
Approach for Simulating Acidification and the Carbon Cycle in ...
Upload
independentCategory
view
0download
0
at SciVerse ScienceDirect
Estuarine, Coastal and Shelf Science 124 (2013) 34e43
Contents lists available
Estuarine, Coastal and Shelf Science
journal homepage: www.elsevier .com/locate/ecss
Reduced marine survival of hatchery-reared Atlantic salmon post-smolts exposedto aluminium and moderate acidification in freshwater
Eva B. Thorstad a,*, Ingebrigt Uglem a, Bengt Finstad a, Frode Kroglund b, Ingibjörg Eir Einarsdottir c,Torstein Kristensen d, Ola Diserud a, Pablo Arechavala-Lopez e, Ian Mayer f, Andy Moore g, Rune Nilsen h,Björn Thrandur Björnsson c, Finn Økland a
aNorwegian Institute for Nature Research (NINA), P.O. Box 5685 Sluppen, NO-7485 Trondheim, NorwaybNorwegian Institute for Water Research (NIVA), Televeien 3, NO-4879 Grimstad, Norwayc Fish Endocrinology Laboratory, Department of Biological and Environmental Sciences, University of Gothenburg, Box 463, S-40530 Gothenburg, SwedendNorwegian Institute for Water Research (NIVA), NMU Bygget, Mørkvedbukta, NO-8020 Bodø, NorwayeDepartment of Marine Sciences and Applied Biology, University of Alicante, PO Box 99, 03080 Alicante, SpainfNorwegian School of Veterinary Science, P.O. Box 8146 Dep, NO-0033 Oslo, NorwaygCentre for Environment, Fisheries and Aquaculture Science, Pakefield Road, Lowestoft, Suffolk NR33 0HT, United Kingdomh Institute of Marine Research (IMR), P.O. Box 6404, NO-9294 Tromsø, Norway
a r t i c l e i n f o
Article history:Received 3 June 2012Accepted 23 March 2013Available online 3 April 2013
Keywords:Salmo salaracidificationacid precipitationaluminiumsmolt and post-smolt migrationtelemetry
Regional index terms:EuropeNorwayMøre and Romsdal CountyRomsdalsfjord
* Corresponding author.E-mail address: [email protected] (E.B. Thorst
0272-7714/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.ecss.2013.03.021
a b s t r a c t
Short-term Al-exposure and moderate acidification increased initial marine mortality in migrating post-smolts, and can thereby reduce viability of Atlantic salmon stocks. The delayed impact of short-termaluminium (Al) exposure on hatchery-reared Atlantic salmon smolt in moderately acidified freshwater(pH 5.88e5.98) was investigated during the first 37 km of the marine migration. Smolts were taggedwith acoustic tags and exposed to low (28.3 � 4.6 mg l�1 labile Al, 90 h) or high (48.5 � 6.4 mg l�1 labile Al,90 or 48 h) Al concentrations within the hatchery. Thereafter their movements, together with a controlgroup, were monitored throughout the marine fjord. Al-exposure resulted in increased gill-Al andcompromised hypoosmoregulatory capacity, as shown by elevated mortality in laboratory seawaterchallenge tests and reduced Naþ, Kþ-ATPase activity levels. Further, Al-exposure resulted in decreasedplasma concentrations of growth hormone (GH), while the insulin-like growth factor (IGF-I) was unaf-fected. There was a significant mortality in the 90 h high-Al group during exposure, and those survivinguntil release died during the first 3.6 km of the marine migration. Physiological stress and mortality werenot only a result of the Al-concentrations, but also dependent on exposure duration, as shown by resultsfrom the 48 h high-Al group. Elevated mortality was not recorded in freshwater or after entering the seafor this group, which highly contrasts to the 100% mortality in the 90 h high-Al group, despite bothgroups having similarly high gill-Al levels. The low-Al group showed a 20% higher mortality compared tothe control group during the first 10 km of the marine migration, but during the next 28 km, mortalityrates did not differ. Hence, post-smolts surviving the first 10 km subsequently showed no differences inmortality compared to controls. At least one third of the mortality in both the low-Al and control groupswere due to predation by marine fishes, indicating that the proximate cause for elevated mortality due toAl-exposure may have been predation. Migration speeds over 3.6, 9.6 or 37.1 km from the release site wasnot affected by Al-exposure.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The Atlantic salmon is primarily an anadromous species withspawning and juvenile development taking place in freshwater,
ad).
All rights reserved.
followed by a rapid growing phase in seawater, after which thesexually maturing adults migrate back to their native spawninggrounds (Thorstad et al., 2011a). Anadromy is seen as an adaptivestrategy, with individuals using the best suited habitat duringdifferent stages of the life cycle to increase fitness (Lucas and Baras,2001). However, the transition from freshwater to marine habitatsis a challenging phase that involves high mortalities (20e65%mortality in estuaries and fjords; Hvidsten and Møkkelgjerd, 1987;

E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e43 35
Thorstad et al., 2007; Davidsen et al., 2009). Preparatory morpho-logical, biochemical, physiological and behavioural changes, whichtake place during parr-smolt transformation (smoltification), arenecessary to preadapt the fish for life in the saline marine envi-ronment, which involves both long migrations and exposure tonovel prey and predator species (McCormick et al., 1998). Themarine mortality is mainly driven by density-independent factors,and the number of outward migrating post-smolts is positivelycorrelated with the number of returning spawners of a cohort(Jonsson et al., 1998; Milner et al., 2003). Hence, factors affectingmortality during the smolt and post-smolt stages will contribute indetermining population abundance (Milner et al., 2003). With thecontinued decline in wild salmon populations (ICES, 2011), it isimperative that salmon smolts exhibit high survival rates duringtheir seaward migration.
The smoltification processmay be adversely affected by a varietyof anthropogenic factors, including acidification and pollutants(McCormick et al., 1998; Monette and McCormick, 2008). Acidifi-cation has caused the loss or reduction of numerous Atlanticsalmon populations on both sides of the North Atlantic (e.g.Hesthagen and Hansen, 1991; Watt et al., 2000), and many riversare still severely affected by chronic or episodic acid water(Kroglund et al., 2008; McCormick et al., 2009). Increased mortalitydue to acidification may be related to both high levels of Hþ
(reduced pH) and aluminium (Al). During acidification, Al ismobilised from the soil, leading to elevated concentrations in thewater (Teien et al., 2004b). The solubility of Al increases withdecreasing pH, causing increased concentrations of inorganicmonomeric Al (Ali), which is the most toxic form to fish (Gensemerand Playle, 1999). As such, apart from when acidification is severeenough to be directly toxic, toxicity may also occur due to Aliaccumulating on the gill tissue, which may disrupt ion regulationand impair respiration (Rosseland and Staurnes, 1994; Gensemerand Playle, 1999). Loss of seawater tolerance in smolts during acidand Al exposure may be mediated, in part, by negative impacts onthe GH-IGF-I system (Monette et al., 2008).
As long as the smolts remain in freshwater, moderate Al-exposure due to acidification may not be detrimental to the fish.However, as elevated gill-Al content can cause reduced hypo-osmoregulatory ability, sub-lethal exposure effects in freshwatermay become lethal in seawater, causing increased marine mortality(Staurnes et al., 1993, 1996; Kroglund and Finstad, 2003; Kroglundet al., 2007). Such cross-over effects where the stressor is applied inone environment but the effect does not emerge until the fish hasentered the new environment, are difficult to demonstrate in na-ture because contamination levels and mortality of individual fishare difficult to follow over time and through different environ-ments. Previous studies have mainly been based on laboratoryexperiments coupled to limited tagging-recapture data.
This study aimed to use recent advances in telemetry techniquesto test the hypothesis that the survival and migratory behaviour ofacid and aluminium impacted Atlantic salmon smolts (Salmo salar)differ during their migration into the marine environment. Thedelayed impact of Al-exposure in moderately acidified freshwaterwas investigated during the first 37 km of the marinemigration in acombined field and laboratory study. Hatchery-reared smolts weretagged with acoustic tags and exposed to zero (control group), lowor high Al concentrations within the hatchery. After release,movements of the fish were monitored throughout a fjord usingstationary acoustic receiver arrays. The physiological impact of theexposure on smolts (separate sub-groups kept under controlledexperimental conditions in the hatchery) was determined byassessing seawater tolerance, branchial Al concentrations and Naþ,Kþ-ATPase activity, as well as plasma levels of chloride, growthhormone (GH) and insulin-like growth factor (IGF-I).
2. Materials and methods
2.1. Study area
The study was performed in the laboratory of the Statkraft En-ergy AS hatchery situated at the outlet of the River Eira, and in theRomsdalsfjord system in Norway, 62�400N 8�100E (Fig. 1, Finstadet al., 2005). The area is not affected by acidification (Anon, 2011).The River Eira is a soft and clear water river, with a pH 6.7, 1e2 mg Ca l�1, total Al <20 mg l�1, Acid neutralising capacity (ANC)>50 meq l�1, low ion concentration, low conductivity(z2.4 mS m�1) and a low total organic content (<0.6 mg C l�1)(Table 1).
2.2. Salinity and temperature in the fjord
Salinity and temperature were recorded from the surface to30 m depth on 6, 8, 11 and 14 May 2009 at all receiver positions atsite 1, and at three positions at each of sites 2, 3 and 4 (Valeport Ltd.miniCTD, Devon, UK, Fig. 1). In the inner fjord (sites 1e3), there wasa brackish surface layer, with salinity increasing with depth. At0.4 m depth, the average salinity was 21.3 (range 4.1e28.6), withthe lowest salinities close to the river mouth. At 1.0 m depth,average salinity was 26.9 (range 22.8e28.6). A salinity of 30 wasreached at average 8.6 m depth (range 6.0e11.3). In the outer fjord(site 4), salinities were never below 30.5 at any depth. Watertemperature at 0e3.0 m depth, where smolts usually live (Hedgeret al., 2011; Thorstad et al., 2011b), varied between 6.4 and11.4 �C at all sites.
2.3. Fish and acoustic transmitter tagging
First generation hatchery-reared Atlantic salmon of the localRiver Eira stock were used. On 7 April 2009, pre-smolts (age 2þ)were transferred from the Statkraft Energy AS hatchery to anexperimental facility at the same location, where they were housedin tanks supplied with flow-through river water. On 14e15 April,135 pre-smolts were randomly selected and surgically implantedwith individually coded acoustic transmitters. Transmitters usedwere made by VEMCO Ltd., Canada, model V9P-1L-69KHz-S256(dimensions 9 � 39 mm; weight in air/water of 5.2/2.7 g), havinga depth sensor range of 50 m. The fish were anaesthetised (2-phenoxy-ethanol EC No 204-589-7, SIGMA Chemical Co., USA,0.5e1.0 ml l�1 water) and transmitters were surgically implanted inthe body cavity according to methods described by Finstad et al.(2005). Mean body mass of the smolts was 167 g (range 97e283)and mean total length 259 mm (range 213e314). Body size did notdiffer among the treatment groups (one-way ANOVA, mass:F2,132 ¼ 1.09, P ¼ 0.34, length: F2,132 ¼ 1.03, P ¼ 0.36).
2.4. Acid/aluminium exposure
Tagged fish were randomly divided into three experimentalgroups: a control group, a group exposed to moderately acidicwater and low Al levels, and a group exposed to moderately acidicwater and high Al levels (45 fish in each group). Full details of thewater quality characteristics for the three experimental groups, aswell as the background River Eira water are given in Table 1. Each ofthe three experimental groupswas divided into three replicate sub-groups (n ¼ 15), which were released on 5, 7 and 9 May 2009,referred to as release 1, 2 and 3, respectively. This sequential releaseover five days was carried out to reduce the risk of acoustic tagsignal collisions and detection loss caused by several fish simulta-neously passing receivers. Prior to release, each sub-group wastransferred to exposure tanks (details below), acclimated there for

Fig. 1. Map of the study area showing the smolt release site in the mouth of the River Eira and the receiver sites in the fjord. Individual receivers at site 1 in the inner fjord areindicated by C. There was also a receiver at the release site (+). At site 2e4, several receivers were placed in a line across the fjord.
E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e4336
24 h before being exposed to control, low-Al or high-Al conditionsfor 90 h. Due to highmortality observed during exposure in the firsttwo high-Al groups (Table 2), the exposure time for the last high-Algroup was reduced from 90 to 48 h (referred to as high-Al, shortexposure). The fish were fed with commercial fish feed until theywere moved to the exposure tanks, but were not fed duringexposure.
Table 1Water chemistry characteristics of River Eira (May 2009), and the fish exposuretanks (control, low-Al and high-Al) during the study. (Data are given as meanvalues � standard deviation). Water quality in the control tank are for untreatedwater in the laboratory, hence they are representative of the rearing tank where fishwere held prior to experiments and the header tank before addition of chemicals tothe low-Al and high-Al exposures. e indicates that the variable was not measured.
River Eira Control Low-Al High-Al
pH 6.7 � 0 6.00e6.01 5.95e5.98 5.88e5.94Temperature (�C) e 2.9e3.3 2.9e3.3 2.9e3.3Conductivity
(mS/cm)2.4 � 0.2 2 � 0 2.1 � 0.2 2.2 � 0.2
Ca (mg/l) 1.8 � 0.4 1.2 � 0 e e
Mg (mg/l) 0.3 � 0 0.3 � 0 e e
Na (mg/l) 1.7 � 0.1 1.9 � 0.1 e e
K (mg/l) 0.4 � 0.1 0.3 � 0 e e
SO4 (mg/l) 2.5 � 0.6 1.5 � 0 e e
Cl (mg/l) 2.6 � 0.2 3 � 0.1 3.2 � 0.3 3.4 � 0.4NO3-N (mg/l) 119 � 18.8 71 � 48.1 e e
TOC (mg/l) 0.5 � 0.1 0.3 � 0.1 e e
Al/ICP (mg/l) 10.5 � 6 6.8 � 1.7 36.1 � 7.5 65.9 � 11Al/c (mg/l) 3.5 � 1 1 � 0.8 5.3 � 1 8 � 1.4Al/R (mg/l) 10.5 � 4.9 6.3 � 1 33 � 4.4 52.5 � 6.4Al/Il (mg/l) <5 <5 4.8 � 0.5 4 � 0LAl (mg/l) 2.8 � 4.3 2 � 0 28.3 � 4.6 48.5 � 6.4Fe (mg/l) 0 � 0 0 � 0 e e
Cu (mg/l) e <2 e e
ZN (mg/l) e <2 e e
ANC (meq/l) 66.7 � 9.5 50 � 5.8 44.4 � 5.8 38.8 � 5.8
2.5. Exposure tanks and water chemistry
Flow was adjusted to 23 L/min from a header tank with un-treated water into ageing tanks. Before entering the ageing tanks,aluminium (Al) and acid (HCl) was delivered to the flow at a rate of3.1 ml/min from a stock solution of 41 g AlCl3 and 60 ml HCl (low-Al), or 82 g AlCl3 and 125 ml HCl (high-Al) dissolved in 25 L water.Stock solution and water was mixed using an air pump. Nominally,this should increase total Al by 25 mg/l in low-Al exposures, and by50 mg/l in high-Al exposures. The addition of HCL was intended togenerate a target pH of between 5.7 and 5.8. Stock solution wasdelivered using a Watson Marlow pump 505S fitted with greenegreen MARPRENE tubing and running at 12 RPM. Based on reten-tion time, Al was aged for about 1 h. The exposure tanks wereoctahedral with a maximum width of 1.5 � 1.5 m, surface area of2 m2, and water depth of 23e25 cm, giving the fish a water expo-sure volume of 460e510 L. Surplus Al-adjusted water was led to anoverflow. Ageing of water prior to exposure is required to ensure adefinable Al exposure concentration. Inorganic Al (Ali) present inthe stock solution will in a time-dependent manner respond tochanges in pH (and temperature) and reach a semi-stable level
Table 2Mortality (number of fish) of fish tagged with acoustic transmitters and untaggedcontrol fish (fish retained in laboratory, used for determining physiological status ofthe groups) during the laboratory exposure to the experimental treatments.
Treatment Fish with acoustic transmitters Untagged control fish
Release 1n ¼ 15
Release 2n ¼ 15
Release 3n ¼ 15
Release 1n ¼ 40
Release 2n ¼ 40
Release 3n ¼ 40
Control 0 0 0 0 0 0Low-Al 1 0 0 0 0 0High-Al 9 10 0* 1 1 0
* Exposure time reduced from 90 h to 48 h due to high mortality of previousrelease groups.

E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e43 37
within 10 min after addition at a temperature of 4e8 �C (Kroglundet al., 2001a; Teien et al., 2004a). Based on this, we assumed that a20 min ageing period would stabilise Al to the target level in theexposure waters, and that the additional ageing and mixing in theexposure tanks would ensure stable exposure conditions.
Water temperature, pH and conductivity were measured in situin all tanks using a WTW pH/Cond 340i meter fitted with a Radi-ometer pHC2005 electrode (Radiometer; Denmark), a TFK 325temperature sensor and a 325 Tetracon conductivity cell. Watersamples for determination of main ionic composition were takenfrom the control tank. Al was fractionated in all water types withrespect to charge and reactivity. Total-Al was analysed on aninductively-coupled plasma atomic emission spectrometer ICP-AES(Perkin Elmer), and total reactive Al (RAl) and total non-labile Al(nLAl) were analysed by flow injection analyses (FIA) using thepyrocatechol-violet (PCV) method (Røgeberg and Henriksen, 1985).nLAl is the fraction of Al that passed through a cation-exchangecolumn (Amberlite). The labile Al fraction (cationic or inorganicmonomeric, LAl) was calculated as the difference between RAl andnLAl. Total organic carbon (TOC) was determined after oxidation byperoxydisulfate and UV-irradiation. Major base cations (Ca, Mg, Na,K, Si) were analysed using standard laboratory protocols(Kristensen et al., 2009). Acid neutralising capacity (ANC) is ameasure of water sensitivity to acidification. The ANC-value wascalculated as S base cations minus S strong acid anions.
2.6. Monitoring of fish with acoustic transmitters after release
After exposure, the tagged fish were quickly transported(<5 min) to the release site in the river mouth (surface watertemperature 4.5e5.7 �C). The release site was in the brackishtransition zone, with salinity of 0.8 at the surface and 23.5 at thebottom at 1.5 m depth. Hence, the released fish could either chooseto remain in freshwater for some time, or immediately move downinto the more saline water and initiate the marine migration. Fishmovements were recorded by 28 receivers (VEMCOVR2)moored atfour sites in the fjord system (Fig. 1). Signals from the transmitterswere detected as the fish passed through the receiver ranges. Thetransmitter identification code, detection date, time and depthwere then recorded. At site 1, a grid of 11 receivers was placed in theinner part of the fjord close to the river mouth. The distance fromthe river mouth to the outermost receiver was 2.2 km. At site 2,3.6 km from the rivermouth, four receivers were distributed evenlyin a line across the fjord, being 1.5 km wide. At site 3, 9.6 km fromthe river mouth, five receivers were distributed evenly in a lineacross the fjord, being 1.5 kmwide. At site 4, 37.1 km from the rivermouth, eight receivers were distributed evenly across the fjord,being 2.6 km wide. The receiver range for the transmitters wastypically 200e450 m for transmitters at 0.5e3.0 m depth, butvaried with factors such as wave action, salinity and depth. Hence,post-smolts were unlikely to pass receiver sites without beingrecorded, which was supported by the observation that all fishdetected at a site had been detected passing previous sites, exceptfor a single fish that passed site 2 undetected. Sea depths at thereceiver sites were 26e288 m, and receivers were attached 3e5 mbelow surface to a mooring. Data were downloaded on 15 July2009.
Post-smolt mortality was presumed when individually taggedfish disappeared between two receiver sites and were not laterrecorded, or the transmitter remained stationary without anymovements for the entire study period. As described by Thorstadet al. (2011b; 2012b), depth recordings were used to distinguishbetween live post-smolt, and post-smolt that had been eaten, i.e.,with the transmitter remaining within a moving fish predator.Atlantic cod (Gadus morhua) and saithe (Pollachius virens), the main
fish predators in the fjord (Jepsen et al., 2006), stay at much greaterdepths (down to a maximum of 43e103 m), only periodicallymoving to the surface during feeding, while salmon post-smoltstypically remain in the upper 1e3 m (Hedger et al., 2011;Thorstad et al., 2011b). Hence, transmitters descending from thesurface to great depths were interpreted as predation mortality.
2.7. Physiological status of the exposure groups
To assess physiological status of the groups, untagged fish weretreated in the same way as the tagged fish, being simultaneouslyexposed in the same tanks and water quality, but separated fromthe tagged fish by a net wall dividing the tanks into two similarcompartments (Table 2). At the end of the exposure, the untaggedfish were sampled for determination of gill Naþ, Kþ-ATPase activity,total gill aluminium content (gill-Al), and plasma levels of chlorideions, GH and IGF-I. To assess seawater adaptability, seawater chal-lenge tests were performed on 14e15 fish for each group, byexposing them for 24 h to a salinity of 33e34 at 6e8 �C, after whichthe fish were sacrificed and plasma obtained for analysis of sodium(Naþ) and calcium (Ca2þ) levels.
To examine physiological recovery after Al-exposure, six un-tagged fish were retained in each of the exposure tanks in un-treated water for three days (release group 3) or six days (releasegroup 2) after which terminal samples were obtained for analysis ofgill Naþ, Kþ-ATPase activity and total gill Al-content.
To monitor the general status of the hatchery-fish during thestudy period, fish from the rearing tank from which all experi-mental fish were taken were also regularly sampled (referred to asrearing tank control) for physiological analyses to examine if thefish used for experiments had a different development than theremaining hatchery fish. Prior to experiments, gill Naþ, Kþ-ATPaseactivity and total gill-Al content were determined on 25 and 29April and blood Cl- on 27, 29 and 30 April for the rearing tankcontrol fish. Plasma GH and IGF-I levels were determined on 16, 25and 29 April. Seawater challenge tests (salinity 33e34 and watertemperature 3e8 �C for 24 h) were performed on 27 and 30 April,and on 4 May. However, determination of plasma Naþ and Ca2þ
levels as well as analysis of gill Naþ, Kþ-ATPase activity was onlycarried out on fish from the two first exposure dates. Further, ateach release date, similar sampling and seawater challenge testswere performed for the rearing tank control.
Fish used for physiological analyses had a mean body mass of83 g (range 26e250 g) and a mean body length of 209 mm (range151e304 mm). Thus, they were smaller than the tagged fish (one-way ANOVA, mass: F1,546 ¼ 593, P < 0.001, length: F1,546 ¼ 743,P < 0.001), as the larger fish were selected for tagging due to thesize of the transmitters.
For physiological sampling, fish were first anaesthetised usingclove oil (0.5 ml/l), and then killed by a blow to the head. Bloodsamples were taken from the caudal vein using 1-ml heparinisedsyringes. Plasma Cl- levels were analysed on site using whole bloodin an I-STAT blood analyser using a blood cartridge EG8þ (Abbott,Chicago, USA). The remaining blood was transferred to 1.5 mlEppendorf tubes and plasma obtained by centrifugation (3000� gfor 5min) and stored at�28 �C for subsequent analyses. The secondgill arch on the right hand side of each fish was dissected out andfrozen in pre-weighed, acid-washed polyethylene vials for freeze-drying and acid digesting for analysis of total gill-Al content,while the second gill arch on the left hand side was dissected outand frozen in 1.5 ml Eppendorf tubes containing 1 ml SEI buffersolution for analysis of gill Naþ, Kþ-ATPase activity (Zaugg, 1982).
The digested gill samples were analysed with respect to Al, iron(Fe), copper (Cu) and zinc (Zn) on an ICP-AES (Teien et al., 2004a).The tissue metal content (mg metal per g dry weight) thus includes

Distance from river mouth (km)
0 10 20 30 40
Su
rvival (p
ercen
tag
e o
f fish
)
0
20
40
60
80
100
Control, n = 45
Low Al , n = 44
High Al short exp, n = 15
High Al, n = 11
Site 2
Site 3
Site 4
0 0 0
14
12
9
6 6
4
22 21
15
Fig. 2. Survival (%) during the marine migration of acoustically tagged salmon smoltsat receiver sites 2, 3 and 4, between 3.6 and 37.1 km from the release site in the rivermouth. Sample sizes (n) given in the legend indicate the number of fish released, andnumbers in the figure indicate number of fish recorded alive at each site.
E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e4338
metal bound to both mucus and cell surfaces as well as metalincorporated inside the gill cells. Iron, copper and zinc wereincluded in the analyses to identify whether Al was the only metalcontaminant in the experiment. Metals have previously beenmeasured on non-anaesthetised fish. As fish were anaesthetised inthis experiment, some gill-metals were measured on both anaes-thetised and non-anaesthetised fish in order to verify that anaes-thesia per se had no effect on gill Al levels. Based on comparisons ofgill-Al values of anaesthetised and non-anaesthetised smolt fromthe same treatment groups (two-sample t-tests, all P > 0.16), therewas no difference in gill-Al levels between fish sampled with andwithout anesthetisation, which is consistent with the results foundby McCormick et al. (2009).
Gill Naþ, Kþ-ATPase activity was analysed using the method ofMcCormick (1993) as described in Kroglund et al. (2007). PlasmaGH was measured by homologous radioimmunoassay (RIA) asoutlined by Björnsson et al. (1994), and plasma IGF-I was measuredby RIA as outlined by Moriyama et al. (1994). Plasma sodium (Naþ)and calcium (Ca2þ) levels were measured by flame photometry(Eppendorf ELEX 6361).
Mortality and migration speeds of the tagged fish exposed to alow-Al level and the untreated controls could be compared as theywere released in similar numbers. Due to the high mortality of thehigh-Al group at release 1 and 2, and a reduced exposure time atrelease 3, the high-Al groupwas not included in statistical analyses,except in a comparisonwithin release 3 with the low-Al and controlgroups. Crosstabulations were used to compare mortality amonggroups (Fisher’s exact test and Pearson chi square). The high-Al,short exposure group was not included in tests of migrationspeeds due to a small sample size. Results from physiological an-alyses after three- and six-day recovery periods are descriptivelypresented due to small sample sizes. All tests were performed usingthe VUE database 1.6.4. (AMIRIX Systems Inc.), SPSS 17.0 (SPSS Inc.),and the software R (R Development Core Team, 2011).
3. Results
3.1. Water chemistry in the exposure tanks
The addition of AlCl3 and HCl affected the Hþ, Al and Cl con-centrations, reducing pH and ANC and increasing Al. The measuredincrease was 29 and 59 mg Al/l in the low-Al and high-Al groupsrespectively, of which 26 and 47 mg/l was recovered as LAlrespectively. The high proportion recovered as LAl was probablydue to the low content of organic matter in the treated water(0.6 mg C/l). Conductivity was in the range of 2.0e2.2 mS/m due tothe low concentration of all the major ions (Table 1), which rendersthe source water sensitive to pollutants.
3.2. Mortality of the tagged fish during exposure and marinemigration
Rapid and elevated mortality occurred during the treatmentperiod in the high-Al groups, totalling 60 and 67% for release groups1 and 2, respectively (Table 2). For release group 2, but not releasegroup 1, the fish that died during exposure were larger than thosesurviving (ManneWhitney U-test, body length, release 1: U ¼ 22.0,P¼ 0.061, release 2: U¼ 9.0, P¼ 0.055). The mortality of the taggedfish was higher than that of the untagged control fish in the high-Algroup during exposure, as only two untagged control fish died(Table 2). All the high-Al fish released (for both releases 1 and 2)died during the first 3.6 km of the marine migration (Fig. 2).
A larger proportion of the low-Al group than the tagged controlgroup died during the first 10 km of the marine migration (Fisher’sexact test, site 2 at 3.6 km: one-sided P¼ 0.077, site 3 at 9.6 km: one-
sided P ¼ 0.047, Fig. 2). Thus, by site 3, the cumulative mortality ofthe low-Al group was 73% while that of the control group was 53%.The mortality during the next 28 km of the marine migration, untilsite 4, did not differ between the low-Al and control group (Fisher’sexact test, one-sided P¼ 0.58, Fig. 2). Themortality of the fish in thehigh-Al group with a short exposure time (release 3) that had sur-vived the initial experimental treatment did not differ from thesimultaneously released low-Al or control fish during any stage ofthe migration (Pearson chi-square, c2 ¼ 0.68e3.59, P ¼ 0.17e0.71).
In all groups, the largest mortality occurred during the first3.6 km of the marine migration (Fig. 2). The total mortality duringthe first 37.1 km of the marine migration was 67% for the controlgroup, 80% for the low-Al group and 73% for the high-Al shortexposure group. Based on shifts to abnormal behavioural patternsand depth distributions, about one third of all mortalities in thesegroups (24 of 76 fish) was assigned to predation by marine fishes.This cause of death was proportionally similar between the low-Aland control group (Fisher’s exact test P ¼ 0.78), and between thehigh-Al short exposure group and the control group (Pearson chi-square, c2 ¼ 1.19, P ¼ 0.55). None of the fish in the high Al-groupcould be attributed to having succumbed to marine fish predation.
3.3. Migration speeds during the marine migration
Migration speeds from release until 3.6, 9.6 or 37.1 km from therelease site did not differ between the low-Al group and the controlgroup (ManneWhitney U-tests U ¼ 57e130, P ¼ 0.56e0.70). Thefish spent on average 11 h (range 1.9e86) from release until theirfirst detections at site 2 (3.6 km), 24 h (range 4.9e98) at site 3(9.6 km) and 67 h (range 24.8e441.7) at site 4 (37.1 km).
3.4. Gill-Al content
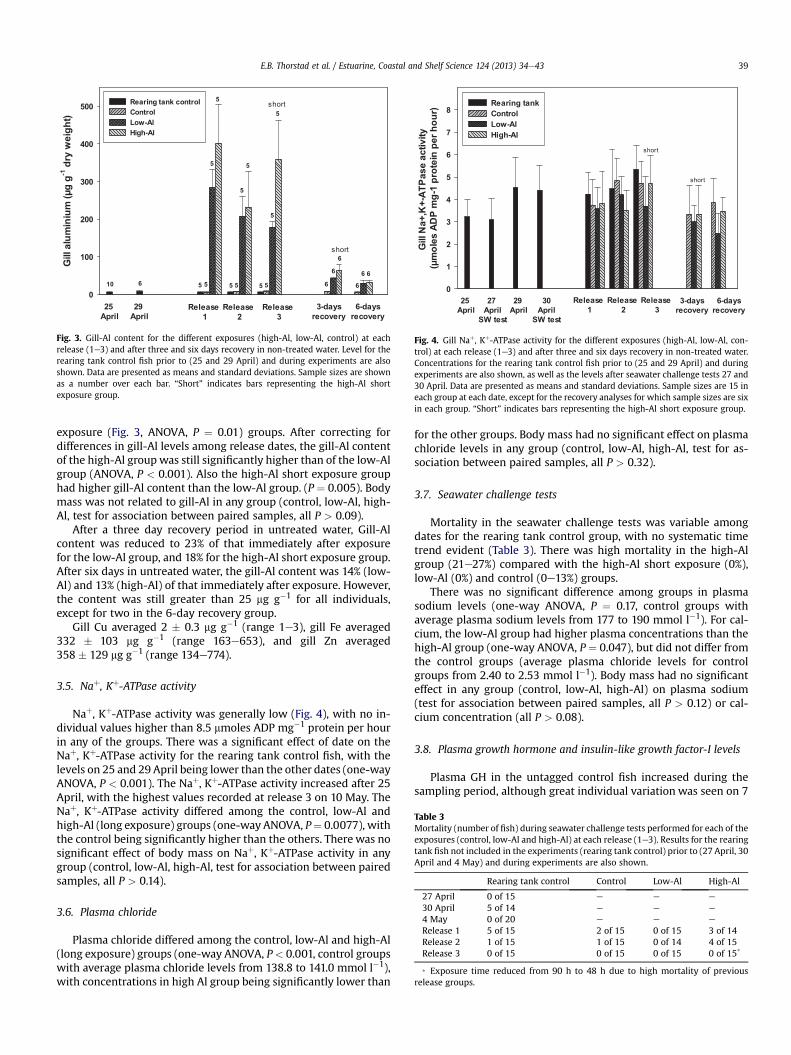
Al-exposure caused highly elevated gill-Al content in all expo-sure groups. Values averaged 223 � 60 (range 150e328), 317 � 128(range 128e527) and 358 � 105 mg g�1 (range 257e480) for thelow-Al, high-Al and high-Al short exposure groups, respectively(Fig. 3). By contrast, values for the untagged control group averaged7� 1.7 mg g�1 (range 4e12). There was a significant effect of releasedate on gill-Al, with decreasing levels evident for later releases forboth the low-Al (one-way ANOVA, P < 0.001) and high-Al long
Gill a
lum
iniu
m (
µg
g-1
dry
we
igh
t)
0
100
200
300
400
500Rearing tank control
Control
Low-Al
High-Al
25
April
29
April
Release
1
Release
2
Release
3
3-days
recovery
6-days
recovery
10 6
5
55
5
5
5
5
5 5
5
5 5 6
6
6
6 6
6
short
short
Fig. 3. Gill-Al content for the different exposures (high-Al, low-Al, control) at eachrelease (1e3) and after three and six days recovery in non-treated water. Level for therearing tank control fish prior to (25 and 29 April) and during experiments are alsoshown. Data are presented as means and standard deviations. Sample sizes are shownas a number over each bar. “Short” indicates bars representing the high-Al shortexposure group.
Gill N
a+
,K+
-A
TP
as
e a
ctiv
ity
(µ
mo
les
AD
P m
g-1
pro
te
in p
er h
ou
r)
0
1
2
3
4
5
6
7
8
Rearing tank
Control
Low-Al
High-Al
25
April
27
April
SW test
Release
1
Release
2
Release
3
3-days
recovery
6-days
recovery
29
April
30
April
SW test
short
short
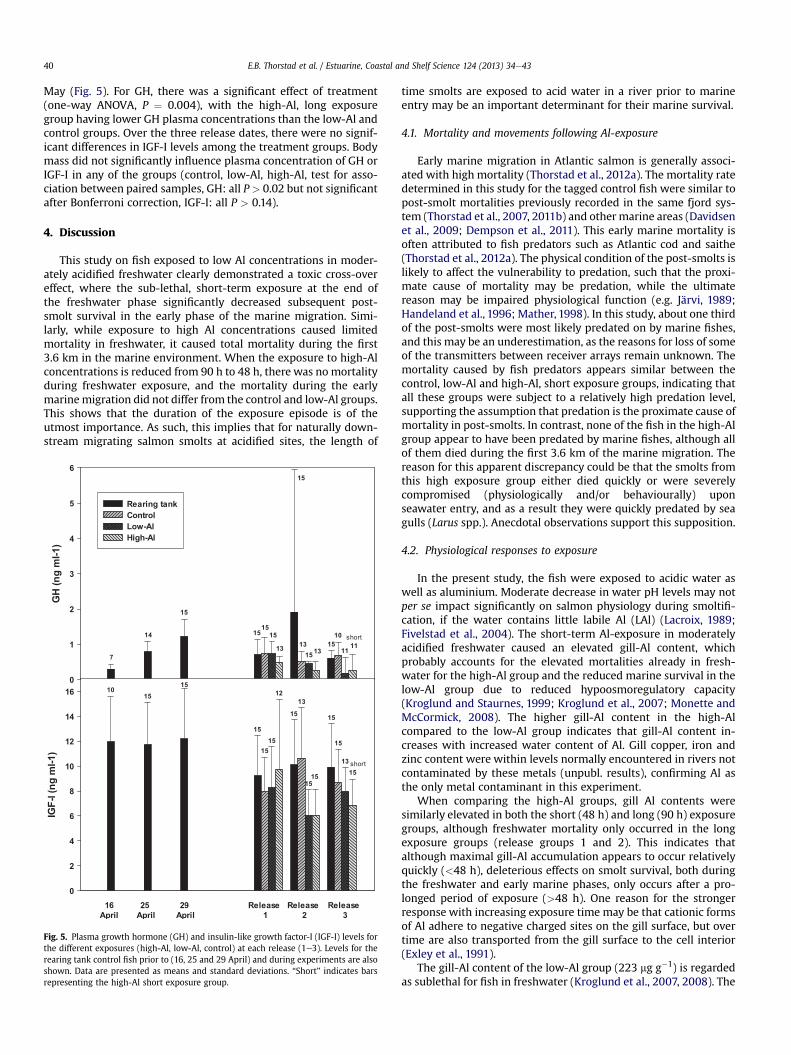
Fig. 4. Gill Naþ, Kþ-ATPase activity for the different exposures (high-Al, low-Al, con-trol) at each release (1e3) and after three and six days recovery in non-treated water.Concentrations for the rearing tank control fish prior to (25 and 29 April) and duringexperiments are also shown, as well as the levels after seawater challenge tests 27 and30 April. Data are presented as means and standard deviations. Sample sizes are 15 ineach group at each date, except for the recovery analyses for which sample sizes are sixin each group. “Short” indicates bars representing the high-Al short exposure group.
Table 3Mortality (number of fish) during seawater challenge tests performed for each of theexposures (control, low-Al and high-Al) at each release (1e3). Results for the rearingtank fish not included in the experiments (rearing tank control) prior to (27 April, 30April and 4 May) and during experiments are also shown.
Rearing tank control Control Low-Al High-Al
27 April 0 of 15 e e e
30 April 5 of 14 e e e
4 May 0 of 20 e e e
Release 1 5 of 15 2 of 15 0 of 15 3 of 14Release 2 1 of 15 1 of 15 0 of 14 4 of 15Release 3 0 of 15 0 of 15 0 of 15 0 of 15*
* Exposure time reduced from 90 h to 48 h due to high mortality of previousrelease groups.
E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e43 39
exposure (Fig. 3, ANOVA, P ¼ 0.01) groups. After correcting fordifferences in gill-Al levels among release dates, the gill-Al contentof the high-Al group was still significantly higher than of the low-Algroup (ANOVA, P < 0.001). Also the high-Al short exposure grouphad higher gill-Al content than the low-Al group. (P ¼ 0.005). Bodymass was not related to gill-Al in any group (control, low-Al, high-Al, test for association between paired samples, all P > 0.09).
After a three day recovery period in untreated water, Gill-Alcontent was reduced to 23% of that immediately after exposurefor the low-Al group, and 18% for the high-Al short exposure group.After six days in untreated water, the gill-Al content was 14% (low-Al) and 13% (high-Al) of that immediately after exposure. However,the content was still greater than 25 mg g�1 for all individuals,except for two in the 6-day recovery group.
Gill Cu averaged 2 � 0.3 mg g�1 (range 1e3), gill Fe averaged332 � 103 mg g�1 (range 163e653), and gill Zn averaged358 � 129 mg g�1 (range 134e774).
3.5. Naþ, Kþ-ATPase activity
Naþ, Kþ-ATPase activity was generally low (Fig. 4), with no in-dividual values higher than 8.5 mmoles ADP mg�1 protein per hourin any of the groups. There was a significant effect of date on theNaþ, Kþ-ATPase activity for the rearing tank control fish, with thelevels on 25 and 29 April being lower than the other dates (one-wayANOVA, P < 0.001). The Naþ, Kþ-ATPase activity increased after 25April, with the highest values recorded at release 3 on 10 May. TheNaþ, Kþ-ATPase activity differed among the control, low-Al andhigh-Al (long exposure) groups (one-way ANOVA, P¼ 0.0077), withthe control being significantly higher than the others. There was nosignificant effect of body mass on Naþ, Kþ-ATPase activity in anygroup (control, low-Al, high-Al, test for association between pairedsamples, all P > 0.14).
3.6. Plasma chloride
Plasma chloride differed among the control, low-Al and high-Al(long exposure) groups (one-way ANOVA, P< 0.001, control groupswith average plasma chloride levels from 138.8 to 141.0 mmol l�1),with concentrations in high Al group being significantly lower than
for the other groups. Body mass had no significant effect on plasmachloride levels in any group (control, low-Al, high-Al, test for as-sociation between paired samples, all P > 0.32).
3.7. Seawater challenge tests
Mortality in the seawater challenge tests was variable amongdates for the rearing tank control group, with no systematic timetrend evident (Table 3). There was high mortality in the high-Algroup (21e27%) compared with the high-Al short exposure (0%),low-Al (0%) and control (0e13%) groups.
There was no significant difference among groups in plasmasodium levels (one-way ANOVA, P ¼ 0.17, control groups withaverage plasma sodium levels from 177 to 190 mmol l�1). For cal-cium, the low-Al group had higher plasma concentrations than thehigh-Al group (one-way ANOVA, P ¼ 0.047), but did not differ fromthe control groups (average plasma chloride levels for controlgroups from 2.40 to 2.53 mmol l�1). Body mass had no significanteffect in any group (control, low-Al, high-Al) on plasma sodium(test for association between paired samples, all P > 0.12) or cal-cium concentration (all P > 0.08).
3.8. Plasma growth hormone and insulin-like growth factor-I levels
Plasma GH in the untagged control fish increased during thesampling period, although great individual variation was seen on 7
E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e4340
May (Fig. 5). For GH, there was a significant effect of treatment(one-way ANOVA, P ¼ 0.004), with the high-Al, long exposuregroup having lower GH plasma concentrations than the low-Al andcontrol groups. Over the three release dates, there were no signif-icant differences in IGF-I levels among the treatment groups. Bodymass did not significantly influence plasma concentration of GH orIGF-I in any of the groups (control, low-Al, high-Al, test for asso-ciation between paired samples, GH: all P> 0.02 but not significantafter Bonferroni correction, IGF-I: all P > 0.14).
4. Discussion
This study on fish exposed to low Al concentrations in moder-ately acidified freshwater clearly demonstrated a toxic cross-overeffect, where the sub-lethal, short-term exposure at the end ofthe freshwater phase significantly decreased subsequent post-smolt survival in the early phase of the marine migration. Simi-larly, while exposure to high Al concentrations caused limitedmortality in freshwater, it caused total mortality during the first3.6 km in the marine environment. When the exposure to high-Alconcentrations is reduced from 90 h to 48 h, there was no mortalityduring freshwater exposure, and the mortality during the earlymarinemigration did not differ from the control and low-Al groups.This shows that the duration of the exposure episode is of theutmost importance. As such, this implies that for naturally down-stream migrating salmon smolts at acidified sites, the length of
GH
(n
g m
l-1)
0
1
2
3
4
5
6
Rearing tank
Control
Low-Al
High-Al
IGF
-I (n
g m
l-1)
0
2
4
6
8
10
12
14
16
16
April
25
April
Release
1
Release
2
Release
3
29
April
14
7
15
15
15
1313
15
15
13
15
11
10
15 11
10
15
15
12
15
15
15
15
15
13
15
15
13
15
15
short
short
Fig. 5. Plasma growth hormone (GH) and insulin-like growth factor-I (IGF-I) levels forthe different exposures (high-Al, low-Al, control) at each release (1e3). Levels for therearing tank control fish prior to (16, 25 and 29 April) and during experiments are alsoshown. Data are presented as means and standard deviations. “Short” indicates barsrepresenting the high-Al short exposure group.
time smolts are exposed to acid water in a river prior to marineentry may be an important determinant for their marine survival.
4.1. Mortality and movements following Al-exposure
Early marine migration in Atlantic salmon is generally associ-ated with high mortality (Thorstad et al., 2012a). The mortality ratedetermined in this study for the tagged control fish were similar topost-smolt mortalities previously recorded in the same fjord sys-tem (Thorstad et al., 2007, 2011b) and other marine areas (Davidsenet al., 2009; Dempson et al., 2011). This early marine mortality isoften attributed to fish predators such as Atlantic cod and saithe(Thorstad et al., 2012a). The physical condition of the post-smolts islikely to affect the vulnerability to predation, such that the proxi-mate cause of mortality may be predation, while the ultimatereason may be impaired physiological function (e.g. Järvi, 1989;Handeland et al., 1996; Mather, 1998). In this study, about one thirdof the post-smolts were most likely predated on by marine fishes,and this may be an underestimation, as the reasons for loss of someof the transmitters between receiver arrays remain unknown. Themortality caused by fish predators appears similar between thecontrol, low-Al and high-Al, short exposure groups, indicating thatall these groups were subject to a relatively high predation level,supporting the assumption that predation is the proximate cause ofmortality in post-smolts. In contrast, none of the fish in the high-Algroup appear to have been predated by marine fishes, although allof them died during the first 3.6 km of the marine migration. Thereason for this apparent discrepancy could be that the smolts fromthis high exposure group either died quickly or were severelycompromised (physiologically and/or behaviourally) uponseawater entry, and as a result they were quickly predated by seagulls (Larus spp.). Anecdotal observations support this supposition.
4.2. Physiological responses to exposure
In the present study, the fish were exposed to acidic water aswell as aluminium. Moderate decrease in water pH levels may notper se impact significantly on salmon physiology during smoltifi-cation, if the water contains little labile Al (LAl) (Lacroix, 1989;Fivelstad et al., 2004). The short-term Al-exposure in moderatelyacidified freshwater caused an elevated gill-Al content, whichprobably accounts for the elevated mortalities already in fresh-water for the high-Al group and the reduced marine survival in thelow-Al group due to reduced hypoosmoregulatory capacity(Kroglund and Staurnes, 1999; Kroglund et al., 2007; Monette andMcCormick, 2008). The higher gill-Al content in the high-Alcompared to the low-Al group indicates that gill-Al content in-creases with increased water content of Al. Gill copper, iron andzinc content were within levels normally encountered in rivers notcontaminated by these metals (unpubl. results), confirming Al asthe only metal contaminant in this experiment.
When comparing the high-Al groups, gill Al contents weresimilarly elevated in both the short (48 h) and long (90 h) exposuregroups, although freshwater mortality only occurred in the longexposure groups (release groups 1 and 2). This indicates thatalthough maximal gill-Al accumulation appears to occur relativelyquickly (<48 h), deleterious effects on smolt survival, both duringthe freshwater and early marine phases, only occurs after a pro-longed period of exposure (>48 h). One reason for the strongerresponse with increasing exposure time may be that cationic formsof Al adhere to negative charged sites on the gill surface, but overtime are also transported from the gill surface to the cell interior(Exley et al., 1991).
The gill-Al content of the low-Al group (223 mg g�1) is regardedas sublethal for fish in freshwater (Kroglund et al., 2007, 2008). The

E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e43 41
present data on the low Al group support this assumption, eventhough salinities 21e29 were lower than full strength seawater inthis part of the fjord. The control fish had gill-Al levels that have notpreviously been associated with any reduction in seawater per-formance (Kroglund and Finstad, 2003; Fivelstad et al., 2004).
A 3-day recovery period in good quality freshwater led to aconsiderable reduction in gill-Al content in the exposed groups,with a further recovery during the following three days, in linewithKroglund et al. (2001b; 2012). This indicates that if juvenile salmonare exposed to episodes of acid water, linked to rainfall, during thefreshwater phase, the timing of such exposures in relation to thesmolt migration period is of great importance for long-term sur-vival. Thus, the closer such episodes occur in relation to seawaterentry, the greater the mortality is likely to be. However, after threeand six days recovery, many individuals still had a gill-Al content atlevels that may reduce post-smolt survival (>25 mg g�1, Kroglundet al., 2008), emphasising that even acid exposure episodes priorto the smolt run may have a negative impact on marine survival.
The present study did not reveal any behavioural effects linkedto the high gill-Al content, as the migration speeds did not differbetween the control and Al-exposed groups. Hence, there were noindications that the Al-exposed smolts lingered in freshwater at therelease site. The migration speeds were similar to, or faster than,those recorded for hatchery post-smolt in the same fjord system inprevious years (Finstad et al., 2005; Thorstad et al., 2007), andcomparable to migration speeds recorded in other areas (e.g.Hedger et al., 2008; Davidsen et al., 2009).
High gill-Al content was associated with reduced gill Naþ, Kþ-ATPase activity in the Al-exposed groups, similar to other studies(Staurnes et al., 1993; Magee et al., 2003; Kroglund et al., 2007;Monette et al., 2008). The activity of this enzyme increases rapidlyduring smoltification as a key component of the physiologicaladaptation to seawater (Wedemeyer et al., 1980), and it has beenreported that lowered Naþ, Kþ-ATPase activity following Al-exposure reduces seawater tolerance and marine survival (Mageeet al., 2001, 2003; Kroglund and Finstad, 2003; Kroglund et al.,2007). As expected, gill Naþ, Kþ-ATPase activity in the hatchery-smolts used in the present study increased from 25 April until 10May, just prior to the observed peak smolt run in the River Eira,which occurred during 8e19 May in 2009 (Hedger et al., 2011).However, measured Naþ, Kþ-ATPase activity was generally low inall groups compared with other studies (Finstad et al., 2007;Kroglund et al., 2007), which may indicate a relatively poor smoltquality and reduced seawater tolerance even for the control fish.This was also indicated by high plasma sodium levels measured(fish with <170 mM Naþ plasma levels following a 24 h seawaterchallenge are regarded as seawater tolerant smolts, Blackburn andClarke,1987) and elevatedmortalities in some of the control groupsduring seawater challenge tests. In contrast, migration speeds andsurvival of the control fish released in the fjord were similar tomigration speeds and survival rates recorded in previous studies inthe same area when the smolt quality was assessed as acceptable(Thorstad et al., 2007, 2011b), which does not indicate a poor smoltquality. However, as the post-smolts did not encounter full strengthseawater during at least the first 10 km of themarinemigration, butprobably experienced salinities around 21e29 during this stage, itis possible that they could maintain survival and migration speedseven with a poorer smolt quality.
The present results predict reduced adult return rates aftermoderate acid water and Al-exposure, as indicated in a tagging-recapture study by Kroglund et al. (2007). In addition, we showthat smolt mortality due to acidification only occurred during thefirst 10 km of seawater migration, while those surviving the first10 km seemed to recover. This agrees with the finding that Al-exposed smolts that survive the marine migration and return to
the river as adults do not have reduced marine growth comparedwith the unexposed control fish (Kroglund and Finstad, 2003;Kroglund et al., 2008). Monette and McCormick (2008) also foundthat if smolts survive the initial toxic effects of acid/Al exposure, theymay be able to recover ion homeostasis. Plasma chloride levels forthe smolts in the present experiment during their time in freshwater followed a similar pattern to that reported by Kroglund et al.(2007, 2012), where the high-Al group had significantly lowerplasma chloride, coinciding with the highest mortality in these twogroups. For the other groups, hydromineral balance appearednormal.
Al-exposure resulted in decreased plasma GH levels in the high-Al group, whereas no effect was seen on IGF-I levels. In contrast,Monette et al. (2008) found that Al exposure and moderate acidi-fication decreased plasma IGF-I levels, but not GH levels. Thus,while the studies are not in full agreement, both indicate delete-rious effects of Al exposure on the GH-IGF-I system, resulting inmodulated GH and IGF-I levels. These two hormones are intimatelylinked through stimulatory/inhibitory feedback mechanisms(Björnsson, 1997; Björnsson et al., 2002). GH and IGF-I are bothinvolved in development of seawater tolerance in teleost fishes(Agustsson et al., 2001). In salmonids, the GH-IGF-I system is ofimportance not just for osmoregulation, but also in the regulationof growth, protein and lipid metabolism as well as behaviour(Björnsson, 1997; Björnsson et al., 2002; Jönsson and Björnsson,2002). Monette et al. (2008) suggested that the Al-induced reduc-tion in IGF-I levels may be through an indirect mechanism, i.e. as aresult of a down-regulation of salt secretory mechanisms, orthrough a direct mechanism, i.e. specific toxic actions of Al on IGF-Isecretion. Together, these studies indicate a more general impact ofthe GH-IGF-I system, but whether this is due to direct or indirecteffects, remains to be elucidated. In agreement with Monette et al.(2008), the present study supports the hypothesis that the loss ofseawater tolerance in smolts during acid and Al exposure may bemediated, in part, by negative impacts on the GH-IGF-I system.
Even though there was a high mortality in the high-Al groupduring their experimental exposure in freshwater, this only appliedfor the tagged fish, and not for the control fish which had been keptunder the same husbandry conditions, and later sampled for theevaluation of physiological parameters. This may indicate thattagged fish were more susceptible to Al-exposure compared tountagged fish. Due to the implantation procedure, the tagged fishwere unavoidably somewhat larger than the untagged fish, andlarger fish aremore sensitive to Al exposure (Rosseland et al., 2001).Hence, the difference in mortality between tagged and untaggedfish may have been a size effect. This is supported by the fact thatthe tagged fish that died in the high-Al exposure tank were largerthan those surviving until release. No body size effects were foundin any analyses of the physiological parameters, but this does notexclude a possible size effect, as there was little overlap in sizerange between the tagged and untagged groups. Therefore, a po-tential tagging effect cannot be separated from a size effect in thisstudy. The transmitters and tagging method used was not expectedto cause any adverse effects on swimming speeds, and the recoverytime of more than two weeks has been deemed sufficient for thefish to recover from any tagging effects before exposure (Peakeet al., 1997).
This study was performed using hatchery-reared smolts,because it was difficult to capture a sufficient number of wildsmolts large enough to be implanted with a transmitter. Even withgenerally larger hatchery-reared smolts, due to the transmitter sizeonly the largest fish could be used for implantation. Using wildsmolts may also have been ethically questionable due to thethreatened status of many Atlantic salmon stocks. However, animportant question is whether the results are representative also

E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e4342
for wild smolts. First generation hatchery-reared smolts of the localRiver Eira stock were used, and it may be assumed that these fishdid not differ from thewild smolts in the susceptibility to Al, exceptthe size difference, as discussed above. Previous studies haveshown that hatchery-reared and wild post-smolts did not differ intheir migration behaviour during their seaward migration (Øklandet al., 2006; Thorstad et al., 2007), indicating that results fromhatchery-reared smolts during this migration stage may be repre-sentative also for wild post-smolts.
4.3. Concluding remarks
This study simulated an episode with elevated Al associatedwith moderate acidification in freshwater during the last few daysprior to sea entry of the smolts. While fish will be exposed to amuchmore complex variation inwater chemistry in time and spacein a river, the results indicated that short term increases inbioavailable Al may have population consequences by increasingthe marine post-smolt mortality. The high proportion of Al recov-ered as LAl, was a stronger response than expected from theamount of added Al, probably because the water in the hatcherywas cold, and had a low conductivity and low content of organicmatter. Soft and clear water rivers are sensitive to disturbances, anda specific dose of a contaminant will cause stronger responses infish than occur in waters having a higher ionic or organic content(Rosseland and Kroglund, 2011).
Short-term Al-exposure in moderately acidified freshwaterresulted in increased gill-Al, compromised smolt hypoosmor-egulatory capacity, reduced seawater tolerance and increased earlymarine mortality. Responses to short-term Al-exposure are notonly a result of absolute Al-concentrations, but are also highlydependent on the duration of exposure. This also emphasises theimportance of the timing of such episodes in relation to the peaksmolt run, since only a few days recovery in good quality water mayreduce gill-Al content considerably. In summary, short-term Al-exposure in moderately acidified freshwater increased immediatemortality during the first 10 km of the marine migration by 20%compared to the control group, but thereafter did not affect themigration speeds of the survivors. Hence, the exposure eithercaused a quick death, or post-smolts surviving the first 10 kmseemed to recover and had a normal migration speed and survivalthereafter. The proximate cause for elevated mortality may havebeen predation by marine fishes. Post-smolt mortality caused bysuch Al-exposure episodes may contribute further in the decline inwild Atlantic salmon populations.
Acknowledgements
We thank the staff at the Statkraft hatchery in Eresfjord forextensive help and co-operation during the project. We also thankOle Kristian Bjølstad and John Kofoed for assistance during fieldwork. The project was financed by the Norwegian Research Councilproject number 183992/S30, Statkraft Energy AS and the Norwe-gian Directorate for Nature Management. The experiment wasapproved by the Norwegian Animal Research Authority.
References
Anon, 2011. The Status of Norwegian Salmon Stocks in 2010. Report from theNorwegian Scientific Advisory Committee for Atlantic Salmon Management 3,pp. 1e285 (in Norwegian).
Agustsson, T., Sundell, K., Sakamoto, T., Johansson, V., Ando, M., Björnsson, B. Th.,2001. Growth hormone endocrinology of Atlantic salmon (Salmo salar): pitui-tary gene expression, hormone storage, secretion and plasma levels duringparr-smolt transformation. Journal of Endocrinology 170, 227e234.
Björnsson, B. Th., 1997. The biology of salmon growth hormone: from daylight todominance. Fish Physiology and Biochemistry 17, 9e24.
Björnsson, B. Th., Taranger, G.L., Hansen, T., Stefansson, S.O., Haux, C., 1994. Theinterrelation between photoperiod, growth-hormone, and sexual maturation ofadult Atlantic salmon (Salmo salar). General and Comparative Endocrinology93, 70e81.
Björnsson, B. Th., Johansson, V., Benedet, S., Einarsdottir, I.E., Hildahl, J.,Agustsson, T., Jönsson, E., 2002. Growth hormone endocrinology of salmonids:regulatory mechanisms and mode of action. Fish Physiology and Biochemistry27, 227e242.
Blackburn, J., Clarke, W.C., 1987. Revised procedure for the 24 hour seawater chal-lenge test to measure seawater adaptability of juvenile salmonids. CanadianTechnical Report of Fisheries and Aquatic Sciences 1515, 1e35.
Davidsen, J.G., Rikardsen, A.H., Halttunen, E., Thorstad, E.B., Økland, F., Letcher, B.H.,Skarðhamar, J., Næsje, T.F., 2009. Migratory behaviour and survival rates of wildnorthern Atlantic salmon Salmo salar post-smolts: effects of environmentalfactors. Journal of Fish Biology 75, 1700e1718.
Dempson, J.B., Robertson, M.J., Pennell, C.J., Furey, G., Bloom, M., Shears, M.,Ollerhead, L.M.N., Clarke, K.D., Hinks, R., Robertson, G.J., 2011. Residency time,migration route and survival of Atlantic salmon Salmo salar smolts in a Cana-dian fjord. Journal of Fish Biology 78, 1976e1992.
Exley, C., Chappell, J.S., Birchall, J.D., 1991. A mechanism for acute aluminiumtoxicity in fish. Journal of Theoretical Biology 151, 417e428.
Finstad, B., Økland, F., Thorstad, E.B., Bjørn, P.A., McKinley, R.S., 2005. Migration ofhatchery-reared Atlantic salmon and wild anadromous brown trout post-smolts in a Norwegian fjord system. Journal of Fish Biology 66, 86e96.
Finstad, B., Kroglund, F., Strand, R., Stefansson, S.O., Bjørn, P.A., Rosseland, B.O.,Nilsen, T.O., Salbu, B., 2007. Salmon lice or suboptimal water quality e reasonsfor reduced postsmolt survival? Aquaculture 273, 374e383.
Fivelstad, S., Olsen, A.B., Stefansson, S., Handeland, S., Waagbø, R., Kroglund, F.,Colt, J., 2004. Lack of long-term sublethal effects of reduced freshwater pHalone on Atlantic salmon (Salmo salar) smolts subsequently transferred toseawater. Canadian Journal of Fisheries and Aquatic Sciences 61, 511e518.
Gensemer, R.W., Playle, R.C., 1999. The bioavailability and toxicity of aluminium inaquatic environments. Critical Reviews in Environmental Science and Tech-nology 29, 315e450.
Handeland, S.O., Järvi, T., Fernö, A., Stefansson, S.O., 1996. Osmotic stress, anti-predator behavior, and mortality of Atlantic salmon (Salmo salar) smolts. Ca-nadian Journal of Fisheries and Aquatic Sciences 53, 2673e2680.
Hedger, R.D., Martin, F., Hatin, D., Caron, F., Whoriskey, F., Dodson, J., 2008. Activemigration of wild Atlantic salmon Salmo salar through a coastal embayment.Marine Ecology Progress Series 355, 235e246.
Hedger, R.D., Uglem, I., Thorstad, E.B., Finstad, B., Chittenden, C.M., Arechavala-Lopez, P., Jensen, A.J., Nilsen, R., Økland, F., 2011. Behavioural strategies ofAtlantic cod, a marine fish predator during Atlantic salmon post-smolt migra-tion. ICES Journal of Marine Science 68, 2152e2162.
Hesthagen, T., Hansen, L.P., 1991. Estimates of the annual loss of Atlantic salmon,Salmo salar L., in Norway due to acidification. Aquaculture and Fisheries Man-agement 22, 85e91.
Hvidsten, N.A., Møkkelgjerd, P.I., 1987. Predation on salmon smolts, Salmo salar L., inthe estuary of the River Surna. Journal of Fish Biology 30, 273e280.
ICES (International Council for the Exploration of the Sea), 2011. Report of theWorking Group on North Atlantic Salmon (WGNAS). ICES 2011/ACOM 09.
Järvi, T., 1989. Synergistic effect on mortality in Atlantic salmon, Salmo salar, smoltcaused by osmotic stress and presence of predators. Environmental Biology ofFishes 26, 149e152.
Jepsen, N., Holthe, E., Økland, F., 2006. Observations of predation on salmonand trout smolts in a river mouth. Fisheries Management and Ecology 13,341e343.
Jönsson, E., Björnsson, B. Th., 2002. Physiological functions of growth hormone infish with special reference to its influence on behaviour. Fisheries Science 68(Suppl. I), 742e748.
Jonsson, N., Jonsson, B., Hansen, L.P., 1998. The relative role of density-dependentand density-independent survival in the life cycle of Atlantic salmon Salmosalar. Journal of Animal Ecology 67, 751e762.
Kristensen, T., Åtland, Å., Rosten, T., Urke, H.A., Rosseland, B.O., 2009. Importantinfluent-water quality parameters at freshwater production sites in two salmonproducing countries. Aquacultural Engineering 41, 53e59.
Kroglund, F., Finstad, B., 2003. Low concentrations of inorganic monomericaluminium impair physiological status and marine survival of Atlantic salmon.Aquaculture 222, 119e133.
Kroglund, F., Staurnes, M., 1999. Water quality requirements of smolting Atlanticsalmon (Salmo salar) in limed acid rivers. Canadian Journal of Fisheries andAquatic Sciences 56, 2078e2086.
Kroglund, F., Teien, H.C., Rosseland, B.O., Salbu, B., 2001a. Time and pH-dependentdetoxification of aluminum in mixing zones between acid and non-acid rivers.Water, Air, and Soil Pollution 130, 911e916.
Kroglund, F., Teien, H.C., Rosseland, B.O., Salbu, B., Lucassen, C.H.E.T., 2001b. Waterquality dependent recovery from aluminium stress in Atlantic salmon smolt.Water, Air, and Soil Pollution 130, 905e910.
Kroglund, F., Rosseland, B.O., Teien, H.-C., Salbu, B., Kristensen, T., Finstad, B., 2008.Water quality limits for Atlantic salmon (Salmo salar L.) exposed to short termreductions in pH and increased aluminum simulating episodes. Hydrology andEarth System Sciences 12, 491e507.
Kroglund, F., Finstad, B., Stefansson, S.O., Nilsen, T.O., Kristensen, T., Rosseland, B.O.,Teien, H.C., Salbu, B., 2007. Exposure to moderate acid water and aluminumreduces Atlantic salmon post-smolt survival. Aquaculture 273, 360e373.

E.B. Thorstad et al. / Estuarine, Coastal and Shelf Science 124 (2013) 34e43 43
Kroglund, F., Finstad, B., Pettersen, K., Teien, H.-C., Salbu, B., Rosseland, B.O.,Nilsen, T.O., Stefansson, S., Ebbesson, L.O.E., Nilsen, R., Bjørn, P.A., Kristensen, T.,2012. Recovery of Atlantic salmon smolts following aluminium exposuredefined by changes in blood physiology and seawater tolerance. Aquaculture362-363, 232e240.
Lacroix, G.L., 1989. Ecological and physiological responses of Atlantic salmon inacidic organic rivers of Nova Scotia, Canada. Water, Air, and Soil Pollution 46,375e386.
Lucas, M., Baras, E., 2001. Migration of Freshwater Fishes. Blackwell Science, Oxford.Magee, J.A., Haines, T.A., Kocik, J.F., Beland, K.F., McCormick, S.D., 2001. Effects of
acidity and aluminium on the physiology and migratory behaviour of Atlanticsalmon smolts in Maine, USA. Water, Air, and Soil Pollution 130, 881e886.
Magee, J.A., Obedzinski, M., McCormick, S.D., Kocik, J.F., 2003. Effects of episodicacidification on Atlantic salmon (Salmo salar) smolts. Canadian Journal ofFisheries and Aquatic Sciences 60, 214e221.
Mather, M.E., 1998. The role of context-specific predation in understanding patternsexhibited by anadromous salmon. Canadian Journal of Fisheries and AquaticSciences 55 (Suppl. 1), 232e246.
McCormick, S.D., 1993. Methods for nonlethal gill biopsy and measurement of Naþ,Kþ-ATPase activity. Canadian Journal of Fisheries and Aquatic Sciences 50, 656e658.
McCormick, S.D., Hansen, L.P., Quinn, T.P., Saunders, R.L., 1998. Movement, migra-tion, and smolting of Atlantic salmon (Salmo salar). Canadian Journal of Fish-eries and Aquatic Sciences 55 (Suppl. 1), 77e92.
McCormick, S.D., Keyes, A., Sislow, K.H., Monette, M.Y., 2009. Impacts of episodicacidification on in-stream survival and physiological impairment of Atlanticsalmon (Salmo salar) smolts. Canadian Journal of Fisheries and Aquatic Sciences66, 394e403.
Milner, N.J., Elliott, J.M., Armstrong, J.D., Gardiner, R., Welton, J.S., Ladle, M., 2003.The natural control of salmon and trout populations in streams. FisheriesResearch 62, 111e125.
Monette, M.Y., McCormick, S.D., 2008. Impacts of short-term acid and aluminumexposure on Atlantic salmon (Salmo salar) physiology: a direct comparison ofparr and smolts. Aquatic Toxicology 86, 216e226.
Monette, M.Y., Björnsson, B. Th., McCormick, S.D., 2008. Effects of short-term acidand aluminum exposure on the parr-smolt transformation in Atlantic salmon(Salmo salar): disruption of seawater tolerance and endocrine status. Generaland Comparative Endocrinology 158, 122e130.
Moriyama, S., Swanson, P., Nishii, M., Takahashi, A., Kawauchi, H., Dickhoff, W.W.,Plisetskaya, E.M., 1994. Development of a homologous radioimmunoassay forcoho salmon insulin-like growth-factor-I. General and Comparative Endocri-nology 96, 149e161.
Økland, F., Thorstad, E.B., Finstad, B., Sivertsgård, R., Plantalech, N., Jepsen, N.,McKinley, R.S., 2006. Swimming speeds and orientation of wild Atlantic salmonpost-smolts during the first stage of the marine migration. Fisheries Manage-ment and Ecology 13, 271e274.
Peake, S., McKinley, R.S., Scruton, D.A., Moccia, R., 1997. Influence of transmitterattachment procedures on swimming performance of wild and hatchery-rearedAtlantic salmon smolts. Transactions of the American Fisheries Society 126,707e714.
R Development Core Team, 2011. R: a Language and Environment for StatisticalComputing. R Foundation for Statistical Computing, Vienna, Austria, ISBN 3-900051-07-0. URL: http://www.R-project.org/.
Røgeberg, E.J.S., Henriksen, A., 1985. An automatic method for fractionation anddetermination of aluminium species in freshwaters. Vatten, Water 41, 48e53.
Rosseland, B.O., Staurnes, M., 1994. Physiological mechanisms for toxic effects andresistance to acidic water: an ecophysiological and ecotoxicological approach.In: Steinberg, C.E.W., Wright, R.F. (Eds.), Acidification of Freshwater Systems:Implications for the Future. John Wiley & Sons Ltd., pp. 227e246.
Rosseland, B.O., Kroglund, F., 2011. Ecological consequences of pollution: lessonsfrom acidification and pesticides. In: Aas, Ø., Einum, S., Klemetsen, A., Skurdal, J.(Eds.), Atlantic Salmon Ecology. Wiley-Blackwell, pp. 387e408.
Rosseland, B.O., Kroglund, F., Staurnes, M., Hindar, K., Kvellestad, A., 2001. Toleranceto acid water among strains and life stages of Atlantic salmon (Salmo salar L.).Water, Air, and Soil Pollution 130, 899e904.
Staurnes, M., Blix, P., Reite, O.B., 1993. Effects of acid water and aluminium on parr-smolt transformation and seawater tolerance in Atlantic salmon, Salmo salar.Canadian Journal of Fisheries and Aquatic Sciences 50, 1816e1827.
Staurnes, M., Hansen, L.P., Fugelli, K., Haraldstad, O., 1996. Short-term exposure toacid water impairs osmoregulation, seawater tolerance, and subsequent marinesurvival of smolts of Atlantic salmon (Salmo salar). Canadian Journal of Fisheriesand Aquatic Sciences 53, 1695e1704.
Teien, H.C., Salbu, B., Kroglund, F., Rosseland, B.O., 2004a. Transformation of posi-tively charged aluminium-species in unstable mixing zones following liming.Science of the Total Environment 330, 217e232.
Teien, H.C., Standring, W.J.F., Salbu, B., Marskar, M., Kroglund, F., Hindar, A., 2004b.Mobilization of aluminium and deposition on fish gills during sea salt episodese catchment liming as countermeasure. Journal of Environmental Monitoring 6,191e200.
Thorstad, E.B., Whoriskey, F., Rikardsen, A.H., Aarestrup, K., 2011a. Aquatic nomads:the life and migrations of the Atlantic salmon. In: Aas, Ø., Einum, S.,Klemetsen, A., Skurdal, J. (Eds.), Atlantic Salmon Ecology. Wiley-Blackwell,pp. 1e32.
Thorstad, E.B., Uglem, I., Arechavala-Lopez, P., Økland, F., Finstad, B., 2011b. Lowsurvival of hatchery-released Atlantic salmon smolts during initial river andfjord migration. Boreal Environment Research 16, 115e120.
Thorstad, E.B., Whoriskey, F., Uglem, I., Moore, A., Rikardsen, A.H., Finstad, B., 2012a.A critical life stage of the Atlantic salmon Salmo salar: behaviour and survivalduring the smolt and initial post-smolt migration. Journal of Fish Biology 81,500e542.
Thorstad, E.B., Økland, F., Finstad, B., Sivertsgård, R., Plantalech, N., Bjørn, P.A.,McKinley, R.S., 2007. Fjord migration and survival of wild and hatchery-rearedAtlantic salmon and wild brown trout post-smolts. Hydrobiologia 582, 99e107.
Thorstad, E.B., Uglem, I., Finstad, B., Chittenden, C.M., Nilsen, R., Økland, F., Bjørn, P.-A., 2012b. Stocking location and predation by marine fishes affect survival ofhatchery-reared Atlantic salmon smolts. Fisheries Management and Ecology 19,400e409.
Watt, W.D., Scott, C.D., Zamora, P.J., White, W.J., 2000. Acid toxicity levels in NovaScotia rivers have not declined in synchrony with the decline in sulfate levels.Water, Air, and Soil Pollution 118, 203e229.
Wedemeyer, G., Saunders, R., Clarke, W., 1980. Environmental factors affectingsmoltification and early marine survival of anadromous salmonids. MarineFisheries Review 42, 1e14.
Zaugg, W.S., 1982. A simplified preparation for adenosine-triphosphatase deter-mination in gill tissue. Canadian Journal of Fisheries and Aquatic Sciences 39,215e217.
Copyright © 2022 FDOKUMEN




















