
Records of murine rodents (Mammalia, Rodentia) in the Pleistocene localities of Tan Vinh and Ma...
17
Original article Records of murine rodents (Mammalia, Rodentia) in the Pleistocene localities of Tan Vinh and Ma U’Oi (Northern Vietnam) and their implications to past distribution Présence de rongeurs murinae (Mammalia, Rodentia) dans les sites Pléistocènes de Tan Vinh et de Ma U’Oi (nord du Viêt-Nam) et leurs implications paléogéographiques Anne-Marie Bacon a, * , Fabrice Demeter b , Philippe Duringer c , Stéphane Rousse c , Yukio Dodo d , Hirofumi Matsumura e , Vu The Long f , Nguyen Kim Thuy f , Nguyen Thi Mai Huong f , Tomoko Anezaki g a UPR 2147 du CNRS, 44, rue de l’Amiral Mouchez, 75014 Paris, France b Unité scientifique du muséum « Ecoanthropologie et ethnobiologie », musée de l’homme, 17, place du Trocadéro, 75116 Paris, France c Université Louis-Pasteur, institut de géologie (EOST-CGS), UMR 7517, 1, rue Blessig, 67084 Strasbourg cedex, France d Department of Anatomy and Anthropology, Tohoku University School of Medicine, 2-1 Seiryo-machi Aoba-ku, Sendai 980-8575, Japan e Department of Anatomy, Sapporo Medical University, S1, W17, Chuou-Ku, Sapporo 060-8556, Japan f Institute of Archaeology, 61, Phan Chu Trinh, Hanoi, Vietnam g Systematics and Phylogeny Section, Primate Research Institute, Kyoto University, Inuyama, Aichi 484-8506, Japan Received 20 July 2005; accepted 28 March 2006 Available online 29 November 2006 Abstract We describe two Vietnamese fossiliferous sites located in Hoà Binh province, northern Vietnam. The site of Tan Vinh, first described here, is a quarry opened in Triassic limestone which exposes a karstic network and its filling. One of the fissure filling contained 104 molars of Leopoldamys sabanus (Roden- http://france.elsevier.com/direct/ANNPAL/ Annales de Paléontologie 92 (2006) 367–383 * Corresponding author. E-mail address: [email protected] (A.-M. Bacon). 0753-3969/$ - see front matter © 2006 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.annpal.2006.03.019
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Records of murine rodents (Mammalia, Rodentia) in the Pleistocene localities of Tan Vinh and Ma...

http://france.elsevier.com/direct/ANNPAL/
Annales de Paléontologie 92 (2006) 367–383
Original article
Records of murine rodents (Mammalia, Rodentia)in the Pleistocene localities of Tan Vinh
and Ma U’Oi (Northern Vietnam)and their implications to past distribution
Présence de rongeurs murinae (Mammalia, Rodentia)dans les sites Pléistocènes de Tan Vinh et de Ma U’Oi
(nord du Viêt-Nam) et leurs implicationspaléogéographiques
Anne-Marie Bacona,*, Fabrice Demeterb, Philippe Duringerc,Stéphane Roussec, Yukio Dodod, Hirofumi Matsumurae, Vu The Longf,
Nguyen Kim Thuyf, Nguyen Thi Mai Huongf, Tomoko Anezakig
aUPR 2147 du CNRS, 44, rue de l’Amiral Mouchez, 75014 Paris, FrancebUnité scientifique du muséum « Ecoanthropologie et ethnobiologie », musée de l’homme,
17, place du Trocadéro, 75116 Paris, FrancecUniversité Louis-Pasteur, institut de géologie (EOST-CGS), UMR 7517,
1, rue Blessig, 67084 Strasbourg cedex, FrancedDepartment of Anatomy and Anthropology, Tohoku University School of Medicine, 2-1 Seiryo-machi Aoba-ku,
Sendai 980-8575, JapaneDepartment of Anatomy, Sapporo Medical University, S1, W17, Chuou-Ku, Sapporo 060-8556, Japan
f Institute of Archaeology, 61, Phan Chu Trinh, Hanoi, Vietnamg Systematics and Phylogeny Section, Primate Research Institute, Kyoto University, Inuyama, Aichi 484-8506, Japan
Received 20 July 2005; accepted 28 March 2006Available online 29 November 2006
Abstract
We describe two Vietnamese fossiliferous sites located in Hoà Binh province, northern Vietnam. Thesite of Tan Vinh, first described here, is a quarry opened in Triassic limestone which exposes a karsticnetwork and its filling. One of the fissure filling contained 104 molars of Leopoldamys sabanus (Roden-
* Corresponding author.E-mail address: [email protected] (A.-M. Bacon).
0753-3969/$ - see front matter © 2006 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.annpal.2006.03.019

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383368
tia, Muridae). The biochronological age of these deposits can be estimated from late Middle Pleistoceneto Holocene. The site of Ma U’Oi is a cave opened in a karstified limestone which yielded 56 molars offour species of Muridae in two assemblages: Niviventer andersoni, L. sabanus, Niviventer fulvescensamong a mammal fauna dated from late Middle Pleistocene to Late Pleistocene, and L. sabanus,N. fulvescens and Hapalomys delacouri among a microvertebrate fauna dated to the late Middle Pleisto-cene. First, we present a description of the two sites in terms of sedimentology. Secondly, we comparemolars dimensions of the most common murids, L. sabanus and N. fulvescens, recovered at Ma U’Oiand Tan Vinh, with those recovered in a Thai site. The size difference between the specimens of thetwo species, especially those of L. sabanus in great numbers, from the northern sites and those from theThai site suggests a cline which can be due to latitudinal differences in palaeotemperatures. The occur-rence of N. andersoni, L. sabanus, N. fulvescens in Pleistocene Vietnamese sites allow to update the pastdistribution of these species.© 2006 Elsevier Masson SAS. All rights reserved.
Résumé
Deux localités fossilifères mises au jour dans la province de Hoà Binh, au nord du Viêt-Nam, sontdécrites. Le site de Tan Vinh, pour la première fois présenté ici, est une carrière formée de calcaire duTrias dont les fronts de taille exposent d’innombrables réseaux karstiques et leurs remplissages. Dans unde ces remplissages, nous avons trouvé 104 molaires de Leopoldamys sabanus (Rodentia, Muridae) dontl’âge biochronologique peut être estimé entre le pléistocène moyen récent et l’holocène. Le site de MaU’Oi est une grotte tapissée d’une brèche typique de caverne qui a livré 56 dents appartenant à quatreespèces de Muridae. Ces espèces ont été trouvées dans deux assemblages fauniques : Niviventer ander-soni, Niviventer fulvescens et L. sabanus associées à une faune de grands mammifères dont l’âge peutêtre estimé entre le pléistocène moyen récent et le pléistocène supérieur ; L. sabanus, N. fulvescens etHapalomys delacouri associée à une faune de microvertébrés datée du pléistocène moyen récent. En pre-mier lieu une description des deux sites en terme de sédimentologie est présentée. Puis, nous comparonsles dimensions des molaires des deux muridés les plus communs trouvés dans les sites vietnamiens(L. sabanus et N. fulvescens), avec celles des mêmes muridés provenant d’un site fossile de Thaïlande.La différence de taille observée entre les spécimens des deux espèces provenant de sites éloignés (parti-culièrement L. sabanus), permet de suggérer un cline qui peut s’expliquer par les variations latitudinalesdes paléotempératures. Enfin, la découverte de N. andersoni, L. sabanus et N. fulvescens dans le pléisto-cène du Viêt-Nam nous amène à réviser la répartition passée de ces espèces.© 2006 Elsevier Masson SAS. All rights reserved.
Keywords: Vietnam; Pleistocene; Holocene; Leopoldamys sabanus; Niviventer fulvescens; Distribution;Palaeoenvironment
Mots clés : Viêt-Nam ; Pléistocène ; Holocène ; Leopoldamys sabanus ; Niviventer fulvescens ; Biogéographie ;Paléoenvironnement
1. Introduction
The work that follows is based on Muridae (Rodentia) recovered in 2001 and 2002 in twosites, Ma U’Oi and Tan Vinh, in northern Vietnam, by a Vietnamese–French–Japanese team.
The site of Ma U’Oi is a limestone karst cave (Roussé et al., 2003; Bacon et al., 2004;Demeter et al., 2004; Bacon et al., 2006). The landscape of this area is characterised by a typi-cal morphology of karst peaks called “tower-karst”, developed in monotonous limestone bedsthick of several hundred metres and dated to Carboniferous and Triassic. The site of Ma U’Oiyielded in two different loci inside the cave remains of murids (Bacon et al., 2004, 2006).

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 369
From the walls of the cave, the breccia produced the in situ mammal assemblage (rhinocerotid,cervid, primate, suid, rodent) with two mandibles and 4 molars of the chesnut rat Niviventerfulvescens (Gray, 1847), the Chinese rat Niviventer andersoni (Thomas, 1911) and the long-tailed giant rat Leopoldamys sabanus (Thomas, 1887). The biochronological age of this assem-blage suggests an age from late Middle Pleistocene to Late Pleistocene and the U/Th dating aminimal age of 49 ± 4 kyr (data of Falguères cited in Bacon et al., 2006).
The following year, the breccia of the roof produced the in situ microvertebrate fauna(small-sized cervid, primate, insectivore, reptile, amphibian, rodent) with the muridsL. sabanus (14 molars, two incomplete mandibles), N. fulvescens (12 molars, two incompletemandibles) and the marmoset mouse Hapalomys delacouri (Thomas, 1927) (1 molar). This as-semblage is dated to 193 ± 17 kyr, the late Middle Pleistocene (data of Falguères cited in Ba-con et al., 2006).
The site of Tan Vinh was discovered while prospecting fissure fillings in quarries for theirfossiliferous potential. In Southeast Asia, fissure fillings are known as sources of vertebratefossils, especially those from the Pleistocene and Holocene (Chaimanee et al., 1993). TheTan Vinh quarry corresponds to a Triassic limestone. As the quarry is still being activelymined, we undertook a 3-day rescue excavation. The findings from Tan Vinh consist of 104well preserved molars of a single murid L. sabanus. This abundance could be explained bythe reworking of pellets regurgitated by birds of prey at the time of the filling. Taking intoaccount this species, the biochronological age of these deposits could be from late MiddlePleistocene to Holocene (Chaimanee et al., 1993; Chaimanee, 1998).
Regarding Pleistocene Vietnamese sites, these new data are important because of the scar-city of rodents in faunal lists. The aim of this brief article is threefold. First, we describe thetwo fossiliferous localities (Ma U’Oi and Tan Vinh) in terms of sedimentology, with a focuson Tan Vinh first mentioned here. Second, the presence of isolated molars in great numbers,especially at Tan Vinh, allows a statistical comparison with those recovered in other MiddlePleistocene localities from Thailand (Ginsburg et al., 1982; Chaimanee and Jaeger, 1993;Chaimanee et al., 1993; Tougard, 1998, 2001; Esposito et al., 1998; Chaimanee, 1998). Third,we discuss implications of N. fulvescens, N. andersoni and L. sabanus occurrences in thePleistocene of Vietnam in terms of biochronology and past distribution.
2. Description of Vietnamese sites
2.1. The Tan Vinh site
2.1.1. Location and geological setting
The Tan Vinh quarry is located in the Luong Son district, Hoà Binh province, about 50 kmWNW from Hanoi, in northern Vietnam (coordinates: 20°51′38″N–105°30′02″E) (Fig. 1).Many quarries are situated along the right border of the Red River delta in the first escarpmentthat dominates the river plain. The Tan Vinh quarries, several metres high, contain fracturedgrey-dark micritic limestone attributed to the marine Triassic (Martini et al., 1998).
A dense network of caves and galleries was developed through time in this geological fra-mework (Fenart et al., 1999). Many quarries around the Tan Vinh village expose several fis-sures with fillings in the karstic network (Fig. 2). Most part of the fissures is full of brown tored-brown argillaceous material. Some fissures and cave fillings are several metres large and

Fig. 1. Location of Thai and Vietnamese sites.Fig. 1. Localisation des sites thaïlandais et vietnamiens.
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383370
usually larger toward the top of outcrops. Some of them pass laterally or vertically to cavesespecially when quarries are open in ancient natural cliffs.
2.1.2. Description of the fissure fillingThe fissure filling is composed of two kinds of material (Fig. 2). The most abundant is a
brown argillaceous pelite with often a quartzitic arenitic proportion and granule-sized Fe–Mn-rich pisoliths concretions. This material is generally poorly consolidated. This argillaceous fa-cies contains modern fauna. The second filling component (5% of the filling) is a yellow-brown carbonate breccia characterised by numerous calcitic veins. It consists of typical fossili-ferous cave breccia cemented on walls of the fissures, with a mixing of weathered clays with

Fig. 2. Scenario for the formation of the Tan Vinh deposits: A. Complete or partial filling of a fissure in the karsticnetwork with the fossiliferous cave breccia facies (Late Pleistocene); B. Erosion of the precedent filling, some relicts ofbreccia stay caught on the wall or ground of the fissure; C. Complete filling of the karst by the recent brown clay facies(Holocene).Fig. 2. Scénario simplifié illustrant la formation des dépôts du gisement de Tan Vinh : A. Remplissage complet ou partieldu réseau karstique par la brèche fossilifère (pléistocène supérieur) ; B. Érosion du remplissage précédent, seulesquelques reliques de brèche restent accrochées sur les parois ou au fond de la fissure ; C. Remplissage complet desfissures par de l’argile brune récente (holocène).
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 371
granule-sized Fe–Mn-rich pisoliths, reworked speleothems, clasts of dark limestone and fossilremains of mammals. All murid teeth come from this breccia. Some clasts of the breccia aresometimes reworked in the argillaceous facies.
2.1.3. InterpretationThe breccia is in fact a relict facies and the mode of deposit could be the following: (1)
Filling of the fissures with the fossiliferous breccia, perhaps during an uplift of the base level(Fig. 2A). (2) Opening of the fissure and clearing out of the most part of the breccia during anew water circulation phase. The cave filling is karstified in the same way as the former karstsystem (Fig. 2B). During this clearing, 95% of the material into the fissure is eroded. Somebreccia stays cemented on the wall of the fissure. (3) New filling of fissures and cave withthe brown argillaceous material produced by weathering of surrounding soils (Fig. 2C).
2.2. The Ma U’Oi site
2.2.1. Location and geological settingThe Ma U’Oi cave is situated in the Man Duc village, Tan Lac district, Hoà Binh province,
25 km SSW from Hoà Binh city, northern Vietnam (coordinates: 20°37′22″N, 105°16′40″E)(Fig. 1). Around the site, the bedrock consists of grey-dark micritic marine limestone attributed

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383372
to the Triassic (Martini et al., 1998). A dense network of caves and galleries developedthrough the time in this framework, partially driven by faults and fractures (Fenart et al.,1999).
In the neighbourhood of Man Duc village, several caves expose the karstic network and itsfossiliferous filling (Fig. 3A). The caves are partly filled with brown to red-brown argillaceousand breccia material with numerous iron pisoliths. The largest karstic fillings reach severalmetres thick.
2.2.2. Description of the filling of Ma U’Oi caveThe Ma U’Oi cave consists of two distinct corridors (A and B) composed of several rooms
(Fig. 3B). The fossiliferous facies shows a quite constant composition in all rooms. The facies,a calcaro-pelitic fossiliferous breccia, is characterised by a relative apparent monotony, a short-scale lateral and vertical extension and a variable potential of preservation. In all excavatedrooms, the fossiliferous level forms a thick layer (ranges from 0.5 to 1.5 m) covering the vault,the upper part of the walls and the floor.
All murids have been recovered in the breccia from the roof of the room A1 (in situ micro-vertebrate fauna) and from the walls of the rooms A1 and A2 (in situ mammal assemblage)(Fig. 3). U/Th datings have been performed on two samples. The first one from the upper partof the fossiliferous breccia (coarse-grained sparite from stalagmitic floor) reveals an age of193 ± 17 kyr, i.e. late Middle Pleistocene for the microvertebrate fauna. The second sample,sparite from speleothems partially covering the fossiliferous breccia, gives for filling alterationan age of 49 ± 4 kyr (i.e. late Late Pleistocene). This estimate corresponds to the minimal ageof the mammal assemblage.
Fig. 3. A. Synthetic scenario of the Ma U’Oi site showing the location of the excavated fossiliferous level and the twoalluvial terraces (Terrace 1 and 2, lying respectively at 92 and 43 m above the contemporary alluvial plain). Both ofalluvial terrace depositional units are closely associated to endokarstic deposits (fossiliferous cave breccia andspeleothems). The crosses indicate the location of the breccia from which were extracted rodents teeth, in the rooms A1(in situ microvertebrate fauna) and A2 (in situ mammal assemblage). B. Plan of Ma U’Oi cave.Fig. 3. A. Aperçu schématique du site de Ma U’Oi montrant la position du niveau fossilifère étudié, ainsi quel’association entre les dépôts endokarstiques (brèche fossilifère et spéléothèmes) et les niveaux étagés des terrassesalluviales (terrasses 1 et 2, reposant respectivement à 92 et 43 m au dessus du niveau de la plaine actuelle). Les croixindiquent la localisation de la brèche d’où ont été extraites les dents de rongeurs, dans la salle A1 (faune in situ demicrovertébrés) et dans la salle A2 (assemblage in situ de mammifères). B. Plan de la grotte de Ma U’Oi.

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 373
2.2.3. InterpretationThe present-day disposition of the breccia up to the vault of the cave seems to indicate first
a quite complete filling of the cave by sediments (at least 3 or 4 m thick) before being partiallyeroded. The outcropping of the breccia (i.e. thin relicts on walls, roof or ground) suggests atleast one episode of removing of karstic sediments since first deposit of the breccia (i.e. fossi-liferous facies). Such features are now well-known in karstic environment (Bacon et al., 2004)and suggest that karst filling is the result of several episodes of sedimentation interrupted bynon-deposition and erosion periods (Roussé et al., 2003).
Inside the breccia, close imbrication of clastic materials (limestone, Fe–Mn-rich pisoliths,speleothems) with bones and teeth, attest to a sedimentological origin for the breccia (down-stream water circulations inside the caves).
General aspects of the fossiliferous breccia and relations between endokarstic (activity in-side the cave) and exokarstic (alluvial terraces) environments are described in Roussé et al.(2003); Bacon et al. (2004).
3. Molars description of L. sabanus and N. fulvescens
3.1. Material and methods
At Tan Vinh, we collected several kilos of breccia and dissolved them with formic acid(10%). After washing the sediment with running water and drying, we sorted it using a sieveof 2 mm. At Ma U’Oi, the breccia was crushed in small fragments, washed in the water of thenearby river, dried and screened with same sieves. We used a binocular and a digital counterfor measuring teeth. Two dimensions have been taken: maximal length (mesiodistal) and max-imal width (buccolingual). A statistical analysis has been realised on raw data by using rangeof variation, mean and standard deviation (Tables 1 and 2). Microphotographs of isolated teethwere realised with the Scanning Electron Microscope (JEOL JSM-25SIII). Teeth have notbeen metallised.
Fossil data (description, statistical parameters) of rodents from Thai localities are takenfrom Chaimanee (1998) and compiled in Tables 1 and 2 (raw data are not available). Theyconcern localities of Snake cave (main and upper layers), Kanchanaburi 2, Khao Toi 1 andKhao Rupchang. Data of modern species are also taken from this reference, but their origin isunknown.
3.2. Results
We attributed 14 molars and two fragmentary mandibles (one with m1–m2 and one withm1) from Ma U’Oi and all of the isolated 104 molars from Tan Vinh to L. sabanus on thebasis of descriptions of comparable fossil specimens in Chaimanee (1998). Upper and lowermolars are large (Fig. 4). In M1 and M2, central cusps are larger than labial ones. We foundthe same features as those described in Chaimanee (1998) (p. 115): On the first row (of M1),cusp t1 is separated from t2 by a deep groove. Cusp t3 is reduced and fused with t2. On thesecond row, cusp t4 and t6 are more distal than the central cusp t4 is separated from t5 by adeep groove. On the third row, cusp t8 is large (…). There is no posterior cingulum. In M2,the cusp t3 is absent and the cusp t4 is rounded and separated from t5 by a deep groove. Lin-

Table 1
Measurements (length and width) of molars of modern and fossil L. sabanus. Measurements from Ma U’Oi and TanVinh have been taken by the authors; those from Snake cave and Kanchanaburi 2 are from Chaimanee (1998). Livingspecimens are also compiled in Chaimanee (1998) but their origin is unknown. N = number of specimens; x = mean;S.D. = standard deviation (only for sample sizes superior or equal to 10)Tableau 1Mesures (longueurs et largeurs) des molaires de L. sabanus fossile et actuelle. Les mesures des spécimens de Ma U’Oi etde Tan Vinh ont été prises par les auteurs ; celles de Snake cave et de Kanchanaburi 2 sont extraites du travail deChaimanee (1998). Les mesures des spécimens actuels proviennent également de Chaimanee (1998) mais leur origineest inconnue. N = nombre de molaires ; x = moyenne ; S.D. = Ecart-type (uniquement pour les effectifs supérieurs ouégaux à 10)
Localités N Length (mm) Width (mm)R x S.D. R x S.D.
M1 Ma U’Oi 4 4.52–5.05 4.85 – 3.06–3.62 3.21 –Tan Vinh 16 4.21–5.36 4.92 0.28 2.85–3.31 3.09 0.13SC main layer 36 4.29–5.18 4.74 0.25 2.56–3.00 2.79 0.12SC upper layer 11 4.21–4.82 4.49 0.22 2.49–2.79 2.65 0.08Kanchanaburi 2 2 4.12–4.40 4.26 – 2.53–2.61 2.61 –Living 2 4.63–5.20 4.92 – 2.72–3.00 2.86 –
M2 Ma U’Oi 4 3.74–4.26 3.98 – 2.88–3.04 2.93 –Tan Vinh 19 2.92–4.55 3.78 0.35 2.77–3.23 2.95 0.12SC main layer 24 2.98–3.62 3.37 0.16 2.48–2.90 2.63 0.10SC upper layer 10 3.11–3.58 3.33 0.16 2.29–2.96 2.55 0.06Kanchanaburi 2 3 2.75–3.06 2.87 – 2.39–2.70 2.51 –Living 3 3.24–3.80 3.45 – 2.74–3.27 2.99 –
M3 Tan Vinh 3 2.33–2.69 2.48 – 2.29–2.47 2.36 –m1 Ma U’Oi 6 3.99–4.63 4.33 – 2.49–2.99 2.70 –
Tan Vinh 29 4.07–4.69 4.39 0.17 2.53–3.29 2.77 0.16SC main layer 27 3.45–4.00 3.77 0.18 2.11–2.72 2.41 0.15SC upper layer 5 3.68–4.04 3.85 – 2.35–2.58 2.43 –Living 3 3.89–4.04 3.96 – 2.20–2.84 2.52 –
m2 Ma U’Oi 4 3.17–3.73 3.50 – 2.76–3.18 2.98 –Tan Vinh 30 3.08–3.93 3.46 0.21 2.49–3.41 3.07 0.48SC main layer 27 2.61–3.49 2.96 0.18 2.39–2.82 2.62 0.11SC upper layer 10 2.78–3.16 2.97 0.13 2.46–2.87 2.64 0.11Living 3 2.93–3.22 3.09 – 2.18–2.93 2.59 –
m3 Tan Vinh 7 2.43–3.04 2.9 – 2.26–2.75 2.51 –
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383374
gual cusps of m1 are more mesial than labial cusps. The second and third rows are chevron-shaped. Cv5 is present on m1 and m2.
Among the microvertebrate fauna from Ma U’Oi, 12 isolated teeth and two mandibles withm1–m2 are affiliated to N. fulvescens (Fig. 5). The morphological differences with L. sabanusare tenuous. However, concerning the size, there is no overlap between the two species,N. fulvescens being markedly smaller than L. sabanus (Tables 1 and 2). We affiliated thesesmall teeth to N. fulvescens on the basis of description of Chaimanee (1998) (p. 122). In short,M1, long and narrow, display central cusps larger than labial and lingual ones. In unworn M1,the lingual cusps t1 and t4 are clearly separated from the central cusps. The second row ischevron-shaped and the third row is characterised by an enlarged cusp t8 connected to areduced cusp t9. In M2, the cusps t1 and t8 are rounded and large and t9 is absent. m1 aresmall and narrow with lingual cusps more mesial than labial cusps. The second and third rowsof cusps show chevron-shaped laminae. m2 are slightly larger than m1 (Table 2). The presenceof Cv5 is variable on m1 and m2.

Table 2
Measurements (length and width) of molars of modern and fossil N. fulvescens. Measurements from Ma U’Oi and TanVinh have been taken by the authors; those from Snake cave, Khao Toi 1, Khao Rupchang are from Chaimanee (1998).Living specimens are also compiled in Chaimanee (1998) but their origin is unknown. N = number of specimens;x = mean; S.D. = standard deviation (only for sample sizes superior or equal to 10)Tableau 2Mesures (longueurs et largeurs) des molaires de N. fulvescens fossile et actuelle. Les mesures des spécimens de Ma U’Oiet Tan Vinh ont été prises par les auteurs ; celles de Snake cave, Khao Toi 1, Khao Rupchang sont extraites du travail deChaimanee (1998). Les mesures des spécimens actuels ont été trouvées également chez Chaimanee (1998) mais leurorigine est inconnue. N = nombre de spécimens ; x = moyenne ; S.D. = Écart-type (uniquement pour les effectifssupérieurs ou égaux à 10)
Localités N Length (mm) Width (mm)R x S.D. R x S.D.
M1 Ma U’Oi 2 3.31–3.62 3.46 – 2.03–2.05 2.05 –SC main layer 47 2.99–3.50 3.20 0.12 1.56–1.85 1.71 0.07SC upper layer 17 3.04–3.34 3.15 0.09 1.54–1.74 1.65 0.06Khao Toi 1 4 2.83–3.25 3.06 – 1.67–1.84 1.76 –Khao Rupchang 3 2.89–3.06 2.95 – 1.57–1.61 1.60 –Living 7 3.13–3.46 3.20 – 1.59–1.77 1.68 –
M2 Ma U’Oi 2 2.19–2.75 2.47 – 1.62–2.29 1.95 –SC main layer 17 1.98–2.35 2.17 0.09 1.50–1.82 1.68 0.10SC upper layer 3 2.04–2.18 2.12 – 1.55–1.70 1.63 –Khao Toi 1 3 1.96–2.13 2.03 – 1.40–1.52 1.45 –Khao Rupchang 3 2.05–2.12 2.09 – 1.52–1.63 1.58 –Living 8 1.99–2.33 2.11 – 1.52–1.69 1.64 –
m1 Ma U’Oi 5 2.63–2.96 2.81 – 1.57–1.85 1.72 –SC main layer 20 2.40–2.76 2.59 0.08 1.34–1.71 1.51 0.09SC upper layer 9 2.53–2.72 2.61 – 1.44–1.63 1.50 –Living 7 2.38–2.71 2.54 – 1.39–1.73 1.55 –
m2 Ma U’Oi 7 1.75–2.44 2.05 – 1.65–1.99 1.74 –SC main layer 29 1.77 2.18 1.99 0.11 1.53–1.76 1.64 0.06SC upper layer 5 1.98–2.23 2.11 – 1.61–1.77 1.70 –Living 5 1.88–2.14 1.98 – 1.50–1.65 1.60 –
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 375
We report ranges of variation, means and standard deviations of molars dimensions ofL. sabanus (Table 1) and N. fulvescens (Table 2) found in Vietnamese (Ma U’Oi and TanVinh) and Thai sites (Snake cave (SC) upper layer and main layer, Kanchanaburi 2, KhaoToi 1, Khao Rupchang). Except the sample size of Tan Vinh and Snake Cave (superior to 15specimens), all other data (modern and fossil species) are only represented by small samples(less than three specimens), which limit the meaning of statistical parameters and their compar-ison.
Nevertheless, we can notice some variations between L. sabanus of different localities. Allspecimens considered, upper and lower molars from Vietnamese sites (especially Tan Vinh)tend to be slightly greater than those from the Thai site Snake cave (data from Kanchanaburiare too sporadic) for both dimensions (Fig. 6). Concerning N. fulvescens, molars from MaU’Oi are also slightly greater than those from Thai sites, but samples are small.
If we consider the two late Middle Pleistocene localities, Ma U’Oi (193 ± 17 kyr) andSnake Cave (169 ± 15 kyr, Esposito et al., 1998; Chaimanee, 1998), these small size differ-ences between individuals of same species could be interpreted as a cline. Indeed, these twosites are clearly located at different latitudes, the northern site Ma U’Oi (and also the closeTan Vinh) and the southern site (Snake Cave) being around 1000 kilometres apart. Thus, we
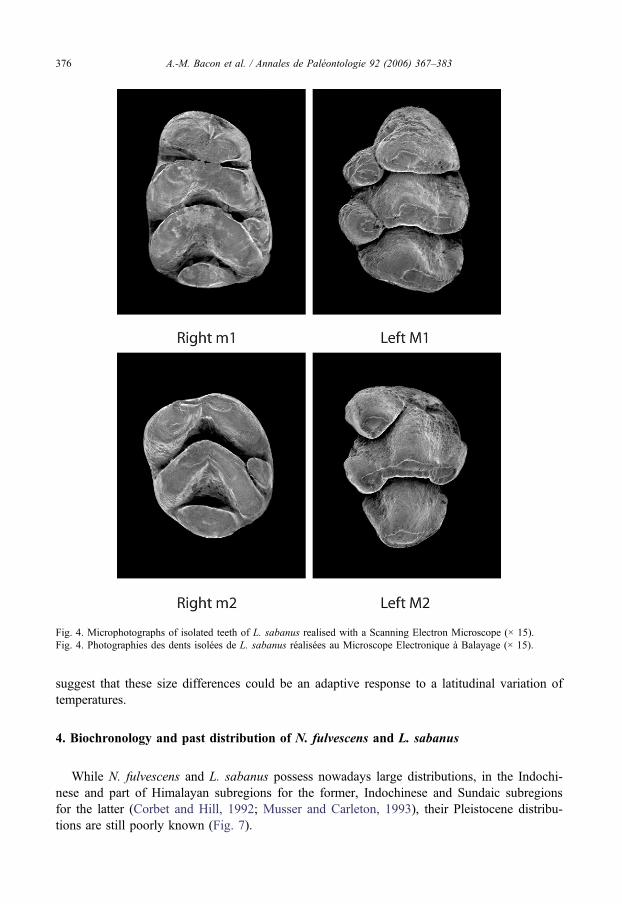
Fig. 4. Microphotographs of isolated teeth of L. sabanus realised with a Scanning Electron Microscope (× 15).Fig. 4. Photographies des dents isolées de L. sabanus réalisées au Microscope Electronique à Balayage (× 15).
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383376
suggest that these size differences could be an adaptive response to a latitudinal variation oftemperatures.
4. Biochronology and past distribution of N. fulvescens and L. sabanus
While N. fulvescens and L. sabanus possess nowadays large distributions, in the Indochi-nese and part of Himalayan subregions for the former, Indochinese and Sundaic subregionsfor the latter (Corbet and Hill, 1992; Musser and Carleton, 1993), their Pleistocene distribu-tions are still poorly known (Fig. 7).

Fig. 5. Microphotographs of isolated teeth of N. fulvescens realised with a Scanning Electron Microscope (× 20).Fig. 5. Photographie des dents isolées de N. fulvescens réalisées au microscope électronique à balayage (× 20).
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 377
Concerning Vietnam, the findings from Tan Vinh (late Middle Pleistocene to Holocene)and Ma U’Oi (late Middle Pleistocene) bring new data about this past distribution. Indeed,L. sabanus had never been found so far north in the Indochinese subregion and also so far inthe Pleistocene of Vietnam (Tham Kuyen and Keo Leng produced two unidentified speciesRattus sp. and Mus sp. (Cuong, 1985; Olsen and Ciochon, 1990) (Table 3). Only, the LangTrang fauna (cave II, breccia 5) dated to Late Pleistocene yielded the species Rattus sabanus(Long et al., 1996) (Fig. 7B). Concerning N. fulvescens, its presence at Ma U’Oi is the firstever mentioned in the Pleistocene of Vietnam. However, its presence so far north in the Indo-chinese area is not new as it has been already listed at Baiyanjiao, Yanhui and Washuwan insouthern China for the Middle Pleistocene (Zheng, 1993) (Fig. 7A).

Fig. 6. Graphics showing statistical results of Table 1 (mean and variation of each dimension (L: length, W: width) calculated for molars of L. sabanus from northernVietnamese sites (Ma U’Oi and Tan Vinh) and the southern Thai site (Snake Cave, main layer (SCml) and upper layer (SCul)). Means of each group are linked with a line for abetter comparison.Fig. 6. Graphique montrant les résultats statistiques du tableau 1 (moyenne et intervalle de variation pour chaque dimension (L : longueur, W : largeur) réalisés sur les molairesde L. sabanus provenant des sites vietnamiens au nord (Ma U’Oi et Tan Vinh) et du site thailandais localisé plus au sud (Snake Cave, main layer (SCml) et upper layer (SCul)).Les moyennes de chaque groupe sont reliées par un trait pour une meilleure comparaison.
A.-M
.Bacon
etal.
/Annales
dePaléontologie
92(2006)
367–383
378

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 379
Outside of Vietnam, the only well documented murid assemblages come from Pliocene toHolocene sites of Thailand (Chaimanee, 1998). This author reported the presence ofL. sabanus in Thai sites dated from Middle Pleistocene to Holocene (Fig. 7). Among all mur-ids found at Snake cave [fauna dated to late Middle Pleistocene (Tougard, 1998, 2001) and to169 ± 15 kyr by U/Th method (Esposito et al., 1998)], L. sabanus is rather abundant (withN. fulvescens, Rattus sikkimensis and R. rattus) (Chaimanee, 1998), that is also confirmed byMa U’Oi and Tan Vinh data. It is also present in other sites in Central and Peninsular Thai-land, but the data are sporadic (Chaimanee, 1998). This author also showed that N. fulvescenswas present southwards into the Malay peninsula since the Middle Pleistocene (Fig. 7, BanNasan, Khao Toi 1 and Khao Rupchang sites). However, the reconstruction of past distributionof these species varies in accordance with systematics of the modern taxa (Lekagul andMcNeely, 1977; Corbet and Hill, 1992; Nowak, 1999). Indeed, if modern N. fulvescens andN. bukit are considered as separated species (as represented in Fig. 7), the record of this fossilspecies behind the limit of the Isthmus of Kra is new for the Pleistocene, as it extends south-wards. On the contrary, if N. fulvescens and N. bukit are considered as conspecific andgrouped in a single species N. fulvescens with a larger distribution as far as the Sondaic sub-region (Java, Sumatra, Bali), the fossil specimens do not question the past distribution of thetaxon (see Chaimanee (1998) for exhaustive references about this question).
No fossil records of L. sabanus and N. fulvescens older than the Middle Pleistocene havebeen listed in Southeast Asia, but the oldest occurrence of the genus Leopoldamys(L. minutus n. sp.) is mentioned at Saraburi in Thailand, a site dated to Late Pliocene (Chaima-nee, 1998) and both genera Leopoldamys and Niviventer are present in the Indochinese area atthe Longgupo locality, dated to the Early Pleistocene (Zheng, 1993). According to a recentphylogeographical analysis based on mitochondrial DNA (Gorog et al., 2004), L. sabanusmay have dispersed throughout its current Indochinese and Sundaic areas in the early Pliocene“before the long-standing connections between Indochina, Java, Sumatra, Borneo and the Ma-lay Peninsula were severed” (Gorog et al., 2004: p. 103), suggesting a vicariance history ratherthan a dispersal. The fossil record tends to reflect a similar conclusion (Chaimanee, 1998).
N. andersoni is an endemic species present only in limited Chinese areas, from East Tibetto southern and central provinces (Yunnan, Sichuan, South Gansu and Shaanxi) (Corbet andHill, 1992; Musser and Carleton, 1993). Until now, it was only mentioned in different ChinesePleistocene localities, from the limit Pliocene/Pleistocene to the beginning of the Holocene, be-tween 1.8 myr and 10 kyr (Zheng, 1993). So, the Vietnamese record of N. andersoni at MaU’Oi is the first mention of this species in Quaternary deposits outside China (Bacon et al.,2004). It attests of a southernmore distribution of this species in the Indochinese subregion inthe past.
5. Contribution to reconstruction of past environments
The murids record of Ma U’Oi is far for being exhaustive and their interest in palaeoenvir-onmental reconstruction can only be succinct. Among the in situ mammal fauna, three muridsare listed (Bacon et al., 2004). L. sabanus is a large-bodied arboreal rat, still today largelywidespread in Southeast Asia, Indochinese (Vietnam, Cambodia, Laos, Thailand and Bangla-desh) and Sundaic subregions (Java, Malay Peninsula, Sumatra, Borneo and many smallislands) (Medway, 1972; Musser, 1982; Corbet and Hill, 1992; Van der Meulen and Musser,1999). It is adapted to all types of forests and is common in lowlands and foothills of ever-

Fig. 7. Current distributions of N. fulvescens and L. sabanus represented by grey and N. andersoni represented bytriangles from Corbet and Hill (1992). The numbers correspond to Pleistocene localities (Bacon et al., 2004; Chaimanee,1998; Zheng, 1993; Long et al., 1996). Map A: 1-Ma U’Oi (late Middle Pleistocene), 2-Snake cave (late MiddlePleistocene), 3-Kanchanaburi 2 (Middle Pleistocene), 4-Khao Songto, 5-Ban Nasan (Middle Pleistocene), 6-Khao Toi 1(Middle Pleistocene), 7-Khao Rupchang (Middle Pleistocene), 8-Baiyanjiao (Middle Pleistocene). Map B: 1-Ma U’Oi(late Middle Pleistocene), 2-Tan Vinh, 3-Lang Trang (Late Pleistocene), 4-Snake cave (three localities, late MiddlePleistocene), 5-Kanchanaburi 1 (Holocene) et Kanchanaburi 2 (Middle Pleistocene), 6-Khao Tinpet (MiddlePleistocene), 7-Khao Toi 1 (Middle Pleistocene).Fig. 7. Distributions géographiques actuelles de N. fulvescens et L. sabanus représentées en grisé, et de N. andersonireprésentée par des triangles d’après Corbet et Hill (1992). Les chiffres correspondent aux localités pléistocènes (Baconet al., 2004 ; Chaimanee, 1998 ; Zheng, 1993 ; Long et al., 1996). Carte A : 1-Ma U’Oi (Pléistocène moyen récent),
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383380

Table 3
List of murids in Pleistocene Vietnamese (Cuong, 1985; Bacon et al., 2004, 2006) and Thai sites (Chaimanee, 1998).Snake cave (ML: Main Layer; UP: Upper Layer); K2: Kanchanaburi 2Tableau 3Listes des Muridae découverts dans les sites pléistocènes vietnamiens (Cuong, 1985 ; Bacon et al., 2004, 2006) etthaïlandais (Chaimanee, 1998). Snake cave (ML : Main Layer ; UP : Upper Layer) ; K2 : Kanchanaburi 2
Taxa Ma Tan Tham Keo Snake Snake K 2U’Oi Vinh Kuyen Leng Cave ML cave UL
Rattus sp. X X XR. sikkimensis X X XR. rattus XMus sp. X ?M. shortridgei X X XM. pahari X XM. cooki X XM. cervicolor X X XVandeleuria oleracea X XHapalomys delacouri X X XChiropodomys gliroides X XBandicota savilei XB. indica X XHadromys humei X X XBerylmys berdmorei XMaxomys surifer X XNiviventer fulvescens X X X XN. andersoni XN. gracilis XLeopoldamys sabanus X X X X X
A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 381
green forests (Corbet and Hill, 1992). So, it does not provide any pertinent environmentalinformation, as it is present in northern seasonal forest as well in southern rain forest.N. fulvescens occurs in various kinds of forests in both lowlands and mountains in the northof the Isthmus of Kra as far as the Himalayan forest zone, following also the eastern boundaryof the Palaearctic Region (Corbet and Hill, 1992). It seems adapted to a more seasonal climateor to a climate with other seasonality. The presence of N. andersoni at Ma U’Oi is more infor-mative. According to Musser and Chiu (1979), “Both andersoni and excelsior inhabit the highmountains along the eastern edge of the Tibetan Plateau and the Himalayas” and, farther,“Examples of andersoni have been collected from elevations ranging from 6000 to 10,000 ft”(2000 to 3300 m). Corbet and Hill (1992) confirm that N. andersoni can be observed at alti-tudes of 1800–3000 m. The presence of this murid at Ma U’Oi which attests a more southerndistribution of this species in the past, also suggests a more southern boundary of montane for-est vegetation with drier conditions (and that before 49 ± 4 kyr, the minimal age of the fauna).It also reflects that the montane vegetation zone was probably lower in altitude due to coolertemperatures. This seems to be in accordance with palaeoenvironmental reconstruction (Vander Kaars and Dam, 1995), which show a marked climatic change between 74,000 yrs andthe last glacial peak around 20,000 yrs with drier conditions.
2-Snake cave (pléistocène moyen récent), 3-Kanchanaburi 2 (pléistocène moyen), 4-Khao Songto, 5-Ban Nasan(pléistocène moyen), 6-Khao Toi 1 (pléistocène moyen), 7-Khao Rupchang (pléistocène moyen), 8-Baiyanjiao(pléistocène moyen). Carte B : 1-Ma U’Oi (pléistocène moyen), 2-Tan Vinh, 3-Lang Trang (pléistocène supérieur),4-Snake cave (trois localités du pléistocène moyen), 5-Kanchanaburi 1 (holocène) et Kanchanaburi 2 (pléistocènemoyen), 6-Khao Tinpet (pléistocène moyen), 7-Khao Toi 1 (pléistocène moyen).

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383382
The microvertebrate in situ fauna at Ma U’Oi seems to indicate a more forested environ-ment, not contradictory with the late Middle Pleistocene (a period corresponding to a shortinterglacial episode). N. fulvescens and L. sabanus are not the only rodents present in thislocality. The third murid rodent listed at Ma U’Oi H. delacouri is a specialised arboreal ratwhich inhabits today tropical forest and bamboo forest in small zones of the Indochinese area(Central Vietnam, Northern Laos, Hainan) Corbet and Hill (1992). The fauna is also composedof sciurid rodents with three taxa of flying squirrels Hylopetes phayrei, Hylopetes sp., and aPetauristinae indet., which reflect evergreen forest habitat. Shrews were present at Ma U’Oiwith Crocidura sp. which inhabits lowland and montane forests. This record can be seen asan “impoverished” version of the microvertebrate fauna found at Snake cave (Ginsburg et al.,1982; Chaimanee et al., 1993; Chaimanee, 1998), the Ma U’Oi species (Bacon et al., 2004,2006), especially flying squirrels, suggesting a comparable habitat with the presence of ever-green forest.
6. Conclusion
These original data from Ma U’Oi and Tan Vinh bring the first information concerningrecord and past distribution of the two murids L. sabanus and N. fulvescens in Vietnam. Thecomparison of molar dimensions between rodents of these Vietnamese sites and those of thesouthernmore thai site (Snake Cave) emphasizes the possibility of clines. However, we cannot go deeper in the discussion as the sample size for each species has to be increased.
Acknowledgments
The authors want to present their gratitude to people who gave them the possibility to un-dertake these fieldworks in Vietnam: Bui Thi Xuan, Bui Van Hop and Quach Van Ach, QuachDinh Thi from the Hoà Binh Museum, Bui Giang Huong, Bui Manh Hung and Bui Van Khaifrom the Commune Department of Culture and Mrs. Tran Thi Lan Anh from the NationalCentre for Social Sciences and Humanities (CNSSH). Thanks to C. Falguères for datings (In-stitut de Paléontologie humaine, Paris); A. Gorog, J. de Vos, S. Sen and the referee M. Mi-chaux for correction of the manuscript; D. Fouchier (UPR 2147) and A. Bouzeghaia (Instituteof Geology, Strasbourg) who realised drawings; C. Chancogne (National Museum of NaturalHistory, Paris) for SEM photographs. This mission was financed by the Collège de France(Chaire de Paléoanthropologie et de Préhistoire); UPR 2147 of CNRS; DRI of CNRS (projectsno. 12746, no. 13669); the Earth Sciences Doctoral School of Louis Pasteur University (F.Beck and M. Cara), Strasbourg; the Department of Anatomy and Anthropology, Tohoku Uni-versity School of Medicine, Sendai, Japan; the Department of Anatomy, Sapporo Medical Uni-versity, Japan.
References
Bacon, A.-M., Demeter, F., Roussé, S., Long, V.T., Duringer, P., Antoine, P.-O., Thuy, N.K., Huong, N.T.M., Dodo,Y., Matsumura, H., Schuster, M., Anezaki, T., 2006. New palaeontological assemblage, sedimentological andchronological data from the Pleistocene Ma U’Oi cave (Northern Vietnam). Palaeogeography, Palaeoclimatology,Palaeoecology 230, 280–298.
Bacon, A.M., Demeter, F., Schuster, M., Long, V.T., Thuy, N.K., Antoine, P.O., Sen, S., Nga, H.H., Huong, N.T.M.,2004. The Pleistocene Ma U’Oi cave, northern Vietnam: palaeontology, sedimentology and palaeoenvironments.Geobios 37, 305–314.

A.-M. Bacon et al. / Annales de Paléontologie 92 (2006) 367–383 383
Chaimanee, Y., 1998. Plio–Pleistocene rodents of Thailand. Thai studies in Biodiversity 3, 1–103.Chaimanee, Y., Jaeger, J.J., 1993. Pleistocene mammals of Thailand and their use in the reconstruction of the pa-
leoenvironments of Southeast Asia. SPAFA Journal 3, 4–10.Chaimanee, Y., Jaeger, J.J., Suteethorn, V., 1993. Pleistocene microvertebrates from fissure-fillings in Thailand. Jour-
nal of Southeast Asian Earth Sciences 8, 45–48.Corbet, G.B., Hill, J.E., 1992. The mammals of the Indomalayan region. Natural History Museum Publications, Ox-
ford University Press, New York.Cuong, N.L., 1985. Fossile Menschenfunde aus Nordvietnam. In: Herrmann, J., Ullrich, H. (Eds.), Menschwerdung—
Biotischer und gesellschaftlicher Entwicklungsprozess. Akademieverlag, Berlin, pp. 96–102.Demeter, F., Bacon, A.M., Thuy, N.K., Long, V.T., Matsumura, H., Nga, H.H., Schuster, M., Huong, N.T.M., Cop-
pens, Y., 2004. An archaic Homo molar from northern Vietnam. Current Anthropology 45, 535–541.Esposito, M., Chaimanee, Y., Jaeger, J.J., Reyss, J.L., 1998. Datation des concrétions carbonatées de la « Grotte du
serpent » (Thaïlande) par la méthode Th/U. Comptes Rendus de l’Académie des Sciences de Paris 326, 603–608.Fenart, P., Cat, N.N., Drogue, C., Canh, D.V., Pistre, S., 1999. Influence of tectonics on the morphogenesis of the
peak karst of Halong Bay, Vietnam. Geodinamica Acta 12, 193–200.Ginsburg, L., Ingavat, R., Sen, S., 1982. A Middle Pleistocene (Loagian) cave fauna in Northern Thailand. Comptes
Rendus de l’Académie des Sciences de Paris 294, 295–297.Gorog, A.J., Sinaga, M.H., Engstrom, M.D., 2004. Vicariance or dispersal? Historical biogeography of three Sunda
shelf murine rodents (Maxomys surifer, Leopoldamys sabanus and Maxomys whiteheadi). Biological Journal ofLinnean Society 81, 91–109.
Lekagul, B., McNeely, J.A., 1977. Mammals of Thailand, first ed. Association for the conservation of wildlife, Bang-kok.
Long, V.T., de Vos, J., Ciochon, R.S., 1996. The fossil mammal fauna of the Lang Trang caves, Vietnam, comparedwith Southeast Asian fossil and recent mammal faunas: the geographical implications. Bulletin of the indo-pacificPrehistory Association 14, 101–109.
Martini, R., Zaninetti, L., Cornée, J.J., Villeneuve, M., Tran, N., Thang Ta, T., 1998. Occurrence of Triassic forami-nifers in carbonate deposits from the Ninh Binh Area (North Vietnam). Comptes Rendus de l’Académie desSciences de Paris 326, 113–119.
Medway, L., 1972. Niah Cave bone VII: size changes in the teeth of two rats, Rattus sabanus Thomas andR. muelleri Jentink. Sarawak Museum Journal 11, 616–623.
Musser, G.G., 1982. The Trinil rats. Modern Quaternary Research SE Asia 7, 65–85.Musser, G.G., Carleton, M.D., 1993. Family Muridae. In: Wilson, D.E., Reeder, D.M. (Eds.), Mammal Species of the
World. A Taxonomic and Geographic Reference. Smithsonian Institution Press, Washington, pp. 501–755.Musser, G.G., Chiu, S., 1979. Notes on taxonomy of Rattus andersoni and R. excelsior, murid endemic to western
China. Journal of Mammalogy 60, 581–592.Nowak, R.M., 1999. Walker’s Mammals of the World. Johns Hopkins University Press, Baltimore and London (6e
Edition volume II, 837–1936).Olsen, J.W., Ciochon, R.L., 1990. A review of evidence for postulated Middle Pleistocene occupations in Viet Nam.
Journal of Human Evolution 19, 761–788.Roussé, S., Bacon, A.M., Demeter, F., Duringer, P., Schuster, M., Long, V.T., Thuy, N.K., Huong, N.T.M., 2003.
Dynamique des « creusements-colmatages » des réseaux karstiques du Pléistocène nord-vietnamien. Approche sé-dimentologique et implications tectono-climatiques. 9th French Congress on Sedimentology, Abstracts book,2003. ASF Publication, Bordeaux (38, 438–440).
Tougard, C., 1998. Les faunes de grands mammifères du pléistocène moyen terminal de Thaïlande dans leur cadrephylogénétique, paléoécologique et biochronologique. Thèse de Doctorat. Université de Montpellier II, Montpel-lier.
Tougard, C., 2001. Biogeography and migration routes of large mammal faunas in South-East Asia during the lateMiddle Pleistocene: focus on the fossil and extant faunas from Thailand. Palaeogeography, Palaeoclimatology, Pa-laeoecology 168, 337–358.
Van Der Kaars, W.A., Dam, M.A.C., 1995. A 135,000-year record of vegetational and climatic change from the Ban-dung area, West Java, Indonesia. Palaeogeography, Palaeoclimatology, Palaeoecology 177, 55–72.
Van Der Meulen, A.J., Musser, G.G., 1999. New paleontological data from the continental Plio–Pleistocene of Java.In: Reumer, J.W.F., de Vos, J. (Eds.), Elephants have a Snorkel! Papers in Honor of Paul Sondaar, vol. 7. DEIN-SEA, Rotterdam, pp. 361–368.
Zheng, S., 1993. Quaternary Rodents of Sichuan-Guizhou Area, China. Science Press.




















