
Effects of two dietary protein levels on energy balance and digestive capacity of Octopus maya
16
ORIGINAL RESEARCH Effects of two dietary protein levels on energy balance and digestive capacity of Octopus maya Carlos Rosas • Ariadna Sa ´nchez • Cristina Pascual • Josue ´ Aguila • Teresita Maldonado • Pedro Domingues Received: 8 February 2010 / Accepted: 12 May 2010 / Published online: 28 May 2010 Ó Springer Science+Business Media B.V. 2010 Abstract Octopus maya is a carnivorous species and protein is the main energy source. During the present study, two different dietary protein levels (40 and 60% CP) were offered to octopuses as specifically designed artificial diets, to determine protein needs and the effects on metabolism. Frozen crab (Callinectes spp.) was used as control. Results obtained demonstrated that crab remains as one of the best diets for O. maya. The artificial diet with 60% CP produced a low but positive growth rate, and at times, a physiological response similar to that observed in octopuses fed crabs. The present results show the capacity of O. maya juveniles to adjust their digestive enzymes to different types of food and protein level, and this appears to be well correlated with octopus growth. General proteases and trypsin from the pancreas were well correlated with growth rates. A low activity was observed in octopuses fed 40% PC diet (negative growth rate), while a high activity was present in octopuses fed 60% CP diet and crabs (low and high growth rate, respectively). In contrast, these same enzymes were inducted in the salivary glands of octopuses fed with the diet that promoted weight loss (40% CP diet), while a reduced activity was observed in octopuses fed crabs. Energy budget indicates that the animals ingested more than 1,000 kJ week -1 kg -1 ; with such energy, octopuses should satisfy their physiological demands such as was observed when animals were fed crab C. Rosas A. Sa ´nchez C. Pascual Facultad de Ciencias, Universidad Nacional Auto ´noma de Me ´xico, Unidad Multidisciplinaria de Docencia e Investigacio ´n, Administracio ´n de correos 1080312003, C.P. 97350 Hunucma ´, Yucata ´n, Me ´xico J. Aguila Posgrado en Ciencias del Mar, Facultad de Ciencias, UNAM, Puerto de abrigo s/n, Sisal, Yucata ´n, Me ´xico T. Maldonado Facultad de Ciencias Quı ´mico Biolo ´gicas, Universidad Autonoma de Campeche, Av. Agustı ´n Melgar s/n, C.P. 24030 Campeche, Me ´xico P. Domingues (&) Centro Oceanogra ´fico de Vigo, IEO, Canido, Cabo Estai, 36390 Vigo, Spain e-mail: [email protected] 123 Aquacult Int (2011) 19:165–180 DOI 10.1007/s10499-010-9350-7
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effects of two dietary protein levels on energy balance and digestive capacity of Octopus maya

ORI GINAL RESEARCH
Effects of two dietary protein levels on energy balanceand digestive capacity of Octopus maya
Carlos Rosas • Ariadna Sanchez • Cristina Pascual • Josue Aguila •
Teresita Maldonado • Pedro Domingues
Received: 8 February 2010 / Accepted: 12 May 2010 / Published online: 28 May 2010� Springer Science+Business Media B.V. 2010
Abstract Octopus maya is a carnivorous species and protein is the main energy source.
During the present study, two different dietary protein levels (40 and 60% CP) were
offered to octopuses as specifically designed artificial diets, to determine protein needs and
the effects on metabolism. Frozen crab (Callinectes spp.) was used as control. Results
obtained demonstrated that crab remains as one of the best diets for O. maya. The artificial
diet with 60% CP produced a low but positive growth rate, and at times, a physiological
response similar to that observed in octopuses fed crabs. The present results show the
capacity of O. maya juveniles to adjust their digestive enzymes to different types of food
and protein level, and this appears to be well correlated with octopus growth. General
proteases and trypsin from the pancreas were well correlated with growth rates. A low
activity was observed in octopuses fed 40% PC diet (negative growth rate), while a high
activity was present in octopuses fed 60% CP diet and crabs (low and high growth rate,
respectively). In contrast, these same enzymes were inducted in the salivary glands of
octopuses fed with the diet that promoted weight loss (40% CP diet), while a reduced
activity was observed in octopuses fed crabs. Energy budget indicates that the animals
ingested more than 1,000 kJ week-1 kg-1; with such energy, octopuses should satisfy
their physiological demands such as was observed when animals were fed crab
C. Rosas � A. Sanchez � C. PascualFacultad de Ciencias, Universidad Nacional Autonoma de Mexico, Unidad Multidisciplinaria deDocencia e Investigacion, Administracion de correos 1080312003, C.P. 97350 Hunucma, Yucatan,Mexico
J. AguilaPosgrado en Ciencias del Mar, Facultad de Ciencias, UNAM, Puerto de abrigo s/n, Sisal, Yucatan,Mexico
T. MaldonadoFacultad de Ciencias Quımico Biologicas, Universidad Autonoma de Campeche,Av. Agustın Melgar s/n, C.P. 24030 Campeche, Mexico
P. Domingues (&)Centro Oceanografico de Vigo, IEO, Canido, Cabo Estai, 36390 Vigo, Spaine-mail: [email protected]
123
Aquacult Int (2011) 19:165–180DOI 10.1007/s10499-010-9350-7

(I = 1,300 kJ week-1 kg-1; P = 834 kJ week-1 kg-1). However, a very low digested
energy was observed in octopuses the fed artificial diets, indicating that these could have a
factor that limits digestibility.
Keywords Energy balance � Enzymes � Lipids � Metabolism � Octopus maya � Protein
Introduction
Octopus maya is a carnivorous species and protein is the main energy source (Van
Heukelem 1983; Segawa and Hanlon 1988). As for other cephalopods, this species is
characterized by a low O/N ratio, indicating protein as the main metabolic substrate
(Segawa and Hanlon 1988). Higher growth rate and assimilated energy was observed in
individualized O. maya fed on frozen crab, in comparison with octopuses fed on shrimp
pellet with 21% lipid (Domingues et al. 2007). Domingues et al. (2010) reported growth
rates *1% body weight day-1 at culture densities ranging between 4 and 25 kg m-3.
Similar results were reported for O. vulgaris (Segawa and Hanlon 1988; Iglesias et al.
2000; Aguado and Garcıa-Garcıa 2003) and Sepia officinalis (Domingues and Marquez
2010).
To elaborate an acceptable artificial diet for cephalopods, it is necessary to know the
digestive route physiology, characterizing the food route through the digestive tract; this
was previously studied on octopus species (Bouche-Rodoni 1975; Boucaud-Camou et al.
1976; Boucher-Rodoni et al. 1987). Once ingested food is ducted by peristaltic movements
to the stomach, it is decomposed by digestive enzymes. The chymo passes to the crop
where digestion is finished and then absorbed into the digestive gland, where intracellular
digestion occurs. The main organs that segregate proteolytic enzymes are the salivary
glands, digestive gland (through pancreas), crop and digestive channels (Morishita 1974;
Boucaud-Camou et al. 1976). Morishita (1974) showed that besides the pancreas, posterior
salivary glands have an important role in octopus protein digestion. This is common to
several cephalopods species (Koslovskaya and Vaskovsky 1970). Nutrients, mainly pro-
tein, are absorbed and stored in the digestive gland and muscle, where they are used for
growth. (Houlihan et al. 1990) indicated that at a maximum growth rate of 6% day-1, over
90% of synthesized protein was retained for growth in O. vulgaris, demonstrating that a
high proportion of ingested protein is stored in the tissues. Therefore, cephalopod energetic
reserves depend on dietary proteins which, through amino acid metabolism and other
molecules such as glycogen and lactate, are involved in growth (Akagi and Ohmori 2004).
Besides fast growth, O. maya easily adapts to captivity and has a high market price and
high feeding efficiency (Rosas et al. 2007). Although O. maya has low fecundity, the
holobenthic hatchlings allow the obtention of juveniles without the massive mortality
observed in other species with paralarvae phases (Van Heukelem 1983). O. maya also
accepts artificial diets (Domingues et al. 2007; Rosas et al. 2007). Our research group
firstly used an artificial for O. maya based on a shrimp pellet enriched with squid paste
(Domingues et al. 2007; Rosas et al. 2007). This diet had 40% CP and 21% lipids and
covered maintenance energy requirements. Lipids were the limiting factor of octopus
growth with that diet. Based on those results and taking into consideration protein needs for
metabolism, two different dietary protein levels (40 and 60% CP) were assayed in the
present study and offered to octopuses as specifically designed artificial diets. Frozen crab
(Callinectes spp.) was used as a control diet. The form in which dietary protein level
166 Aquacult Int (2011) 19:165–180
123

modulates the digestive capacity, the accumulated energy in tissue and energy budget of
octopuses was evaluated through digestive enzyme activity (proteases in salivary glands:
anterior and posterior, pancreas and crop), energy content (muscle and digestive gland),
food ingestion, oxygen consumption, ammonia excretion and growth of juveniles of
O. maya. Glycogen and total lipids accumulated in muscle and digestive gland were
measured in order to determine how nutrients modulate reserves at tissues level.
Materials and methods
Animals
Specimens of O. maya were caught using artisanal lines, with live crabs as bait, in front of
Sisal harbour (Yucatan, Mexico). Octopuses were transported to the laboratory situated
300-m inland, in 120-l circular tanks with sea water. A total of 30 octopuses were used for
the experiments. In the laboratory, octopuses were placed in individual flow-through 80-l
tanks with aerated sea water. A PVC tube (400 in diameter) was placed in each tank as a
refuge. Ten octopuses were fed the diet 40% PC, 10 were fed the diet 60% PC, and 10 were
fed frozen crabs (Callinectes spp.). All groups had similar weights (P [ 0.05) at the start of
the experiment: 446.1 ± 41.3, 458.7 ± 40.7, and 458.8 ± 64.4 g, for 40% PC and 60%
PC artificial diets, and frozen crab, respectively. Natural filtered sea water (5 lm) flowed at
120 l h-1, allowing a complete renewal in each tank every 40 min. During the experiment,
sea water in the tanks was maintained at 28 ± 2�C, salinity at 36 ppt, dissolved oxygen
[5 mg l-1 and pH [ 8. Each tank was covered with a green solid net sowed around the
top of the tank, to prevent octopuses from escaping. The experiment lasted for 40 days.
Diets preparation
The experimental diets were prepared by thoroughly mixing ingredients with oil and then
adding water until a semi-humid diet resulted (Table 1). The artificial diets were prepared
on a weekly basis and frozen until utilization. Energy content was measured in a calori-
metric pump (Parr@) calibrated with benzoic acid. Octopuses were fed twice a day (0900 h
and 1700 h) between 7 and 10% animal dw day-1 for all regimes. This ration is considered
to be an adequate feeding rate for this species and cephalopods in general, at these rearing
temperatures (Van Heukelem 1983).
Enzymatic activity
At the end of the experiment, enzymatic activity was measured individually for each diet.
Salivary glands (anterior and posterior), pancreas and crop were dissected and placed at
-40�C until analysis. All the animals were fasted 12 h before sampling. Frozen samples
were homogenized at 4�C in 500-ll ice-cold pyrogen free water. Homogenates were
centrifuged at 13,200 rpm for 20 min at 4�C. Supernatant was diluted in ten volumes of
ice-cold pyrogen-free water. Homogenates were immediately used for enzyme analysis.
The soluble-protein content was measured in diluted homogenates (Bradford 1976) using
the Bio-Rad protein determination kit (Biorad@-500-0006). Samples were read in a Biorad
model 550 micro plates reader at 495 nm. Assays were triplicated for each sample. General
proteases activity was measured in homogenates using azocoll (Sigma A-4341) as substrate
Aquacult Int (2011) 19:165–180 167
123

in phosphate buffer, with pH 7. Absorbance was read in a spectrophotometer Spectronic
model 21D at 520 nm. One unit is defined as the amount of enzyme which catalyze the
release of azo dye causing a DA/Dt = 0.001 min-1. Assays were triplicated for each
sample. Trypsin activity was measured in homogenate diluted (1:100) using benzoil-
arginina-paranitro-anilide (BAPNAl 100 mM) as a substrate in buffer pH 7.5, 4�C (0.1 M
TRIS, NaCl 0.05 M, pH 7.5). Absorbance was read at 405 nm. Leucin amino peptidase
was measured in an homogenate diluted (1:10) using L-Leucine-p-nitroanilide (LPNA)
100 mM as a substrate in buffer pH 8 (0.1 M TRIS, pH 8; 25�C). Absorbance was read at
410 nm.
Glycogen, total lipids and energy content of the digestive gland and muscle
After measuring the total weight, the muscle and digestive gland were homogenized at 4�C
for 5 min. Glycogen in the digestive gland and muscle was extracted in the presence of
sulfuric acid and phenol (Dubois et al. 1965). Tissues were first homogenized in ice-cold
trichloroacetic acid (TCA, 5%) at 3,340g. After centrifugation (7,000g), the supernatant
was quantified; this procedure was done twice. One milliliter of supernatant was pipetted
into a tube and mixed with 5 volumes of 95% ethanol. Tubes were placed in an oven at 37–
40�C for 3 h. After precipitation, the tubes were centrifuged at 7,000g for 15 min. Gly-
cogen pellet was dissolved by adding 0.5 ml of boiling water. Then, 5 ml of concentrated
sulfuric acid and phenol (5%) were added and mixed. Tubes content were transferred to
micro plates and read at 490 nm in a micro plate reader (Bio-Rad 550). Extraction of total
lipids followed the (Bligh and Dyer 1959) method. After phase equilibration, the lower
Table 1 Feeds composition forO. maya. g/100 g
a Vitamin premix withoutvitamin C, provided byAgribrands de Mexicob Ascorbyl phosphate (stay-C-35%; Roche)
Ingredients Protein level, %
40 60 Crab meat
Menhaden fish meal 27.3 41.0
CPSP70, fish hidrolizate: BIOCP@ 3.3 5.0
PROFINE 6.7 10.0
Squid meal 6.7 10.0
Dried whey 13.2 20.0
Soybean lecithin 1.0 1.0
Fish oil 1.6 0.0
Wheat starch 28.8 4.0
Vitamin mixa 1.0 1.0
Vitamin Cb 0.5 0.5
Carophill red 0.0 0.0
Filler 8.5 6.5
Alginate Na 1.0 1.0
Dry base, %
Protein 40.0 60.0 58.0
Lipids 6.0 6.1 4.7
Ash 6.2 9.4 22.0
Fiber 0.8 1.1 2.5
kJ g-1 9.0 13.0 16.8
168 Aquacult Int (2011) 19:165–180
123

chloroform layer (total lipids) was removed, concentrated, and weighed. Total lipids were
expressed as mg of lipids g-1 fresh tissue analyzed.
Energy content of the digestive gland and muscle was determined using a calorimetric
pump (Parr@) calibrated with benzoic acid. Samples of both tissues were dried at 60�C
until constant weight.
Energy budget
Food ingestion and absorption efficiency
Every day, uneaten food was removed, dried and weighed after 4 h in the tanks to
determine ingestion and food conversion. The crab given to the octopuses was calculated in
a dry weight basis considering that 100-g fresh crab corresponded to 18 g of dry biomass
without shell. Therefore, both artificial diets and fresh crab were offered in similar rations,
considering the real dry biomass.
Feces were collected once a day, early in the morning, and immediately dried in the
oven for determination of water content and calorimetric value. The ingestion rate (IR) was
calculated as the difference between the delivered and the food remaining after 4 h and
corrected by the percentage of leached nutrients. The leaching percentage of the pelletized
diets was measured using the shaking method (Obaldo et al. 2002). For this, 1 g of pelleted
feed was placed in 250-ml flask on a horizontal shaker at 25�C during 2 h. Afterward, all
the water was filtered through a Whatman filter number 3 (5 l) to separate the remaining
pellet from leached water. The leached and original feed samples were dried in a con-
vection oven at 105�C for 24 h and then cooled in a desecrator. Dried feed samples were
weighed and analyzed for dry matter retention. Pellet stability was calculated as the ratio of
dry matter retention after leaching and dry matter of original samples, expressed as a
percentage.
The absorption efficiency of the diet (AE) was determined by using feces recovered
from octopuses fed each diet. Feces were dried at 60�C until constant dry weight; part of
the dry feces was used to determine caloric value and part to determine ash content. The
AE was determined using the equation: AE = [(DWAF/DWd – DWAF/DWf) / ((1 –
DWAF/DWf) DWAF/DWd)] 9 100, where DWAF/DWd is the ratio between the dry
weight ash free and the dry weight of the diet, and the DWAF/DWf is the ratio between dry
weight ash free and dry weight of feces. The energy absorbed (kJ week-1 kg-1 live)
by octopuses was calculated as: Ab = AE 9 I 9 FE where I is the ingestion rate (g dry
weight-1 week-1 kg-1 live) and FE is the food energy content expressed as kJ g-1 dry
weight. Food and feces energy content were determined with a calorimetric pump (Parr@)
calibrated with benzoic acid.
All feces were pooled by diet and dried at 60�C until analysis. Feces energy was
obtained from three samples per diet burned in the calorimetric pump. Total feces energy
was expressed as kJ g-1 of dry weigh feces.
Oxygen consumption
The effect of the all diets on the physiological condition was measured in nine animals
from each diet. Octopuses were placed in individual respirometric chamber (80 l) in order
to measure oxygen consumption and nitrogen excretion. Animals were deprived of food for
12 h and placed individually in 80-l chambers. Tanks were sealed and connected to a flow-
through sea water recirculation system (Rosas et al. 1993). To prevent handling stress to
Aquacult Int (2011) 19:165–180 169
123

affect readings, octopuses were conditioned in the chambers for 12 h. Oxygen consump-
tion was calculated as the difference in concentrations of the dissolved oxygen between the
input and output of each chamber, timing the water flow. Dissolved oxygen was measured
using the polarographic electrode of a digital oxygen meter (YSI 50B ± 0.01 mg l-1). The
readings were corrected with respect to the control chamber, without octopuses. Oxygen
consumption was obtained every hour (between 0930 and 1830 h) during a 2-day period.
The first day was considered as a fasting condition, and the second day animals were fed
after the first (0930 h) oxygen consumption measurement. Animals and the control
chamber were fed with the frozen crab or the artificial diet, using a ration between 7 and
10% of ww day-1. At the end of the second day, octopuses were weighed. Routine
metabolism (Rrout) was estimated from VO2 (mg g-1 h-1) of unfed octopuses considering
the time during the day in which octopuses were not fed. Apparent heat increase (RAHI;
J g-1 h-1) was estimated from the difference between VO2 of unfed octopuses and
maximum value attained after feeding and considering the time needed for peak oxygen
consumption after feeding and the number of rations fed to octopuses per day (n = 2)
during the previous experiment. A conversion factor of oxygen consumption of
14.3 J mg-1 was used to transform unfed and fed VO2 to J g-1 wet weight (ww) (Lucas
1993). The AHI coefficient was calculated as RAHI/I 9 100. Respiration (R) was calculated
as Rrout ? RAHI and expressed as joules day-1 g-1 ww, considering that octopuses were
fed twice a day during the growth trials.
Nitrogen excretion was measured both before and after feeding, at the same time as
oxygen consumption. Ammonia concentration was analyzed using the colorimetric indo-
phenol method (APHA 1995). The U values were obtained transforming ammonia values
using the value of 20.5 J mg-1 of ammonia excretion (Lucas 1993). The atomic ratio of
O:N was estimated for both fasting and feeding animals, and recorded values of oxygen
consumption and ammonia excretion transformed to lg At h-1 g-1 dw. Feeding O:N ratio
was obtained using the maximum oxygen consumption and nitrogen excretion during the
experimental period.
Energy budget was estimated using the following equations (Lucas 1993):
I = Ab = U ? R ? P, where I is the ingested gross energy, Ab is the energy absorbed or
digestive energy (DE), U the energy lost in nitrogen products, R indicates respiration
(R = Rrout ? RAHI), and P is energy invested in biomass production. Assimilated energy
(AS) was calculated as R ? P. All values were expressed as kJ week-1 kg-1. Energy
produced (P) was calculated using the actual growth rate of the octopuses obtained during
the experimental time (40 days). The value of 10.09 ± 1.3 kJ g-1 dw was used to trans-
form the growth data into production units (P; kJ week-1 kg-1 live). This value was
obtained from analyzing energy content applied to 25 octopuses by means of a calorimeter
(Parr@), previously calibrated with benzoic acid. Assimilated, respiratory and production
gross efficiencies were calculated as As/C 9 100, R/C 9 100 and P/C 9 100, respec-
tively. Respiratory (R) and production net efficiencies (PE) were calculated as R/As 9 100
and P/As 9 100, respectively.
Statistical analysis
One-way ANOVA’s were applied to results of growth rate, survival and energetic balance.
Two-way ANOVA’s were applied to results of enzyme activity, glycogen, total lipids and
energy content to determine the effects of all diets in each analyzed tissue. A visual
inspection of the variances with the purpose of verifying its homogeneity was made
170 Aquacult Int (2011) 19:165–180
123

previously; on the contrary, the data were transformed. An arc sin transformation was used
before processing percentage data (Zar 1974).
Results
Growth rate and survival
Survival was of 70% for octopuses fed the 40 CP diet, 90% for octopuses fed the 60 CP
diet and 80% for octopuses fed crabs. Octopus fed crabs had a higher growth rate
(SGR = 1.77% day-1) than those fed the artificial diets (SGR = -0.43% day-1 and
0.08% day-1, for octopuses fed the 40% CP and the 60% CP diets, respectively) (Table 2;
P \ 0.05).
Enzyme activity
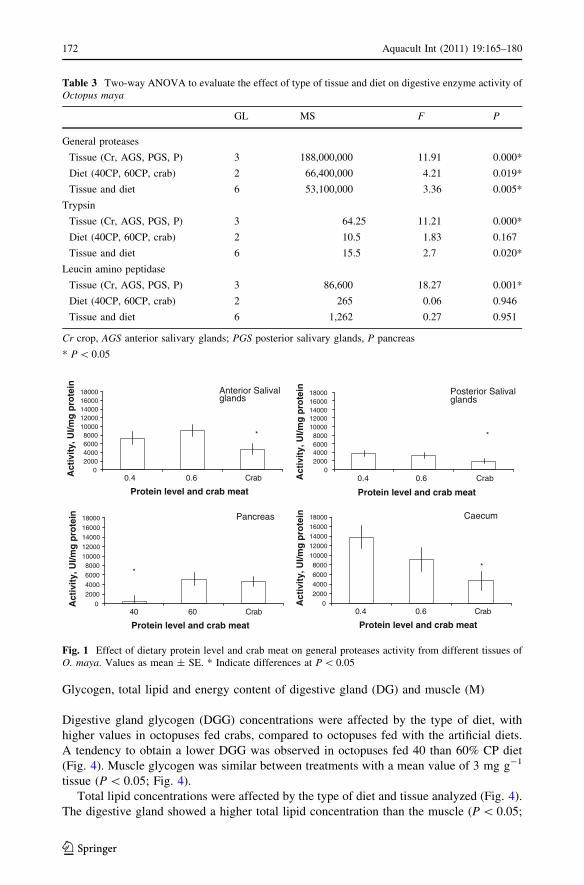
General protease activity was influenced both by diets and by tissues analyzed (Table 3).
Protease activity in the anterior salivary glands and crop was higher than in the pancreas
and posterior salivary glands. Octopuses fed with the compounded diets had higher pro-
tease general activity than octopuses fed with crabs. Interaction observed indicates that
general proteases in the different tissues were affected in different ways by each diet.
A low general proteases activity was observed in salivary glands (anterior and posterior)
and crop of octopuses fed with crabs. However, in the pancreas, an inverse relationship was
observed: a high protease activity was registered for octopuses fed 60% CP and crab
(Fig. 1).
A significant interaction between diet and trypsin activity was observed (Table 3).
A higher trypsin activity was found in cecum in comparison with that obtained for the
anterior salivary glands, pancreas and posterior salivary glands (P \ 0.05). A low trypsin
activity was observed in the posterior salivary glands and cecum of octopuses fed crabs,
while in the pancreas an inverse relationship was observed: a high proteases activity was
registered in octopuses fed crab (Fig. 2).
Tissues had different leucine amino peptidase activity, but they were not affected by the
diet. Leucine amino peptidase activity was found higher in crop than in other tissues
(P \ 0.05; Fig. 3).
Table 2 Effect of dietary protein level and fresh crab meat on specific growth rate and survival of O. mayawild juveniles. Mean ± SE
Dietary protein level, %
40 60 Crab
Initial weight, g 446.1 ± 41.3a 458.7 ± 40.7a 458.8 ± 64.4a
Final weight, g 375.7 ± 29.1a 473.8 ± 17.3b 931.3 ± 72.3c
N 10 10 10
Time, days 40 40 40
Survival, % 70 90 80
SGR, % day-1 -0.43 ± 0.05a 0.08 ± 0.05b 1.77 ± 0.05c
Different letter between columns means P \ 0.05
Aquacult Int (2011) 19:165–180 171
123
Glycogen, total lipid and energy content of digestive gland (DG) and muscle (M)
Digestive gland glycogen (DGG) concentrations were affected by the type of diet, with
higher values in octopuses fed crabs, compared to octopuses fed with the artificial diets.
A tendency to obtain a lower DGG was observed in octopuses fed 40 than 60% CP diet
(Fig. 4). Muscle glycogen was similar between treatments with a mean value of 3 mg g-1
tissue (P \ 0.05; Fig. 4).
Total lipid concentrations were affected by the type of diet and tissue analyzed (Fig. 4).
The digestive gland showed a higher total lipid concentration than the muscle (P \ 0.05;
Table 3 Two-way ANOVA to evaluate the effect of type of tissue and diet on digestive enzyme activity ofOctopus maya
GL MS F P
General proteases
Tissue (Cr, AGS, PGS, P) 3 188,000,000 11.91 0.000*
Diet (40CP, 60CP, crab) 2 66,400,000 4.21 0.019*
Tissue and diet 6 53,100,000 3.36 0.005*
Trypsin
Tissue (Cr, AGS, PGS, P) 3 64.25 11.21 0.000*
Diet (40CP, 60CP, crab) 2 10.5 1.83 0.167
Tissue and diet 6 15.5 2.7 0.020*
Leucin amino peptidase
Tissue (Cr, AGS, PGS, P) 3 86,600 18.27 0.001*
Diet (40CP, 60CP, crab) 2 265 0.06 0.946
Tissue and diet 6 1,262 0.27 0.951
Cr crop, AGS anterior salivary glands; PGS posterior salivary glands, P pancreas
* P \ 0.05
02000400060008000
1000012000140001600018000
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Anterior Salival glands
02000400060008000
1000012000140001600018000
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Posterior Salival glands
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Caecum
0
2000
4000
6000
800010000
12000
14000
16000
18000
0.4 0.6 Crab 0.4 0.6 Crab
0.4 0.6 Crab40 60 Crab
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Pancreas
Fig. 1 Effect of dietary protein level and crab meat on general proteases activity from different tissues ofO. maya. Values as mean ± SE. * Indicate differences at P \ 0.05
172 Aquacult Int (2011) 19:165–180
123

Fig. 4). Total lipid in octopuses fed crab was higher than in octopuses fed with the artificial
diets (P \ 0.05; Fig. 4). The interaction observed in the ANOVA showed that the type of
diet affected the total lipids in both tissues in a different way: while DG was affected by the
type of diet, M total lipids showed similar values in all dietary treatments (Table 4).
Tissue energy content was affected by the type of diet, with higher values in octopuses
fed crabs (P \ 0.05). Higher values were observed in the digestive gland (DG) compared
to the muscle, in octopuses fed the 40% CP diet and crabs (P \ 0.05). There was no
interaction between energy content in the different tissues for octopuses fed the different
types of diet (P [ 0.05; Table 4).
012345678
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein Anterior Salival
Glands
012345678
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Posterior Salival Glands
012345678
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Caecum
012345678
0.4 0.6 Crab
Protein level and crab meat
Act
ivit
y, U
I/mg
pro
tein
*
Pancreas
0.4 0.6 Crab
0.4 0.6 Crab 0.4 0.6 Crab
Fig. 2 Effect of dietary protein level and crab meat on trypsin activity from different tissues of O. maya.Values as mean ± SE. * Indicate differences at P \ 0.05
0
50
100
150
200
250
Protein level and crab meat
Act
ivit
y m
UI/m
g p
rote
in Anterior Salival Gland
0
50
100
150
200
250
Protein level and crab meat
Act
ivit
y m
UI/m
g p
rote
in
Posterior Salival Gland
0
50
100
150
200
250
Protein level and crab meat
Act
ivit
y m
UI/m
g p
rote
in Caecum
0
50
100
150
200
250
0.4 0.6 Crab 0.4 0.6 Crab
0.4 0.6 Crab40% 60% Crab
Protein level and crab meatAct
ivit
y m
UI/m
g p
rote
in
Pancreas
Fig. 3 Effect of dietary protein level and crab meat on leucine amino peptidase activity from differenttissues of O. maya. Values as mean ± SE. * Indicate differences at P \ 0.05
Aquacult Int (2011) 19:165–180 173
123

Energy budget
Ingestion rate was affected by the type of diet, with lower values for octopuses fed the 60%
CP diet (P \ 0.05) (Table 5). The absorbed energy (Ab) was also affected by the type of
diet, with high values in octopuses fed crabs and the 40% CP diet, and low in animals fed
the 60% CP diet (P \ 0.05; Table 5). The proportion of the ingested food that was digested
(Ab/I, %) was 61, 51, and 82%, for the 40% CP diet, the 60% CP diet and crabs,
respectively (Table 5).
Total ammonia excretion was also affected by the type of diet, with high values in
octopuses fed the 40% CP diet, and low in animals fed crabs. Intermediate values were
registered in octopuses fed the 60% CP diet (P \ 0.05; Table 5). The proportion of
ingested energy that was lost as ammonia excretion (U/I, %) was of 3.7, 2.6 and 1.3%, for
0
20
40
60
80
100
120
0.4 0.6 Crab 0.4 0.6 Crab
UMGD
Protein level and crab meat
Tota
l lip
ids,
mg
/g t
issu
e
c
a a a
bb
0
2
4
6
8
10
12
40% 60% Crab 40% 60% Crab
MGD
Type of diet and tissue
Gly
cog
en, m
g/g
tis
sue
a
aa
a
a
b
10000
12000
14000
16000
18000
20000
22000
24000
40% 60% Crab 40% 60% Crab
MGD
Type of food and tissue
En
erg
y co
nte
nt,
jou
les/
g d
w
a a
b
a a
b
Fig. 4 Effect of dietary protein level on glycogen, total lipids and energy content of digestive gland (DG)and muscle (M) of O. maya. Values as mean ± SE. Different letters mean statistical differences at P \ 0.05
174 Aquacult Int (2011) 19:165–180
123

Table 4 Two-way ANOVA to evaluate the effect of type of tissue and diet on glycogen, total lipids andenergy content on digestive gland and muscle of Octopus maya
GL MS F P
Glycogen
Tissue (DG vs. M) 1 3.56 0.369 0.548
Diet (40CP, 60CP, crab) 2 77.9 8.068 0.001*
Tissue and diet 2 57.63 5.969 0.006*
Total lipids
Tissue (DG vs. M) 1 24,800 121.1 0.000*
Diet (40CP, 60CP, crab) 2 6,725 32.9 0.000*
Tissue and diet 2 5,427 26.5 0.000*
Energy content
Tissue (DG vs. M) 1 20,900,000 5.09 0.029*
Diet (40CP, 60CP, crab) 2 70,000,000 17.04 0.001*
Tissue and diet 2 4,230,000 1.03 0.365
DG digestive gland; M muscle
* P \ 0.05
Table 5 Effect of dietary protein level and fresh crab meat on energetic balance of O. maya. Values asKJ week-1 kg-1 live. Mean ± SE
40% 60% Crab
I 1,381.8 ± 35b 1,065.4 ± 56a 1,300.6 ± 385b
Ab 842.1 ± 62.2b 544.7 ± 124.18a 1,066.1 ± 303.1b
U 50.9 ± 7.3c 27.7 ± 2.8b 16.6 ± 1.26a
As 795.2 ± 119.7b 500.0 ± 34.37a 1,152.3 ± 431.8c
Rahi 560.5 ± 57.5c 179.2 ± 7.42b 100.1 ± 27.4a
Rrut 359.3 ± 35c 294.2 ± 28b 217.8 ± 70a
Rtot 919.8 ± 131.1c 473.4 ± 90.44b 317.9 ± 124.3a
P -124.6 ± 637a 26.6 ± 287b 834.4 ± 441c
Ab/I 60.9 51.1 82.0
AS/I, % 57.6 46.9 88.6
AS/Ab, % 94.4 91.8 108.1
U/I, % 3.7 2.6 1.3
U/Ab, % 6.0 5.1 1.6
R/I, % 26.0 27.6 16.7
R/Ab, % 42.7 54.0 20.4
R/As, % 45.2 58.8 18.9
P/I, % -9.0 2.5 64.2
P/Ab, % -14.8 4.9 78.3
P/As, % -15.7 5.3 72.4
Different letters mean statistical differences at P \ 0.05
I ingested energy, Ab absorbed energy, U energy loose through ammonia excretion, As assimilated energy,Rahi energy loose in apparent heat increment, Rrut routine metabolism, Rtot Rahi ? Rtot, P production
Aquacult Int (2011) 19:165–180 175
123
the 40% CP diet, the 60% CP diet and crabs, respectively (Table 5). The proportion of the
digestible energy that was lost as U was 6.0, 5.0 and 1.6% for the 40% CP diet, the 60% CP
diet and crabs, respectively (Table 5).
Fasting and feeding oxygen consumption were affected by the type of diet, with high
values in octopuses fed the 40% CP diet, and low in animals fed crabs (P \ 0.05; Table 5).
A value three times higher of Rtot was registered in octopuses fed the 40% CP diet,
compared to animals fed crabs (P \ 0.05). An intermediate value was observed in animals
fed the 60% CP diet (P \ 0.05; Table 5). The proportion of the ingested energy that was
used for metabolism (Rtot/I, %) was between 26 and 27% for octopuses fed the artificial
diets and of 17% for animals fed crabs. A higher proportion (R/As, %, between 45 and
58%) was used for respiratory metabolism by octopuses fed the artificial diets, than for
octopuses fed crabs (R/As, % = 19%) (Table 5).
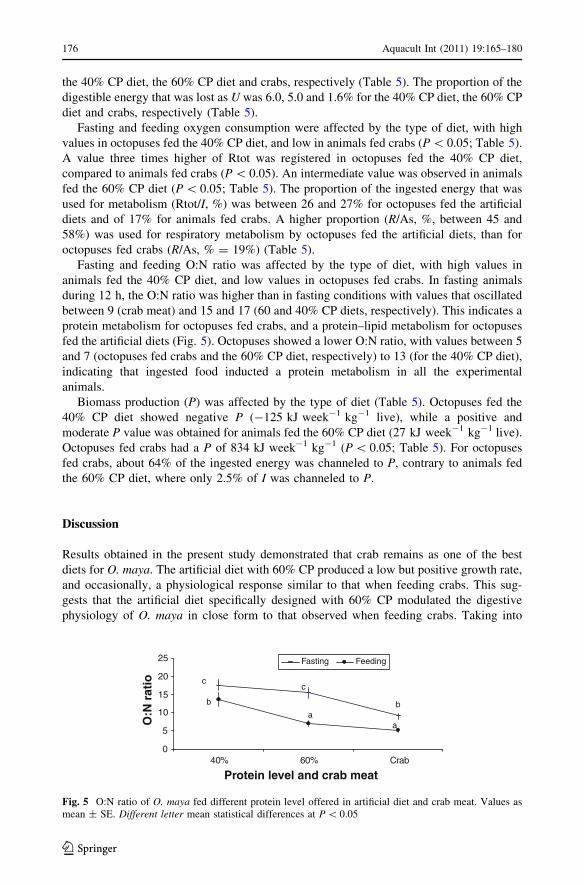
Fasting and feeding O:N ratio was affected by the type of diet, with high values in
animals fed the 40% CP diet, and low values in octopuses fed crabs. In fasting animals
during 12 h, the O:N ratio was higher than in fasting conditions with values that oscillated
between 9 (crab meat) and 15 and 17 (60 and 40% CP diets, respectively). This indicates a
protein metabolism for octopuses fed crabs, and a protein–lipid metabolism for octopuses
fed the artificial diets (Fig. 5). Octopuses showed a lower O:N ratio, with values between 5
and 7 (octopuses fed crabs and the 60% CP diet, respectively) to 13 (for the 40% CP diet),
indicating that ingested food inducted a protein metabolism in all the experimental
animals.
Biomass production (P) was affected by the type of diet (Table 5). Octopuses fed the
40% CP diet showed negative P (-125 kJ week-1 kg-1 live), while a positive and
moderate P value was obtained for animals fed the 60% CP diet (27 kJ week-1 kg-1 live).
Octopuses fed crabs had a P of 834 kJ week-1 kg-1 (P \ 0.05; Table 5). For octopuses
fed crabs, about 64% of the ingested energy was channeled to P, contrary to animals fed
the 60% CP diet, where only 2.5% of I was channeled to P.
Discussion
Results obtained in the present study demonstrated that crab remains as one of the best
diets for O. maya. The artificial diet with 60% CP produced a low but positive growth rate,
and occasionally, a physiological response similar to that when feeding crabs. This sug-
gests that the artificial diet specifically designed with 60% CP modulated the digestive
physiology of O. maya in close form to that observed when feeding crabs. Taking into
0
5
10
15
20
25
40% 60% Crab
Protein level and crab meat
O:N
rat
io
Fasting Feeding
b
cc
b
aa
Fig. 5 O:N ratio of O. maya fed different protein level offered in artificial diet and crab meat. Values asmean ± SE. Different letter mean statistical differences at P \ 0.05
176 Aquacult Int (2011) 19:165–180
123

account results obtained in the present study to day O. maya production is based in a crab
paste bound with gelatine and vitamins as a food (Rosas et al. 2008), because with that
food, it is possible to obtain juveniles of between 30 and 50 g in 130 days after hatch. The
artificial food was well ingested, demonstrating that it is accepted by octopuses in cap-
tivity. In the near future, new formulations will be prepared taking into account that
O. maya could have a protein requirement around 60%.
The present results show the capacity of O. maya juveniles to adjust their digestive
enzymes to different types of food and protein level, and this appears to be well correlated
with octopus growth. Also, the digestive adaptation could involve different reactions,
depending on whether the food satisfies the nutritional requirement or not. Van Worm-
houdt et al. (1980) showed that digestive enzymes can be inducted as a response related to
growth, or in an attempt to obtain more nutrients from a nutritional deficient meal. Results
obtained in the present study show that general proteases and trypsin from the pancreas
were well correlated with growth rates. A low activity was observed in octopuses fed 40%
PC diet (negative growth rate), while a high activity was present in octopuses fed 60% CP
diet and crabs (low and high growth rate, respectively). In contrast, these same enzymes
were inducted in the salivary glands of octopuses fed with a food that promoted weight loss
(40% CP diet), while a reduced activity was observed in octopuses fed crabs. This suggests
that processes involved in digestive enzyme production in different tissues are related to
the octopus capacity to recognize the nutritional characteristics of the ingested food. There
are internal and external stimuli that regulate the activity of the digestive tract of
O. vulgaris (Best and Wells 1983, 1984). Presence of food is by itself enough to stimulate
activity in the gut. When an octopus sees food, the digestive gland begins to secrete,
initiating a nervous stimulus directed to enzyme production. These authors also observed
that salivary glands are stimulated at the same time, producing enzymes that are used to
initiate the external digestion. These authors classified these reactions as external stimuli.
Additionally, with external stimuli, the presence of food in the crop invoked activity in
the salivary glands, suggesting that there are mechanical stimuli in the crop that regulate
the salivary enzyme production. According to the same authors, saliva production was
independent of the type of food because it was produced even when cellulose was used
as inert food. Interestingly, they observed that cellulose did not stimulate activity in the
digestive gland, indicating that the chemical characteristics of the meal could regulate
the secretion of enzymes in the form of proteinaceous ‘‘boules’’. For O. maya, a similar
mechanism could induct the enzymes in the digestive and the salivary glands. The higher
food ingestion registered in octopuses fed with the artificial diet with 40% CP could
mechanically stimulate more saliva secretion, inducing a higher production of enzymes,
and consequently a higher activity of the salivary glands. This could explain why the
salivary glands showed a higher enzyme activity than the other analyzed tissues.
However, due to the nutritional deficiency of the diet actually used, crop could not send
the chemical signals to adequately stimulate the digestive gland, reducing the enzyme
production in the pancreas and presumably in the digestive gland, as observed when
octopuses were fed with the 40% CP. Contrary, an adequate ingestion rate coupled with
a nutritionally complete food (crab meal) could induct a moderated production of
enzymes in the salivary glands and send chemical signals to the digestive gland to induce
enzyme production that allows high assimilation of nutrients and high growth. Only
activities of amino peptidases were not affected by the type of diet, indicating that this
enzyme is not regulated at the same level of trypsin. Interestingly, all digestive enzymes
measured showed a high activity in the cecum, where absorption occurs (Boucaud-
Camou et al. 1976).
Aquacult Int (2011) 19:165–180 177
123

In the present study, O. maya digestive gland reserves correlated well with growth,
accumulating more glycogen, lipid and energy in octopuses fed crabs, compared to the
ones fed the artificial diets. Glycogen is normally produced through a glycogenesis
pathway, via protein metabolism, and stored in the digestive gland (Boucaud-Camou et al.
1976). Cephalopods have the ability to utilize carbohydrate, lipid and protein as metabolic
substrates, with lipid and protein fueling routine aerobic metabolism and carbohydrate
providing energy for burst (anaerobic) activity (Wells and Clark 1996). Glycogen may also
be important for the maturation process and embryo genesis (Rosa et al. 2004). In fact,
there is a correlation between glycogen synthesis and dietary protein level in invertebrates
(Pascual et al. 2006), which suggests in the case of O. maya fed with crabs that crab protein
was adequately digested and absorbed in the digestive gland, promoting synthesis of
reserves. The direct use of protein as an energy reserve may occur in the lack of major
glycogen and lipid reserves in the digestive gland and muscle (O’Dor et al. 1983). For this
reason, it is reasonable to assume that the low glycogen concentration registered in the
digestive gland of octopuses fed 40% CP diet could be associated with the use of reserves
directed to satisfy energy demand that 40% CP feed could not satisfy.
As for O. vulgaris and O. defilipi (Rosa et al. 2004), O. maya had higher lipid con-
centration in the digestive gland than in muscle. As glycogen, lipids are stored in the
digestive gland where they are used also to be supplied as essential fatty acids and
membrane components as phospholipids and cholesterol (Petza et al. 2006). Proximal
analysis revealed that crab meat had only 4% total lipids, indicating that a high growth rate
can be achieved with diets with low lipid levels. The absorption of lipids in cuttlefish Sepiaofficinalis as in octopus is carried out in the digestive gland but is a limited process
(Takahashi 1963). Lee (1994) reported a lipid digestibility for O. vulgaris of 46%, indi-
cating that cephalopods metabolism has evolved to use protein as a source of energy.
Lipids do not play a significant role in energy catabolism and have a mainly structural
function in the tissues. This could explain the limited capacity of cephalopods to oxidize
lipids and the low digestibility observed in O. vulgaris fed fatty fish (O’Dor et al. 1983;
Garcıa and Gimenez 2002; Petza et al. 2006).
Energy budget indicate that animals ingested more than 1,000 kJ week-1 kg-1 and that
with such energy octopuses should satisfy its physiological demands such as was observed
when animals were fed crab (I = 1,300 kJ week-1 kg-1; P = 834 kJ week-1 kg-1).
Rosas et al. (2007) reported an ingested energy of 1,957 kJ week-1 kg-1 and a biomass
production of 1,334 kJ week-1 kg-1 for O. maya feeding on crabs, indicating that such
ingested energy could be enough to produce octopus growth rates averaging 2% day-1.
Although the protein content of the diet could be assumed as the principal responsible of
the differences observed between experimental diets, other differences must be affecting
the use of the nutrients and energy in octopuses fed the artificial diets. According to (Lucas
1993), energy absorbed is a measure of the digestibility of the diet. In the present study, a
digestibility (Ab/I) between 51 and 60% was observed for octopuses fed with the artificial
diets, in contrast to that observed in animals fed with crabs (82%). This suggests that the
actual artificial diets specifically designed for O. maya have a factor that limits digest-
ibility. No similar studies exist in which artificial diets specifically designed for octopuses
were tested, but in a recent study, a digestibility of 71% for E. megalocyathus fed crus-
taceans and a negative growth when octopuses were fed fat mussel were reported (Perez
et al. 2006).
A higher value of R was observed in animals fed 40% CP diet, compared to the ones fed
60% CP diet and crabs. These results can be interpreted as a compensation strategy of
octopuses to invest more energy in an attempt to obtain more nutrients from food, via and
178 Aquacult Int (2011) 19:165–180
123

increment in ingestion rate and/or increment in enzyme production in the salivary glands.
A 10% higher intake energy was invested in respiration (R/I, %) by octopuses fed the
artificial diets, compared to crab, indicating that the processes used to obtain food were
costly in animals fed the artificial diets. Apparent heat increment (Rahi) was high in
animals fed with the artificial diets (diet with 40% CP = 40% of the ingested energy; diet
with 60% CP = 16% of the ingested energy) compared to octopuses fed crab (7% of the
ingested energy).
Results indicate that artificial diets specifically were partially digested, limiting nutri-
ents absorption and growth. A compensatory strategy was observed through the induction
of salivary enzymes production. However, that compensatory mechanism was not enough
to stimulate the physiological digestion in the digestive gland and in consequence, gly-
cogen reserves and lipids were presumably used to satisfy energy demand, which affected
growth. Values of O:N obtained in octopuses fed the 40% CP diet indicate that a protein–
lipid metabolism could be used by these animals, explaining the low lipid reserves
obtained. Therefore, other formulations must be done in attempt to enhance diet digest-
ibility, possibly using more digestible protein sources that provide peptides and amino
acids required for O. maya growth.
Acknowledgments The present study was partially financed by DGAPA-UNAM project No. IN 216006and CONACYT-Basico 2007-24743. Authors also thank to CONACYT for the fellowship awarded to JosueAguila, and appreciate the technical assistance of Darwin Chay, and Julia Tut.
References
Aguado F, Garcıa-Garcıa B (2003) Growth and food intake models in Octopus vulgaris Cuvier (1797):influence of body weight, temperature, sex and diet. Aquacult Int 10:361–377
Akagi S, Ohmori S (2004) Threonine is the best substrate for D-lactate formation in octopus tentacle. AminoAcids 26:169–174
APHA (1995) Standard methods for the examination of water and wastewater, 19th edn. American PublicHealth Association, Washington, DC
Best EMH, Wells MJ (1983) The control of digestion in Octopus. I. The anticipatory response and theeffects of severing the nerves to the gut. Vie Millieu 33:135–142
Best EMH, Wells MJ (1984) The control of digestion in Octopus. II. The role of internal stimulus. VieMillieu 34:1–7
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol37:911–931
Boucaud-Camou E, Boucher-Rodoni R, Mangold K (1976) Digestive absorption in Octopus vulgaris(Cephalopoda: Octopoda). J Zool 179:261–271
Bouche-Rodoni R (1975) Vitesse de digestion chez les cephalopodes Eledone cirrosa (Lamarck) et Illexillecebrosus (Lesueur). Cah Biol Mar 16:159–175
Boucher-Rodoni R, Boucaud-Camou E, Mangold K (1987) Feeding and digestion. In: Boyle P (ed)Cephalopod life cycles. Academic Press, London, pp 85–108
Bradford MM (1976) A refined and sensitive method for the quantitation of microgram quantities of proteinutilizing the principle of protein-dye binding. Anal Biochem 72:248
Domingues P, Marquez L (2010) Effects of culture density and bottom area on growth and survival of thecuttlefish Sepia officinalis (Linnaeus, 1758). Aquacult Int 18:361–369. doi:10.1007/s10499-009-9249-3
Domingues PM, Lopez N, Munoz JA, Maldonado T, Gaxiola G, Rosas C (2007) Effects of an artificial dieton growth and survival of the Yucatan octopus Octopus maya. Aquac Nutr 13:278–280
Domingues P, Garcia S, Garrido D (2010) Effects of three culture densities on growth and survival ofOctopus vulgaris. Aquac Int 18:165–174
Dubois MK, Lilles LA, Hamilton JC, Rebers PA, Smith F (1965) Cholorimetric method for determination ofsugars and related substances. Anal Chem 28:350–356
Aquacult Int (2011) 19:165–180 179
123

Garcıa BG, Gimenez FA (2002) Influence of diet on growing and nutrient utilization in the common octopus(Octopus vulgaris). Aquaculture 211:173–184
Houlihan DF, McMillan DN, Agnisola C, Genoino TI, Foti L (1990) Protein synthesis and growth inOctopus vulgaris. Mar Biol 106:251–259
Iglesias J, Sanchez FJ, Otero JJ, Moxica C (2000) Culture of octopus (Octopus vulgaris, Cuvier): presentknowledge, problems and perspectives. Recent advances in Mediterranean marine aquaculture finifishspecies diversification. Cah Options Mediterr 47:313–322
Koslovskaya EP, Vaskovsky VE (1970) A comparative study of proteinases of marine invertebrates. CompBiochem Physiol 34(137):142
Lee PG (1994) Nutrition of cephalopods: fueling the system. Mar Freshw Behav Physiol 25:35–51Lucas A (1993) Bioenergetique Des Animaux Aquatiques. Masson, ParisMorishita T (1974) Participation in digestion by the proteolotic enzymes of the posterior salivary gland in
Octopus. I. Confirmation of the existence of protein digestive enzymes in the posterior salivary gland.Nippon Suisam Gakkaishi (Bull Jpn Soc Sci Fish) 40:601–607
O’Dor RK, Mangold K, Boucher-Rodoni R, Wells MJ, Wells J (1983) Nutrient absorption, storage andremobilization in Octopus vulgaris. Mar Behav Physiol 11:239–258
Obaldo LG, Divakaran S, Tacon AG (2002) Method for determining the physical stability of shrimp feeds inwater. Aquac Res 33(5):369–377
Pascual C, Sanchez A, Zenteno E, Cuzon G, Gaxiola G, Brito R, Gelabert R, Hidalgo E, Rosas C (2006)Biochemical, physiological, and immunological changes during starvation in juveniles of Litopenaeusvannamei. Aquaculture 251:416–429
Perez MC, Lopez DA, Aguila K, Gonzalez ML (2006) Feeding and growth in captivity of the octopusEnteroctopus megalocyathus. Aquac Res 37:550–555
Petza D, Katsanevakis S, Verriopoulos G (2006) Experimental evaluation of the energy balance in Octopusvulgaris, fed ad libitum on a high-lipid diet. Mar Biol 148:827–832
Rosa R, Costa PR, Nunes L (2004) Effect of sexual maturation on the tissue biochemical composition ofOctopus vulgaris and O. defilippi (Mollusca: Cephalopoda). Mar Biol 145:563–574
Rosas C, Vanegas C, Tabares I, Ramirez J (1993) Energy balance of Callinectes rathbunae Contreras 1930in floating cages in a tropical coastal lagoon. J World Aquac Soc 24:71–79
Rosas C, Cuzon G, Pascual C, Gaxiola G, Chay D, Lopez N, Maldonado T, Domingues PM (2007) Energybalance of Octopus maya fed crab and artificial diet. Mar Biol 152:371–381
Rosas C, Tut J, Baeza J, Sanchez A, Sosa V, Pascual C, Arena L, Domingues P, Cuzon G (2008) Effect oftype of binder on growth, digestibility, and energetic balance of Octopus maya. Aquaculture 275:291–297
Segawa S, Hanlon RT (1988) Oxygen consumption and ammonia excretion rates in Octopus maya, Loligoforbesi and Lolliguncula brevis (Molluscs: Cephalopoda). Mar Behav Physiol 13:389–400
Takahashi T (1963) Studies on the viscera enzymes of cuttlefish Omastrephes sloani pacificus. J Fac FishPrefect Univ Mie 5:384–411
Van Heukelem WF (1983) Octopus maya. Cephalopod life cycles. Academic Press, London, pp 311–323Van Wormhoudt A, Ceccaldi JH, Martin BJ (1980) Adaptation de la teneur en enzymes digestives de
l0hepatopancreas de Palaemon serratus (Crustacea Decapoda) a la composition daliments experi-mentaux. Aquaculture 21:63–78
Wells MJ, Clark A (1996) Energetics: the costs of living and reproducing for an individual cephalopod.Philos Trans R Soc Lond B Biol Sci 351:1083–1104
Zar JH (1974) Biostatistical analysis. Prentice Hall, Englewood Cliff
180 Aquacult Int (2011) 19:165–180
123




















