
Postsynaptic D2 dopamine receptor supersensitivity in the striatum of mice lacking TAAR1

Reduction of Zinc-Positive TerminalFields in Striatum of Mouse after
1-Methyl-4-Phenylpyridinium NeurotoxicityPatricia Rojas 1,3,*, Javier E. Franco-Perez 1, Carolina Rojas 2, Julio Rojas-Castaneda 4,
Manuchair Ebadi 3, Francisca Fernandez-Valverde 1, Norma Serrano-Garcıa 1
1Laboratory of Neurotoxicology, Instituto Nacional de Neurologıa y Neurocirugıa, Manuel Velasco Suarez, SS, Av.
Insurgentes Sur No. 3877, C.P. 14269 Mexico D.F., Mexico2 Instituto de Investigaciones Biomedicas, Department of Cellular Biology and Physiology, Universidad Nacional
Autonoma de Mexico Apartado Postal 70228, C.P. 04510 Mexico D.F., Mexico3Department of Pharmacology, University of North Dakota, School of Medicine and Health Sciences, 501 North
Columbia Road, Grand Forks, ND 58203, USA4Laboratory of Histomorphology, Instituto Nacional de Pediatrıa, Av. Insurgentes Sur No.3700-C, Col. Insurgentes
Cuicuilco, C.P. 04530 Mexico D.F., Mexico
Received 4 October 2004; accepted 4 April 2005
Available online 9 June 2005
Abstract
Zinc is an essential trace element in the central nervous system and is located in three distinct pools: free zinc,
vesicular zinc and protein-bound zinc. Zinc may serve as an endogenous neuromodulator and has been associated with
neuropathologies. This study was undertaken to determine whether levels of vesicular zinc in neuronal terminals would
decrease in response to the dopaminergic neurotoxin 1-methyl-4-phenylpyridinium ion (MPP+). Adult male C-57 black
mice were injected with MPP+ (0.72 mg/kg) into their right lateral ventricle. All animals were killed at 1, 2, 24 h and 7
days after MPP+ or saline administration. The brains were stained for zinc sulfides and the density of zinc-positive
terminal fields was evaluated after MPP+ administration. The relative optical density analysis of zinc-positive terminal
fields showed significant decreases in the striatum at 1, 2 and 24 h (24, 18 and 14%, respectively, versus control) and
ventricular epithelium (1, 2, 24 h and 7 days). The hippocampus showed increase in the stratum oriens and stratum
radiatum at different times. MPP+ administration reduced dopamine levels at 24 h and 7 days (36 and 40%, respectively,
versus control) as a result of the neurotoxic action of MPP+. The decrease of zinc-positive neuronal terminal fields in the
striatum after MPP+ administration is most likely due to a neuronal release of vesicular zinc in response to its
dopaminergic neurotoxicity.
# 2005 Elsevier Inc. All rights reserved.
Keywords: Zinc; 1-Methyl-4-phenylpyridinium; Striatum; Histochemistry; Timm’s method
NeuroToxicology 26 (2005) 959–968
INTRODUCTION
Parkinson’s disease (PD) is a neurological disorder
characterized by degeneration and death of the dopa-
minergic neurons of the nigrostriatal pathway in the
* Corresponding author. Tel.: +52 55 5606 4040;
fax: +52 55 5424 0808.
E-mail address: [email protected] (P. Rojas).
0161-813X/$ – see front matter # 2005 Elsevier Inc. All rights reserv
doi:10.1016/j.neuro.2005.04.007
brain (Jenner, 1989). Death of these neurons produces a
decrease in striatal dopamine (DA) content. Although
several molecular mechanisms have been proposed for
the pathogenesis of this disorder, its etiology remains
unclear. However, there is increasing evidence suggest-
ing that oxidative stress may be involved in the neu-
ronal death (Owen et al., 1996).
1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MP-
TP) is regarded as the best available experimental model
ed.

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968960
for the neurochemical sequelae of PD (Gerlach et al.,
1991). MPTP is a compound, which selectively affects
the dopaminergic (DAergic) neurons of the nigrostriatal
pathway. The neurotoxic mechanism of the drug is not
well understood, but its toxicity has been related to
oxidative stress via the production of reactive oxygen
species (Adams and Odunze, 1991). MPTP toxicity
depends on its conversion to the toxic metabolite 1-
methyl-4-phenylpyridinium (MPP+) by monoamine
oxidase (MAO) type B (Chiba et al., 1984). The high
concentration of MPP+ inside mitochondria blocks com-
plex I activity in the respiratory chain. MPP+ induces
free radical production in mitochondria, a process that
can further inhibit the function of complex I in mito-
chondria (Adams et al., 1993).
On the other hand, manganese and copper constitute
part of the active sites of the superoxide dismutase, an
enzyme that catalyzes the destruction of superoxide
free radicals. We found depletion of manganese and
copper following MPTP administration (Rıos et al.,
1995). Pretreatment with these metals protected
against MPTP/MPP+ neurotoxicity (Rojas and Rıos,
1995; Alcaraz-Zubeldia et al., 2001).
Assuming the great ability of MPTP to produce free
radicals by a metal-catalyzed reaction, the effects of
metals on this process would seem particularly useful
for a better understanding of both the mechanisms
underlying the neurotoxicity of MPTP/MPP+ and the
suggested involvement of this neurotoxin in the patho-
genesis of PD (Kienzl et al., 1995).
In particular, with the exception of calcium and
magnesium, zinc is the most abundant cation in the
brain. There are three main sources of zinc in the brain,
namely: (a) a vesicular pool localized in the synaptic
vesicles of nerve terminals; (b) a membrane-bound,
metalloprotein, or protein–metal complex pool
involved in both metabolic reactions and non-meta-
bolic reactions, such as structural support for biomem-
branes and protein folds; (c) an ionic pool of free or
loosely bound ions in the cytoplasm (Frederickson and
Danscher, 1990).
In the brain, in addition to its general functions as a
cofactor of several enzymes (Vallee and Falchuk,
1993), zinc is present in nerve terminals throughout
the mammalian central nervous system, where it plays
important roles as an endogenous signaling substance
(Walsh et al., 1994). They are more prominent parti-
cularly in the neocortex, hippocampal formation, piri-
form cortex, amygdaloid complex, striatum and
septum (Perez-Clausell and Danscher, 1985). Zinc is
actively taken up by central neurons, stored in their
synaptic vesicles (Palmiter et al., 1996) and released
into the brain interstitial space after neuronal depolar-
ization (Frederickson and Danscher, 1990).
Zinc can affect numerous membrane-spanning pro-
teins besides the glutamate receptors, including the
GABA receptors, the DA pump and some types of Na+
and K+ channels (for a review, see Frederickson and
Mocrieff, 1994).
Zinc is a non-transition metal ion, which has often
been associated with the pathogenesis of PD (Forsleff
et al., 1999). Zinc plays a protective role in physiolo-
gical conditions and its antioxidant properties in rela-
tion to brain oxidative stress have been reported (Ebadi
et al., 1996; Tate et al., 1999).
This study was undertaken to determine whether
levels of vesicular zinc in striatum would decrease in
response to MPP+ administration using Timm’s method.
MATERIALS AND METHODS
Animals
All experiments were conducted on male C-57 black
mice (25–30 g) aged 11–13 weeks and 6–8 animals for
each experimental group, obtained from Harlan (Mex-
ico City, Mexico). Animals were maintained in stan-
dard conditions (12-h light:12-h dark cycle, 21 � 2 8C,
relative humidity 40%) and allowed access to food and
water ad libitum. All animals were treated humanely to
minimize discomfort in accordance with the ethical
principles and regulations specified by Animal Care
and Use Committee of our Institution and the standards
of the National Institutes of Health of Mexico.
Fifty-four male C-57 black mice were allocated in
one of the eight groups: group I (n = 6), saline solution
(intracerebroventricular, i.c.v.) for 1 h treatment; group
II (n = 6), MPP+ (i.c.v.) group for 1 h treatment; group
III (n = 6), saline solution (i.c.v.) for 2 h treatment;
group IV (n = 6), MPP+ (i.c.v.) group for 2 h treatment;
group V (n = 8), saline solution (i.c.v.) for 24 h treat-
ment; group VI (n = 6), MPP+ (i.c.v.) group for 24 h
treatment; group VII (n = 8), saline solution (i.c.v.) for
7 days treatment; group VIII (n = 8), MPP+ (i.c.v.)
group for 7 days treatment.
Chemicals
MPP+ iodine was obtained from Research Biochem-
icals Incorporated (Wayland, MA), sodium sulfide
from Fisher Scientific (Pittsburgh, PA) and parafor-
maldehyde, glutaraldehyde and other reagents from
Sigma Chemical Co. (St. Louis, MO).

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968 961
MPP+ Administration and Tissue Preparation
MPP+ at a dose of 0.72 mg/kg was injected into the
right lateral ventricle of mice as we previously
described (Rojas and Rıos, 1993). This MPP+ dose
has been shown to produce significant damage to
dopaminergic neurons (Mihatsch et al., 1988). Animals
similarly injected with saline solution, served as con-
trols. At 1, 2, 24 h and 7 days mice from both groups
were anesthetized with ether and perfused transcar-
dially with 0.1 M Na2S in 0.2 M phosphate buffer
followed by 1% paraformaldehyde and 1.25% glutar-
aldehyde in 0.1 M phosphate buffer at pH 7.4 (West
and Hodges-Savola, 1983; West and Dewey, 1984).
Sulfide perfusion produces precipitation of endogenous
zinc, which is silver-amplified later on tissue sections.
After perfusion, the brains were removed and post-
fixed in the same fixative to which has been added 30%
sucrose. They remained in this solution until they sank
to the bottom of the container. Brains were sectioned in
the coronal and frontal planes on a cryostat (Jung
Frigocut 2800 N, Leica). At every 240 mm interval,
three consecutive sections were cut at a thickness of
20 mm. The series were collected on gelatin-coated
slides and processed using a modification of Timm’s
sulfide–silver technique (Cintra et al., 1997). Devel-
opment was performed in the dark at 26 8C for 60 min.
Two series were counterstained with cresyl violet or
Toluidine Blue.
Relative Optical Density
Densitometric analysis of zinc sulfide-positive
Timm-stained terminals was done using a Metamorph
Imaging System (Version 4.5, Metamorph Imaging
Corporations, PA, USA). The pictures were converted
to a standard video signal using a black and white CCD
video camera (Sony CCD-Iris), connected to a Nikon
Eclipse E600 microscope. The video signal was digi-
tized using an eight bit flash converter with an imagen
size of 512 pixels � 480 pixels frames, providing grey
scale values between 0 and 255. The density of Timm
stain was examined in striatum, hippocampus, hypotha-
lamus and ventricles at 10� magnification. The optical
density was measured by placing 50 circular cursors (5
for each section), each 40 mm2 in area, along 10 digi-
tized images from each animal. For each group, tissue
equivalent sections were identified, corresponding
approximately to anteroposterior �0.62 mm from
bregma for striatum, �0.46 mm for choroids plexus
and ventricular epithelium, �1.46 mm for arcuate
hypothalamic nucleus and �1.82 for hippocampus.
Circles in central region of striatum were placed with
distance of 300 mm each one. Circles in choroids plexus
and ventricular epithelium were placed at random, in
arcuate hypothalamic nucleus circles were placed two to
each side of the third ventricle and other below the third
ventricle. Circles in hippocampus were placed along of
each region.
Electron Microscopy
Five control and five experimental male mice at 24 h
of treatment were used to confirm the ultrastructural
localization of zinc in hippocampus following the
procedure described by Danscher (1981) with modifi-
cations. Animals were anesthetized with ether and
perfused transcardially as described above. After per-
fusion, the brains were removed and placed in the same
fixative for 1 h. The left striatal and hippocampal
formation were isolated and stained using a modifica-
tion of Timm’s sulfide–silver technique as described
above. The tissue was subsequently dehydrated and
finally embedded in Epon. The blocks were polymer-
ized for 16–18 h at a temperature of 60 8C. From each
block, 1-mm semi-thin sections were cut and stained
with Toluidine Blue. This procedure allowed unambig-
uous identification of the area of interest prior to
ultrathin sectioning, which was made from striatum
and stratum radiatum of hippocampal field CA2. From
each block 11–13 serial ultrathin sections (60–80 nm)
were cut with a diamond knife. The sections were
mounted on single-slot grids, which had been coated
with formvar film and contrasted with uranyl acetate
and lead citrate.
Determination of Dopamine
Additional 54 mice were allocated in one of the
eight groups as previously described. Animals were
treated with MPP+ or saline solution and included to
analyze DA content in striatum. Mice were sacrificed
by cervical dislocation at 1, 2, 24 h and 7 days after
MPP+ administration. Mice brains were immediately
removed and corpora striata were dissected out as
described above. DA concentrations were analyzed
according to the method described previously (Rojas
and Rıos, 1993). An aliquot (500 ml) of perchloric
acid–sodium metabisulfite solution (0.1%, w/v) was
added to the weighed tissue and sonicated with a Lab-
line ultratip labsonic system (Lab-line instruments,
Melrose Park, IL). Samples were then centrifuged at
4,000 g for 10 min, and the supernatants were kept at
�70 8C until analyzed.

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968962
24
h7
day
s
Sal
ine
MP
P+
Sal
ine
MP
P+
�0
.00
20
.35
8�
0.0
14
0.3
20�
0.0
04
0.3
37�
0.0
19
0.3
63�
0.0
33
�0
.00
50
.34
5�
0.0
12
0.3
39�
0.0
13
0.3
45�
0.0
09
0.3
81�
0.0
22
�0
.03
60
.46
7�
0.0
23
0.4
92�
0.0
18
0.4
23�
0.0
19
0.4
42�
0.0
27
�0
.01
40
.25
4�
0.0
15
0.2
75�
0.0
03
0.2
25�
0.0
22
0.2
13�
0.0
06
�0
.03
30
.44
8�
0.0
26
0.5
00�
0.0
09
0.4
27�
0.0
26
0.4
36�
0.0
17
�0
.01
40
.24
4�
0.0
11
0.2
69�
0.0
02
0.2
18�
0.0
11
0.2
19�
0.0
20
�0
.02
40
.49
7�
0.0
09
0.5
53�
0.0
31
0.4
94�
0.0
32
0.4
38�
0.0
07
�0
.01
50
.23
8�
0.0
16
0.2
68�
0.0
05
0.2
28�
0.0
09
0.2
14�
0.0
15
�0
.03
60
.59
1�
0.0
58
0.6
96�
0.0
37
0.5
97�
0.0
35
0.5
92�
0.0
27
mm
2o
fd
iam
eter
,ea
cho
ne
for
10
tiss
ue
sect
ion
sp
erre
gio
nan
dan
imal
,n
=6–8;
stat
isti
cal
The striatal content of DA was analyzed using an
HPLC system (LC 250 Perkin-Elmer) with an electro-
chemical detector (Methrom 656) and a Hewlett-Pack-
ard 3396-II integrator. The detector potential was
adjusted to 0.8 V versus Ag/AgCl reference electrode.
The mobile phase consisted of aqueous phosphate buffer
(pH 3.1), which contained 0.2 mM sodium octyl sulfate,
0.1 mM EDTA and 15% (v/v) of methanol. An Alltech
Associates Inc. (Deerfield, IL) adsorbosphere catecho-
lamine analytical column of 100 mm � 4.8 mm with
3 mm particle diameter was used. Calibration curves
were constructed for DA and concentration was
obtained by interpolation of the respective standard
curve. The results were expressed as micrograms of
compound per gram of tissue.
Statistical Analysis
Results were analyzed using two-way analysis of
variance (ANOVA), with Tukey’s test as post hoc.
Values of P < 0.05 or P < 0.01 were considered to
be of statistical significance.
Table
1
Zin
cp
osi
tive
term
inal
sin
dif
fere
nt
bra
inre
gio
ns
of
mo
use
afte
rM
PP
+ad
min
istr
atio
n
Bra
inst
ruct
ure
Su
bre
gio
ns
Rel
ativ
eo
pti
cal
den
sity
,m
ean�
S.E
.M
1h
2h
Sal
ine
MP
P+
Sal
ine
MP
P+
Ven
tric
les
Ch
oro
idp
lex
us
0.3
10�
0.0
14
0.3
33�
0.0
08
0.3
27�
0.0
22
0.2
83
Hy
po
thal
amu
sA
rcu
ate
nu
cleu
s0
.33
1�
0.0
04
0.3
10�
0.0
06
0.3
34�
0.0
09
0.3
41
Hip
po
cam
pu
sC
A1
–st
ratu
mp
yra
mid
ale
0.3
80�
0.0
11
0.4
33�
0.0
28
0.4
85�
0.0
24
0.4
09
CA
1–
stra
tum
mo
lecu
lare
0.2
10�
0.0
18
0.2
18�
0.0
18
0.2
74�
0.0
04
0.2
55
CA
2–
stra
tum
py
ram
idal
e0
.45
3�
0.0
15
0.4
65�
0.0
10
0.4
94�
0.0
22
0.4
31
CA
2–
stra
tum
mo
lecu
lare
0.2
03�
0.0
19
0.2
23�
0.0
20
0.2
53�
0.0
07
0.2
47
CA
3–
stra
tum
py
ram
idal
e0
.52
5�
0.0
06
0.5
25�
0.0
17
0.5
16�
0.0
21
0.4
69
CA
3–
stra
tum
mo
lecu
lare
0.2
17�
0.0
19
0.2
27�
0.0
18
0.2
51�
0.0
08
0.2
44
Den
tate
gy
rus
0.5
56�
0.0
16
0.5
29�
0.0
14
0.5
89�
0.0
32
0.5
71
Reg
ion
sw
ere
loca
lize
du
nd
erb
rig
ht
fiel
dli
gh
t-m
icro
sco
py,
and
rela
tive
op
tica
ld
ensi
tyw
aso
bta
ined
infi
ve
circ
les
of
40
anal
ysi
so
ftw
o-w
ayA
NO
VA
;n
och
ang
eso
bse
rved
.
RESULTS
Zinc-rich terminal fields were observed in various
telencephalic areas: striatum, hippocampus, hypotha-
lamus and ventricles (Fig. 1; Table 1). The highest
zinc-rich terminal field in control animals was recorded
in hippocampus (black > dark brown), such as dentate
gyrus > CA3–field pyramidale > CA2–field pyrami-
dale > CA1–field pyramidale. The lowest zinc-rich
terminal field in control animals was seen in some
layers of hippocampus (light brown), such as CA2–
field moleculare > CA1–field moleculare and stria-
tum.
Examination at high magnification in the light
microscope showed that the precipitate was virtually
exclusively in the neuropil (presumably in axonal
boutons). Electron microscopic examination showed
that silver grains were almost always in boutons as seen
in control and experimental group (Fig. 1E and F), as
has been reported before (Friedman and Price, 1984).
Striatum
Statistical two-way ANOVA test revealed signifi-
cant effects for the treatment in the striatum
(F = 99.32, P < 0.01). The measured mean density
of zinc sulfide–silver precipitate in the Timm staining
was significantly lower in the striatum after MPP+

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968 963
Fig. 1. Representative microscopic images of striatum (A and B) and hippocampus (C and D) after MPP+ neurotoxicity showing loss or enhancement of zinc-
positive Timm-stained terminal fields, magnification 4�: (A) frontal section of the striatum demonstrating intense Timm’s stain for vesicular zinc in the control
group and loss in MPP+ group at 2 h (B) compared to that in control mice; (C) frontal section of the hippocampus demonstrating intense Timm’s stain for
vesicular zinc in the control group and MPP+ at 7 days (D). MPP+ administration showed increased of zinc staining in stratum oriens and stratum radiatum of
CA1 and CA2. The reaction was heavier in stratum radiatum of CA3 after MPP+ neurotoxicity. Sections were counterstained with cresyl violet or Toluidine Blue
and the color of the images correspond to the intensity of zinc staining. The lowest zinc-rich terminal field is shown by light brown color and highest zinc-rich
terminal field is represented by black > dark brown color. Results were repeated with five independent experiments. Electron micrographs demonstrating
axonal boutons of the hippocampus at 24 h of control group (E) and hippocampus at 24 h after MPP+ administration (F) of neurons containing zinc precipitated
as zinc sulfide in the Timm staining. Compare the heavier distribution of silver grain in MPP+ group. Magnification (E and F) 21,600�. MPP+, 1-methyl-4-
phenylpyridinium ion; CA1, CA1 field of the hippocampus; CA2, CA2 field of the hippocampus; CA3, CA3 field of the hippocampus; DG, dentate gyrus; Rad,
stratum radiatum; Or, stratum oriens; Py, stratum pyramidale; Mol, stratum moleculare. Ax, axonal boutons.

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968964
Fig. 2. Decrease on relative optical density of zinc-positive Timm-stained
terminal fields in striatum (A) and ventricular epithelium (B). Results are
expressed as mean � S.E.M. of n = 6–8 independent experiments: (A)
**statistically significant effect of treatment (F = 99.32, P < 0.01), two-
way ANOVA followed by Tukey’s test; (B) **statistically significant effect
of treatment (F = 147.74, P < 0.01), two-way ANOVA followed by Tukey’s
test. MPP+, 1-methyl-4-phenylpyridinium ion.
administration at 1 h (Fig. 2A). This decrease was
significant in zinc-positive terminal staining continued
through all times examined up to 24 h (Figs. 1A, 1B
and 2A) after MPP+ administration. The largest
decrease was at 2 h (24%), followed by 1 h (18%)
and 24 h (14%) after MPP+ neurotoxicity when com-
pared with control groups.
Ventricles
The two-way ANOVA test showed an important
significant effect for the treatment in ventricular
epithelium (F = 147.74, P < 0.01), as well as the
interaction between treatment and time (F = 8.61,
P < 0.01). Staining in ventricular epithelium shows
a drastic decrease at 2 h (44%) after MPP+ admin-
istration (Fig. 2B). At 1 h, 24 h and 7 days the
decrease was 21, 29 and 15%, respectively, after
MPP+ administration compared to respective control
groups.
In the choroid plexus, statistical two-way ANOVA
test revealed no changes in staining after MPP+ neu-
rotoxicity versus control group (Table 1).
Hypothalamus
In the hypothalamus, no changes were observed in
arcuate nucleus after MPP+ neurotoxicity versus con-
trol group (Table 1; two-way ANOVA test).
Hippocampus
Density changes were found after Timm staining in
the hippocampus in two strata (oriens and radiatum) of
the hippocampus (CA1, CA2 and CA3; Figs. 1C, 1D
and 3).
The two-way ANOVA statistical analysis revealed a
significant effect for the treatment in CA1 stratum
oriens (F = 12.48, P < 0.01) and the interaction
between treatment and time (F = 7.22, P < 0.01). Zinc
staining in CA1 stratum oriens after MPP+ adminis-
tration enhanced at 24 h (50%) and 7 days (41%) when
compared to control group (Fig. 3A).
The two-way ANOVA statistical analysis revealed a
significant effect for the treatment in CA1 stratum
radiatum (F = 11.72, P < 0.01), as well as for the
interaction between treatment and time (F = 5.77,
P < 0.01). The reaction in CA1 stratum radiatum after
MPP+ administration shows an enhancement at 24 h
(39%) and 7 days (47%) as compared to control values
(Fig. 3B).
Statistical two-way ANOVA test shows effect for the
treatment in CA2 stratum oriens (F = 8.14, P < 0.01),
as well as for the interaction treatment and time
(F = 3.98, P < 0.01). In CA2, the stratum oriens shows
an increase at 7 days (25%) after MPP+ administration
versus control group (Fig. 3C).
Statistical analysis showed a clear effect for the
treatment in CA2 stratum radiatum (F = 12.03,
P < 0.01) as well as the interaction treatment and time
(F = 4.55, P < 0.01). The stratum radiatum in CA2
presented higher staining at 24 h (41%) and 7 days
(42%) as compared to control group (Fig. 3D).
In the stratum oriens lighter staining was observed in
CA3 but not significant changes versus control group
(Fig. 3E).
The two-way ANOVA showed an important effect in
the treatment in CA3 stratum radiatum (F = 5.80,
P < 0.01) and the interaction between treatment and
time (F = 4.11, P < 0.01). In the stratum radiatum, the
reaction was heavier in CA3 at 7 days (39%) after
MPP+ neurotoxicity versus control group (Fig. 3F).
Administration of MPP+ did not produce significative
changes in dentate gyrus, CA1 (strata pyramidale and
moleculare), CA2 (strata pyramidale and moleculare)
and CA3 (strata pyramidale and moleculare).
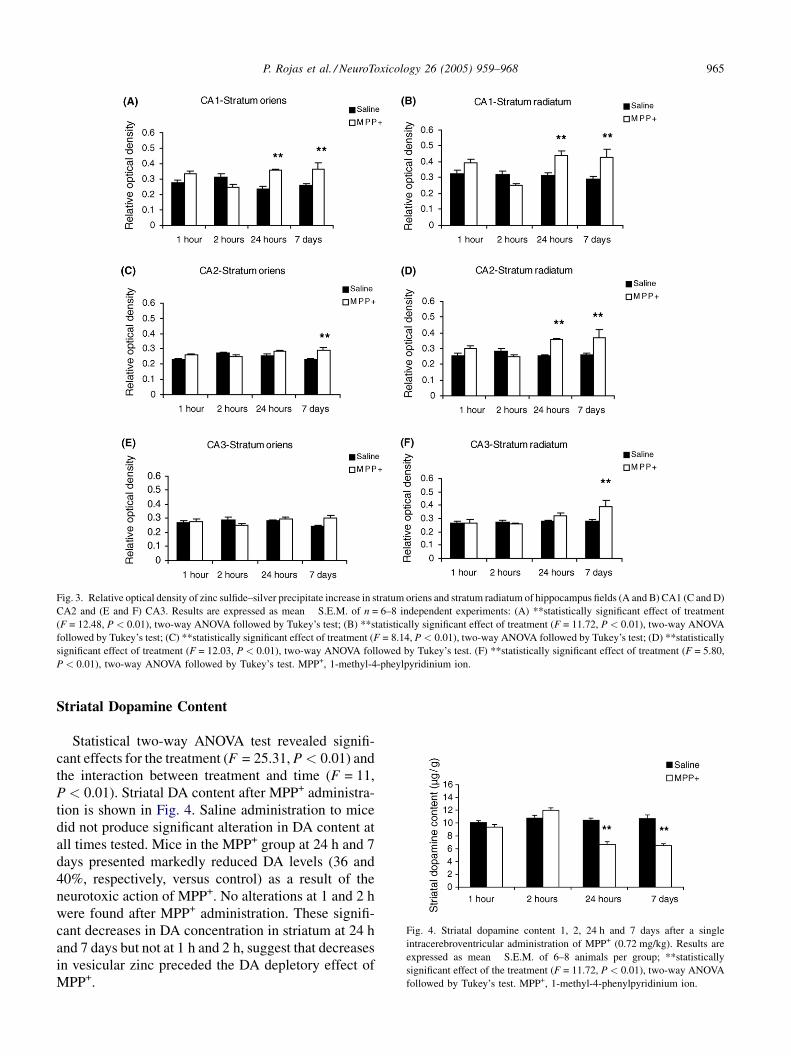
P. Rojas et al. / NeuroToxicology 26 (2005) 959–968 965
Fig. 3. Relative optical density of zinc sulfide–silver precipitate increase in stratum oriens and stratum radiatum of hippocampus fields (A and B) CA1 (C and D)
CA2 and (E and F) CA3. Results are expressed as mean � S.E.M. of n = 6–8 independent experiments: (A) **statistically significant effect of treatment
(F = 12.48, P < 0.01), two-way ANOVA followed by Tukey’s test; (B) **statistically significant effect of treatment (F = 11.72, P < 0.01), two-way ANOVA
followed by Tukey’s test; (C) **statistically significant effect of treatment (F = 8.14, P < 0.01), two-way ANOVA followed by Tukey’s test; (D) **statistically
significant effect of treatment (F = 12.03, P < 0.01), two-way ANOVA followed by Tukey’s test. (F) **statistically significant effect of treatment (F = 5.80,
P < 0.01), two-way ANOVA followed by Tukey’s test. MPP+, 1-methyl-4-pheylpyridinium ion.
Fig. 4. Striatal dopamine content 1, 2, 24 h and 7 days after a single
intracerebroventricular administration of MPP+ (0.72 mg/kg). Results are
expressed as mean � S.E.M. of 6–8 animals per group; **statistically
significant effect of the treatment (F = 11.72, P < 0.01), two-way ANOVA
followed by Tukey’s test. MPP+, 1-methyl-4-phenylpyridinium ion.
Striatal Dopamine Content
Statistical two-way ANOVA test revealed signifi-
cant effects for the treatment (F = 25.31, P < 0.01) and
the interaction between treatment and time (F = 11,
P < 0.01). Striatal DA content after MPP+ administra-
tion is shown in Fig. 4. Saline administration to mice
did not produce significant alteration in DA content at
all times tested. Mice in the MPP+ group at 24 h and 7
days presented markedly reduced DA levels (36 and
40%, respectively, versus control) as a result of the
neurotoxic action of MPP+. No alterations at 1 and 2 h
were found after MPP+ administration. These signifi-
cant decreases in DA concentration in striatum at 24 h
and 7 days but not at 1 h and 2 h, suggest that decreases
in vesicular zinc preceded the DA depletory effect of
MPP+.

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968966
DISCUSSION
Approximately 90% of the brain’s total zinc is in
metalloproteins. The rest is in the presynaptic vesicles
and histochemically reactive as revealed by Timm’s
sulfide–silver staining method (Frederickson, 1989).
Vesicular zinc, probably ionic zinc, may play a role in
synaptic neurotransmission in the mammalian brain
including the DA pump (for a review, see Frederickson
and Mocrieff, 1994). Zinc is transported into the brain
via not only the blood brain barrier but also the blood–
CSF barrier.
Zinc is taken up by neurons, which may have two
zinc uptake sites, i.e. the soma and the neuron terminal
and also by glia cells, and it is then incorporated into
zinc-binding proteins.
We undertook the present study to examine whether
the levels of Timm stainable vesicular zinc in neuronal
terminals would decrease in response to MPP+ neuro-
toxicity. The intensity of the staining reflects the
number of zinc-rich boutons in a terminal field or
the intensity of labeling enclosed in these boutons as
reported previously (Martınez-Guijarro et al., 1987).
Our results show reduction of zinc-positive terminals in
striatum following MPP+ administration. This occurs
within 1 h after MPP+ administration and continues up
to 24 h after MPP+ administration. Our electron micro-
scopy studies show that the reaction for zinc is confined
within synaptic boutons as reported previously (Fried-
man and Price, 1984). Therefore, the terminal field
staining observed in our results was considered to be
located in synaptic boutons.
Our results showed a loss of zinc-positive terminal
fields in striatum and decreases in DA levels after
MPP+ administration. This suggests that loss of zinc
is associated with the occurrence of DA decrease in
striatum. The decrease in the density of Timm stainable
terminal zinc found in the striatum, most probably
represents a release of vesicular zinc from the synaptic
terminals by MPP+ neurotoxicity. This free zinc
induced by MPP+ could avoid DA reuptake and might
produce reduction in DA release in synaptic cleft and
its production. These results suggest that this alteration
in zinc could be related to the DAergic neurotoxicity of
MPP+. Our results showed a loss of zinc-positive
terminal fields in striatum at 1–24 h after MPP+ admin-
istration and also showed decreases in DA levels but
starting at 24 h. This difference could be because MPP+
produces its action showing reduction in zinc-rich
boutons in striatum starting at 1 h and this effect is
not affecting the synaptic neurotransmission and the
DA production at this time analyzed. The reduction of
zinc and DA content occur at 24 h and 7 days when the
DAergic neurotoxin produces cell death and /or reduc-
tion in DA production after MPP+ injection (for a
review, see Gerlach et al., 1991).
It has been shown that zinc and MPP+ inhibits
uptake of DA (Richfield, 1993; Scholze et al.,
2002), an effect that is accounted for by direct binding
of zinc to human DA transporter (Norregaard et al.,
1998). Thus, there is some regulation of DA transport
by zinc. It has been suggested that the release of
glutamate depolarizes the dendrites, resulting in trans-
port reversal and subsequent release of DA non-exo-
cytotically from the dendritic region (Falkenburger
et al., 2001). Thus, when zinc is co-released with
glutamate, it may greatly augment the efflux of DA.
The enhancement of zinc-positive terminal fields
found in oriens and radiatum strata of hippocampus, is
thought to be due to uptake of zinc released by
surrounding zinc-positive terminals. This neuronal
uptake of zinc in hippocampus could represent an
attempt to buffer the concentration increase in extra-
cellular zinc that presumably takes place in relation to
the MPP+ neurotoxicity in the striatum. Another alter-
native explanation is that the neuronal zinc represents a
pathologic influx of zinc into the hippocampus, as
neuronal accumulation of zinc has been shown to
precede neurodegeneration (Koh et al., 1996).
Staining reduction was observed in ventricular
epithelium. This decrease in staining may reflect the
reduction of the passage of zinc through the blood-
brain barrier, the main route of entry for zinc into the
brain (Takeda, 2000). The mechanism of zinc secretion
from the brain capillary endothelial cells into the brain
extracellular fluid and the CSF, respectively, is
unknown.
Zinc is an important metal, because it is necessary
for the formation of a vast associational network of
neurons, while a portion of zinc is stored in the synaptic
vesicles in zinc-containing neuron terminals in the
telencephalon (Frederickson and Mocrieff, 1994).
Zinc is known to be accumulated dramatically and
selectively in the region of substantia nigra of the brain,
which has often been related to the pathogenesis of PD
(Forsleff et al., 1999). Regarding zinc, antioxidant
properties in relation to brain oxidative stress have
been reported (Ebadi et al., 1996; Tate et al., 1999). It
has been suggested that zinc does not promote the
formation of free radicals since zinc is not redox active
(Berg and Shi, 1996). Although it has been suggested
that these antioxidant properties may be related to the
capacity shown by zinc to induce the synthesis of
metallothionein, a stable endogenous cysteine-rich

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968 967
protein that may participate in scavenging free radicals
(Ebadi et al., 1996), the precise mechanism of this
effect is still unclear. Also, the mobilization of zinc
from metallothionein under conditions of oxidative
stress (Maret, 1995) can also happen in MPP+ neuro-
toxicity (Rojas et al., 1996). However, it must be kept
in mind that synaptic zinc is just a small fraction of the
total zinc pool in the brain (for a review, see Ebadi
et al., 1996).
Our findings show that the decrease of zinc-positive
terminal fields in the striatum after MPP+ administra-
tion is most likely due to a neuronal release of vesicular
zinc in response to MPP+ neurotoxicity.
ACKNOWLEDGEMENTS
The authors thank Dr. Sultan Habeebu for his com-
ments. Partially supported by the National Council of
Science and Technology of Mexico (CONACyT)
28605-M Grant.
REFERENCES
Adams JD Jr, Odunze IN. Biochemical mechanisms of 1-methyl-4-
phenyl-1,2,3,6-biochemical mechanisms of 1-methyl-4-phenyl-
1,2,3,6-tetrahydropyridine toxicity. Could oxidative stress be
involved in the brain?. Biochem Pharmacol 1991;41:1099–105.
Adams JD, Klaidman LK, Leung A. MPP+ and MPDP+ induced
oxygen radical formation with mitochondrial enzymes. Free
Radic Biol Med 1993;15:181–6.
Alcaraz-Zubeldia M, Rojas P, Boll C, Rıos C. Neuroprotective
effect of acute and chronic administration of copper(II) sulfate
against MPP+ neurotoxicity in mice. Neurochem Res
2001;26:59–64.
Berg JM, Shi Y. The galvanization of biology: a growing apprecia-
tion for the role of zinc. Science 1996;271:1081–5.
Chiba K, Trevor A, Castagnoli N. Metabolism of the neurotoxic
tertiary amine MPTP, by brain monoamine oxidase. Biochem
Biophys Res Commun 1984;120:574–8.
Cintra L, Granados L, Aguilar A, Kemper T, DeBassio W, Galler J,
et al. Effects of prenatal protein malnutrition on mossy fibers of
the hippocampal formation in rats of tour age groups. Hippo-
campus 1997;7:184–91.
Danscher G. Histochemical demonstration of heavy metals. His-
tochemistry 1981;71:1–16.
Ebadi M, Leuschen MP, El Refaey H, Hamada FM, Rojas P. The
antioxidant properties of zinc and metallothionein. Neurochem
Int 1996;29:159–66.
Falkenburger BH, Barstow KL, Mintz IM. Dendrodendritic inhibi-
tion through reversal of dopamine transport. Science 2001;293:
2465–70.
Forsleff L, Schauss AG, Bier ID, Stuart S. Evidence of functional
zinc deficiency in Parkinson’s disease. J Altern Complem Med
1999;5:57–64.
Frederickson CJ. Neurobiology of zinc and zinc-containing neu-
rons. Int Rev Neurobiol 1989;31:145–238.
Frederickson CJ, Danscher G. Zinc-containing neurons in hippo-
campus and related CNS structures. Prog Brain Res
1990;83:71–84.
Frederickson CJ, Mocrieff DW. Zinc-containing neurons. Biol
Signals 1994;3:127–39.
Friedman B, Price JL. Fiber systems in the olfactory bulb and
cortex: a study in adult and developing rats, using Timm method
with the light and electron microscope. J Comp Neurol
1984;223:88–109.
Gerlach M, Riederer P, Przuntek H, Youdim MBH. MPTP mechan-
isms of neurotoxicity and their implications for Parkinson’s
disease. Eur J Pharmacol Mol Pharmacol 1991;208:273–86.
Jenner P. Clues to the mechanism underlying dopamine cell death
in Parkinson’s disease. J Neurol Neurosurg Psychiatry 1989;22–
8 [Special Suppl.].
Kienzl E, Puchinger L, Jellinger K, Linert W, Stachelberger H,
Jameson RF. The role of transition metals in the pathogenesis of
Parkinson’s disease. J Neurol Sci 1995;134:69–78.
Koh JY, Suh SW, Gwag BJ, He YY, Hsu CY, Choi DW. The role of
zinc in selective neuronal death after transient global cerebral
ischemia. Science 1996;272:1013–6.
Maret W. Metallothionein/disulfide interactions, oxidative stress,
and the mobilization of cellular zinc. Neurochem Int
1995;27:111–7.
Martınez-Guijarro FJ, Molowny A, Lopez Garcıa C. Timm-stain-
ing intensity is correlated with the density of Timm-positive
presynaptic structures in the cerebral cortex of lizards. Histo-
chemistry 1987;86:315–9.
Mihatsch W, Russ H, Przuntek H. Intracerebroventricular adminis-
tration of MPP+ in mice: effects of simultaneously administered
nomifensine, deprenyl, and 1-butyl-4, 4-diphenylpiperidine. J
Neural Transm 1988;71:177–88.
Norregaard L, Frederiksen D, Nielsen EO, Gether U. Delineation of
an endogenous zinc-binding site in the human dopamine trans-
porter. EMBO J 1998;17:4266–73.
Owen AD, Schapira AHV. Jenner P, Marsden CD. Oxidative stress
and Parkinson’s disease. Ann NY Acad Sci 1996;786:217–23.
Palmiter RD, Cole TB, Quaife CJ, Findley SD. ZnT-3, a putative
transporter of zinc into vesicles. Proc Natl Acad Sci USA
1996;93(25):1434–9.
Perez-Clausell J, Danscher G. Intravesicular localization of zinc in
rat telencephalic boutons: a histochemical study. Brain Res
1985;337:91–8.
Richfield EK. Zinc modulation of drug binding, cocaine affinity
states, and dopamine uptake on the dopamine uptake complex.
Mol Pharmacol 1993;43:100–8.
Rıos C, Alvarez-Vega R, Rojas P. Depletion of copper and man-
ganese in brain after MPTP treatment of mice. Pharmacol
Toxicol 1995;76:348–52.
Rojas P, Rıos C. Increased striatal lipid peroxidation after intracer-
ebroventricular MPP+ administration to mice. Pharmacol Tox-
icol 1993;72:364–8.
Rojas P, Rıos C. Short-term manganese pretreatment partially
protects against 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine
neurotoxicity. Neurochem Res 1995;20(10):1217–23.
Rojas P, Cerutis DR, Happe HK, Hao R, Murrin LC, Pfeiffer RF, et
al. 6-Hydroxydopamine mediated induction of rat brain
metallothionein I mRNA. Neurotoxicology 1996;17(2):
323–34.

P. Rojas et al. / NeuroToxicology 26 (2005) 959–968968
Scholze P, Nørregaard L, Singer E, Freissmuth M, Gether U, Sittle
HH. The role of zinc ions in reverse transport mediated by
monoamine transporters. J Biol Chem 2002;277(24):21505–13.
Takeda A. Movement of zinc and its functional significance in the
brain. Brain Res Rev 2000;34:137–48.
Tate DJ, Miceli M, Newsome DA. Zinc protects against oxidative
damage in cultured human retinal pigment epithelial cells. Free
Radic Biol Med 1999;26:704–13.
Vallee VL, Falchuk KH. The biochemical basis of zinc physiology.
Physiol Rev 1993;73:79–117.
Walsh CT, Sandstead HH, Prasad AS, Newberne PM, Fraker PJ.
Zinc: health effects and research priorities. Environ Health
Perspect 1994;102(Suppl. 2):5–46.
West JR, Hodges-Savola CA. Permanent hippocampal mossy fiber
hyperdevelopment following prenatal ethanol exposure. Neu-
robehav Toxicol Teratol 1983;5:139–50.
West JR, Dewey SL. Mossy fiber sprouting in the fascia dentate
after unilateral entorhinal lesions: quantitative analysis using
computer-assisted image processing. Neuroscience 1984;13:
337–84.
Copyright © 2022 FDOKUMEN




















