagrass species, the thresholds of lightreduction are not known, hampering the ability of coastal resource
All rights reserved.
ollier).
managers to identify impacts and apply appropriate mitigationprocedures (Ralph et al., 2007).
Seagrasses have adapted somewhat to their highly variable lightenvironment and they respond in ways that provide tolerance toshort-term periods of light reduction. This is partially achievedby balancing carbon supply and respiratory requirements (Ralphet al., 2007). There are a number of possible changes that can helpto achieve this balance: both photosynthetic light capture and pho-tosynthetic efficiency can be increased (Abal et al., 1994; Durakoet al., 2003; Enríquez, 2005); carbohydrate reserves can be re-allo-cated from storage organs, such as the rhizome, to respiration andgrowth; and respiratory requirements can be reduced by slowinggrowth rates and reducing the amount of respiratory-demandingbiomass through morphological and meadow-scale changes(Collier et al., 2009; Fourqurean and Zieman, 1991). At sub-lethallight levels, these changes may be permanent enabling long-termsurvival, but at lethal light levels (below minimum light require-ments) die-off is inevitable. The sequence of change is outlinedin Fig. 1. This broad strategy for tolerating low light is adopted tosome extent in all species studied thus far; however, the thresholdlevels of light reduction (both duration and intensity of impact)leading to these responses, as well as the specific nature and mag-nitude of the adaptations are not known for many seagrass speciesincluding the tropical species of the Indo-West Pacific (Ralph et al.,2007; Walker et al., 1999; Waycott et al., 2005).
There are ecological consequences of the plants adaptating tolocal conditions. For example, as productive carbon fixers, seagrassmeadows support numerous foraging animals including the obli-gate seagrass feeders, dugongs; loss of productivity will reducecapacity to support these foragers. Seagrass productivity alsodrives nutrient cycling and nutrient regeneration: these place the

Increasing stress e.g. light reduction
Healthy population
Primary sub-lethalstress response
State change /population decline
Population extinction
Reducing growth rates
Changing tissue nutrients
Adjusting photosystems
Decreasing density of reproductive structures
Changing plant morphology
Changing species composition
Reducing abundance and meadow area
Seagrass population status (declining)
Pla
nt
resp
on
se
Ecophysiologicalresponse
Morphologicalresponse
Meadowresponse
Fig. 1. Seagrass light response model outlining the sequence of changes that occur in response to light reduction. This paper has sought to place timelines on the physiologicaland morphological changes. (Adapted from Waycott et al. (2005) and Ralph et al. (2007).)
C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354 343
economical value of seagrasses amongst the highest of the world’secosystems (Costanza et al., 1997; Romero et al., 2006) and reduc-tion in their capacity to capture inorganic nutrients will influencethe nutrient balance of other nearby ecosystems such as coral reefs(cf. Fabricius, 2011). Furthermore, meadow density and morphol-ogy are critical in the role of seagrass as a habitat and in preda-tor–prey relations (Heck and Orth, 2006; Hyndes et al., 2003).Due to the dependency of these ecological processes on meadowproductivity and structure, light reduction can have significantecological consequences (Hyndes et al., 2003; Unsworth andCullen, 2010). However, many of the ecological roles of seagrassesare species-specific, with some being more important as a directfood source and others in their role as habitat (Heck et al., 2008).Therefore, the ecological impacts of light reduction will dependon the seagrass species, or the mix of species that are impacted.
Species-specific thresholds depend on species differences inmorphological plasticity, storage products, life-form, and growthrates (Czerny and Dunton, 1995; Grice et al., 1996; Olesen et al.,2002; Ralph et al., 2007; Walker et al., 1999). For example, mor-phologically large and slow-growing species, such as Posidoniaspp. tolerate prolonged periods of low light by implementing thebroad strategy outlined above, but if a severe impact results, theyare slow to recover. In contrast, small, rapidly growing species,such as Halophila spp., cannot tolerate extended periods of lowlight, largely because of limited storage reserves, but they can re-cover from such impacts quickly if conditions improve (Longstaffet al., 1999; Waycott et al., 2005).
The Great Barrier Reef (GBR) supports some 15 species of sea-grass most of which are common throughout the Indo-West Pacific(Waycott et al., 2004). Light availability is the dominant factordriving seagrass distribution and composition in the GBR withlight-related stress resulting from run-off, wind-driven resuspen-
sion and coastal modification including dredging (Carrutherset al., 2002; Waycott et al., 2005). Cymodocea serrulata is commonin subtidal environments, particularly on muddy reef tops. Halod-ule uninervis (which also includes H. pinifolia until species demar-cations are clarified is common throughout northern Australiaand is the dominant species in many of the shallow subtidal andintertidal coastal regions (Waycott et al., 2004). It is one of the pre-ferred species for dugong and can rapidly re-colonize after distur-bances (Aragones et al., 2006). Thalassia hemprichii is morecommon in reef environments where influences from freshwaterand turbidity are less common. It has high rates of leaf production,but thick rhizomes that are slow to extend laterally compared tomost other tropical species (Marbà and Duarte, 1998). Zosteramuelleri is common in tropical and temperate eastern Australiaand New Zealand and is dominant in the river estuaries and pro-tected bays of coastal northern Australia where it experiencespulsed run-off during summer rainfall and episodes of low lightare common (Carruthers et al., 2002; McKenzie, 1994).
Here we examine shade responses in these four tropical seag-rasses, which are foundation species in the GBR, with a particularemphasis on timelines of response. The aim of this experimentwas to describe the time taken for light reduction to impact ecolog-ically important traits of seagrass meadows. These timelines canguide environmental managers in their quest to establish environ-mental management guidelines and to minimize impacts to coastalenvironments from light reduction. An aquarium-based lightreduction experiment was carried out at four light intensities:two below and two above their theoretical minimum light require-ments (e.g. Longstaff and Dennison, 1999). We hypothesized thatthe species with larger rhizomes, and larger storage capacity(C. serrulata and T. hemprichii), would be more tolerant to the lightreduction, despite their predominance in environments that are

344 C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354
less exposed to severe low light events. Impacts of the low lighttreatments would therefore be delayed in these species relativeto the other species.
2. Materials and methods
2.1. Experimental setup
The seagrasses C. serrulata, H. uninervis, T. hemprichii and Z. muel-leri were collected from Magnetic Island, off the coast of Townsvillein the northern Great Barrier Reef, Queensland, Australia 2–4weeks prior to the beginning of the experiments: T. hemprichiiwas collected 2 weeks prior to the experiment, and all other spe-cies were collected 4 weeks prior. All species were collected byinserting a 17 cm i.d. stainless steel corer into the sediment to adepth below the root zone: typically this was 10–20 cm. The ex-tracted core was placed into a 17 cm plastic plant pot lined witha plastic bag. The plastic bag was pulled up over the seagrass andtied at the top to hold in moisture while in transit. The wholepot was placed in large plastic containers with a small amount ofwater in the bottom, and a lid to retain moisture for transport toaquaria within 6 h.
The plants were kept in large 1000 L outdoor plastic aquariawith constant water flow and aerators at 66% surface irradiance(SI). The experiment was initiated on the 12th March 2009. Therewere four light treatments applied by placing shade screens overthe seagrass: the screens were supported by the rim of the tankand hung down into the water to block sideward penetration oflight. There were five replicate tanks, each tank containing one ofeach treatment (n = 5). Water was filtered through a 10 lm filterto remove invertebrate grazers such as amphipods then retainedin a closed circulation system. Coris wrasse were added to eachtank to feed on any remaining small crustaceans, such as thoseintroduced with the seagrass.
2.2. Environmental characteristics
Ambient light was measured using 2p light loggers (Submersi-ble Odyssey Photosynthetic Irradiance Recording System, DataflowSystems Pty. Ltd., New Zealand), which were deployed on siteabove the aquaria. On average, the ambient surface light was31 mol m�2 d�1. The light reducing properties of the shade screenswas verified using a 2p quantum sensor (LiCorTM Li-192SA, Ne-braska, USA) held at canopy height. Daily light reaching the sea-grass was calculated from the ambient light and the lightreducing properties of the shade screens.
The light treatments were 66%, 31%, 14% and 1% of surface light(SI), equating to an average of 20.3, 9.5, 4.4 and 0.2 mol m�2 d�1 atcanopy height, however, light intensity declined throughout thestudy period as it was initiated at the beginning of autumn. Thetwo highest light treatments are at or above the minimum lightrequirements for long-term survival: H. uninervis has a minimumlight requirement (MLR) of more than 14% SI (Longstaff and Denn-ison, 1999); Z. muelleri has an MLR of 16–36% SI (Longstaff, 2002);and MLR has not been quantified for C. serrulata or T. hemprichii butit is probably greater than 20% based on their occurrence in shal-low and/or high light environments. The two lower treatmentsare below MLR and seagrass meadows can be exposed to such con-ditions during flood plume events.
2.3. Leaf dimensions and demography
Leaf morphological dimensions were measured before the lighttreatments were applied (day 0) and 25, 46 and 74 d after thetreatments were applied. On five shoots in each replicate pot the
following were measured on the youngest mature leaf: leaf length(from base of sheath to leaf tip), leaf width at centre of leaf (usingMitutoyo digital calipers) and leaf thickness at centre of leaf (usingMitutoyo digital calipers). The number of shoots per pot and thenumber of leaves per shoot were measured before the experimentstarted (day 0) and after 14, 20, 25, 46, 61, 74 and 102 (shoot den-sity only at 102 d). At the end of the experiment (102 d) the sea-grass was collected, rinsed and stored at �18 �C prior to furtheranalysis. The samples were later sorted into above-ground biomass(leaves and live sheath) and below-ground biomass (roots and rhi-zomes), with the dead sheath being excluded from biomass mea-surements. Between the last morphological measures (day 74)and the termination of the experiment (day 102), an outbreak ofsmall crustaceans, primarily amphipods, had occurred in one ofthe replicate tanks. They had partially eaten the leaves of most spe-cies and therefore, this replicate was excluded from biomass mea-sures (i.e. n = 4 for biomass).
Leaf area index (LAI, m2 m�2) was calculated as:
LAI ¼ LL� LW � Lves1;000;000
� �� Shoots ð1Þ
where LL is leaf length (mm), LW is leaf width (mm), Lves is leavesper shoot (total number of leaves less 0.5 to account for a singlejuvenile growing leaf on each shoot) and Shoots is shoot density(shoots, m�2).
2.4. Leaf growth
Leaf growth was measured according to the method of Shortand Duarte (2001). Ten shoots from each pot were marked witha needle for growth. After 6–11 d of growth, the length of growth(mm) was measured on the shoots without removing them fromthe pots on day 7, 21 and 54. Leaf production time, leaf turnovertime and relative leaf growth rate were calculated from leaf growthrates and leaf dimension data.
Areal leaf growth rate (ArGrLf, cm m�2 d�1) was calculated as:
ArGrLf ¼GrLf � Shoots
100ð2Þ
where GrLf is growth rate of the leaves (mm d�1).Leaf production time (ProdLf, d) was calculated as
ProdLf ¼LL
GrLfð3Þ
Leaf turnover time (TLf, d) as:
TLf ¼ ProdLf � Lves ð4Þ
and Relative leaf growth rate (RelGrLf, mm mm�1 d�1) as:
RelGrLf ¼GrLf
LL� Lvesð5Þ
2.5. Chlorophyll
A single leaf was collected from each replicate pot 0, 8 and 22 dafter the initiation of shading for analysis of chlorophyll concentra-tion. Collection was not continued as the shade treatments hadstarted to affect the plants and leaves were becoming increasinglyscarce with further collection of leaves likely to impact the plants.The leaves were placed in an ice slurry prior to storage at �18 �Cand analysis. For determination of chlorophyll concentration, a10 or 20 mm (depending on the species) section of leaf was cutfrom the middle of a fully mature leaf and the width of the leaf seg-ment was measured using InTech digital callipers. The leaves wereblotted dry then ground in a chilled mortar. Chlorophyll was thenextracted from the sample in 4 ml cold (4 �C) 90% acetone. The

C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354 345
sample was gently shaken, then left to extract for 12 h at 4 �C. Theextract was measured for chlorophyll concentration according toGranger and Izumi (2002).
2.6. Carbohydrates
The below-ground biomass, collected after 102 d, of the highlight treatments were analyzed for total soluble sugars and totalnon-structural carbohydrates (TNSC). Only the high light treat-ments were analyzed as we sought to establish a comparisonamongst species of the CHO storage capacity of the below-groundmaterial, furthermore the low and very low light treatments hadinsufficient biomass for such analysis. Soluble sugars were ex-tracted in 80% ethanol at 80 �C for 3 min (repeated 3 times andcentrifuged at 2500 rpm for 5 min between each extraction). Thesupernatant was retained for soluble CHO determination. TNSCremaining in the pellet was then solubilized by mixing in deion-ized water and heating at 95 �C for 1 h. The TNSC was then digestedwith amylase enzyme and incubated at 55 �C for 2 h. The samplewas then centrifuged and the supernatant filtered. The sampleswere then analyzed colorimetrically using a ferricyanide reagent.The values for soluble and TNSC components were summed for to-tal carbohydrates. This was then expressed in terms of the above-ground biomass supported by the carbohydrates by multiplyingthe carbohydrate concentration in the rhizome by the below-ground biomass and comparing weight of carbohydrates againstweight of leaf material.
2.7. Statistical analysis
Time series data was analyzed with two-way repeated measuresANOVA with species and treatment as fixed effects (between-subjecteffects) over time (within-subjects effects). Data were checked forhomogeneity of variance using Levene’s test and if they failed, datawere square root or log transformed. If the transformation was notsuccessful at improving the variance in the data, the ANOVA was per-formed but with significant p-values set to 0.01 to minimize the risk ofa Type 1 error (Underwood, 1997). Transformations and p-values areshown for all data. The variance–covariance matrices were testedusing Mauchly’s test of sphericity, and if the assumption was notmet, the Greenhouse–Geisser epsilon adjustment was applied to thedegrees of freedom (non-adjusted degrees of freedom are presentedin the tables). Treatment and species effects identified using RM ANO-VA were interpreted using Tukey’s post hoc analysis. Where no speciesinteraction was identified in the RM ANOVA, all species were pooledfor further post hoc interrogation and where a species interactionwas identified, the species were further analyzed separately.
Leaf production and turnover time, relative leaf growth rate,biomass and carbohydrates were analyzed after 54 d (growth)and 102 d (biomass and carbohydrates) using a two-way ANOVAwith both Species and Treatment as fixed effects. The data weretested and adjusted as per above. All statistical analyses wereundertaken in SPSS PASWStatistics 18.0.
3. Results
3.1. Leaf dimensions
Leaf length was affected by Treatment, but it depended on bothSpecies and Time (Time � Species � Treatment, p < 0.001, Table 1),so Treatment and Time effects were further analyzed separatelyfor each species. For C. serrulata, leaves were slightly, but signifi-cantly longer in the high light treatment than the low and verylow on day 0. Following this, it was 74 d before leaves in the verylow light treatment were significantly reduced in length compared
to the high light treatment (reduced to just 52%, Fig. 2). Leaves ofH. uninervis, responded more quickly, taking 46 d in the very lowand 74 d in the low light treatment to be significantly different fromthe high light treatment (after 74 d they were 30% and 42% of thelength in the high light treatment, respectively). T. hemprichii leaveswere also significantly reduced after 46 d and they remained shorterafter 74 d (leaves were 67% of the length of the high light treatmentafter 74 d). Z. muelleri leaves responded quickly in the very low lighttreatment with leaves shorter than the moderate and low light treat-ments after 25 d while it took 74 d for leaves in the low light treat-ment to be significantly reduced in length (after 74 d leaves in thelow and very low were 60% and 23% the length of the high light treat-ment). Leaf dimensions were not different in the moderate lighttreatment compared to the high light treatment for all morphologi-cal parameters and all species.
Leaf width and leaf thickness were the slowest variables tochange following the light treatment with a treatment effect occur-ring that depended on time (Time � Treatment, p < 0.001, Table 1).For all species, leaf width was significantly narrower than all othertreatments in the very low light treatment and narrower in the lowcompared to the moderate and high than the high light treatmentafter 74 d (Fig. 2). Leaf thickness was reduced on day 74 whenleaves were significantly thinner in the very low and low comparedto the moderate and high light treatments for all species (Fig. 2).The magnitude of difference between treatments was the greatestin Z. muelleri, where thickness of the leaves in the very low werejust 54% of the high light treatment followed by H. uninervis(66%), T. hemprichii (70%) and C. serrulata (81%) after 74 d.
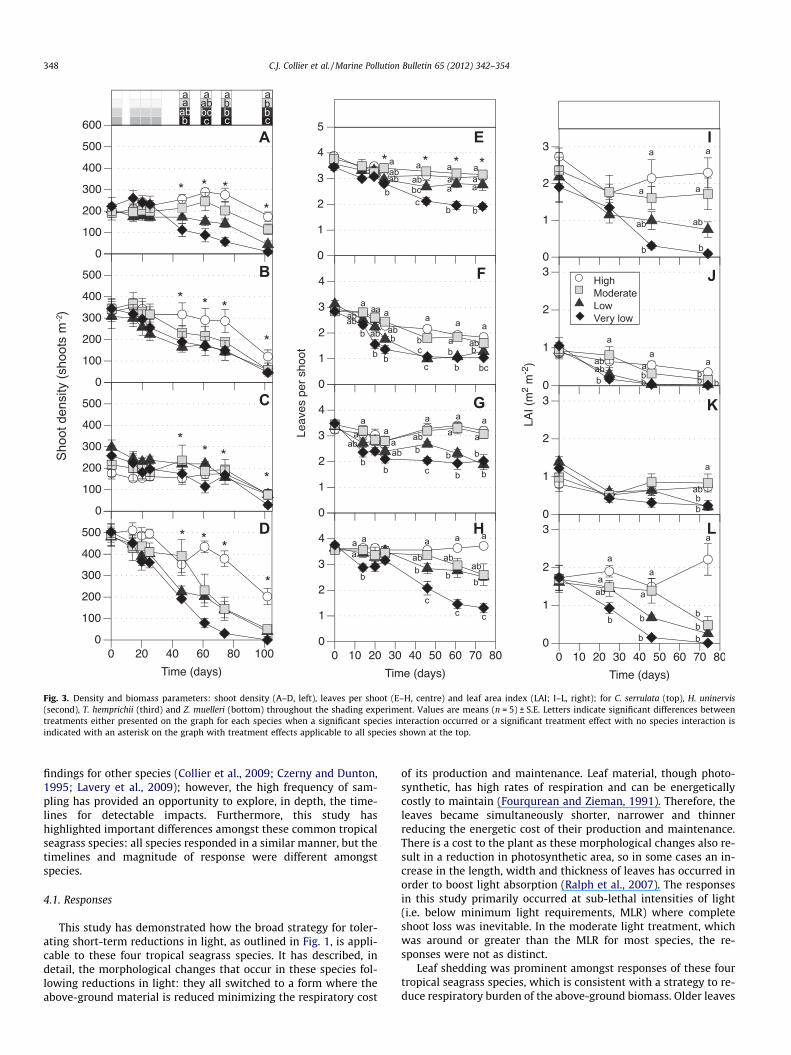
3.2. Density
There was an effect of the light Treatment on shoot density inall species but it was dependant on Time (Time � Treatment,p < 0.001, Table 1). Shoot density was significantly reduced in thevery low light treatment after 46 d, in the low light treatment after61 d, and reduced in the moderate light treatment after 74 d(Fig. 3). The magnitude of the effect of the light treatment on shootdensity was highly variable between species with Z. muelleri beingthe most profoundly impacted: shoot density in the very low treat-ment compared to the high light treatment on day 102 was 0% (Z.muelleri), 6% (C. serrulata), 35% (T. hemprichii), 38% (H. uninervis).There was also a general reduction in shoot density in all treat-ments throughout the course of the shading experiment, includingthe high light treatment, which may have been due to reductionsin ambient air and water temperature toward the later part ofthe experiment which was completed at the beginning of winter.Taking a conservative estimate (i.e. assuming linear loss) of thetime taken for complete loss of shoots in the low and very low lighttreatments, all are predicted to have been lost within 130 d ofshading (i.e. after 28 more days).
The number of leaves on each shoot responded quickly andstrongly to the light treatment, with the treatment effect depen-dant on Time (Time � Treatment, p < 0.001, Table 1). The numberof leaves per shoot was reduced in the very low compared to allother treatments after 14 d and was reduced in both the low andvery low compared to the moderate and high after 46 d (Fig. 3).After 61 d, the very low was reduced compared to all other treat-ments, but because of ongoing declines in the moderate treatment,the low was then reduced compared to the high light treatmentonly. By day 74, only the very low was significantly reduced com-pared to the high light treatment. The magnitude of change wasgreatest for Z. muelleri as there were an average of just 1.3 leavesin the very low compared to 3.7 leaves in the high light treatmentwhile for other species there were 1.9 compared to 3.1 (C. serrula-ta), 1 compared to a 1.8 (H. uninervis) and 2 compared to 3.2(T. hemprichii).

Table 1Results of a two-way repeated measures ANOVA testing for the within-subjects effects of time, species and treatment on the leaf dimension parameters leaf length, width andthickness, and on the density parameters shoot density, leaves per shoot and leaf area index (LAI). Also shown are any transformations applied to the data for statistical analysisand the p-value that was considered significant (adjusted if variances were not homogenous).
Leaf length (mm) Leaf width (mm) Leaf thickness (mm) Shoot density (shoots, m�2) Leaves per shoot LAI (m2 m�2)
MS p MS p MS p MS p MS p MS p
Within-subjects effectsTime 4967.6 <0.001 0.603 <0.001 0.005 <0.001 741138.6 <0.001 11.313 <0.001 0.167 <0.001Time � Species 1088.0 <0.001 0.048 <0.001 0.001 <0.001 83216.4 <0.001 1.246 <0.001 0.006 <0.001Time � Treatment 3013.1 <0.001 0.168 <0.001 0.002 <0.001 39,013 <0.001 1.054 <0.001 0.019 <0.001Time � Species � Treatment 550.2 <0.001 0.013 n.s. 0.0003 n.s. 5696.1 n.s. 0.190 <0.05 0.004 <0.001
Between-subjects effectsSpecies 30072.3 <0.001 17.210 <0.001 0.039 <0.001 700214.3 <0.001 38.337 <0.001 0.220 <0.001Treatment 12151.8 <0.001 0.405 <0.001 0.005 <0.001 175224.1 <0.01 14.079 <0.001 0.087 <0.001Species � Treatment 1003.6 n.s. 0.021 n.s. 0.001 n.s. 58803.8 n.s. 0.250 n.s. 0.016 <0.01
Transformation None Natural log None SqRt None Natural logSignificance level p < 0.01 p < 0.01 p < 0.01 p < 0.01 p < 0.01 p < 0.01
346 C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354
The leaf area index (LAI) was affected by the light treatments,but it was dependant on both Species and Time (Time � Spe-cies � Treatment, p < 0.001, Table 1), so post hoc analysis of treat-ment effects were performed for each species separately. ForC. serrulata, LAI was significantly reduced in the very low lighttreatment after 46 d and in the low and very low after 74 d(Fig. 3). For H. uninervis, LAI was significantly reduced in the verylow compared to the moderate after just 25 d, and reduced in boththe low and very low after 46 d. For T. hemprichii, there was a sig-nificant treatment effect only after 74 d while for Z. muelleri, LAIwas reduced after just 25 d. After 74 d, LAI was reduced the mostin Z. muelleri (0.2% compared to the high light treatment) followedclosely by H. uninervis (3.2%) and C. serrulata (4%) but was less im-pacted in T. hemprichii (37.2%).
Above-ground and below-ground biomasses were measuredafter 102 d. Both were affected by the light treatments, but notequally, resulting in a switch in the ratio of below-ground:above-ground biomass (Fig. 4). For the below-ground biomass, there wasa significant effect of the shade treatment for all species (Treatment,p < 0.001, MS = 111.462, F = 7.591): biomass was highest in the highlight treatment and lowest in the very low light treatment and onlythese two treatments were significantly different from each other(p < 0.001). The magnitude of difference was very different betweenspecies: the reduction was largest in Z. muelleri (just 7% of biomassremained in the very low light compared to the high light treatment)and C. serrulata (11%) followed by H. uninervis (47%) and T. hemprichii(68%). There was also a significant difference between species(p < 0.001, MS = 319.884, F = 21.785): T. hemprichii had a signifi-cantly larger below-ground biomass than all other species followedby C. serrulata, which was also significantly larger than H. uninervisand Z. muelleri. For the above-ground biomass, there was also a sig-nificant effect of the light treatment, but it was dependant on species(Species � Treatment, p < 0.05, MS = 5.741, F = 2.726): for C. serrula-ta and T. hemprichii, biomass was significantly reduced in the verylow compared to the high and moderate treatments, for H. uninervis,biomass was smaller in the very low compared to the high, while forZ. muelleri, biomass was largest in the high light, followed by themoderate and low, and then the very low treatment, where above-ground biomass was reduced to zero after 102 d. The above-groundbiomass generally reduced more in the low and very low light treat-ments, than the below-ground biomass, leading to an increase in theratio below-ground:above-ground biomass. This was least pro-nounced in C. serrulata and Z. muelleri where the ratio changed from4.4 to 11.9 and from 0.9 to 2.7, respectively. The change in ratio wasthe most pronounced in T. hemprichii where the ratio from 6.5 to 47.5due to large reductions in above-ground biomass with little changein the below-ground biomass. Complete loss of seagrass waspredicted in the very low light treatment after 76 d for Z. muelleri
(no shoots were recovered after 102 d when the biomass was ex-tracted), 100 d for C. serrulata, 119 d for H. uninervis and 133 d forT. hemprichii. These timelines extended to 126 and 128 d for Z. muel-leri and H. uninervis in the low light treatment.
3.3. Growth
Leaf growth rates declined in response to the light treatments,but the nature of the response was dependant on time (Time �Treatment, p < 0.001, Table 2): within the first 7 d, leaf extensionwas slower in the very low light treatment compared to all othersand this trend continued up to 21 d (Fig. 5). After 54 d, leaf extensionwas slower in the low and very low light treatments compared tothe moderate and high light treatments. Shading had the most pro-nounced effect on Z. muelleri (where leaf growth in the very lowtreatment was just 9% of that in the high light treatment after 54d) followed by H. uninervis (18%), T. hemprichii (33%) and C. serrulata(49%). Areal growth rate was affected by the light treatments but theeffect was dependant on both the Time and Species (Time � Spe-cies � Treatment, p < 0.01, Table 2). Areal growth responded thefastest in Z. muelleri — taking just 7 d — and the slowest in T. hempri-chii — taking 54 d (Fig. 5). After 54 d, areal leaf extension rate in thevery low treatment was less than 5% of the extension rate in the highlight treatment for all species, with the most pronounced effectagain on Z. muelleri (0.3%).
Leaf production time was affected by treatment, but not for allspecies (Species � Treatment, p < 0.01, MS = 0.240, F = 35.859):there was no effect of treatment for C. serrulata, and the largesttreatment effects were for T. hemprichii and Z. muelleri with ratesin the very low light treatment being 2.5 and 4.4 times slowercompared to the high light treatments, respectively (Table 3). Incontrast, there was no significant treatment effect on the rate ofleaf turnover (Species � Treatment, p > 0.05, MS = 273.525,F = 2.026, Treatment, p > 0.05, MS = 324.104, F = 2.400), thoughthere was a difference between species with a quicker turnovertime in H. uninervis compared to Z. muelleri (Table 3). There wasalso no significant effect of Treatment or Species on relative growthrate (RGR, Treatment � Species, p > 0.05, MS = 0.000, F = 0.562,Treatment, p > 0.05, MS = 0.000, F = 1.512, Species, p > 0.05,MS = 0.000, F = 1.367) with rates ranging from 0.19 to 0.43 for allspecies and treatments (Table 3).
3.4. Carbohydrates
Total carbohydrate concentration in the rhizome was signifi-cantly different amongst species (p < 0.001, MS = 2.062, F = 24.724,SqRt transformation): it was highest in H. uninervis followed by C.serrulata and lowest in both T. hemprichii and Z. muelleri (Table 4).

*
*
*
*
*
*
*
*
Leaf
wid
th (m
m)
Leaf
thic
knes
s (m
m)
aa
a
a
a a
a
aa
a
a
a
aa
a
abab
ab ab
abab
b
b
b
b
c
b
b
b
b
aa
bb
b
ab
aabc
aabb
HighModerateLowVery low
A
B
C
D
E
F
G
H
I
J
K
L
Fig. 2. Leaf dimensions: leaf length (left), leaf width (centre) and leaf thickness (right) for C. serrulata (top), H. uninervis (second), T. hemprichii (third) and Z. muelleri (bottom)throughout the shading experiment. Values are means (n = 5) ± S.E. Asterisks indicate significant treatment effect. Letters indicate significant differences between treatmentseither presented on the graph for each species when a significant species interaction occurred or a significant treatment effect with no species interaction is indicated with anasterisk on the graph with treatment effects applicable to all species shown at the top.
C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354 347
When expressed as weight of leaf material supported by thesereserves (total carbohydrate per leaf biomass), Z. muelleri had signif-icantly lower carbohydrates to support its leaf material (p < 0.001,MS = 0.029, F = 11.327, SqRt transformation): it was an order ofmagnitude lower than all other species (Table 4).
3.5. Chlorophyll
There was a difference amongst species in the concentration ofchlorophyll on a leaf area basis (p < 0.001, Table 2); C. serrulata andH. uninervis had a lower concentration than T. hemprichii and Z.muelleri, but they were not different from each other (Fig. 6). Therewas also a treatment effect that was dependant on time(p < 0.001): on day 0 there were no differences amongst treat-ments; on day 8 chlorophyll concentration was increased in thelow light treatment compared to the high and very low light treat-ment and was increased in the moderate compared to the high
light treatment only; on day 22 chlorophyll concentration was in-creased in the moderate compared to the high and increased in thelow light treatment compared to all other treatments. There wasalso a significant effect of treatment on the chlorophyll a/b ratiothat was dependant on time (Time � Treatment, p < 0.001, Table2): on day 0 there were no differences amongst treatments; onday 8 chlorophyll a/b was reduced in the low and very low lighttreatment compared to the high light treatment, and; and on day22 there were no significant differences amongst treatments(Fig. 6). There was also a significant difference amongst species(Species, p < 0.001), with only C. serrulata and H. uninervis havinga similar a/b ratio.
4. Discussion
Light reduction affected the four Great Barrier Reef tropicalseagrass species in a manner that was consistent with previous
*
* * **
* *
*
** *
*
* * *
*
* * * *
Shoo
t den
sity
(sho
ots
m-2)
aa
abb
aabbcc
abbc
abbc
a a aa a
a a
aa a
aaab
ab ab
b
b
b b
bc
bc
abab
a a a a a aa
aa
a
aa
a
aa
a
a
a
b
b b
bb
b b bb
b
b b
b bbb
b
bb
b
b
b
ababab ab
ab
abab
ab
ab
c
c
c
bc
aaa aaa
ababab
bbb
b
ccc
aaa
a a
aa
a a
b
bb
b
b b
b
abab
ab
A
B
C
D
E
F
G
H
I
J
K
L
HighModerateLowVery low
Fig. 3. Density and biomass parameters: shoot density (A–D, left), leaves per shoot (E–H, centre) and leaf area index (LAI; I–L, right); for C. serrulata (top), H. uninervis(second), T. hemprichii (third) and Z. muelleri (bottom) throughout the shading experiment. Values are means (n = 5) ± S.E. Letters indicate significant differences betweentreatments either presented on the graph for each species when a significant species interaction occurred or a significant treatment effect with no species interaction isindicated with an asterisk on the graph with treatment effects applicable to all species shown at the top.
348 C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354
findings for other species (Collier et al., 2009; Czerny and Dunton,1995; Lavery et al., 2009); however, the high frequency of sam-pling has provided an opportunity to explore, in depth, the time-lines for detectable impacts. Furthermore, this study hashighlighted important differences amongst these common tropicalseagrass species: all species responded in a similar manner, but thetimelines and magnitude of response were different amongstspecies.
4.1. Responses
This study has demonstrated how the broad strategy for toler-ating short-term reductions in light, as outlined in Fig. 1, is appli-cable to these four tropical seagrass species. It has described, indetail, the morphological changes that occur in these species fol-lowing reductions in light: they all switched to a form where theabove-ground material is reduced minimizing the respiratory cost
of its production and maintenance. Leaf material, though photo-synthetic, has high rates of respiration and can be energeticallycostly to maintain (Fourqurean and Zieman, 1991). Therefore, theleaves became simultaneously shorter, narrower and thinnerreducing the energetic cost of their production and maintenance.There is a cost to the plant as these morphological changes also re-sult in a reduction in photosynthetic area, so in some cases an in-crease in the length, width and thickness of leaves has occurred inorder to boost light absorption (Ralph et al., 2007). The responsesin this study primarily occurred at sub-lethal intensities of light(i.e. below minimum light requirements, MLR) where completeshoot loss was inevitable. In the moderate light treatment, whichwas around or greater than the MLR for most species, the re-sponses were not as distinct.
Leaf shedding was prominent amongst responses of these fourtropical seagrass species, which is consistent with a strategy to re-duce respiratory burden of the above-ground biomass. Older leaves

4.4 4.5 5.8 11.9 5.6 12.1 11.6 22.6 6.5 7.0 17.1 47.5 0.9 3.7 2.7
C. serrulata H. uninervis T. hemprichii Z. muelleri
Abov
e-gr
ound
bio
mas
s (g
m-2)
Belo
w-g
roun
d bi
omas
s (g
m-2)
–
a
a
a
aab ab
a
a
a
a
aab
a
ab
ab
ab
ab
ab
b
b
b
b
bbb c
b b
b
b
b
b
10
5
00
5
10
15
20
25
30
35
40
High
Moderate
Low
Very low
Fig. 4. Above-ground biomass (top) and below-ground biomass (bottom) after 102 d shading in C. serrulata, H. uninervis, T. hemprichii and Z. muelleri. Values are means(n = 4) ± S.E. The ratio of below-ground:above-ground biomass is shown along the bottom of the figure.
Table 2Results of a two-way repeated measures ANOVA testing for the within-subjects effects of time, species and treatment on leaf extension and areal leaf extension. Also shown areany transformations applied to the data for statistical analysis and the p-value that was considered significant (adjusted if variances were not homogenous).
Leaf extension (mm d�1) Areal leaf extension (cm m�2 d�1) Chlorophyll conc. (lg cm�2) Chlorophyll a/b
MS p MS p MS p MS p
Within-subjects effectsTime 84.630 <0.001 35.243 <0.001 850.894 <0.001 1.514 <0.001Time � Species 11.161 <0.001 0.632 <0.001 22.542 n.s. 0.049 n.s.Time � Treatment 11.276 <0.001 1.733 <0.001 95.780 <0.001 0.093 <0.01Time � Species � Treatment 2.570 n.s. 0.181 <0.01 16.700 n.s. 0.016 n.s.
Between-subjects effectsSpecies 90.214 <0.001 10.395 <0.001 288.477 <0.001 2.892 <0.001Treatment 159.278 <0.001 6.763 <0.001 194.798 <0.001 0.373 <0.001Species � Treatment 3.095 n.s. 0.478 <0.05 5.679 n.s. 0.040 n.s.
Transformation None Natural log None NoneSignificance level p < 0.01 p < 0.05 p < 0.01 p < 0.01
C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354 349
have reduced photosynthetic rates (partially due to higher loads ofepiphytes) and high respiration rates (Bulthuis and Woelkerling,1983; Libes and Boudouresque, 1987), so by shedding these olderleaves only the most photosynthetically active material is retained.Because leaves are shed more quickly, all species maintained theirhigh rates of leaf turnover, despite slowed leaf extension rates. Thiscan also assist in the light response as epiphyte accumulation is re-duced with faster rates of leaf turnover (Peterson et al., 2007).Whole shoots were also shed, reducing overall shoot density whichis indicative of a meadow-scale response to the low light condi-tions (Collier et al., 2009). These combined findings indicate a‘gearing down’ strategy (qv Grubb, 1998) in which changes to re-duce respiration rates enable the seagrasses to tolerate light depri-vation. The morphological and meadow-scale changes further
benefit light penetration through the canopy to saturate the leafsurface with light (Collier et al., 2009; Enríquez et al., 2002).
There were also physiological changes, including pigment con-tent, which improved light capture and its conversion to chemicalenergy. Chlorophyll concentration was increased, which can in-crease the efficiency of light capture under low light conditions(Ralph et al., 2007), though, due to the constraints of packing extrachlorophyll into a single layer of epidermal cells, this increase inchlorophyll does not correspond linearly to increased light capture(Enríquez, 2005). However, reductions in the a/b ratio in all shadetreatments (significant on day 8 only) compared to the high lighttreatment, can also improve the capture of light at longer wave-lengths, which are proportionally more dominant in low light con-ditions. Interestingly, the concentration of chlorophyll was not

* * *
**
*
* * *
***
A
B
C
D
E
F
G
H
a a aaa
a aabbb b
a
a
a
a
a
a
aa
aa
a
aa
ab
ab
ab
abab
ab
ab
abb
b
b
b
b
b bbc
bb
Area
l gro
wth
rate
(cm
m-2 d
-1)
HighModerateLowVery low
Fig. 5. Leaf extension parameters: leaf extension (A–D, left) and areal leaf extension (E–H, right) for C. serrulata (top), H. uninervis (second), T. hemprichii (third) and Z. muelleri(bottom) throughout the shading experiment. Values are means (n = 5) ± S.E. Asterisks indicate significant treatment effect. Letters indicate significant differences betweentreatments either presented on the graph for each species when a significant species interaction occurred or at the top of the graph when results apply to all species.
350 C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354
increased in the very low light treatment compared to the highlight treatment, a trend previously observed for Z. muelleri (Abalet al., 1994); perhaps under these extreme low light conditions,it is not energetically worthwhile to boost chlorophyll concentra-tion. Given this light intensity-dependant response, caution wouldhave to be implemented if interpreting changes in the environ-ment based on chlorophyll concentration. Other pigments can re-spond to changing light intensity, including the photoprotectivepigments, the xanthophylls; however, their potential applicationas a indicator of reductions in light remains uncertain and as yetfully untested (Collier et al., 2008; Ralph et al., 2002).
4.2. Timelines
The high sampling frequency of this study has provided theopportunity to describe the timelines of response to shading forthese four seagrass species. The responses to reduced light oc-
curred in the following sequence: metabolic changes, sheddingand production of new morphologically altered tissue, with thetiming dependant on the intensity of the treatment. This broad se-quence concurs with findings of other light deprivation experi-ments on seagrasses (Collier et al., 2009; Lavery et al., 2009;Longstaff and Dennison, 1999; Waycott et al., 2005).
Metabolic changes can occur within seconds or minutes follow-ing changes in the environment. For example, photosynthetic effi-ciency changes throughout the day follow diurnal patterns in solarirradiance (Beer et al., 2006; Belshe et al., 2007). In this experi-ment, changes in the rate of leaf extension also occurred withinthe first 7 d. In few cases, growth does not immediately reduce fol-lowing light reduction probably because growth is supported bycarbohydrate reserves (Collier et al., 2009); however, these smalltropical species have fast leaf growth and turnover times and com-parably small storage capacities and growth is sensitive to changesin light. Leaves were shed soon afterwards, with significant loss of

Table 3Leaf production time, leaf turnover time and relative growth rate of C. serrulata, H.uninervis, T. hemprichii and Z. muelleri after 54 d in high, moderate, low and very lowlight treatments. n = 5 with standard error shown in parentheses.
Treatment C. serrulata H. uninervis T. hemprichii Z. muelleri
Leaf production time (d)High 13.9 (0.9) 19.3 (1.1)a 12.2 (0.7)a 11.7 (0.8)a
Moderate 13.6 (0.8) 25.8 (3.7)a 12.3 (1.4)a 16.3 (2.5)ab
Low 21.9 (5.4) 44.6 (1.2)b 26.1 (4.6)b 23.9 (2.3)b
Very low 22.4 (3.5) 33.5 (5.7)a,b 30.0 (5.9)b 51.7 (5.6)c
Leaf turnover time (d)High 32.2 (3.8) 28.9 (1.7) 32.5 (0.9) 38.5 (3.5)Moderate 37.1 (2.4) 22.3 (5.5) 33.6 (3.9) 38.3 (2.6)Low 46.2 (5.9) 38.7 (0.6) 51.6 (5.8) 48.7 (4.9)Very low 29.9 (7.2) 16.7 (2.9) 39.9 (7.6) 48.7 (12.1)
Leaf relative growth rate (mm cm�1 d�1)High 0.030 (0.004) 0.032 (0.002) 0.028 (0.001) 0.024 (0.002)Moderate 0.025 (0.002) 0.026 (0.004) 0.029 (0.003) 0.024 (0.001)Low 0.021 (0.003) 0.033 (0.008) 0.019 (0.002) 0.019 (0.002)Very low 0.042 (0.015) 0.036 (0.017) 0.026 (0.005) 0.023 (0.005)
Note: Identical letters in the same row are not significantly different (p > 0.05).
C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354 351
leaves per shoot after just 14 d in three out of the four species.Shoot density losses followed some time later after 46 d and com-plete loss was predicted within 76–133 d. The timeline for shootloss concurs with findings for H. uninervis (described as H. pinifoliain the earlier publication) from northern Australia in the Gulf ofCarpentaria where shoot loss started after 38 d and complete losswas predicted after 100 d (Longstaff and Dennison, 1999).
Leaf morphological changes coincided with approximately 1–2complete cycles of leaf turnover. Turnover time was not signifi-cantly different amongst species or treatments and ranged from20 to 50 d. Leaf morphological changes were first detected at day46 but at this time were restricted to leaf length changes: this doesnot require new leaf production as senescence of leaf tips can re-sult in shorter leaves. The exception to this was Z. muelleri whichhad an earlier record of shorter leaf length after just 25 d. It tookalmost two leaf replacement cycles for the other leaf morphologi-cal changes, leaf width and thickness, to occur after 74 d.
Shading intensity affected the time taken for seagrass plants torespond to the reduced light conditions. The very low treatmentsresponded first in the sequence described above. The low lighttreatment generally took longer, for example leaves per shoot re-duced after 46 d, not 14 d and leaf extension and areal leaf exten-sion took 54 d in the low light treatment, not 7 d. The additionallight available in the low light compared to the very low light treat-ment enabled extra photosynthetic production to support growthand delay a draw-down on carbohydrate reserves. This signifi-cantly delayed many of the responses even though both treatmentswere below the minimum light requirements for the species. Therewere few significant changes in the moderate light treatment (LAIand shoot density) and these were only apparent after 74 d. Thesefindings highlight the importance of shading intensity: even belowminimum light requirements the intensity of a light deprivationevent is critical to the time taken for impacts to occur.
The seasonal timing of a light reduction event can be a criticalfactor in the response of seagrasses to shading and may affectthe timelines and magnitude of response (Lavery et al., 2009). Car-
Table 4Carbohydrate concentration in the below-ground biomass of the high light treatment afte
Treatment C. serrulata
Total carbohydrate concentration (%DW) 7.89 (0.73)a
Total carbohydrate relative to leaf biomass ðg g�1leavesÞ 0.34 (0.06)a
Note: Identical letters in the same row are not significantly different (p > 0.05).
bohydrate reserves are at a maximum at the end of summer(Alcoverro et al., 2001; Collier et al., 2009) suggesting that a lightreduction impact at this time would have a delayed effect as theplants can draw on the reserves and delay other responses; thishypothesis is supported by slower responses which occurred dur-ing shading of Posidonia sinuosa when initiated in summer com-pared to winter (Collier et al., 2009; Gordon et al., 1994;Neverauskas, 1988). However, in Amphibolis griffithii, shading im-posed at the end of summer had a greater effect because watertemperatures were high post-summer resulting in a high compen-sation irradiance while ambient light intensities dropped consider-ably, exacerbating the intensity of shading (Lavery et al., 2009).With these seasonal considerations, the results of this study willbe most applicable to shading events occurring during late summerand autumn; this is the wet season in the GBR when flood plumesand low light events are the most common (Devlin et al., 2012).
4.3. Species differences
All species responded to the light treatments, but there weresmall differences amongst species in the rate they responded andlarge differences amongst species in the magnitude of response.Z. muelleri generally changed its morphology and growth ratesthe most rapidly and C. serrulata and T. hemprichii, the most slowly.The magnitude of the response also differed between species: Z.muelleri morphology and growth was the most ‘plastic’, with largeresponses in morphology and growth, generally followed by H.uninervis, and with the smallest responses in T. hemprichii and C.serrulata. These differences amongst species could reflect a numberof key differences in the life strategy and habitats of the differentspecies (Walker et al., 1999).
The species which responded the fastest and with the largestdifferences — Z. muelleri — had the smallest below-ground bio-mass, carbohydrate concentration as well as below-ground bio-mass and carbohydrate concentration relative to weight of leafmaterial. This could result in rapid utilization of reserves, makingit necessary to implement structural and meadow-scale changesto balance metabolic processes. The below-ground biomass of thelarger species C. serrulata and T. hemprichii provides a larger capac-ity to store carbohydrates reserves (Alcoverro et al., 2001; Long-staff et al., 1999) and this could delay the need to makestructural changes to balance metabolic processes. H. uninervishad the highest concentration of carbohydrates in the rhizomeand least leaf material to be supported by it and yet, despite this,H. uninervis was the second most responsive to the shade treat-ments. This species is known to have large morphological variabil-ity that is, in part, related to environmental conditions (Waycottet al., 2004).
Though these four species can overlap in distribution, there aredifferences in their dominant habitat that can affect their responseto shading. Z. muelleri and H. uninervis tend to inhabit coastalenvironments that are highly dynamic and face a range of regulardisturbances including episodic light reduction caused by pulsedrun-off and wind-driven resuspension of the fine coastal sediments(Carruthers et al., 2002; Rasheed et al., 2008). Z. muelleri, in partic-ular, often occurs in muddy estuarine habitats that other speciescannot inhabit, since, it can be quickly impacted, an essential
r 102 d. n = 4.
H. uninervis T. hemprichii Z. muelleri
10.47 (1.43)b 4.87 (0.36)c 3.36 (0.33)c
0.85 (0.33)a 0.42 (0.18)a 0.03 (0.01)b

Time (days)
0 5 10 15 20 25
Chl
orop
hyll
conc
entra
tion
(µg
cm-2)
10
20
30
10
20
30
10
20
30
10
20
30
40
Chl
orop
hyll a
/b
1.5
2.0
2.5
3.0
1.5
2.0
2.5
Time (days)
0 5 10 15 20 251.5
2.0
2.5
1.5
2.0
2.5
abcbac
abcab
aabbb
aaaa
A
B
C
D
E
F
G
H
HighModerateLowVery low
Fig. 6. Chlorophyll concentration (left) and chlorophyll a/b ratio (right) for C. serrulata (top), H. uninervis (second), T. hemprichii (third) and Z. muelleri (bottom) throughout theshading experiment. Values are means (n = 5) ± S.E. Asterisks indicate significant treatment effect. Letters indicate significant differences between treatments eitherpresented on the graph for each species when a significant species interaction occurred or at the top of the graph when results apply to all species.
352 C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354
habitat will be lost. However, these species have high productivi-ties and rapid turnover, and opportunistically grow when condi-tions are favourable (Marbà and Duarte, 1998; Walker et al.,1999). This can enable them to recover quickly from such distur-bances: following large-scale flood-related loss of Z. muellerimeadows (90% loss) in 1999 in southern Queensland, there wascomplete recovery of the meadows after just 3 years (Campbelland McKenzie, 2004). In contrast, both C. serrulata and T. hemprichiialso inhabit coastal areas, but are more common in subtidal andreef habitats (Waycott et al., 2004) where there are fewer distur-bances and light-related levels of stress are reduced. They alsohave high leaf productivity and leaf turnover rates but they tooklonger to respond to the light treatments and the changes werenot as pronounced. If severe impacts occur, the cycle of recovery
is much slower, and can take up to a decade (Birch and Birch,1984) so tolerance of low light is more critical in these species.
5. Conclusions
These Great Barrier Reef seagrass species all responded to thelow light treatments in the following sequence: metabolicchanges; shedding; and production of new, altered tissue. All spe-cies demonstrated meadow-scale impacts (shoot die-off) after46 d, with complete loss expected after 76–133 d at 1% of surfacelight, which is well below minimum light requirements. Leaf mor-phological characteristics took just as long or even longer thanshoot density to significantly change, and would therefore beunsuitable as early-warning indicators of light-related loss. In

C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354 353
contrast, physiological changes, including growth rates were rapidin onset. All of these changes are ecologically significant as produc-tion and habitat structure are ecologically important attributes ofseagrass meadows. The timelines and magnitude of response weredifferent amongst species, with Z. muelleri being impacted the fast-est and with the greatest magnitude. It is dominant in estuarineenvironments where exposure to low light events through run-off and wind-driven resuspension are common, and it can recoverin a matter of years from intense disturbances (Campbell andMcKenzie, 2004). In contrast C. serrulata and T. hemprichii werethe slowest to respond and exhibited the smallest changes. Giventheir slower recovery rates (up to 10 years) an increased toleranceto these disturbances is critical to their long-term survival. Despitesimilar projections for complete shoot loss in the low and the verylow light treatments for two of the species, the intensity of lightreduction was none-the-less critical for the degree and rate of im-pact: seagrasses in the low light treatment were impacted moreslowly and to a lesser degree than the very low light treatment.This highlights the importance of environmental management thataims to minimize declines in water quality.
References
Abal, E.G., Loneragan, N.R., Bowen, P., Perry, C.J., Udy, J.W., Dennison, W.C., 1994.Physiological and morphological responses of the seagrass Zostera capricorniAschers. to light intensity. J. Exp. Mar. Biol. Ecol. 178, 113–129.
Alcoverro, T., Manzanera, M., Romero, J., 2001. Annual metabolic carbon balance ofthe seagrass Posidonia oceanica: the importance of carbon reserves. Mar. Ecol.Prog. Ser. 211, 105–116.
Aragones, L., Lawler, I.R., Foley, W.J., Marsh, H., 2006. Dugong grazing and turtlecropping: grazing optimization in tropical seagrass ecosystems. Oecologia 149,635–647.
Beer, S., Mtolera, M., Lyimo, T., Bjork, M., 2006. The photosynthetic performance ofthe tropical seagrass Halophila ovalis in the upper intertidal. Aquat. Bot. 84,367–371.
Belshe, E.F., Durako, M.J., Blum, J.E., 2007. Photosynthetic rapid light curves (RLC) ofThalassia testudinum exhibit diurnal variation. J. Exp. Mar. Biol. Ecol. 342, 253–268.
Birch, W.R., Birch, M., 1984. Succession and pattern of tropical intertidal seagrassesin Cockle Bay, Queensland, Australia: a decade of observations. Aquat. Bot. 19,343–367.
Bulthuis, D.A., Woelkerling, W.J., 1983. Biomass accumulation and shading effects ofepiphytes on leaves of the seagrass, Heterozostera tasmanica, in Victoria,Australia. Aquat. Bot. 16, 137–148.
Campbell, S.J., McKenzie, L.J., 2004. Flood related loss and recovery of intertidalseagrass meadows in southern Queensland, Australia. Estuar. Coast. Shelf Sci.60, 477–490.
Carruthers, T.J.B., Dennison, W.C., Longstaff, B.J., Waycott, M., Abal, E.G., McKenzie,L.J., Lee Long, W.J., 2002. Seagrass habitats of northeast Australia: models of keyprocesses and controls. Bull. Mar. Sci. 71, 1153–1169.
Collier, C.J., Lavery, P.S., Ralph, P.J., Masini, R.J., 2008. Physiological characteristics ofthe seagrass Posidonia sinuosa along a depth-related gradient of lightavailability. Mar. Ecol. Prog. Ser. 353, 65–79.
Collier, C.J., Lavery, P.S., Masini, R.J., Ralph, P.J., 2009. Shade-induced response andrecovery of the seagrass Posidonia sinuosa. J. Exp. Mar. Biol. Ecol. 370, 89–103.
Costanza, R., d’Arge, R., de Groot, R., Farber, S., Grasso, M., Hannon, B., Limburg, K.,Naeem, S., O’Neill, R.V., Paruelo, J., Raskin, R.G., Sutton, P., van den Belt, M., 1997.The value of the worlds ecosystem services and natural capital. Nature 387,253–260.
Czerny, A.B., Dunton, K.H., 1995. The effects of in situ light reduction on the growthof two subtropical seagrasses, Thalassia testudinum and Halodule wrightii.Estuaries 18, 418–427.
Devlin, M.J., Harkness, P., McKinna, L.I.W., Abott, B., Brodie, J., 2012. Mapping thepollutants in surface river plume waters in the Great Barrier Reef, Australia.Marine Pollution Bulletin 65, 224–235.
Durako, M.J., Kunzelman, J.I., Kenworthy, W.J., Hammerstrom, K.K., 2003. Depth-related variability in the photobiology of two populations of Halophila johnsoniiand Halophila decipiens. Mar. Biol. 142, 1219–1228.
Enríquez, S., 2005. Light absorption efficiency and the package effect in the leaves ofthe seagrass Thalassia testudinum. Mar. Ecol. Prog. Ser. 289, 141–150.
Enríquez, S., Merino, M., Iglesias-Prieto, R., 2002. Variations in the photosyntheticperformance along the leaves of the tropical seagrass Thalassia testudinum. Mar.Biol. 140, 891–900.
Erftemeijer, P.L.A., Robin Lewis Iii, R.R., 2006. Environmental impacts of dredging onseagrasses: a review. Mar. Pollut. Bull. 52, 1553–1572.
Fabricius, K.E., 2011. Factors determining the resilience of coral reefs toeutrophication: a review and conceptual model. In: Dubinsky, Z., Stambler, N.(Eds.), Coral Reefs: An Ecosystem in Transition. Springer Science + BusinessMedia, p. 552.
Fourqurean, J.W., Zieman, J.C., 1991. Photosynthesis, respiration and whole plantcarbon budget of the seagrass Thalassia testudinum. Mar. Ecol. Prog. Ser. 69,161–170.
Gordon, D.M., Grey, K.A., Chase, S.C., Simpson, C.J., 1994. Changes to the structureand productivity of a Posidonia sinuosa meadow during and after imposedshading. Aquat. Bot. 47, 265–275.
Granger, S., Izumi, H., 2002. Water quality measurement methods for seagrasshabitats. In: Short, F.T., Coles, R.G. (Eds.), Global Seagrass Research Methods.Elsevier Science, Amsterdam, p. 482.
Grice, A.M., Loneragan, N.R., Dennison, W.C., 1996. Light intensity and theinteractions between physiology, morphology and stable isotope ratios in fivespecies of seagrass. J. Exp. Mar. Biol. Ecol. 195, 91–110.
Grubb, P.J., 1998. A reassessment of the strategies of plants which cope withshortages of resources. Perspect. Plant Ecol. Evol. Syst. 1, 3–31.
Heck, K.L., Carruthers, T.J.B., Duarte, C.M., Hughes, A.R., Kendrick, G., Orth, R.J.,Williams, S.W., 2008. Trophic transfers from seagrass meadows subsidizediverse marine and terrestrial consumers. Ecosystems 11, 1198–1210.
Heck, K.L.J., Orth, R.J., 2006. Predation in seagrass beds. In: Larkum, A.W.D., Orth, R.J.,Duarte, C.M. (Eds.), Seagrasses: Biology, Ecology and Conservation. Springer,Dordrecht.
Hyndes, G.A., Kendrick, A.J., MacArthur, L.D., Stewart, E., 2003. Differences in thespecies- and size-composition of fish assemblages in three distinct seagrasshabitats with differing plant and meadow structure. Mar. Biol. 142, 1195–1206.
Lavery, P.S., McMahon, K., Mulligan, M., Tennyson, A., 2009. Interactive effects oftiming, intensity and duration of experimental shading on Amphibolis griffithii.Mar. Ecol. Prog. Ser. 394, 21–33.
Libes, M., Boudouresque, C.-F., 1987. Uptake and long-distance transport of carbonin the marine phanerogam Posidonia oceanica. Mar. Ecol. Prog. Ser. 38, 177–186.
Longstaff, B.J., 2002. Investigations into the Light Requirements of Seagrasses inNortheast Australia. Ph.D. Thesis, University of Queensland Brisbane,Department of Botany, 152p.
Longstaff, B.J., Dennison, W.C., 1999. Seagrass survival during pulsed turbidityevents: the effects of light deprivation on the seagrasses Halodule pinifolia andHalophila ovalis. Aquat. Bot. 65, 101–121.
Longstaff, B.J., Loneragan, N.R., O’Donohue, M.J., Dennison, W.C., 1999. Effects oflight deprivation on the survival and recovery of the seagrass Halophila ovalis(R.Br.) Hook. J. Exp. Mar. Biol. Ecol. 234, 1–27.
Marbà, N., Duarte, C.M., 1998. Rhizome elongation and seagrass clonal growth. Mar.Ecol. Prog. Ser. 174, 269–280.
McKenzie, L.J., 1994. Seasonal changes in biomass and shoot characteristics of aZostera capricorni Aschers. dominant meadow in Cairns Harbour, northernQueensland. Aust. J. Mar. Freshwat. Res. 45, 1337–1352.
Neverauskas, V.P., 1988. Response of a Posidonia community to prolongedreduction in light. Aquat. Bot. 31, 361–366.
Olesen, B., Enríquez, S., Duarte, C.M., Sand-Jensen, K., 2002. Depth-acclimation ofphotosynthesis, morphology and demography of Posidonia oceanica andCymodocea nodosa in the Spanish Mediterranean Sea. Mar. Ecol. Prog. Ser. 236,89–97.
Onuf, C.P., 1996. Seagrass responses to long-term light reduction by brown tide inupper Laguna Madre, Texas: distribution and biomass patters. Mar. Ecol. Prog.Ser. 138, 219–231.
Peterson, B.J., Frankovich, T.A., Zieman, J.C., 2007. Response of seagrass epiphyteloading to field manipulations of fertilization, gastropod grazing and leafturnover rates. J. Exp. Mar. Biol. Ecol. 349, 61–72.
Preen, A.R., Long, W.J.L., Coles, R.G., 1995. Flood and cyclone related loss, and partialrecovery, of more than 1000 km2 of seagrass in Hervey Bay, Queensland,Australia. Aquat. Bot. 52, 3–17.
Ralph, P.J., Polk, S.M., Moore, K.A., Orth, R.J., Smith, W.O.J., 2002. Operation of thexanthophyll cycle in the seagrass Zostera marina in response to variableirradiance. J. Exp. Mar. Biol. Ecol. 271, 189–207.
Ralph, P.J., Durako, M.J., Enriquez, S., Collier, C.J., Doblin, M.A., 2007. Impact of lightlimitation on seagrasses. J. Exp. Mar. Biol. Ecol. 350, 176–193.
Rasheed, M.A., Unsworth, R.K.F., 2011. Long-term climate-associated dynamics of atropical seagrass meadow: implications for the future. Mar. Ecol. Prog. Ser. 422,93–103.
Rasheed, M.A., Dew, K.R., McKenzie, L.J., Coles, R.G., Kerville, S.P., Campbell, S.J.,2008. Productivity, carbon assimilation and intra-annual change in tropical reefplatform seagrass communities of the Torres Strait, north-eastern Australia.Cont. Shelf Res. 28, 2292–2303.
Romero, J., Lee, K.-S., Pérez, M., Mateo, M.A., Alcoverro, T., 2006. Nutrient dynamicsin seagrass ecosystems. In: Larkum, A.W.D., Orth, R.J., Duarte, C.M. (Eds.),Seagrasses: Biology, Ecology and Conservation. Springer, Dordrecht.
Short, F.T., Duarte, C.M., 2001. Methods for the measurement of seagrass growthand production. In: Short, F.T., Coles, R. (Eds.), Global Seagrass ResearchMethods. Elsevier Science, Amsterdam, p. 473.
Sofonia, J.J., Unsworth, R.K.F., 2009. Development of water quality thresholds duringdredging for the protection of benthic primary producer habitats. J. Environ.Monit. 12, 159–163.
Underwood, A.J., 1997. Experiments in Ecology: Their Logical Design andInterpretation using Analysis of Variance. Cambridge University Press,Cambridge.
Unsworth, R.K.F., Cullen, L.C., 2010. Recognising the necessity for Indo-Pacificseagrass conservation. Conserv. Lett. 1, 11.
Walker, D.I., Dennison, W.C., Edgar, G., 1999. Status of Australian seagrass researchand knowledge. In: Butler, A., Jernakoff, P. (Eds.), Seagrass in Australia: StrategicReview and Development of an R&D Plan. CSIRO Publishing, Victoria.

354 C.J. Collier et al. / Marine Pollution Bulletin 65 (2012) 342–354
Waycott, M., Longstaff, B.J., Mellors, J., 2005. Seagrass population dynamics andwater quality in the Great Barrier Reef region: a review and future researchdirections. Mar. Pollut. Bull. 51, 343–350.
Waycott, M., McMahon, K., Mellors, J., Calladine, A., Kleine, D., 2004. A Guide to TropicalSeagrasses of the Indo-West Pacific. James Cook University, Townsville, 72p.
Waycott, M., Duarte, C.M., Carruthers, T.J.B., Orth, R.J., Dennison, W.C., Olyarnik, S.,Calladine, A., Fourqurean, J.W., Heck, K.L., Hughes, A.R., Kendrick, G.A.,Kenworthy, W.J., Short, F.T., Williams, S.L., 2009. Accelerating loss ofseagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci.USA 106, 12377–12381.




















