
Response to flavonoids as a factor influencing competitiveness and symbiotic activity of Rhizobium...
11
Microbiological Research 165 (2010) 50—60 Response to flavonoids as a factor influencing competitiveness and symbiotic activity of Rhizobium leguminosarum Dominika Maj, Jerzy Wielbo, Monika Marek-Kozaczuk, Anna Skorupska Department of Genetics and Microbiology, Maria Curie-Skłodowska University, Akademicka 19 st., 20-033 Lublin, Poland Received 20 March 2008; received in revised form 3 June 2008; accepted 30 June 2008 KEYWORDS Rhizobium; Flavonoids; Competitiveness; Symbiotic activity Summary Flavonoids play a crucial role as signal molecules in promoting the formation of nodules by symbiotic bacteria commonly known as rhizobia. The early interaction between flavonoids and NodD regulatory protein activates nod gene transcription and the synthesis of Nod factor that initiates nodule primordium. In this study, we assessed response to flavonoids as factors influencing competitiveness of rhizobia and their symbiotic activity. Rhizobium leguminosarum nodule isolates belonging to three biovars, trifolii, viciae and phaseoli characterized earlier as competitive or uncompetitive relative to native rhizobia, were used. Investigating nodA promoter induction using plasmid lacZ fusion, we found that competitive strains more readily responded to a wide range of synthetic flavonoids and seed exudates in comparison to uncompetitive strains, albeit some exceptions were noticed. Of all the synthetic flavonoids and seed exudates studied, naringenin, hespertin and clover and vetch exudates were the most effective inducers of nodA promoter in competitive strains. Only one of the nine examined uncompetitive strains was highly induced by clover seed exudate. Subsequently, the effect of preinduction of R. leguminosarum bv. trifolii with clover exudate was assessed. Out of 18 pre-activated strains, nine strains (including competitive ones) increased clover wet mass of shoots and nodule number when used as inoculants. Our results demonstrate a plausible approach of isolating and characterizing flavonoid-responsive field isolates that could be further developed into relevant legume inoculants. & 2008 Elsevier GmbH. All rights reserved. ARTICLE IN PRESS www.elsevier.de/micres 0944-5013/$ - see front matter & 2008 Elsevier GmbH. All rights reserved. doi:10.1016/j.micres.2008.06.002 Corresponding author. Tel.: +48 81 537 5972; fax: +48 81 537 5959. E-mail address: [email protected] (A. Skorupska).
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Response to flavonoids as a factor influencing competitiveness and symbiotic activity of Rhizobium...

ARTICLE IN PRESS
Microbiological Research 165 (2010) 50—60
0944-5013/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/micres
Response to flavonoids as a factor influencingcompetitiveness and symbiotic activity ofRhizobium leguminosarum
Dominika Maj, Jerzy Wielbo, Monika Marek-Kozaczuk, Anna Skorupska�
Department of Genetics and Microbiology, Maria Curie-Skłodowska University, Akademicka 19 st.,20-033 Lublin, Poland
Received 20 March 2008; received in revised form 3 June 2008; accepted 30 June 2008
KEYWORDSRhizobium;Flavonoids;Competitiveness;Symbiotic activity
ee front matter & 2008micres.2008.06.002
ing author. Tel.: +48 81ess: [email protected]
SummaryFlavonoids play a crucial role as signal molecules in promoting the formation ofnodules by symbiotic bacteria commonly known as rhizobia. The early interactionbetween flavonoids and NodD regulatory protein activates nod gene transcriptionand the synthesis of Nod factor that initiates nodule primordium. In this study, weassessed response to flavonoids as factors influencing competitiveness of rhizobiaand their symbiotic activity. Rhizobium leguminosarum nodule isolates belonging tothree biovars, trifolii, viciae and phaseoli characterized earlier as competitive oruncompetitive relative to native rhizobia, were used. Investigating nodA promoterinduction using plasmid lacZ fusion, we found that competitive strains more readilyresponded to a wide range of synthetic flavonoids and seed exudates in comparisonto uncompetitive strains, albeit some exceptions were noticed. Of all the syntheticflavonoids and seed exudates studied, naringenin, hespertin and clover and vetchexudates were the most effective inducers of nodA promoter in competitive strains.Only one of the nine examined uncompetitive strains was highly induced by cloverseed exudate. Subsequently, the effect of preinduction of R. leguminosarum bv.trifolii with clover exudate was assessed. Out of 18 pre-activated strains, ninestrains (including competitive ones) increased clover wet mass of shoots and nodulenumber when used as inoculants. Our results demonstrate a plausible approach ofisolating and characterizing flavonoid-responsive field isolates that could be furtherdeveloped into relevant legume inoculants.& 2008 Elsevier GmbH. All rights reserved.
Elsevier GmbH. All rights reserved.
537 5972; fax: +48 81 537 5959.s.lublin.pl (A. Skorupska).

ARTICLE IN PRESS
Response to flavonoids in R. leguminosarum 51
Introduction
Rhizobia are symbiotic bacteria that have theability to establish a N2-fixing symbiosis withleguminous plants. These bacteria are taxonomi-cally and physiologically diverse members of thea and b subclasses of the Proteobacteria, and mostlycomprise the genera Rhizobium, Bradyrhizobium,Mesorhizobium, Sinorhizobium and Azorhizobium.The establishment of the symbiosis requires co-ordination of plant and bacterial gene expression,which is regulated through the mutual exchange ofmolecular signals (Perret et al., 2000; Spaink, 2000;Jones et al., 2007). The recent completion ofnucleotide sequence of seven rhizobial genomesand several earlier studies revealed that bacterialgenes responsible for nodulation and nitrogenfixation are located on the megaplasmids (pSym)in Rhizobium and Sinorhizobium, or on the chromo-some within the symbiosis islands in Mesorhizo-bium, Bradyrhizobium and Azorhizobium (forreview, see MacLean et al., 2007; Jones et al.,2007). Nodulation genes have been classified intothree categories: (i) the common nodulation genesnodABC, which are essential for nodulation and themutations in which lead to Nod� phenotype (Jacobset al., 1985; Debelle et al., 1986); (ii) host-specificity genes (nodFE, nodH, nodG, nodPQ andothers) that determine the host range of micro-symbionts and influence the rate and frequency ofnodule formation (Horvath et al., 1986; Debelleet al., 1986; Schwedock and Long, 1989) and (iii) afamily of regulatory nodD genes (Spaink, 2000).
The first signals to be exchanged between thehost plant and its rhizobial symbiont are flavonoidcompounds (2-phenyl-1.4-benzopyrone derivatives)produced by plants, which in addition to otherfunctions are required for nod gene induction(Mulligan and Long, 1985; Rossen et al., 1985).Flavonoids influence DNA binding of NodD, proteinsbelonging to the LysR family of transcriptionalregulators (Hong et al., 1987; Fisher et al., 1988;Kondorosi et al., 1989). NodD, in complex with aspecific flavonoid, binds conservative sequencesupstream of nod operons, called nod boxes, actingas a transcriptional activator of several nod genepromoters. Recent research suggests that thepresence of the appropriate flavonoids induceschanges in DNA topology at the location of NodDbinding in the promoter region, thereby allowingRNA polymerase to initiate gene transcription(Chen et al., 2005). The nod genes control thebiosynthesis of lipochitooligosaccharide Nod fac-tors, LCOs (for review, see Spaink, 2000; D’Haezeand Holsters, 2002). Nod factors are secreted bybacteria and elicit multiple responses required for
the nodulation of the appropriate host plants. Therhizobia invade the roots through the production oftubular structures called infection threads, whichpenetrate root cortex and stimulate the noduleprimordium formation (Jones et al., 2007). Primor-dial cell division is preceded by the inhibition ofauxin transport caused by endogenous root flavo-noids, indicating that these compounds play acrucial role as developmental signals in the initia-tion of nodule primordia, as evidenced in Medicagotruncatula and soybean roots (Mathesius et al.,1998; Wasson et al., 2006; Subramanian et al.,2006).
The interaction between NodD and flavonoids isassumed to be specific. For example, luteolin(30,40,5,7-tetrahydoxyflavone) is a specific nod geneinducer in Sinorhizobium meliloti (Peters et al.,1986; Peck et al., 2006), naringenin (5,7,40-trihy-droxyflavanone) induces nod genes in Rhizobiumleguminosarum bv. viciae (Zaat et al., 1987, 1988)and 7,40-dihydroxyflavone activates nod genes inR. leguminosarum bv. trifolii and Bradyrhizobiumsp. (Redmond et al., 1986; Djordjevic et al., 1987;Zaat et al., 1988, 1989; Kosslak et al., 1987). Onthe other hand, nod genes from different rhizobiamay respond to different sets of flavonoids (seereview, Cooper, 2004, 2007). It has been documen-ted that some flavonoids present in the root or seedextracts may be symbiotically ineffective or eveninhibit nodulation. Mixtures of flavonoids and othercompounds that are exuded by plant roots arethought to act as signals that influence the abilityof rhizobia to colonize the roots, survive in therhizosphere, and affect the competitiveness ofrhizobia and symbiotic interactions with legumes(Cooper, 2004).
Evidence exist that introduction of exogenousnod gene inducers increases nodulation of somelegume species. For example, the addition ofluteolin to certain alfalfa cultivars significantlyincreased nodulation (Kapulnik et al., 1987). Like-wise, bean nodulation by Rhizobium etli or Rhizo-bium tropici was enhanced by the addition ofquercetin (Hungria and Phillips, 1993); pre-treat-ment of Bradyrhizobium japonicum with genisteinincreased nodulation and grain yield of soybean(Glycine max) (Zhang and Smith, 1995, 1996); andpre-induction of R. leguminosarum with hesperetinand naringenin was found to stimulate nodulationand plant dry matter accumulation of pea and lentilplants (Begum et al., 2001).
In this study, we investigated the role offlavonoid responsiveness as a factor possibly affect-ing the competitiveness of rhizobia in nodulatinglegume plants. We assessed the response ofPnodA–lacZ transcriptional fusion introduced into

ARTICLE IN PRESS
D. Maj et al.52
R. leguminosarum bv. trifolii, bv. viciae and bv.phaseoli strains characterized earlier as competi-tive and uncompetitive to several different flavo-noids and seed exudates. The competitive strainswere selected from among the R. leguminosarumisolates of agronomic important legumes (clover,pea, vetch, tick bean and bean) on the basis oftheir ability to plant growth promotion, colonizehost nodules and their metabolic characteristics(Wielbo et al., 2007). The isolates tagged with gusAgene were examined in relation to indigenousrhizobia and were classified as competitive basedon the range of nodule occupation from 7% to 42%.Uncompetitive strains occupied less than 1% of thenodules.
The aim of the study was to determine whetherthe responsiveness of rhizobia to flavonoid induc-tion was correlated with strain competitiveness.We hypothesized that the greater efficiency of theearly interactions between rhizobia and the rootexudates enhances the symbiotic activity of themicrosymbionts. Subsequently, the effect of pre-activation of several R. leguminosarum bv. trifoliiisolates with host seed exudates on their ability tonodulate clover and fix nitrogen was also assessed.
Materials and methods
Bacterial strains and growth conditions
All rhizobial strains had been derived fromlegume plants growing on different types ofcultivated soils in the region of Lublin, Poland(Wielbo et al., 2007). R. leguminosarum bv. trifoliistrains KO17, KO24, KO25 and other isolatesused for plant tests were isolated from nodulesof red clover (Trifolium pratense L. cv. Dajana);R. leguminosarum bv. viciae strains GR08, GR09,PE07, PE08, PE13 and PE15 from pea (Pisumsativum L. cv. Ramrod), BO03, BO07 and BO14from tick bean (Vicia faba ssp. minor L. cv. Titus)and WY05 from vetch (Vicia sativa L. cv. Wista),respectively. R. leguminosarum bv. phaseoli strainFA02, FA03, FA21, FA22 and FA23 were isolated fromnodules of bean (Phaseolus vulgaris L. cv. ZlotaSaxa). All strains were maintained and grown onmannitol yeast extract agar 79CA medium (Vincent1970). To study the induction activity of flavonoids,M1 medium containing 18.7mM NH4Cl, 11.5mMK2HPO4, 3.7mM KH2PO4, 1.7mM NaCl, 0.8mMMgSO4 �H2O, 0.01mM FeCl3, 1% glycerol, 1 mg/mlthiamine HCl, 0.5 mg/ml biotin and 1 mg/ml pantho-tenate was used (Janczarek et al., 1997). Escheri-chia coli strains were cultured in LB medium(Sambrook et al., 1989). When needed, tetracy-
cline (Tc) was added to 79CA or LB media at theconcentration of 10 mgml�1.
Construction of plasmid vector containingnodA promoter–lacZ fusion
Standard techniques were used for plasmidisolation, restriction enzyme digestion, agarosegel electrophoresis and DNA cloning (Sambrooket al., 1989). DNA of the pMP220 vector (Spaink et al.,1987) was isolated and digested with EcoRI/PstIrestriction enzymes. A 125 bp fragment, con-taining nodA promoter, was amplified by PCRusing genomic DNA of R. leguminosarum bv. viciaestrain BO03 as template and primers NodA1:50 AAATCGGGCCCCTGCAGGGC 30, and NodA2:50 ATGGGTTGAATTCCCATTCCATAGATGAT 30 (Chenet al., 2005). The recognition sites for PstI andEcoRI in the primers were incorporated into theprimer sequences (underlined). The amplifiedfragment was inserted upstream of a promoterlesslacZ gene in the respective restriction sites ofplasmid pMP220 and the resulting construct,pMPA221, was transferred into several rhizobialisolates by electroporation. To 90 ml (about 5� 108)of electrocompetent cells (Garg et al., 1999), 5 ml(about 50 ng) of plasmid DNA was added andincubated 30min on ice. Following this, thebacterial cells were electroporated (at 1800 V,200O, 50 mF), and then incubated for 24 h at 28 1Cin liquid 79CA medium and plated on 79CA agarmedium with tetracycline. The Tcr colonies werepurified by successive isolation and passages in theselective medium.
Preparation of exudates from sprouted seeds
After surface sterilization, clover, pea, vetch,bean and tick bean seeds were shaken in sterilewater, in darkness, for 4 days at 28 1C, within whichtime the seeds germinated. After removal of thesprouted seeds, the supernatant was extractedwith ethyl acetate in the ratio 10:1 (vol/vol). Ethylacetate was evaporated and the obtained pelletresolubilized in 95% ethanol (Janczarek et al.,1997) and stored at 4 1C. To determine the amountof flavonoids, the ethanol extracts were dried andweighed. The approximate flavonoid concentrationin seed exudates was calculated relative to themolecular weight of authentic flavone. The sourcesof commercially available flavonoids were asfollows: hespertin from Sigma-Aldrich (Germany),quercetin, apigenin, 7,40-dihydroxyflavone, narin-genin, luteolin, genistein and daidzein from

ARTICLE IN PRESS
Response to flavonoids in R. leguminosarum 53
Indofine (USA). The compounds were prepared as5mM stocks in 70% ethanol.
Assay of PnodA–lacZ promoter fusion activity
The derivatives of R. leguminosarum bv. trifolii,bv. viciae and bv. phaseoli harbouring pMPA221plasmid with PnodA–lacZ fusion were used to studythe effect of synthetic flavonoids including apigen-in, 7,40-dihydroxyflavone, luteolin, daidzein, genis-tein, hespertin, naringenin, quercetin and seedexudates of clover, vetch, pea, bean and tick bean.Overnight cultures of rhizobia in 79CA mediumsupplemented with tetracycline were centrifuged,washed twice with M1 medium and resuspended inM1 to OD550 of 0.04. The synthetic flavonoids orexudates from sprouted seeds were added to M1culture at concentrations of 1, 2, 5 and 10 mM. Thecultures were grown for additional 24 h at 28 1C andtheir growth was monitored as the optical densityat 600 nm. The level of PnodA transcriptionalactivity was determined by b-galactosidase activitymeasurements with the o-nitrophenyl-b-D-galacto-side (ONPG, Sigma) assay (Miller, 1972). The resultswere expressed as b-galactosidase (Miller) units.
Plant tests
Red clover seeds (T. pratense L. cv. Dajana) weresurface sterilized, germinated and grown on nitro-gen-free F(ahraeus medium slants (Vincent, 1970).Logarithmic cultures of R. leguminosarum bv.trifolii strains grown in M1 medium were treatedwith sterile sprouted clover seed exudates at thefinal concentration of 2 mM and incubated at 28 1Cwith shaking for 16 h. Following incubation, cloverseedlings growing on agar slants were inoculatedwith 0.2ml of a cell suspension at approximatedensity of 1.0� 109 cells/ml. Clover (one plant pertube) was grown in a greenhouse under naturallight supplemented with artificial light (14 h day/10 h night, at 24/19 1C). After 5 weeks, the plantswere harvested for nodule counts and the effi-ciency of nitrogen fixation was estimated byweighing fresh mass of shoots and roots. For eachstrain, 30 clover plants were used and threeindependent experiments were conducted.
Figure 1. Genetic map of plasmid pMP221. The arrowabove the map shows direction of PnodA promoter-drivingtranscription of lacZ gene. Restriction sites used inplasmid pMP221 construction are shown.
Statistical data analysis
Data on the shoot and root matter yield andnodule quantity of red clover were subjected toanalysis of variance using the ANOVA module of theSTATISTICA program.
Results
Induction of PnodA–lacZ activity by flavonoids
In a previous study, several nodule isolates ofcommonly cultivated legumes, such as clover,vetch, pea, bean and tick bean, were selected ascompetitive and highly effective in plant growthpromotion (Wielbo et al., 2007). Currently, weproceeded to answer the question whether acorrelation exists between strain competitivenessand the level of flavonoid-induced nodA promoteractivity, i.e., whether the strains selected ascompetitive or uncompetitive differ in the levelof PnodA response to flavonoid inducers. nodA is thefirst gene in nodABC operon preceded by a typicalinducible nod promoter containing the conservednod box (Spaink et al., 1987; Chen et al., 2005).The expression of nodA gene is positively regulatedby the trans-activator NodD protein that binds tonod box and upon interaction with the flavonoidactivates transcription of the nodABC operon (Honget al., 1987).
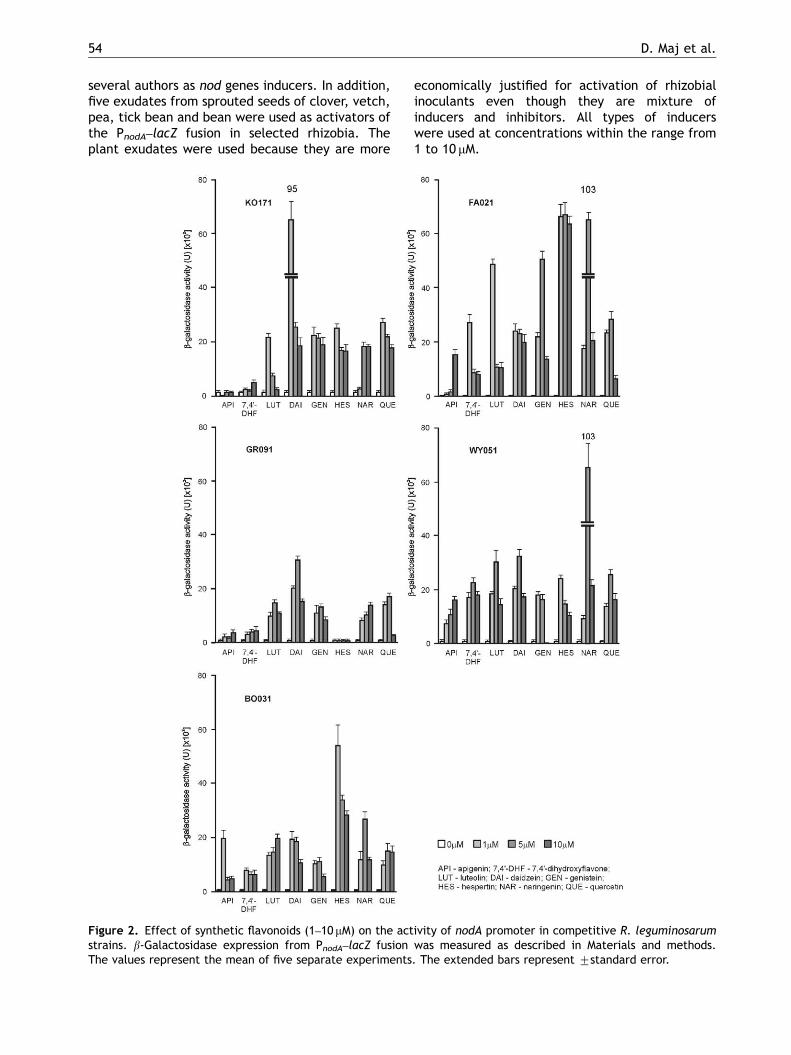
To assess the flavonoid nodA promoter activationpotential, several competitive or uncompetitiveR. leguminosarum bv. viciae, bv. trifolii and bv.phaseoli strains (Wielbo et al., 2007) harbouringplasmid pMPA221 (Figure 1) were used. We inves-tigated eight authentic, structurally differentflavonoids, including flavones (7,40-dihydroxyfla-vone, apigenin, luteolin), isoflavones (daidzein,genistein), flavanones (hespertin, naringenin) andflavonols (quercetin) that were earlier described by
ARTICLE IN PRESS
D. Maj et al.54
several authors as nod genes inducers. In addition,five exudates from sprouted seeds of clover, vetch,pea, tick bean and bean were used as activators ofthe PnodA–lacZ fusion in selected rhizobia. Theplant exudates were used because they are more
Figure 2. Effect of synthetic flavonoids (1–10 mM) on the actstrains. b-Galactosidase expression from PnodA–lacZ fusionThe values represent the mean of five separate experiments
economically justified for activation of rhizobialinoculants even though they are mixture ofinducers and inhibitors. All types of inducerswere used at concentrations within the range from1 to 10 mM.
ivity of nodA promoter in competitive R. leguminosarumwas measured as described in Materials and methods.. The extended bars represent 7standard error.
ARTICLE IN PRESS
Response to flavonoids in R. leguminosarum 55
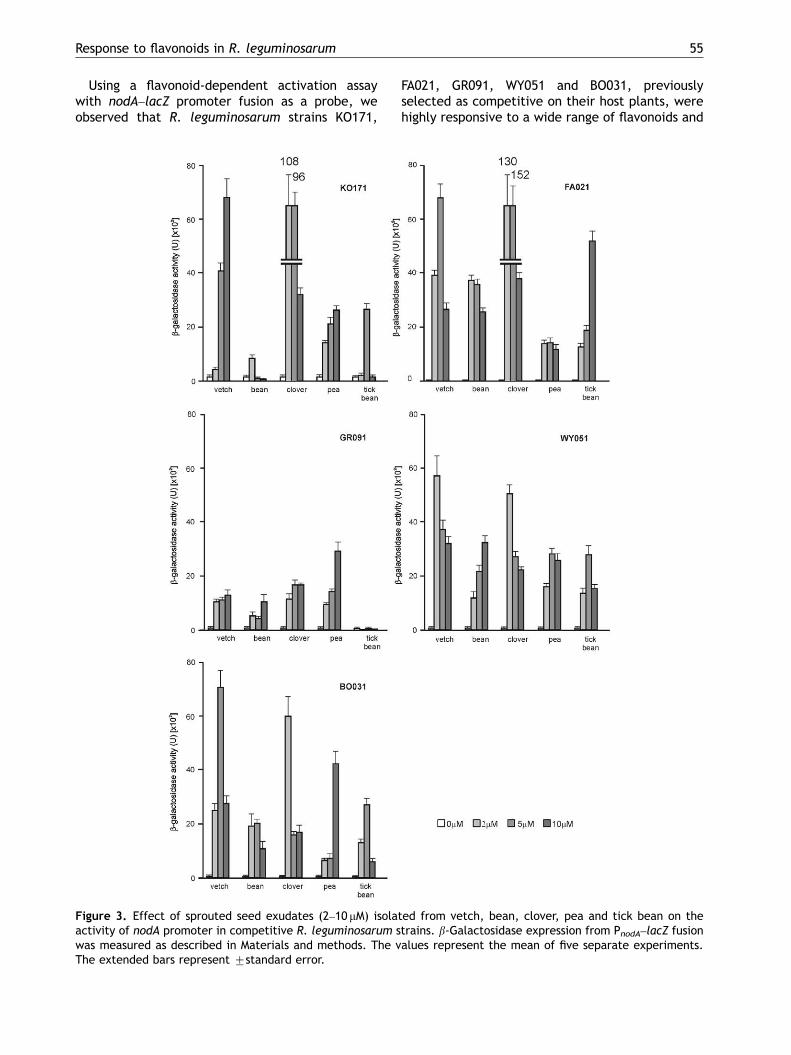
Using a flavonoid-dependent activation assaywith nodA–lacZ promoter fusion as a probe, weobserved that R. leguminosarum strains KO171,
Figure 3. Effect of sprouted seed exudates (2–10 mM) isolaactivity of nodA promoter in competitive R. leguminosarum swas measured as described in Materials and methods. The vThe extended bars represent 7standard error.
FA021, GR091, WY051 and BO031, previouslyselected as competitive on their host plants, werehighly responsive to a wide range of flavonoids and
ted from vetch, bean, clover, pea and tick bean on thetrains. b-Galactosidase expression from PnodA–lacZ fusionalues represent the mean of five separate experiments.
ARTICLE IN PRESS
D. Maj et al.56
the best inducer and its optimal concentration wasdetermined in each case (Figure 2). Unexpectedly,the R. leguminosarum bv. trifolii KO171 was mostefficiently induced by 2 mM daidzein, to a lesserextent by genistein, hespertin, naringenin andquercetin, but poorly in the presence of 7,40-dihydroxyflavone, which was earlier described asthe specific nod gene inducer in R. leguminosarumbv. trifolii (Djordjevic et al., 1987; Redmond et al.,1986). Moreover, this compound seemed to be arelatively weak PnodA inducer of R. leguminosarumbv. viciae GR091 or BO031 strains (Figure 2). GR091was poorly activated by all flavonoids used in thisstudy in comparison to other strains. BO031 andWY051 strains were moderately induced by allflavonoids with the exception of high induction ofWY051 in the presence of 5 mM naringenin. In thecase of R. leguminosarum bv. phaseoli strain FA021,the level of b-galactosidase activity was very highespecially in the presence of hespertin and 5 mMnaringenin (Figure 2).
The same competitive strains were treated withexudates prepared from sprouted seeds of clover,vetch, pea, bean and tick bean (Figure 3).Generally, the PnodA activity was much higher inresponse to exudates relative to synthetic flavo-noids. The clover and vetch exudates proved to bethe most potent inducer for all but one (GR091)competitive rhizobial strains (Figure 3). We haveobserved a nearly 50-fold induction of the PnodA–
lacZ fusion in the R. leguminosarum bv. phaseolistrain FA021 in the presence of 5 mM clover exudate(Figure 3). The host-specific bean exudate eliciteda much lower induction. Clover exudate was the
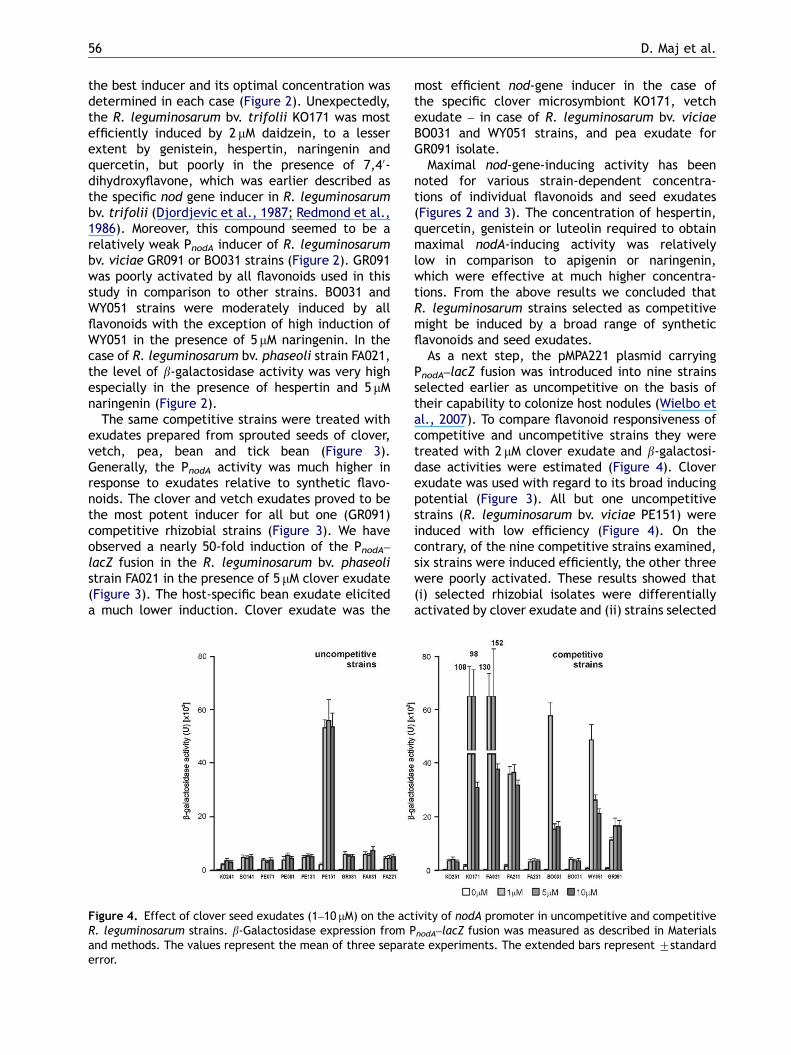
Figure 4. Effect of clover seed exudates (1–10 mM) on the actR. leguminosarum strains. b-Galactosidase expression from Pand methods. The values represent the mean of three separaerror.
most efficient nod-gene inducer in the case ofthe specific clover microsymbiont KO171, vetchexudate – in case of R. leguminosarum bv. viciaeBO031 and WY051 strains, and pea exudate forGR091 isolate.
Maximal nod-gene-inducing activity has beennoted for various strain-dependent concentra-tions of individual flavonoids and seed exudates(Figures 2 and 3). The concentration of hespertin,quercetin, genistein or luteolin required to obtainmaximal nodA-inducing activity was relativelylow in comparison to apigenin or naringenin,which were effective at much higher concentra-tions. From the above results we concluded thatR. leguminosarum strains selected as competitivemight be induced by a broad range of syntheticflavonoids and seed exudates.
As a next step, the pMPA221 plasmid carryingPnodA–lacZ fusion was introduced into nine strainsselected earlier as uncompetitive on the basis oftheir capability to colonize host nodules (Wielbo etal., 2007). To compare flavonoid responsiveness ofcompetitive and uncompetitive strains they weretreated with 2 mM clover exudate and b-galactosi-dase activities were estimated (Figure 4). Cloverexudate was used with regard to its broad inducingpotential (Figure 3). All but one uncompetitivestrains (R. leguminosarum bv. viciae PE151) wereinduced with low efficiency (Figure 4). On thecontrary, of the nine competitive strains examined,six strains were induced efficiently, the other threewere poorly activated. These results showed that(i) selected rhizobial isolates were differentiallyactivated by clover exudate and (ii) strains selected
ivity of nodA promoter in uncompetitive and competitive
nodA–lacZ fusion was measured as described in Materialste experiments. The extended bars represent 7standard
ARTICLE IN PRESS
Response to flavonoids in R. leguminosarum 57
as competitive more readily responded to flavonoidactivation than uncompetitive.
Effect of clover seed exudate on nodulationand growth promotion
In the next stage of our investigation, wedetermined whether flavonoid pre-treatment ofrhizobia influences their symbiotic activity. Wechose 18 R. leguminosarum bv. trifolii strains fromour collection of clover nodule isolates, among themthe KO171 and KO181 strains earlier selected ascompetitive, and KO241 and KO271 characterized asuncompetitive, all harbouring pMPA221 plasmid. Therhizobial cultures were treated with 2 mM cloverseed exudate and the bacteria were used asinoculants of clover seedlings. Clover shoot and rootwet mass and nodule number were assessed as themost important symbiotic parameters and measureof symbiotic efficiency (Table 1). On the basis ofability to plant growth promotion, 18 flavonoid pre-activated isolates could be divided into two groups.
Table 1. Effect of clover seed exudate pre-activation of R.promotion, nodulation and nodA promoter activity
Strain Wet mass of shoots (mg) Wet mass of root
Non-induced
Induced Non-induced
Ind
Group 1KO181 97.479.8 109.977.8 52.877.6 52KO171 51.0*73.7 96.1*710.6 16.471.9 38K4.91 68.9713.3 92.477.5 47.5711.1 34K8.81 70.377.8 76.175.5 41.378.8 31K5.61 59.176.9 73.373.4 28.474.0 34K3.81 58.777.5 72.679.2 21.273.2 22K4.111 58.778.3 65.378.3 24.374.4 36K10.41 60.579.8 69.276.8 25.773.0 23K2.41 47.975.8 53.677.2 13.971.5 22
Group 2KO241 120.576.4 111.877.8 58.176.7 63K3.221 106.177.2 104.276.3 61.978.2 52K1.131 94.4712.2 92.579.3 41.479.5 38K10.81 82.075.0 73.978.3 35.172.9 27K3.161 61.178.3 63.178.1 34.075.3 27K10.71 55.675.4 56.675.0 15.372.1 16KO271 66.775.7 49.175.3 27.374.5 19K3.131 44.573.9 37.276.3 24.574.0 14K4.171 30.774.2 22.872.0 21.872.2 26
The values represent the means from three independent experiments7 Represent standard error. Stars denote significant differences (Ppof plants inoculated with induced and non-induced strains.ab-Galactosidase activity of R. leguminosarum bv. trifolii strains waThe values represent means of four b-galactosidase assays after sub
Nine strains (group 1) stimulated green wet massgain and nodule quantity, as compared to the non-treated strains. However, in this group, a statisti-cally significant increase was observed only withregard to shoot mass with the competitive strainKO171, and for nodule formation with strain K4.111.In the remaining nine pre-activated strains(group 2), wet shoot mass and nodule number werecomparable or lower than untreated control. It isworth noting that the uncompetitive KO241 andKO271 strains were found in group 2 which did notrespond to pre-activation.
We also studied the responsiveness of PnodA–lacZfusion to clover exudate in 18 clover isolates usedin this experiment (Table 1). A high level ofinduction (above 1000 Miller U) was observed infive of nine strains of group 1 and only in three ofnine strains of group 2. In conclusion, although, wedid not observe a direct relationship between thelevel of flavonoid responses of rhizobial isolates andefficiency of symbiosis, we showed that strains thathighly responded to exudates more frequentlyincreased symbiotic activities.
leguminosarum bv. trifolii strains on clover plant growth
s (mg) Number of nodules b-Galactosidaseactivity (Millerunits)a
uced Non-induced
Induced
.075.7 7.571.2 11.171.2 301731
.774.9 6.371.0 9.071.2 20707390
.074.7 7.171.4 6.971.0 339749
.774.7 6.570.8 7.270.9 396753
.473.4 5.470.8 10.671.2 35357821
.174.0 7.371.0 8.971.6 47337519
.973.8 8.6*71.1 22.3*73.6 541753
.473.3 7.371.0 6.771.3 54817248
.173.2 4.670.7 5.271.1 23247514
.177.9 10.671.5 11.772.0 346743
.676.1 9.571.5 8.170.7 279718
.576.3 6.370.9 8.671.0 26947601
.175.0 7.570.7 8.471.6 358743
.073.0 21.972.8 17.671.6 17527609
.672.4 8.471.0 6.670.8 623745
.173.9 6.971.0 5.270.6 460737
.372.4 7.871.3 4.570.9 369738
.174.0 16.172.2 18.573.2 50597442
. Thirty plants were used for each treatment at each experiment.0.05, t test) in wet mass of shoots and nodule number in the case
s determined after 16 h exposure to 2mM clover seed exudate.traction of background activity; 7 represent standard error.

ARTICLE IN PRESS
D. Maj et al.58
Discussion
Individual flavonoids and seed exudates com-prised by a mixture of various compounds, includingflavonoids, function as nod gene inducers involvedin early signalling (Hungria and Phillips, 1993). Seedexudates, however, also contain flavonoids that actas anti-inducers (antagonists) of nod gene expres-sion (Cooper, 2004, 2007). Recently, Peck et al.(2006) have shown that non-inducing flavonoids actas competitive inhibitors of the inducing flavonoidsin S. meliloti, preventing NodD1 from activatingnod gene transcription. The authors proposed thatboth the inducing and non-inducing flavonoidsstabilize the NodD1 conformers bound to nod genepromoters but that only luteolin-bound NodD1 iscapable of undergoing the conformational changesrequired to initiate nod gene transcription (Peck etal., 2006). The authors confirmed the specificity ofnod gene induction by flavonoids (see also reviewby Cooper, 2007).
Based on these studies, we hypothesized thatamong other factors affecting the competitivenessof Rhizobium strains, the rate of response toflavonoids could play a role and thus affect earlyphases of legume–microbe interactions. In ourearlier work (Wielbo et al., 2007), we selectedR. leguminosarum nodule isolates that were com-petitive in soil environment and were distinguishedfrom other isolates by high rate of symbiotic plantgrowth promotion and the capability to utilize awide range of carbon and energy substrates. Fromthe set selected for the analysis, five competitivestrains (belonging to three biovars of R. legumino-sarum) harbouring PnodA–lacZ fusion readily re-sponded to a wide range of synthetic flavonoidsand seed exudates. Naringenin, hespertin andclover and vetch exudates were the most effectiveinducers of nodA promoter. Unexpectedly, theisoflavonoid daidzein was also found to be aneffective inducer of nodA promoter, particularlyin the case of R. leguminosarum bv. trifolii. Thiscompound that derives from the flavone intermedi-ate naringenin was previously reported to be aninhibitor of nod genes in R. leguminosarum bv.viciae (Begum et al., 2001; Cooper, 2004) and aninducer in Rhizobium spp. (Bolanos-Vasquez andWerner, 1997). Daidzein and naringenin weredescribed as the major component of exudates ofthe common bean seedlings that were uninfected,as well as those infected by specific strains ofR. leguminosarum bv. phaseoli. However, the mostpotent nodA promoter inducers in the competitivestrains of R. leguminosarum were clover and vetchexudates, indicating the possibility of synergisticeffects between several nod-gene-activating sub-
stances (Figures 2 and 3). This would not beunexpected as combinations of several inducerspresent in plant exudates may be more effectivethan isolated flavonoid compounds (Zaat et al.,1989; Begum et al., 2001). On the other hand,the specificity of nodA promoter induction wasnot apparent using experimental approachesemployed in this study. A comparable high level ofb-galactosidase induction was achieved in the caseof FA021 strain using a broad spectrum of suchflavonoids as naringenin, luteolin, hespertin andgenistein, and clover, vetch or bean seed exudates.Stimulation of nodulation genes with multiple in-ducers may be perceived as an advantageous featureof the symbiotic interaction (Cooper, 2004). Thiswould not exclude the specific interaction of NodDprotein with individual flavonoids to increase bindingof this protein to nod promoter (Peck et al., 2006).
Treating a set of competitive and uncompetitivestrains with clover exudates to assess the respon-siveness of nodA promoter, we found only one ofnine uncompetitive strains to be highly induced(Figure 4). In contrast, under the same conditions,five of nine competitive strains were highlyactivated by clover exudate. Taken together, weconcluded that the level of PnodA induction wasstrain-specific and it might constitute one of thefactors influencing strain competitiveness. Compe-tition for nodulation in the natural environment(i.e. the soil) could depend on greater efficiencyand speed of the interaction between a parti-cular bacterial strain and flavonoid compounds,the crucial signal molecules in rhizobium–legumesymbiosis.
The potential of seed exudates as nod-geneinducers could be exploited in the symbioticactivation of selected rhizobia prior to their useas inoculants. Flavonoid inducers act in low con-centrations and the pre-activation of rhizobia usedas inoculants (biofertilizers) is undoubtedly eco-nomically justified. Pre-activation of strains mightincrease rhizobial competitiveness in the soilenvironment (Zhang and Smith, 1995, 1996).Indeed, it has been shown that flavonoid pre-activated B. japonicum increased soybean nodulequantity and weight (about 30%), the seasonallevels of N2 fixation (35%) and yields (10–40%) whencompared to conventional inoculants (Zhang andSmith, 2002). Likewise, field pea and lentil plantsdisplayed increased nodulation and biomass pro-duction when inoculated with R. leguminosarumpre-induced with hesperetin (Begum et al., 2001).Improving legume inoculation efficiency is extre-mely important when considering the economic andenvironmental costs associated with the use ofchemical nitrogen fertilizers in agriculture.

ARTICLE IN PRESS
Response to flavonoids in R. leguminosarum 59
In our study, pre-treatment of R. leguminosarumbv. trifolii strains with 2 mM clover seed exudatestimulated wet mass of shoots and increased nodulenumber with nine of 18 studied strains. However,the extent of the increase of symbiotic activity incomparison with the uninduced strains varied withthe rhizobial strains and, in most cases, was notstatistically significant. The pre-induced competi-tive strains KO181 and KO171, contrary to the non-competitive strains KO241 and KO271, stimulatedbiomass production and nodule number. Never-theless, we did not observe a direct correlationbetween nodA induction and stimulation of sym-biosis. The level of nodA promoter induction wasstrain-dependent, and the isolates that more read-ily responded to exudate treatment were alsofound to more frequently stimulate symbioticactivities of R. leguminosarum bv. trifolii. Inconclusion, although we were unable to unambigu-ously ascertain whether pre-activation of rhizobiawith flavonoids invariably stimulates symbiosis, weestablished an experimental approach sufficient forselection of field isolates, which repeatedly re-spond to flavonoid pre-activation and that can befurther developed into legume inoculants.
Acknowledgement
This research was supported by the Ministry ofEducation and Science, Grant no. PBZ-KBN-112/PO6/2005.
References
Begum AA, Leibovitch S, Migner P, Zhang F. Specificflavonoids induced nod gene expression and pre-activated nod genes of Rhizobium leguminosarumincreased pea (Pisum sativum L.) and lentil (Lensculinaris L.) nodulation in controlled growth chamberenvironments. J Exp Bot 2001;52:1537–43.
Bolanos-Vasquez MC, Werner D. Effects of Rhizobiumtropici, R. etli, and R. leguminosarum bv. phaseoli onnod gene-inducing flavonoids in root exudates ofPhaseolus vulgaris. Mol Plant-Microbe Interact1997;10:339–46.
Chen H-C, Feng J, Hou B-H, Li F-Q, Li Q, Hong G-F.Modulating DNA bending affects NodD-mediated tran-scriptional control in Rhizobium leguminosarum.Nucleic Acid Res 2005;33:2540–8.
Cooper JE. Multiple responses of rhizobia to flavonoidsduring legume root infection. Adv Bot Res2004;41:1–62.
Cooper JE. Early interactions between legumes andrhizobia: disclosing complexity in a molecular dialo-gue. J Appl Microbiol 2007;103:1355–65.
Debelle F, Rosenberg C, Vasse J, Maillet F, Martinez E,Denarie J, et al. Assignment of symbiotic develop-mental phenotypes to common and specific nodulation(nod) genetic loci of Rhizobium meliloti. J Bacteriol1986;168:1075–86.
D’Haeze W, Holsters M. Nod factor structures, responses,and perception during initiation of nodule develop-ment. Glycobiology 2002;12:79–105.
Djordjevic MA, Redmond JW, Batley M, Rolfe BG. Cloverssecrete specific phenolic compounds which eitherstimulate or repress nod gene expression in Rhizobiumtrifolii. EMBO J 1987;6:1173–9.
Fisher RF, Egelhoff TT, Mulligan JT, Long SR. Specificbinding of Rhizobium meliloti extracts containingnodD to DNA sequences upstream of inducible nodula-tion genes. Genes Dev 1988;2:282–93.
Garg B, Dogra RC, Shama PK. High-efficiency transforma-tion of Rhizobium leguminosarum by electroporation.Appl Environ Microbiol 1999;65:2802–4.
Hong GF, Burn JE, Johnston AWB. Evidence that DNAinvolved in the expression of nodulation (nod) genes inRhizobium binds to the product of the regulatory genenodD. Nucleic Acids Res 1987;15:9677–90.
Horvath B, Kondorosi E, John M, Schmidt J, Torok I,Gyorgypal Z, et al. Organization, structure andsymbiotic function of Rhizobium meliloti nodulationgenes determining host specificity for alfafa. Cell1986;46:335–43.
Hungria M, Phillips DA. Effects of a seed color mutationon rhizobial nod-gene-inducing flavonoids and nodula-tion in common bean. Mol Plant-Microbe Interact1993;6:418–22.
Jacobs TW, Egelhoff TT, Long SR. Physical and geneticmap of a Rhizobium meliloti nodulation gene regionand nucleotide sequence of nodC. J Bacteriol1985;162:469–76.
Janczarek M, Urbanik-Sypniewska T, Skorupska A. Effectof authentic flavonoids and the exudate of clover rootson growth rate and inducing ability of nod genes ofRhizobium leguminosarum bv. trifolii. Microbiol Res1997;152:93–8.
Jones KM, Kobayashi H, Davies BW, Taga ME, Walker GC.How symbionts invade plants: the Sinorhizobium-Medicago model. Nature 2007;5:619–33.
Kapulnik Y, Joseph CM, Phillips DA. Flavone limitations toroot nodulation and symbiotic nitrogen fixation inalfalfa. Plant Physiol 1987;84:1193–6.
Kondorosi E, Gyuris J, Schmidt J, John E, Hofmann DB,Schell J, et al. Positive and negative control of nodgene expression in Rhizobium meliloti is reqiured foroptimal nodulation. EMBO J 1989;8:1331–41.
Kosslak RM, Bookland R, Barkei R, Paaren HE, AppelbaumER. Induction of Bradyrhizobium japonicum commonnod genes by isoflavones isolated from Glycine max.Proc Natl Acad Sci USA 1987;84:7428–32.
MacLean AM, Finan TM, Sadowsky MJ. Genomes of thesymbiotic nitrogen-fixing bacteria of legumes. PlantPhysiol 2007;144:615–22.
Mathesius U, Schlaman HRM, Spaink HP, Sauter C, RolfeBG, Djordjevic MA. Auxin transport inhibition precede

ARTICLE IN PRESS
D. Maj et al.60
root nodule formation in white clover roots and isregulated by flavonoid and derivatives of chitinoligosaccharides. Plant J 1998;14:23–34.
Miller J. Experiments in molecular genetics. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory; 1972.
Mulligan JT, Long SR. Induction of Rhizobium melilotinodC expression by plant exudate requires nodD. ProcNatl Acad Sci USA 1985;82:6609–13.
Peck MC, Fisher RF, Long SR. Diverse flavonoids stimulateNodD1 binding to nod gene promoters in Sinorhizo-bium meliloti. J Bacteriol 2006;188:5417–27.
Perret X, Staehelin C, Broughton WJ. Molecular basis ofsymbiotic promiscuity. Microbiol Mol Biol Rev 2000;64:180–201.
Peters NK, Frost JW, Long SR. A plant flavone, luteolin,induces expression of Rhizobium meliloti nodulationgenes. Science 1986;233:917–1008.
Redmond JW, Batley M, Djordjevic MA, Innes RW,Kuempel PL, Rolfe BG. Flavones induce expression ofnodulation genes in Rhizobium. Nature 1986;323:632–4.
Rossen L, Shearman CA, Johnston AWB, Downie JA. ThenodD gene of Rhizobium leguminosarum is autoregu-latory and in the presence of plant exudate inducesthe the nodABC genes. EMBO J 1985;4:3369–73.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: alaboratory manual. Cold Spring Harbor LaboratoryPress: Cold Spring Harbor, NY; 1989.
Schwedock J, Long SR. Nucleotide sequence and proteinproducts of two new nodulation genes of Rhizobiummeliloti, nodP and nodQ. Mol Plant-Microbe Interact1989;2:181–94.
Spaink HP. Root nodulation and infection factors pro-duced by rhizobial bacteria. Annu Rev Microbiol2000;54:25–288.
Spaink HP, Okker RJH, Wijffelman CA, Pees E, LugtenbergBJJ. Promoters in the nodulation region of theRhizobium leguminosarum Sym plasmid pRL1JI. PlantMol Biol 1987;9:29–37.
Subramanian S, Stacey G, Yu O. Endogenous isoflavonesare essential for the establishment of symbiosis
between soybean and Bradyrhizobium japonicum.Plant J 2006;48:261–73.
Vincent JM. A manual for the practical study of root-nodule bacteria. IBP Handbook no. 15. Oxford: Black-well Scientific; 1970. p. 29–50.
Wasson AP, Pellerone FI, Mathesius U. Silencingthe flavonoid pathway in Medicago truncatulainhibits root nodule formation and prevents auxintransport regulation by rhizobia. Plant Cell 2006;18:1617–29.
Wielbo J, Marek-Kozaczuk M, Kubik-Komar A, SkorupskaA. Increased metabolic potential of Rhizobium spp. isassociated with bacterial competitiveness. Can JMicrobiol 2007;53:957–67.
Zaat SA, Wijffelman CA, Spaink HP, van Brussel AAN,Okker RJ, Lugtenberg BJJ. Induction of the nodApromoter of Rhizobium leguminosarum Sym plasmidpRL1JI by plant flavanones and flavones. J Bacteriol1987;169:198–204.
Zaat SA, Wijffelman CA, Mulders IHM, van Brussel AAN,Lugtenberg BJJ. Root exudates of various host plantsof Rhizobium leguminosarum contain different sets ofinducers of Rhizobium nodulation genes. Plant Physiol1988;86:1298–303.
Zaat SA, Schripsema J, Wijffelman CA, van Brussel AAN,Lugtenberg BJJ. Analysis of the major inducers of theRhizobium nodA promoter from Vicia sativa rootexudate and their activity with defferent nodD genes.Plant Mol Biol 1989;13:175–88.
Zhang F, Smith DL. Preincubation of Bradyrhizobiumjaponicum with genistein accelerates nodule devel-opment of soybean at suboptimal root zone tempera-tures. Plant Physiol 1995;108:961–8.
Zhang F, Smith DL. Inoculation of soybean (Glycine max.(L.) Merr.) with genistein-preincubated Bradyrhizo-bium japonicum or genistein directly applied into soilincreases soybean protein and dry matter yield undershort season condition. Plant Soil 1996;179:233–41.
Zhang F, Smith DL. Interorganismal signaling in subopti-mum environments: the legume – rhizobia symbiosis.Adv Agron 2002;76:125–61.


















![Comparison between the reference [i]Rhizobium - Archive ...](https://static.fdokumen.com/doc/165x107/63134e0faca2b42b580d2177/comparison-between-the-reference-irhizobium-archive-.jpg)

