
Molecular prey identification in wild Octopus vulgaris paralarvae
Upload
camden-rutgersCategory
view
1download
0
Cell Tissue Res. 204, 463-472 (1979) Cell and Tissue Research �9 by Springer-Verlag 1979
Relationship Between Photoreceptor Terminations and Centrifugal Neurons in the Optic Lobe of Octopus
William M. Saidel
Neurobiology Unit, Scripps Institution of Oceanography and Department of Neurosciences, School of Medicine, University of California, San Diego, La Jolla, USA
Summary. Retinal bundles, connecting the retina of the octopus to the ipsilateral optic lobe, contain both retinal photoreceptor axons that terminate in the optic lobe and centrifugal axons whose cell bodies lie within the lobe. Staining axonal elements in proximal stubs of individual retinal bundles by cobalt diffusion and subsequent sulphide treatment reveals the topographic relationship between afferent terminals and centrifugal cell bodies. At the outer border of the plexiform layer, stained terminal bags (photoreceptor axon enlargements), an indicator of photoreceptor terminal spread within this layer, overlap stained centrifugal cell bodies located within the inner granule layer. The details of this overlap indicate a dorsoventral representation of each retinal bundle within the optic lobe cortex.
Key words: Optic lobe cortex - Centrifugal cells - Photoreceptor terminals - Cobalt staining - Octopus bimaculoides and O. bimaculatus (Cephalopoda).
Many retinal nerve bundles connect an octopus eye with its central ganglia. These bundles interweave in a vertical chiasm within the sinus just medial to the eye (Kopsch, 1899; Cajal, 1917; Young, 1962b, 1971). They continue through the sinus surrounded by the white-body tissue to reach the optic lobe in which they terminate (Young, 1971; Wells, 1978). The bundles contain both centripetally-directed photoreceptor axons that terminate in the cortex of the optic lobe (Lenhossek, 1896; Cajal, 1917; Young, 1962b, 1971; Dilley et al., 1963; Case et al., 1972; Cohen, 1973a, b; Case, 1976) and axons of centrifugal cells located in the inner granule layer (Cajal, 1917; Young, 1962 a, 1971) that terminate near the proximal segments of photoreceptors or photoreceptor axon collaterals (Young, 1962a; Lund, 1966; Gray, 1970; Cohen, 1973a) and/or intraretinal muscle fibers (Alexandrowicz, 1927).
Send offprint requests to: Dr. William M. Saidel, Department of Neurosciences A-001, University of California, San Diego, La Jolla, CA 92093, USA
Acknowledgements: I gratefully acknowledge the helpful advice of Drs. T.H. Bullock and R.G. Northeutt. This work was supported by a grant from N.I.N.C.D.S. to Dr. T.H. Bullock
0302-766X/79/0204/0463/$02.00

464 W.M. Saidel
Topographic analysis of the afferent te rminat ions and centrifugal cell bodies within the optic lobe has no t been subject to prior invest igat ion because the methods of classical cephalopod neuroana tomy, Golgi impregnat ion and silver stains (see Young, 1971), do no t lend themselves to control led localization. The experimental method of axonal degenerat ion has been used most ly to conf i rm pathways inferred from descriptive ana tomy.
This study utilized cobal t diffusion through proximal stubs of sectioned retinal bundles to examine the locations within the optic lobe of s imul taneously-s ta ined photoreceptor te rminat ions and centrifugal cell bodies. In addit ion, the stained afferent te rminat ions demonst ra te that retinal regions corresponding to single retinal bundles are represented in mediolateral ly oriented slabs within the optic lobe cortex.
Materials and Methods
Octopus bimaculoides and O. bimaculatus (80-150g) were anesthetized in cold sea water containing 3-5 % alcohol until sucker reflexes and respiration disappeared. Both optic lobes were removed and placed in Petri dishes. The ends of one or more cut retinal bundles were placed into small polyethylene cups (made from Intramedic polyethylene tubing, No's. PE 90 and 160). A cup was first filled with distilled water to dilate the end of the cut nerve, then drained and filled with 0.3 M to 1.0 M CoCI 2. In some cases, individual grains of CoC12 were placed in a cup containing distilled water. Co 2§ ions were allowed to travel passively along the fibers for 4-20 hours at 10-12 ~ C. After rinsing in sea water for five minutes, the lobe was immersed in 10 % ammonium sulfide mixed in sea water for 45 min to 1 h, rinsed in sea water, and fixed with 10 700 formalin orAFA (90 cc of 80 7o ethanol, 5 cc glacial acetic acid, 5 cc formalin). Lobes were dehydrated in graded ethanols, cleared in xylene, embedded in Paraplast, and sectioned in various orientations at 8-30~tm. A later modification, dehydration in a tert.-butyl series, proved more suitable for embedding and sectioning. Some sectioned lobes were treated with Goodman's (1976) modification of Timm's intensification procedure (Tyrer and Bell, 1974). A further modification, lowering the incubation temperature from 60~ to 50~ proved useful. Identification of cobalt-stained areas was facilitated by comparing the experimental material with optic lobe sections stained by the Bodian method (Humason, 1979).
Density of inner granule centrifugal cells was determined by counting all inner granule cells stained by one retinal nerve bundle; the volume was determined from the lateral extent of these cells and the depth along the dorsoventral axis. Depth was computed from the number and thickness of serial sections from a horizontal tissue series. Linear dimensions measured from sectioned material were uncorrected for paraffin embedding. Final cell density was determined in terms of a 100 ~tm cube.
External reference to various axes in the octopus nervous system is complicated by the many postural complexities demonstrated by this animal. The following terminology was adapted for consistency. Medial and lateral directions are referred to a sagittal section of the central ganglia. Anterior (towards the head); posterior (towards the mantle); ventral (towards the side containing the funnel and mantle opening); and dorsal (towards the side opposite ventral) are used in accordance with Young (1971).
Results
Both afferent fiber terminals and centrifugal neurons in the optic lobe were successfully stained by s imul taneous or thodromic and ant idromic cobalt move- ment . Individual , stained fiber fascicles subdivided as they coursed through the 50-60 Ixm-thick, outer granule layer after leaving a retinal bundle (Fig. 1 A-B). At the outer border of the plexiform layer, most photoreceptor axons formed a conical- shaped terminal bag 8 ~tm in length (Insert, Fig. 1 B). Darkened terminal bags

Cortical Organization in Octopus Optic Lobe 465
Fig. 1A and B. Localization of cobalt-stained elements in the optic lobe cortex of Octopus bimaculoides. A Divisions of cortex of Tele optic lobe. Outer granule layer (OGL); plexiform layer (PL); and inner granule layer (IGL). (Prepared by a Bodian stain.) B Elements within the cortex stained by immersing a retinal bundle in 0.3 M cobalt chloride. Retinal afferents course through the outer granule layer to form enlargements, terminal bags (insert), at the outer margin of the plexiform layer (tb). Many afferents terminate in plexiform tangential lamina 2 (t2). Centrifugal cells are located predominantly within the inner granule layer. Scale: 50 ~tm
defined the afferent terminal zone at the optic lobe surface (Figs. 1 B, 2). When a lobe was sectioned in an anteroposterior series, the stained region varied little in width in adjacent sections. The region tapered as it approached the medial edge of the lobe (Fig. 2B-D).
Many afferent fibers terminated within the plexiform layer near its distal border; others terminated closer to its proximal border. These layers apparently correspond to the first radial and second tangential layers respectively (Young, 1962b) (Fig. 1 B). A few fibers continued without forming a terminal bag through the cortex to end in deep cell islands, neuropil or in the optic tract. (This last location prevented identification of the particular fiber as centrifugal or centripetal.) Terminations in the lobe medulla were uncommon.
Other fibers branched through the plexiform layer before terminating in their cell bodies located primarily in the inner granule layer (Figs. 1B, 2B-D, 3).

Fig.
2A
-D.
Pro
ject
ion
zone
wit
hin
the
cort
ex o
f th
e op
tic
lobe
fro
m s
tain
ing
a si
ngle
ret
inal
bun
dle.
A S
chem
atic
dra
win
g sh
owin
g th
e m
edio
late
ral
aspe
ct a
fter
st
aini
ng a
sin
gle
reti
nal b
undl
e. B
-D P
hoto
mic
rogr
aphs
illu
stra
ting
the
ant
erop
oste
rior
asp
ect o
f a
slab
sta
ined
by
a si
ngle
reti
nal
bund
le.
Eac
h se
ctio
n is
200
~tm
m
ore
late
ral
than
the
pre
cedi
ng o
ne.
Sec
tion
thic
knes
s is
20
Ixm
. Sca
le:
B-D
: 10
0 ~t
m. A
bbre
viat
ions
: A
ant
erio
r; L
lat
eral
; M
med
ial;
P p
oste
rior
; ch
chi
asm
; p-o
lf
pedu
ncle
-olf
acto
ry l
obes
com
plex
; ot
opt
ic t
ract
. Se
e al
so l
egen
d of
Fig
. 1
.... /~_/_--f--~ ~ -c:
r ~
a
| , /" ~/ J / /S // ~ J
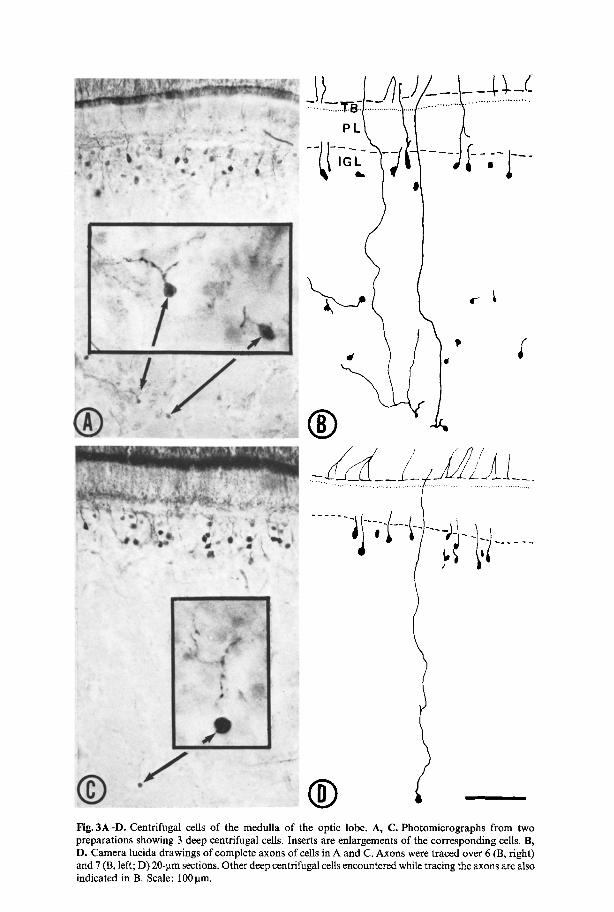
| Fig. 3A-D. Centrifugal cells of the medulla of the optic lobe. A, C. Photomicrographs from two preparations showing 3 deep centrifugal cells. Inserts are enlargements of the corresponding cells. B, D. Camera lucida drawings of complete axons of cells in A and C. Axons were traced over 6 (B, fight) and 7 (B, left; D) 20-gm sections. Other deep centrifugal cells encountered while tracing the axons are also indicated in B. Scale: 100 lam.

468 W.M. Saidel
|
/
/// 2-, )_
Fig. 4A-G. Centrifugal cells of the inner granule layer with emphasis on lateral dendritic projections within the plexiform layer. A-C. Three cells illustrating branches of the exiting neurite located especially near plexiform lamina t 2 (arrow). D--G. Camera lucida drawings of four additional cells showing characteristic lateral branches at the boundary between plexiform and inner granule layers (dashed line) and at plexiform lamina t 2 (broken line). Scale: 10 Ima
Scat tered ind iv idua l cells were also loca ted deeper in cell is lands o f the medul la . F igure 3 A - D shows cells loca ted more than 300 ~tm below the ou te r bo rde r o f the inner granule layer.
A compar i son between the te rminal afferent zone and centr i fugal cell loci suggests tha t componen t s o f ind iv idua l re t inal bundles are loca ted in discrete slabs l imited in the an te ropos te r ior plane and l imited at the slab ends a long a mediola tera l c i rcumference (Fig. 2). The te rmina l bag zone, however , ex tended fur ther a long a

Cortical Organization in Octopus Optic Lobe 469
mediolateral circumference than did the region of underlying stained cells. Centrifugal cell density tended to be sparcer at the medial and lateral borders of a slab than at its center.
All nerve bundles intertwining at a point in the chiasm were dipped in cobalt in one preparation. The stained area of the lobe traversed an entire mediolateral circumference without exceeding an anteroposterior spread of 300 gm. Chiasm intertwinings of retinal bundles apparently define an entire mediolateral slab of the lobe cortex.
Stained cells within the inner granule layer appeared in one of three predominant shapes. A majority were unipolar with the centrifugal process projecting radially outward to the surface of the lobe or laterally and then turning outward. Cells with two processes were the next most frequent type. Types could be distinguished depending upon the opposed or adjacent positions of the processes. More than two processes projected from a third type of cell. Only one exited the lobe via a retinal bundle. The others extended laterally within the inner granule layer, centrally to the medulla or laterally within the plexiform layer.
Axons of the centrifugal cells branched conspicuously within two plexiform laminae (Fig. 4A-G). These branches were most probably dendrites, not axon collaterals (Young, 1962b). Many branched tangentially at the level where deep photoreceptor terminals ended (second tangential layer). Others branched at the plexiform-inner granule layer border (fourth tangential layer). Branches in other tangential layers were also identified.
The lateral extent of darkened terminal bags with two exceptions overlapped the stained inner granule layer cell bodies. A few cells just outside the outer margin of stained terminal bags were the first exception. In many of those cases, a fiber projected laterally into the stained terminal bag area to eventually exit the lobe. Secondly, only centrifugal cells stained when the staining time was short (less than 8 h). This latter result is consistent with a passive model of cobalt diffusion through the larger fibers of these cells.
The density of cobalt-stained centrifugal cells was determined in four separately stained lobes in which the terminal bags formed exceptionally distinct slabs. These lobes were cut along the mediolateral axis to minimize oblique sectioning of the stained area. (Density of stained cells increased by 50 % or more per section when the stained area was not radially sectioned.) A cell was counted only when a unique axon passing through the entire plexiform layer could be identified regardless of intensity of cell staining. Cobalt-stained cell densities in these lobes (1.8-2.5 cm long) were 30.6, 32.4, 34.8 and 39.3 cells per 100 gm 3.
Discussion
This paper demonstrates the topographic relationship between locations of centrifugal cells in the optic lobe and retinal photoreceptor terminations by the first use of cobalt staining in the octopus central nervous system. Retinal bundles partition the lobe into a series of parallel slabs oriented along a mediolateral circumference. A circumferential slab is defined by the intertwining of all retinal bundles at a point in the chiasm. A portion of one slab is defined by staining only a

470 W.M. Saidel
single retinal bundle. Within a slab or a portion of one, photoreceptor terminals overlap centrifugal cell bodies in the plexiform and inner granule layers respectively.
Similar cell densities and uninterrupted afferent terminals displayed in the four measured lobes suggest that this method does stain all or nearly all of the axons in each bundle. However, with respect to the method, two points must be mentioned. Short staining times did not reveal the smaller afferent fibers indicating some relationship between axon diameter, time and depth of staining. The various degrees to which centrifugal cells filled agree with the observation by Strausfeld and Obermayer (1976) that large axon diameter is not a guarantee of complete filling. Some lightly stained, inner granule cells could be identified as centrifugal rather than as transynaptically stained because an axon was located traversing the entire plexiform layer. If these lightly stained cells had been transynaptically-stained amacrine cells, no such fiber could have been found (Young, 1962b). Depth of staining, although related to axon diameter, is not a simple function of axon diameter. Interestingly, nearly all the deep centrifugal cells stained only a light or dark brown. They never approached the intensity of the darkest inner granule centrifugal cells.
Many details regarding photoreceptor terminations and centrifugal cell structure first elaborated by silver and Golgi methods have been confirmed by cobalt staining. Three classes of photoreceptor terminations within the optic lobe, (types I, II and III) have been described (Young, 1962b). Types I and II both form enlargements (terminal bags) (Insert, Fig. 1 B) when they enter the plexiform layer, and terminate in the first radial or second tangential laminae respectively (Young, 1962b; Dilly et al., 1963; Case et al., 1972; Cohen, 1973b; 1976). Cobalt-stained photoreceptor axons duplicate this terminal stratification in the plexiform laminae.
Type-III photoreceptor endings were as elusive in this material as in other reports (Young, 1962b, 1971). It is possible that a) the time required for staining types I and II and the centrifugal cells was not sufficient for type III, b) type-III fibers are rarer in this species than in Octopus vulgaris of c) fibers previously identified as type III were incomplete branches of deep medulla or more central centrifugal cells. Not enough information is available from this or previous reports to resolve this point.
Centrifugal cells of the inner granule layer identified in this study have four characteristics that compare favorably with Golgi-identified centrifugal cells (Young, 1962b, 1971, 1974): (1) Location in the inner granule layer; (2) Increasing cell body diameter with depth from the surface (of the inner granule layer); (3) Variations in cell shape, i.e., unipolar, etc. (compare Fig. 4 with Figs. 36-38 in Young, 1962b); (4) Characteristic dendritic branching in the inner tangential layers and especially the layer containing Type II photoreceptor endings (Fig. 4A-G) (Young, 1962b, 1971). However, the fine details, such as total dendritic spread, synaptic enlargements, etc. were difficult to completely discern due to a background coloration that results from Timm's intensification.
Retinal nerves terminate in dorsoventrally oriented wedge-shaped slabs of various widths within the cortex of the optic lobe (Fig. 2). It had already been suspected that retinal bundles project to discrete areas in the plexiform layer because a sharp border between normal and degenerated terminals in the plexiform

Cortical Organization in Octopus Optic Lobe 471
layer was produced after cutting several bundles (Young, 1962b). Each slab corresponds to the projection area from a single retinal nerve bundle. The slabs are not identically wide for all retinal nerve bundles because the number of fascicles per bundle is not identical. Parceling a retinal nerve bundle into individual fascicles would only decrease the size of the slab because individual facicles pass radially, not laterally, through the outer granule cell layer (Young, 1971). Intertwining of fascicles from adjacent retinal nerve bundles apparently does not occur. As predicted from the decussation of retinal nerve bundles at the chiasma, a region of dorsal retina terminates in a region of ventral lobe (young, 1971).
Growth in the octopus retina adds to the absolute number of equal-size photoreceptors (Packard, 1969). Optic lobes also enlarge with age. The growth patterns of neither photoreceptor nor optic lobe have been determined. The organization of the cortex of the optic lobe demonstrated in this paper places a constraint on possible modes of development. Terminals of new photoreceptors must be added at the ends of a lobe or inserted into a pre-existing topographic retinal map in the lobe. Centrifugal cells must develop in concert to maintain the overlapping organization between the two.
Very little has been published regarding the physiological properties of these centrifugal cells. Lettvin and Maturana (1961) found that changes in state of visual, equilibrium, and somatosensory systems activated different units recordable from proximal branches of a retinal nerve bundle. Lange and Hartline (1974) recorded from their isolated optic-lobe preparations from squid eyes both visual centrifugal units with large (30 ~ receptive fields and other units non-visually driven. I have also confirmed in the intact animal centrifugal activity in proximal stumps of cut retinal nerve bundles responsive to light, vibratory and tactile stimulation (manuscript in prep.). It would be particularly interesting to correlate the different cell morphologies with the different sensory modalities, but a detailed form-function study has not yet been performed in this system.
Each retinal nerve bundle appears to define within the optic lobe an area where afferent photoreceptor input to centrifugal cells occurs. The characteristic branch in the second tangential layer emphasizes an apparent retinal type II input to the centrigual cells. The physiological reports of centrifugal activity alluded to above (Lettvin and Maturana, 1961; Saidel, pers. obs.) were obtained by recording from axons found in proximal portions of cut retinal nerve bundles. Since in these reports, direct retinal input to the centrifugal cell being recorded is absent due to severing the nerve bundle from the retina, visually active centrifugal cells must also be gathering photoreceptor information via an indirect path. The locus of this path is unknown although one might hypothesize that it lies in interconnections within the inner granule layer or perhaps dendro-dendritic interactions within the plexiform layer. Visually responsive optic lobe centrifugal cells than have the ability to signal to their postsynaptic sites within the retina both the photoreceptor activity from a restricted region of the retina, as defined by the overlapping photoreceptor terminals within a slab, and a diffuse retinal area.

472 W.M. Saidel
References
Alexandrowicz, J.S.: Contribution a l'&ude des muscles, des nerfs et du m~canisme de l'accomodation de l'oeil des ctphalopodes. Archs. Zool. Exp. Gen. 66, 71-134 (1927)
Cajal, S.R.: Contribucitn al conocimiento de la retina y centros 6pticos de los cefaloptdos. Trab. Lab. Invest. Biol. Univ. Madr. 15, 1-82 (1917)
Case, N.M.: Ultrastructural observations on the cortex of the optic lobe of the brain of Octopus and Eledone. In: Principles in Vision. (R. Zettler and R. Weiler, eds.) Berlin-Heidelberg-New York: Springer-Verlag 1976
Case, N.M., Gray, E.G., Young, J.Z.: Ultrastructure and synaptic relations in the optic lobe of the brain of Eledone and Octopus. J. Ultrastruct. Res. 39, 115-123 (1972)
Cohen, A.I.: An ultrastructural analysis of the photoreceptors of the squid and their synaptic connections. II. Intraretinal synapses and plexus. J. Comp. Neurol. 147, 379-398 (1973a)
Cohen, A.I.: An ultrastructural analysis of the photoreceptors of the squid and their synaptic connections. III. Photoreceptor terminations in the optic lobes. J. Comp. Neurol. 147, 399-426 (1973b)
Dilly, P.N., Gray, E.G., Young, J.Z.: Electron microscopy of optic nerves and optic lobes of Octopus and Eledone. Proc. Roy. Soc. B. 158, 44~456 (1963)
Goodman, C.: Anatomy of the ocellar interneurons of acridid grasshoppers. I. The large interneurons. Cell Tissue Res. 175, 183-202 (1976)
Gray, E.G.: A note on synaptic structure of the retina of Octopus vulgaris. J. Cell Sci. 7, 203-215 (1970) Humason, G.L. :Animal Tissue Techniques. 4th edition, San Francisco: W.H. Freeman and Co. 1979 Kopsch, F.: Mitteilungen tiber das Ganglion opticum der Cephalopoden. Int. Mschr. Anat. Physiol. 16,
33-54 (1899) Lange, G.D., Hartline, P.H.: Retinal responses in squid and octopus. J. Comp. Physiol. 93, 19-36 (1974) Lenhossek, M. von: Histologische Untersuchungen der Cephalopoden. Arch. Mikr. Anat. 47, 45-120
(1896) Lettvin, J.Y., Maturana, H.R.: Octopus vision. Quart. Prog. Rept. of the Res. Lab. Elec., M.I.T. No. 61,
194-209 (1961) Lund, R.D.: Centrifugal fibers to the retina of Octopus rulgaris. Exp. Neurol. 15, 100-i 12 (1966) Packard, A.: Visual acuity and eye growth in Octopus vulgaris (Lamarck). Monit. Zool. Ital. (N.S.) 3,19-
32 (1969) Strausfeld, N.I., Obermayer, M.: Resolution of intraneuronal and transynaptic migration of cobalt in
the insect visual and central nervous systems. J. Comp. Physiol. 110, 1-12 (1976) Tyrer, N.M., Bell, E.M.: The intensification of cobalt-filled neurone profiles using a modification of
Timm's sulphide-silver method. Brain Res. 73, 151-155 (1974) Wells, M.J.: Octopus: Physiology and Behavior of an Advanced Invertebrate. London: Chapman and
Hall 1978 Young, J.Z.: The retina of cephalopods and its degeneration after optic nerve section. Phil. Trans. B. 245,
1-18 (1962a) Young, J.Z.: The optic lobes of Octopus vulgaris. Phil. Trans. B. 245, 19-58 (1962b) Young, J.Z.: The Anatomy of the Nervous System of Octopus vulgaris. Oxford: Clarendon Press 1971 Young, J.Z.: The central nervous system of Loligo. I. The optic lobe. Phil. Trans. B. 267, 263-302 (1974)
Accepted October 16, 1979
Copyright © 2022 FDOKUMEN




















