
reponse des cellules endotheliales a un flux hémodynamique ...
96
PATRICK CHARRON REPONSE DES CELLULES ENDOTHELIALES A UN FLUX HÉMODYNAMIQUE DANS UN SUBSTITUT VASCULAIRE CONSTRUIT PAR GÉNIE TISSULAIRE Mémoire présenté à la Faculté des études supérieures de l'Université Laval dans le cadre du programme de maîtrise en biologie cellulaire et moléculaire pour l'obtention du grade de Maître ès sciences (M. Se.) DÉPARTEMENT DE BIOLOGIE CELLULAIRE ET MOLÉCULAIRE FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL QUÉBEC 2008 © Patrick Charron, 2008
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of reponse des cellules endotheliales a un flux hémodynamique ...

PATRICK CHARRON
REPONSE DES CELLULES ENDOTHELIALES A UN FLUX HÉMODYNAMIQUE DANS UN SUBSTITUT
VASCULAIRE CONSTRUIT PAR GÉNIE TISSULAIRE
Mémoire présenté à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en biologie cellulaire et moléculaire pour l'obtention du grade de Maître ès sciences (M. Se.)
DÉPARTEMENT DE BIOLOGIE CELLULAIRE ET MOLÉCULAIRE FACULTÉ DE MÉDECINE
UNIVERSITÉ LAVAL QUÉBEC
2008
© Patrick Charron, 2008

Résumé L'environnement hémodynamique des cellules endothéliales est un stimulus capital
pour l'activation et le maintien de leurs fonctions. Les vaisseaux sanguins sont
constamment soumis aux forces hémodynamiques sous forme de tension et de forces de
cisaillement et ces forces font partie intégrante de l'environnement physiologique des
cellules endothéliales. La force de cisaillement présente à l'interface sang-paroi vasculaire
agit au niveau de l'endothélium et influence la biologie des cellules endothéliales. La
présence de ces forces est souvent négligée dans les modèles in vitro traditionnels d" étude
de l'endothélium. Une simple couche de cellules, en deux dimensions, dans un
environnement statique s'éloigne de l'environnement natif des cellules endothéliales. La
méthode d'autoassemblage développée dans notre laboratoire a permis de mettre au point
un modèle in vitro d'étude de l'endothélium humain dans des conditions plus
physiologiques. Ce modèle appelé matrice fibroblastique tubulaire est formé de matrice
extracellulaire endogène, tridimensionnelle et de forme tubulaire. Ces caractéristiques
permettent la culture de cellules endothéliales humaines sur une matrice endogène en plus
de les exposer à un flux hémodynamique physiologique. L'objectif de ces travaux était
d'évaluer la réponse au flux hémodynamique des cellules endothéliales cultivées dans ce
modèle. Après une période d'exposition de 4 heures au flux, les observations
microscopiques de l'endothélium démontrent des changements de morphologie des cellules
endothéliales qui se rapproche de la réalité in vivo. Nous avons observé un alignement et un
étirement des cellules parallèles à la direction du flux et un réarrangement du cytosquelette
par la formation de fibres de stress également parallèles à la direction du flux. De plus,
nous n'avons pas observé de décollement des cellules par le flux, indiquant l'affinité du
substrat de matrice extracellulaire. Nous avons analysé l'influence de l'exposition au flux
sur l'expression de 7 gènes impliqués dans la biologie endothéliale : PECAM, eNOS, ET-
B l , PAI-1, tPA, VECAD et vWF. Nous n'avons pas observé de différences significatives
des niveaux d'expression de ces gènes entre la condition statique et dynamique. Le modèle
de la matrice fibroblastique tubulaire possède un potentiel certain pour l'étude des cellules
endothéliales dans un environnement hémodynamique physiologique. Son utilisation
pourrait profiter à plusieurs domaines d'étude impliquant les cellules endothéliales.

Abstract The hemodynamic environment of endothelial cells (EC) is a major stimulus for their
functions and maintenance. Blood vessels are constantly exposed to hemodynamic forces in
form of tension and shear stress. Thèse forces are part of the natural envrionment of blood
vessels and EC. The shear stress présent at the blood-vessel wall interface acts directly on
the endothelium and modulate its fonctions at several levels. The hemodynamic
environment is oiten neglected with traditional in vitro models of the endothlium. A simple
layer of EC. in two dimensions, in a static environment is far away from the native
physiological conditions. The self-assembly method. developed in our laboratory. open the
door for a new in vitro model of the human endothelium which can be study under more
physiological conditions. This model is composed of fibroblastic three dimensional tubular
extracellular matrix (FTM). Its features allow us to culture human EC on an endogenous
matrix and to expose them to a physiological flow. In order to study the response of human
EC under shear stress in our FTM, the endothelialized FTMs were submitted to static and
flow conditions for 4 hours in an hemodynamic bench. Following flow experiments,
microscopic observations of the endothelium show changes of cell morphology which
approach in vivo reality. We observed an alignment and a stretching of the cells parallel to
the flow direction. We also observed a rearrangement of the cytoskeleton by actin stress
fibers formation parallel to flow direction. Moreover, we did not observe any cell removed
by the flow indicating the affinity for the extracellular matrix. We analyzed the influence of
hemodynamic flow on the expression of 7 gènes involved in EC biology: PECAM, eNOS,
ET-B1, PAI-1, tPA, VECAD and vWF. We did not observe any signifîcant différences
induced by the flow in the expression levels of thèse gènes. The FTM model has an
undeniable potential for EC studies under a physiological hemodynamic environment and
several fields involving EC could benefit from it.

Avant-Propos Ces travaux ont été réalisés au Laboratoire d'Organogénèse Expérimentale (LOEX)
de l'hôpital du Saint-Sacrement de Québec, sous la direction du Docteur François A.
Auger.
Au cours de ma maîtrise, la majorité de mon travail à porté sur la mise au point des
techniques d'utilisation de nos models in vitro. L'adaptation des techniques pour la biologie
moléculaire (RT-PCR, PCR et PCR quantitative), l'extraction de TARN, l'augmentation du
temps d'exposition au flux, les méthodes d'ensemencement des cellules endothéliales dans
la MFT, la validation du flux utilisé et le traitement des échantillons pour la microscopie ne
sont que quelques exemples. Quoique nous n'ayons obtenu que peu de résultats et qu'il
reste du travail à faire, j e considère avoir appris énormément lors de mon cheminement,
tant du point de vue scientifique que personnel. La planification d'expériences scientifiques
complexes m'a amené à développer mon autonomie, mes aptitudes à la recherche et au
travail d'équipe. Je suis reconnaissant de l'opportunité d'avoir évolué au sein d'un
laboratoire dynamique et visionnaire.
Je tiens à remercier tous les gens qui ont contribué de près ou de loin à mon projet de
maîtrise. Tout d'abord, Dr Auger pour m'avoir accueilli dans son laboratoire et de m'avoir
rappeler : « Patrick, si tu savais le nombre de fois qu'on m'a dit que ce que je faisais
n'allait pas fonctionner ! ». Ensuite, Dr Dan Lacroix, qui a toujours été là pour moi et sur
qui j 'a i toujours pu compter. Également, Cindy Perron avec ses doigts de fée et Todd
Galbraith avec ses mains de velours qui ont été d'une aide plus que précieuse pour toutes
mes manipulations. Dr Maud Vallée qui avec une patience et une gentillesse remarquable
m'a pratiquement tout appris sur l'ARN et l'ADN. Dr Caroline Basoni et Valérie Cattan,
deux postdocs en or. En plus d'être sympathiques et très agréables à côtoyer, elles m'ont
éclairé par leurs expériences et leurs connaissances. L'inoubliable Jean-François Oligny,
qui est à l'origine de tout mon savoir-faire avec le montage du banc hémodynamique et des
MFT. Dr Pierre-Luc Tremblay, qui a été pour moi un exemple à suivre et la source de
nombreux conseils. Marie-Christine Fiola, qui en plus d'être une merveilleuse « coloc » a
passé beaucoup de temps avec moi au confocal. Jean Dubé, qui lui aussi a été un exemple à
suivre et une ressource on ne peut plus polyvalente. L'incarnation de la gentillesse.

Kathleen Baker, pour ces efforts de transfert du savoir-faire et du maintien de Tordre dans
le monde des cellules congelées. Karl-Stéphane Rossignol, pour son aide en mécanique des
fluides. Tous les membres du groupe vasculaire et du LOEX qui forment une formidable
équipe. Finalement, Vicky Gagnon qui en plus de m'avoir aidé au confocal est devenue ma
plus grande source de motivation. A vous tous : MERCI !

/ only know îhat I was born and I know that 111 die.
The in between is mine. (Eddie Vedder)

Table des matières Résumé i Abstract ii Avant-Propos iii Table des matières vi Liste des tableaux viii Liste des figures viii Liste des abréviations x Introduction 11
1.1 Système vasculaire 11 1.2 Vaisseaux sanguins 11
1.2.1 Artères 13 1.2.2 Capillaires 14 1.2.3 Veines 15
1.3 Cellules endothéliales 16 1.3.3 Barrière hémato-encéphalique 17 1.3.4 Implication des cellules endothéliales dans les pathologies 20
1.4 Forces hémodynamiques et circulation sanguine 22 1.4.1 Pression transmurale 23 1.4.2 Force de cisaillement 24 1.4.3 Écoulement laminaire et turbulent 26
1.5 Forces hémodynamiques et biologie de l'endothélium 26 1.5.1 Mécanotransduction 28 1.5.2 Expression génique 31 1.5.3 Fonctions des cellules endothéliales 34
1.6 Modèles d'étude des cellules endothéliales 37 1.6.1 Substitut vasculaire construit par génie tissulaire comme modèle d'étude in vitro 38
1.7 Problématique et objectif 42 Matériels et Méthodes 44
2.1 Culture cellulaire 44 2.1.1 HUVEC 44 2.1.2 Fibroblastes 44 2.1.3 Cellules d'astrocytome 45
2.2 Matrice Fibroblastique Tubulaire (MFT) 46 2.3 Banc hémodynamique 47 2.4 Microscopie 49
2.4.1 Contraste de phase 49 2.4.2 Microscopie électronique à balayage 49 2.4.3 Immunofluorescence 50
2.5 Biologie Moléculaire 51 2.5.1 Extraction d'ARN total 51 2.5.2 RT-PCR 52 2.5.3 PCR 52 2.5.4 Séquençage 53

2.5.5 PCR quantitative 53 2.6 Culture en chambre de Boyden 54 2.7 Test de perméabilité transendothéliale 55
Résultats 57 3.1 Morphologie 57 3.2 Expression génique 64 3.3 Barrière hémato-encéphalique 70
Discussion 76 4.1 Réponses des cellules endothéliales au flux hémodynamique 76 4.2 Barrière hématoencéphalique 81
Conclusion 83 Annexe 1 85 Bibliographie 86

Liste des tableaux
Tableau 1. Liste des anticorps utilisés pour immunofluorescence 51
Tableau 2. Liste des amorces utilisées 53
Tableau 3. Résultats du séquençage des produits PCR purifiés 66
Tableau 4. Expression relative des gènes PECAM eNOS, ET-B1. PAI-1. tPA. VECAD
et vWF 66
Liste des figures
Figure 1. Schéma de la circulation sanguine systémique et des types de vaisseaux 12
Figure 2. Schéma de vaisseau sanguin composé des trois couches distinctes 13
Figure 3. Vue microscopique de capillaires sanguins 15
Figure 4. Schéma des forces de pression et de tension 24
Figure 5. Schéma de la force de cisaillement 25
Figure 6. Résumé schématique de la réponse des CE à la force de cisaillement laminaire..28
Figure 7. Étapes de la production d'un équivalent vasculaire humain reconstruit par génie
tissulaire selon la technique d'autoassemblage 39
Figure 8. Vaisseaux sanguin humain reconstruit par génie tissulaire selon la technique
d'autoassemblage 40
Figure 9. Étapes de la production d'une matrice fibroblastique tubulaire (MFT)
endothélialisée 42
Figure 10. Enroulement d'un feuillet de fibroblastes autour d'un mandrin tubulaire 46

Figure 11. Montage d'une MFT pour endothélisation 48
Figure 12. Montage du banc hémodynâmique relié aux pompes péristaltiques 49
Figure 13. Principe de l'immunofluorescence 51
Figure 14. Schéma d'une chambre de Boyden 56
Figure 15. Coupes transversales de MFT 57
Figure 16. Microscopie électronique à balayage d'une MFT sans cellule 59
Figure 17. Microscopie électronique à balayage d'une MFT endothélialisée en condition
statique 59
Figure 18. Microscopie électronique à balayage de MFT endothélialisées 60
Figure 19. Microscopie confocale du marquage immunohistochimique des fibres
d'actine 61
Figure 20. Microscopie confocale du marquage immunohistochimique de VECAD 62
Figure 21. Microscopie confocale du marquage immunohistochimique de PECAM 63
Figure 22. Gel d'agarose des produits de PCR pour différents gènes 65
Figure 23. Graphique de l'expression relative moyenne des gènes PECAM, eNOS,
ET-B1, PAI-1, tPA, VECAD et vWF 67
Figure 24 Graphique des données individuelles de l'expression relative des gènes
PECAM, eNOS, ET-B1, PAI-1, tPA, VECAD et vWF 68
Figure 25. Test de perméabilité transendothéliale au BSA-FITC en chambres de
Boyden 71
Figure 26. Marquage immunohistochimique pour GFAP 73
Figure 27. Marquage immunohistochimique de ZO-1 75

Liste des abréviations
ADNc: Acide Désoxyribonucléique complémentaire
ATCC : American Type Culture Collection
BHE : Barrière Hémato-Encéphalique
BSA: Bovine Sérum Albumin
CE : Cellule Endothéliale
CML : Cellule Musculaire Lisse
ECGS: Endothelial Cell Growth Supplément
eNOS : Endothelial Nitric Oxide Synthase
ET-B : Endothelin Type B Receptor
FC : Force de Cisaillement
FITC: Fluoresceine Iso Thio Cyanate
GFAP: Glial Fibrillary Acidic Protein
HUVEC : Human Umbilical Vein Endothelial Cell
LOEX: Laboratoire d'Organogénèse Expériementale
MEC : Matrice Extracellulaire
MFT: Matrice Fibroblastique Tubulaire
PAI-1 :Plasminogen Activator Inhibitor
PECAM : Patelet Endothelial Cell Adhésion Molécule
tPA : Tissue Plasminogen Activator
VECAD : Vascular Endothelial Cadherin (VE-Cadherin)
VEGF : Vascular Endothelial Growth Factor
vWF : Von Willebrand Factor
ZO-1 : Zona Occludens 1

Introduction 1.1 Système vasculaire
L'évolution des organismes unicellulaires vers des organismes pluricellulaires
nécessite d'approvisionner les cellules en nutriments et en oxygène en plus de les
débarrasser des déchets métaboliques. Pour permettre ces échanges entre les cellules et
l'environnement, le système vasculaire a évolué avec l'augmentation en volume des
organismes [1]. Chez les mammifères, le système cardio-vasculaire est tellement développé
que presque toutes les cellules du corps ont accès aux échanges avec le sang. Par exemple,
dans le muscle squelettique, la densité des capillaires sanguins est d'environs 500/mm 2[2.
3] et chez l'humain adulte, mis bout à bout, les vaisseaux sanguins couvrent une distance de
100 000 km [4]. Ceci démontre bien l'étendue et l'importance du système vasculaire. Cette
voie de distribution du sang est essentielle au maintien de l'homéostasie de l'organisme. En
plus de permettre le transport des éléments essentiels tels les gaz et les nutriments, le
système vasculaire est le support de transport des systèmes immunitaire et endocrinien. Il
joue des rôles centraux dans l'osmorégulation et la thermorégulation. Les vaisseaux
sanguins se divisent en trois principales catégories : les artères, les capillaires et les veines.
1.2 Vaisseaux sanguins
Chez les mammifères, la circulation sanguine systémique part du ventricule gauche
d'où le sang oxygéné passe part l'aorte pour ensuite se jeter dans les différentes artères.
Ensuite, le sang emprunte les artérioles de plus petit calibre pour aboutir au réseau
capillaire. C'est à cet endroit que les échanges entre le sang et le milieu interstitiel ont lieu.
Le sang désoxygéné retombe dans des veinules, puis dans les veines et revient finalement
au cœur par les veines caves.

Réseau capillaire
Artérioles Veinules
Figure 1. Schéma de la circulation sanguine systémique et des types de vaisseaux. © Patrick Charron
Les artères et les veines sont composées de trois couches concentriques. En partant du
centre, on retrouve le vide central où circule le sang, soit le lumen (lumière). Directement
en contact avec le sang se trouve l'endothélium (intima). L'endothélium est un épithélium
pavimenteux simple qui tapisse la lumière de tous les vaisseaux sanguins et du muscle
cardiaque (endocarde). Les cellules endothéliales (CE) de l'intima reposent sur une
membrane basale composée de matrice extracellulaire (MEC). La couche centrale, la
média, est formée par des anneaux concentriques de cellules musculaires lisses (CML)
incluses dans une MEC complexe de collagènes, d'élastine, de fibrine et de protéoglycanes
[5]. Cette couche est le principal siège de la régulation de la circulation sanguine par son
pouvoir de vasoconstriction et de vasodilatation. La dernière couche, l'adventice (externa),
est principalement composée de fibres de collagènes provenant de fibroblastes résidants et
d'élastine [5]. L'adventice protège les vaisseaux et les ancres aux tissus environnants.
Lorsque les vaisseaux présentent une épaisseur importante, la diffusion des gaz sanguins
n'est plus suffisante pour atteindre les cellules des couches externes. Dans cette situation,
on retrouve à travers l'adventice des petits vaisseaux, un réseau de capillaires, des
neurofibres et des vaisseaux lymphatiques. Ces petits vaisseaux qui nourrissent l'extérieur
des gros vaisseaux sont appelés vasa-vasorum. Cet agencement en trois couches distinctes
est commun aux artères et aux veines. Cependant, leur composition varie selon le calibre et
la fonction des vaisseaux [1, 4. 6. 7]. Il sera maintenant discuté des principales
caractéristiques des différents types de vaisseaux sanguins.
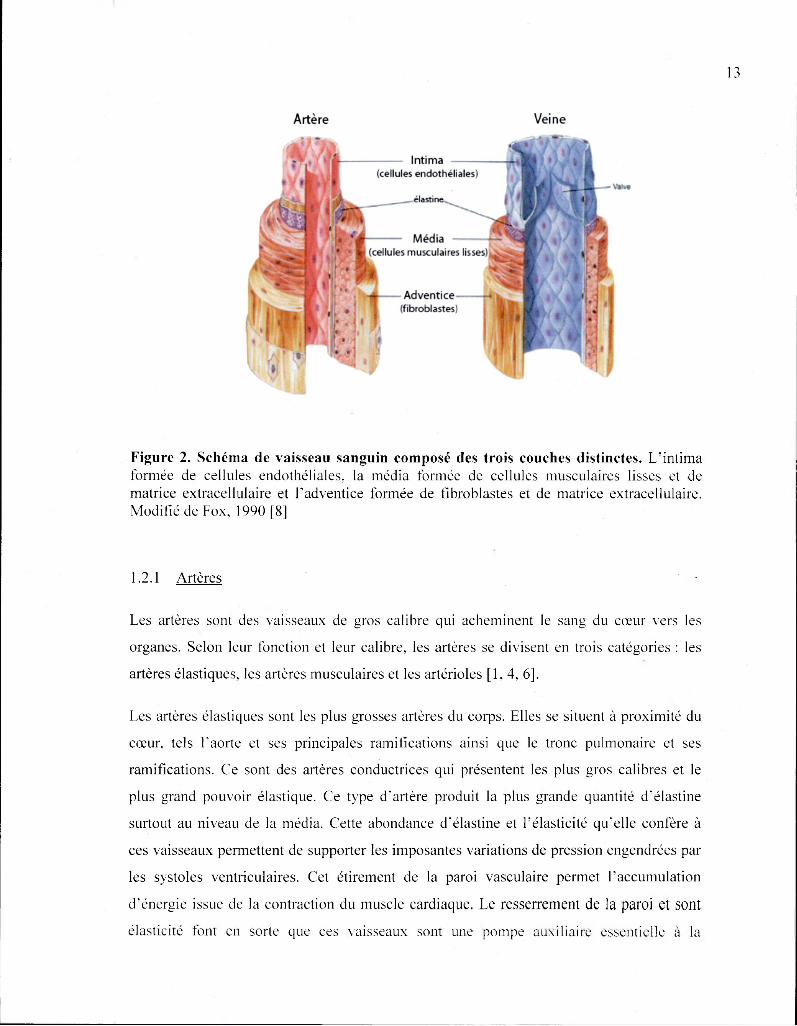
Figure 2. Schéma de vaisseau sanguin composé des trois couches distinctes. L'intima formée de cellules endothéliales, la média formée de cellules musculaires lisses et de matrice extracellulaire et l'adventice formée de fibroblastes et de matrice extracellulaire. Modifié de Fox, 1990 [8]
1.2.1 Artères
Les artères sont des vaisseaux de gros calibre qui acheminent le sang du cœur vers les
organes. Selon leur fonction et leur calibre, les artères se divisent en trois catégories : les
artères élastiques, les artères musculaires et les artérioles [1, 4, 6].
Les artères élastiques sont les plus grosses artères du corps. Elles se situent à proximité du
cœur, tels l'aorte et ses principales ramifications ainsi que le tronc pulmonaire et ses
ramifications. Ce sont des artères conductrices qui présentent les plus gros calibres et le
plus grand pouvoir élastique. Ce type d'artère produit la plus grande quantité d'élastine
surtout au niveau de la média. Cette abondance d'élastine et l'élasticité qu'elle confère à
ces vaisseaux permettent de supporter les imposantes variations de pression engendrées par
les systoles ventriculaires. Cet étirement de la paroi vasculaire permet l'accumulation
d'énergie issue de la contraction du muscle cardiaque. Le resserrement de la paroi et sont
élasticité font en sorte que ces vaisseaux sont une pompe auxiliaire essentielle à la

circulation sanguine [6]. Elles offrent donc la circulation du sang avec un minimum de
résistance.
Les artères musculaires tiennent leur nom de leur média qui est la plus épaisse de tous les
vaisseaux. Leur média est pourvue d'un grand nombre de CML compris dans une MEC
moins riche en élastine par rapport aux artères élastiques. Ainsi, les artères musculaires ont
une capacité de vasoconstriction plus importante. Elles jouent un rôle de distribution du
sang aux divers organes du corps. Leur diamètre varie de moins de 1 mm jusqu'aux
environs de 1 cm [4].
Les artérioles sont les plus petites artères puisque leur diamètre est inférieur à 0,3 mm [4].
Les plus grosses artérioles possèdent les trois différentes couches vasculaires. mais leur
média est constituée principalement de cellules musculaires lisses et de très peu d'élastine.
Les plus petites ne possèdent qu'une seule couche de cellules musculaires lisses autour de
l'intima. La circulation sanguine dans les lits capillaires est régulée principalement par la
vasoconstriction des artérioles.
1.2.2 Capillaires
Les capillaires sont les plus petits vaisseaux sanguins de l'organisme. Ils sont constitués
seulement de cellules endothéliales et de leur membrane basale. Les capillaires mesurent en
moyenne 1 mm et leur calibre moyen n'est que de 8 à 10 um [6, 9]. Ils sont le siège des
échanges entre le sang et le milieu interstitiel. Mis à part quelques exceptions, on les
retrouve dans toutes les parties du corps [4]. Il existe deux types de capillaires : les
capillaires continus et fenestrés. Les continus sont les plus communs et répandus. Ils sont
appelés continus du fait que leur endothelium recouvre entièrement la lumière du capillaire
et que les cellules endothéliales adjacentes sont jointes par des jonctions adhérentes et
serrées [4, 9]. Cependant, on y retrouve aussi des fentes intercellulaires permettant les
échanges de molécules et de liquides entre le sang et le milieu interstitiel. La présence des
ces fentes est absente au niveau de la barrière hémato-encéphalique qui sera discutée plus
loin. Pour leur part, les capillaires fenestrés présentent certaines cellules endothéliales

munies de pores appelés fenestrations. La présence de ces fenestrations augmente
grandement la perméabilité de l'endothélium. On retrouve donc ce type de capillaires dans
les organes où les échanges sont importants tels que l'intestin grêle, le foie, les glandes
endocrines et les reins [4, 10].
Figure 3 Vue microscopique de capillaires sanguins. A) Photographie d'un capillaire par microscopie électronique à transmission. Le capillaire est formé d'une seule cellule endothéliale entourée de sa membrane basale. B) Photographie d'un capillaire par microscopie électronique à balayage. Modifié de Alberts et al 2002 [9].
1.2.3 Veines
En poursuivant l'itinéraire du sang, l'union des capillaires forme les veinules. Le diamètre
de ces dernières varie entre 8 et 100 u,m. Les plus petites veinules sont composées
simplement d'un endothelium entouré de quelques fibroblastes alors que les plus grosses
présentent les trois couches distinctes de cellules vasculaires. La perméabilité des veinules
est très réactive, c'est pourquoi les cellules de l'immunité utilisent souvent ce type de
vaisseau pour sortir de la circulation [11]. Des veinules, le sang rejoint ensuite les veines.
Tout comme les artères, la première mission des veines est le transport du sang, mais dans
la direction inverse, des organes vers le cœur. Les vaisseaux du système veineux sont
caractérisés par leur grand diamètre interne par rapport aux artères correspondantes. Les
parois des veines sont plus fines et contiennent moins de CML. Elles sont moins élastiques
comparativement aux artères puisqu'elles contiennent plus de fibres de collagènes que de
'Cellule tndotheliak
s Lumen

fibres élastiques. Pour permettre le retour veineux malgré la pression sanguine plus faible
dans les veines, on y retrouve des valvules. Celles-ci empêchent le reflux sanguin dans la
mauvaise direction. On les retrouve majoritairement dans les veines des membres inférieurs
de façon à contrer la force de gravité qui s'exerce sur le sang. Les valvules collaborent à
contrer l'intolérance orthostatique qui consiste à une incapacité à maintenir une posture
debout provoquée par l'accumulation de sang dans les membres inférieurs par la force de
gravité. Cette accumulation peut même aller jusqu'à provoquer la perte de conscience. Ces
caractéristiques des veines permettent au réseau veineux de jouer le rôle de réservoir
sanguin. En effet, chez les mammifères, plus de 50 % du volume total de sang est contenu
dans les veines [6]. Lorsqu'une hémorragie survient, le volume veineux diminue, mais pas
le volume artériel, ce qui contribue à maintenir à un niveau suffisant la pression artérielle et
le débit au niveau des lits capillaires. Cette diminution de volume engendrée par la
vasoconstriction des CML compense la perte de volume sanguin.
1.3 Cellules endothéliales
Maintenant que l'anatomie du système sanguin a été brièvement décrite, il sera
question du type cellulaire central de cet ouvrage : la cellule endothéliale. Les CE dérivent
du mésoderme embryonnaire [9, 12] et recouvrent le système vasculaire en entier. Chez
l'humain, le volume total des CE est comparable à celui du foie [7]. L'endothélium est bien
plus qu'une simple couche de cellules qui tapisse le lumen du système vasculaire. Il
contrôle le passage des substances de part et d'autre de la circulation sanguine et du milieu
interstitiel. Les CE jouent un rôle central dans le développement, le maintien et le
remodelage de la vasculature [13]. Elles participent à la régulation de la circulation
sanguine et au maintien de son homéostasie par la production de plusieurs facteurs et
l'interaction avec les CML [14]. Par exemple, l'expression d'angiotensine II [15, 16],
d'oxyde d'azote (NO) [17], d'endothéline [18] et d'autres prostaglandines affecte le
maintien de la pression sanguine. L'endothélium est également impliqué dans la
conservation de la viscosité normale du sang et de la coagulation, entre autres, par
l'expression des enzymes plasminogen activator inhibitor-1 (PAI-1) [19] et de tissue
plasminogen activator (tPA) [20]. Les CE sont également impliquées dans des phénomènes

inflammatoires et immunitaires [11, 21]. Elles participent au contrôle de l'extravasion des
cellules immunitaires par l'expression de molécules d'adhérence comme les sélectines. les
intégrines, des immunoglobulines ainsi que par la production de chimiokines.
Les CE ont une remarquable capacité à s'ajuster aux conditions environnantes. Bien qu'on
les retrouve partout dans l'organisme et qu'elles possèdent des caractéristiques communes,
elles diffèrent selon leurs emplacements, le type de vaisseau et leurs rôles. Cette plasticité
notable des CE entraîne la spécialisation locale du système vasculaire qui doit répondre aux
besoins des différents tissus et organes nécessitant une grande variété de flux et de pression.
Les CE forment donc une population cellulaire hétérogène [10. 22-24]. Elles expriment
différents gènes selon le vaisseau dans lequel elles se trouvent et l'état environnant. Ainsi,
les CE des artères diffèrent des CE des veines et des capillaires. De plus, l'endothélium des
capillaires est spécialisé et diffère selon son origine. Par exemple, les CE micros
vasculaires des reins, du foie ou du cerveau arborent des caractéristiques différentes.
1.3.3 Barrière hémato-encéphalique
Un exemple frappant de cette spécialisation des CE se retrouve au niveau de la micro-
vasculature du système nerveux central. En effet, l'endothélium du cerveau forme une
barrière sélective entre la circulation sanguine et le milieu interstitiel cérébral. Ce
phénotype particulier de l'endothélium est appelé barrière hémato-encéphalique (BHE) [4,
6].
Cette barrière entre la circulation systémique et le milieu interstitiel nerveux est un
phénomène très conservé au cours de l'évolution. Dès l'apparition d'une céphalisation, on
retrouve en parallèle l'évolution d'une séparation des environnements nerveux et
systémiques [6, 25]. Les mécanismes précis qui permettent cette séparation ont divergé au
cours de l'évolution. Néanmoins, la protection du contenu du milieu interstitiel cérébral
demeure universelle chez les animaux dotés d'une céphalisation [26-29]. Les mécanismes à
la base des fonctions nerveuses nécessitent un environnement dont le contenu est stable et
contrôlé. Les neurones sont pourvus de propriétés particulières qui leur permettent de

recevoir et de transmettre de l'information aux autres cellules. D'une part, la polarisation de
la membrane cellulaire via le transport d'ions permet la formation de potentiel d'action. La
composition et la concentration ionique du milieu interstitiel cérébral doivent donc être
stables et propices aux fonctions nerveuses. D'autre part, la communication neuronale par
synapses chimiques utilise des neurotransmetteurs. Ces molécules qui jouent le rôle de
neurotransmetteur dans le système nerveux central se retrouvent également en périphérie et
dans la circulation sanguine où elles accomplissent d'autres fonctions. Le contrôle de la
circulation de ces molécules entre le sang et le milieu interstitiel cérébral devient donc
essentiel pour ne pas interférer avec les fonctions neuronales. Ainsi, la BHE protège le
cerveau des fluctuations ioniques et des substances neuroactives qu'on retrouve dans le
sang [29]. La BHE protège également le cerveau de certaines substances neurotoxiques. En
outre, elle confère le statut de site immunologique privilégié au cerveau en empêchant le
passage des lymphocytes naïfs [11]. Malgré la restriction des échanges entre le sang et le
milieu cérébral, l'endothélium de la BHE transporte les nutriments essentiels et les déchets
métaboliques de part et d'autre de la circulation sanguine.
La BHE se situe directement au niveau des CE des micros vaisseaux cérébraux. On peut la
subdiviser en trois types de barrière : physique, de transport et enzymatique [25, 28].
L'endothélium de la BHE agit en barrière physique grâce à la présence de jonctions serrées
entre les CE adjacentes. Ces jonctions forcent le trafic des molécules par la voie
transcellulaire (au travers les membranes cellulaires) plutôt que par la voie paracellulaire
(entre les CE) comme dans la plupart des endothéliums. Les jonctions serrées de
l'endothélium du cerveau sont plus complexes et réduisent plus efficacement le passage
paracellulaire par rapport aux endothéliums de la périphérie [30-32]. Elles réduisent même
le passage de très petites molécules comme les ions sodium et chlore. Parmi les molécules
qui contribuent de façon importante aux jonctions serrées de la BHE, on retrouve les
protéines transmembranaires de la famille des occludines (3, 5 et 12) et des claudines. Ces
protéines sont souvent liées à des protéines adaptatrices zona occludens (ZO-1, 2 et 3) qui
les relient au cytosquelette [33]. On retrouve également chez les CE de la BHE des
molécules de jonctions adhérentes (JAM-A, B et C) qui sont impliquées dans la formation
et le maintient des jonctions serrées [32]. La partie cytoplasmique de ces protéines
transmembranaires interagit avec une multitude de protéines à la périphérie de la membrane

formant de larges complexes protéiques [34]. Ces complexes interagissent avec différentes
protéines kinases, ce qui engendre l'activation de voies de transduction et de facteurs de
transcription. Les jonctions serrées de la BHE sont donc actives et modulables. La
réduction du passage paracellulaire n'est pas Tunique fonction des jonctions serrées, elles
permettent également une polarisation apicale-basale des CE. Cette propriété est
indispensable pour la barrière de transport de la BHE.
Un système de transport spécifique sur la surface luminale et abluminale régule le trafique
transcellulaire des petites molécules hydrophiles, créant ainsi une barrière de transport
sélective. Ceci permet l'entrée des nutriments requis et l'exclusion des composés
potentiellement dommageables. Les grosses molécules hydrophiles comme les peptides et
les protéines sont généralement exclues sauf lorsqu'elles sont transportées par transcytose
spécifique ou d'absorption. Toutefois, ce type de transport est beaucoup plus rare au niveau
des CE de la BHE [25]. Les CE cérébrales expriment plusieurs transporteurs
transmembranaires. Par exemple, le transporteur du glucose GLUT-1 [35], plusieurs
transporteurs d'acides aminés comme LAT-1 [36], des transporteurs de nucléosides et bien
d'autres. On retrouve également des transporteurs de la famille ABC (ATP Binding
Cassette) [37] qui agissent contre un gradient de concentration, des transporteurs d'efflux
comme P-glycoprotéine [38] et des protéines de la famille MRP (Multidrug Resistance-
related Protein). Finalement, on retrouve aussi des transporteurs dépendant du sodium
(Na +).
Une combinaison d'enzymes intra- et extra-cellulaires forme la barrière métabolique [39].
Par exemple, des peptidases et des nucléotidases sont en mesure de métaboliser des
peptides extracellulaires alors que des enzymes intracellulaires comme la monoamine-
oxydase et le cytochrome P450 peuvent inactiver plusieurs composés neurotoxiques ou
neuroactifs.
Qu'est-ce qui cause cette spécialisation aussi évoluée des vaisseaux sanguins du cerveau ?
Depuis longtemps, les études histologiques ont démontré l'association et l'enroulement très
serré des capillaires avec d'autres types cellulaires incluant des astrocytes, des péricytes,
des cellules microgliales et neuronales [40]. Il y a maintenant beaucoup d'études qui
démontrent que les astrocytes ont la capacité de réguler les caractéristiques de la BHE chez

les CE [41], Des preuves expérimentales soutiennent que la présence des astrocytes est
nécessaire à l'apparition du phénotype de barrière physique [42]. de transport [38] et
métabolique [43] chez les CE. Étant donné l'énorme complexité de la BHE. il n'est pas
surprenant que d'autres types cellulaires jouent un rôle dans son induction. Malgré que les
astrocytes demeurent les principaux inducteurs du phénotype de la BHE, des travaux ont
souligné les contributions d'autres types cellulaires comme les péricytes [44]. les
macrophages périvasculaires [45] et les neurones [46].
Étant donné son implication dans plusieurs pathologies [28. 47-52]. la biologie de la BHE
fait l'objet d'un nombre impressionnant d'études. L'imposante complexité de la BHE
limite les conclusions qu'il est possible de tirer des études in vivo. Dans un contexte in vivo,
il est impossible de contrôler tous facteurs en mesure d'influencer les caractéristiques de la
BHE. A l'opposé, les conditions de culture cellulaire classiques in vitro sont trop loin des
conditions natives. Le développement de modèles in vitro de BHE arborant des
caractéristiques plus physiologiques serait un outil précieux pour ce domaine d'étude [25.
26, 28, 41 , 42, 49, 53, 54]. Il sera question de l'élaboration d'un tel modèle plus loin dans
ce mémoire.
1.3.4 Implication des cellules endothéliales dans les pathologies
Les implications des CE dans la physiopathologie de maladies connues sont multiples. Il
serait impossible d'en dresser une liste exhaustive dans ce travail. En effet, comme les CE
sont présentes dans tous les tissus de l'organisme, leur implication dans le développement
de conditions pathologies n'est pas surprenante. Dans le cadre de cet ouvrage, nous allons
nous concentrer sur seulement deux exemples : l'athérosclérose et le cancer.
L'athérosclérose est une réponse inflammatoire chronique de la paroi vasculaire des artères
et artérioles due principalement à l'accumulation de lipoprotéines telles que les
triglycérides et les LDL (Low Density Lipoprotein) [55]. Cette situation mène à la
formation de multiples plaques athéromateuses formées de LDL oxydées, de cholestérol et
de macrophages, à l'intérieur des parois vasculaires artériels. L'accumulation de plaques

athéromateuses, bien que compensée par un remodelage vasculaire, mène éventuellement à
une sténose artérielle provoquant à son tour un apport sanguin insuffisant aux organes
touchés. De plus, la rupture de plaques provoque la formation de thrombus qui est souvent
à l'origine d'infarctus d'artères coronaires. Un autre effet adverse possible de
l'accumulation de ces plaques est élargissement compensatoire excessif de la paroi
vasculaire menant à un anévrisme. Les CE sont au centre de la physiopathologie de
l'athérosclérose [56]. Dans un premier temps, les CE sont directement en contact avec les
lipoprotéines circulantes et sont le siège de leur absorption. Lorsque les LDL s'accumulent
dans les CE, elles sont oxydées par de complexes réactions biochimiques à l'intérieur des
CE, principalement dues à la présence de radicaux libres. Il en résulte des dommages à
l'endothélium ce qui mène à une réaction inflammatoire des CE [57, 58]. Ces dernières
expriment des facteurs inflammatoires et des cytokines qui recrutent des monocytes
circulant via l'expression de molécules d'adhésion comme VCAM-1 (Vascular Cell
Adhésion Molécule 1). Ces monocytes sortent de la circulation sanguine et se différencient
en macrophages. À la suite de cette extravasation, les macrophages phagocytent les LDL
oxydées et deviennent de grosses cellules remplies de lipides. Ces cellules finissent par
mourir et amplifient la réaction inflammatoire. Malgré toute la recherche consacrée à ce
sujet, les mécanismes moléculaires et cellulaires à l'origine de ce phénomène sont encore
mal connus. Des travaux ont démontré que les localisations susceptibles de développer des
plaques d'athérome coïncident avec les endroits où le flux sanguin présente des
caractéristiques hémodynamiques perturbées ou turbulentes [59-61]. Les forces
hémodynamiques de la circulation sanguine seront discutées plus loin. En somme, les CE
sont directement impliquées dans la physiopathologie de l'athérosclérose.
L'apparition de cancer nécessite inévitablement l'implication des CE. En effet, la
croissance d'une tumeur est limitée par l'augmentation de sa taille qui atteint un point où la
diffusion des gaz et des nutriments est insuffisante. Pour que la croissance tumorale se
poursuive, il doit y avoir un processus d'angiogenèse tumorale pour contrer le manque
d'apport en oxygène et nutriments. Ce phénomène d'angiogenèse est provoqué par
l'expression de facteurs angiogéniques par les cellules cancéreuses tels que VEGF
(Vascular Endothelial Growth Factor) et bFGF (basic Fibroblast Growth Factor) [62]. Ces
facteurs agissent sur les CE environnantes qui sont à l'origine de la néo-vasculature

tumorale. L'angiogenèse tumorale fait l'objet de nombreuses études puisque l'arrêt de
celle-ci pourrait être un traitement efficace contre le cancer [63]. Un autre niveau
d'implication des CE dans le cancer est l'émergence des métastases. Le processus
métastatique consiste au détachement de cellules cancéreuses de la tumeur primaire,
l'entrée dans la circulation sanguine, l'extravasation de ces cellules dans un site secondaire,
la prolifération dans ce nouveau site et l'induction de néo-vascularisation. Ce processus
implique donc des interactions entre l'endothélium et les cellules cancéreuses circulantes
[64]. Pour permettre l'extravasation des cellules cancéreuses, elles doivent être en mesure
d'adhérer aux CE. Ceci nécessite l'expression de molécules d'adhésion par les CE [65]
telles que e-sélectine, VCAM-1 et ICAM-1 (Intercellular Adhésion Molécule). Ce
phénomène complexe d'interactions entre les cellules cancéreuses circulantes et
l'endothélium fait aussi l'objet de plusieurs études [66-68].
En somme, l'étude de la biologie des CE est capitale pour la compréhension de pathologies
variées. Nous en décrivons brièvement que deux exemples, mais les champs d'intérêt sont
nombreux. Nous pourrions citer les domaines de la sclérose en plaque [48], de
l'hypertension [14], du diabète [69, 70] et bien d'autres. Tous ces domaines d'études
profitent de différentes expériences in vitro impliquant des CE et bénéficieraient de
modèles d'études in vitro physiologiques.
1.4 Forces hémodynamiques et circulation sanguine
La circulation sanguine engendrée par les contractions du cœur implique la présence
de forces hémodynamiques dans l'ensemble de l'appareil circulatoire. Ces forces
mécaniques issues du mouvement du sang se divisent en deux types : des forces
circonférentielles et tangentielles à la paroi vasculaire, engendrées par la pression sanguine
et des forces longitudinales à la paroi, agissant à l'interface sang-endothélium et issues de
l'écoulement sanguin [6, 7, 14, 23, 71, 72].

1.4.1 Pression transmurale
L'action de pompage du cœur provoque l'écoulement du sang grâce à un gradient de
pression systémique. La contraction des ventricules tente de comprimer le sang qui est
incompressible comme tout liquide. Le sang est alors expulsé des ventricules en suivant un
gradient de pression engendré par les contractions cardiaques. Le sang se déplace donc
d'une zone de haute pression vers une zone de basse pression. Puisque le système
vasculaire est un circuit de conduits fermés, le sang y circule sous pression. Lorsqu'il est
question de pression sanguine, on réfère généralement à la pression transmurale, c'est-à-
dire la différence entre la pression externe et interne qui s'exerce de part et d'autre de la
paroi du vaisseau. La pression transmurale est exprimée en millimètres de mercure (mm
Hg) et la pression systolique (pression maximale au moment de la systole ventriculaire) et
diastolique (pression minimale lors de la diastole) sont donnée dans l'ordre. Par exemple,
chez un humain adulte au repos la pression normale se situe aux environs de 120/80 mm
Hg. Ceci provoque inévitablement des forces mécaniques de pression et de tension au
niveau des parois des vaisseaux. La pression s'exerce perpendiculairement à la paroi du
vaisseau alors que les forces de tension sont parallèles et en circonférence par rapport à la
paroi. L'action de la pression provoque l'étirement et l'augmentation du diamètre des
vaisseaux. Pour équilibrer ces forces et ne pas aboutir à un éclatement de la paroi, les
vaisseaux sont pourvus d'une certaine élasticité et la contraction des CML de la média
empêche la dilatation excessive de la paroi. La pression et les tensions sont donc
compensées par les propriétés de la média et de l'adventice des vaisseaux et c'est
principalement à ces niveaux qu'elles agissent.

Jens
Intima
Membrane Menbrane élastique élastique interne externe
Média Adventice
Figure 4. Schéma des forces de pression et de tension. La pression transmurale est tangentielle à la paroi alors que les forces de tension sont parallèles et circonférentielles. Ces forces agissent surtout au niveau de la média et de l'adventice (en rouge). L'intima est formé de CE. La média est composé de CML et de MEC. L'adventice est composée de fibroblastes et de MEC. © Patrick Charron. LOEX
1.4.2 Force de cisaillement
L'écoulement sanguin engendre aussi des forces parallèles à la paroi vasculaire au niveau
de l'interface sang-endothélium. Cette force produite par la friction issue du déplacement
sanguin est appelée force de cisaillement (FC) ou plus communément dans la littérature,
shear stress.

Intima
Membrane élastique interne
Menbrane élastique externe
Média Adventice
Figure 5. Schéma de la force de cisaillement. L'écoulement laminaire du sang induit des force de cisaillement à l'interface sang-paroi vasculaire. Ces forces sont appliquées directement au niveau de l'endothélium. A gauche, les flèches parralèles représentent le profil d'écoulement laminaire. À droite, la flèche de la force de cisaillement à l'interface sang-endothélium. © Patrick Charron, LOEX
Cette force a pour effet d'entraîner les CE dans la même direction que l'écoulement
sanguin. Les CE doivent donc s'adapter à la présence de cette force pour demeurer en
situation d'équilibre, il sera question plus loin de l'effet des FC sur l'endothélium.
Les valeurs théoriques de la FC sont basées sur la loi de Poiseuille qui décrit l'écoulement
laminaire dans les tubes [73] :
T = 4r| q
nx3
Où T - Force mécanique de cisaillement (dynes/cm2)
n, = Viscosité du milieu (Poises)
q = Flux (cm*s _ 1)
r = Rayon du lumen (cm)

Les variables qui influencent l'importance de la FC sont donc la viscosité du sang, la
vélocité du flux et le diamètre du vaisseau. Étant à la puissance 3, cette dernière variable est
très importante puisque, pour un flux donné, une légère diminution du diamètre vasculaire
entraîne une importante augmentation de la FC. La viscosité du sang des mammifères se
situe entre 3 et 4 [6] par rapport à l'eau qui est de 1, ce qui veut dire que le sang est de 3 à 4
fois plus visqueux que Leau. Le facteur qui influence le plus la viscosité du sang est
rhématocrite. Chez l'homme, en condition physiologique normale, la valeur moyenne de la
FC dans les artères est environ de 10 à 15 dynes/cm2 [74-76] et cette valeur semble
constante le long du circuit artériel [77-79] excepté aux endroits où le flux n'est pas
laminaire. Compte tenu de la nature puisée du flux, on parle de valeur moyenne puisque la
valeur de la FC varie en fonction du cycle cardiaque, elle atteint un maximum à la systole et
un minimum à la diastole.
1.4.3 Écoulement laminaire et turbulent
L'écoulement sanguin normal s'effectue par le déplacement parallèle de lames de liquide
concentriques par rapport à Taxe du vaisseau. La vitesse d'écoulement est minimale au
niveau de la paroi et maximale au centre du vaisseau. Ce profil parabolique des vitesses
d'écoulement est appelé laminaire. Lorsque les déplacements des différentes parties du
liquide ne sont pas parallèles à l'axe, on réfère à un écoulement turbulent. L'écoulement
laminaire pulsatile est caractéristique de la circulation périphérique [6, 7, 72]. L'écoulement
turbulent y est rare bien que possible dans certaine situation comme lorsque la vitesse est
très élevée et au niveau des ramifications où des boucles tourbillonnaires peuvent se former
[6, 7 , 7 1 , 7 2 ] .
1.5 Forces hémodynamiques et biologie de l'endothélium
Les vaisseaux sanguins sont constamment soumis aux forces hémodynamiques sous
forme de tension et de forces de cisaillement. Toute variation de ces contraintes mécaniques
est perçue au niveau des cellules vasculaires et se traduit par des modifications

fonctionnelles et structurales [14, 23, 71. 80, 81], La pression sanguine, qui est à l'origine
des forces de tension, affecte tous les types cellulaires du vaisseau. Parallèlement, les FC
agissent à l'interface sang-paroi vasculaire et affectent principalement l'endothélium [23].
Bien que la pression soit un important stimulus des vaisseaux, ce travail se concentre sur
les effets des FC sur F endothelium.
Tel que mentionné précédemment, la contrainte de cisaillement soumise à l'endothélium
artériel est relativement constante aux environs de 10-15 dynes/cm2. Cette constance est
assurée par des mécanismes humoraux autocrines et paracrines qui permettent au système
vasculaire de s'ajuster aux modifications hémodynamiques [6. 7. 15. 76]. Les changements
locaux des forces hémodynamiques induisent des transformations de la paroi du vaisseau
de manière à s'ajuster aux nouvelles conditions et de rétablir un climat physiologique de
tension et de FC. Par exemple, lorsque le flux augmente, les CE produisent de l'oxyde
d'azote (NO) [17, 82]. Ceci induit le relâchement des CML de la média et du même coup
l'augmentation du diamètre du vaisseau, rétablissant du même coup les valeurs des forces
hémodynamiques. Toutefois, lorsque les variations du tonus vasomoteur sont insuffisantes
pour compenser les modifications des contraintes mécaniques, les vaisseaux s'adaptent à
leur nouvel environnement hémodynamique en ajustant leur calibre et la structure de leur
paroi pour équilibrer les FC imposées à la surface endothéliale [23, 80, 83-85]. La présence
des FC à la surface endothéliale est un stimulus primordial pour les CE. Ces forces agissent
sur toutes les facettes de la biologie cellulaire des CE [80, 83, 86, 87]. La réponse des CE
aux FC est complexe et implique une multitude de phénomènes cellulaires et moléculaires.

Force de cisaillement
Intégrines
Canaux ioniques
Protéines G
Récepteurs membranaires
M e m b r a n e lipidique
G l y c o c a l i x
Transduction : F A K N O Akt
M A P K
Facteurs de trancription
Expression génique
Propriétés mécaniques
Alignement
Perméabilité
Migration
Prolifération
Apoptose
Figure 6. Résumé schématique de la réponse des CE à la force de cisaillement laminaire. Des mécanismes moléculaires sensibles au flux activent des voies de transduction ce qui mènent à la modulation de l'expression génique. La FC est un stimulus complexe qui module plusieurs facettes de l'activité des CE. © Patrick Charron, LOEX.
1.5.1 Mécanotransduction
Pratiquement tous les organismes vivants sont sensibles aux forces mécaniques. Mais
comment une force mécanique provoque-t-elle une réponse biologique ? Les cellules sont
munies de système mécanosensibles permettant de transformer une force physique en
information biochimique. Ce phénomène appelé mécanotranduction est essentiel au
développement des organes et à plusieurs processus physiologiques [88]. Outre le système
vasculaire, la mécanotransduction est impliquée dans la modulation de processus
physiologiques des systèmes pulmonaire, otique, osseux, tégumentaire, sensitif et est
nécessaire au contrôle du cycle cellulaire [89, 90]. Généralement, la mécanotransduction est
occasionnée par des changements de conformation de protéines qui activent ensuite
différents sentiers de transduction de signaux intracellulaires.

Plusieurs mécanismes de mécanotransduction impliqués dans la réponse des CE aux FC
sont suggérés dans la littérature [23, 80, 83, 86, 88, 91-93]. Dans le cadre de ce mémoire,
seulement une brève description des plus communes sera présentée.
Intégrines :
Les intégrines sont une famille de plus de 20 hétérodimères transmembranaires composés
d'une sous unité a et p [9. 94]. Le domaine extracellulaire des intégrines se lie à des
domaines spécifiques des composants de la MEC tels que la fibronectine. la vitronectine et
le collagène. Le domaine intracellulaire des intégrines interagit avec des molécules de
signalisation aux sites d'adhésions focaux. Par exemple, les kinases FAK (Focal Adhésion
Kinase), c-Src et des protéines du cytosquelette telles que a-actinine et taline. Lorsque les
CE sont exposées aux FC, les intégrines sont activées et cette activation est très importante
pour les fonctions endothéliales [81, 95]. De plus, la présence et la nature du ligand des
intégrines (MEC) joue aussi un important rôle dans la réponse des CE aux FC [96].
Canaux ioniques :
Des travaux ont démontré que les FC entraînent une hyperpolarisation des CE en culture
via l'activation des canaux potassiques [97]. Ceci entraîne une augmentation de l'entrée du
Ca 2 + ce qui à son tour régule plusieurs voies de signalisation et des fonctions des CE [98].
Bien que les mécanismes précis par lesquels les FC provoquent l'ouverture de ces canaux
ne sont pas connus, il ait tout de même établi que les canaux ioniques jouent un rôle
significatif dans la conversion des FC en signaux biochimiques.
Protéines G :
Plusieurs études ont démontré l'activation directe de certaines protéines G par les FC chez
les CE en plus de certains récepteurs couplés aux protéines G [99]. Cette activation
déclenche plusieurs autres voies de transduction en aval.

Récepteur de type tyrosinc kinase
Lors de l'exposition au flux des CE, le récepteur de type tyrosine kinase (RTK) au VEGF.
Flk-1, est phosphorylé et s'associe avec la protéine adaptatrice Shc et certaines intégrines
[100]. Ceci semble être inhérent à l'activation de la voie des ERK (Extracellular Regulated
Kinases) et JNK (c-Jun N-terminal Kinases) ainsi que du PI3K (Phosphoinositide 3-
Kinase).
Membrane lipidique :
Lorsque les CE sont exposées au flux, la FC a pour effet d'augmenter la fluidité de la
double couche de phospholipides de la membrane cellulaire [101]. Cette modulation de la
fluidité de la bicouche lipidique semble être un autre moyen de ressentir les FC puisqu'une
augmentation de la fluidité est couplée à l'activation des MAPK [102].
Glycocalix :
Les CE sont recouvertes du côté luminal par un mince réseau de protéoglycanes. de
glycosaminoglycanes (GAG) et de protéines plasmatiques [103]. Ce réseau, appelé
glycocalix, semble jouer un rôle fondamental dans la mécanotransduction des FC [91. 104].
puisque lorsque le glycocalix est altéré, la réponse des CE aux FC est déficiente.
Bien qu'il s'agisse des mécanismes les plus communs de mécanotransduction des FC chez
les CE, d'autres mécanismes sont aussi suspectés de jouer un rôle, notamment,
l'implication de PECAM (Patelet Endothelial Cell Adhésion Molécule) [105, 106] et des
radicaux libres oxygénés (ROS) [107]. Cela dit, il est évident qu'il existe plusieurs
mécanismes moléculaires permettant aux CE de ressentir les forces hémodynamiques. Ces
mécanismes interagissent entre eux et permettent une réponse très finement régulée des CE
aux FC. Les études portant sur les CE en relation avec les FC ont souligné que les
mécanismes de mécanotransduction sont en mesure d'activer plusieurs voies de
signalisation intracellulaires, entre autres les voies de PKC (Protein Kinase C) [108], la
famille Rho [109], PI3K [110] et les MAPK [92]. En plus de moduler plusieurs processus
cellulaires, l'activation de ces voies régule l'expression génique [23, 100, 111-114]. Par
exemple, les FC activent le facteur de transcription N F - K B (Nuclear Factor-kappa B ) [95.

115] qui contrôle un large éventail de gènes [9, 94], particulièrement les gènes impliqués
dans les processus d'inflammation et de la réponse immunitaire. Ainsi, les FC mènent à
l'activation et la propagation de signaux intracellulaires à travers un réseau complexe de
transduction ce qui module l'activité cellulaire et l'expression génique des CE. Ceci fait en
sorte que les FC modulent de façon importante l'activité des CE.
1.5.2 Expression génique
Le profil d'expression génique des CE varie selon l'environnement hémodynamique. En
effet, la présence d'un flux laminaire augmente ou diminue l'expression de plusieurs gènes.
Les changements d'expression sont nombreux et affectent des gènes qui codent pour
différent types de protéines impliquées dans la physiologie des CE. On y retrouve, entre
autres, des facteurs de croissance, des molécules d'adhésion, des substances vasomotrices.
des facteurs impliqués dans la coagulation et beaucoup d'autres types. L'évolution des
micros puces à ADN (DNA microarray) a permis d'évaluer le profil d'expression génique
in vitro des CE dans différente condition de flux hémodynamique [100, 111, 112, 114]. Le
profil d'expression génique des CE en condition de flux diffère de celui qui est en condition
statique, mais on remarque également des différences selon le temps d'exposition au flux et
selon la nature du flux, c'est-à-dire laminaire ou turbulent et puisé ou continu [116, 117].
La présence de FC physiologique induit l'expression de gènes nécessaires au maintien de
l'homéostasie des CE et des vaisseaux sanguins. Il est impossible de survoler tous les gènes
dont l'expression est influencée par les FC. Néanmoins, nous présentons une brève
description de gènes impliqués dans la physiologie endothéliale et étudiés dans le cadre de
ce travail.
PECAM
Aussi connu sous le nom de CD-31, PECAM est une glycoprotéine exprimée dans les CE,
les plaquettes et les leucocytes. Elle est souvent utilisée comme marqueur endothelial.
PECAM a des fonctions d'adhésion cellulaire et de signalisation. On la retrouve aux sites
des jonctions entre les CE avoisinantes et confluentes. Elle joue un important rôle dans les

interactions entre les CE et les éléments circulants du sang, notamment dans l'agrégation et
l'extravasation des leucocytes [9, 11, 94]. L'influence de l'exposition des CE à un flux
physiologique sur le niveau d'expression du gène de PECAM semble différer selon les
études [111, 114. 116, 118-120]. Toutefois, la tendance penche vers une diminution de
l'expression.
VECAD
VECAD est aussi une protéine d'adhésion cellulaire qui est également un marqueur
endothelial commun étant donné sa spécificité aux CE. Elle fait partie de la super famille
des cadhérines qui sont des protéines d'adhésion cellule-cellule dépendantes du calcium
[121]. On la retrouve aux jonctions d'adhésion cellulaire et son côté cytoplasmique peut se
lier avec plusieurs partenaires comme des caténies et des desmoplaquines qui permettent de
faire un lien avec les microfilaments d'actine et les filaments intermédiaires. VECAD joue
un rôle indispensable à la vasculogenèse, au remodelage vasculaire et est impliquée dans la
régulation de la perméabilité endothéliale. Elle fait aussi partie du même complexe
protéique mécano sensible que PECAM [106]. Encore une fois, l'influence des FC sur le
niveau d'expression du gène de VECAD diffère dans la littérature. Les résultats changent
dépendamment du substrat sur lequel les CE sont ensemencées [118], la nature du flux et le
type de CE [100, 111, 119].
vWF
Le facteur de Von Willebrand est une glycoprotéine plasmatique synthétisée de façon
constitutive par les CE. Bien qu'il ne soit pas spécifiquement produit par les CE, vWF est
aussi utilisé comme marqueur endothelial. vWF a des fonctions primordiales pour les
processus de coagulation. Sa fonction principale consiste à la liaison d'autres protéines
impliquées dans le mécanisme de coagulation. Sous sa forme active, le vWF est un gros
polymère formé de plusieurs monomères. Chaque monomère possède plusieurs domaines
spécifiques permettant l'interaction avec d'autres protéines comme le facteur VIII, le
récepteur des plaquettes gplb, l'héparine, le collagène et des intégrines. Ces interactions
sont nécessaires au déroulement normal de la coagulation sanguine. Les FC semble

diminuer l'expression du gène du vWF, mais encore une fois, les résultats ne sont pas
constants d'une étude à l'autre [100, 111, 118. 119, 122. 123].
tPA
Le gène du tPA code pour une protéase qui a comme principale cible la proenzyme
plasminogène. ce qui conduit à sa conversion en plasmine. Cette dernière dégrade plusieurs
protéines sanguines, notamment les caillots de fibrine [124]. Le tPA est donc une enzyme
antithrombotique qui est produite par l'endothélium. En plus de son rôle dans la fibrinolyse,
elle est impliquée dans la migration cellulaire et le remodelage vasculaire. L'imposition de
FC aux CE produit une élévation de l'expression du gène du tPA [100, 125. 126].
PAI-1
PAI-1 est le principal inhibiteur des activateurs du plasminogène tPA et urokinase. Il est
donc un inhibiteur de protéases à activité pro thrombotique [127]. Il est sécrété
principalement par les CE mais il peut aussi être exprimé dans le tissu adipeux. L'influence
des FC sur l'expression du gène PAI-1 est encore une fois mitigée. Certaines études
montrent une augmentation [114, 116] alors que d'autres indiquent une diminution de
l'expression [128].
eNOS
eNOS est une enzyme exprimée constitutivement par les CE dont l'activité produit de
l'oxyde d'azote (NO) à partir de L-arginine en présence d'oxygène et de NADPH
(Nicotinamide Adenine Dinucleotide Phosphate). eNOS joue un rôle majeur dans
l'homéostasie cardiovasculaire [129]. Elle est impliquée dans la régulation de la pression
sanguine, fangiogenèse et le remodelage vasculaire. L'influence positive du flux sanguin et
des FC sur l'activité d'eNOS et la production de NO est bien connue [82]. Néanmoins, le
rôle des FC dans l'expression du gène d'eNOS est moins clair [111, 125], même si certains
travaux ont observé une augmentation de l'expression de l'ARNm d'eNOS lorsque les CE
sont exposées aux FC [129-131].

ET-B
Chez les mammifères, il existe trois isoformes d'endothéline : ET-1. ET-2 et ET-3. Les
récepteurs membranaires qui transmettent leurs effets sont divisés en deux types : ET A et
E T B [132]. Les endothélines. principalement produites par les CE. sont de puissants
vasoconstricteurs. Les interactions entre les types d'endothéline et leurs récepteurs sont au
cœur de la régulation du tonus vasculaire [133]. Les récepteurs des endothélines sont
présents chez plusieurs types cellulaires, notamment les fibroblastes, les cellules
musculaires lisses vasculaires et les CE. Bien qu'on retrouve aussi les récepteurs des
endothélines dans le cœur, les reins et les poumons, il sera discuté seulement de leurs
actions dans les vaisseaux sanguins. Lorsque le récepteur se lie à l'endothéline. il peut
activer plusieurs types de protéine G qui à leur tour activent la voie de la phospholipase C
(PLC). L'effet de la liaison des endothélines au récepteur ET-B diffère selon le type
cellulaire. Chez les CML, elle provoque la vasoconstriction alors que chez les CE, elle
induit la production de NO qui a des effets vasodilatateurs. Malgré que des études aient
démontré l'influence des FC sur l'augmentation de l'expression du gène de l'endothéline-1
(ET-1), elles ne semblent pas avoir d'effet sur l'expression du gène du récepteur de type B
(ET-B) [100, 111, 114]. Cependant, une étude a démontré une augmentation de
l'expression du gène de ET-B avec des HUVEC exposées pendant 24 heures aux FC [134].
1.5.3 Fonctions des cellules endothéliales
Nous avons vu que les FC ont l'habileté de déclencher des signaux de transduction et de
modifier l'expression génique chez les CE. Par conséquent, la présence des FC dans
l'environnement des CE influence plusieurs facettes de leur physiologie. Nous présentons
une brève description de l'effet des FC sur la morphologie, la prolifération, la migration et
la perméabilité des CE.

Morphologie
L'effet le plus facilement observable des FC sur 1"endothelium est sans contre dit le
changement de morphologie des CE. En condition de culture traditionnelle, c'est-à-dire
dans un environnement hémodynamique statique, les CE ont une allure polygonale
communément appelée cobblestone. Lorsque que les contraintes de cisaillement sont
présentes, la morphologie des CE se transforme, passant de la forme polygonale à une
forme allongée dans l'axe [135]. Les CE s'allongent de façon parallèle à la direction du
flux. Cette morphologie correspond à celle qu'on retrouve in vivo. Ce changement de
morphologie est également observable au niveau des éléments du cytosquelette [81. 136].
En plus de l'allongement cellulaire, on observe la formation de microfilaments et de fibres
de stress parallèles au flux [137]. Ce phénomène de remodelage s'applique également aux
microtubules et aux filaments intermédiaires [138]. Au début de l'exposition au flux, durant
environ les trois premières heures, on observe le changement de morphologie, le
remodelage du cytosquelette et aussi un démembrement des jonctions cellulaires adhérentes
[139] et serrées [140]. Après une période d'ajustement (12 à 24 h), les CE ont formé un
dense réseau d'épaisses fibres de stress et rétablissent les jonctions cellulaires qui co
localisent avec les fibres de stress d'actine. D'un point de vue physique, ce remodelage de
la morphologie cellulaire est très important pour le maintien de l'équilibre des forces
mécaniques [141] et de l'homéostasie.
Prolifération et migration
Plusieurs études ont démontré que les FC influencent le cycle cellulaire et le taux de
renouvellement de l'endothélium [142, 143]. Un flux laminaire puisé ou continu réduit le
taux de prolifération des CE par rapport aux conditions statiques alors qu'un flux turbulent
en provoque l'augmentation. Sous un régime laminaire, on observe un nombre réduit de
cellules qui entre dans le cycle cellulaire et une majorité de cellules arrêtées en phase GO ou
Gl [144]. L'activation de la protéine p53 ainsi que la protéine du rétinoblastome (Rb) sont
impliquées dans cet arrêt du cycle cellulaire. En plus de diminuer le taux de prolifération
des CE, les FC physiologiques exercent un effet de suppression de l'apoptose via une
multitude de mécanismes [145-147]. L'addition de la diminution de la prolifération et de la

suppression de l'apoptose contribue grandement au maintien de l'intégrité de l'endothélium
par les FC.
Dans le même ordre d'idées, les FC influencent aussi la migration des CE. Sous un flux
laminaire, la migration est largement favorisée par rapport aux conditions statiques et
turbulentes [148-150]. Bien que plusieurs mécanismes soient impliqués dans cette
augmentation de la motilité. on peut souligner l'importance de l'augmentation des
interactions cellule-MEC et de la signalisation engendrée par la voie des FAK (Focal
Adhésion Kinase) [151, 152]. Dans un contexte de réparation de blessures et de remodelage
de la paroi vasculaire. la présence de FC constitue un stimulus important pour la migration
des CE.
Perméabilité
En temps normal, le passage des macromolécules est restreint par les jonctions
intercellulaires des CE. Lorsque les cellules entrent en mitose et se divisent, ces jonctions
sont altérées et mènent à l'augmentation de la perméabilité de Fendothélium [153]. Étant
donné que les FC provoquent la diminution du taux de division des CE, elles induisent du
même coup la diminution de la perméabilité. Des études ont lié avec une forte corrélation le
renouvellement des CE et le passage transendothélial de macromolécules [100. 144. 154].
Plusieurs études in vivo ont également associé les zones de flux turbulent aux zones de plus
grande perméabilité endothéliale [155]. Cette modulation de la perméabilité par les FC doit
être prise en compte leur d'études in vitro de la perméabilité endothéliale, notamment les
études de la biologie de la BHE [156-158].
Nous avons survolé quelques phénomènes physiologiques influencés par les FC, mais cette
revue n'est pas exhaustive. D'autres mécanismes sont aussi modulés par les forces
hémodynamiques tels que le métabolisme [159] et l'organogenèse du système
cardiovasculaire [160]. Néanmoins, il est indéniable que la présence des FC joue un rôle
prépondérant dans la physiologie vasculaire et endothéliale. Cette importance de l'impact
des FC sur le phénotype des CE doit donc être prise en compte lors des études in vitro de la
biologie endothéliale.

1.6 Modèles d'étude des cellules endothéliales
Dans un contexte de recherche portant sur la biologie cellulaire, l'expérimentation
in vivo est souhaitable, voire indispensable. Cependant, dans certains cas. ce type
d'expérimentation est souvent très complexe, voire même impossible, comme dans le cadre
de la recherche sur les interactions entre les forces hémodynamiques et l'endothélium. En
effet, la circulation sanguine est peu accessible à l'expérimentation et la quantification
précise des forces hémodynamiques in vivo reste particulièrement difficile. De plus, les
procédures d'expérimentation sur la circulation entraînent des conditions d'hypoxie et
d'hyponutrition qui rendent ardues les analyses phénotypiques. Dans cette optique,
l'expérimentation in vitro constitue un excellent substitut.
La plupart des études in vitro impliquant des CE utilisent des techniques de culture
cellulaire classique dans un environnement hémodynamique statique. La simplicité de ces
méthodes est certes un avantage. Toutefois l'interprétation et l'extrapolation des résultats
doivent tenir compte de cet environnement physique moins physiologique. Le domaine de
la biologie vasculaire exploite beaucoup l'étude de l'influence des forces hémodynamiques
sur la physiologie des différents types cellulaires vasculaires. ce qui permet une meilleure
compréhension de leur fonctionnement. Les travaux portant sur les CE en condition
dynamique sont nombreux. Bien qu'il s'agisse d'un pas vers un environnement plus
physiologique, il n'en demeure pas moins que les modèles in vitro utilisés sont souvent
encore loin des conditions in vivo. Effectivement, la majorité des études en condition
dynamique utilisent des chambres à flux laminaires qui exposent des monocouches de CE
en deux dimensions sur une surface recouverte de protéines permettant l'adhésion des
cellules comme la gélatine ou la fibronectine. Comparativement, in vivo, les CE sont dans
un environnement tridimensionnel composé d'une MEC complexe accompagnée d'autres
types cellulaires (CML et fibroblastes). L'utilisation du génie tissulaire pour développer des
modèles d'études in vitro possédant des caractéristiques physiologiques se rapprochant des
conditions in vivo est un outil particulièrement intéressant pour la biologie vasculaire [161-
163]. Dans le cadre de cet ouvrage, nous utilisons un tel modèle.

1.6.1 Substitut vasculaire construit par génie tissulaire comme modèle d'étude in vitro
Le laboratoire d'organogénèse expérimentale (LOEX) a développé une technique de
fabrication par génie tissulaire d'un substitut vasculaire complètement biologique [164]. La
technique de construction de ces vaisseaux reconstruits est appelée l'autoassemblage et elle
sera exposée en détail au chapitre matériel et méthodes. Le but ultime de cette méthode est
l'application clinique. Cependant, elle permet également la production de modèles d'études
in vitro élaborés et par conséquent plus près des conditions physiologiques in vivo. Ils sont
composés de MEC endogène sans aucun biomatériau artificiel, ils sont tridimensionnels et
formés des trois tuniques cellulaires présentes dans les vaisseaux sanguins : intima, média
et adventice. En plus de l'histologie homologue aux réalités in vivo [164, 165], ces
vaisseaux reconstruits possèdent également des propriétés mécaniques similaires aux
vaisseaux natifs, notamment pour la résistance à l'éclatement et la vasomotricité [164, 166-
168]. Ces caractéristiques de notre modèle en font un excellent outil pour des études de la
biologie vasculaire, notamment en environnement hémodynamique. Relativement aux
vaisseaux natifs humains isolés, les vaisseaux reconstruits par autoassemblage sont plus
facilement accessibles et plusieurs exemplaires peuvent être produits à partir des cellules
d'un même individu, réduisant ainsi la variabilité des résultats. D'une part, ils offrent
certains des avantages de la culture cellulaire comme l'accessibilité, la reproductibilité et
des enjeux éthiques plus simples. D'autre part, ils offrent également certains avantages des
vaisseaux ex vivo en étant plus près physiologiquement du tissu d'origine, d'un point de
vue cellulaire et fonctionnel.
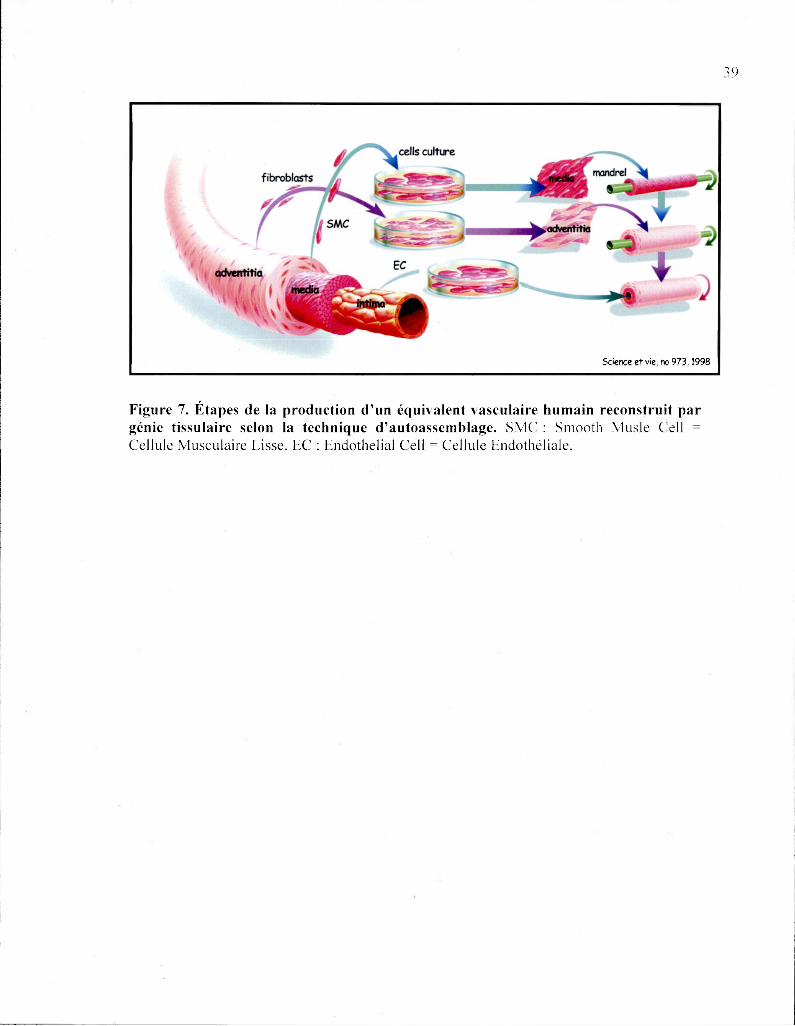
Science et vie. no 9 7 3 , 1 9 9 8
Figure 7. Étapes de la production d'un équivalent vasculaire humain reconstruit par génie tissulaire selon la technique d'autoassemblage. SMC : Smooth Musle Cell = Cellule Musculaire Lisse. EC : Endothelial Cell = Cellule Endothéliale.
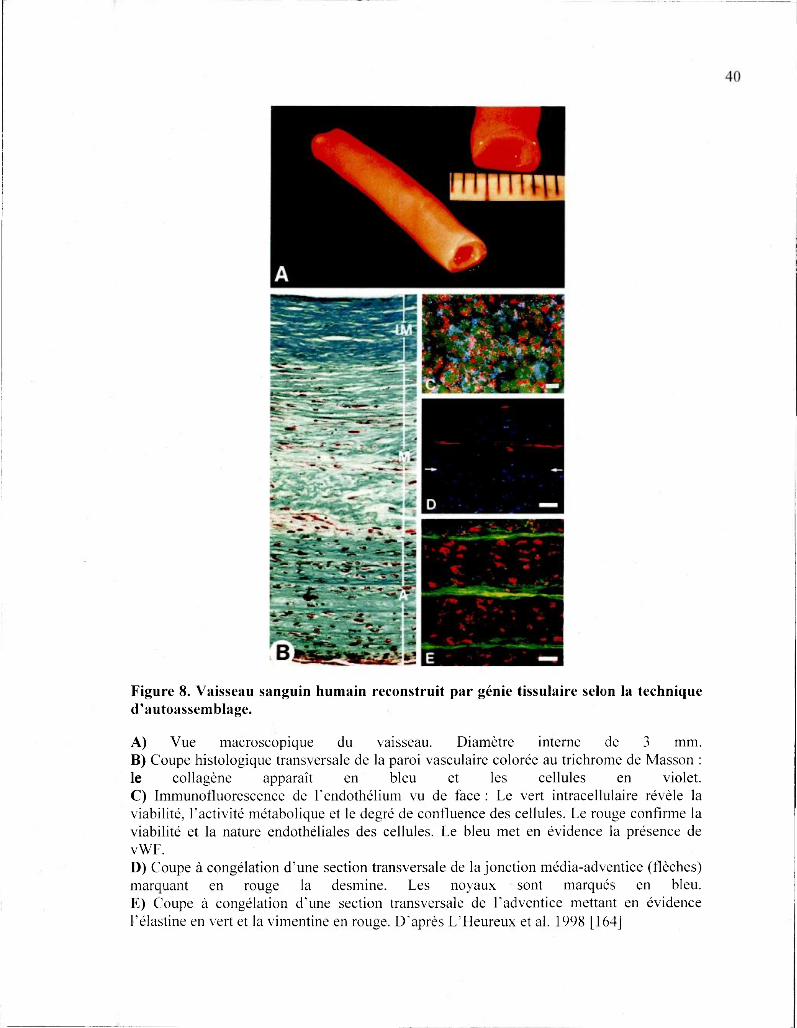
Figure 8. Vaisseau sanguin humain reconstruit par génie tissulaire selon la technique d'autoassemblage.
A) Vue macroscopique du vaisseau. Diamètre interne de 3 mm. B) Coupe histologique transversale de la paroi vasculaire colorée au trichrome de Masson : le collagène apparaît en bleu et les cellules en violet. C) Immunofluorescence de l'endothélium vu de face : Le vert intracellulaire révèle la viabilité, l'activité métabolique et le degré de confluence des cellules. Le rouge confirme la viabilité et la nature endothéliales des cellules. Le bleu met en évidence la présence de vWF. D) Coupe à congélation d'une section transversale de la jonction média-adventice (flèches) marquant en rouge la desmine. Les noyaux sont marqués en bleu. E) Coupe à congélation d'une section transversale de l'adventice mettant en évidence félastine en vert et la vimentine en rouge. D'après L'Heureux et al. 1998 [164]

À partir de la technique d'autoassemblage de vaisseaux reconstruits, notre laboratoire a
développé un modèle simplifié pour l'étude spécifique de fendothélium. Brièvement, la
production de MEC par des fibroblastes est stimulée par l'ajout d'acide ascorbique au
milieu de culture. Les fibroblastes synthétisent et organisent les protéines de la MEC de
façon à former un feuillet de cellules vivantes entourées de MEC. Après environ trois
semaines de maturation, les feuillets présentent une solidité suffisante pour être manipulés
et enroulés autour d'un mandrin tubulaire pour obtenir une adventice reconstruite. Ensuite,
les fibroblastes vivants sont éliminés par déshydratation pour obtenir une MEC
fibroblastique acellularisée tubulaire (MFT). Le support tubulaire est par la suite retiré afin
d'injecter les CE dans la lumière de la MFT. La MEC sécrétée par les fibroblastes fournie
la résistance mécanique nécessaire pour exposer les CE à un flux hémodynamique puisé à
des valeurs physiologiques. Cette version de substitut vasculaire construit par génie
tissulaire (MFT) est simplifiée du fait que la média et les fibroblastes de l'adventice sont
absents. En plus d'être plus simple techniquement, la MFT nous permet d'étudier
spécifiquement l'endothélium sans l'intervention des autres types cellulaires. Il est donc
possible d'extraire les protéines et l'ARN des CE en étant assuré de ne pas avoir de
contaminations des autres types cellulaires (CML et fibroblastes). En somme, la MFT est
un environnement tridimensionnel tubulaire composé de MEC endogène qui nous permet
d'étudier l'endothélium en condition de flux hémodynamique. Dans le cadre de ces
travaux, nous avons utilisé la MFT pour l'étude de l'endothélium en condition de flux
hémodynamique.

Figure 9. Étapes de la production d'une matrice fibroblastique tubulaire (MFT) endothélialisée. © Danielle Larouche, Patrick Charron. LOEX
1.7 Problématique et objectif
Bien plus qu'une simple couche de cellules pavimenteuses qui tapisse l'intérieur des
vaisseaux sanguins, l'endothélium est une composante centrale du système circulatoire.
Les CE sont impliquées dans une multitude de processus physiologiques allant de la
régulation de la circulation sanguine au contrôle de l'extravasation des cellules de
l'immunité. Étant donné cette grande implication dans divers processus biologiques, il
n'est pas surprenant que les CE soient présentes dans une panoplie de travaux de recherche
de tous les domaines. Cependant, les études impliquant les CE se font majoritairement
dans des conditions in vitro excluant une importante caractéristique de l'environnement
physique in vivo : les forces hémodynamiques. Nous avons vu dans l'introduction de cet
ouvrage que les forces hémodynamiques constituent d'importants stimuli pour les
fonctions endothéliales. L'étude des CE dans un environnement soustrait de ces stimuli
n'est donc pas idéal puisqu'il n'est pas fidèle aux conditions in vivo. Pour plusieurs
domaines d'études, il est déjà bien établi que la présence de ces forces change la réponse
des CE. La présence de ces forces est significative non seulement pour l'étude des CE par
rapport aux forces hémodynamiques mais aussi pour les domaines de l'artériosclérose, du
remodelage vasculaire [169, 170], de l'angiogenèse [171], de la pression artérielle [172],
des prothèses vasculaires artificielles [118, 173], du génie tissulaire [161. 163. 165]. du

cancer [65, 174], de la BHE [157, 158, 175] et bien d'autres champs d'étude impliquant les
CE. Etant donné que l'étude des CE humaines in vivo ou ex vivo est très complexe, tous
ces domaines de recherche bénéficieraient d'un modèle d'étude in vitro permettant l'étude
des CE dans un environnement plus physiologique. Nous supposons que notre modèle de
la MFT répond bien à cette problématique puisqu'il permet d'exposer les CE à un flux
hémodynamique sur une MEC endogène tridimensionnelle.
Objectif
Notre objectif principal est d'évaluer la réponse des CE ensemencées dans notre modèle de
la MFT, à suite d'une exposition de 4 heures à un flux hémodynamique physiologique. La
réponse a été évalué d'une part par la morphologie des CE analysée par microscopie.
d'autre part, par les changements du niveau d'expression de gènes impliqués dans la
biologie des CE. Nous avons comparé le niveau d'expression des gènes CD-31. VECAD.
vWF, tPA, PAI-1, eNOS et ET-B par PCR quantitative pour des conditions statiques et
avec flux hémodynamique.
Comme il a été démontré que la présence d'un flux hémodynamique permet de conserver
plus longtemps le phénotype de BHE chez les CE microvasculaires cérébrales in vitro [25,
156, 158, 175], nous croyons que notre modèle de la MFT pourrait être la base d'un
modèle in vitro innovateur de BHE construit par génie tissulaire. Notre objectif secondaire
est donc d'évaluer la possibilité de diminuer la perméabilité transendothéliale in vitro
d'HUVEC (Human Umbilical Vein Endithelial Cell) cultivées dans un milieu conditionné
d'astrocytome. L'utilisation des HUVEC plutôt que des CE cérébrales pour les étapes
préliminaires de développement d'un modèle in vitro de BHE s'explique par leur facilité
d'obtention et d'utilisation en culture. Dans le même ordre d'idées, les cellules
d'astrocytome sont aussi beaucoup plus accessibles et propices à la culture in vitro par
rapport aux cultures primaires d'astrocytes humains. Comme il a été rapporté dans la
littérature qu'il est possible de diminuer la perméabilité d'HUVEC [98, 144, 176, 177] par
l'influence de facteurs astrocytaires [25, 41 , 43, 178], nous voulons déterminer s'il est
possible de diminuer la perméabilité des HUVEC via l'exposition à du milieu conditionné
d'astrocytome humain. La perméabilité des HUVEC a été mesurée à l'aide de chambres de
Boyden et d'un marqueur moléculaire conjugué à un fluorochrome.

Matériels et Méthodes
2.1 Culture cellulaire
2.1.1 HUVEC
L'extraction des CE de veine de cordon ombilical humaine (HUVEC) a été effectuée tel
que décrit précédemment [164]. Brièvement, des cordons ombilicaux obtenus de nouveau
nés en bonne santé ont été conservés dans du milieu de transport à 4°C moins de 12 heures.
La veine a été canulée. à chaque extrémité pour y faire circuler du tampon HEPES (ICN
Biomedicals, Saint-Laurent, Québec. Canada) sans calcium à 10 mmol/L HEPES, 119
mmol/L NaCl, 6,7 mmol/L KC1 et à pH 7.35. Une solution de thermolysine 0.250 mg/mL
(Sigma, Oakville, On, Canada) à 37°C a ensuite été injectée pour rincer et remplir la veine.
Après une période d'incubation de 30 minutes à 37°C, la veine a été perfusée avec du
milieu de culture et les HUVEC ont été cultivé dans du M199 (Sigma. Oakville. ON,
Canada) contenant 20 % de sérum de veau fœtal (FBS, Hyclone, Logan, UT. USA). 20
fig/mL d'ECGS (Endothelial Cell Growth Supplément) (Sigma, Oakville, ON, Canada),
2,28 mM de glutamine (Invitrogen), 40 U/mL d'héparine (Léo Laboratories, Pickering.
Canada) 100 U/mL de pénicilline G (Sigma Oakville, ON, Canada) et 25 ug/mL de
gentamicine (Schering, Pointe-Claire, QC, Canada). Les HUVEC étaient utilisées à leur
cinquième passage dans les MFT. Les cellules ont été obtenues conformément à
l'approbation du comité éthique de l'Université Laval.
2.1.2 Fibroblastes
L'extraction des fibroblastes humains a été effectuée à partir d'un morceau de peau
provenant d'une réduction mammaire, tel que décrit précédemment [179]. Brièvement, les
morceaux de peau ont été incubés 3 h à 37°C dans une solution de thermolysine (Sigma,
Oakville, ON, Canada) à une concentration de 500 |ag/ml dans de l'HEPES (ICN
Biomedicals, Saint-Laurent, QC, Canada). L'épiderme a ensuite été séparé mécaniquement

du derme à l'aide de pinces. Le derme a été incubé 16 h à 37°C dans 0.125 U/ml de
collagénase H (Boehringer Mannheim. Laval, QC, Canada) pour récupérer les fibroblastes.
Les fibroblastes humains ont été cultivés dans du milieu DME complet, soit du Dulbecco-
Vogt modification of Eagle 's médium (DMEM; Gibco. Burlington, ON, Canada) contenant
10% de sérum de veau fœtal (SVF; H>clone. Logan. UT, USA). 100 U/ml de pénicilline G
(Sigma) et 25 ug/ml de gentamicine (Schering, Pointe-Claire, QC, Canada). Les
fibroblastes ont été utilisés à leur quatrième passage. Les cellules ont été incubées à 37°C
dans 8% de C O 2 et le milieu de culture a été changé tous les 2 jours. Les cellules ont été
obtenues conformément à l'approbation du comité éthique de l'Université Laval.
2.1.3 Cellules d'astrocytome
Les cellules d'astrocytome ont été obtenues de ATCC (Manassas, VA, USA).
CRL-1718 est une lignée isolée d'un astrocytome humain de grade IV. Selon ATCC
(American Type Culture Collection), ces cellules ont une morphologie astrocytaire et 70 à
80% des cellules expriment le GFAP (Glial Fibrillary Acidic Protein).
CCL-107 est une lignée isolée d'un astrocytome C 6 de rat. {Rat tus novegicus). Selon
ATCC, ces cellules ont une morphologie fibroblastique et expriment la protéine S-100
spécifique aux cellules gliales.
Ces cellules ont été cultivées dans du milieu DME complet, soit du Dulbecco-Vogt
modification of Eagle's médium (DMEM; Gibco, Burlington, ON, Canada) contenant 10%
de sérum de veau fœtal (SVF; Hyclone, Logan, UT, USA), 100 U/ml de pénicilline G
(Sigma) et 25 |ug/ml de gentamicine (Schering, Pointe-Claire, QC. Canada). Les
fibroblastes ont été utilisés à leur quatrième passage. Les cellules ont été incubées à 37°C
dans 8% de C O 2 et le milieu de culture a été changé tous les 2 jours.

2.2 Matrice Fibroblastique Tubulaire (MFT)
La technique d*autoassemblage utilisée pour la confection des MFT, tel que décrit
précédemment [164], s'est faite à partir de fibroblastes dermiques humains isolés telle que
décrite précédemment. Brièvement. 10 4 cellules/cm2 de fibroblastes humains sont cultivés
dans du milieu tel que décrit précédemment. Le milieu de culture est supplémenté de 50
ug/mL de sodium d'ascorbate (Sigma, Oakville, ON, Canada) pour stimuler la synthèse de
MEC. Après environ 28 jours de culture, les cellules ont formé un feuillet manipulable
comprenant cellules et leur propre MEC. Le feuillet a été détaché du flacon de culture de 75
cm 2 (Falcon, Becton Dickinsin Labware. Franklin Lakes, NJ, USA) et enroulé autour d'un
mandrin tubulaire de PTFE (Falcon. Becton Dickinsin Labware. Franklin Lakes. NJ. USA)
d'un diamètre externe de 4 mm. Les feuillets ont été enroulés trois ou quatre tours autour
du mandrin formant ainsi une épaisseur de 3 ou 4 couches. Les feuillets enroulés ont été
cultivés pendant un autre 28 jours dans des tubes de 50 mL (Falcon, Becton Dickinsin
Labware, Franklin Lakes, NJ, USA). Le milieu de culture supplémenté de sodium
d'ascorbate a été changé trois fois par semaine. Après cette période de maturation, les
couches de feuillets ont adhéré fermement les unes aux autres, formant un tissu tubulaire
cohésif. Ensuite, les tissus tubulaires ont été décellularisés. déshydratés et conservés à
-80°C jusqu'à utilisation. Nous avons ainsi obtenu des matrices fibroblastiques tubulaires
acellularisées.
Figure 10. Enroulement d'un feuillet de fibroblastes autour d'un mandrin tubulaire.
© Patrick Charron, LOEX.

2.3 Banc hémodynamique
Les MFT ont été décongelées et réhydratées dans du milieu M199 (Sigma. Oakville,
ON, Canada) à 4°C pendant 24 heures. Pour procéder à l'endothélisation. les MFT ont été
canulées à chaque extrémité et reliées à une tubulure pouvant être reliée à une pipette de 5
mL (Falcon, Becton Dickinsin Labware. Franklin Lakes. NJ. USA). Des HUVEC au
cinquième passage ont été ensemencé dans le lumen des MFT à raison d'environ 3 à 4
millions de cellules selon la longueur de la MFT. Les MFT endothélialisées ont été ensuite
placées à 37°C pendant trois heures pour laisser les cellules adhérer à la MEC. Pendant
cette période, les MFT sont placées dans des tubes spéciaux de 100 mL dans un dispositif
permettant une rotation lente des MFT pour assurer une endothélisation uniforme du lumen.
Après cette période d'adhésion, les cellules qui n'ont pas adhéré ont été rincées avec du
M199 (Sigma, Oakville, On, Canada). Les MFT endothélialisées ont été ensuite transférées
dans un banc hémodynamique de plexiglas stérile. Les MFT ont été reliées à une tubulure
stérile reliée à une pompe péristaltique (MasterFlex L/S, Cole-Parmer Instrument
Company, Barrington. IL, USA). Les MFT contrôle statique ont été placées dans le même
banc hémodynamique excepté qu'elles n'ont pas été reliées à la pompe. Le milieu de
culture dans lequel baigne les MFT est du M199 avec antibiotiques 100 U/mL de
pénicilline G (Sigma Oakville, ON. Canada) et 25 (ig/mL de gentamicine (Schering.
Pointe-Claire, Qc, Canada) et 5.96 g/L d'HEPES (ICN Biomedicals, Saint-Laurent, QC.
Canada). Le milieu qui circule dans la lumière des MFT est du M199 5 % de sérum de veau
fœtal (FBS, Hyclone, Logan, UT, USA), 2,28 mM de glutamine (Invitrogen), 40 U/mL
d'héparine (Léo Laboratories, Pickering, Canada) 100 U/mL de pénicilline G (Sigma
Oakville, ON, Canada) et 25 jag/mL de gentamicine (Schering, Pointe-Claire, QC, Canada).
5.96 g/L d'HEPES, 1 g/L de BSA (Sigma, Oakville, ON, Canada) et 80 g/L de Dextran
(Sigma, Oakville, ON, Canada)de façon à obtenir une viscosité de 0,04 Poise soit
l'équivalent du sang humain [6]. Le montage a été placé à 37°C et la pompe a fourni un
débit minimum pour faire circuler le milieu de culture à l'intérieur des MFT sans engendrer
une force de cisaillement significative. Les MFT ont maturé de cette façon pendant la nuit.
Le lendemain matin, le débit de la pompe a été élevé graduellement jusqu'à 170 mL/min,
de façon à obtenir un flux de 10 dynes/cm2 pendant 4 heures.

Après la période d'exposition au flux, les MFT ont été retirées du banc hémodynamique.
Par la suite, elles ont été ouvertes sur la longueur avec des instruments stériles et sur une
surface stérile sur glace, puis nous avons conservé un morceau d'environ 1 cm 2 pour les
analyses en microscopie. Les restes des MFT ont été placés dans des tubes stériles de 2 mL
(Eppendorf). puis plongés immédiatement dans de l'azote liquide et ensuite conservés à
-80°C.
Une étude de la mécanique des fluides a été effectuée pour notre montage de banc
hémodynamique dans le but de s'assurer que le flux est bel et bien laminaire et que la force
de cisaillement équivaut à 10 dynes/cm2. Cette étude a été réalisée par un l'étudiant au
doctorat en mécanique des fluides Karl Stéphane Rossignol.
Figure 11. Montage d'une MFT pour endothélisation. Une MFT est reliée à la tubulure
pour ensemencer des CE dans le lumen. © Patrick Charron, LOEX.

Figure 12. Montage du banc hémodynamique relié aux pompes péristaltiques. Les MFT endothélialisées sont installés dans une cuve de plexiglas stérile. Les MFT en condition dynamique sont reliées à la pompe. Les MFT en condition statique sont déposées dans le fond de la cuve. © Jean-François Oligny, LOEX
2.4 Microscopie
2.4.1 Contraste de phase
Les MFT ont été fixées au moins 24 h dans une solution d'Histochoice (Amresco, Solon.
OH, USA), puis enrobées dans la paraffine. Des coupes de 5 um ont finalement été
colorées au trichrome de Masson (TM) pour l'analyse histologique des tissus. Les images
ont été prises avec un microscope Nikon Eclipse E600 (Nikon, Tokyo, Japon) et une
caméra.
2.4.2 Microscopie électronique à balayage
Les biopsies de MFT ont été placées dans une solution de 2,5% glutaraldéhyde (M.E.C.A.
LAB) dans un tampon cacodylate 0,1 M, pH 7,5 pendant une nuit. Le lendemain, les
échantillons ont été rincés avec du tampon cacodylate 0,1 M à quatre reprises. La
déshydratation et la coloration ont été effectuées au service de microscopie électronique de
l'Université Laval. Les images ont été prises avec l'assistance de M. Richard Janvier.

2.4.3 Immuno fluorescence
Suite à l'expérience du banc hémodynamique, les biopsies de MFT ont été rincées avec du
tampon salin phosphate (PBS) et immédiatement transférées sur des lamelles de verre et
fixées dans du formol 3.7%. Les biopsies ont été perméabilisées avec du TritonR 0,1%)
(Bio-Rad Laboratories). Les images ont été obtenues et analysées avec un microscope
confocal Nikon Eclipse E800 combiné au système Nikon D-Eclipse Cl (Nikon).
Des cellules d'astrocytomes humain CRL-1718 ou de rat CCL-107 (ATCC) ont été
cultivées sur des lamelles de verre recouvertes de bromure de poly-L-lysine 0.01 mg/mL
(Sigma. Oakville. ON, Canada). Ces cellules ont été fixées avec du paraphormaldéhyde 4 %
(Baker) et perméabilisées avec de Féthanol 70% (Fisher). Les lamelles ont ensuite été
marquées pour GFAP. Des cerveaux de souris ont été enrobées dans une solution d'OCT
(Somagen, Edmonton. Ab. Canada), puis congelées à -80°C. Des coupes de 5 u.m
d'épaisseur ont subi le même traitement que les cellules sur lamelles et ont servies de
contrôles positifs.
Des HUVEC au cinquième passage ont été cultivées sur des lamelles de verre recouvertes
de gélatine en présence ou non de milieu conditionné de cellules d'astrocytomes (voir
culture en chambre de Boyden). Ces cellules ont été fixées avec du paraphormaldéhyde 4 %
(Baker) et perméabilisées avec de Féthanol 70% (Fisher). Les lamelles ont ensuite été
marquées pour ZO-1.
Les immunofluorescences pour GFAP et ZO-1 ont été visualisées avec des microscopes à
épifluorescence Nikon Eclipse E600 (Nikon, Tokyo, Japon) et Nikon Eclipse TE2000-U
(Nikon).
Pour toutes les immunofluorescences, les anticorps primaires ont été d'abord incubés 45
minutes sur les échantillons. Puis, les anticorps secondaires couplés à un fluorochrome ont
été incubés 30 minutes. Les anticorps primaires et secondaires ont été dilués avec du PBS
1% BSA (Sigma, Oakville, ON, Canada). Un marquage des noyaux cellulaires au Hoechst
33258 (Sigma) a aussi été effectué par une incubation de 10 minutes. Les contrôles négatifs
pour les immunomarquages ont été obtenus par omission du premier anticorps. La
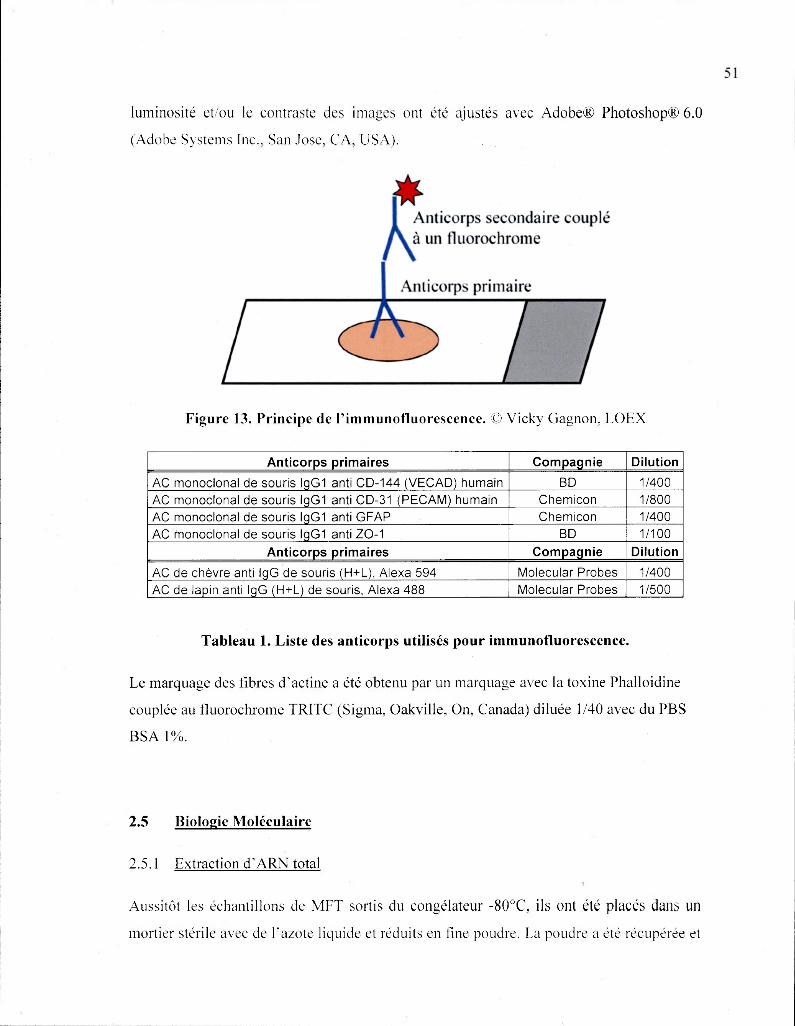
luminosité et/ou le contraste des images ont été ajustés avec Adobe® Photoshop® 6.0
(Adobe Systems Inc., San José. CA. USA).
Figure 13. Principe de rimmunofluorescence. © Vicky Gagnon, LOEX
Anticorps primaires Compagnie Dilution
AC monoclonal de souris lgG1 anti CD-144 (VECAD) humain BD 1/400 AC monoclonal de souris lgG1 anti CD-31 (PECAM) humain Chemicon 1/800 AC monoclonal de souris lgG1 anti GFAP Chemicon 1/400 AC monoclonal de souris lgG1 anti ZO-1 BD 1/100
Anticorps primaires Compagnie Dilution
AC de chèvre anti IgG de souris (H+L), Alexa 594 Molecular Probes 1/400 AC de lapin anti IgG (H+L) de souris, Alexa 488 Molecular Probes 1/500
Tableau 1. Liste des anticorps utilisés pour immunofluorescence.
Le marquage des fibres d'actine a été obtenu par un marquage avec la toxine Phalloidine
couplée au rluorochrome TRITC (Sigma, Oakville, On, Canada) diluée 1/40 avec du PBS
BSA 1%.
2.5 Biologie Moléculaire
2.5.1 Extraction d'ARN total
Aussitôt les échantillons de MFT sortis du congélateur -80°C, ils ont été placés dans un
mortier stérile avec de l'azote liquide et réduits en fine poudre. La poudre a été récupérée et

placée dans du tampon de lyse. L'ARN total a été extrait à l'aide de mini-colonnes
d'extraction RNeasy* (Qiagen, Mississauga, On. Canada). Le protocole recommandé par le
fabriquant a été suivi. La concentration et la pureté des extractions d'ARN ont été
effectuées à l'aide d'un spectrophotomètre ND-1000 (NanoDrop. Wilmington, DE. USA).
La pureté des extractions a été évaluée par les rapports d'absorbance des longueurs d'onde
260/280 nm et 260/230 nm.
2.5.2 RT-PCR
Les réactions de transcription inverse ont été effectuées avec 200 UI de l'enzyme
SuperScript 7/7® Reverse Transcriptase (Invitrogen, Burlington. On. Canada). Les réactions
ont été faites à partir de 1 ug d'ARN et selon le protocole recommandé par le fabriquant de
l'enzyme. Les réactions ont été effectuées dans un thermocycleur Icycler (Bio-Rad).
2.5.3 PCR
Toutes les réactions de PCR ont été effectuées avec l'enzyme Taq DNA Polymerase (New
England BioLabs, Pickering, On, Canada). Les réactions ont été effectuées dans un
thermocycleur Icycler (Bio-Rad) et selon le protocole recommandé par le fabriquant de
l'enzyme. Les réactions ont été effectuées avec 35 cycles d'amplification. Les amorces ont
été choisies à l'aide des logiciels en ligne Primer3 (http://frodo.wi.mit.edu/cgi-
bin/primer3/primer3_www.cgi) et IDT Oligo Analyser 3.0
(http://www.idtdna.com/analyzer/Applications/QligoAnalyzer/).
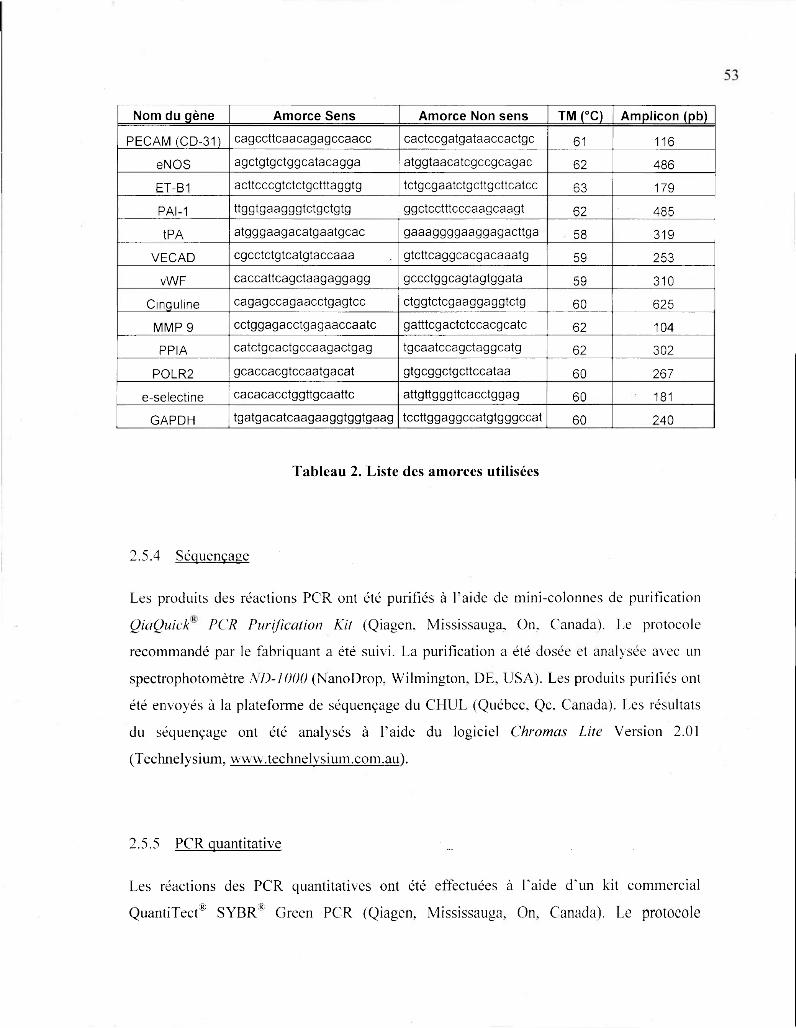
Nom du gène Amorce Sens Amorce Non sens T M (°C) Ampl icon (pb)
PECAM (CD-31) cagcct tcaacagagccaacc cactccgatgataaccactgc 61 116
eNOS agctgtgctggcatacagga atggtaacatcgccgcagac 62 486
ET-B1 acttcccgtctctgctttaggtg tctgcgaatctgcttgcttcatcc 63 179
PAI-1 t tggtgaagggtctgctgtg ggctcctt tcccaagcaagt 62 485
tPA atgggaagacatgaatgcac gaaaggggaaggagact tga 58 319
VECAD cgcctctgtcatgtaccaaa gtct tcaggcacgacaaatg 59 253
vWF caccat tcagctaagaggagg gccctggcagtagtggata 59 310
Cinguline cagagccagaacctgagtcc ctggtctcgaaggaggtctg 60 625
MMP 9 cctggagacctgagaaccaatc gatt tcgactctccacgcatc 62 104
PPIA catctgcactgccaagactgag tgcaatccagctaggcatg 62 302
POLR2 gcaccacgtccaatgacat gtgcggctgcttccataa 60 267
e-selectine cacacacctggttgcaattc attgttgggttcacctggag 60 181
GAPDH tgatgacatcaagaaggtggtgaag tcct tggaggccatgtgggccat 60 240
Tableau 2. Liste des amorces utilisées
2.5.4 Séquencage
Les produits des réactions PCR ont été purifiés à l'aide de mini-colonnes de purification
QiaQuick PCR Purification Kit (Qiagen, Mississauga, On, Canada). Le protocole
recommandé par le fabriquant a été suivi. La purification a été dosée et analysée avec un
spectrophotomètre ND-1000 (NanoDrop, Wilmington, DE, USA). Les produits purifiés ont
été envoyés à la plateforme de séquencage du CHUL (Québec, Qc, Canada). Les résultats
du séquencage ont été analysés à l'aide du logiciel Chromas Lite Version 2.01
(Technelysium, www.technelysium.com.au).
2.5.5 PCR quantitative
Les réactions des PCR quantitatives ont été effectuées à l'aide d'un kit commercial
QuantiTect® SYBR® Green PCR (Qiagen. Mississauga. On. Canada). Le protocole

recommandé par le fabriquant a été suivi. Les amorces utilisées figurent au tableau 2. Les
réactions ont été effectuées dans un thermocycleur Rotor-Gene RG-3000 (Corbett
Research, Kirkland, QC, Canada) et les résultats ont été analysés à Laide du logiciel Rotor-
Gene Version 6.0 (Corbett Research). Toutes les réactions pour un même gène ont été
effectuées lors de la même séance d'amplification. Toutes les réactions ont été faites à la
même température d'hybridation de 58°C. Pour chaque réaction, les données ont été
rapportées aux valeurs d'expression du gène rapporteur GAPDH [111. 114, 116. 119. 120,
180. 181]. L'expression de ce dernier n'était pas influencée par les conditions
expérimentales (Annexe 1). Les valeurs d'expression des gènes analysés ont été exprimées
en valeurs relatives par rapport au gène rapporteur GAPDH. Toutes les réactions ont été
effectuées à partir d'une quantité égale d'ADNc. Toutes les réactions ont été effectuées en
duplicata. Pour chaque gène, des courbes standard ont été obtenues à partir de dilutions des
produits de PCR purifiés (voir séquencage). Un duplicata d'une concentration connue
(environ 5 x 10° ng/rx) de chaque gène a été utilisé lors des réactions pour rapporter la
courbe standard respective.
2.6 Culture en chambre de Boyden
Des inserts ou chambres de Boyden adaptés pour des plaques de culture de 12 puits
(Falcon, Becton Dickinsin Labware, Franklin Lakes, NJ, USA) avec des membranes de
polyéthylène téréphtalate (PTE) et une porosité de 0,4 micron ont été enduites de gélatine
0,2 %. Des HUVEC au cinquième passage ont ensuite été ensemencées à l'intérieur des
inserts à raison de 30 000 cellules par insert. Les cellules ont maturé jusqu'à confluence
avant de faire le test de perméabilité transendothéliale. Le milieu de culture utilisé est du
M199 (Sigma, Oakville, On, Canada) contenant 20 % de sérum de veau fœtal (FBS.
Hyclone, Logan, UT, USA), 20 |ag/mL d'ECGS (Sigma, Oakville, On, Canada ), 2,28 mM
de glutamine (Invitrogen), 40 U/mL d'héparine (Léo Laboratories, Pickering, Canada)
100 U/mL de pénicilline G (Sigma Oakville, On, Canada) et 25 jag/mL de gentamicine
(Schering, Pointe-Claire, Qc. Canada). Les cellules ont été incubées à 37°C dans 8% de
C 0 2 et le milieu a été changé pendant 72 heures. Le milieu a été changé deux fois pendant
cette période. 24 heures avant le test de perméabilité, le milieu a été changé pour du M199

avec les mêmes constituants excepté FECGS et avec seulement 5 % de sérum de veau
foetal.
Dans le fond des puits, soit la chambre inférieure des chambres de Boyden, des cellules
d'astrocytomes CRL-1718 ou CCL-107 (ATCC) ont été ensemencées à raison de 100 000
cellules par puits recouvert de poly-L-lysine hydrobromide 0,01 mg/mL (Sigma. Oakville,
On, Canada). Les cellules ont été cultivées dans du milieu DME complet, soit du Dulbecco-
Vogt modification of Eagle 's médium (DMEM; Gibco. Burlington, ON. Canada) contenant
10% de sérum de veau fœtal (SVF; Hyclone, Logan, UT, USA), 100 U/ml de pénicilline G
(Sigma) et 25 |ig/ml de gentamicine (Schering, Pointe-Claire. QC. Canada). Les
fibroblastes ont été utilisés à leur quatrième passage. Les cellules ont été incubées à 37°C
dans 8% de C O 2 et le milieu de culture a été changé tous les 2 jours.
2.7 Test de perméabilité transendothéliale
Les tests de perméabilité ont été effectués avec des inserts tels que décrits
précédemment. Au moment du test, le milieu de culture de la chambre supérieure a été
changé pour du M199 (Sigma, Oakville, On. Canada) contenant 5 % de sérum de veau
fœtal (FBS, Hyclone, Logan, UT. USA), 2,28 mM de glutamine (Invitrogen). 40 U/mL
d'héparine (Léo Laboratories, Pickering, Canada) 100 U/mL de pénicilline G (Sigma
Oakville, On, Canada), 25 jag/mL de gentamicine (Schering, Pointe-Claire, QC, Canada) et
50 |ig/mL de BSA conjuguée au fluorochrome FITC (Molecular Probes. Eugène. OR.
USA). Les cellules ont été incubées à 37°C dans 8%o de C O 2 . A toutes les heures, pendant 6
heures et après 24 heures, un échantillon de 50 uL de milieu des chambres inférieures a été
recueilli et conservé à l'abri de la lumière à 4°C. La quantité de BSA-FITC ayant traversé
la monocouche d'HUVEC a été mesurée à l'aide d'un fluoromètre Varioskan Flash
(Thermo Electron Corporation. Finlande) et les résultats ont été analysé à l'aide du logiciel
Skanlt RE for Varioskan Flash version 2.4.1 (Thermo Electron Corporation, Finlande).
Les tests ont été effectués avec des inserts en présence de cellules d'astrocytome (CRL-
1718 ou CCL-107) dans la chambre inférieure ou avec du milieu conditionné de ces

Membrane
Chambre supérieure
Chambre inférieure
Cel ules endothéliales
nsert
Puit
Membrane
Figure 14. Schéma d'une chambre de Boyden. À gauche, on voit l'allure macroscopique
d'un insert. À droite, une coupe schématisée de Finsert à l'intérieur d'un puit d'une plaque
de culture. Adapté de http://www.bd.com (Falcon, Beckton Dickinson Labware, Franklin
Lakes, NJ, USA)
cellules ou encore avec seulement des HUVEC. Les contrôles étaient constitués d'inserts
sans cellules recouverts de gélatine 0,2%.
Le milieu conditionné de cellules d'astrocytome a été obtenu en récoltant le surnageant de
cellules CRL-1718 ou CCL-107 confluente dans des flacons de culture de 75 cm 2 (Falcon,
Becton Dickinson Labware, Franklin Lakes. NJ. USA). Le surnageant a été filtré avec un
filtre Milex GV (Millipore) de 0,22 u,m et mélangé avec du milieu frais dans un rapport un
pour un. Le milieu conditionné a été fait avec du M199 complet ou du DME complet avec
5 ou 20 % de sérum de veau foetal. Le temps de culture des HUVEC avec du milieu
conditionné a varié entre 24, 48, 72 et 96 heures.

Résultats
3.1 Morphologie
Pour évaluer la réponse des CE exposées à un flux hémodynamique physiologique
dans notre modèle d'étude de la MFT. nous avons observé l'endothélium suite à une
exposition de 4 heures à un flux de 10 dynes/cm2. De cette façon, il nous est possible
d'observer la confluence, la morphologie et l'expression de certaines protéines des CE
ensemencées dans une MFT avec et sans flux hémodynamique.
Figure 15. Coupes transversales de MFT. A et C) Coupes transversales de MFT sans cellule B et D) Coupe transversales de MFT endothélialisées et exposées à une flux de 10 dynes/cm2 pendant 4h. Coloration hématoxyline-éosine : La MEC est colorée en bleu et les cellules en rose. La barre de mesure = 100 |um

La MFT est composée d'une grande quantité de MEC synthétisée par les fibroblastes et
enroulée en plusieurs couches consécutives autour d'un mandrin. Une fois les fibroblastes
éliminés, nous conservons la MEC tubulaire dans laquelle nous ensemençons des HUVEC.
On peut observer en bleu la volumineuse MEC par rapport au revêtement de CE en rose
(Fig. 15 B et D). En microscopie électronique à balayage, on observe les fibres de MEC qui
composent la MFT.

Figure 17. Microscopie électronique à balayage d'une MFT endothélialisée en condition statique. Vue en pente de l'endothélium.
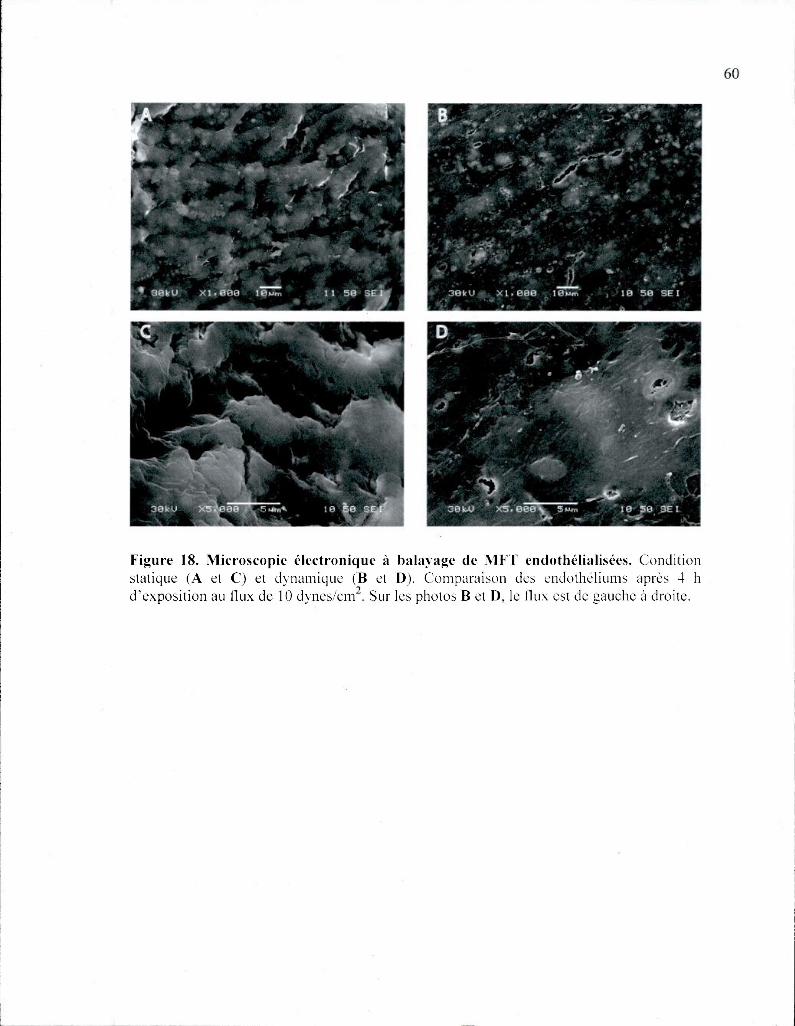
Figure 18. Microscopie électronique à balayage de MFT endothélialisées. Condition statique (A et C) et dynamique (B et D). Comparaison des endothéliums après 4 h d'exposition au flux de 10 dynes/cm . Sur les photos B et D, le flux est de gauche à droite.
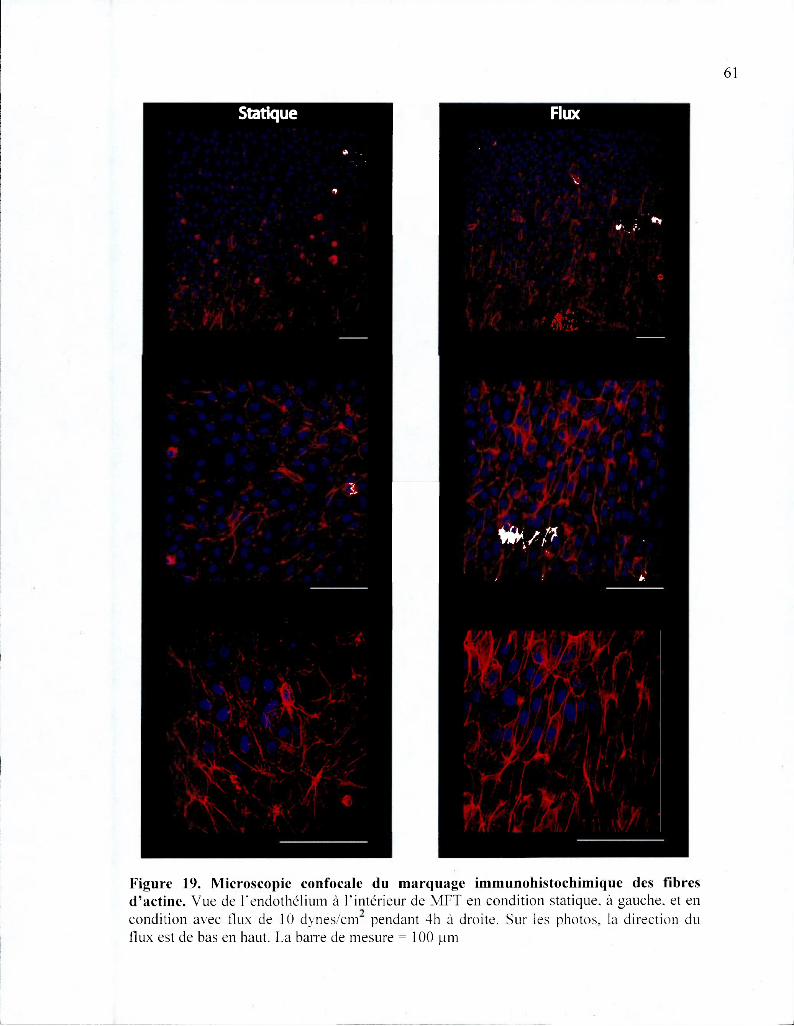
Statique mÊÊ Flux 9 H S I
•
I
Figure 19. Microscopie confocale du marquage immunohistochimique des fibres d'actine. Vue de l'endothélium à l'intérieur de MFT en condition statique, à gauche, et en condition avec flux de 10 dynes/cm pendant 4h à droite. Sur les photos, la direction du flux est de bas en haut. La barre de mesure = 100 (tm
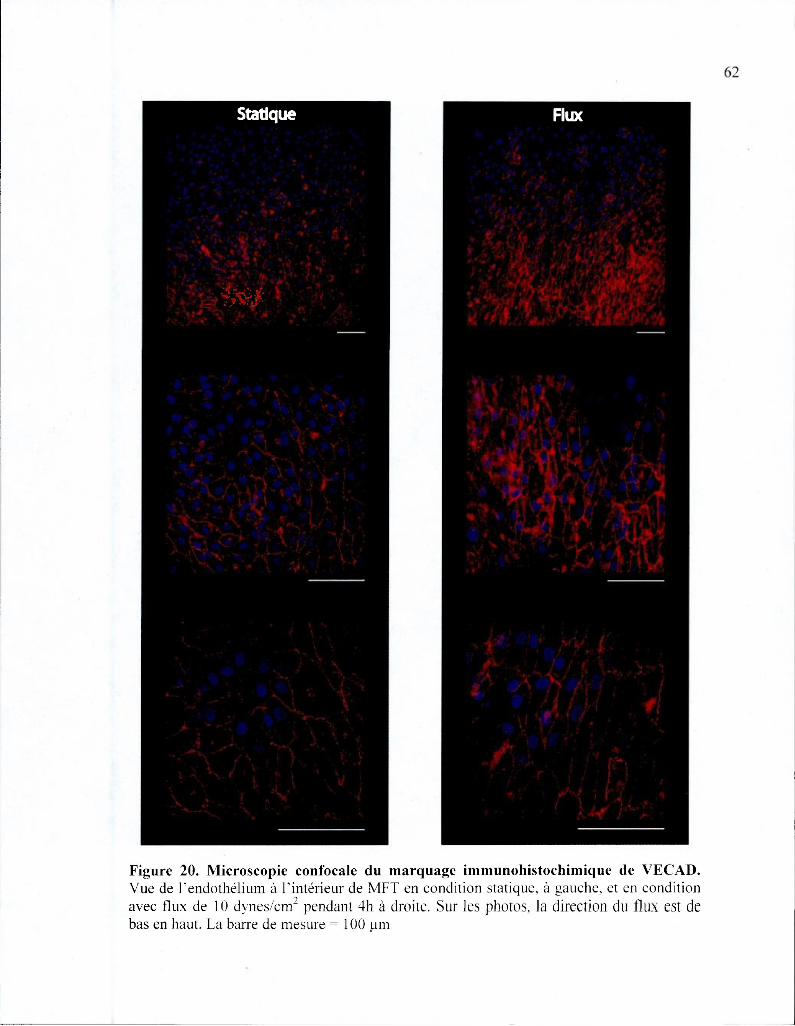
Statique Flux
•
Figure 20. Microscopie confocale du marquage immunohistochimique de VECAD. Vue de l'endothélium à l'intérieur de MFT en condition statique, à gauche, et en condition avec flux de 10 dynes/cm2 pendant 4h à droite. Sur les photos, la direction du flux est de bas en haut. La barre de mesure = 100 \im

Statique
Figure 21. Microscopie confocale du marquage immunohistochimique de PECAM. Vue de l'endothélium à l'intérieur de MFT en condition statique, à gauche, et en condition avec flux de 10 dynes/cm2 pendant 4h à droite. Sur les photos, la direction du flux est de bas en haut. La barre de mesure = 100 um

On remarque clairement que l'endothélium qui a été exposé au flux présente des CE
alignées et allongées parallèlement à la direction du flux comparativement à la morphologie
polygonale en condition statique. Malgré l'exposition aux forces hémodynamiques, les CE
présentent une confluence quasi parfaite à l'intérieur de la MFT. On doit mentionner que le
relief de la MFT fait en sorte qu'il est difficile d'obtenir un champ de visualisation aplani
qui permet de mettre au foyer l'ensemble des cellules. Ceci fait en sorte qu'on peut croire
qu'il y a des endroits sans cellules, en réalité ces endroits sont plus profonds ou plus élevés
par rapport au reste du champ qui est au foyer. En plus de l'alignement des cellules par le
flux, la microscopie électronique à balayage nous permet même d'observer l'aplatissement
des CE exposées au flux par rapport aux CE en condition statique (Fig. 18). Au niveau du
marquage des fibres d'actine, on remarque à plus fort grossissement, la présence de fibres
de stress parallèles à la direction du flux comparativement à la condition statique où elles
n'ont pas de direction communes (Fig. 19). Les marquages de VECAD (Fig. 20) et
PECAM (Fig. 21) montrent également l'alignement des CE en condition dynamique. On
remarque également, au niveau des marquages de VECAD et PECAM, une localisation
cytoplasmique plus prononcée chez la condition dynamique.
3.2 Expression génique
Pour complémenter l'analyse de la réponse des CE exposées à un flux
hémodynamique dans notre modèle de la MFT, nous voulions observer les changements
d'expression génique provoqués par le flux. Pour ce faire, nous avons extrait l'ARN total
des CE exposées ou non au flux dans une MFT. Après avoir converti l'ARN en ADN
complémentaire (ADNc), nous avons vérifié l'expression de certains gènes par PCR.

S S F F
VECAD I 1 1 1 I
Figure 22. Gel d'agarose des produits de PCR pour différents gènes. Réalisé à partir de l'ADNc des CE ensemencées dans une MFT. S = Condition Statique. F = Condition avec flux de 10 dynes/cm2 pendant 4 h. CE activées avec Finterleukine (3.
En dépit du fait que sur les gels d'agarose utilisés pour faire migrer les produits
d'amplification PCR nous avons observé une seule bande visible pour chaque paire
d'amorces utilisée, nous voulions tout de même nous garantir de la spécificité de la réaction
pour les amorces destinées à être utilisées ultérieurement en PCR quantitative. Pour nous
assurer de la spécificité de nos amorces, nous avons séquence les produits des réactions de
PCR.
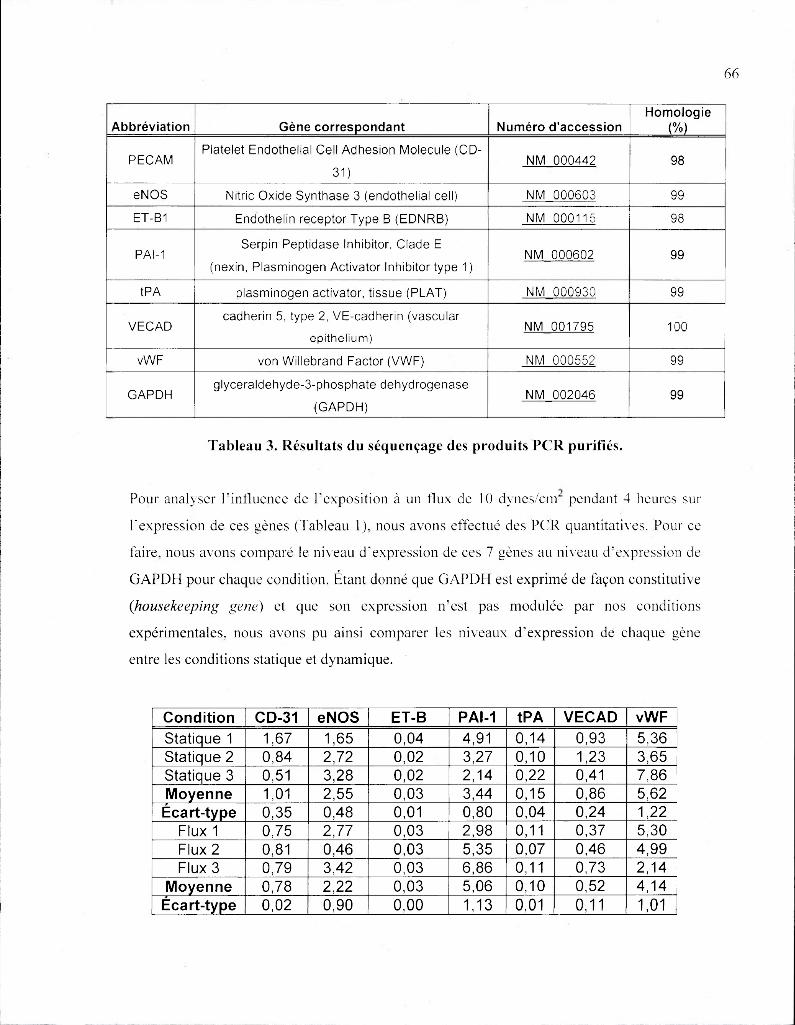
a b r é v i a t i o n L Gène correspondant Numéro d'accession Homologie
(%)
PECAM Platelet Endothelial Cell Adhésion Molécule (CD-
31) NM 000442 98
eNOS Nitric Oxide Synthase 3 (endothelial cell) NM 000603 99
ET-B1 Endothelin receptor Type B (EDNRB) NM 000115 98
PAI-1 Serpin Peptidase Inhibitor, Clade E
(nexin, Plasminogen Activator Inhibitor type 1) NM 000602 99
tPA plasminogen activator, t issue (PLAT) NM 000930 99
VECAD cadherin 5, type 2, VE-cadher in (vascular
epithelium) NM 001795 100
v W F von Wil lebrand Factor (VWF) NM 000552 99
GAPDH glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) NM 002046 99
Tableau 3. Résultats du séquencage des produits PCR purifiés.
Pour analyser 1" influence de F exposition à un flux de 10 dynes/cm pendant 4 heures sur
l'expression de ces gènes (Tableau 1). nous avons effectué des PCR quantitatives. Pour ce
faire, nous avons comparé le niveau d'expression de ces 7 gènes au niveau d'expression de
GAPDH pour chaque condition. Etant donné que GAPDH est exprimé de façon constitutive
(housekeeping gene) et que son expression n'est pas modulée par nos conditions
expérimentales, nous avons pu ainsi comparer les niveaux d'expression de chaque gène
entre les conditions statique et dynamique.
Condition CD-31 eNOS ET-B PAI-1 tPA VECAD vWF Statique 1 1,67 1,65 0,04 4,91 0,14 0,93 5,36 Statique 2 0,84 2,72 0,02 3,27 0,10 1,23 3,65 Statique 3 0,51 3,28 0,02 2,14 0,22 0,41 7,86 Moyenne 1,01 2,55 0,03 3,44 0,15 0,86 5,62
Ecart-type 0,35 0,48 0,01 0,80 0,04 0,24 1,22 Flux 1 0,75 2,77 0,03 2,98 0,11 0,37 5,30 Flux 2 0,81 0,46 0,03 5,35 0,07 0,46 4,99 Flux 3 0,79 3,42 0,03 6,86 0,11 0,73 2,14
Moyenne 0,78 2,22 0,03 5,06 0,10 0,52 4,14 Ecart-type 0,02 0,90 0,00 1,13 0,01 0,11 1,01
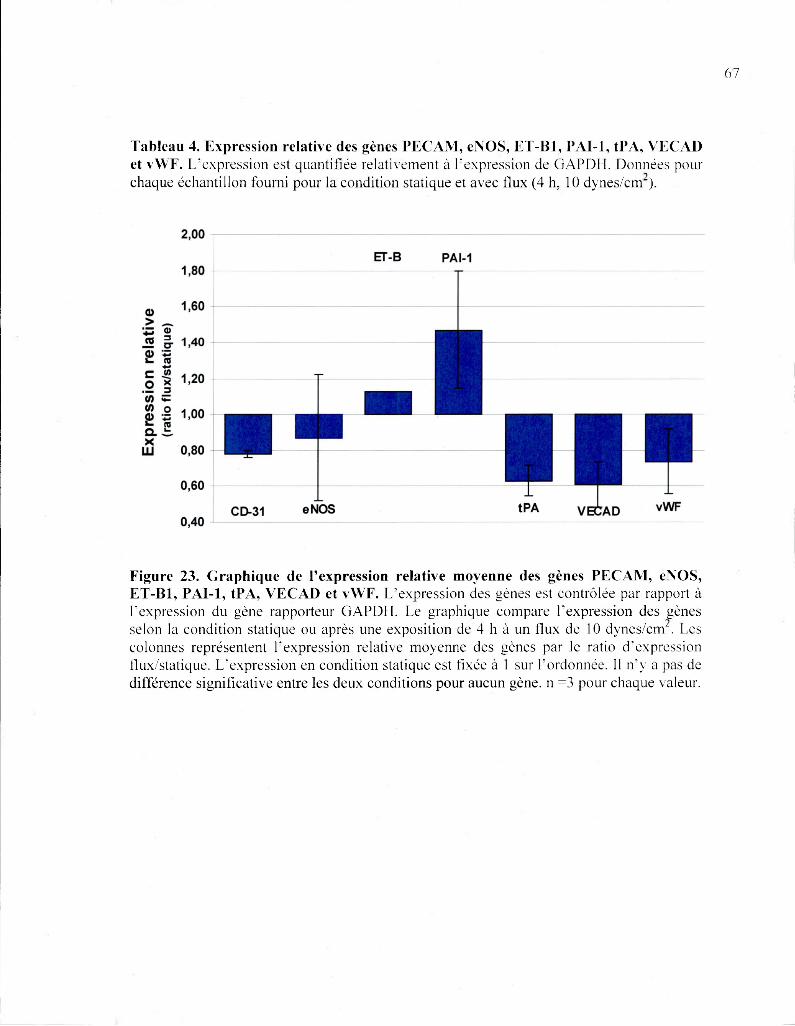
Tableau 4. Expression relative des gènes PECAM, eNOS, ET-B1, PAI-1, tPA, VECAD et vWF. L'expression est quantifiée relativement à l'expression de GAPDH. Données pour chaque échantillon fourni pour la condition statique et avec flux (4 h, 10 dynes/cm ).
Figure 23. Graphique de l'expression relative moyenne des gènes PECAM, eNOS, ET-B1, PAI-1, tPA, VECAD et vWF. L'expression des gènes est contrôlée par rapport à l'expression du gène rapporteur GAPDH. Le graphique compare l'expression des gènes selon la condition statique ou après une exposition de 4 h à un flux de 10 dynes/cm". Les colonnes représentent l'expression relative moyenne des gènes par le ratio d'expression flux/statique. L'expression en condition statique est fixée à 1 sur l'ordonnée. Il n'y a pas de différence significative entre les deux conditions pour aucun gène, n =3 pour chaque valeur.
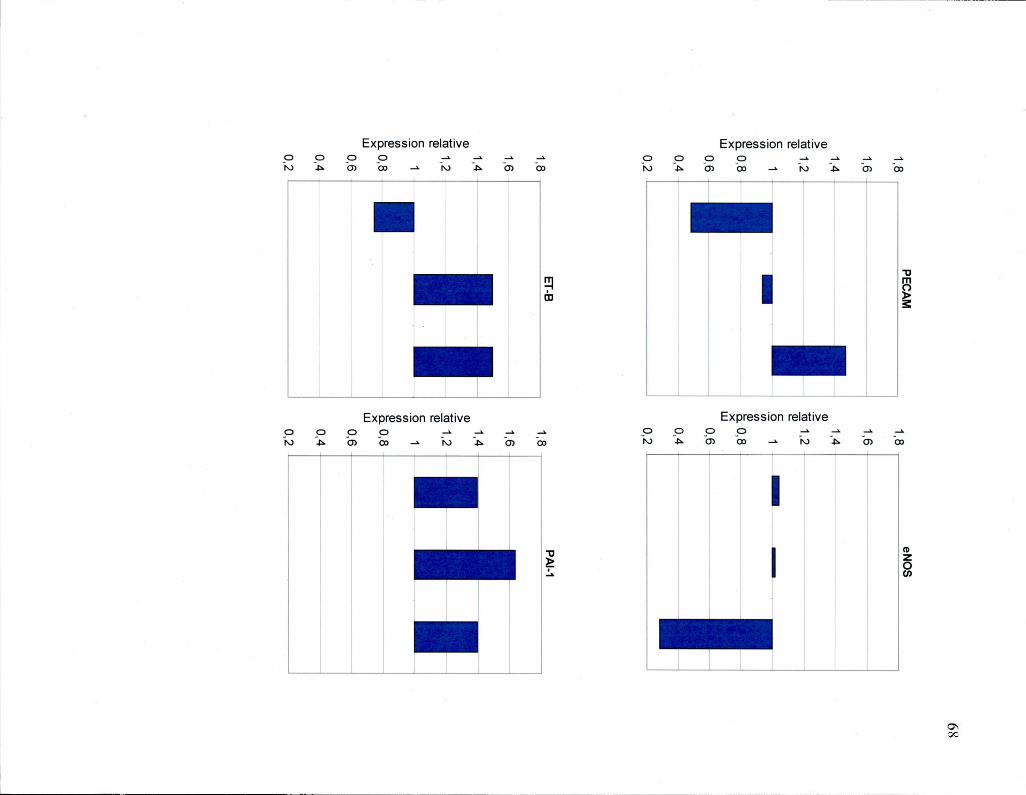
oc
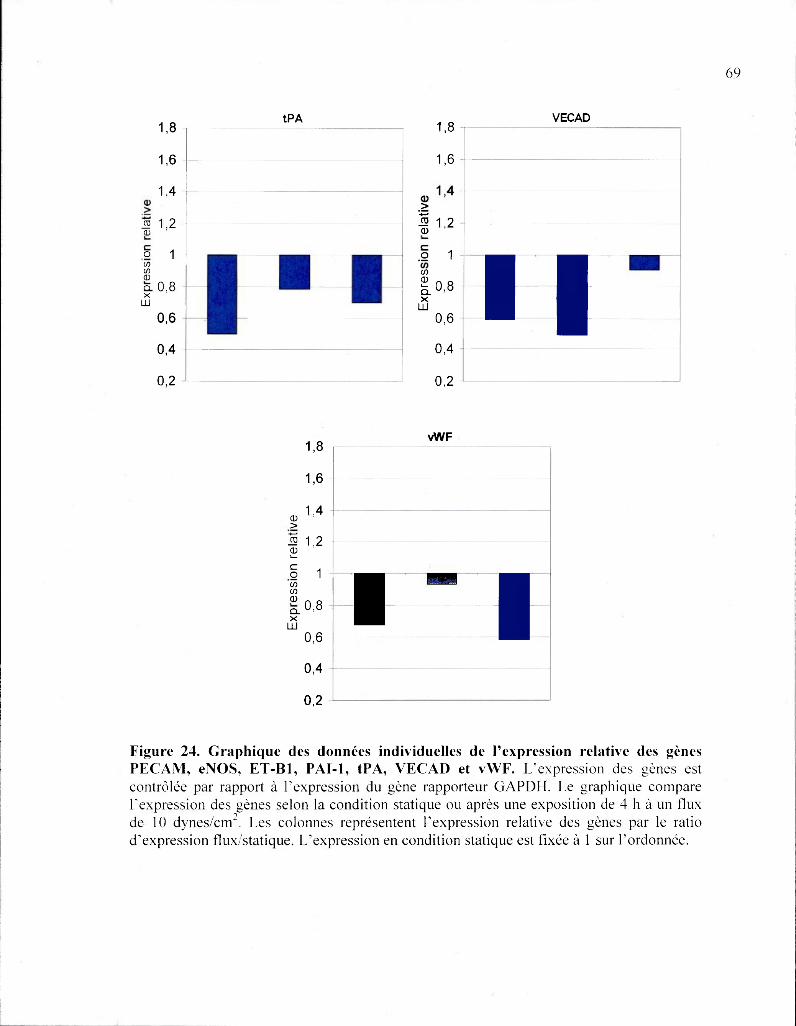
1,8
1,6
1,4
tPA
M 1,2 (D
o 1 CO co Q) Q. 0,8 X LU
0,6
0,4
0,2
1,8
1,6
> 5 1,2 CD
VECAD
1 g
co
| 0 . 8 X
LU
0,6
0,4
0,2
II 1,8
1,6
1,4 CD ' >
5 1,2 CD
i 1 co co CD CL x LU
vWF
0,8
0,6
0,4
0,2
• I I 0 Figure 24. Graphique des données individuelles de l'expression relative des gènes PECAM, eNOS, ET-B1, PAI-1, tPA, VECAD et vWF. L'expression des gènes est contrôlée par rapport à l'expression du gène rapporteur GAPDH. Le graphique compare l'expression des gènes selon la condition statique ou après une exposition de 4 h à un flux de 1 0 dynes/cm2. Les colonnes représentent l'expression relative des gènes par le ratio d'expression flux/statique. L'expression en condition statique est fixée à 1 sur l'ordonnée.

Sur les figures 23 et 24, une expression relative inférieure à 1 sur l'ordonnée signifie que
l'expression du gène en condition dynamique est inférieure à celle en condition statique qui
est fixée à 1. Par contre, une expression relative supérieure à 1 signifie que l'expression du
gène en condition dynamique est supérieure à celle en condition statique. Par exemple, une
expression relative égale à 2 signifierait que l'expression de gène est 2 fois plus élevée en
condition dynamique par rapport à la condition statique. Dans le même ordre d'idées, une
expression relative égale à 0.5 signifierait que l'expression du gène est 2 fois plus basse en
condition dynamique par rapport à la condition statique. Nous n'avons observé aucune
différence significative (p>0,05) entre les niveaux relatifs moyens d'expression mesurés en
condition statique et dynamique pour les 7 gènes analysés. On remarque une grande
variabilité des valeurs d'expression pour certains gènes (eNOS et PAI-1) et une meilleure
stabilité pour d'autres (PECAM et ET-B). On peut tout de même relever une certaine
tendance à une augmentation de l'expression en condition de flux pour ET-B1 et PAI-1
comparativement à une tendance à la baisse pour tPA, VECAD et vWF.
3.3 Barrière hémato-encéphalique
Dans le but d'évaluer la perméabilité des HUVEC en monocouche, nous avons
ensemencé des HUVEC dans la chambre supérieure de chambres de Boyden. Nous avons
ensuite suivi le passage d'un marqueur protéique au travers la couche de CE. Pour évaluer
l'influence sur la perméabilité endothéliale que pourraient avoir des facteurs solubles
astrocytaires, nous avons ensemencé des cellules d'astrocytome humain ou de rat dans la
chambre inférieure des chambres de Boyden. Le passage du marqueur BSA-FITC de la
chambre supérieure à la chambre inférieure a été mesuré toutes les heures pendant 4 heures
et après 24 heures.
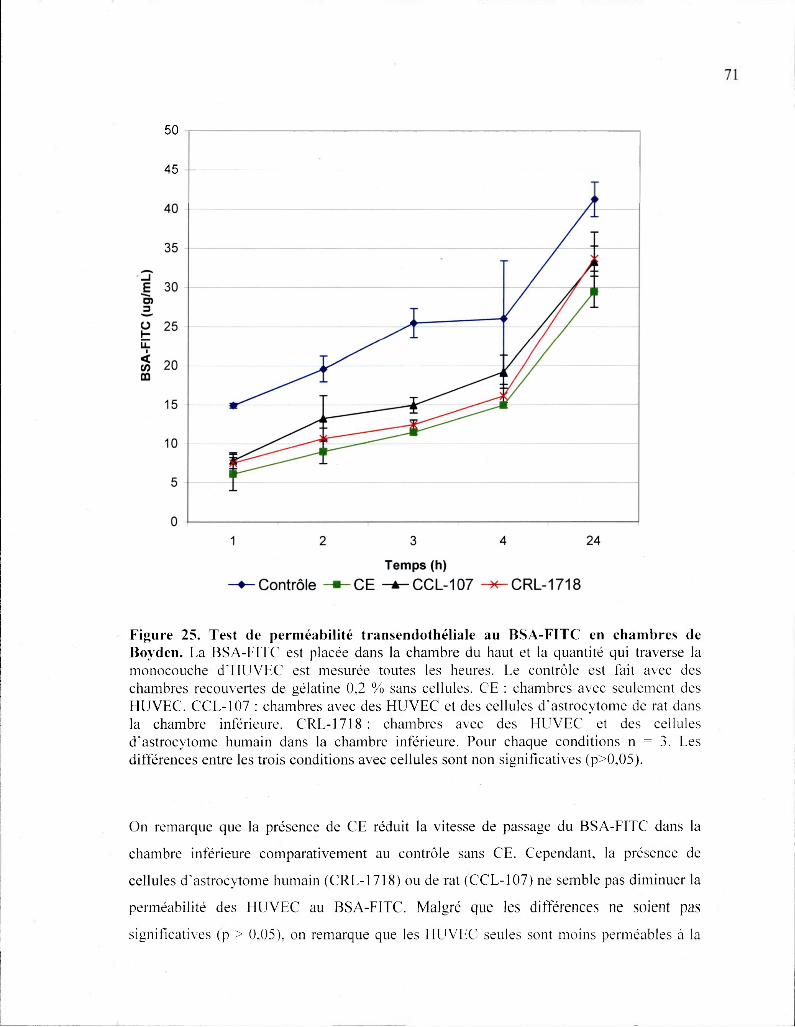
45
Figure 25. Test de perméabilité transendothéliale au BSA-FITC en chambres de Boyden. La BSA-FITC est placée dans la chambre du haut et la quantité qui traverse la monocouche d'HUVEC est mesurée toutes les heures. Le contrôle est fait avec des chambres recouvertes de gélatine 0,2 % sans cellules. CE : chambres avec seulement des HUVEC. CCL-107 : chambres avec des HUVEC et des cellules d'astrocytome de rat dans la chambre inférieure. CRL-1718 : chambres avec des HUVEC et des cellules d'astrocytome humain dans la chambre inférieure. Pour chaque conditions n = 3. Les différences entre les trois conditions avec cellules sont non significatives (p>0,05).
On remarque que la présence de CE réduit la vitesse de passage du BSA-FITC dans la
chambre inférieure comparativement au contrôle sans CE. Cependant, la présence de
cellules d'astrocytome humain (CRL-1718) ou de rat (CCL-107) ne semble pas diminuer la
perméabilité des HUVEC au BSA-FITC. Malgré que les différences ne soient pas
significatives (p > 0,05), on remarque que les HUVEC seules sont moins perméables à la

BSA-FITC par rapport aux HUVEC cultivées avec les cellules d'astrocytomes. Nous avons
répété cette expérience à plusieurs reprises avec différentes conditions de culture. Nous
avons utilisé différents milieux de culture (M 199 et EBM2) différents temps de culture
dans les chambres avant de faire le test de perméabilité (24, 48. 72 et 96h) et différentes
expositions des HUVEC aux facteurs solubles des cellules d'astrocytome, soit en utilisant
du milieu conditionné ou par la culture directe de ces cellules dans la chambre inférieure.
Malgré ces différentes conditions, nous avons toujours obtenu des résultats similaires,
c'est-à-dire pas de différence significative entre la perméabilité des HUVEC seules et de
celles cultivées en présence de milieu conditionné d'astrocytome.
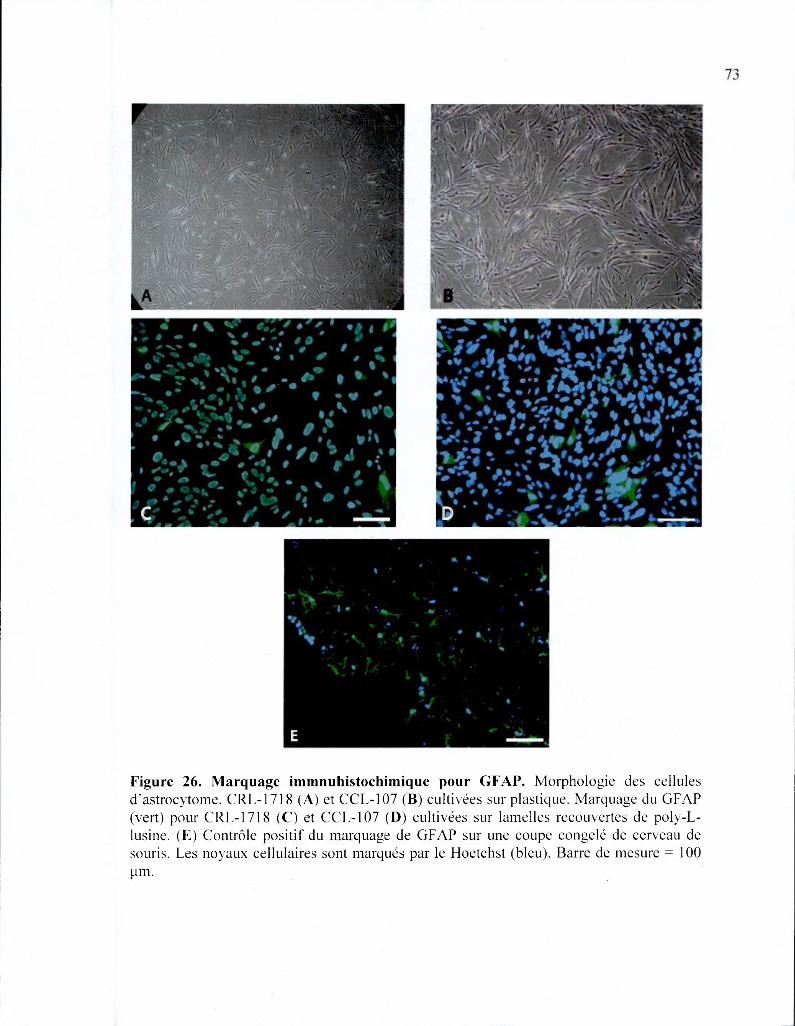
Figure 26. Marquage immnuhistochimique pour GFAP. Morphologie des cellules d'astrocytome. CRL-1718 (A) et CCL-107 (B) cultivées sur plastique. Marquage du GFAP (vert) pour CRL-1718 (C) et CCL-107 (D) cultivées sur lamelles recouvertes de poly-L-lusine. (E) Contrôle positif du marquage de GFAP sur une coupe congelé de cerveau de souris. Les noyaux cellulaires sont marqués par le Hoetchst (bleu). Barre de mesure = 100 uni.

Les cellules d'astrocytome ne présentent pas une morphologie similaire à des astrocytes
primaires. On remarque aussi que seulement un faible pourcentage de cellules CRL-1718 et
CCL-107 est positif pour le marquage du GFAP. Le fait de cultiver des HUVEC avec du
milieu de culture conditionné par les cellules d'astrocytome C6 CCL-107 ne semble pas
changer la présence et la quantité de ZO-1.
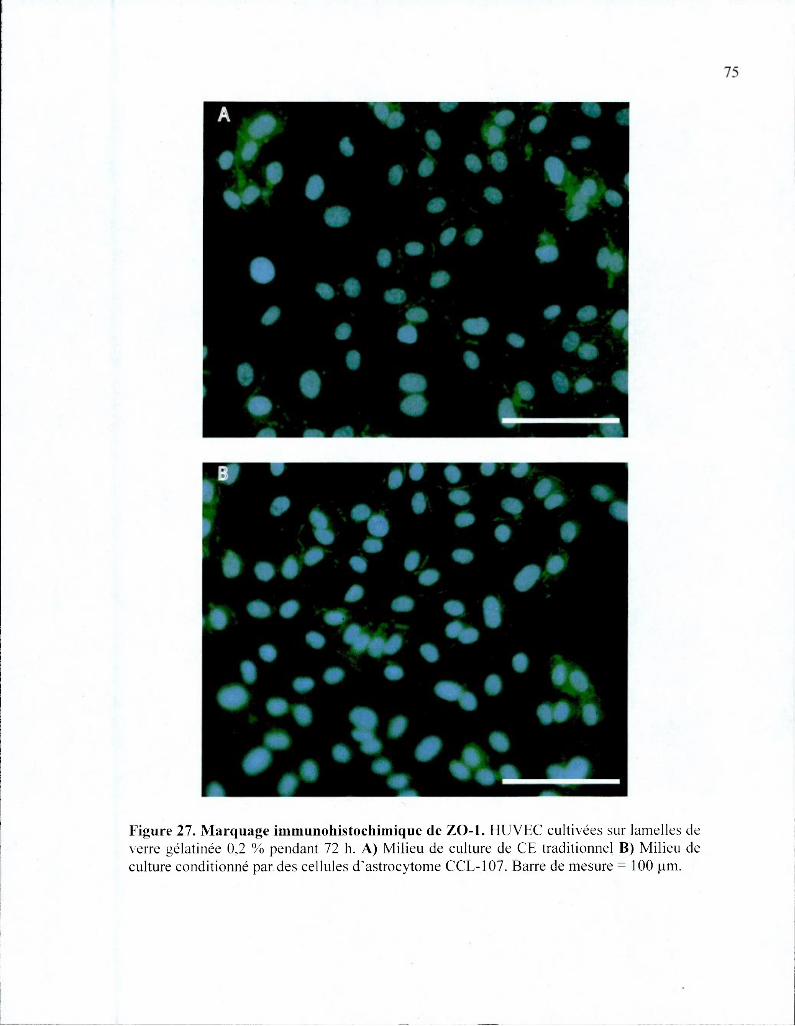
Figure 27. Marquage immunohistochimique de ZO-1. HUVEC cultivées sur lamelles de verre gélatinée 0,2 % pendant 72 h. A) Milieu de culture de CE traditionnel B) Milieu de culture conditionné par des cellules d'astrocytome CCL-107. Barre de mesure = 100 um.

Discussion
4.1 Réponses des cellules endothéliales au flux hémodynamique
Les observations microscopiques de l'endothélium des MFT en condition statique
ou dynamique démontrent bien que notre modèle de la MFT constitue un bon
environnement pour T étude des CE en condition hémodynamique. D'une part, la
conservation d'une confluence quasi parfaite et d'autre part le changement notable de
morphologie suite à une exposition de 4 heures à un flux hémodynamique physiologique
suppose que la MFT, composée de MEC endogène, constitue un support adéquat pour les
cellules. Comme prévu, les CE en condition dynamique se sont allongées et alignées
parallèlement au flux [135] en plus de présenter un remodelage du cytosquelette par la
formation de fibres de stress d'actine également parallèles au flux [137]. Ce changement de
morphologie et la réorganisation du cytosquelette sont essentiels au maintien des CE en
condition hémodynamique [141]. Les liens établis entre les cellules et la MEC semblent
être suffisamment forts pour permettre aux CE de résister aux forces engendrées par le flux.
Ces liens sont particulièrement importants dans un contexte de forces hémodynamiques
puisqu'en plus de maintenir les cellules en place, la liaison des intégrines à leurs ligands de
la MEC permet l'activation de signaux intracellulaires importants dans la réponse des CE
aux forces hémodynamiques [81, 95, 96]. La présence d'une localisation cytoplasmique de
VECAD et PECAM en condition dynamique nous amène à supposer qu'après 4 heures
d'exposition au flux, les cellules soient toujours en période d'ajustement. En effet, nous
supposons que cette localisation cytoplasmique concorde avec la disruption des jonctions
adhérentes [139] qui, après la période d'ajustement, se reformeront et établiront des liens
avec les éléments du cytosquelette remodelé. Ainsi, la préservation de la confluence
additionnée au remodelage du cytosquelette et au changement de morphologie nous pousse
à croire que les CE ensemencées dans notre modèle de la MFT sont en mesure de répondre
de façon adéquate aux forces induites par un flux hémodynamique physiologique. Bien que
ces résultats ne nous surprennent pas, ils appuient la validité de notre modèle de la MFT
pour l'étude des CE en condition hémodynamique.

Quoique l'observation des CE semble démontrer qu'elles sont en mesure de bien répondre
aux forces hémodynamiques, l'étape suivante était d'analyser si l'exposition aux FC
provoque des changements d'expression de certains gènes. Nous avons choisi d'analyser
l'influence du flux sur le niveau d'expression par PCR quantitative de 7 gènes impliqués
dans la physiologie endothéliale : PECAM. eNOS. ET-B1. PAI-1. tPA. VECAD et vWF.
Pour ce faire, nous avons soumis les CE ensemencées dans des MFT à un flux puisé
physiologique de 10 dynes/cm2 pendant 4 heures. En parallèle, nous avons traité
exactement de la même façon d'autres MFT, excepté qu'elles n'ont pas été exposées à un
flux. Nous avons ensuite extrait l'ARN total de ces CE pour analyser les différences
d'expression des 7 gènes entre les deux conditions, soit avec (dynamique) ou sans flux
(statique).
La PCR classique ne nous permet pas d'évaluer de façon précise le niveau d'expression des
gènes. Néanmoins, elle nous permet d'optimiser les conditions de la réaction PCR en plus
de valider la performance et la spécificité des amorces utilisées. A la suite d'une migration
sur gel d'agarose des produits de réactions PCR, nous n'avons observé qu'une seule bande
par paire d'amorces, ce qui nous laisse croire que la réaction est bel et bien spécifique.
Cependant, pour nous assurer que le produit d'amplification est réellement la cible visée,
nous avons isolé les produits de PCR et nous les avons fait séquencer. En somme, les
résultats de séquences obtenus concordent avec la séquence des gènes que nous avions
ciblés (tableau 3). Ceci confirme la spécificité des amorces utilisées pour l'analyse de
l'expression génique par PCR quantitative.
La réalisation de PCR quantitatives avec l'ADNc de CE ensemencées dans des MFT en
condition statique ou dynamique ne nous a pas permis d'observer des différences
significatives des expressions relatives moyennes des 7 gènes analysés. Cette absence de
différences statistiquement significatives est fort probablement due à la grande variabilité
entre les échantillons (figure 24 et tableau 4). En raison du nombre limité de MFT par
expérience, il est techniquement difficile d'avoir un grand échantillonnage. Dans ce cas ci,
nous avons seulement trois MFT par condition. On remarque que la majorité de la
variabilité provient d'un seul échantillons sur trois. Ceci fait en sorte que la moyenne des
trois échantillons ne représente pas nécessairement la situation observée et engendre des

écarts types élevés. Pour pallier à cette situation, il faudrait augmenter le nombre
d'échantillons afin de diminuer l'impact des données aberrantes et du même coup permettre
une analyse beaucoup plus juste de l'influence du flux sur l'expression génique.
Bien que les différences des niveaux d'expression ne soient pas significatives, on peut tout
de même observer certaines tendances. L'exposition pendant 4 heures à un flux de 10
dynes/cm2 semble induire une augmentation de l'expression de ET-B1 et PAI-1 et une
diminution pour PECAM. eNOS. tPA. VECAD et vWF. Il est très difficile de faire un lien
avec nos résultats et ceux qu'on retrouve dans la littérature puisque les conditions
expérimentales diffèrent énormément. Entre autres, la force et la nature du flux, le temps
d'exposition, le substrat et le type de cellules sont tous des facteurs qui altèrent les résultats
et leurs analyses. Malgré ces différences notables, on peut tout de même souligner que les
changements d'expression apportés par l'exposition au flux sur PECAM [118. 119], ET-
Bl[134] et vWF [118, 123] correspondent à la tendance dans la littérature alors qu'ils ne
correspondent pas pour eNOS [129, 131] et tPA [118. 125, 126]. Pour ce qui est de
VECAD et PAI-1. il n'y a pas de consensus sur l'effet du flux sur leur niveau d'expression,
cependant Brooks et al. ont aussi observé une diminution pour VECAD et une
augmentation pour PAI-1 après une exposition de 24 heures au flux [116]. L'influence du
flux ne semble pas avoir d'effet sur l'expression de ET-B1 conformément à ce que d'autres
études ont observé [111, 120].
Les effets antithrombotiques d'un flux hémodynamique sont bien connus [71, 86, 87, 128,
150, 159, 182]. C'est pour quoi à la suite de l'exposition au flux hémodynamique
physiologique, on pouvait s'attendre à une diminution de l'expression des gènes pros
thrombotiques PAI-1 et vWF et à une augmentation du gène antithrombotique tPA. À
l'exception de vWF, nous avons observé le contraire. Encore une fois, le fait que nos
résultats ne soient pas significatifs fait en sorte qu'on ne peut tirer de conclusions valables
et il serait important de reprendre ces expériences avec un plus grand nombre
d'échantillons. Dans le même ordre d'idées, il est bien établi que le flux provoque une
production d'oxyde d'azote (NO) chez les CE [82]. Cependant l'effet sur l'expression du
gène eNOS est beaucoup moins clair [111, 125], même si certains travaux ont démontré
une augmentation de son expression [129-131]. Dans notre cas. nous observons une très

grande variabilité de l'expression d'eNOS entre les échantillons, ce qui nous ne permet pas
de faire ressortir une tendance.
Il est intéressant de voir que les tendances d'une diminution de l'expression de PECAM et
VECAD correspondent à l'observation d'une localisation plus cytoplasmique après
l'exposition au flux chez les marquages immunohistochimiques (figures 20 et 21). Tel qu'il
a été mentionné précédemment, ceci nous laisse croire qu'il est possible que les cellules
soient en période d'adaptation au flux et qu'elles soient en réorganisation des éléments du
cytosquelette et des jonctions adhérentes [139]. On peut supposer qu'après cette période
d'ajustement les cellules rétabliront les jonctions adhérentes avec la nouvelle structure du
cytosquelette de façon à rétablir un nouvel état d'équilibre avec l'environnement
hémodynamique.
Le choix des 7 gènes s'est basé d'une part sur leur implication dans la physiologie
endothéliale et d'autre sur l'influence possible du flux sur leur expression, telle qu'il a été
rapporté dans la littérature. Cela dit, le fait que nous n'observons pas de différences
significatives d'expression pour ces gènes n'indique en rien que les cellules n'ajustent pas
leur expression génique en réponse au flux. En effet, bien des facteurs peuvent expliquer
cette absence de différences significatives. Bien sûr. il faut augmenter le nombre
d'échantillons et répéter l'expérience, mais il faut également préciser qu'il est possible que
le flux, aux valeurs utilisées, n'ait pas d'influence sur l'expression de ces gènes précis.
Également, le temps d'exposition au flux n'était peut-être pas suffisant pour atteindre un
état d'équilibre vis-à-vis le changement d'expression de certains gènes. Le fait que les
cellules pourraient être en période d'ajustement au flux et qu'elles ne soient pas totalement
revenues en état d'équilibre par rapport à l'environnement hémodynamique pourrait
expliquer en partie la variabilité de nos résultats. Manifestement, les changements
d'expression génique ne sont pas instantanés et il est fort possible que les échantillons ne
soient pas tous au même stade. Cependant, les variabilités sont également présentes entre
les échantillons qui n'ont pas été soumis au flux. On peut donc suspecter que les techniques
peuvent être en partie à l'origine de la variabilité. Néanmoins, toutes les précautions ont été
prises pour avoir des conditions constantes d'extraction de l'ARN, de RT-PCR et de PCR
quantitative. Les échantillons ont été analysés en duplicata pour la PCR quantitative, mais il

aurait été préférable de faire deux réactions RT-PCR à partir d'une extraction d'ARN pour
s'assurer de l'uniformité de la réaction. Toutefois, la faible quantité d'ARN recueillie à
partir d'une MFT endothélialisée ne le permet pas selon les conditions de RT-PCR que
nous nous sommes fixées, soit 1 ug d'ARN total par réaction. De plus, chaque MFT est
construite à la main, ce qui occasionne des variations inévitables entre elles. Leurs
caractéristiques peuvent différer quelque peu et ces différences peuvent être à l'origine
d'une certaine variabilité des résultats. Par exemple, une petite différence dans la taille de la
MFT peut engendrer des différences significatives des FC. Des différences au niveau des
forces hémodynamiques peuvent aussi provenir du montage du banc hémodynamique. Par
exemple, si la pompe péristaltique ne fournit pas un flux équivalent pour chaque MFT ou
s'il y a des fuites de liquide non perceptibles, des FC différentes seront imposées aux CE.
Il est important de spécifier que nous avons analysé l'effet du flux hémodynamique sur
l'expression génique. Tel qu'il a été spécifier en introduction de ce travail, la FC est la
force qui agit au niveau de l'endothélium, mais nous n'excluons pas que les autres types de
forces engendrées par le flux puisé peuvent aussi avoir un effet sur la réponse des CE. C'est
pour quoi nous référons à l'effet du flux et non pas à l'effet de la FC uniquement. Au cours
de ces expériences, nous avons utilisé un flux physiologique de 10 dynes/cm2 et nous
n'avons pas mesuré les valeurs de pression transmurale.
En somme, pour améliorer la pertinence de ces travaux, il faut absolument répéter
l'expérience, augmenter le nombre d'échantillons et idéalement faire l'expérience en
fonction du temps. C'est-à-dire qu'on devrait analyser les changements d'expression
génique après 2, 4, 8 et 24 heures par exemple. Il serait également logique d'étendre
l'analyse à plusieurs gènes à l'aide de puces à ADN. Cela dit, il faut mentionner la lourdeur
des manipulations reliées aux modèles construits par génie tissulaire. Il serait
techniquement très difficile de produire une grande quantité de vaisseaux reconstruits et de
les exposer simultanément à un flux dans notre appareillage de banc hémodynamique. De
plus, il faut continuer à peaufiner les méthodes d'extraction d'ARN et étudier la pertinence
d'utiliser des systèmes de RT-PCR qui exigent moins de matériel de départ. De cette
manière, nous pourrions obtenir des échantillons de meilleure qualité et en quantité
suffisante pour travailler aisément.

4.2 Barrière hématoencénhalique
Dans le but de mettre au point un éventuel modèle in vitro de BHE humaine, nous
voulions évaluer la possibilité de maintenir l'intégrité la barrière endothéliale d'HUVEC
par la présence de cellules d'astrocytomes. Le matériel biologique nécessaire à l'utilisation
de cultures primaires de CE micros vasculaires cérébrales et d'astrocytes humains est très
difficile à obtenir pour des questions techniques et éthiques. Effectivement, l'obtention de
tissus cérébraux humains est complexe, sans compter que les astrocytes adultes ne se
conservent pas en culture. C'est pourquoi nous voulions nous rabattre sur les HUVEC et
des cellules d'astrocytomes qui sont beaucoup plus facile d'utilisation et à obtenir.
Quelques études ont rapporté avoir provoqué un phénotype de BHE chez des CE d'origine
non cérébrale [98, 144, 176, 177] suite à une coculture avec des astrocytes. D'ailleurs,
compte tenu des difficultés reliées à l'utilisation des cultures primaires d'astrocytes. il est
commun d'utiliser des cellules d'astrocytomes dans des modèles in vitro de BHE [53. 54.
144, 183, 184]. Étant donné qu'une éventuelle mise au point d'un modèle in vitro de BHE
demandera une quantité appréciable de cellules, nous voulions vérifier la possibilité de
provoquer des caractéristiques de BHE chez des HUVEC cultivées en présence de facteurs
solubles de cellules d'astrocytome. Nous avons vérifié la faculté de conservation de la
barrière endothéliale des HUVEC par deux lignées d'astrocytome : une humaine (CRL-
1718) et une de rat C6 (CCL-107). Aucune des deux lignées n'a permis de diminuer la
perméabilité transendothéliale des monocouches d'HUVEC au marqueur protéique BSA-
FITC. L'utilisation de ce marqueur est très répandue dans la mesure de la perméabilité
transendothéliale puisque que l'albumine est la protéine sanguine la plus commune et la
plus abondante. La diminution de la perméabilité transendothéliale est certainement la
caractéristique indispensable de la BHE et c'est pourquoi nous nous sommes précisément
attardés à cette dernière. Sans cette diminution de la perméabilité, l'analyse des autres
attributs de la BHE serait vaine. Nous avons tout de même vérifié si les HUVEC cultivées
avec du milieu conditionné d'astrocytome exprimaient une plus grande quantité de
jonctions serrées par le marquage immunohistochimique de la protéine ZO-1 [30, 31, 34].
Nous n'avons observé aucune différence entre les contrôles et les HUVEC cultivées avec
du milieu conditionné d'astrocytome. Nous avons choisi les deux lignées d'astrocytome du
fait que ATCC stipule qu'elles ont conservé une morphologie astrocytaire et qu'elles

exprimaient le marqueur glial GFAP (Glial Fibrillary Acidic Protein). Cependant, nous
avons été incapables d'observer un marquage immunohistochimique non équivoque de
GFAP pour les deux lignées. L'efficacité de notre anticorps et de notre méthode de
marquage a été validée sur des coupes congelées de cerveaux de souris (figure 26). A la
suite de ces observations, nous pouvons supposer que l'incapacité de réduire la
perméabilité transendothéliale des HUVEC par le milieu conditionné d'astrocytome peut-
être dû au fait que ces cellules cancéreuses présentent un état dédifférencié et qu'elles
n'expriment plus les facteurs nécessaires à l'induction du phénotype de BHE chez les CE.
Il serait donc intéressant de répéter les tests de perméabilité des HUVEC avec d'autres
cellules d'astrocytome et surtout avec un contrôle positif comme des astrocytes primaires
de fœtus de souris et des CE micros vasculaires cérébrales humaines pour valider les
résultats obtenus. Même si nous avons été incapables de démonter une diminution de la
perméabilité des HUVEC, nos résultats ne nous permettent pas de conclure qu'il n'est pas
possible de diminuer la perméabilité transendothéliale des HUVEC. Dans la mesure où
aucune composante de la BHE n'était présente dans ces expériences, il n'est pas surprenant
de ne pas avoir observé de diminution de la perméabilité transendothéliale. Le
développement d'un modèle in vitro de BHE humaine par génie tissulaire nécessitera fort
probablement la présence d'au moins un type cellulaire de la BHE native. Il est possible
qu'il soit plus efficace de mettre le modèle au point avec des cellules de rongeurs avant de
passer à des cellules humaines, et ce, en dépit des différences physiologiques interespèces
et du fait que les conditions établies avec des cellules de rongeurs devront être adaptées
pour les appliquer aux cellules humaines.

Conclusion
Notre objectif principal était d'évaluer la réponse des CE. ensemencées dans notre
modèle de la MFT. à la suite d'une exposition de 4 heures à un flux hémodynamique
physiologique. La réponse a été évalué d'une part par la morphologie des CE analysée par
microscopie et d'autres parts par les changements du niveau d'expression de gènes
impliqués dans la biologie des CE. La morphologie des CE soumises au flux est très
positive. L'adaptation du cytosquelette et de la morphologie cellulaire suppose que la MFT
constitue un substrat adéquat pour l'étude des CE en condition de flux hémodynamique.
Malgré que des tendances aient été perçues, nous n'avons pas observé de changements
significatifs des niveaux d'expression des gènes analysés. La grande variabilité de nos
résultats a rendu l'analyse de ces derniers problématique et plus ou moins pertinente.
Cependant, la poursuite des expériences et l'amélioration des techniques permettront sans
aucun doute d'en arriver à des conclusions plus solides. La répétition de l'expérience et
l'augmentation des échantillons sont les perspectives immédiates de ce travail.
Notre objectif secondaire était d'évaluer la possibilité de diminuer la perméabilité
tansendothéliale in vitro d'HUVEC cultivées dans un milieu conditionné d'astrocytome.
Nous n'avons pas été en mesure de provoquer cette diminution de la perméabilité des
HUVEC par la présence de cellules d'astrocytome. Le développement d'un modèle in vitro
de BHE humaine par génie tissulaire demandera une source de cellules possédant les
caractéristiques nécessaires à l'induction du phénotype de BHE. Nous en sommes au tout
premier pas du développement d'un tel modèle et les avenues pour y arriver restent à
explorer. Bien qu'il soit compréhensible de tenter d'utiliser des types cellulaires plus
facilement accessibles et plus susceptibles à la culture cellulaire, il ne s'agit peut-être pas
d'une solution possible. Il reste énormément de pain sur la planche pour en arriver à un
modèle humain in vitro, tridimensionnel et perfusé. Cependant, la contribution que pourrait
avoir un tel modèle à la recherche mérite d'y investir des efforts.
L'application du génie tissulaire au développement de modèles d'étude in vitro innovateurs
est certainement un outil de taille pour les chercheurs. Dans des contextes où la recherche

in vivo est ardue, voire même impossible, les modèles in vitro conçus par génie tissulaire
constituent une évolution par rapport à la culture cellulaire traditionnelle qui est souvent
loin des conditions physiologiques naturelles. Notre modèle de la MFT possède plusieurs
caractéristiques lui conférant un potentiel d'utilisation manifeste pour la recherche. Sa
composition en MEC endogène tridimensionnelle, la possibilité d'y faire circuler un flux
hémodynamique et sa relative simplicité font en sorte qu'il pourrait servir à plusieurs
champs d'études. Mise à part l'étude de la biologie des CE et des forces hémodynamiques,
on peut citer l'exemple de l'étude de l'artériosclérose, de l'extravasation des cellules
cancéreuses [65]. de l'angiogenèse. du remodelage vasculaire. de la pression artérielle, des
prothèses vasculaires artificielles, du génie tissulaire. de la BHE et bien d'autres champs
d'études impliquant les CE.

Annexe 1
Graphique de l'expression du gène GAPDH par PCR quantitative. Les colonnes représentent la valeur de cycle du seuil en nombre de cycles (Ct : cycle threshold). Ces valeurs sont inversement proportionnelles à la quantité d'ADN initial et l'incertitude sur la mesure est minimisée (<5%). Il n'y a pas de différence significative entre la condition statique et avec flux (4 h. 10 dynes/cm ) p>0,05. n = 3 pour chaque condition et mesures faites en duplicata.

Bibliographie
1. Farmer, C.G., Evolution of the vertebrate cardio-pulmonary System. Annu Rev Physiol, 1999. 61: p. 573-92.
2. Krogh, A., The number and distribution of capillaries in muscles with calculations of the oxygen pressure head necessary for supplying the tissue. J Physiol, 1919. 52(6): p. 409-15.
3. Schmidt-Nielsen, K. and P. Pennycuik, Capillary density in mammals in relation to body size and oxygen consumption. Am J Physiol, 1961. 200: p. 746-50.
4. Marieb, E.N., Anatomie et physiologie humaines. 2 ed. 1993, Saint-Laurent: Édition du renouveau pédagogique inc. 1014.
5. Bou-Gharios, G., et al., Extra-cellular matrix in vascular networks. Cell Prolif. 2004. 37(3): p. 207-20.
6. Eckert, R., et al.. Animal Physiology: Mechanisms and Adaptations. 4th ed. 1999. Paris: De Boeck Université. 822.
7. Loscalzo, J . , M.A. Creager, and V.J. Dzau, Vascular Medicine. lst ed. 1992, Boston: Little, Brown and Company. 1211.
8. Fox, S.I., Human Physiology. 4th ed. 1990, Burr Ridge: WM. C. Brown Publishers. 574.
9. Alberts, B., et al., Molecular Biology of the Cell 4th ed. 2002, New York: Garland Science. 1463.
10. Aird. W.C., Phenotypic heterogeneity of the endothelium: I. Structure, function, and mechanisms. Cire Res, 2007. 100(2): p. 158-73.
11. Janeway, C.A.. et al., Immunobiology: The Immune System in Health and Disease. 5th ed. 2001, New York: Garland Publishing. 782.
12. Folkman, J . and C. Haudenschild, Angiogenesis in vitro. Nature, 1980. 288(5791): p. 551-6.
13. Risau, W., Mechanisms of angiogenesis. Nature, 1997. 386(6626): p. 671-4. 14. Cohen, J.D., Overview of physiology, vascular biology, and mechanisms of
hypertension. J Manag Care Pharm, 2007. 13(5 Suppl): p. S6-8. 15. Ardaillou, R. and J .B . Michel, The relative rôles of circulating and tissue renin-
angiotensin Systems. Nephrol Dial Transplant, 1999. 14(2): p. 283-6. 16. Ardaillou, R., Active fragments of angiotensin II: enzymatic pathways of synthesis
and biological effects. Curr Opin Nephrol Hypertens, 1997. 6(1): p. 28-34. 17. Palmer, R.M., A.G. Ferrige, and S. Moncada, Nitric oxide release accounts for the
biological activity of endothelium-derived relaxingfactor. Nature, 1987. 327(6122): p. 524-6.
18. Yanagisawa, M., et al., A novelpotent vasoconstrictor peptide produced by vascular endothelial cells. Nature, 1988. 332(6163): p. 411-5.
19. Kohler, H.P. and P.J. Grant, Plasminogen-activator inhibitor type 1 and coronary artery disease. N Engl J Med, 2000. 342(24): p. 1792-801.
20. van den Eijnden-Schrauwen, Y., et al., Involvement of calcium and G proteins in the acute release of tissue-type plasminogen activator and von Willebrand factor from cultured human endothelial cells. Arterioscler Thromb Vase Biol, 1997. 17(10): p. 2177-87.

21. Muller. W.A. and GJ . Randolph. Migration ofleukocytes across endothelium and beyond: molécules involved in the transmigration and fate of monocytes. J Leukoc Biol, 1999. 66(5): p. 698-704.
22. Aird, W.C., Phenotypic heterogeneity of the endothelium: IL Représentative vascular beds. Cire Res, 2007. 100(2): p. 174-90.
23. Lehoux, S., Y. Castier, and A. Tedgui, Molecular mechanisms of the vascular responses to haemodynamic forces. J Intern Med. 2006. 259(4): p. 381-92.
24. Chi, J.T., et al.. Endothelial cell diversity revealed by global expression profiling. Proc Natl Acad Sci U S A. 2003. 100(19): p. 10623-8.
25. Nag, S., The Blood-Brain Barrier: Biology and Research Protocols. Methods in Molecular Medicine. ed. S. Nag. 2003. Totowa: Humana Press. 549.
26. Hawkins, B.T. and T.P. Davis, The blood-brain barrier/neurovascular unit in health and disease. Pharmacol Rev. 2005. 57(2): p. 173-85.
27. Daneman. R. and B.A. Barres, The blood-brain barrier—lessons from moody /lies. Cell, 2005. 123(1): p. 9-12.
28. Abbott, N.J., Dynamics of CNS Barriers: Evolution, Differentiation, and Modulation. Cell Mol Neurobiol, 2003. 25(1).
29. Cserr, H.F. and M. Bundgaard, Blood-brain interfaces in vertebrates: a comparative approach. Am J Physiol, 1984. 246(3 Pt 2): p. R277-88.
30. Liebner, S., et al.. Corrélation of tight junction morphology with the expression of tight junction proteins in blood-brain barrier endothelial cells. Eur J Cell Biol. 2000. 79(10): p. 707-17.
31. Kniesel, U. and H. Wolburg, Tight junctions of the blood-brain barrier. Cell Mol Neurobiol, 2000. 20(1): p. 57-76.
32. Balda, M.S. and K. Matter, Transmembrane proteins of tight junctions. Semin Cell Dev Bio l 2000. 11(4): p. 281-9.
33. Fanning, A.S., T.Y. Ma, and J.M. Anderson, Isolation andfunctional characterization of the actin binding région in the tight junction protein ZO-1. Faseb J , 2002. 16(13): p. 1835-7.
34. Wolburg, H. and A. Lippoldt, Tight junctions ofthe blood-brain barrier: development, composition and régulation. Vascul Pharmacol, 2002. 38(6): p. 323-37.
35. Maher, F., S.J. Vannucci, and LA. Simpson, Glucose transporter proteins in brain. Faseb J , 1994. 8(13): p. 1003-11.
36. Hawkins, R.A., et al., Structure of the blood-brain barrier and its rôle in the transport of amino acids. J Nutr, 2006. 136(1 Suppl): p. 218S-26S.
37. Schinkel, A.H. and J.W. Jonker, Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: an overview. Adv Drug Deliv Rev, 2003. 55(1): p. 3-29.
38. Schinkel, A.H., P-Glycoprotein, a gatekeeper in the blood-brain barrier. Adv Drug Deliv Rev, 1999. 36(2-3): p. 179-194.
39. el-Bacha, R.S. and A. Minn, Drug metabolizing enzymes in cerebrovascular endothelial cells afford a met abolie protection to the brain. Cell Mol Biol (Noisy-le-grand), 1999. 45(1): p. 15-23.
40. Davson, H. and W.H. Oldendorf, Symposium on membrane transport. Transport in the central nervous System. Proc R Soc Med. 1967. 60(4): p. 326-9.

41. Abbott. N.J., L. Ronnback. and E. Hansson, Asîrocyîe-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci, 2006. 7(1): p. 41-53.
42. Cecchelli. R.. et al.. Modelling of the blood-brain barrier in drug discovery and development. Nat Rev Drug Discov. 2007. 6(8): p. 650-61.
43. Haseloff, R.F., et al., In search of the astrocytic factor (s) modulating blood-brain barrier functions in brain capillary endothelial cells in vitro. Cell Mol Neurobiol. 2005. 25(1): p. 25-39.
44. Ramsauer, M., D. Krause. and R. Dermietzel. Angiogenesis of the blood-brain barrier in vitro and the function of cérébral pericytes. Faseb J . 2002. 16(10): p. 1274-6.
45. Zenker. D., et al.. Human blood-derived macrophages enhance barrier function of culturedprimary bovine and human brain capillary endothelial cells. J Physiol, 2003. 551(Pt 3): p. 1023-32.
46. Schiera, G., et al., Synergistic effects of neurons and astrocytes on the differentiation of brain capillary endothelial cells in culture. J Cell Mol Med. 2003. 7(2): p. 165-70.'
47. de Boer, A.G. and P.J. Gaillard. Blood-brain barrier dysfunction and recovery. J Neural Transm, 2006. 113(4): p. 455-62.
48. Prat, A. and J . Antel, Pathogenesis of multiple sclerosis. Curr Opin Neurol. 2005. 18(3): p. 225-30.
49. Deli. M.A., et al.. Permeability studies on in vitro blood-brain barrier models: physiology, pathology, andpharmacology. Cell Mol Neurobiol, 2005. 25(1 ): p. 59-127.
50. Berger. J.R. and M. Avison. The blood brain barrier in HIV infection. Front Biosci. 2004. 9: p. 2680-5.
51. Farkas, E. and P.G. Luiten, Cérébral microvascular pathology in aging and Alzheimer's disease. Prog Neurobiol, 2001. 64(6): p. 575-611.
52. Calabria. A.R. and E.V. Shusta, Blood-brain barrier genomics andproteomics: elucidatingphénotype, identifying disease targets and enabling brain drug delivery. Drug Discov Today, 2006. 11(17-18): p. 792-9.
53. Reichel, A., D.J. Begley, and N J . Abbott, An overview of in vitro techniques for blood-brain barrier studies. Methods Mol Med, 2003. 89: p. 307-24.
54. Pardridge, W.M., Molecular biology of the blood-brain barrier. Methods Mol Med, 2003. 89: p. 385-99.
55. Mattes, R.D., Fat taste and lipidmetabolism in humans. Physiol Behav, 2005. 86(5): p. 691-7.
56. Mineo, C , et al., Endothelial and antithrombotic actions of HDL. Cire Res, 2006. 98(11): p. 1352-64.
57. Stevens, R.J., et al., Inflammation and atherosclerosis in rheumatoid arthritis. Expert Rev Mol Med, 2005. 7(7): p. 1-24.
58. Raines, E.W. and N. Ferri, Thematic review séries: The immune System and atherogenesis. Cytokines affecting endothelial and smooth muscle cells in vascular disease. J Lipid Res, 2005. 46(6): p. 1081-92.
59. Yoshizumi, M., et al., Stress and vascular responses: atheroprotective effect of laminar fluid shear stress in endothelial cells: possible rôle of mitogen-activated protein kinases. J Pharmacol Sci, 2003. 91(3): p. 172-6.

60. Frangos, S.G., V. Gahtan. and B. Sumpio. Localization of atherosclerosis: rôle of hemodynamics. Arch Surg, 1999. 134(10): p. 1142-9.
61. Traub. O. and B.C. Berk, Laminar shear stress: mechanisms by which endothelial cells transduce an atheroprotective force. Arterioscler Thromb Vase Biol. 1998. 18(5): p. 677-85.
62. Folkman, J . , Fundamental concepts of the angiogenic process. Curr Mol Med. 2003. 3(7): p. 643-51.
63. Folkman, J . , Antiangiogenesis in cancer therapy—endostatin and its mechanisms of action. Exp Cell Res, 2006. 312(5): p. 594-607.
64. Gout, S., P.L. Tremblay, and J . Huot, Selectins and selectin ligands in extravasation of cancer cells and organ selectivity of metastasis. Clin Exp Metastasis. 2007.
65. Tremblay, P.L., F. A. Auger, and J . Huot, Régulation of tr ans endothelial migration of colon cancer cells by E-selectin-mediated activation ofp38 and ERK MAP kinases. Oncogene, 2006. 25(50): p. 6563-73.
66. Miles, F.L., et al., Stepping out of the flow: capillary extravasation in cancer metastasis. Clin Exp Metastasis, 2007.
67. Kobayashi, H., K.C. Boelte, and P.C. Lin, Endothelial cell adhésion molécules and cancer progression. Curr Med Chem. 2007. 14(4): p. 377-86.
68. Iiizumi, M., et al., Tumor-endothelial cell interactions: therapeuticpotential. Microvasc Res, 2007. 74(2-3): p. 114-20.
69. Sowers, J.R., Insulin résistance and hypertension. Am J Physiol Heart Cire Physiol. 2004. 286(5): p. H1597-602.
70. Kendall, D.M. and A.P. Harmel, The metabolic syndrome, type 2 diabètes, and cardiovascular disease: understanding the rôle of insulin résistance. Am J Manag Care, 2002. 8(20 Suppl): p. S635-53; quiz S654-7.
71. Gimbrone, M. A., Jr., et al., Endothelial dysfunction, hemodynamic forces, and atherogenesis. Ann N Y Acad Sci, 2000. 902: p. 230-9; discussion 239-40.
72. Nerem, R.M., Vascular fluid mechanics, the arterial wall, and atherosclerosis. J Biomech Eng, 1992. 114(3): p. 274-82.
73. Poiseuille, J . , Recherches expérimentales sur le mouvement des liquides dans les tubes de très petits diamètres. Crit. Rev. Acad. Sci., 1840. 11: p. 961-967.
74. Samijo, S.K., et al., Wall shear stress in the human common carotid artery as function of âge and gender. Cardiovasc Res, 1998. 39(2): p. 515-22.
75. Gnasso, A., et al., Association between intima-media thickness and wall shear stress in common carotid arteries in healthy maie subjects. Circulation, 1996. 94(12): p. 3257-62.
76. Kamiya, A., R. Bukhari, and T. Togawa, Adaptive régulation of wall shear stress optimizing vascular tree function. Bull Math Biol, 1984. 46(1): p. 127-37.
77. Pries, A.R., T.W. Secomb, and P. Gaehtgens, Design principles of vascular beds. Cire Res, 1995. 77(5): p. 1017-23.
78. Kassab, G.S. and Y.C. Fung, Thepattern of coronary arteriolar bifurcations and the uniform shear hypothesis. Ann Biomed Eng, 1995. 23(1): p. 13-20.
79. LaBarbera, M., Principles of design offluid transport Systems in zoology. Science, 1990. 249(4972): p. 992-1000.
80. Reneman, R.S., T. Arts, and A.P. Hoeks, Wall shear stress—an important déterminant of endothelial cell function and structure—in the arterial System in vivo. Discrepancies with theory. J Vase Res, 2006. 43(3): p. 251-69.

81. Helmke. B.P., Molecular control of cyioskeleîal mechanics by hemodynamic forces. Physiology (Bethesda), 2005. 20: p. 43-53.
82. Dimmeler. S., et al., Activation of nitric oxide synihase in endothelial cells by Akt-dependentphosphorylation. Nature. 1999. 399(6736): p. 601-5.
83. Li, Y.S., J.H. Haga, and S. Chien, Molecular basis of the effects of shear stress on vascular endothelial cells. J Biomech, 2005. 38(10): p. 1949-71.
84. Tronc, F., et al.. Rôle ofNO in flow-induced remodeling of the rabbit common carotidartery. Arterioscler Thromb Vase Biol. 1996. 16(10): p. 1256-62.
85. Glagov, S., Intimai hyperplasia. vascular modeling, and the restenosis problem. Circulation, 1994. 89(6): p. 2888-91.
86. Davies, P.F., J.A. Spaan, and R. Krams, Shear stress biology of the endothelium. Ann Biomed Eng, 2005. 33(12): p. 1714-8.
87. Gimbrone, M.A., Jr., et al., Spécial communieationthe critical rôle of mechanical forces in blood vessel development, physiology andpathology. J Vase Surg, 1999. 29(6): p. 1104-51.
88. Orr. A.W.. et al., Mechanisms of mechanotransduction. Dev Cell, 2006. 10(1): p. 11-20.
89. Ernstrom, G.G. and M. Chalfie, Genetics of sensory mechanotransduction. Annu Rev Genêt. 2002. 36: p. 411-53.
90. Gillespie, P.G. and R.G. Walker, Molecular basis of me chano sensory transduction. Nature, 2001. 413(6852): p. 194-202.
91. Tarbell. J.M.. S. Weinbaum. and R.D. Kamm, Cellular fluid mechanics and mechanotransduction. Ann Biomed Eng, 2005. 33(12): p. 1719-23.
92. Jalali, S., et al., Shear stress activâtesp60src-Ras-MAPK signalingpathways in vascular endothelial cells. Arterioscler Thromb Vase Biol. 1998. 18(2): p. 227-34.
93. Davies. P.F., Flow-mediated endothelial mechanotransduction. Physiol Rev. 1995. 75(3): p. 519-60.
94. Lodish. H., et al.. Molecular Cell Biology. 4th ed. 2001. New York: W.H. Freeman and Company. 1084.
95. Sun, H.W., et al., Involvement of integrins, MAPK, and NF-kappaB in régulation of the shear stress-induced MMP-9 expression in endothelial cells. Biochem Biophys Res Commun, 2007. 353(1): p. 152-8.
96. Jalali, S., et al., Integrin-mediated mechanotransduction requires its dynamic interaction with spécifie extracellular matrix (ECM) ligands. Proc Natl Acad Sci U S A, 2001. 98(3): p. 1042-6.
97. Hoger, J.H., et al., Shear stress régulâtes the endothelial Kir2.1 ion channel. Proc Natl Acad Sci U S A , 2002. 99(11): p. 7780-5.
98. Hayashi, Y., et al., Induction of various blood-brain barrier properties in non-neural endothelial cells by close apposition to co-cultured astrocytes. Glia, 1997. 19(1): p. 13-26.
99. Gudi, S.R., C.B. Clark, and J.A. Frangos, Fluid flow rapidly activâtes G proteins in human endothelial cells. Involvement of G proteins in mechanochemical signal transduction. Cire Res, 1996. 79(4): p. 834-9.
100. Li, B. , et al., Involvement of Rho/ROCK signalling in small cell lung cancer migration through human brain microvascular endothelial cells. FEBS Lett, 2006. 580(17): p. 4252-60.

101. Haidekker, M.A., N. L'Heureux, and J.A. Frangos, Fluid shear stress increases membrane fluidity in endothelial cells: a study with DCVJ fluorescence. Am J Physiol Heart Cire Physiol, 2000. 278(4): p. H1401-6.
102. Butler, P.J., et al.. Rate sensitivity of shear-induced changes in the latéral diffusion of endothelial cell membrane lipids: a rôle for membrane perturbation in shear-induced MAPK activation. Faseb J . 2002. 16(2): p. 216-8.
103. Luft, J.H., Fine structures of capillary and endocapillary layer as revealed by ruthénium red. Fed Proc. 1966. 25(6): p. 1773-83.
104. Weinbaum, S., et al., Mechanotransduction and flow across the endothelial glycocalyx. Proc Natl Acad Sci U S A , 2003. 100(13): p. 7988-95.
105. Fujiwara, K.. et al., Is PECAM-J a mechanoresponsive molécule? Cell Struct Funct. 2001. 26(1): p. 11-7.
106. Tzima, E., et al., A mechanosensory complex that médiates the endothelial cell response to fluid shear stress. Nature, 2005. 437(7057): p. 426-31.
107. Silacci, P., et al., Flow pulsatility is a critical déterminant of oxidative stress in endothelial cells. Hypertension, 2001. 38(5): p. 1162-6.
108. Ni, C.W., et al., Activation of PKC-epsilon and ERK1/2 participâtes in shear-induced endothelial MCP-1 expression that is repressed by nitric oxide. J Cell Physiol, 2003. 195(3): p. 428-34.
109. Tzima, E., et al., Activation of integrins in endothelial cells by fluid shear stress médiates Rho-dependent cytoskeletal alignment. Embo J , 2001. 20(17): p. 4639-47.
110. Go, Y.M., et al., Phosphatidylinositol 3-kinase gamma médiates shear stress-dependent activation ofJNK in endothelial cells. Am J Physiol. 1998. 275(5 Pt 2): p. H1898-904.
111. McCormick, S.M., et al., DNA microarray reveals changes in gene expression of shear stressed human umbilical vein endothelial cells. Proc Natl Acad Sci U S A , 2001. 98(16): p. 8955-60.
112. Garcia-Cardena, G., et al., Mechanosensitive endothelial gene expression profiles: scripts for the rôle of hemodynamics in atherogenesis? Ann N Y Acad Sci, 2001. 947: p. 1-6.
113. Resnick, N., et al., Endothelial gene régulation by laminar shear stress. Adv Exp Med Biol, 1997. 430: p. 155-64.
114. Wasserman, S. M., et al., Gene expression profile of human endothelial cells exposed to sustainedfluid shear stress. Physiol Genomics, 2002. 12(1): p. 13-23.
115. Hay, D.C., et al., Activation of NF-kappaB nuclear transcription factor by flow in human endothelial cells. Biochim Biophys Acta, 2003. 1642(1-2): p. 33-44.
116. Brooks, A.R., P.I. Lelkes, and G.M. Rubanyi, Gene expression profiling of human aortic endothelial cells exposed to disturbed flow and steady laminar flow. Physiol Genomics, 2002. 9(1): p. 27-41.
117. Fisher, A.B., et al., Endothelial cellular response to altered shear stress. Am J Physiol Lung Cell Mol Physiol, 2001. 281(3): p. L529-33.
118. Fernandez, P., et al., Gene Response in Endothelial Cells Cultured on Engineered Surfaces Is Regulated by Shear Stress. Tissue Eng, 2007.
119. Vara, D. S., et al., The effect of shear stress on human endothelial cells seeded on cylindrical viscoelastic conduits: an investigation of gene expression. Biotechnol Appl Biochem, 2006. 45(Pt 3): p. 119-30.

120. Wasserman, S.M. and J.N. Topper, Adaptation of the endothelium to fluid flow: in vitro analyses of gene expression and in vivo implications. Vase Med. 2004. 9(1): p. 35-45.
121. Angst. B.D.. C. Marcozzi, and A.I. Magee, The cadherin superfamily: diversity in form and function. J Cell Sci, 2001. 114(Pt 4): p. 629-41.
122. Gomes, N., C. Legrand, and F. Fauvel-Lafeve, Shear stress induced release of von Willebrand factor and thrombospondin-1 in HUVEC extracellular matrix enhances breast tumour cell adhésion. Clin Exp Metastasis. 2005. 22(3): p. 215-23.
123. Sun, R.J., et al.. Régulation of von willebrand factor of human endothelial cells exposed to laminar flows: an in vitro study. Clin Hemorheol Microcirc, 2000. 23(1): p. 1-11.
124. Cesarman-Maus, G. and K.A. Hajjar, Molecular mechanisms of fibrinolysis. Br J Haematol. 2005. 129(3): p. 307-21.
125. Topper. J.N.. et al.. Identification of vascular endothelial gènes differentially responsive to fluid mechanical stimuli: cyclooxygenase-2, manganèse superoxide dismutase, and endothelial cell nitric oxide synthase are selectively up-regulated by steady laminar shear stress. Proc Natl Acad Sci U S A . 1996. 93(19): p. 10417-22.
126. Diamond, S.L., et al., Tissue plasminogen activator messenger RNA levels increase in cultured human endothelial cells exposed to laminar shear stress. J Cell Physiol, 1990. 143(2): p. 364-71.
127. Binder, B.R., et al., Plasminogen activator inhibitor 1: physiological and pathophysiological rôles. News Physiol Sci, 2002. 17: p. 56-61.
128. Kawai, Y., et al., Hemodynamic forces modulate the effects of cytokines on fibrinolytic activity of endothelial cells. Blood, 1996. 87(6): p. 2314-21.
129. Wu, K.K., Régulation of endothelial nitric oxide synthase activity and gene expression. Ann N Y Acad Sci, 2002. 962: p. 122-30.
130. Ranjan, V., Z. Xiao, and S.L. Diamond, Constitutive NOS expression in cultured endothelial cells is elevated by fluid shear stress. Am J Physiol, 1995. 269(2 Pt 2): p. H550-5.
131. Prasad, C.K., K. Jayakumar, and L.K. Krishnan, Phénotype gradation of human saphenous vein endothelial cells from cardiovascular disease subjects. Endothelium, 2006. 13(5): p. 341-52.
132. Arai, H., et al., Cloning and expression of a cDNA encoding an endothelin receptor. Nature, 1990. 348(6303): p. 730-2.
133. Schneider, M.P., E.I. Boesen, and D.M. Pollock, Contrasting actions of endothelin ET (A) and ET(B) receptor s in cardiovascular disease. Annu Rev Pharmacol Toxicol, 2007. 47: p. 731-59.
134. Morawietz, H., et al., Régulation of the endothelin System by shear stress in human endothelial cells. J Physiol, 2000. 525 Pt 3: p. 761-70.
135. Nerem, R.M., M.J. Levesque, and J .F. Cornhill, Vascular endothelial morphology as an indicator of the pattern of blood flow. J Biomech Eng, 1981. 103(3): p. 172-6.
136. Galbraith, C.G., R. Skalak, and S. Chien, Shear stress induces spatial reorganization of the endothelial cell cytoskeleton. Cell Motil Cytoskeleton, 1998. 40(4): p. 317-30.
137. Franke, R.P., et al., Induction of human vascular endothelial stress fibres by fluid shear stress. Nature, 1984. 307(5952): p. 648-9.

138. Helmke, B.P., R.D. Goldman, and P.F. Davies. Rapiddisplacement of vimentin intermediate filaments in living endothelial cells exposed to flow. Cire Res, 2000. 86(7): p. 745-52.
139. Noria, S., et al., Transient and steady-state effects of shear stress on endothelial cell adherens junctions. Cire Res. 1999. 85(6): p. 504-14.
140. Sill, H.W., et al., Shear stress increases hydraulic conductivity of cultured endothelial monolayers. Am J Physiol. 1995. 268(2 Pt 2): p. H535-43.
141. Barbee, K.A., et al.. Suhcellular distribution of shear stress at the surface offlow-aligned and nonaligned endothelial monolayers. Am J Physiol, 1995. 268(4 Pt 2): p. Hl765-72.
142. Levesque, M.J., R.M. Nerem, and E.A. Sprague, Vascular endothelial cell prolifération in culture and the influence of flow. Biomaterials. 1990. 11(9): p. 702-7.
143. Akimoto. S., et al., Laminar shear stress inhibits vascular endothelial cell prolifération by inducing cyclin-dependent kinase inhibitor p21(Sdil/Cipl/Wafl). Cire Res, 2000. 86(2): p. 185-90.
144. Garberg, P., et al., In vitro models for the blood-brain barrier. Toxicol In Vitro, 2005. 19(3): p. 299-334.
145. Urbich, C., et al., Laminar shear stress upregulates integrin expression: rôle in endothelial cell adhésion and apoptosis. Cire Res, 2000. 87(8): p. 683-9.
146. Dimmeler, S., et al., Upregulation of super oxide dismutase and nitric oxide synthase médiates the apoptosis-suppressive effects of shear stress on endothelial cells. Arterioscler Thromb Vase Biol, 1999. 19(3): p. 656-64.
147. Dimmeler, S., et al., Fluid shear stress stimulâtes phosphorylation of Akt in human endothelial cells: involvement in suppression of apoptosis. Cire Res, 1998. 83(3): p. 334-41.
148. Tardy, Y., et al., Shear stress gradients remodel endothelial monolayers in vitro via a cellproliferation-migration-loss cycle. Arterioscler Thromb Vase Biol, 1997. 17(11): p. 3102-6.
149. Sprague, E.A., J . Luo, and J.C. Palmaz, Human aortic endothelial cell migration onto stent surfaces under static and flow conditions. J Vase Interv Radiol, 1997. 8(1 Pt l ) : p . 83-92.
150. Wu, M.H., et al., Effect of differential shear stress onplatelet aggregation, surface thrombosis, and endothelialization of bilatéral carotid-fémoral grafts in the do g. J Vase Surg, 1995. 22(4): p. 382-90; discussion 390-2.
151. Gilmore, A.P. and L.H. Romer, Inhibition of focal adhésion kinase (FAK) signaling in focal adhésions decreases cell motility and prolifération. Mol Biol Cell, 1996. 7(8): p. 1209-24.
152. Hsu, P.P., et al., Effects of flow patterns on endothelial cell migration into a zone of mechanical denudation. Biochem Biophys Res Commun, 2001. 285(3): p. 751-9.
153. Weinbaum, S., et al., Effect of cell turnover and leaky junctions on arterial macromole cul ar transport. Am J Physiol, 1985. 248(6 Pt 2): p. H945-60.
154. Chuang, P.T., et al., Macromolecular transport across arterial and venous endothelium in rats. Studies with Evans blue-albumin and horseradish peroxidase. Arteriosclerosis, 1990. 10(2): p. 188-97.

155. Herrmann. R.A., R.A. Malinauskas. and G.A. Truskey, Char acier ization of sites with elevated LDL permeability at intercostal, celiac, and iliac branches of the normal rabbit aorta. Arterioscler Thromb, 1994. 14(2): p. 313-23.
156. Krizanac-Bengez, L., et al., Loss of'Flow Induces Leukocyte-mediated MMP/TIMP Imbalance in Dynamic in vitro Blood-Brain Barrier Model: Rôle of Pro-inflammatory Cytokines. Am J Physiol Cell Physiol, 2006.
157. Krizanac-Bengez, L., et al., Loss of shear stress induces leukocyte-mediated cytokine release and blood-brain barrier failure in dynamic in vitro blood-brain barrier model. J Cell Physiol 2006. 206(1): p. 68-77.
158. Santaguida, S., et a l , Side by side comparison between dynamic versus static models of blood-brain barrier in vitro: A permeability study. Brain Res. 2006.
159. Busse, R. and I. Fleming, Vascular endothelium and blood flow. Handb Exp Pharmacol, 2006(176 Pt 2): p. 43-78.
160. Sidi, S. and F.M. Rosa, [Mechanotransduction of hemodynamic forces régulâtes organogenesis]. Med Sci (Paris), 2004. 20(5): p. 557-6 L
161. Khetani, S.R. and S.N. Bhatia, Engineering tissues for in vitro applications. Curr Opin Biotechnol, 2006. 17(5): p. 524-31.
162. Stoclet, J.C., et a l . [Human blood vessels created with tissue engineering]. Med Sci (Paris), 2004. 20(6-7): p. 675-8.
163. Moulin. V., et a l , [Tissue engineering: a tool to under stand the physiological mechanisms]. Med Sci (Paris), 2003. 19(10): p. 1003-10.
164. L'Heureux, N., et a l , A completely biological tissue-engineered human blood vessel. Faseb J , 1998. 12(1): p. 47-56.
165. Remy-Zolghadri, M., et a l , Endothelium properties of a tissue-engineered blood vessel for small-diameter vascular reconstruction. J Vase Surg, 2004. 39(3): p. 613-20.
166. Auger, F.A., P. D'Orleans-Juste, and L. Germain, Adventitia contribution to vascular contraction: hints provided by tissue-engineered substitutes. Cardiovasc Res, 2007. 75(4): p. 669-78.
167. Laflamme, K., et a l , Tissue-engineered human vascular média produced in vitro by the self-assembly approach présent functional properties similar to those of their native blood vessels. Tissue Eng, 2006. 12(8): p. 2275-81.
168. Laflamme, K., et a l , Adventitia contribution in vascular tone: insights from adventitia-derived cells in a tissue-engineered human blood vessel. Faseb J , 2006. 20(8): p. 1245-7.
169. Helderman, F., et a l , Effect of shear stress on vascular inflammation and plaque development. Curr Opin Lipidol, 2007. 18(5): p. 527-33.
170. Chatzizisis, Y.S., et a l , Rôle of endothelial shear stress in the natural history of coronary atherosclerosis and vascular remodeling: molecular, cellular, and vascular behavior. J Am Coll Cardiol, 2007. 49(25): p. 2379-93.
171. Li, S., N.F. Huang, and S. Hsu, Mechanotransduction in endothelial cell migration. J Cell Biochem, 2005. 96(6): p. 1110-26.
172. Haga, J.H., Y. S. Li, and S. Chien, Molecular basis of the effects of mechanical stretch on vascular smooth muscle cells. J Biomech, 2007. 40(5): p. 947-60.
173. Fernandez, P., et a l , Endothelial cells cultured on engineered vascular grafts are able to transduce shear stress. Tissue Eng, 2006. 12(1): p. 1-7.

174. Bockhorn, M., R.K. Jain, and L.L. Munn, Active versus passive mechanisms in metastasis: do cancer cells crawl into vessels, or are they pushed? Lancet Oncol. 2007. 8(5): p. 444-8.
175. Krizanac-Bengez, L., M.R. Mayberg, and D. Janigro, The cérébral vasculature as a therapeutic target for neurological disorders and the rôle of shear stress in vascular homeostatis andpathophysiology. Neurol Res. 2004. 26(8): p. 846-53.
176. Hurst, R.D. and I.B. Fritz, Properties of an immortalised vascular endothelial/glioma cell co-culture model of the blood-brain barrier. J Cell Physiol, 1996. 167(1): p. 81-8.
177. Scism, J.L., et al.. Evaluation of an in vitro coculture model for the blood-brain barrier: comparison ofhuman umbilical vein endothelial cells (ECV304) and rat glioma cells (C6) from two commercial sources. In Vitro Cell Dev Biol Anim. 1999. 35(10): p. 580-92.
178. Prat, A., et al., Glial cell influence on the human blood-brain barrier. Glia, 2001. 36(2): p. 145-55.
179. Auger. F.A., et al., Skin équivalentproduced with human collagen. In Vitro Cell Dev Biol Anim, 1995. 31(6): p. 432-9.
180. Dussault, A.A. and M. Pouliot, Rapid and simple comparison of messenger RNA levels using real-time PCR. Biol Proced Online, 2006. 8: p. 1-10.
181. Radonic, A., et a l , Guideline to référence gene sélection for quantitative real-time PCR. Biochem Biophys Res Commun, 2004. 313(4): p. 856-62.
182. Esper, R.J., et al., Endothelial dysfunction: a comprehensive appraisal. Cardiovasc Diabetol, 2006. 5: p. 4.
183. Neuhaus, W., et al., A novel flow based hollow-fiber blood-brain barrier in vitro model with immortalised cell line PBMEC/C1-2. J Biotechnol, 2006.
184. Boveri, M., et al., Induction of blood-brain barrier properties in cultured brain capillary endothelial cells: comparison between primary glial cells and C6 cell line. Glia, 2005. 51(3): p. 187-98.




















