
The ultrastructure of transplanted rabbit retinal epithelium
Upload
independentCategory
view
7download
0
Field Crops Research, 26 ( 1991 ) 327-345 327 Elsevier Science Publishers B.V., Amsterdam
Relationships between ripening-phase productivity and crop duration, canopy
photosynthesis and senescence in transplanted and direct-seeded lowland rice
M. Dingkuhn a, H.F. Schnier a, S.K. De Datta a'~, K. Dorffling b and C. Javellana a aInternational Rice Research Institute (IRRI), P.O. Box 933, Manila, The Philippines bInstitut fuer Allgemeine Botanik, Ohnhorststrasse 18, D-2000 Hamburg 52, Germany
(Accepted 23 April 1990)
ABSTRACT
Dingkuhn, M., Schnier, H.F., De Datta, S.K., Dorffling, K. and Javellana, C., 1991. Relationships between ripening-phase productivity and crop duration canopy photosynthesis and senescence in transplanted and direct-seeded lowland rice. Field Crops Res., 26: 327-345.
A field experiment was conducted to analyze the effect of crop duration on growth and the yield- formation process in tropical lowland rice. Three semidwarf rices (IR58, IR64, and IR29723-143-3- 2-1 ) of different growth durations were either transplanted or broadcast-seeded, and grown under different nitrogen fertilizer regimes. Dry-matter accumulation and N concentration of various plant organs, leaf-area index (L), tissue death, tiller number, plant height, and canopy photosynthesis were recorded periodically. Crop duration increased plant biomass but did not significantly affect grain- yield. Plant N uptake was not affected by crop duration. Direct seeding gave grain yields superior to those of transplanted rice using short-duration IR58, hut equal or lower grain-yield using the medium- and long-duration varieties. Growth and development were delayed, and tillering and foliage growth were reduced in transplanted rice due to transplant shock. Tiller production and abortion were a function of relative growth rate. Tissue death occurred during the reproductive phase and depended on L and foliage N concentration. IR58 had the highest L (up to I 1.6) and IR64 the lowest. Direct- seeded rice had higher L and dry-matter during vegetative and reproductive growth, but lower foliage N concentration, than had transplanted rice. Growth during ripening was negatively affected by high biomass and L at flowering but enhanced by high foliage N concentration. It is concluded that grain- yield is impeded by high biomass at flowering in direct-seeded rice, particularly in long-duration va- rieties. High biomass at flowering, however, may enable further yield improvement ifN partitioning and foliage expansion patterns are modified.
INTRODUCTION
The growth durat ion of a rice crop can vary between 90 and 200 days, de- pending on temperature, photoperiod, cultural factors and cultivar. A farm-
IAuthor to whom correspondence should be addressed.
0378-4290/91/$03.50 © 1991 - - Elsevier Science Publishers B.V.

328 M. DINGKUHN ET AL.
er's choice of a long- or short-duration cultivar depends on the cropping pat- tern of his choice, the seasonal characteristics of the location, and the grain- yield he can expect. Grain-yield potential, however, is not a simple function of growth duration. Even though a long-duration crop can harvest more cu- mulative solar radiation and can extract soil nutrients over a longer period, the highest tropical rice grain-yields (9-11 t ha- ~ ) have been achieved with short- to medium-duration cultivars (115-125 days) such as IR8 or IR64 (Yoshida, 1983).
Long-duration cultivars are often superior to short-duration rices if soil fer- tility is low (Kawano and Tanaka, 1968). Short-duration rices can give high grain-yields if well-fertilized and direct-seeded or densely planted (Yoshida, 1981 ). The shorter the growth duration, the more vegetative growth limits yield. Transplanting restricts vegetative growth through transplanting shock. Direct-seeding, therefore, enables the production of more tillers and higher plant biomass (Dingkuhn et al., 1990a; Schnier et al., 1990a).
Whether high biomass production can translate into high grain-yield de- pends on numerous factors, including cultural practices, climate and geno- type (Dingkuhn et al., 1990a,b). The present study compares growth patterns and yield of three high-yielding rices that are of different growth duration but of similar plant type (high-tillering, erect indica semidwarfs). Emphasis is given to effects of growth duration and crop establishment technique on dry- matter accumulation, and its conversion into grain-yield.
M A T E R I A L S A N D M E T H O D S
A field experiment was conducted during the 1988 dry season (January to June) at the Maligaya Rice Research and Training Center, Mufioz, Nueva Ecija, The Philippines. The soil was a Vertic Tropaquept with pH 5.9, 14 g organic C kg- ~, 1.1 g total N kg- 1, fEE 38 cmol kg- 1, 45 g clay kg- 1, and 5 g sand kg- ~. The experimental design was a two-factorial randomized complete block with three rice varieties and six treatments (combinations of planting method and N application; Table 2), replicated four times. Plot size was 35 m 2"
Short-statured IR58 (short duration, 95-105 days), IR64 (medium dura- tion, 110-120 days), and IR29723-143-3-2-1 (long duration, 130-140 days) were either transplanted as 20-day-old seedlings at 4 plants hill- ~ and at a hill spacing of 0.2 m × 0.2 m (TP), or pregerminated seeds were broadcast seeded by hand at 90 kg dry seed ha- 1. Broadcast seeded plots were either left unal- tered (BS), or rows 0.25 m apart were established 10 days after seeding (DAS) using a rake with 0.125-m-wide prongs (BSR).
Phosphorus (26 kg P ha- l ) as single superphosphate, potassium (75 kg K ha- ~ ) as KC1, and zinc (4.5 kg Zn ha- ~ ) as zinc sulfate heptahydrate were broadcast and incorporated before seeding or transplanting. Nitrogen was

GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 329
either not applied (control treatments) or applied at 90 kg N ha-~ as urea using different methods:
- In BS and one TP treatment, 60 kg N ha-~ as prilled urea was basaUy broadcast with subsequent incorporation (B&I) before transplanting or broadcast seeding. Another 30 kg N ha -~ was broadcast (2 /3 -1 /3 split) in the floodwater at 5-7 days before panicle initiation (PI).
- In BSR and one TP treatment, liquid urea solution was band-placed using a two-row, push-type injector (Schnier et al., 1990a). The first N application of 30 kg ha-L was at 14 days after transplanting (DAT) in TP and at 20 DAS in BSR treatments, followed by 60 kg N ha-~ ( 1 /3-2/3 split) applied at 5-7 days before PI.
The plots were flooded one day after transplanting and 5 days after broad- cast-seeding, and a water depth of 0.05 m was kept throughout the growth period. The subplots were individually drained two weeks before maturity. Weed control was by hand-weeding, and insect control was optimum, follow- ing standard recommendations.
Destructive plant sampling was conducted weekly between 08:00 and 10:00 h during the entire growth period to determine dry-matter (stems and leaf sheaths, leaf blades, panicles, and dead material separately), tiller number, leaf area index (L), plant height, total N uptake, and N concentration (stems and leaf sheaths, leaf blades, and panicles separately). Plant sample size was 0.24 m 2 (equivalent to six hills in TP ) per subplot during the vegetative growth stage and 0.16 m-2 (four hills in TP) per subplot during the reproductive growth stage. Samples were collected at a minimum distance of 0.8 m from the plot border to avoid border effects. Leaf area was measured with a leaf- area meter (Hayashi Denkoh, Tokyo, Japan). Tissue dry-matter was deter- mined after oven-drying at 70°C until constant weight was observed. Tissue N was determined by the Kjeldahl method. At maturity, grain and straw from a 7.2-m 2 sample area at the center of the plots were harvested and analyzed separately. Grain-yields were expressed at 14% moisture.
Analysis of yield components was done on plant samples taken from a 0.32- m 2 area per subplot. Harvest index (HI) was determined from the grain:straw ratio of the harvest area (7.2 m 2 ). Canopy CO2 exchange was measured weekly with a mobile gas-exchange system (Walz, Effeltrich, Germany), using a technique similar to that described by Morgan (1988). Transparent mylar chambers (0.6 m × 0 . 6 m × 1.0 m or 0.5 m × 1.0 mX 1.2 m) were placed on top of the canopy for less than 1 min while a continuously flowing air sample was collected through a 20-m teflon tubing. Air concentrations of CO2 and H20 were analyzed with a BINOS-1 (Heraeus, Germany) infrared gas ana- lyzer. Gas mixing and wind generation in the chamber were achieved using two fans located above the canopy. The rate of CO2 concentration change in the chamber served as a measure of the photosynthetic rate after it was cor- rected through separate measurements for floodwater CO2 exchange. Canopy

330 M. DINGKUHN ET AL.
C02 assimilation rates measured before noon (10:00-12:00) under cloudless conditions were termed Pn .... indicating that they represented the daily max- imum (diurnal data not presented).
Meteorological data (relative humidity, photosynthetically active radia- tion (Qpa), and temperature ) were recorded simultaneously with the gas ex- change measurements, and supplemented by daily weather data recorded in a meteorological station 300 m from the experimental site.
RESULTS A N D DISCUSSION
Crop duration and plant height
Cultivars IR58, IR64, and IR29723-143-3-2-1 differed markedly in crop duration and plant height but were similar in plant type (high-tillering sem- idwarf with erect leaves). In the transplanted treatments with N fertilization, crop duration was 105 days in IR58, 117 days in IR64, and 140 days in IR29723-143-3-2-1, including 20 days of seedbed cultivation. In the direct- seeded treatments, however, crop duration was 8-12 days shorter regardless of variety, due to the absence of a transplanting shock (Anonymous, 1987; Dingkuhn et al., 1990a). Control plants matured 2-4 days earlier than N- fertilized plants.
Differences in crop duration were due to varying duration of the vegetative growth phase, while the reproductive and ripening phases each lasted about 28 days. In a given environment, varietal differences in crop duration usually depend on the length of the vegetative growth phase (Vergara and Chang, 1985 ). Plant height increased almost linearly until shortly after flowering and stabilized thereafter (data not presented). The three cultivars did not differ significantly in plant height at a given crop age. Plant height at maturity
TABLE 1
Plant height (cm) at maturity
Trt. N applied Cultivar LSD0.o5 (kg ha-1 )
IR58 IR64 IR29723- 143-3-2-1
TP 0 68 79 89 7 90, B&I a 80 92 103 4 90, BW 87 88 109 5
BS 0 56 64 74 10 90, B&I 73 86 97 6
BSR 90, BP 85 91 105 3 LSDo.o5 6 5 5
aB&I, Broadcast & incorporated; BP, band-placed.
GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 331
( H (cm); Table 1 ) was significantly correlated with crop duration ( CD, days ) (linear regressions of pooled treatment means):
N-fertilized: H = 22.0 + 0.60 CD r= 0.9 l**
Controls: H = 6.43 + 0.57 CD r= 0.866*
Consequently, the varietal differences in growth duration led to differences in plant height at maturity. Nitrogen fertilization increased plant height sig- nificantly. The late split (1/3-2/3, band-placed) resulted in higher plants than did the 2/3-1/3 (B&I) split.
Plant height during ripening is an important factor for lodging, which rep- resents a major constraint to yields of direct-seeded rice in the tropics.
Grain-yield, harvest index, and agronomic efficiency
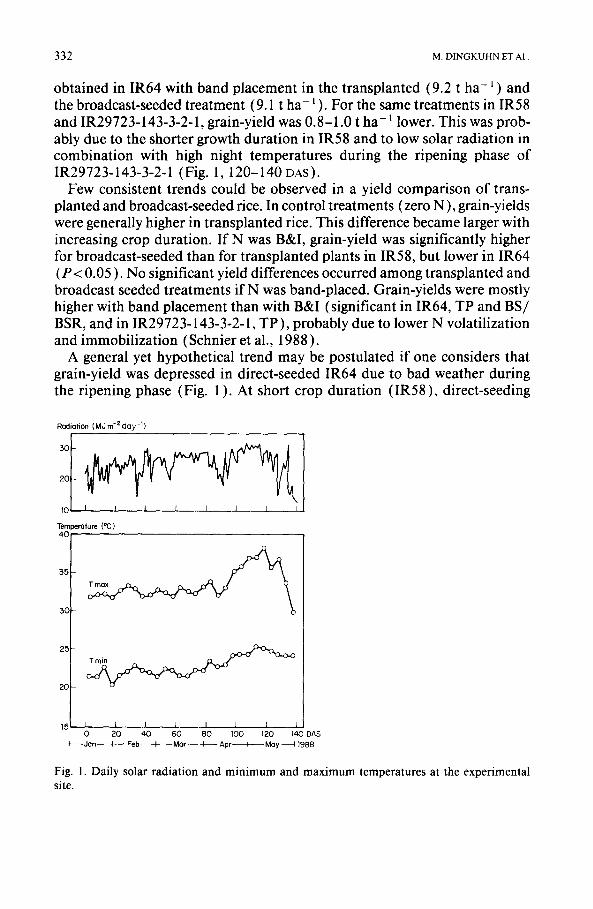
Grain-yield (Table 2 ) was significantly affected by N fertilization but dif- fered less among cultivars and planting techniques. Highest grain-yields were
TABLE2
Grain-yield (GV) at 14% moisture (t h a - t ) ; agronomic efficiency (AE) (kg grain/kg N appl.); har- vest index (HI); straw (NS), grain (NG), and total N content (TN) at maturity (kg ha - ~ ); and nitro- gen harvest index (Nm) (means of four replications)
Treatment GY AE HI NS NG TN NHI
I R 5 8
TP no fertilizer N 4.9 - 0.562 21.6 44.7 66.3 0.67 TP B&I, 2 / 3 - 1 / 3 split 8.0 35 0.575 31.4 81.1 112.5 0.72 TP band-placed, 1 / 3 - 2 / 3 8.1 36 0.571 37.5 97.0 134.5 0.72 BS no fertilizer N 4.7 - 0.525 20.1 35.3 55.4 0.64 BS B&I, 2 / 3 - 1 / 3 split 8.5 43 0.556 32~6 78.6 111.2 0.71 BSR band-placed, 1 / 3 - 2 / 3 8.3 40 0.564 41.0 107.9 148.9 0.72 I R 6 4
TP no fertilizer N 5.5 - 0.538 20.8 48.6 69.4 0.70 TP B&I, 2 / 3 - 1 / 3 split 8.8 37 0.513 43.7 93.4 137.1 0.68 TP band-placed, 1 / 3 - 2 / 3 9.2 42 0.502 59.3 107.6 166.9 0.64 BS no fertilizer N 4.3 - 0.493 18.7 34.6 53.3 0.65 BS B&I, 2 / 3 - 1 / 3 split 8.0 42 0.516 36.2 81.4 117.6 0.69 BSR band-placed, 1 / 3 - 2 / 3 9.1 54 0.535 49.2 104.0 153.2 0.68 IR29723-143-3-2-1 TP no fertilizer N 4.8 - 0.505 16.0 40.3 56.3 0.72 TP B&I, 2 / 3 - 1 / 3 split 8.0 36 0.456 36.8 75.3 112.1 0.67 TP band-placed, 1 / 3 - 2 / 3 8.4 40 0.427 60.4 91.1 151.5 0.60 BS no fertilizer N 3.8 - 0.390 17.0 30.8 47.8 0.64 BS B&I, 2 / 3 - 1 / 3 split 8.0 47 0.437 37.9 74.5 112.4 0.64 BSR band-placed, 1 / 3 - 2 / 3 8.2 50 0.448 41.7 85.9 127.6 0.67
LSD0.o5 0.4 - 0.020 8.7 9.4 14.2 -
332 M. DINGKUHN ET AL.
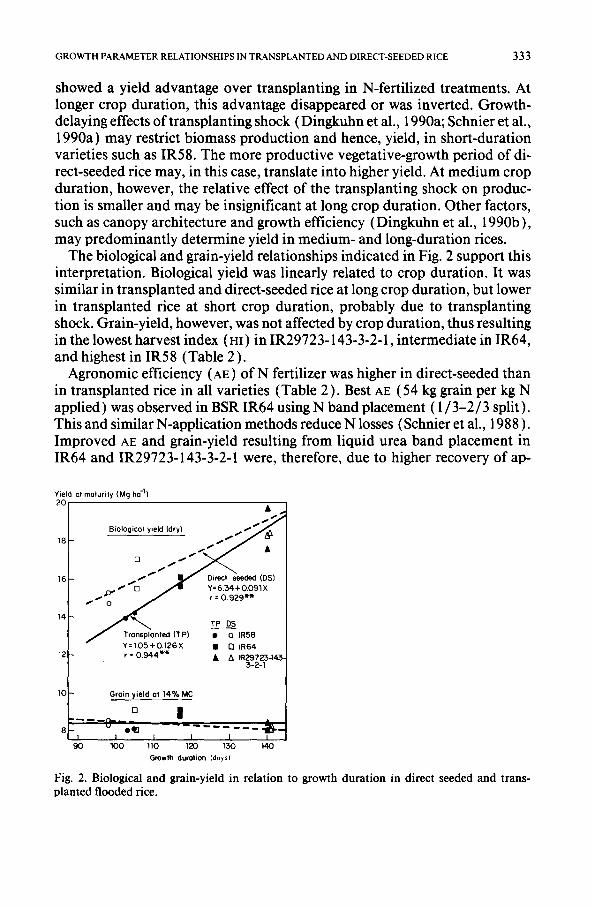
obtained in IR64 with band placement in the transplanted (9.2 t ha - l ) and the broadcast-seeded treatment (9.1 t ha- ~ ). For the same treatments in IR58 and IR29723-143-3-2-1, grain-yield was 0.8-1.0 t ha-1 lower. This was prob- ably due to the shorter growth duration in IR58 and to low solar radiation in combination with high night temperatures during the ripening phase of IR29723-143-3-2-1 (Fig. 1,120-140 DAS).
Few consistent trends could be observed in a yield comparison of trans- planted and broadcast-seeded rice. In control treatments (zero N ), grain-yields were generally higher in transplanted rice. This difference became larger with increasing crop duration. If N was B&I, grain-yield was significantly higher for broadcast-seeded than for transplanted plants in IR58, but lower in IR64 (P < 0.05 ). No significant yield differences occurred among transplanted and broadcast seeded treatments if N was band-placed. Grain-yields were mostly higher with band placement than with B&I (significant in IR64, TP and BS/ BSR, and in IR29723-143-3-2-1, TP), probably due to lower N volatilization and immobilization (Schnier et al., 1988 ).
A general yet hypothetical trend may be postulated if one considers that grain-yield was depressed in direct-seeded IR64 due to bad weather during the ripening phase (Fig. 1). At short crop duration (IR58), direct-seeding
Radiation (Md m" z d a y - l )
3°f 20,
10 I I I t I I I I
Temperature (°C) 4O
3.5
30
25
20
15 I I . 1 I I I I l 0 20 4 0 6 0 8 0 100 120 140 DAS
I - ~ J o n I • Feb I Mar I -- Apr I May - - - I 1988
Fig. 1. Daily solar radiation and minimum and maximum temperatures at the experimental site.
GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 3 3 3
showed a yield advantage over transplanting in N-fertilized treatments. At longer crop duration, this advantage disappeared or was inverted. Growth- delaying effects of transplanting shock (Dingkuhn et al., 1990a; Schnier et al., 1990a) may restrict biomass production and hence, yield, in short-duration varieties such as IR58. The more productive vegetative-growth period of di- rect-seeded rice may, in this case, translate into higher yield. At medium crop duration, however, the relative effect of the transplanting shock on produc- tion is smaller and may be insignificant at long crop duration. Other factors, such as canopy architecture and growth efficiency (Dingkuhn et al., 1990b), may predominantly determine yield in medium- and long-duration rices.
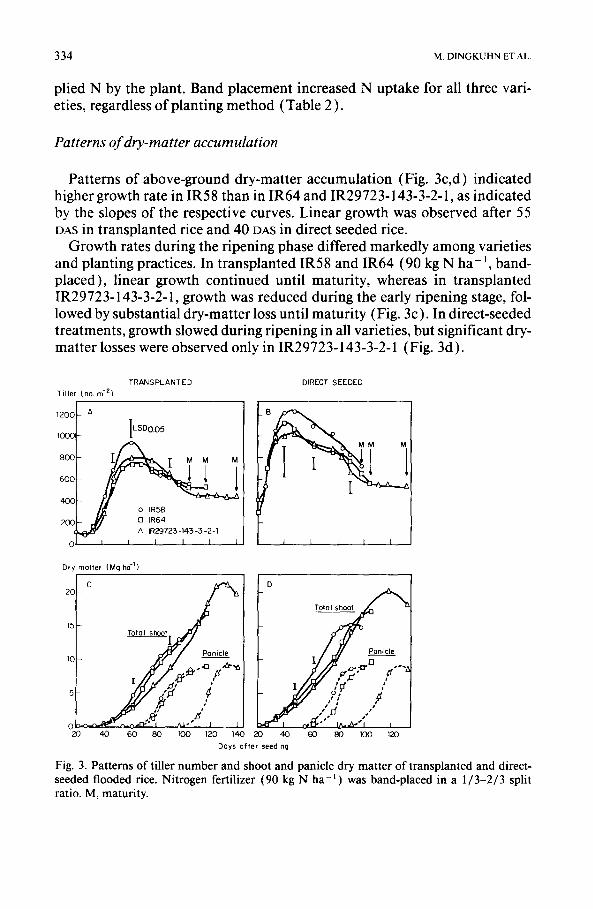
The biological and grain-yield relationships indicated in Fig. 2 support this interpretation. Biological yield was linearly related to crop duration. It was similar in transplanted and direct-seeded rice at long crop duration, but lower in transplanted rice at short crop duration, probably due to transplanting shock. Grain-yield, however, was not affected by crop duration, thus resulting in the lowest harvest index (HI) in IR29723-143-3-2-1, intermediate in IR64, and highest in IR58 (Table 2).
Agronomic efficiency (AE) of N fertilizer was higher in direct-seeded than in transplanted rice in all varieties (Table 2). Best AE (54 kg grain per kg N applied) was observed in BSR IR64 using N band placement ( 1 / 3-2 / 3 split ). This and similar N-application methods reduce N losses (Schnier et al., 1988 ). Improved AE and grain-yield resulting from liquid urea band placement in IR64 and IR29723-143-3-2-1 were, therefore, due to higher recovery of ap-
Yield at maturity (M(J ha -1) 20
18
16
14
10
Biological yield (dry) ~ s* j`'p&S*
#
- - - - j, ~, . , s J
~ " o'~ J ,-- 0929.*
Y=l.05+O.126X I E] lR64 r = 0944 W'~ • /% IR297'2.3-14,3
3-2-I
Groin yield at 14% MC
° I
I I I I I I 90 100 110 120 130 140
Growth duration (days)
Fig. 2. Biological and grain-yield in relation to growth duration in direct seeded and trans- planted flooded rice.
334 M . D I N G K U H N E T A L .
plied N by the plant. Band placement increased N uptake for all three vari- eties, regardless of planting method (Table 2 ).
Patterns of dry-matter accumulation
Patterns of above-ground dry-matter accumulation (Fig. 3c,d) indicated higher growth rate in IR58 than in IR64 and IR29723-143-3-2-1, as indicated by the slopes of the respective curves. Linear growth was observed after 55 DAS in transplanted rice and 40 DAS in direct seeded rice.
Growth rates during the ripening phase differed markedly among varieties and planting practices. In transplanted IR58 and IR64 (90 kg N ha- 1, band- placed), linear growth continued until maturity, whereas in transplanted IR29723-143-3-2-1, growth was reduced during the early ripening stage, fol- lowed by substantial dry-matter loss until maturity (Fig. 3c ). In direct-seeded treatments, growth slowed during ripening in all varieties, but significant dry- matter losses were observed only in IR29723-143-3-2-1 (Fig. 3d).
TRANSPLANTED DIRECT SEEDED Tiller (no.m -z )
120(
IOCX
800
600
40C
20C
A
I LSDo.05
T M M M
/,~ o ~ 1~/ fl IR64
~ " A IR29723 -14~-3-2-1 I I I I I 1 [ [ 1 [
Dry molter (Mg ha . I)
Total
r 7 / / . - . . - = i / .~ , ,,~# ,r ' f . / / I / . . . Y /
. ~ ~' • / / o ~ . . 4 ~ > . o . . - ' ~ ~ . - ' ~ , , k= ~', ~ " " ,^--~-'; ,
Days after seeding
Fig. 3. Patterns of tiller number and shoot and panicle dry matter of transplanted and direct- seeded flooded rice. Nitrogen fertilizer (90 kg N ha - l ) was band-placed in a 1 /3 -2 /3 split ratio. M, maturity.

GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 335
If N was B&I, dry-matter accumulation patterns were similar to those with band placement but growth rates during ripening were further reduced (data not presented).
Plant N uptake and foliage N concentration
Total plant N uptake varied greatly among treatments but little among va- rieties (Table 2). Total N uptake was not correlated with growth duration ( r= -0 .323 n's', controls; r= -0 .06 ns, 90 kg N ha - l ) . Grain-yield, however, was highly correlated with total N uptake ( r = 0.955"* ).
If no N fertilizer was applied, N uptake was higher in transplanted rice than in direct-seeded rice, although not significantly, in all varieties. At 90 kg N ha -I, plant N uptake depended on N application method and split. Broad- cast-and-incorporation (2 /3 -1 /3 split) raised N uptake by 46 to 68 kg ha-1 over control, while band placement (1 /3 -2 /3 split) led to additional N up- take of 68-100 kg ha -1. Schnier et al. (1988) studied the causes of higher N efficiency in band placement. In the present study, band placement not only increased grain-yield but also led to significantly (P< 0.05 ) higher N concen- tration in both straw and grain (overall means: 0.580% N (BP) vs. 0.462% N (B&I) in straw; 1.157% N (BP) vs. 0.982% N (B&I) in grain). Nitrogen harvest index (Table 2 ) was similar in IR64 and IR29723-143-3-2-1 (means: 0.67 and 0.66, respectively) but was higher in IR58 (0.71 ).
Temporal patterns of foliage N concentration are shown in Figs. 4e and f. Initial foliage N concentrations were low for transplanted rice, presumably due to transplanting shock, but increased during crop establishment. As fre- quently observed in crops (Kumura, 1975; Van Keulen and Seligman, 1987 ), N concentrations decreased after crop establishment until maturity due to depletion and tissue expansion, followed by N dilution (Schnier et al., 1990a) and translocation. The decline of foliar N concentration was continuous in transplanted rice, but was interrupted by a brief recovery due to the second N application in direct-seeded rice. At all developmental stages after crop es- tablishment, foliage N concentration was less in direct-seeded than in trans- planted rice. Similar observations have been reported for IR64 (Dingkuhn et al., 1990a; Schnier et al., 1990b). These could be explained by the earlier onset of exponential growth in direct-seeded rice resulting from the absence of a transplanting shock, followed by tissue N dilution. Higher plant density of broadcast-seeded rice further promoted N dilution.
Patterns of maximum canopy photosynthesis and leaf area index
Maximum canopy photosynthesis (Pnm,x) reached similar peak values ( 50 pmol CO2 m -2 s -I ) in all N-fertilized treatments and varieties (Fig. 4a,b). In transplanted treatments (Fig. 4a), canopy photosynthesis was low until 35
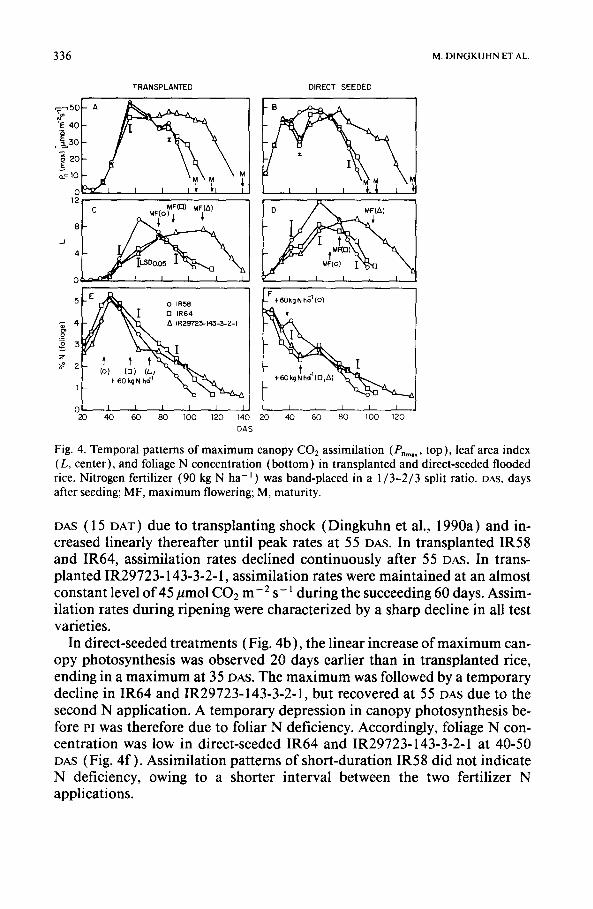
336 M . D I N G K U H N E T A L .
TRANSPLANTED DIRECT SEEDED
'~ 4Ol- I / [ ~ - i 0F ! % \
o~9/" i i I i l h
C MF(o ) ~,
5
4
3
i i
2O
2972;3--1,~'3 ..~- 2 -1
[ I I I I I 40 60 80 100 120 1,40
DAS
I D / ~ MF(A)
_F" +60kg N h6~(o )
l I 1 I I 20 40 coo 80 100 120
Fig. 4. Temporal patterns of maximum canopy C O 2 assimilation (P, .... top), leaf area index (L, center), and foliage N concentration (bottom) in transplanted and direct-seeded flooded rice. Nitrogen fertilizer (90 kg N ha -l) was band-placed in a 1/3-2/3 split ratio. DAS, days after seeding; MF, maximum flowering; M, maturity.
DAS (15 DAT) due to transplanting shock (Dingkuhn et al., 1990a) and in- creased linearly thereafter until peak rates at 55 DAS. In transplanted IR58 and IR64, assimilation rates declined continuously after 55 DAS. In trans- planted IR29723-143-3-2- l, assimilation rates were mainta ined at an almost constant level of 45/tmol C02 m -2 s-1 during the succeeding 60 days. Assim- ilation rates during ripening were characterized by a sharp decline in all test varieties.
In direct-seeded t reatments (Fig. 4b ), the linear increase of m ax i m um can- opy photosynthesis was observed 20 days earlier than in transplanted rice, ending in a m a x i m u m at 35 DAS. The max imum was followed by a temporary decline in IR64 and IR29723-143-3-2-1, but recovered at 55 DAS due to the second N application. A temporary depression in canopy photosynthesis be- fore Pi was therefore due to foliar N deficiency. Accordingly, foliage N con- centrat ion was low in direct-seeded IR64 and IR29723-143-3-2-1 at 40-50 DAS (Fig. 4f ) . Assimilation patterns of short-duration IR58 did not indicate N deficiency, owing to a shorter interval between the two fertilizer N applications.

GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 337
Patterns of leaf-area index (L) did not mirror canopy assimilation patterns (Fig. 4c,d). While maximum L coincided with peak canopy photosynthesis in both transplanted and direct-seeded IR58, IR29723-143-3-2-1 reached its highest assimilation rates clearly before L was at its maximum.
Unlike plant height (Table 1 ) and dry-matter production (Fig. 2), maxi- mum L was not a function of growth duration. Medium-duration IR64 had the lowest L but did not differ significantly in assimilation rates from the other test varieties. That IR64 had the lowest leaf area but highest grain-yield and agronomic efficiency among the test varieties (Table 2) even suggests a negative effect of high leaf area on yield in the N-fertilized treatments. The L values observed in this experiment were high, and may therefore have ex- ceeded optimum (Dingkuhn et al., 1990b). An optimum L of 7 has been postulated by Tanaka et al. (1966). Murty and Murty ( 1981 ) found a posi- tive correlation between L and yield for an L range of 2-6.
Peak L was generally observed at the heading stage. Highest L was observed in IR58 (transplanted, 8.6; direct-seeded, 11.6). In all test varieties and N regimes, direct-seeded treatments had higher L than transplanted treatments.
Tillering patterns
Active tillering was observed as early as 20 DAS in direct-seeded treatments, but was delayed in transplanted rice until 30-35 DAS (10-15 OAT), due to transplanting shock (Schnier et al., 1990a). Maximum tiller number and tiller number at maturity (Fig. 3a,b) were higher in direct-seeded than in trans- planted rice as a result of higher plant density ( 100 plants m-2, TP; 220 m-2, BS; and 205 m -2, BSR) and a longer tillering phase. Maximum tiller number was observed as early as 40 DAS (IR58) and 50 DAS (IR64 and IR29723-143- 3-2-1 ) in direct-seeded treatments compared with 60 hAS (IR58 ) and 70 DAS (IR64 and IR29723-143-3-2-1 ) in transplanted treatments. Tillering was fol- lowed by a tiller abortion phase which was particularly pronounced in direct- seeded rice.
Cultivar IR58, with the shortest growth duration among the test varieties, produced and maintained the highest tiller number. The varietal differences in tillering activity, however, may not necessarily represent genetic variation in tillering ability. Relative tillering and tiller abortion were a function of rel- ative growth rate (RGR) (Schnier et al., 1990a). This linear relationship was identical among the three varieties and agreed with observations made at the same location in the previous year (Fig. 5 ). Relative growth of 5.5-6% day- represented the threshold for both tiller production and tiller abortion. Schnier et al. (1990a) have shown that the RGR threshold for active tillering is con- stant during the vegetative and early reproductive growth phases. High till- ering activity in IR58 was therefore a result of high RGR, probably caused by the short interval between the two N-fertilizer applications.
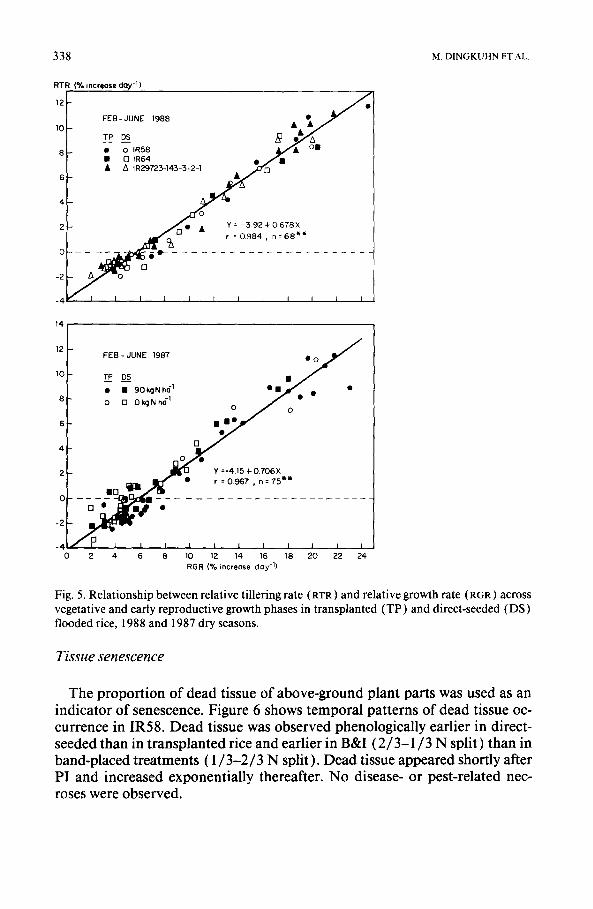
338 M. D I N G K U H N ET AL.
RTR (% increase dOy -I)
12
10
81
6
4
2
0
-2
_4
FEB-JUNE ]988 • ~
T P D_S ~ • o IR58 ~"~A on
A ~ t ~ U -- r --0.984, n--68 ~* . . . . . . . . . . . . . . . . . . . . .
. ~ ' - o
I 1 I I 1 I 1 I I I I 1
14
12
10
8
6
4
2
0
-2
- 4 0
/ FEB- JUNE 1987 0 0 7 "
/ - r P DS • / • • 90kgNho-I Omm j,,," • •
0 E] OkQNho-' 0 / j -
m m~y"
J~'U Y :-4.15 + 0706X . ~ - O r : 0.967 , n :75 ~*
I n ~ . . . . . . . . . . . . . . . . . .
z 1 I I 1 I I , , l I l I I 2 4 6 8 10 12 14 16 18 20 22 24
RGR (% increase day "I)
Fig. 5. Relationship between relative tiilering rate (RTR) and relative growth rate (RGR) across vegetative and early reproductive growth phases in transplanted (TP) and direct-seeded (DS) flooded rice, 1988 and 1987 dry seasons.
Tissue senescence
The proportion of dead tissue of above-ground plant parts was used as an indicator of senescence. Figure 6 shows temporal patterns of dead tissue oc- currence in IR58. Dead tissue was observed phenologically earlier in direct- seeded than in transplanted rice and earlier in B&I ( 2 / 3 - 1 / 3 N split) than in band-placed treatments ( 1 / 3 - 2 / 3 N split). Dead tissue appeared shortly after PI and increased exponentially thereafter. No disease- or pest-related nec- roses were observed.
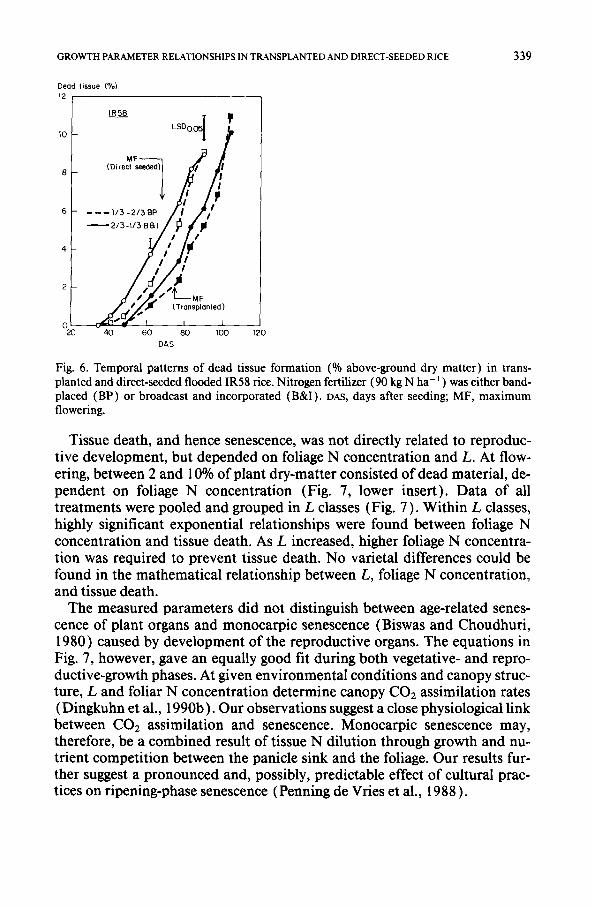
GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 339
Dead tissue (%) 12
10'
20 120
IR58 LSD 0.05[ i ~
2/ (Direct seeded) 1
7'13-1/3 B & I
/ # / "/j/,¢ I ,
I I 40 60 BO lO0
DAS
Fig. 6. Temporal patterns of dead tissue formation (% above-ground dry matter) in trans- planted and direct-seeded flooded IR58 rice. Nitrogen fertilizer (90 kg N ha-t ) was either band- placed (BP) or broadcast and incorporated (B&I). DAS, days after seeding; MF, maximum flowering.
Tissue death, and hence senescence, was not directly related to reproduc- tive development, but depended on foliage N concentration and L. At flow- ering, between 2 and 10% of plant dry-matter consisted of dead material, de- pendent on foliage N concentration (Fig. 7, lower insert). Data of all treatments were pooled and grouped in L classes (Fig. 7). Within L classes, highly significant exponential relationships were found between foliage N concentration and tissue death. As L increased, higher foliage N concentra- tion was required to prevent tissue death. No varietal differences could be found in the mathematical relationship between L, foliage N concentration, and tissue death.
The measured parameters did not distinguish between age-related senes- cence of plant organs and monocarpic senescence (Biswas and Choudhuri, 1980) caused by development of the reproductive organs. The equations in Fig. 7, however, gave an equally good fit during both vegetative- and repro- ductive-growth phases. At given environmental conditions and canopy struc- ture, L and foliar N concentration determine canopy CO2 assimilation rates (Dingkuhn et al., 1990b). Our observations suggest a close physiological link between CO2 assimilation and senescence. Monocarpic senescence may, therefore, be a combined result of tissue N dilution through growth and nu- trient competition between the panicle sink and the foliage. Our results fur- ther suggest a pronounced and, possibly, predictable effect of cultural prac- tices on ripening-phase senescence (Penning de Vries et al., 1988 ).
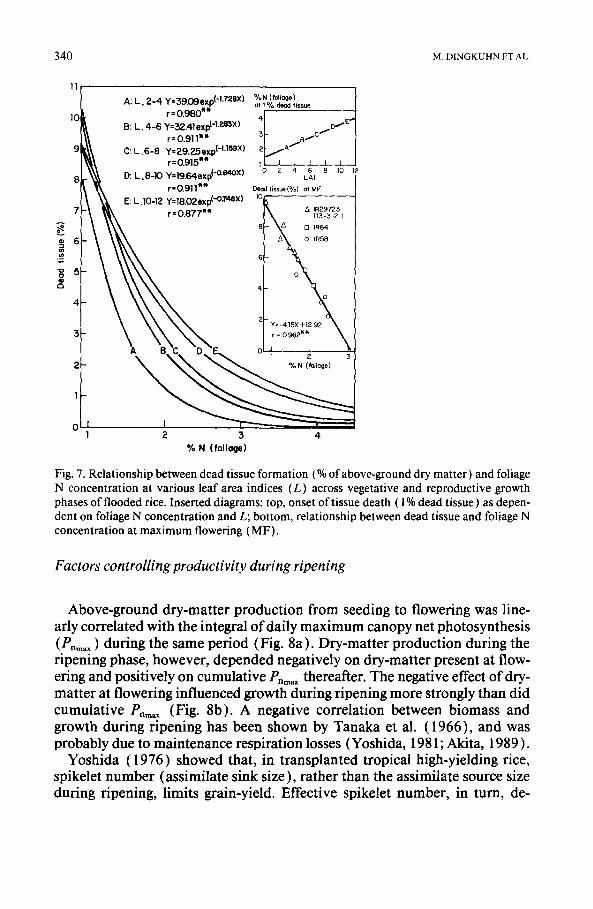
11 % N (foliage) Y=39.09=_xy ( ' l '72ex)~ at 1% dead tissue A: L. 2-4
# o=
3 4 0 M. DINGKUHN ET AL.
1 2 3 4 % N (foliage)
Fig. 7. Relationship between dead tissue formation (% of above-ground dry matter) and foliage N concentration at various leaf area indices (L) across vegetative and reproductive growth phases of flooded rice. Inserted diagrams: top, onset of tissue death ( 1% dead tissue) as depen- dent on foliage N concentration and L; bottom, relationship between dead tissue and foliage N concentration at maximum flowering (MF).
Factors controlling productivity during ripening
Above-ground dry-matter production from seeding to flowering was line- arly correlated with the integral of daily maximum canopy net photosynthesis (Pnm,x) during the same period (Fig. 8a). Dry-matter production during the ripening phase, however, depended negatively on dry-matter present at flow- ering and positively on cumulative Pn~ax thereafter. The negative effect of dry- matter at flowering influenced growth during ripening more strongly than did cumulative Pnm,x (Fig. 8b). A negative correlation between biomass and growth during ripening has been shown by Tanaka et al. (1966), and was probably due to maintenance respiration losses (Yoshida, 1981; Akita, 1989).
Yoshida (1976) showed that, in transplanted tropical high-yielding rice, spikelet number (assimilate sink size), rather than the assimilate source size during ripening, limits grain-yield. Effective spikelet number, in turn, de-
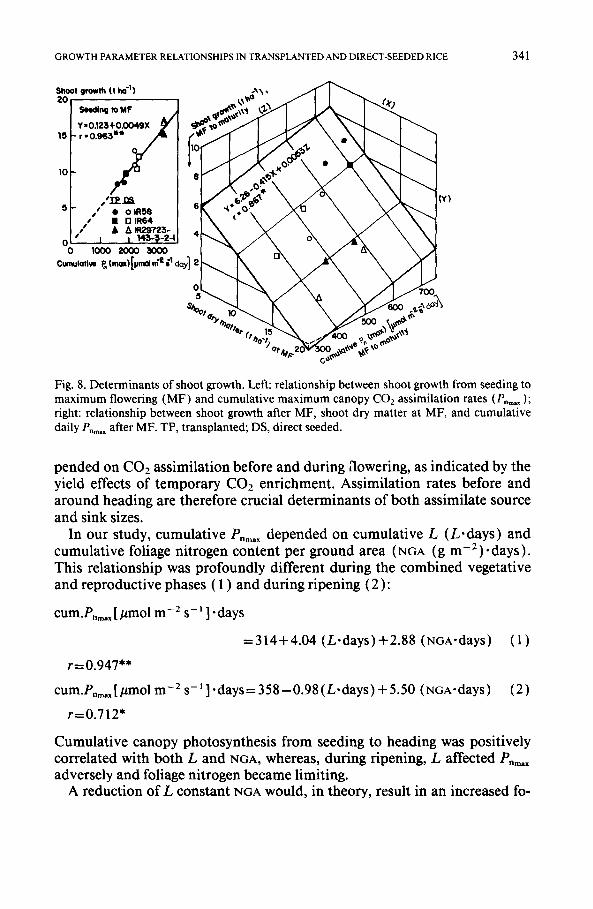
GROWTH PARAMETER RELATIONSHIPS 1N TRANSPLANTED AND DIRECT-SEEDED RICE 341
Shoot itOwth (! ha "1) 20 t-- ~ k~ x~c
I,.om÷o.oo x ¥ / - to" S
j t °
| • • 01R64 ~% ~ ' / , ' • ~ ~fmn~- 4- \ /
c . , , , ° , , .
%rte, 0
?OO
Fig. 8. Determinants of shoot growth. Left: relationship between shoot growth from seeding to maximum flowering (MF) and cumulative maximum canopy CO2 assimilation rates (P.=.~); right: relationship between shoot growth after MF, shoot dry matter at MF, and cumulative daily P,~.x after MF. TP, transplanted; DS, direct seeded.
pended o n C O 2 assimilation before and during aowering, as indicated by the yield effects of temporary CO2 enrichment. Assimilation rates before and around heading are therefore crucial determinants of both assimilate source and sink sizes.
In our study, cumulative P-max depended on cumulative L (L.days) and cumulative foliage nitrogen content per ground area (NGA (gm -2) -days). This relationship was profoundly different during the combined vegetative and reproductive phases ( 1 ) and during ripening (2):
cum.P,m~ [ #mol m- 2 g- - l ] . days
=314+4.04 (L.days)+2.88 (NGA'days) (1)
r=0.947"*
cum.P,~x[#molm-2s-1].days=358-0.98(L.days)+5.50 (NGA'days) (2)
r=0.712"
Cumulative canopy photosynthesis from seeding to heading was positively correlated with both L and NGA, whereas, during ripening, L affected P,m,x adversely and foliage nitrogen became limiting.
A reduction of L constant NGA would, in theory, result in an increased fo-
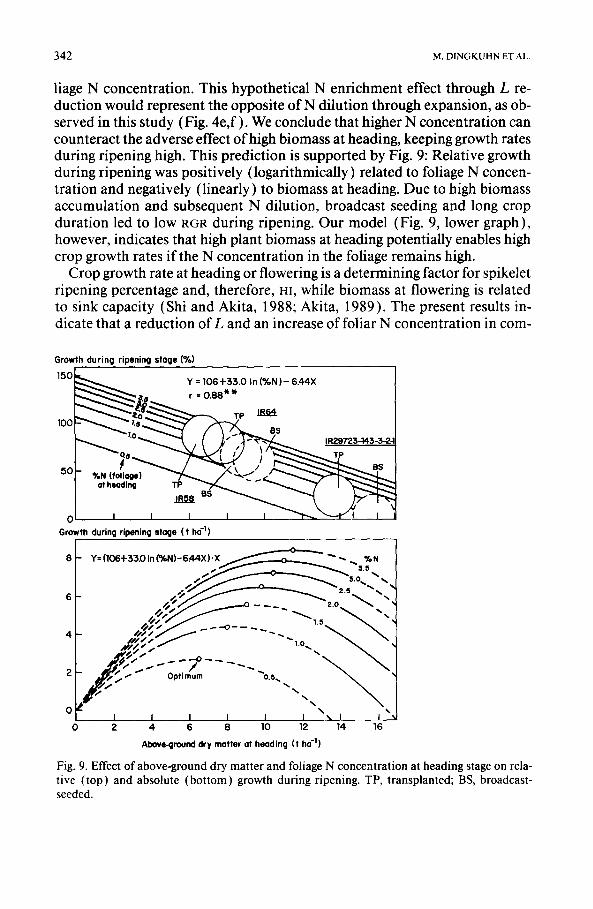
342 M. D1NGKUHN ET AL.
liage N concentration. This hypothetical N enrichment effect through L re- duction would represent the opposite of N dilution through expansion, as ob- served in this study (Fig. 4e,f) . We conclude that higher N concentration can counteract the adverse effect of high biomass at heading, keeping growth rates during ripening high. This prediction is supported by Fig. 9: Relative growth during ripening was positively (logarithmically) related to foliage N concen- tration and negatively (linearly) to biomass at heading. Due to high biomass accumulation and subsequent N dilution, broadcast seeding and long crop duration led to low RGR during ripening. Our model (Fig. 9, lower graph), however, indicates that high plant biomass at heading potentially enables high crop growth rates if the N concentration in the foliage remains high.
Crop growth rate at heading or flowering is a determining factor for spikelet ripening percentage and, therefore, m, while biomass at flowering is related to sink capacity (Shi and Akita, 1988; Akita, 1989). The present results in- dicate that a reduction of L and an increase of foliar N concentration in com-
Growth during ripening stage (%]
1 5 0 ~ _ _ _ ~ = Y = 106+33.0 In (%N) - 6,44X
~ ~ r = 0 . 8 8 " "
I otheading TP ~ - ~ ~ / J " ~ _ ~
OI I I I I I " ~ " ~ " - - - ~ f {
Growth during ripening stage ( t ha -11
Y = ( 1 0 6 + 3 3 0 I n ( % N ) - 6 . 4 4 X ) . X ~ " " ~. ,.. %N 8 • , , ~ , . . - ~ - - ~ ~ 3.5
~ 10 •
2 - ~ , ~ Optimum O.S.,, ~ •
l I I I I I ~'% I I ~ , 0 2 4 6 8 10 12 14 16
Above-ground dry matter at heading (t ha -1)
Fig. 9. Effect of above-ground dry matter and foliage N concentration at heading stage on rela- tive (top) and absolute (bottom) growth during ripening. TP, transplanted; BS, broadcast- seeded.

GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 343
bination with high biomass at flowering may raise grain-yield of tropical low- land rice. Direct-seeded rice, in this experiment, exhibited higher L (Fig. 4c,d ) and lower foliage N concentration (Fig. 4e,f) than transplanted rice, while having higher biomass at flowering (Fig. 3c,d ). The inability of direct-seeded and long-duration rice to convert high biomass at heading into grain-yield may be improved in future cultivars with lower L, higher foliar N concentra- tion, a modified N-partitioning pattern, and an expanded storage ability of the stems for carbohydrates (Chaturvedi and Ingram, 1990).
CONCLUSIONS
Of the numerous significant varietal differences observed, most could be traced back to common physiological or agronomic response patterns:
- biomass at maturity and plant height were a function of crop duration; - tiller production and abortion were related to relative growth rate; - tissue death was related to foliage N concentration and L; - growth during ripening was a function of biomass at flowering and foli-
age N concentration, which in turn depended on L; - peak values of canopy net photosynthesis were similar among the vari-
eties, their patterns being determined by planting technique, density, and N fertilization rather than by varietal differences;
- total N uptake depended on N application technique and not on cultivar or crop duration.
The only varietal differences that could not be explained by cultural factors were growth duration and leaf area. Little is known about the physiological basis of genotypic differences in growth duration, and no new clues can be offered here. Our observation that foliage growth capacity appears to be high- est in IR58 and lowest in IR64 may be of agronomic importance.
A high plant biomass at flowering may offer a good basis for high grain- yield potential, but poses a serious constraint to growth during ripening. This constraint is especially significant in direct-seeded medium- to long-duration rice, due to a very productive vegetative-growth phase and subsequent dilu- tion of foliar N. Consequently, tropical rice cropping systems that use direct- seeding and medium- to long-duration cultivars require plants with moderate leaf area and high foliar N concentration during the reproductive growth stages, enabling high growth rates in spite of a high respiring biomass. As our study showed, this concept can only partly be realized through modified cul- tural practices, and hence should be further addressed by breeders.
ACKNOWLEDGEMENT
This research is supported by the German Agency for Technical Coopera- tion (GTZ), Germany.

344 M. DINGKUHN ET AL.
REFERENCES
Akita, S., 1989. Improving yield potential in tropical rice. In: Progress in Irrigated Rice Re- search. International Rice Research Institute, Manila, The Philippines, pp. 41-76.
Anonymous, 1987. Canopy photosynthesis of direct seeded and transplanted rice. In: Annual Report for 1986. International Rice Research Institute, Manila, The Philippines, pp. 368- 370.
Biswas, A.K. and Choudhuri, M.A., 1980. Mechanism of monocarpic senescence in rice. Plant Physiol., 65: 340-345.
Chaturvedi, G.S. and Ingram, K.T., 1990. Assimilate translocation of lowland rice in response to water deficit and shade. Philipp. J. Crop Sci. (in press).
Dingkuhn, M., Schnier, H.F., De Datta, S.K., Wijangco, E. and Dorffling, K., 1990a. Diurnal and developmental changes in canopy gas exchange in relation to growth in transplanted and direct seeded flooded rice. Aust. J. Plant Physiol., 17:119-134.
Dingkuhn, M., Schnier, H.F., De Datta, S.K., Dorffling, K., Javellana, C. and Pamplona, R., 1990b. Response of direct seeded and transplanted flooded rice to nitrogen fertilizer appli- cation. II. Effect of leaf area and nitrogen status on foliage expansion, senescence, canopy photosynthesis and growth. Crop Sci. (in press).
Kawano, K. and Tanaka, A., 1968. Growth duration in relation to yield and nitrogen response in the rice plant. Jpn. J. Breed., 18: 46-52.
Kumura, A., 1975. Comparison of growth characteristics between species. In: Y. Murata (Edi- tor), Crop Productivity and Solar Energy Utilization in Various Climates in Japan. JIBP Synthesis Vol. 11, University of Tokyo Press, Tokyo, Japan, pp. 221-234.
Morgan, A., t988. Growth and canopy carbon dioxide exchange rate of spring wheat as affected by nitrogen status. Crop Sci., 28: 95-100.
Murty, P.S.S. and Murty, M.S., 1981. Influence of LAI at flowering on the productive efficiency of long duration rice cultures. Oryza, 18: 35-38.
Penning de Vries, F.W.T., van Keulen, H. and Alagos, J.C., 1988. Nitrogen redistribution and potential production in rice. In: S.K. Sinha, P.V. Sane, S.C. Bhargava and P.K. Agrawahl (Editors), Proc. Int. Congress Plant Physiology, New Delhi, India, 15-20 February 1988. Society for Plant Physiology and Biochemistry, New Delhi, Vol. 1, pp. 513-520.
Schnier, H.F., De Datta, S.K., Mengel, K., Marqueses, E.P. and Faronilo, J.E., 1988. Nitrogen use efficiency, floodwater properties, and nitrogen- 15 balance in transplanted lowland rice as affected by liquid urea band placement. Fert. Res., 16:241-255.
Schnier, H.F., Dingkuhn, M., De Datta, S.K., Mengel, K., Wijangco, E. and Javellana, C., 1990a. Nitrogen economy and canopy CO2 assimilation in tropical lowland rice. Agron. J., 82:451- 459.
Schnier, H.F., Dingkuhn, M., De Datta, S.K., Mengel, K. and Faronilo, J.E., 1990b. Response of direct seeded and transplanted flooded rice to nitrogen fertilizer application. I. Plant ni- trogen uptake, canopy photosynthesis, growth and yield. Crop Sci. (in press).
Shi, Q. and Akita, S., 1988. Biomass production and grain yield of lR cultivars in high nitrogen water culture. Jpn. J. Crop Sci., 57 (extra iss. 1 ): 23-24.
Tanaka, A., Kawano, K. and Yamaguchi, J., 1966. Photosynthesis, respiration, and plant type of the tropical rice plant. IRRI Tech. Bull. 7.
Van Keulen, H. and Seligman, N.G., 1987. Simulation of Water Use, Nitrogen Nutrition and Growth of a Spring Wheat Crop. Pudoc, Wageningen, The Netherlands.
Vergara, B.S. and Chang, T.T., 1985. Flowering response of the rice plant to photoperiod. A review of Literature (4th edition). International Rice Research Institute, Manila, The Philippines.
Yoshida, S., 1976. Carbon dioxide and yield of rice. In: Climate and Rice. International Rice Research Institute, Manila, The Philippines, pp. 221-222.

GROWTH PARAMETER RELATIONSHIPS IN TRANSPLANTED AND DIRECT-SEEDED RICE 345
Yoshida, S., 1981. Fundamentals of Rice Crop Science. International Rice Research Institute, Manila, The Philippines.
Yoshida, S., 1983. Growth and yield of field crops. In: Potential Productivity of Field Crops under Different Environments. International Rice Research Institute, Manila, The Philip- pines, pp. 103-128.
Copyright © 2022 FDOKUMEN




















