
Ecto-protein kinases and phosphatases: an emerging field for translational medicine

Neurochemistry International 61 (2012) 681–688
Contents lists available at SciVerse ScienceDirect
Neurochemistry International
journal homepage: www.elsevier .com/locate /nci
Regulation of ecto-50-nucleotidase (CD73) in cultured cortical astrocytesby different inflammatory factors
Dusica Brisevac a, Ivana Bjelobaba b, Aleksandar Bajic a,c, Tim Clarner d, Mirjana Stojiljkovic b,Cordian Beyer d, Pavle Andjus c, Markus Kipp d, Nadezda Nedeljkovic a,⇑a Institute for Physiology and Biochemistry, Faculty of Biology, University of Belgrade, Serbiab Department of Neurobiology, Institute for Biological Research ‘‘Sinisa Stankovic’’, University of Belgrade, Serbiac Center for Laser Microscopy, Faculty of Biology, University of Belgrade, Serbiad Institute of Neuroanatomy, Faculty of Medicine, RWTH Aachen University, Germany
a r t i c l e i n f o a b s t r a c t
Article history:Received 24 December 2011Received in revised form 12 June 2012Accepted 21 June 2012Available online 29 June 2012
Keywords:Ecto-50-nucleotidase (CD73)AstrocytesATPAdenosineInflammation
0197-0186/$ - see front matter � 2012 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.neuint.2012.06.017
⇑ Corresponding author. Address: Institute for PFaculty of Biology, University of Belgrade, StudentskiTel./fax: +381 11 3032 356.
E-mail address: [email protected] (N. Nedeljkov
Ecto-50-nucleotidase (e-5NT) is a cell-surface located, rate-limiting enzyme in the extracellular metabo-lism of ATP, catalyzing the final step of the conversion of AMP to adenosine. Since this enzyme shifts thebalance from pro-inflammatory ATP to anti-inflammatory adenosine, it is considered to be an importantregulator of inflammation. Although up-regulation of e-5NT was repeatedly reported in several in vivomodels of brain injury, the regulation of its expression and function remains largely unknown. We havestudied effects of several pro-inflammatory factors, namely, bacterial endotoxin lipopolysaccharide (LPS),tumor necrosis factor-a (TNF-a), interferon-c (IFN-c), glutamate (Glu) and hydrogen peroxide (H2O2) one-5NT (i) activity, (ii) mRNA expression and (iii) membrane protein abundance in primary cultured cor-tical astrocytes. We are clearly able to demonstrate a stimulus-specific regulation of the e-5NT pathway.IFN-c, LPS, Glu and H2O2 decrease, while TNF-a increases e-5NT activity. The analysis of e-5NT geneexpression and e-5NT membrane protein levels revealed that tested factors regulate e-5NT at differentlevels and by employing different mechanisms. In summary, we provide evidence that e-5NT activityis tightly regulated in a stimulus-specific manner.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Neuroinflammation is a complex homeostatic response of thecentral nervous system (CNS), devised to protect its structuraland functional integrity against endogenous or exogenous noxiousagents. Microglia are the major resident immune cells in the brain,where they constantly survey the microenvironment and producefactors that influence surrounding cells, such as neurons or astro-cytes. Astrocytes are second key players in inflammatory responses(Aloisi et al., 2000; Farina et al., 2007), intimately involved in virtu-ally every form of neuropathology (Verkhratsky et al., 2009).Microglia sense the presence of pathogen-associated molecularpatterns–PAMPs (Janeway, 1992) or danger-associated molecularpatterns–DAMPs found on proteins or released from damaged cells(Matzinger, 2002). These entities are recognized by microglial pat-tern recognition receptors (PRRs), whose activation induces imme-diate microglial activation and secretion of pro-inflammatorycytokines and chemokines, such as TNF-a, IL-1b and IFN-c (Herx
ll rights reserved.
hysiology and Biochemistry,trg 3, 11001 Belgrade, Serbia.
ic).
and Young, 2001; Selmaj et al., 1990), that disturb astrocyte func-tioning (Zhang et al., 2009) and induce astrogliosis. Activation ofastrocytes is characterized, among others, by hypertrophia andhyperplasia of astrocytic processes, increased expression of GFAPand vimentin and functional changes that are still poorly under-stood. Under pathological conditions or after in vitro exposure todifferent neurotoxins, activated astrocytes contribute to neuroin-flammation by releasing pro-inflammatory cytokines and chemo-kines (Farina et al., 2007) and by synthesizing prostaglandines,thromboxanes and leukotrienes (Johann et al., 2008). On the otherhand, astrocytes contribute to protective and regenerative pro-cesses in the brain by releasing anti-inflammatory factors thatattenuate microglial response (Kim et al., 2010) and by releasinggrowth factors, that support neuronal survival and stability (Braunet al., 2009). Therefore, inflammation is not a deleterious processper se but if balanced, it regulates regenerative processes (Kippet al., 2011).
The role of purinergic signaling is a rapidly expanding area ofinterest in neuroinflammation research. An important aspect inthe induction of an inflammatory response is release of signalsfrom damaged tissues that activate microglia and astrocytes.Among many other endogenous factors, extracellular purine nucle-otides and nucleosides, such as ATP and adenosine are now

682 D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688
regarded as true inflammatory mediators (for review see Virgilioet al., 2009). In this regard, results from several groups stronglysuggest that extracellular ATP, derived from the injured tissue,orchestrates microglia and astrocyte activation (Bolego et al.,1997; Cavaliere et al., 2003; Le Feuvre et al., 2002). By acting attwo types of receptors, ionotropic (P2X) and metabotropic (P2Y),ATP modulates various cellular responses (Gudipaty et al., 2003;Witting et al., 2004) and induces the secretion of pro-inflammatorycytokines and chemokines by microglia (Ferrari et al., 2006; Kat-aoka et al., 2009; Suzuki et al., 2004) and astrocytes (Kucher andNeary, 2005; Liu et al., 2011).
Tissue damage and inflammation are also accompanied by in-creased level of extracellular adenosine (Frengueli et al., 2007). Incontrast to ATP, adenosine acts as a potent anti-inflammatoryagent (Hasko and Cronstein, 2004; Sitkovsky et al., 2004). In addi-tion, adenosine induces microglial taxis and reactivity (Gyonevaet al., 2009), controls neuronal viability (Dall’Igna et al., 2004), reg-ulates astrogliosis (Brambilla et al., 2003; Hindley et al., 1994) andastrocytic release of glutamate (Li et al., 2006), IL-6 (Fiebich et al.,1996; Schwaninger et al., 1997), NGF and S-100b (Ciccarelli et al.,1999). Adenosine also fine tunes the action of several other recep-tor systems in astrocytes, including metabotropic glutamate (Cor-mier et al., 2001), histamine (Peakman and Hill, 1995) and P2Yreceptors (Alloisio et al., 2004).
Adenosine is the end product of sequential enzymatic break-down of extracellular ATP, catalyzed by ecto-50-nucleotidase(CD73; EC 3.1.3.5), which converts AMP to adenosine. Ecto-50-nucleotidase (e-5NT) is a 70 kDa GPI-anchored glycoprotein,widely expressed on the membrane of different cell types through-out the brain (Bjelobaba et al., 2007; Langer et al., 2008). The en-zyme is considered to be the key regulator of extracellular ATPmetabolism and inflammation, as it shifts the balance from pro-inflammatory ATP to anti-inflammatory adenosine. In severalin vivo models of brain injury characterized by sustained inflam-matory astrogliosis, e-5NT expression undergoes biphasic change,with an initial decrease (Bjelobaba et al., 2011; Braun et al.,1998; Nedeljkovic et al., 2008), followed by a prominent up-regu-lation during the post-injury period (Braun et al., 1998; Bonanet al., 2000a,b; Nedeljkovic et al., 2006). It has been demonstratedthat activated astrocytes predominantly account for the observedshift in the total e-5NT activity and protein levels (Bjelobabaet al., 2011).
In the light of our and others previous findings, the aim of thepresent study was to investigate whether pro-inflammatory fac-tors, that are the hallmark of neuroinflammation in several neuro-logical disorders, alter the expression of e-5NT in corticalastrocytes in vitro. Astrocytic cultures were treated with factorsthat mimic a variety of pathological events in the CNS (Glasset al., 2010), such as bacterial lipopolysaccharide (LPS), the proto-type endotoxin that induces inflammatory response (Pitossi et al.,1997), tumor necrosis factor-a (TNF- a) and interferon-c (IFN-c), both the master cytokines involved in glial inflammatory re-sponses (Barcia et al., 2011; Clark et al., 2010), glutamate, an exci-totoxic factor (Werner et al., 2000) and hydrogen peroxide (H2O2),a mediator of oxidative stress (Watt et al., 2004).
2. Materials and methods
2.1. Cortical astrocytes culture and treatments
Primary astrocytes were prepared from 3-day old neonatal Wis-tar pups in accordance with Institutional Animal Care and UseCommittee Guideline. All efforts were made to minimize animalsuffering and to reduce the number of animals used. Cerebral cor-tices were dissected and meninges peeled off in ice-cold
phosphate-buffered saline (PBS). Tissue was subsequentlymechanically dispersed in the culture medium containing 1% tryp-sin, 0.02% EDTA, filtered through a 40-lm nylon mesh and centri-fuged at 400�g for 5 min. Culture medium was Dulbecco’smodified Eagles’s medium with addition of 10% heat-inactivatedfetal calf serum (FCS; both from PAA Laboratories GmbH, Ger-many), 25 mmol/l glucose, 2 mmol/l glutamine, 1 mmol/l so-dium-pyruvate (all from Sigma–Aldrich, St. Louis, USA), 100 IU/ml penicillin and 100 lg/ml streptomycin (both from Invitrogen,Carlsbad, USA). Resuspended mixed glial cells were subsequentlyseeded in 75 cm2 tissue culture flasks (Sarstedt, Nümbrecht, Ger-many) and grown at 37 �C in a humified incubator with 5% CO2.Culture medium was replaced every third day. After 7–10 days inculture, confluence was reached and primary microglial cells andoligodendrocytes were removed by moderate shaking. Adherentprimary astrocytes were washed with PBS, trypsinized (0.25% tryp-sin and 0.02% EDTA, Sigma–Aldrich, St. Louis, USA), and resus-pended in the culture medium. Astrocytes were replated andgrown until confluence. This resulting final astroglial culture wascharacterized by >95% homogeneity of GFAP positive cells, as de-fined by immunofluorescence labeling.
For e-5NT enzyme assays, semi-quantitative RT-PCR, immunob-loting, imunofluorescence imaging and cell viability analyses, cellswere serum-starved (0.5% heat-inactivated fetal calf serum) for24 h and subsequently treated with TNF-a (0.1 lg/ml), IFN-c(100 U/ml), LPS (0.1 lg/ml), glutamate (4 mmol/l) or H2O2
(20 lmol/l) in the culture medium without FCS for 24 h. Non-trea-ted cultures which underwent the same cell culture procedurewere used as controls.
2.2. Preparation of cortical astrocytes membranes
Twenty-four hours after addition of the stimuli, membraneswere isolated from cultured cortical astrocytes. To this end, cells(2 � 106 cells/well) were washed twice with ice-cold PBS and500 ll RIPA buffer with protease inhibitor cocktail (0.5% w/v; Sig-ma–Aldrich) was added and left for 5 min. The cells were harvestedwith a scraper and homogenized using a Teflon pestle homoge-nizer. Obtained suspensions were kept at 4 �C for 1 h with constantmild shaking and were subsequently centrifuged at 60000�g for60 min at 4 �C in a L8-M ultracentrifuge (Beckman Coulter Inc.).The supernatant was kept and protein concentrations determinedby the method developed by Lowry et al. (1951).
2.3. Cell viability assay
To exclude that the applied stimuli induce cell death, cell viabil-ity was determined by evaluating total mitochondrial activity usingthe MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazoliumbromide, Sigma–Aldrich, St. Louis, USA) assay. To this end, cellswere seeded into 96 wells (15 � 103 cells/well) and serum starved,as described above. Cultures were subsequently treated for 24 hwith indicated concentrations of TNF-a, IFN-c, LPS, Glu and H2O2
in serum-free culture medium. On the next day, new medium con-taining 10% of MTT (5 mg/ml) was added to the cultures. Cells wereincubated for 3 h at 37 �C and formazan generated by total mito-chondrial activity was dissolved in DMSO. The optical densitywas measured at 540 nm with an LKB 5060 multiplate reader. Mea-surements were performed in quadruplicate and the viability oftreated cells was expressed as percentage of untreated controls.
2.4. Ecto-50-nucleotidase assay
Ecto-50-nucleotidase activity was assayed as previously de-scribed (Nedeljkovic et al., 2008). The reaction mixture contained10 mmol/l MgCl2, 0.1 mol/l Tris–HCl, pH 7.4 in the final volume

D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688 683
of 200 ll. Five micrograms of astrocyte membrane proteins wereadded into the reaction mixture and preincubated for 5 min at37 �C. The reaction was initiated by the addition of AMP (Sigma–Aldrich, Germany) in a final concentration of 250 lmol/l. Following30 min of incubation, the reaction was stopped by the addition of3 mol/l PCA and by transferring tests tubes on ice. The release ofinorganic phosphate (Pi) was measured by the malachite greenmethod (Chan et al., 1986), with KH2PO4 as a Pi standard. Incuba-tion times, as well as protein and substrate concentrations weredetermined in pilot studies to ensure the linearity of the reactions.Additional control measurements without astrocytic membranepreparation were performed to correct for non-enzymatic hydroly-sis of the substrates.
In order to elucidate if some of the applied stimuli directly af-fect catalytic efficiency of e-5NT, the activity was assayed in astro-cytic membrane preparation obtained from the untreated culture,in the presence or absence of tested stimuli. To this end, membranepreparations were pre-incubated for 30 min with three raisingconcentrations of each tested stimuli prior to the enzyme assay.Applied concentrations of tested stimuli were as follow: TNF-a(0.002, 0.02; 0.1 lg/ml), IFN-c (2; 20; 100 U/ml), LPS (0.002;0.02; 0.1 lg/ml), H2O2 (0.4; 4; 20 lmol/l) and glutamate (0.08;0.8; 4 mmol/l).
The rate of e-5NT activity was expressed as mean activity in nmolof inorganic phosphate released per mg of protein per min (nmolPi/mg protein/min) from n experiments performed in triplicate.
2.5. Real-time PCR (RT-PCR)
Ecto-50-nucleotidase gene expressions was measured using RT-PCR technology (BioRad, Germany), QTM SYBR Green Supermix(BioRad, Germany) and standardized protocol as described previ-ously (Braun et al., 2009). Isolation of total RNA was performedwith the NucleoSpin kit (Macherey–Nagel, Germany). RNA concen-tration and purity were determined using OD260 and OD260/OD280 ratio, respectively and reverse transcribed using an Invitro-gen M-MLV RT-kit and random hexanucleotide primers. Reactionswere carried out in a mixture consisting of 2 ll cDNA, 2 ll RNAse-free water (UltraPure, Invitrogen, Germany), 5 ll QTM SYBR GreenSupermix (Biolmol, Germany) and 0.5 ll primers (100 pmol/ll).Reactions were conducted in standard tubes using MylQ qRT-PCRdetection system (Bio-Rad, Germany) under following conditions:10 min of enzyme activation at 95 �C, 40 cycles of 15 s denatur-ation at 95 �C, 30 s annealing at individual temperatures, 30 samplification at 72 �C and 5 s fluorescence measurement at 72 �C.Primer sequences and individual annealing temperatures are givenin Table 1. Relative quantification was performed using the DCtmethod which results in ratios between target genes and cyclophi-lin A (CycA) as a reference gene (Braun et al., 2009). As the validityof this method critically depends on the constant expression of thehousekeeping gene, this condition was tested in preliminary exper-iments (data not shown). In each run, internal standard curveswere generated by several fold dilutions of generated cDNA. Rela-tive expression of target genes was calculated by comparing Ct val-ues in each sample with Ct values of the internal standard curveand data were expressed as the ratio of the amount of each tran-script vs. concentration of CycA. Melting curves and gel electropho-resis of the PCR products were routinely performed to determinethis specificity of the PCR reaction.
Table 1Primer sequences used for RT-PCR.
Sense
E-5NT caa atc tgc ctc tgg aaa gcCycA ggc aaa tgc tgg acc aaa cac
2.6. Immunoblotting
Equivalent amounts (12 lg) of membrane preparations ob-tained from untreated and treated cortical astrocytes were re-solved on 4–8% SDS–PAGE gradient gels and transferred to PVDFsupport membrane. After blocking with 3% non-fat dry milk in Trisbuffer saline/Tween 20 (TBST), support membranes were probedwith 1:1000 dilution of goat polyclonal e-5NT antibodies (SantaCruz Biotechnology, Santa Cruz, CA) in the same buffer overnightat 4 �C. Visualization procedure was performed on X-ray films (Ko-dak) with the use of chemiluminescence, after incubating supportmembrane for 1 h at room temperature in donkey anti-goat IgG-horseradish peroxidase conjugated secondary antibodies (1:5000dilution; Santa Cruz Biotechnology). The intensity of the bandswas evaluated by densitometric analysis using ImageJ 1.41 soft-ware. The optical density of each protein band was normalizedagainst the optical density of its own b-actin band used as internalstandard on the same lane. For each experiment, the differencesbetween the cultures treated with different stimuli were expressedrelative to the untreated control (100%) and mean effects ± SEM arepresented graphically.
2.7. Fluorescence immunostaining
Six-well culture plates (2 � 105 cells/dish) containing 25 mmpoly-L-lysine coated glass coverslips (Sigma–Aldrich, St. Louis,USA) were incubated for 30 min at room temperature in a blockingsolution containing 10% normal goat serum (NGS), 1% bovine ser-um albumin (BSA) and 0.3% Triton X-100 in PBS. Primary goat poly-clonal antibodies against e-5NT (Santa Cruz Biotechnology Inc.)were diluted 1:100 in 1% BSA containing 0.3% Triton X-100. Slideswere incubated overnight at 4 �C, rinsed in PBS and secondary don-key anti-goat Alexa Fluor 555 antibodies (1:200 dilution; Invitro-gen, Carlsbad, CA) were applied for 90 min at room temperature.After washing with PBS, slides were incubated with anti-GFAPantibody (1:500; DAKO) and then in secondary donkey anti-mouseAlexa 488 antibody (1:200 dilution; Invitrogen, Carlsbad, CA). Be-tween each antibody application, slides were washed in PBS. Slideswere counterstained for 10 min with DAPI (1:10 000 in 0.01 mol/lPBS; Invitrogen, Carlsbad, CA). Negative controls were performedby the omission of the primary antibody which did not result inspecific labeling. Cells were examined under LSM 520 confocalmicroscope system (Carl Zeiss Inc.). At least five different fieldswere imaged from each dish. Mean per-pixel fluorescence intensi-ties were evaluated from whole images after setting the thresholdlevel to exclude ‘‘out-of-cell’’ pixels. Fluorescence intensities of allimages obtained from a dish were averaged. Relative fluorescenceintensity was calculated by comparing fluorescence obtained in atreated culture dish with fluorescence of the untreated control(100%), obtained on the same experimental day.
3. Results
3.1. Cell viability
In the initial experiments effects of TNF-a (0.1 lg/ml), IFN-c(100 U/ml), LPS (0.1 lg/ml), glutamate (4 mmol/l) and H2O2
(20 lmol/l) on astrocyte viability were examined by MTT assay.As illustrated in Fig. 1 applied stimuli for 24 h in the given
Antisense Annealing temp (�C)
acc ttc cag aag gac cct gt 59ggc aaa tgc tgg acc aaa cac 60

Fig. 2. Effect of different stimuli on (A) e-5NT activity and (B) e-5NT-mRNAexpression in cultured cortical astrocytes following 24 h treatment. Data in (A)represent mean activity (nmol Pi/mg protein/min) from n = 4 independent exper-iments performed in triplicate on separately isolated membrane prepara-tions ± SEM. Dot line indicate mean e-5NT activity in the untreated controlculture ± SEM (gray areas). Data in (B) represent mean effect (%) ± SEM from n = 2independent determinations performed in quintuplicate in respect to the controlvalue obtained for untreated cultures, arbitrarily defined as 100 (dot line) ± SEM(gray areas). Significance level shown inside the graphs: ⁄p < 0.05; ⁄⁄p < 0.01;⁄⁄⁄p < 0.001.
684 D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688
concentrations did not affect cell viability of cortical astrocytes, asdeduced from their stable metabolic activity. In the previous studywe were able to show that applied concentrations after 24 h treat-ment were high enough to activate cultured astrocytes (Braunet al., 2009).
3.2. e-5NT activity
In the next set of experiments we studied the effects of testedstimuli on the rate of AMP hydrolysis, that corresponds to theastrocytic e-5NT activity. Conditions for e-5NT acitivity, such asoptimum amount of cell membrane proteins, substrate concentra-tion and duration of incubation, were determined in preliminaryexperiments in order to ensure linearity of the reaction (data notshown). The rate of e-5NT activity in control cultures was3.12 ± 0.16 nmol Pi/mg protein/min. As illustrated in Fig. 2A, therate of e-5NT activity significantly decreased in the membranepreparations obtained from cultures exposed to IFN-c(34.9 ± 3.4%, p < 0.001), LPS (66.1 ± 8.2%, p < 0.05), glutamate(36.8 ± 3.8%, p < 0.001) and H2O2 (49.5 ± 5.7,%, p < 0.001), whereasTNF-a induced statistically significant increase in the enzymeactivity (138.3 ± 15.9%, p < 0.05) compared to control cultures.
3.3. RT-PCR analysis
To investigate if the observed effects of tested stimuli on astro-cytic e-5NT activity were caused by altered e-5NT gene expression,we determined e-5NT-mRNA levels by RT-PCR. As illustrated inFig. 2B, a significant decrease in e-5NT-mRNA levels was detectedin cultures treated with IFN-c (46.1 ± 7.0, p < 0.001), LPS(53.5 ± 7.3, p < 0.001) and glutamate (68.8 ± 9.8%, p < 0.01),whereas treatment with either TNF-a or H2O2 did not result in sig-nificant alterations of e-5NT-mRNA levels. In summary, reduced e-5NT activities observed in IFN-c, LPS, and glutamate treated cul-tures are paralleled by comparable reduction in e-5NT mRNA levels.In contrast, induction of e-5NT activity by TNF-a and its reductionby H2O2 cannot be related to the respective gene expression levels.
3.4. Direct effect of proinflammatory factors on e-5NT activity
Due to the observed discrepancies in e-5NT activity and mRNAlevels in cultures treated with TNF-a and H2O2, we speculated thatsome of the applied stimuli may directly influence e-5NT enzy-matic activity without affecting the transcription machinery. Tothis end, we performed e-5NT assay on the membrane preparation
Fig. 1. Cultured cortical astrocytes viability and proliferation assessed by MTTassay. Bars represent mean treatment effects (%) ± SEM, from n = 2 experimentsperformed in quintuplicate, in respect to the mean optical density (dot line) ± SEM(gray area) obtained in untreated control culture, arbitrarily defined as 100%.
obtained from untreated cultures. Before the assay, the membranesamples were pre-incubated for 30 min with three serial concen-trations of each tested stimuli. As shown in Fig. 3, e-5NT activitywas significantly reduced by H2O2 at a concentration used for theculture treatment (77.1 ± 2.8%, p < 0.05), suggesting that its inhib-itory action could be due to its direct effect on the enzyme mole-cule or on some membrane components that disturb theenzymatic activity. We have also observed slight inhibitory effectof LPS (85.7 ± 1.2%, p < 0.05) and Glu (84.4 ± 1.6%, p < 0.05), whichimply that both factors may directly affect e-5NT catalytic func-tion. A similar effect was not observed in isolated membranes incu-bated with TNF-a prior to the enzyme assay. Specifically, we didnot observe a direct induction of e-5NT activity by TNF-a.
3.5. Immunoblotting and immunofluorescence
While different results in e-5NT activity and mRNA expressioncan be explained by a direct inhibitory effect of H2O2 on e-5NTactivity, this holds not true for TNF-a. We speculated that observedinduction of e-5NT activity in TNF-a-treated culture was due to a

Fig. 3. Effect of tested stimuli on e-5NT activity in astrocytic membrane prepara-tion obtained from untreated cortical astrocytes. Astrocytic membranes isolatedfrom untreated cultures were pre-incubated for 30 min with serial dilutions oftested factors; initial concentrations used for culture treatment (dark gray bars –TNF-a (0.1 lg/ml), IFN-c (100 U/ml), LPS (0.1 lg/ml), H2O2 (20 lmol/l) or glutamate(4 mmol/l)), fivefold dilution (ligh gray bars) and 50-fold dilution of initialconcentration (white bars), before e-5NT enzyme assay (as described in Section 2).Dot line indicates mean e-5NT activity obtained in untreated control culture ± SEM(gray area). Bars represent mean activity (nmol Pi/mg protein/min) from n = 3independent experiments performed in triplicate ± SEM. Significance level:⁄p < 0.05.
D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688 685
shift of e-5NT from cytoplasmic compartment into the cell mem-brane. To test this, we performed immunoblotting experiments,using isolated membrane preparations and immunofluorescencedouble-labeling experiments. As demonstrated in Fig. 4B, mem-branes isolated from TNF-a-treated culture contained a signifi-cantly greater amount of e-5NT protein compared to controls
Fig. 4. Expression of e-5NT by cultured cortical astrocytes assessed by semi-quantitative immunoblot detection in astrocytic membrane preparations obtainedfrom cultures treated with different stimuli for 24 h. (A) Representative immuno-blot; (B) graph representing mean e-5NT protein abundance (%) ± SEM, from n = 3independent determinations, expressed relative to the mean band intensityobtained for untreated culture (dot line), arbitrarily defined as 100%, ± SEM (grayarea). Significance levels: ⁄p < 0.05; ⁄⁄p < 0.01.
(143.8 ± 7.2%, p < 0.05). Accordingly, we observed a comparable in-crease (128.3 ± 7.8%, p < 0.05) in relative e-5NT immunofluores-cence intensity of TNF-a-treated-GFAP-expressing astrocytes(Fig. 5A and B). Furthermore, e-5NT protein levels (Fig. 4) were sig-nificantly reduced in the membrane preparation obtained fromGlu-treated astrocytes (75.3 ± 3.8%, p < 0.01), whereas in IFN-c,LPS and H2O2-treated astrocytes, e-5NT protein levels (Fig. 4) andrelative immunofluorescence intensities (Fig. 5B) remainedunchanged.
4. Discussion
Our study clearly shows that all tested pro-inflammatory fac-tors affect e-5NT activity in cultured cortical astrocytes. LPS, IFN-c, Glu and H2O2 decrease e-5NT activity, whereas TNF-a inducesan up-regulation of the enzyme activity. Decrease in e-5NT activitycould be linked to alteration in the enzyme catalytic efficiancy and/or change in its membrane expression. Since Glu decreases e-5NTactivity, its mRNA and the enzyme protein levels, it is most likelythat Glu affects e-5NT by activating signaling cascade that leadsto transcriptional down-regulation of the enzyme. On the otherhand, H2O2 attenuates e-5NT activity in astrocytic cultures, as wellas in astrocytic membrane preparation directly submitted to per-oxide action, without any change in mRNA and membrane proteinlevels. These findings together strongly suggest that H2O2 directlyaffect e-5NT activity by inducing oxidative modifications and theenzyme protein carbonylation (Takano et al., 2011). LPS and IFN-c decrease e-5NT activity and its mRNA, but do not alter e-5NTprotein levels and membrane abundance. Since LPS directly modu-lates e-5NT catalytic efficiancy, it is most likely that it acts at thetranscriptional level, as well as directly affecting the enzyme. Onthe other hand, IFN-c acts at the transcriptional level, probablyon the longer time scale. Finaly, TNF-a is unique among testedstimuli, since it stimulates e-5NT activity, increases its membraneabundance, without change in the mRNA level, implying that itacts at post-transcriptional and/or translational levels probablyby mobilizing e-5NT peptides from the pre-existing intracellularpools.
From the mechanicistic point of view, inhibition of e-5NT byLPS, IFN-c, Glu and H2O2 increases AMP levels, while stimulationof e-5NT activity by TNF-a leads to the accumulation of adenosinein the pericellular space. AMP and adenosine oppositely affectastrocytic environment, as AMP promotes (Hide et al., 2000; Sanzand Virgilio, 2000), while adenosine attenuates (Kreckler et al.,2009; Romio et al., 2011) the release of several pro-inflammatorycytokines, including TNF-a and IL-1b. It appears that tested stimuli,i.e., LPS, IFN-c, Glu and H2O2 on one side and TNF-a on the other,exert opposite effects on local adenosine and AMP levels andaccordingly, on astrocyte inflammatory response. The situation,however, appears to be more complex in vivo, with the presenceof different cell types and many inflammatory mediators that actin a combinatorial manner and in a precisely regulated temporalsequence (Yasuda et al., 2011). Namely, it is well documented thatmicroglial and astroglial responses occur separately in time, withmicroglial activation preceding astrogliosis (Graeber and Kreutz-berg, 1998; Schmitt et al., 1998). The canonical LPS receptors,TRL4, are abundantly expressed on microglia, whereas in verylow or undetectable levels at astrocytes and neurons, respectively(Kim et al., 2000). Consistent with this finding, microglia are muchmore responsive to LPS as astrocytes, whereas neurons are virtu-ally unresponsive (Saijo et al., 2009). Therefore, in an in vivo situa-tion microglia are likely the primary responders to LPS thatinitiates a signaling cascade leading to the induction of inflamma-tory response and release of proinflammatory cytokines, such asIFN-c and TNF-a (Hanisch, 2002). Induction of an inflammatory re-
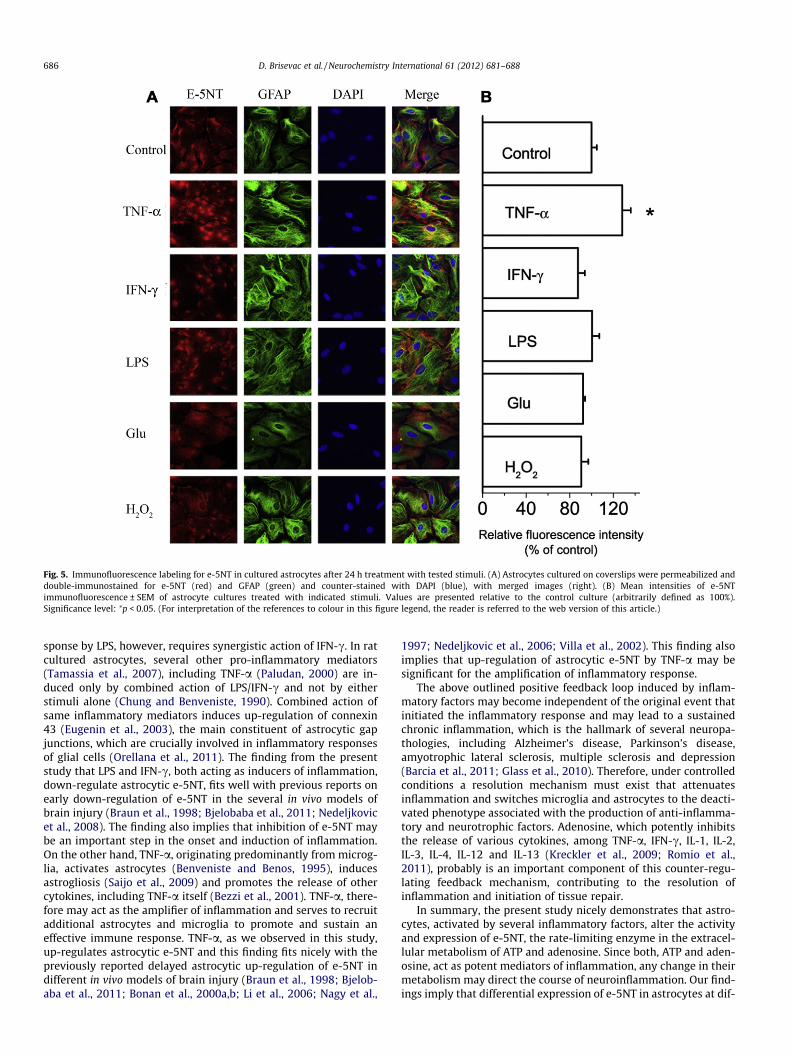
Fig. 5. Immunofluorescence labeling for e-5NT in cultured astrocytes after 24 h treatment with tested stimuli. (A) Astrocytes cultured on coverslips were permeabilized anddouble-immunostained for e-5NT (red) and GFAP (green) and counter-stained with DAPI (blue), with merged images (right). (B) Mean intensities of e-5NTimmunofluorescence ± SEM of astrocyte cultures treated with indicated stimuli. Values are presented relative to the control culture (arbitrarily defined as 100%).Significance level: ⁄p < 0.05. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
686 D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688
sponse by LPS, however, requires synergistic action of IFN-c. In ratcultured astrocytes, several other pro-inflammatory mediators(Tamassia et al., 2007), including TNF-a (Paludan, 2000) are in-duced only by combined action of LPS/IFN-c and not by eitherstimuli alone (Chung and Benveniste, 1990). Combined action ofsame inflammatory mediators induces up-regulation of connexin43 (Eugenin et al., 2003), the main constituent of astrocytic gapjunctions, which are crucially involved in inflammatory responsesof glial cells (Orellana et al., 2011). The finding from the presentstudy that LPS and IFN-c, both acting as inducers of inflammation,down-regulate astrocytic e-5NT, fits well with previous reports onearly down-regulation of e-5NT in the several in vivo models ofbrain injury (Braun et al., 1998; Bjelobaba et al., 2011; Nedeljkovicet al., 2008). The finding also implies that inhibition of e-5NT maybe an important step in the onset and induction of inflammation.On the other hand, TNF-a, originating predominantly from microg-lia, activates astrocytes (Benveniste and Benos, 1995), inducesastrogliosis (Saijo et al., 2009) and promotes the release of othercytokines, including TNF-a itself (Bezzi et al., 2001). TNF-a, there-fore may act as the amplifier of inflammation and serves to recruitadditional astrocytes and microglia to promote and sustain aneffective immune response. TNF-a, as we observed in this study,up-regulates astrocytic e-5NT and this finding fits nicely with thepreviously reported delayed astrocytic up-regulation of e-5NT indifferent in vivo models of brain injury (Braun et al., 1998; Bjelob-aba et al., 2011; Bonan et al., 2000a,b; Li et al., 2006; Nagy et al.,
1997; Nedeljkovic et al., 2006; Villa et al., 2002). This finding alsoimplies that up-regulation of astrocytic e-5NT by TNF-a may besignificant for the amplification of inflammatory response.
The above outlined positive feedback loop induced by inflam-matory factors may become independent of the original event thatinitiated the inflammatory response and may lead to a sustainedchronic inflammation, which is the hallmark of several neuropa-thologies, including Alzheimer’s disease, Parkinson’s disease,amyotrophic lateral sclerosis, multiple sclerosis and depression(Barcia et al., 2011; Glass et al., 2010). Therefore, under controlledconditions a resolution mechanism must exist that attenuatesinflammation and switches microglia and astrocytes to the deacti-vated phenotype associated with the production of anti-inflamma-tory and neurotrophic factors. Adenosine, which potently inhibitsthe release of various cytokines, among TNF-a, IFN-c, IL-1, IL-2,IL-3, IL-4, IL-12 and IL-13 (Kreckler et al., 2009; Romio et al.,2011), probably is an important component of this counter-regu-lating feedback mechanism, contributing to the resolution ofinflammation and initiation of tissue repair.
In summary, the present study nicely demonstrates that astro-cytes, activated by several inflammatory factors, alter the activityand expression of e-5NT, the rate-limiting enzyme in the extracel-lular metabolism of ATP and adenosine. Since both, ATP and aden-osine, act as potent mediators of inflammation, any change in theirmetabolism may direct the course of neuroinflammation. Our find-ings imply that differential expression of e-5NT in astrocytes at dif-

D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688 687
ferent phases during the course of neuroinflammation may have apivotal role in the induction, amplification and counter-regulationof inflammatory response in the CNS. The question remains if mod-ulation of e-5NT and other ectonucleotidases as well, may be aneffective strategy to slow or even reverse the course of an inflam-matory response in neurodegenerative disseases associated withsustained chronic inflammation.
Acknowledgments
This work was funded by Serbian Ministry for Education andScience Project No. III41014 and Bilateral Project between DAAD/Serbian Ministry of Science and Education (2010). D.B. was sup-ported by DAAD financed Scientific Internship in Germany forUndergraduates and Graduates from Serbia.
References
Alloisio, S., Cugnoli, C., Ferroni, S., Nobile, M., 2004. Differential modulation of ATP-induced calcium signaling by A1 and A2 adenosine receptors in cultured corticalastrocytes. Br. J. Pharmacol. 141, 935–942.
Aloisi, F., Ria, F., Adorini, L., 2000. Regulation of T-cell responses by CNS antigen-presenting cells: different roles for microglia and astrocytes. Immunol. Today21, 141–147.
Barcia, C., Ros, C.M., Annese, V., Gomez, A., Ros-Bernai, F., Aguardo-Yera, D.,Martinez-Pagan, M.E., de Pablos, V., Fernandez-Villalba, E., Herrero, M.T., 2011.IFN-c signaling, with the synergistic contribution of TNF-a, mediates cellspecific microglial and astroglial activation in experimental models ofParkinson’s disease. Cell Death Dis.. http://dx.doi.org/10.1038/cddis.2011.17.
Benveniste, E.N., Benos, D.J., 1995. TNF-alpha and IFN-gamma-mediated signaltransduction pathways: effects on glial cell gene expression and function.FASEB J. 9, 1577–1584.
Bezzi, P., Domercq, M., Brambilla, L., Galli, R., Schols, D., De Clercq, E., Vescovi, A.,Bagetta, G., Kollias, G., Meldolesi, J., Volterra, A., 2001. CXCR4-activatedastrocyte glutamate release via TNF-a: amplification by microglia triggersneurotoxicity. Nat. Neurosci. 4, 702–710.
Bjelobaba, I., Stojiljkovic, M., Pekovic, S., Dacic, S., Lavrnja, I., Stojkov, D., Rakic, L.,Nedeljkovic, N., 2007. Immunohistological determination of ecto-nucleosidetriphosphate diphosphohydrolase1 (NTPDase1) and 50-nucleotidase in rathippocampus reveals overlapping distribution. Cell. Mol. Neurobiol. 27, 731–743.
Bjelobaba, I., Parabucki, A., Lavrnja, I., Stojkov, D., Dacic, S., Pekovic, S., Rakic, L.,Stojiljkovic, M., Nedeljkovic, N., 2011. Dynamic changes in the expressionpattern of ecto-50-nucleotidase in the rat model of cortical stab injury. J.Neurosci. Res. 89, 862–873.
Bolego, C., Ceruti, S., Brambilla, R., Puglisi, R., Cattabeni, F., Burnstock, G.,Abbracchio, M.P., 1997. Characterization of the signaling pathway involved inATP and basic fibroblast growth factor induced astrogliosis. Br. J. Pharmacol.121, 1692–1699.
Bonan, C.D., Amaral, O.B., Rockenbach, I.C., Walz, R., Battastini, A.M., Izquierdo, I.,Sarkis, J.J., 2000a. Altered ATP hydrolysis induced by pentylenetetrazol kindlingin rat brain synaptosomes. Neurochem. Res. 25, 775–779.
Bonan, C.D., Walz, R., Pereira, G.S., Worm, P.V., Battastini, A.M., Cavalheiro, E.A.,Izquierdo, I., Sarkis, J.J., 2000b. Changes in synaptosomal ectonucleotidaseactivities in two rat models of temporal lobe epilepsy. Epilepsy Res. 39, 229–238.
Brambilla, R., Cottini, L., Fumagalli, M., Ceruti, S., Abbracchio, M.P., 2003. Blockade ofA2A adenosine receptors prevents basic fibroblast growth factor-inducedreactive astrogliosis in rat striatal primary astrocytes. Glia 43, 190–194.
Braun, N., Zhu, Y., Krieglstein, J., Culmsee, C., Zimmermann, H., 1998. Upregulationof the enzyme chain hydrolyzing extracellular ATP after transient ferebrainischemia in the rat. J. Neurosci. 18, 4891–4900.
Braun, A., Dang, J., Johann, S., Beyer, C., Kipp, M., 2009. Selective regulation ofgrowth factor expression in cultured cortical astrocytes by neuro-pathologicaltoxins. Neurochem. Int. 55, 610–618.
Cavaliere, F., Florenzano, F., Amadio, S., Fusco, F.R., Viscomi, M.T., D’Ambrosi, N.,Vacca, F., Sancesario, G., Bernardi, G., Molinari, M., Volonte, C., 2003.Upregulation of P2X2, P2X4 receptors and ischemic cell death: prevention byP2 antagonists. Neuroscience 120, 85–98.
Chan, K.M., Delfert, D., Junger, K.D., 1986. A direct colorimetric assay for Ca2+-stimulated ATPase activity. Anal. Biochem. 157, 375–380.
Chung, I.Y., Benveniste, E.N., 1990. Tumor necrosis factor-a production byastrocytes. Induction by lipopolysaccharide, IFN-c and IL-1b. J. Immunol. 144,2999–3007.
Ciccarelli, R., Di Iorio, P., Bruno, V., Battaglia, G., D’Alimonte, I., D’Onofrio, M.,Nicoletti, F., Caciagli, F., 1999. Activation of A1 adenosine or mGlu3metabotropic glutamate receptors enhances the release of nerve growthfactor and S-100b protein from cultured astrocytes. Glia 27, 275–281.
Clark, I.A., Alleva, L.M., Vissel, B., 2010. The roles of TNF in brain dysfunction anddisease. Pharmacol. Ther. 128, 519–548.
Cormier, R.J., Mennerick, S., Melbostad, H., Zorumski, C.F., 2001. Basal levels ofadenosine modulate mGluR5 on rat hippocampal astrocytes. Glia 33, 24–35.
Dall’Igna, O.P., Souza, D.O., Lara, D.R., 2004. Caffeine as a neuroprotective adenosinereceptor antagonist. Ann. Pharmacother. 38, 717–718.
Eugenin, E.A., Branes, M.C., Berman, J.W., Saez, J.C., 2003. TNF-a plus IFN-c induceconnexin 43 expression and formation of gap junctions between humanmonocytes/macrophages that enhance physiological responses. J. Immunol.170, 1320–1328.
Farina, C., Aloisi, F., Meinl, E., 2007. Astrocytes are active players in cerebral innateimmunity. Trends Immunol. 28, 138–145.
Ferrari, C.C., Pott Godoy, M.C., Tarelli, R., Chertoff, M., Depino, A.M., Pitossi, F.J., 2006.Progressive neurodegeneration and motor disabilities induced by chronic IL-1bin in the substantia nigra. Neurobiol. Dis. 24, 183–193.
Fiebich, B.L., Biber, K., Gyufko, K., Berger, M., Bauer, J., van Calker, D., 1996.Adenosine A2B receptors mediate an increase in interleukin (IL)-6 mRNA andIL-6 protein synthesis in human astrocytoma cells. J. Neurochem. 66, 1426–1431.
Frengueli, B.G., Wigmore, G., Llaudet, E., Dale, N., 2007. Temporal and mechanisticdissociation of ATP and adenosine release during ischemia in the mammalianhippocampus. J. Neurochem. 101, 1400–1413.
Glass, C.K., Saijo, K., Winner, B., Marchetto, M.C., Gage, F.H., 2010. Mechanismsunderlying inflammation in neurodegeneration. Cell 140, 918–934.
Graeber, M.B., Kreutzberg, G.W., 1998. Delayed astrocyte reaction following facialnerve axotomy. J. Neurocytol. 17, 209–220.
Gudipaty, L., Munetz, J., Verhoef, P.A., Dubyak, G.R., 2003. Essential role for Ca2+ inregulation of IL-1 beta secretion by P2X7 nucleotide receptor in monocytes,macrophages and HEK-239 cells. Am. J. Physiol. Cell Physiol. 285, C286–C299.
Gyoneva, S., Orr, A.G., Traynelis, S.F., 2009. Differential regulation of microglialmotility by ATP/ADP and adenosine. Parkinsonism Relat. Disord. 1553, S195–S199.
Hanisch, U.K., 2002. Microglia as a source and target of cytokines. Glia 40, 140–155.Hasko, G., Cronstein, B.N., 2004. Adenosine: an endogenous regulator of innate
immunity. Trends Immunol. 25, 33–39.Herx, L.M., Young, V.W., 2001. Interleukin-1 beta is required for the early evolution
of reactive gliosis following CNS lesion. J. Neuropathol. Exp. Neurol. 60, 961–971.
Hide, I., Tanaka, M., Inoue, A., Nakajima, K., Kohsaka, S., Inoue, K., Nakata, Y., 2000.Extracellular ATP triggers tumor necrosis factor-alpha release from ratmicroglia. J. Neurochem. 75, 965–972.
Hindley, S., Herman, M.A., Rathbone, M.P., 1994. Stimulation of reactive astrogliosisin vivo by extracellular adenosine diphosphate or an adenosine A2 receptoragonist. J. Neurosci. Res. 38, 399–406.
Janeway, C.A., 1992. The immune system evolved to discriminate infectious nonselffrom noninfectious self. Immunol. Today 13, 111–116.
Johann, S., Kampmann, E., Denecke, B., Arnold, S., Kipp, M., Mey, J., Beyer, C., 2008.Expression of enzymes involved in the prostanoid metabolism by corticalastrocytes after LPS-induced inflammation. J. Mol. Neurosci. 34, 177–185.
Kataoka, A., Tozaki-Saitoh, H., Koga, Y., Tsuda, M., Inoue, K., 2009. Activation of P2X7receptors induces CCL3 production in microglial cells through transcriptionfactor NFAT. J. Neurochem 108, 115–125.
Kim, W.G., Mohney, R.P., Wilson, B., Jeohn, G.H., Liu, B., Hong, J.S., 2000. Regionaldifferences in susceptibility to lipopolysaccharide-induced neurotoxicity in therat brain: role of microglia. J. Neurosci. 20, 6309–6316.
Kim, J.H., Min, K.J., Seol, W., Jou, I., Joe, E.H., 2010. Astrocytes in injury state rapidlyproduce anti-inflammatory factors and attenuate microglial inflammatoryresponses. J. Neurochem. 115, 1161–1171.
Kipp, M., Gingele, S., Pott, F., Clarner, T., van der Valk, P., Denecke, B., Gan, L., Siffrin,V., Zipp, F., Dreher, W., Baumgartner, W., Pfeifenbring, S., Godbout, R., Amor, S.,Beyer, C., 2011. BLBP-expression in astrocytes during experimentaldemyelination and in human multiple sclerosis lesions. Brain Behav.Immunol. 25, 1554–1568.
Kreckler, L.M., Gizewski, E., Wan, T.C., Auchampach, J.A., 2009. Adenosinesupresses lipopolysaccharide-induced tumor necrosis factor-alpha productionby murine macrophages through a protein kinase A and exchange proteinactivated by cAMP-independent signaling pathway. J. Pharmacol. Exp. Ther.331, 1051–1061.
Kucher, B.M., Neary, J.T., 2005. Bi-functional effects of ATP/P2 receptor activation ontumor necrosis factor-alpha release in lipopolysaccharide-stimulatedastrocytes. J. Neurochem. 92, 525–535.
Langer, D., Hammer, K., Koszalka, P., Schrader, J., Robson, S., Zimmermann, H., 2008.Distribution of ectonucleotidases in the rodent brain revisited. Cell. Tissue Res.334, 199–217.
Le Feuvre, R., Brough, D., Rothwell, N., 2002. Extracellular ATP and P2X7 receptors inneurodegeneration. Eur. J. Pharmacol. 447, 261–269.
Li, X., Zhou, T., Zhi, X., Zhao, F., Yin, L., Zhou, P., 2006. Effect of hypoxia/reoxygenation on CD73 (ecto-50-nucleotidase) in mouse microvesselendothelial cell lines. Microvasc. Res. 72, 48–53.
Liu, J., Gao, X.-F., Ni, W., Li, J.-B., 2011. Effects of propofol on P2X7 receptors and thesecretion of tumor necrosis factor-a in cultured astrocytes. Clin. Exp. Med..http://dx.doi.org/10.1007/s10238-011-0139-4.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Matzinger, P., 2002. The danger model: a renewed sense of self. Science 295, 301–305.
Nagy, A.K., Walton, N.Y., Treiman, D.M., 1997. Reduced cortical ecto-ATPase activityin rat brains during prolonged status epilepticus induced by sequential

688 D. Brisevac et al. / Neurochemistry International 61 (2012) 681–688
administration of lithium and pilocarpine. Mol. Chem. Neuropathol. 31, 135–147.
Nedeljkovic, N., Bjelobaba, I., Subasic, S., Lavrnja, I., Pekovic, S., Stojkov, D., Vjestica,A., Rakic, L., Stojiljkovic, M., 2006. Up-regulation of ectonucleotidase activityafter cortical stab injury in rats. Cell. Biol. Int. 30, 541–546.
Nedeljkovic, N., Bjelobaba, I., Lavrnja, I., Stojkov, D., Pekovic, S., Rakic, L., Stojiljkovic,M., 2008. Early temporal changes in ecto-nucleotidase activity after corticalstab injury in rat. Neurochem. Res. 33, 873–879.
Orellana, J.A., Froger, N., Ezan, P., Jiang, J.X., Bennett, M.V., Naus, C.C., Giaume, C.,Saez, J.C., 2011. ATP and glutamate released via astroglial connexin 43hemichannels mediate neuronal death through activation of pannexon 1hemichannels. J. Neurochem. 118, 826–840.
Paludan, S.R., 2000. Synergistic action of pro-inflammatory agents: cellular andmolecular aspects. J. Leukocyte Biol. 67, 18–25.
Peakman, M.C., Hill, S.J., 1995. Adenosine A1 receptor-mediated changes in basaland histamine – stimulated levels of intracellular calcium in primary ratastrocytes. Br. J. Pharmacol. 115, 801–810.
Pitossi, F., Delrey, A., Kabiersch, A., Besedovsky, H., 1997. Induction of cytokinetranscripts in the central nervous system and pituitary following peripheraladministration of endotoxin in mice. J. Neurosci. Res. 48, 287–298.
Romio, M., Reinbeck, B., Bongardt, S., Huls, S., Burghoff, S., Schrader, J., 2011.Extracellular purine metabolism and signaling of CD-73 derived adenosine inmurine Treg and Teff cells. Am. J. Physiol. Cell Physiol. 301, C530–539.
Saijo, K., Winner, B., Carson, C.T., Collier, J.G., Boyer, L., Rosenfeld, M.G., Gage,F.H., Glass, C.K., 2009. A Nurrq/CoREST pathway in microglia and astrocytesprotects dopaminergic neurons from inflammation-induced death. Cell 137,47–59.
Sanz, J.M., Virgilio, F., 2000. Kinetics and mechanism of ATP-dependent Il-1b releasefrom microglial cells. J. Immunol. 164, 4893–4898.
Schmitt, A.B., Brook, G.A., Buss, A., Nacimiento, W., Noth, J., Kreutzberg, G.W., 1998.Dynamics of microglial activation in the spinal cord after cerebral infarctationare revealed by expression of MHC class II antigen. Neuropathol. Appl.Neurobiol. 24, 167–176.
Schwaninger, M., Neher, M., Viegas, E., Schneider, A., Spranger, M., 1997.Stimulation of interleukin-6 secretion and gene transcription in primaryastrocytes by adenosine. J. Neurochem. 69, 1145–1150.
Selmaj, K.W., Farooq, M., Norton, W.T., Raine, C.S., Brosnan, C.F., 1990. Proliferationof astrocytes in vitro in response to cytokines. A primary role for tumor necrosisfactor. J. Immunol. 144, 129–135.
Sitkovsky, M.V., Lukashev, D., Apasov, S., Kojima, H., Koshiba, M., Caldwell, C., Ohta,A., Thiel, M., 2004. Physiological control of immune response and inflammatorytissue damage by hypoxia-inducible factors and adenosine A2A receptors.Annu. Rev. Immunol. 22, 657–682.
Suzuki, T., Hide, I., Ido, K., Kohsaka, S., Inoue, K., Nakata, Y., 2004. Production andrelease of neuroprotective tumor necrosis factor by P2X7 receptor-activatedmicroglia. J. Neurosci. 24, 1–7.
Takano, K., Sugita, K., Moriyama, M., Hashida, K., Hibino, S., Choshi, T., Murakami, R.,Yamada, M., Suzuki, H., Hori, O., Nakamura, Y., 2011. A dibenzoylmethanederivate protects against hydrogen peroxide-induced cell death and inhibitslipopolysaccharide-induces nitric oxide production in cultured astrocytes. J.Neurosci. Res. 89, 955–965.
Tamassia, N., Calzetti, F., Ear, T., Cloutier, A., Gasperini, S., Bazzoni, F., McDonald, P.P.,Casatella, M.A., 2007. Molecular mechanisms underlying the synergisticinduction of CXCL10 by LPS and IFN-c in human neutrophils. Eur. J. Immunol.37, 2627–2634.
Verkhratsky, A., Kristhal, O.A., Burnstock, G., 2009. Purinoreceptors on neuroglia.Mol. Neurobiol. 39, 190–208.
Villa, R.F., Gorini, A., Hoyer, S., 2002. ATPases of synaptic plasma membranes fromhippocampus after ischemia and recovery during aging. Neurochem. Res. 27,861–870.
Virgilio, F.D., Ceruti, S., Bramanti, P., Abbracchio, M.P., 2009. Purinergic signalling ininflammation of the central nervous system. Trends Neurosci. 32, 79–87.
Watt, B.E., Proudfoot, A.T., Vale, J.A., 2004. Hydrogen peroxide poisoning. Toxicol.Rev. 23, 51–57.
Werner, P., Pitt, D., Raine, C.S., 2000. Glutamate excitotoxicity – a mechanism foraxonal damage and oligodendrocyte death in multiple sclerosis? J. Neural.Transm. Suppl. 60, 375–385.
Witting, A., Walter, L., Wacker, J., Moller, T., Stella, N., 2004. P2X7 receptors control2-arachidonoylglycerol production by microglial cells. Proc. Natl. Acad. Sci. USA101, 3214–3219.
Yasuda, Y., Shimoda, T., Uno, K., Tateishi, N., Furuya, S., Tsuchihashi, Y., Kawai, Y.,Naruse, S., Fujita, S., 2011. Temporal and sequential changes of glial cells andcytokine expressine during neuronal degeneration after transient globalischemia in rats. J. Neuroinflamm. 8, 70. http://dx.doi.org/10.1186/1742-2094-8-70.
Zhang, D., Hu, X., Qian, L., Wilson, B., Lee, C., Flood, P., Langenbach, R., Hong, J.S.,2009. Prostaglandin E2 released from activated microglia enhances astrocyteproliferation in vitro. Toxicol. Appl. Pharmacol. 238, 64–70.
Copyright © 2022 FDOKUMEN




















