
Regulation of self-renewal and reprogramming by TCF factors
9
© 2012 Landes Bioscience. Do not distribute. www.landesbioscience.com Cell Cycle 39 Cell Cycle 11:1, 39-47; January 1, 2012; © 2012 Landes Bioscience EXTRA VIEW EXTRA VIEW Key words: Tcf3, Tcf1, Wnt path- way, cell reprogramming, self-renewal, β-catenin, T-cell factor, Tcf7l1, Tcf7 Submitted: 11/04/11 Revised: 11/11/11 Accepted: 11/14/11 http://dx.doi.org/10.4161/cc.11.1.18759 *Correspondence to: Maria Pia Cosma; Email: [email protected] W nt/β-catenin canonical pathway has very important roles both in the regulation of self-renewal and lin- eage differentiation of pluripotent stem cells as well as in the reprogramming of somatic cells. In the absence of Wnt stimulation, the target genes of the path- way are repressed or downregulated. Conversely, when β-catenin is stabilized, there is an accumulation of cytoplasmic and nuclear β-catenin that allows upreg- ulation of its target genes. However, the molecular mechanisms downstream of β-catenin that regulate self-renewal of embryonic stem cells remain unclear. It has been proposed that β-catenin acts through members of the T-cell factor (TCF) family. Recent studies have con- firmed the role of Tcf3 (Tcf7l1) as a tran- scriptional repressor in embryonic stem cells, and they have also proposed a new mechanistic view as to how activation of the Wnt/β-catenin pathway can over- come this repression. Wnt Canonical Pathway Wnt signaling has central roles in many processes during development and disease. Specifically, Wnt pathway participates in bone development, angiogenesis, vascu- lature remodeling, myogenesis, adipo- genesis and stem cell self-renewal, among others processes. 1 WNT genes are highly conserved between vertebrate species, while Wnt proteins comprise a family of secreted glycoproteins (19 in human) that are characterized by several conserved cys- teine residues. Transduction of Wnt sig- nals involves binding of Wnt proteins to Regulation of self-renewal and reprogramming by TCF factors Luigi Ombrato, 1,† Frederic Lluis 1,† and Maria Pia Cosma 1,2, * 1 Centre for Genomic Regulation (CRG) and Universitat Pompeu Fabra (UPF); Barcelona, Spain; 2 Institució Catalana de Recerca i Estudis Avançats (ICREA); Barcelona, Spain † These authors contributed equally to this work. members of two distinct families of cell surface receptors: the Frizzled (Fzd) gene family and the LDL receptor-related pro- tein (LRP) gene family. 2 Historically, the Wnt signaling path- way has been categorized into two major subsets: the canonical and non-canonical Wnt pathways. The canonical Wnt path- way includes a series of subsequent events that results in stabilization and transloca- tion of β-catenin into the nucleus. Non- canonical Wnt signaling involves all of the Wnt-activated cell-signaling path- ways that are not directly correlated with β-catenin stabilization. The activity of the Wnt canonical pathway in stem cell self- renewal and somatic-cell reprogramming is the main focus of this review. In the absence of Wnt binding to its receptor, the cytoplasmic β-catenin protein is degraded by the action of the destruction complex, which is formed by adenomatous polyposis coli (APC), Axin, glycogen synthase kinase 3 (GSK3) and casein kinase I (CK1). Specifically, APC directly binds Axin and β-catenin. In addition to binding APC, Axin inter- acts with GSK3, CK1 and β-catenin. Consequently, the kinases CK1 and GSK3 sequentially phosphorylate β-catenin in its N-terminal region. Phosphorylated β-catenin is recognized by β-TrCP, an E3 ubiquitin ligase subunit, and it can thus be ubiquitinated and then degraded via the 26S proteasome 3,4 (Fig. 1A). The Wnt/β-catenin pathway is acti- vated when a Wnt ligand binds to the Fzd and LRP5/6 cell surface receptors. The formation of the Wnt-Fzd-LRP5/6 com- plex, together with the recruitment of the scaffold protein dishevelled (Dvl), results
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Regulation of self-renewal and reprogramming by TCF factors

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Cell Cycle 39
Cell Cycle 11:1, 39-47; January 1, 2012; © 2012 Landes Bioscience
extra view extra view
Key words: Tcf3, Tcf1, Wnt path-way, cell reprogramming, self-renewal, β-catenin, T-cell factor, Tcf7l1, Tcf7
Submitted: 11/04/11
Revised: 11/11/11
Accepted: 11/14/11
http://dx.doi.org/10.4161/cc.11.1.18759
*Correspondence to: Maria Pia Cosma; Email: [email protected]
Wnt/β-catenin canonical pathway has very important roles both in
the regulation of self-renewal and lin-eage differentiation of pluripotent stem cells as well as in the reprogramming of somatic cells. In the absence of Wnt stimulation, the target genes of the path-way are repressed or downregulated. Conversely, when β-catenin is stabilized, there is an accumulation of cytoplasmic and nuclear β-catenin that allows upreg-ulation of its target genes. However, the molecular mechanisms downstream of β-catenin that regulate self-renewal of embryonic stem cells remain unclear. It has been proposed that β-catenin acts through members of the T-cell factor (TCF) family. Recent studies have con-firmed the role of Tcf3 (Tcf7l1) as a tran-scriptional repressor in embryonic stem cells, and they have also proposed a new mechanistic view as to how activation of the Wnt/β-catenin pathway can over-come this repression.
Wnt Canonical Pathway
Wnt signaling has central roles in many processes during development and disease. Specifically, Wnt pathway participates in bone development, angiogenesis, vascu-lature remodeling, myogenesis, adipo-genesis and stem cell self-renewal, among others processes.1
WNT genes are highly conserved between vertebrate species, while Wnt proteins comprise a family of secreted glycoproteins (19 in human) that are characterized by several conserved cys-teine residues. Transduction of Wnt sig-nals involves binding of Wnt proteins to
Regulation of self-renewal and reprogramming by TCF factors
Luigi Ombrato,1,† Frederic Lluis1,† and Maria Pia Cosma1,2,*1Centre for Genomic Regulation (CRG) and Universitat Pompeu Fabra (UPF); Barcelona, Spain; 2Institució Catalana de Recerca i Estudis Avançats (ICREA);
Barcelona, Spain†These authors contributed equally to this work.
members of two distinct families of cell surface receptors: the Frizzled (Fzd) gene family and the LDL receptor-related pro-tein (LRP) gene family.2
Historically, the Wnt signaling path-way has been categorized into two major subsets: the canonical and non-canonical Wnt pathways. The canonical Wnt path-way includes a series of subsequent events that results in stabilization and transloca-tion of β-catenin into the nucleus. Non-canonical Wnt signaling involves all of the Wnt-activated cell-signaling path-ways that are not directly correlated with β-catenin stabilization. The activity of the Wnt canonical pathway in stem cell self-renewal and somatic-cell reprogramming is the main focus of this review.
In the absence of Wnt binding to its receptor, the cytoplasmic β-catenin protein is degraded by the action of the destruction complex, which is formed by adenomatous polyposis coli (APC), Axin, glycogen synthase kinase 3 (GSK3) and casein kinase I (CK1). Specifically, APC directly binds Axin and β-catenin. In addition to binding APC, Axin inter-acts with GSK3, CK1 and β-catenin. Consequently, the kinases CK1 and GSK3 sequentially phosphorylate β-catenin in its N-terminal region. Phosphorylated β-catenin is recognized by β-TrCP, an E3 ubiquitin ligase subunit, and it can thus be ubiquitinated and then degraded via the 26S proteasome3,4 (Fig. 1A).
The Wnt/β-catenin pathway is acti-vated when a Wnt ligand binds to the Fzd and LRP5/6 cell surface receptors. The formation of the Wnt-Fzd-LRP5/6 com-plex, together with the recruitment of the scaffold protein dishevelled (Dvl), results

© 2012 Landes Bioscience.
Do not distribute.
40 Cell Cycle volume 11 issue 1
Figure 1. the wnt/β-catenin canonical pathway. (a) in the absence of wnt signaling, the destruction complex that is formed by axin, aPC, GSK3 and CKi binds to β-catenin; in turn, CKi and GSK3 phosphorylate β-catenin. this phosphorylation targets β-catenin for proteasome-mediated proteolytic degradation. (B) wnt proteins bind to the Frizzled and LrP5/6 cell surface receptors, which leads to the inhibition of the destruction complex and sup-pression of β-catenin phosphorylation. the stabilized β-catenin can then enter the nucleus and interact with transcriptional regulators, including the tcf family factors. in addition, the cytosolic pool of β-catenin tightly associates with e-cadherin at the cell junctions.

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Cell Cycle 41
provide the greatest benefits reprogram-ming efficiency, suggesting that ad hoc levels of β-catenin are important for the reprogramming, and that different levels of stabilized β-catenin can lead to dif-ferent responses in the same cell through the activation of different genetic pro-grams.25 Interestingly, ESCs cultured with high doses of BIO or CHIR99021, which induce high stabilization levels of β-catenin, and ESC clones expressing high levels of β-catenin both show reduc-tions in cell proliferation,10,26 increased expression of p16 and p1926 and down-regulation of c-Myc.10 This might explain why ESCs that have high β-catenin levels cannot reprogram somatic cell nuclei by cell fusion, given that the inhibition of p53 or of p16INK4a, p15INK4b and p19ARF senes-cence markers causes a significant increase in the efficiency of reprogramming.30-36 However, this cannot completely explain why GSK3-/- ESCs also cannot reprogram somatic cells after fusion;25 indeed, these cells proliferate similarly to wild-type (wt)ESCs.19 These observations suggest two main possibilities: (1) The high β-catenin levels in GSK3-/- ESCs activate gene net-works that are different from those acti-vated in β-catenin-expressing ESC clones; or (2) GSK3-/- ESCs and high-β-catenin-expressing ESC clones share the same mechanism of inhibition of reprogram-ming, which is not reduced proliferation, and which remains to be identified.
All in all, we can conclude that Wnt and LIF pathways are synergistic, even if the activation of the Wnt canonical path-way can maintain self-renewal of ESCs in the absence of LIF,8,16 and β-catenin-/- ESCs express stem-genes when cultured with LIF.22,23 However, how Wnt and LIF pathways crosstalk to maintain self-renewal still needs to be clarified.
Furthermore, Wnt signaling has an essential role in reprogramming. In the presence of LIF, Wnt/β-catenin increases the reprogramming of somatic cells by cell fusion or by direct reprogramming,25,27 which suggests that Wnt signaling might have different activities in the regulation of ESC self-renewal and of reprogramming of somatic cells. Whether or not the repro-gramming process can be accomplished in the absence of β-catenin is an open ques-tion that needs additional studies.
of Wnt signals result in differentiation of ESCs in epiblast stem cells (EpiSCs).18 Furthermore, medium without serum that is supplemented with the MEK inhibitor PD0325901, the GSK3 inhibitor CHIR99021 and LIF (known as 2i-LIF) will also sustain ESC self-renewal.12 Moreover, GSK3-/- ESCs (with a double knockout for the related protein homo-logs GSK3α and GSK3β) show very high levels of stabilized β-catenin and can self-renew in a medium without LIF.19,20 On the other hand, GSK3-/- ESCs are severely compromised in their ability to differ-entiate and cannot form true embryoid bodies or teratomas.19,20 Taken together, these findings show that activation of the Wnt/β-catenin pathway can sustain ESC self-renewal, and that constitutive high activation of this pathway severely reduces differentiation of ESCs.
It has been reported that β-catenin-/- ESCs show defects in the expression of stemness markers.21 However, in sharp contrast, two independent groups showed recently that deletion of β-catenin in ESCs does not affect the self-renewal of ESCs if they are maintained under LIF conditions, while they show minor cell-adhesion defects22,23 (see also below). This suggests that LIF/Stat3 might be the essential signal for the maintenance of self-renewal, while β-catenin might not be essential, if ESCs are cultured in serum-containing medium supplemented with LIF.
Of further interest, when ESCs are cul-tured with Wnt3a or BIO (in a medium with LIF), these ESCs can reprogram somatic cells after polyethyleneglycol (PEG)-mediated fusion or spontaneous fusion.24-26 Similarly, Wnt3a-conditioned medium promotes the reprogramming of mouse embryonic fibroblasts (MEFs) infected with Oct4/Klf4/Sox2 into induced pluripotent stem cells (iPSCs),27 and inhibition of GSK3 by CHIR99021 can increase human somatic cell repro-gramming after Oct4/Klf4 transduc-tion.28,29 However, in cell fusion-mediated reprogramming experiments, it has been shown that reprogramming fails when cell fusion is performed with GSK3-/- ESCs or with ESCs expressing high levels of β-catenin.25 Treatments that caused rela-tively modest increases in β-catenin levels
in LRP5/6 phosphorylation and activa-tion and the recruitment of the destruc-tion complex to the receptors. These events lead to inhibition of the destruc-tion complex and β-catenin phosphoryla-tion, and, thereby, to the accumulation of β-catenin (Fig. 1B).3,4
There are two pools of β-catenin in the cells: one pool is tightly associated with cadherins at cell-cell junctions, and the other is free in the cytosol or nucleus. This latter pool of β-catenin is involved in the regulation of gene transcription. Once β-catenin has been stabilized, it translocates into the nucleus, where it interacts with different Tcf DNA-binding factors; in turn, this complex activates the transcription of its specific target genes5-7 (Fig. 1B).
Small-molecule drugs, such as (2'Z,3'E)-6-bromoindirubin-3'-oxime (BIO) and CHIR99021, can increase β-catenin stabilization and have been used to study the functional role of the Wnt/β-catenin pathway in the regulation of stem cell self-renewal. Both of these compounds inhibit GSK3 phosphoryla-tion activity and thereby act as mimetics of Wnt stimulation.8-11
Regulation of Self-Renewal and Reprogramming
through WNT/β-Catenin and the TCF Factors
The role of the Wnt/β-catenin pathway in self-renewal and reprogramming. Mouse embryonic stem cells (ESCs) can be derived from the inner cell mass of blas-tocysts. They can be maintained under pluripotent conditions by culturing in serum/BMP and cytokine leukemia inhib-itory factor (LIF), which activates signal transducer and activator of transcription 3 (Stat3).12,13 However, the LIF pathway is dispensable for embryonic development before gastrulation, which suggests that there are other signaling cascades that are essential for ESC self-renewal.14,15
Interestingly, addition of Wnt3a to cul-ture medium has been shown to maintain ESC self-renewal.8,16 Likewise, stabiliza-tion of β-catenin with GSK3 inhibitors, such as BIO or lithium chloride, promotes maintenance of both mouse and human ESC self-renewal.8,17 Indeed, inhibition

© 2012 Landes Bioscience.
Do not distribute.
42 Cell Cycle volume 11 issue 1
pluripotency. There are at least three key factors that determine ESC pluripotency: Oct4, Nanog and Sox2. Mouse embryos lacking either Oct4 or Sox2 do not form the epiblast. Overexpression of Nanog enhances ESC self-renewal45 and the expression levels of Oct4 and Sox2 must be maintained within a very narrow range to support pluripotency.46 Importantly, if high levels of Oct4 and Nanog are main-tained, the ability of ESCs to differentiate is reduced,46,47 which means that impor-tant mechanisms that prevent excessive levels of self-renewing factors are neces-sary to allow the cells to differentiate.
Furthermore, genome-wide mapping of the binding sites of these three factors has revealed that they can bind to the pro-moters of several hundred target genes, either alone or cooperatively.48,49
In somatic cells, the overexpression of all or some of these self-renewal factors, such as Oct4, Sox2, Klf4, Nanog and c-Myc, induces the reprogramming of the cells into iPSCs, which have similar fea-tures to ESCs.50-52
One possible effect of the Wnt pathway might be the regulation of the expression of pluripotent genes, such as Oct4 and Nanog. However, β-catenin stabilization by Wnt or BIO treatment, or in GSK3-/- ESCs, has no major effects on the expres-sion of Oct4 and Nanog in ESCs.19,25 Surprisingly, as compared with wtESCs, β-catenin-/- ESCs were shown to have a comparable genome-wide transcriptional profile and, also, comparable Oct4 and Nanog expression and protein levels.23 These data indicate that the regulation of self-renewal by the Wnt pathway and its effects on increased reprogramming are not due to regulation of the transcrip-tional activities of the pluripotent genes.
To define the role of nuclear β-catenin in ESC self-renewal, different groups have focused on the nuclear binding proteins of β-catenin, the Tcf proteins. These are DNA binding factors that can modulate the Wnt canonical sig-naling pathway. Through a highly con-served HMG domain and an N-terminal β-catenin domain, each Tcf protein can regulate the transcription of downstream targets when β-catenin accumulates in the nucleus.53,54
cell-adhesion defects) do not show differ-ences in Oct4 expression with respect to wtESCs when they are maintained under undifferentiated (+LIF) conditions.41 This is in contrast with a recent study in which silencing of E-cadherin in ESCs was per-formed.42 This study showed that reduc-tion in the levels of E-cadherin reduces the levels of Oct4; however, the role or the lev-els of β-catenin and of plakoglobin were not analyzed in this case.
More importantly, β-catenin-/- ESCs show defects during in vitro differen-tiation. Embryoid bodies generated by β-catenin-/- ESCs have adhesion defects during differentiation and defects in endoderm formation, whereas ectoderm induction can occur normally, which indicates that the cell-adhesion function of β-catenin is required at least for endo-derm formation. Similar results were also obtained in the case of embryoid body for-mation by E-cadherin-/- ESCs.41
Whether or not the cytoplasmic form of β-catenin and its binding to cadher-ins contributes to somatic cell repro-gramming has not been established yet. However, recently it was demonstrated that in the absence of E-cadherin expres-sion, reprogramming is impaired.42,43 During the conversion of mouse fibro-blasts into iPSCs by Oct4/Klf4/Sox2/c-Myc infection, reprogramming is accompanied by morphological changes, in which the cells move from a single layer of adherent cells to multilayered epithelial cells.43,44 This process, which is known as mesenchymal-to-epithelial transition, is characterized by upregulation of epithe-lial markers, such as E-cadherin, along with downregulation of mesenchymal markers, such as Snail. This mesenchy-mal-to-epithelial transition was shown to be required for, rather than a consequence of, iPSC formation.43,44 In addition, the efficiency of iPSC generation from MEFs infected with Oct4/Klf4/Sox2/c-Myc is substantially reduced when E-cadherin is deleted,42,43 which suggests that cytoplas-mic cues also participate in the process of reprogramming.
Nuclear regulation of self-renewal and reprogramming by Wnt/β-catenin. A combination of cell culture and gene abla-tion studies in ESCs has led to the identifi-cation of the intracellular determinants of
In all studies to date, the down-stream effects of β-catenin activity have remained poorly investigated. Once stabi-lized, β-catenin localizes both in the cyto-plasm and in the nucleus, with different functions in each of these compartments. Therefore, here we will review β-catenin’s role in self-renewal and reprogramming as related to its cytoplasmic and nuclear effects.
Cytoplasmic regulation of self-renewal and reprogramming by Wnt/β-catenin. One of the functions of β-catenin in the cytoplasm is to form part of the adherent junctions that link cadherins to α-catenin and the actin cytoskeleton, thereby partic-ipating in cell-cell adhesion complexes.37 In the course of pre-implantation develop-ment, β-catenin/E-cadherin contacts are crucial for correct compaction and blasto-cyst formation38 as well as in the deriva-tion of embryoid bodies from pluripotent ESCs.39
The treatment of undifferentiated ESCs with BIO or CHIR99021 leads these cells to grow in more compact clusters, with an increase in intercellular adhesion,10 an effect that is also observed in GSK3-/- ESCs.20 This effect might be due to the modulation of cell adhesions by the β-catenin/E-cadherin association, and indeed, β-catenin co-localizes with E-cadherin in ESCs, and GSK3 inhibition results in an increase in β-catenin binding to E-cadherin.
In agreement with this, ESCs lack-ing β-catenin show minor cell-adhesion defects, with regions lacking cell-cell con-tacts.23 That deletion of β-catenin does not lead to severe defects in cell adhesion of β-catenin-/- ESCs is mainly due to the compensatory effects of plakoglobin, a membrane protein that can bind to cad-herins, and which, thereby, participates in adherens junctions.23,40 Indeed, when plakoglobin expression is also low in β-catenin-/- ESCs, the cells show a flat-tened and dispersed morphology, with a reduction in the levels of the cell-adhesion molecules PECAM1 and E-cadherin. However, self-renewal markers are not altered here, and cell proliferation is only slightly decreased,23 which suggests that cell adhesion is not essential for the main-tenance of self-renewal. In agreement with this, E-cadherin-/- ESCs (which also have

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Cell Cycle 43
form.22 Analogous results using similar but independently generated ESC clones were obtained following the reprogram-ming phenotype (see below).55
Moreover, Yi et al.16 engineered Tcf3ΔN/ ΔN ESCs cells, where endogenous Tcf3 was replaced by the Tcf3ΔN iso-form. The engineered ESCs were growth under +LIF conditions and formed alka-line phosphatase positive (AP+) colonies similarly to wtESCs. In contrast, after Wnt3 stimulation, but in absence of LIF, wtESCs could form AP+ colonies, while Tcf3ΔN/ΔN ESCs showed a reduced, even if not impaired, ability to form AP+ colonies.
Altogether, these results show that Wnt stimulation of self-renewal triggers a Tcf3/β-catenin-dependent mechanism; however, they also suggest that additional Tcf3-independent mechanisms can con-tribute to ESC self-renewal. Alternative Tcf factors (see below) or alternative β-catenin binding factors might also be involved in this process.
Indeed, not all of the nuclear functions of β-catenin rely on its interaction with Tcf, as β-catenin can bind several other nuclear proteins to elicit its biological effects. β-catenin effects in GSK3-/- cells have been shown to be Tcf-independent.20 GSK3-/- ESCs can be maintained indefi-nitely under serum-free conditions, and they show important defects in differen-tiation.19,20 The overexpression of Tcf1ΔN and Tcf4ΔN in GSK3-/- ESCs cannot revert the differentiation defects of the GSK3-/- ESCs, which suggests that there might be a Tcf-independent mechanism.20 It is important to note that these authors20 showed that β-catenin can interact with Oct4, and that this interaction increases the transcriptional activity of an Oct4 reporter construct.
Deletion of Tcf3 can induce high effi-cient reprogramming of somatic nuclei.55 Upon fusion with neural progenitor cells (NPCs), Tcf3-/- ESCs produce a high number of reprogrammed hybrids as compared with the fusion of NPCs with wtESCs. Also, when the levels of Tcf3 in ESCs was reduced by RNA interference, after fusion, these cells were able to repro-gram MEFs.63
The ablation of Tcf3 in ESCs leads to an important change in the epigenome: the cells show an increase in acetylation of
a medium containing serum but lacking LIF stimulation.62 This indicates again that Tcf3 functions by limiting the levels of pluripotent factors. However, in serum-free medium, to maintain Tcf3-/- ESCs under self-renewal conditions, inhibition of at least the ERK pathway is needed.16
Given that stimulation of the stabiliza-tion of β-catenin and ablation of Tcf3 in ESCs give similar results for self-renewal (deletion of Tcf3 replaces the need to inhibit GSK3), and that both molecules are part of the same pathway, it appears natural to speculate that Tcf3 acts as a transcriptional repressor to inhibit self-renewal, and that this effect is directly suppressed by Wnt/β-catenin.
In this respect, using microarray analy-sis, it was shown recently that the expres-sion profile of wtESCs pre-treated with Wnt3a and of Tcf3-/- ESCs are highly cor-related, even if there is also a specific set of differentially expressed genes within each of these cell types.16 Given that Tcf3-/- cells showed increased expression of Nanog, it might be that the effects on self-renewal due to ablation of Tcf3 are, indeed, due to increased Nanog expression. However, by reducing the level of Nanog in Tcf3-/- ESCs or increasing Nanog levels in wtESCs and analyzing the expression profiles of these cells, Yi et al.16 concluded that the self-renewal ability of Tcf3-/- ESCs is not due to Nanog expression levels. On the other hand, in this study, how much Nanog expression was increased in wtESCs was not analyzed, and it might well be that the effects are different if Nanog is upregu-lated to high levels.
Recently, it has been shown that acti-vation of the Wnt signaling pathway induces the direct interaction of β-catenin with Tcf3.22 Wild-type Tcf3 (wtTcf3) and Tcf3 lacking the β-catenin binding domain (Tcf3ΔN) were overexpressed in Tcf3-/- ESCs. In the ESC clones obtained, suppression of TOPFlash activity (which contains binding sites for the T-cell factors; Tcf)10,16,25 and reduction in the expression of some endogenous β-catenin/Tcf target genes, such as Cdx1, were shown. More importantly, activation of the Wnt path-way using CHIR99021 re-established self-renewal of the ES clones that overex-pressed wtTcf3 and not completely that of the clones that overexpressed the Tcf3ΔN
In ESCs, the most highly expressed of the Tcf family members is Tcf3 (Tcf7l1), followed by Tcf1 (Tcf7). Tcf4 (Tcf7l2) and Lef-1 are scarcely expressed.55,56 Mainly for this reason, the most studied Tcf protein in ESCs is Tcf3, which has two different isoforms in ESCs.57,58 In addition, and importantly, Tcf3 is an inte-gral component of the Oct4/Sox2/Nanog self-renewal circuit.5,59 Using chromatin immunoprecipitation sequencing (ChIP-seq), it has been shown that 14,230 sites in the genome are co-occupied by Oct4, Sox2, Nanog and Tcf3. Of these, one quarter of the binding sites are within 8 kb of the transcription start site of 3,289 annotated genes. Considering that some promoters have several binding sites for the same factor, it has been shown that Tcf3 binds to 1,369 known promoters, 942 of which are bound by all four of these factors.59 Similar data have been obtained using ChIP-on-chip.5,60 Furthermore, recently gene-trap screening identified Tcf3 as a pluripotency-controlling factor.61 All of these data show that Tcf3 is a key regulating factor in the self-renewal core in ESCs.
Although Tcf3 is associated with several promoters, deletion of Tcf3 in ESCs does not especially affect their self-renewal. Nanog expression is increased 2.5-fold in Tcf3-/- ESCs with respect to wtESCs. In contrast, Oct4 and Sox2 are similarly expressed in wtESCs and Tcf3-/- ESCs.56 These expression levels are in agreement with those obtained when Wnt signaling is activated in wtESCs. However, the knock-down of Tcf3 by shRNAs increases the expression of Oct4, Sox2 and Nanog.5,60 The differences between these studies can be explained considering that in one case, steady-state Tcf3-/- ESCs were used, while in the other case, the silencing of Tcf3 was transient, and dynamic effects (temporal knockdown) might have occurred.
It is interesting to note that Tcf3 is expressed at higher levels in ESCs than in their differentiated progeny, which might suggests that Tcf3 is required to limit the levels of pluripotent proteins like Nanog to allow correct differentiation. Indeed, in agreement with this hypothesis, Tcf3-
/- ESCs show a delay in differentiation, and more importantly, they can be main-tained under self-renewal conditions in
© 2012 Landes Bioscience.
Do not distribute.
44 Cell Cycle volume 11 issue 1
Interestingly, NPCs infected with Oct4/Klf4/shTcf3 also show an increase in AcH3 and a decrease in H3K9me3, fur-ther demonstrating that these epigenetic changes that arise from a reduction in Tcf3 expression might enhance reprogramming of somatic nuclei. More importantly, these events of chromatin derepression occurred before re-expression of the endogenous stem cell genes.55
We have shown previously that Tcf3-/- ESCs can reprogram NPCs after fusion with great efficiency.55 We then generated Tcf3-/- ESC clones that express wtTcf3 or
was confirmed by the efficient binding of Oct4 to different promoters in Tcf3-/- cells compared with wtESCs.16,55 Interestingly, treatment of wtESCs with BIO can pro-duce the same changes in the chromatin state.55 All of these changes might explain why Tcf3-/- cells are so efficient in the pro-motion of somatic cell reprogramming after fusion.
In addition, Tcf3 is also an important factor for the generation of iPSCs. NPCs transduced with Oct4/Klf4 and shRNA against Tcf3 lead to the rapid genera-tion of a high number of iPSC colonies.
H3 (AcH3) and a reduction in trimethyl-ation of lysine 9 in histone 3 (H3K9me3). Both of these epigenome changes promote an increase in the open and active tran-scriptional state of the chromatin.64 The importance of these epigenetic changes resides in the high number of genes that are co-regulated by Tcf3 and the Oct4/Nanog/Sox2 pluripotent core. If dele-tion of Tcf3 induces an open chromatin state, it might also increase the ability of other factors to bind the chromatin and, thus, increase the self-renewal potential of these Tcf3-/- cells. Indeed, this theory
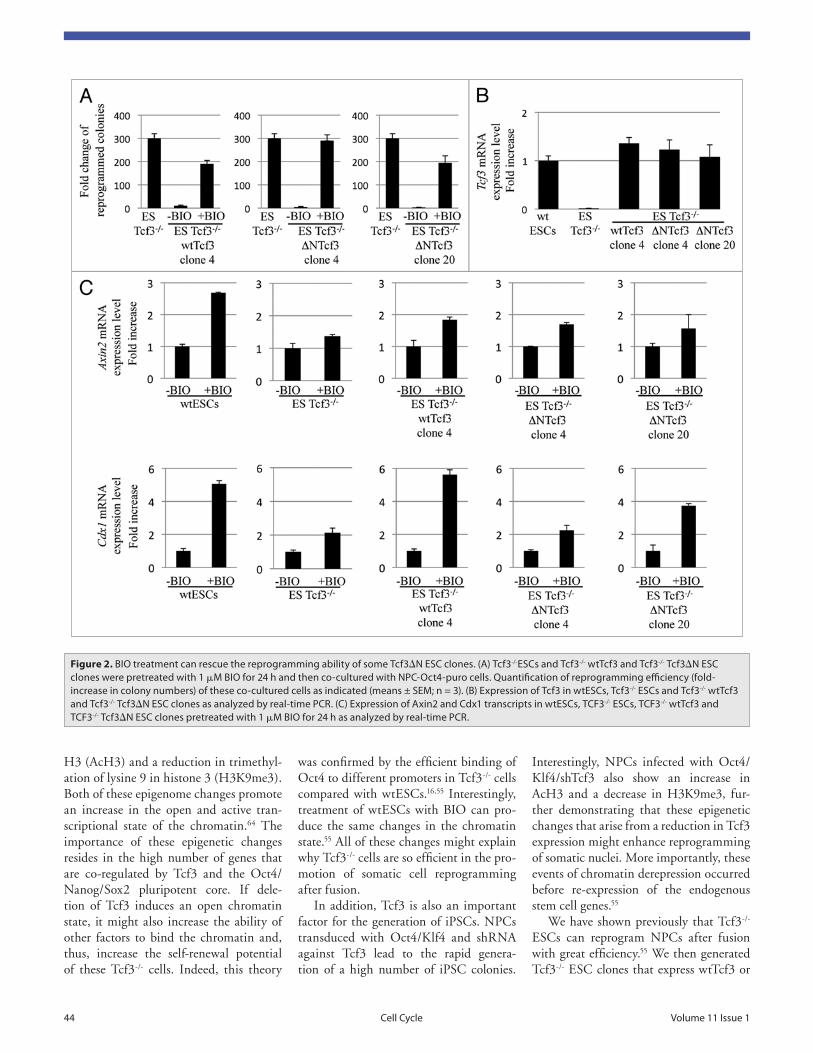
Figure 2. BiO treatment can rescue the reprogramming ability of some tcf3ΔN eSC clones. (a) tcf3-/-eSCs and tcf3-/- wttcf3 and tcf3-/- tcf3ΔN eSC clones were pretreated with 1 μM BiO for 24 h and then co-cultured with NPC-Oct4-puro cells. Quantification of reprogramming efficiency (fold-increase in colony numbers) of these co-cultured cells as indicated (means ± SeM; n = 3). (B) expression of tcf3 in wteSCs, tcf3-/- eSCs and tcf3-/- wttcf3 and tcf3-/- tcf3ΔN eSC clones as analyzed by real-time PCr. (C) expression of axin2 and Cdx1 transcripts in wteSCs, tCF3-/- eSCs, tCF3-/- wttcf3 and tCF3-/- tcf3ΔN eSC clones pretreated with 1 μM BiO for 24 h as analyzed by real-time PCr.

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Cell Cycle 45
more recently, it has been shown that Tcf3 acts mainly as a repressor, and that it can be displaced from promoters after the binding of β-catenin in a complex with different Tcf factors, which can, in turn, activate target gene transcription.65,66 In Xenopus embryos, the decreased binding of Tcf3 to the Vent2 promoter that arises through Wnt stimulation is accompanied by enhanced association of Tcf1 with the same binding site, which leads to upregu-lation of Vent2 expression.67
Tcf1 is the second most expressed Tcf factor in ESCs,56 and it has roles in both ESC self-renewal and reprogramming. In ESCs with Tcf1 silenced by shRNAs, there are no apparent cellular defects. However, in absence of LIF, the silencing of Tcf1 in wtESCs, in Tcf3-/- ESCs and in Tcf3ΔN/ ΔN ESCs cultured in Wnt3a-containing medium induced downregulation of plu-ripotent genes, such as Nanog and Nr5a2 and a decreased ability to form pluripo-tent colonies.16 Furthermore, silencing of Tcf1 expression through shRNA-medi-ated knockdown in Tcf3-/- ESCs greatly reduced their ability to reprogram somatic cells by cell fusion.55
In conclusion, the activity of Tcf1 is also a critical step in the modulation of the Wnt signaling pathway in its role to control somatic cell reprogramming and ESC self-renewal.
levels of Tcf3ΔN comparable to the Tcf3 levels in wtESCs) responded to BIO treat-ment and activated reprogramming of NPCs after fusion (Fig. 2A and B). Even if surprising, this result was not totally unexpected, given that overexpression of Tcf3ΔN in wtESCs or Tcf3-/- ESCs does not completely inhibit the ability of these cells to maintain self-renewal after Wnt stimulation.16,22 Finally, we analyzed the expression of β-catenin target genes in the Tcf3-/- wtTcf3 and Tcf3-/- Tcf3ΔN clones after BIO treatment. Here, Axin2 and Cdx1 were activated in the above-mentioned ΔN4 and ΔN20 clones, even if at lower levels compared with the Tcf3-/- wtTcf3 (Fig. 2C). This confirmed that in the Tcf3-/- Tcf3ΔN clones 4 and 20, the Wnt signaling pathway was activated by BIO treatment independently of Tcf3.
All in all, these observations demon-strate that although ESCs treated with Wnt3a or deleted for Tcf3 can main-tain self-renewal, β-catenin can activate self-renewal or reprogramming not only through Tcf3 de-repression. Indeed, some of the Tcf3-/- Tcf3ΔN clones and Tcf3ΔN/ ΔN cells still show a low response to Wnt stimulation.16,22
Canonical Wnt signaling was proposed to support pluripotency through the con-version of the activity of Tcf3 complexes from repressors to activators.5,65 However,
Tcf3ΔN; these Tcf3-/- wtTcf3 and Tcf3-/- Tcf3ΔN clones did not reprogram NPCs after fusion. This effect was reversed by pretreatment of the Tcf3-/- wtTcf3 clones with BIO for 24 h but not when the Tcf3-/- Tcf3ΔN clones were pretreated with BIO before fusion.55
Given the hypothesis that Tcf3-independent mechanisms contribute to ESC self-renewal (see above), the inhibi-tory effects of Tcf3 and its exclusive function in the repression of reprogram-ming were further studied by generating and analyzing a large number of clones overexpressing wtTcf3 and Tcf3ΔN in Tcf3-/- ESCs. All of the newly generated clones showed a block of the reprogram-ming ability of the original Tcf3-/- cells. As expected, for all of the Tcf3-/- wtTcf3 clones, pretreatment with BIO for 24 h reversed their blocked reprogramming ability after fusion as also previously shown (reviewed in ref. 55; Fig. 2A, wtTcf3 clone 4). However, rescue of the reprogramming ability was also observed in two Tcf3-/- Tcf3ΔN clones (clones ΔN4 and ΔN20) pretreated with BIO before fusion (Fig. 2A). Interestingly, the clones expressing high levels of Tcf3ΔN were the ones that did not respond to BIO treat-ment and showed a lack of reprogram-ming ability after fusion,55 whereas the clones expressing low levels of Tcf3ΔN (or
Figure 3. wnt/β-catenin and tcf regulation of self-renewal and reprogramming. in the absence of wnt activation, tcf3 acts as a repressor of transcrip-tion, as it can bind to several promoters along with the pluripotent gene network (Oct4, Nanog, Sox2). when β-catenin is stabilized, tcf3 is phos-phorylated and dissociates from DNa. this leads to massive epigenome modifications, which determine the establishment of an “open chromatin” conformation. in turn, the binding of tcf1 with β-catenin and the increase in Oct4 binding are favored. alternatively, a tcf-independent mechanism postulates that β-catenin can associate with Oct4, and that it is this complex that can activate the target genes.

© 2012 Landes Bioscience.
Do not distribute.
46 Cell Cycle volume 11 issue 1
16. Yi F, Pereira L, Hoffman JA, Shy BR, Yuen CM, Liu DR, et al. Opposing effects of Tcf3 and Tcf1 control Wnt stimulation of embryonic stem cell self-renewal. Nat Cell Biol 2011; 13:762-70; PMID:21685894; http://dx.doi.org/10.1038/ncb2283.
17. Aubert J, Dunstan H, Chambers I, Smith A. Functional gene screening in embryonic stem cells implicates Wnt antagonism in neural differentiation. Nat Biotechnol 2002; 20:1240-5; PMID:12447396; http://dx.doi.org/10.1038/nbt763.
18. ten Berge D, Kurek D, Blauwkamp T, Koole W, Maas A, Eroglu E, et al. Embryonic stem cells require Wnt proteins to prevent differentiation to epiblast stem cells. Nat Cell Biol 2011; 13:1070-5; PMID:21841791; http://dx.doi.org/10.1038/ncb2314.
19. Doble BW, Patel S, Wood GA, Kockeritz LK, Woodgett JR. Functional redundancy of GSK-3alpha and GSK-3beta in Wnt/beta-catenin signaling shown by using an allelic series of embryonic stem cell lines. Dev Cell 2007; 12:957-71; PMID:17543867; http://dx.doi.org/10.1016/j.devcel.2007.04.001.
20. Kelly KF, Ng DY, Jayakumaran G, Wood GA, Koide H, Doble BW. beta-catenin enhances Oct-4 activity and reinforces pluripotency through a TCF-independent mechanism. Cell Stem Cell 2011; 8:214-27; PMID:21295277; http://dx.doi.org/10.1016/j.stem.2010.12.010.
21. Anton R, Kestler HA, Kuhl M. beta-Catenin signal-ing contributes to stemness and regulates early differ-entiation in murine embryonic stem cells. FEBS Lett 2007; 581:5247-54; PMID:17950287; http://dx.doi.org/10.1016/j.febslet.2007.10.012.
22. Wray J, Kalkan T, Gomez-Lopez S, Eckardt D, Cook A, Kemler R, et al. Inhibition of glycogen synthase kinase-3 alleviates Tcf3 repression of the pluripotency network and increases embryonic stem cell resistance to differentiation. Nat Cell Biol 2011; 13:838-45; PMID:21685889; http://dx.doi.org/10.1038/ncb2267.
23. Lyashenko N, Winter M, Migliorini D, Biechele T, Moon RT, Hartmann C. Differential requirement for the dual functions of beta-catenin in embryonic stem cell self-renewal and germ layer formation. Nat Cell Biol 2011; 13:753-61; PMID:21685890; http://dx.doi.org/10.1038/ncb2260.
24. Lluis F, Cosma MP. Somatic cell reprogramming control: signaling pathway modulation versus tran-scription factor activities. Cell Cycle 2009; 8:1138-44; PMID:19305152; http://dx.doi.org/10.4161/cc.8.8.8206.
25. Lluis F, Pedone E, Pepe S, Cosma MP. Periodic activation of Wnt/beta-catenin signaling enhances somatic cell reprogramming mediated by cell fusion. Cell Stem Cell 2008; 3:493-507; PMID:18983965; http://dx.doi.org/10.1016/j.stem.2008.08.017.
26. Lluis F, Pedone E, Pepe S, Cosma MP. The Wnt/beta-catenin signaling pathway tips the balance between apoptosis and reprograming of cell fusion hybrids. Stem Cells 2010; 28:1940-9; PMID:20827748; http://dx.doi.org/10.1002/stem.515.
27. Marson A, Foreman R, Chevalier B, Bilodeau S, Kahn M, Young RA, et al. Wnt signaling promotes reprogramming of somatic cells to pluripotency. Cell Stem Cell 2008; 3:132-5; PMID:18682236; http://dx.doi.org/10.1016/j.stem.2008.06.019.
28. Li W, Zhou H, Abujarour R, Zhu S, Young Joo J, Lin T, et al. Generation of human-induced pluripotent stem cells in the absence of exogenous Sox2. Stem Cells 2009; 27:2992-3000; PMID:19839055.
29. Zhu S, Ma T, Li J, Ding S. Recent advances in chemically induced reprogramming. Cell Cycle 2011; 10:871-2; PMID:21368574; http://dx.doi.org/10.4161/cc.10.6.15035.
30. Banito A, Rashid ST, Acosta JC, Li S, Pereira CF, Geti I, et al. Senescence impairs successful repro-gramming to pluripotent stem cells. Genes Dev 2009; 23:2134-9; PMID:19696146; http://dx.doi.org/10.1101/gad.1811609.
from HFSP Grant RGP0011/2010 (to M.P.C.). F.L. is funded by CP10/00445 Project “Miguel Servet” of the Instituto de Salud Carlos III.
References1. Logan CY, Nusse R. The Wnt signaling pathway in
development and disease. Annu Rev Cell Dev Biol 2004; 20:781-810; PMID:15473860; http://dx.doi.org/10.1146/annurev.cellbio.20.010403.113126.
2. Miller JR. The Wnts. Genome Biol 2002; 3:3001; PMID:11806834.
3. MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms and diseases. Dev Cell 2009; 17:9-26; PMID:19619488; http://dx.doi.org/10.1016/j.devcel.2009.06.016.
4. Wu D, Pan W. GSK3: a multifaceted kinase in Wnt signaling. Trends Biochem Sci 2010; 35:161-8; PMID:19884009; http://dx.doi.org/10.1016/j.tibs.2009.10.002.
5. Cole MF, Johnstone SE, Newman JJ, Kagey MH, Young RA. Tcf3 is an integral component of the core regulatory circuitry of embryonic stem cells. Genes Dev 2008; 22:746-55; PMID:18347094; http://dx.doi.org/10.1101/gad.1642408.
6. Willert K, Jones KA. Wnt signaling: is the party in the nucleus? Genes Dev 2006; 20:1394-404; PMID:16751178; http://dx.doi.org/10.1101/gad.1424006.
7. Hoppler S, Kavanagh CL. Wnt signalling: vari-ety at the core. J Cell Sci 2007; 120:385-93; PMID:17251379; http://dx.doi.org/10.1242/jcs.03363.
8. Sato N, Meijer L, Skaltsounis L, Greengard P, Brivanlou AH. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat Med 2004; 10:55-63; PMID:14702635; http://dx.doi.org/10.1038/nm979.
9. Silva J, Barrandon O, Nichols J, Kawaguchi J, Theunissen TW, Smith A. Promotion of reprogram-ming to ground state pluripotency by signal inhibi-tion. PLoS Biol 2008; 6:253; PMID:18942890; http://dx.doi.org/10.1371/journal.pbio.0060253.
10. Sineva GS, Pospelov VA. Inhibition of GSK3beta enhances both adhesive and signalling activities of beta-catenin in mouse embryonic stem cells. Biol Cell 2010; 102:549-60; PMID:20626347; http://dx.doi.org/10.1042/BC20100016.
11. Meijer L, Skaltsounis AL, Magiatis P, Polychronopoulos P, Knockaert M, Leost M, et al. GSK-3-selective inhibitors derived from Tyrian purple indirubins. Chem Biol 2003; 10:1255-66; PMID:14700633; http://dx.doi.org/10.1016/j.chem-biol.2003.11.010.
12. Ying QL, Wray J, Nichols J, Batlle-Morera L, Doble B, Woodgett J, et al. The ground state of embry-onic stem cell self-renewal. Nature 2008; 453:519-23; PMID:18497825; http://dx.doi.org/10.1038/nature06968.
13. Niwa H, Burdon T, Chambers I, Smith A. Self-renewal of pluripotent embryonic stem cells is medi-ated via activation of STAT3. Genes Dev 1998; 12:2048-60; PMID:9649508; http://dx.doi.org/10.1101/gad.12.13.2048.
14. Smith AG. Embryo-derived stem cells: of mice and men. Annu Rev Cell Dev Biol 2001; 17:435-62; PMID:11687496; http://dx.doi.org/10.1146/annurev.cellbio.17.1.435.
15. Sato N, Sanjuan IM, Heke M, Uchida M, Naef F, Brivanlou AH. Molecular signature of human embry-onic stem cells and its comparison with the mouse. Dev Biol 2003; 260:404-13; PMID:12921741; http://dx.doi.org/10.1016/S0012-1606(03)00256-2.
Conclusions
Tremendous efforts have been made to identify the components involved in and the molecular mechanisms driving ESC self-renewal and somatic cell reprogram-ming. The function of the Wnt/β-catenin/Tcf pathway in these processes remained puzzling for several years. However, it is now clear that the activation of the Wnt canonical pathway (or inhibition of GSK3) can mimic the deletion of Tcf3 in ESCs, although the resulting effects on self-renewal and reprogramming are not the same. As an important example, stabi-lization of β-catenin by Wnt3a treatment or GSK3 inhibition maintains ESC self-renewal and reprogramming, as does dele-tion of Tcf3 in ESCs; however, Wnt3a and BIO treatments do not increase the expres-sion of Nanog, while Nanog is upregu-lated in Tcf3-/- ESCs. Thus, the effects of β-catenin stabilization on target genes do not necessarily pass through Tcf3.
One important player in this scenario is Tcf1, which has an opposite effect to that of Tcf3, acting primarily as an acti-vator. As a result of reduction of Tcf1 expression in ESCs, there is a reduction in the self-renewal capacity as well as in the reprogramming activity, making Tcf1 an important candidate to counterbalance the repressing effects of Tcf3. However, Oct4/β-catenin interactions have also been reported to increase transcription of target genes independently of Tcf activi-ties20,68 (Fig. 3).
Finally, and importantly, in ESCs, the levels of β-catenin are also crucial for the modulation of self-renewal and repro-gramming after fusion. When β-catenin activity is very high (due to high doses of BIO or in GSK3-/- ESCs), the result-ing effects are Tcf-independent. Indeed, overexpression of Tcf1ΔN or Tcf4ΔN in GSK3-/- ESCs appears not to produce any effects compared with GSK3-/- ESCs.20 Furthermore, treatment of Tcf3-/- ESCs with high concentrations of BIO can block the reprogramming activity of these cells.55
Acknowledgments
This project was financed with funds from the European Research Council Grant, StERC 242630-RERE (to M.P.C.) and

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Cell Cycle 47
56. Pereira L, Yi F, Merrill BJ. Repression of Nanog gene transcription by Tcf3 limits embryonic stem cell self-renewal. Mol Cell Biol 2006; 26:7479-91; PMID:16894029; http://dx.doi.org/10.1128/MCB.00368-06.
57. Salomonis N, Conklin BR. Stem cell pluripotency: alternative modes of transcription regulation. Cell Cycle 2010; 9:3133-4; PMID:20814242; http://dx.doi.org/10.4161/cc.9.16.12888.
58. Salomonis N, Schlieve CR, Pereira L, Wahlquist C, Colas A, Zambon AC, et al. Alternative splic-ing regulates mouse embryonic stem cell pluripo-tency and differentiation. Proc Natl Acad Sci USA 2010; 107:10514-9; PMID:20498046; http://dx.doi.org/10.1073/pnas.0912260107.
59. Marson A, Levine SS, Cole MF, Frampton GM, Brambrink T, Johnstone S, et al. Connecting microR-NA genes to the core transcriptional regulatory cir-cuitry of embryonic stem cells. Cell 2008; 134:521-33; PMID:18692474; http://dx.doi.org/10.1016/j.cell.2008.07.020.
60. Tam WL, Lim CY, Han J, Zhang J, Ang YS, Ng HH, et al. T-cell factor 3 regulates embryonic stem cell pluripotency and self-renewal by the transcriptional control of multiple lineage pathways. Stem Cells 2008; 26:2019-31; PMID:18467660; http://dx.doi.org/10.1634/stemcells.2007-1115.
61. Guo G, Huang Y, Humphreys P, Wang X, Smith A. A PiggyBac-based recessive screening method to identify pluripotency regulators. PLoS ONE 2011; 6:18189; PMID:21533166; http://dx.doi.org/10.1371/journal.pone.0018189.
62. Yi F, Pereira L, Merrill BJ. Tcf3 functions as a steady-state limiter of transcriptional programs of mouse embryonic stem cell self-renewal. Stem Cells 2008; 26:1951-60; PMID:18483421; http://dx.doi.org/10.1634/stemcells.2008-0229.
63. Han J, Yuan P, Yang H, Zhang J, Soh BS, Li P, et al. Tbx3 improves the germ-line competency of induced pluripotent stem cells. Nature 2010; 463:1096-100; PMID:20139965; http://dx.doi.org/10.1038/nature08735.
64. Meshorer E, Misteli T. Chromatin in pluripotent embryonic stem cells and differentiation. Nat Rev Mol Cell Biol 2006; 7:540-6; PMID:16723974; http://dx.doi.org/10.1038/nrm1938.
65. Sokol SY. Maintaining embryonic stem cell pluri-potency with Wnt signaling. Development 2011; 138:4341-50; PMID:21903672; http://dx.doi.org/10.1242/dev.066209.
66. Hikasa H, Ezan J, Itoh K, Li X, Klymkowsky MW, Sokol SY. Regulation of TCF3 by Wnt-dependent phosphorylation during vertebrate axis specification. Dev Cell 2010; 19:521-32; PMID:20951344; http://dx.doi.org/10.1016/j.devcel.2010.09.005.
67. Hikasa H, Sokol SY. Phosphorylation of TCF Proteins by Homeodomain-interacting Protein Kinase 2. J Biol Chem 2011; 286:12093-100; PMID:21285352; http://dx.doi.org/10.1074/jbc.M110.185280.
68. Takao Y, Yokota T, Koide H. Beta-catenin upregu-lates Nanog expression through interaction with Oct-3/4 in embryonic stem cells. Biochem Biophys Res Commun 2007; 353:699-705; PMID:17196549; http://dx.doi.org/10.1016/j.bbrc.2006.12.072.
44. Samavarchi-Tehrani P, Golipour A, David L, Sung HK, Beyer TA, Datti A, et al. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell 2010; 7:64-77; PMID:20621051; http://dx.doi.org/10.1016/j.stem.2010.04.015.
45. Niwa H, Miyazaki J, Smith AG. Quantitative expres-sion of Oct-3/4 defines differentiation, dediffer-entiation or self-renewal of ES cells. Nat Genet 2000; 24:372-6; PMID:10742100; http://dx.doi.org/10.1038/74199.
46. Niwa H. How is pluripotency determined and maintained? Development 2007; 134:635-46; PMID:17215298; http://dx.doi.org/10.1242/dev.02787.
47. Silva J, Nichols J, Theunissen TW, Guo G, van Oosten AL, Barrandon O, et al. Nanog is the gateway to the pluripotent ground state. Cell 2009; 138:722-37; PMID:19703398; http://dx.doi.org/10.1016/j.cell.2009.07.039.
48. Boyer LA, Mathur D, Jaenisch R. Molecular control of pluripotency. Curr Opin Genet Dev 2006; 16:455-62; PMID:16920351; http://dx.doi.org/10.1016/j.gde.2006.08.009.
49. Loh YH, Wu Q, Chew JL, Vega VB, Zhang W, Chen X, et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat Genet 2006; 38:431-40; PMID:16518401; http://dx.doi.org/10.1038/ng1760.
50. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007; 131:861-72; PMID:18035408; http://dx.doi.org/10.1016/j.cell.2007.11.019.
51. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006; 126:663-76; PMID:16904174; http://dx.doi.org/10.1016/j.cell.2006.07.024.
52. Yamanaka S, Blau HM. Nuclear reprogramming to a pluripotent state by three approaches. Nature 2010; 465:704-12; PMID:20535199; http://dx.doi.org/10.1038/nature09229.
53. Brannon M, Brown JD, Bates R, Kimelman D, Moon RT. XCtBP is a XTcf-3 co-repressor with roles throughout Xenopus development. Development 1999; 126:3159-70; PMID:10375506.
54. Molenaar M, van de Wetering M, Oosterwegel M, Peterson-Maduro J, Godsave S, Korinek V, et al. XTcf-3 transcription factor mediates beta-catenin-induced axis formation in Xenopus embryos. Cell 1996; 86:391-9; PMID:8756721; http://dx.doi.org/10.1016/S0092-8674(00)80112-9.
55. Lluis F, Ombrato L, Pedone E, Pepe S, Merrill BJ, Cosma MP. T-cell factor 3 (Tcf3) deletion increases somatic cell reprogramming by inducing epigenome modifications. Proc Natl Acad Sci USA 2011; 108:11912-7; PMID:21730189; http://dx.doi.org/10.1073/pnas.1017402108.
31. Hong H, Takahashi K, Ichisaka T, Aoi T, Kanagawa O, Nakagawa M, et al. Suppression of induced plu-ripotent stem cell generation by the p53-p21 pathway. Nature 2009; 460:1132-5; PMID:19668191; http://dx.doi.org/10.1038/nature08235.
32. Kawamura T, Suzuki J, Wang YV, Menendez S, Morera LB, Raya A, et al. Linking the p53 tumour suppressor pathway to somatic cell reprogramming. Nature 2009; 460:1140-4; PMID:19668186; http://dx.doi.org/10.1038/nature08311.
33. Li H, Collado M, Villasante A, Strati K, Ortega S, Canamero M, et al. The Ink4/Arf locus is a barrier for iPS cell reprogramming. Nature 2009; 460:1136-9; PMID:19668188; http://dx.doi.org/10.1038/nature08290.
34. Marión RM, Strati K, Li H, Murga M, Blanco R, Ortega S, et al. A p53-mediated DNA dam-age response limits reprogramming to ensure iPS cell genomic integrity. Nature 2009; 460:1149-53; PMID:19668189; http://dx.doi.org/10.1038/nature08287.
35. Utikal J, Polo JM, Stadtfeld M, Maherali N, Kulalert W, Walsh RM, et al. Immortalization eliminates a roadblock during cellular reprogramming into iPS cells. Nature 2009; 460:1145-8; PMID:19668190; http://dx.doi.org/10.1038/nature08285.
36. Menendez S, Camus S, Izpisua Belmonte JC. p53: guardian of reprogramming. Cell Cycle 2010; 9:3887-91; PMID:20948296; http://dx.doi.org/10.4161/cc.9.19.13301.
37. Kemler R. From cadherins to catenins: cytoplasmic protein interactions and regulation of cell adhe-sion. Trends Genet 1993; 9:317-21; PMID:8236461; http://dx.doi.org/10.1016/0168-9525(93)90250-L.
38. Larue L, Ohsugi M, Hirchenhain J, Kemler R. E-cadherin null mutant embryos fail to form a trophectoderm epithelium. Proc Natl Acad Sci USA 1994; 91:8263-7; PMID:8058792; http://dx.doi.org/10.1073/pnas.91.17.8263.
39. Dang SM, Gerecht-Nir S, Chen J, Itskovitz-Eldor J, Zandstra PW. Controlled, scalable embryonic stem cell differentiation culture. Stem Cells 2004; 22:275-82; PMID:15153605; http://dx.doi.org/10.1634/stemcells.22-3-275.
40. Fukunaga Y, Liu H, Shimizu M, Komiya S, Kawasuji M, Nagafuchi A. Defining the roles of beta-catenin and plakoglobin in cell-cell adhesion: isolation of beta-catenin/plakoglobin-deficient F9 cells. Cell Struct Funct 2005; 30:25-34; PMID:16357441; http://dx.doi.org/10.1247/csf.30.25.
41. Soncin F, Mohamet L, Eckardt D, Ritson S, Eastham AM, Bobola N, et al. Abrogation of E-cadherin-mediated cell-cell contact in mouse embryonic stem cells results in reversible LIF-independent self-renew-al. Stem Cells 2009; 27:2069-80; PMID:19544408; http://dx.doi.org/10.1002/stem.134.
42. Redmer T, Diecke S, Grigoryan T, Quiroga-Negreira A, Birchmeier W, Besser D. E-cadherin is crucial for embryonic stem cell pluripotency and can replace OCT4 during somatic cell reprogramming. EMBO Rep 2011; 12:720-6; PMID:21617704; http://dx.doi.org/10.1038/embor.2011.88.
43. Li R, Liang J, Ni S, Zhou T, Qing X, Li H, et al. A mesenchymal-to-epithelial transition initi-ates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell 2010; 7:51-63; PMID:20621050; http://dx.doi.org/10.1016/j.stem.2010.04.014.




















