
Past plant use in Jordan as revealed by archaeological and ethnoarchaeological phytolith signatures
Upload
stcloudstateCategory
view
1download
0
Reconstruction of the late Pleistocene grassland ofthe Columbia basin, Washington, USA, based on
phytolith records in loess
Mikhail Blinnikov a;*, Alan Busacca b, Cathy Whitlock c
a Department of Geography, St. Cloud State University, St. Cloud, MN 56301-4498, USAb Department of Crop and Soil Science, Washington State University, Pullman, WA 99164-6420, USA
c Department of Geography, University of Oregon, Eugene, OR 97403-1251, USA
Accepted 16 August 2001
Abstract
Silica phytoliths preserved in three loess sections in southeastern Washington State revealed a 100 000-year historyof the Columbia Basin grassland. Changes in the proportion of different morphotypes indicate large shifts in vegetationcomposition during the last 100 ka. A low-elevation section (677 m asl) near the center of the basin provided a record ofalternating xeric Festuca^Poa and mesic Festuca^Koeleria grassland. The middle-slope section (1095 m asl) supportedPicea^Abies or Pinus ponderosa forest or non-analog parkland at different times. Some trees were present at or near thesite even during the Last Glacial Maximum. The highest site (1220 m asl) supported Stipa-, Festuca- andPoa-dominated grassland with some Artemisia shrub during most of the late Pleistocene, but supports a coniferousforest today. Variations in vegetation can be explained as a response to changes in large-scale climatic controls.Grasslands and shrub steppe were apparently more widespread and forests more restricted than today during themarine isotope stages 2 and 4, probably as a result of cooler and drier conditions. The three new records are wellcorrelated with previously published paleo-reconstructions based on phytolith, cicada burrow and stable isotope datafrom a nearby KP-1 loess section, Carp Lake pollen record, and global ice volume variations. ß 2002 Elsevier ScienceB.V. All rights reserved.
Keywords: Columbia Plateau; grasslands; modern analogs; paleoecology; phytoliths; upper Pleistocene
1. Introduction
The nature and causes of terrestrial environ-mental change on di¡erent spatial and temporalscales during the late Pleistocene remain poorlyknown at the regional level. In the Paci¢c North-
west, terrestrial data are progressively scarcer aswe move back in history beyond the last 15 ka.Existing environmental reconstructions of thePaci¢c Northwest (Barnosky et al., 1987; Heusserand King, 1988; Whitlock, 1992; Thompson etal., 1993) are based mostly on pollen records ob-tained from lakes in forested areas west of theCascade Range, while the dry interior of the re-gion has received little attention due to lack ofsuch sites.
0031-0182 / 02 / $ ^ see front matter ß 2002 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 1 - 0 1 8 2 ( 0 1 ) 0 0 3 5 3 - 4
* Corresponding author. Fax: +1-320-529-1660.E-mail address: [email protected] (M. Blinnikov).
PALAEO 2714 22-1-02
Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101
www.elsevier.com/locate/palaeo

Vegetation history of the eastern part of theColumbia Basin Physiographic Province (Frank-lin and Dyrness, 1988) remains poorly studied.Prior to Euro-American settlement in the 19thcentury, the area supported extensive `Palousegrassland', a distinct endangered grassland typeamong eight recognized in North America (Coup-land, 1992). The grassland was established no lat-er than by the mid-Pliocene, 7^8 million yearsago, probably in response to increasing ariditycreated by the rain shadow e¡ect of the risingCascade Range (Leopold and Denton, 1987).Most of the Columbia Basin remained unglaci-ated during the late Pleistocene, when the Cordil-leran Ice Sheet existed 300 km to the north andnumerous ice caps covered the surroundingmountains. The history of the area in the latePleistocene is known primarily from the geologi-cal record (e.g., Busacca, 1989; Baker et al.,1991), but the character and spatial extent of veg-etation types in the Columbia Basin at that timeremain unknown.
Carp Lake on the western margin of the Co-lumbia Basin at 714 m asl provides a single pollenrecord spanning 130 ka to present (Barnosky,1985a; Whitlock and Bartlein, 1997). Other pollenrecords from the area are considerably shorter,spanning less than 13 ka (Mack et al., 1976,1978a,b,c; Mehringer, 1985). According to theCarp Lake record, the vegetation of the westernColumbia Basin underwent signi¢cant changeduring the last glacial^interglacial cycle, from aclosed forest during the previous interglacial priorto 116 ka BP (calendar years) to an open coldsteppe during the Last Glacial Maximum(LGM) at 21 ka BP. Periods of intermediate con-ditions were characterized as intervals of openforest, parkland, or forest steppe at Carp Lake(Whitlock and Bartlein, 1997). Shorter pollen rec-ords from the area point to the existence of anextensive Artemisia and grass community at theend of the last glacial period ca. 12^13 14C kaBP (Mack et al., 1976; Mehringer, 1985). Longrecords (s 15^20 ka BP) from other parts of thePaci¢c Northwest suggest that large-scale changesin vegetation took place during the late Pleisto-cene, for example in western Washington (Heus-ser, 1985; Heusser and Heusser, 1990), western
Oregon (Worona and Whitlock, 1995), southeast-ern Idaho (Beiswenger, 1991), and southern andcentral British Columbia (Alley et al., 1986;Clague et al., 1992). The regional changes in veg-etation are ascribed to large-scale changes in theglobal climate system caused by variations in thesize of the Laurentide ice sheet and the seasonalcycle of insolation. These continent-wide andhemispheric controls in£uenced temperature andprecipitation gradients within the Paci¢c North-west, as well as the position and strength of thejet stream and other atmospheric circulation fea-tures (COHMAP, 1988; Webb et al., 1993; Bart-lein et al., 1998).
In terms of climate, an interesting interval ofthe last glacial^interglacial cycle was the LGMat 21 ka BP, during marine isotope stage 2 (14^27 ka BP, Martinson et al., 1987). Stage 2 wasapparently the coldest interval of the last glacia-tion worldwide, followed by a rapid warmingeverywhere in the Northern Hemisphere after14 ka BP (GRIP Members, 1993). At the LGM,the Paci¢c Northwest was in£uenced by the pres-ence of the large Laurentide ice sheet over muchof present-day Canada. The ice sheet created astrong anticyclone circulation over northernNorth America, which generated surface easterlywinds at its southern margin, caused a southwardshift in the jet stream, and produced a steep tem-perature gradient south of the ice sheet (COH-MAP, 1988; Bartlein et al., 1998). Based on thepaleoclimate model results, the Paci¢c Northwestwas much colder and drier at the LGM than to-day (5^7³C colder, about 1000 mm less annualprecipitation), which is also corroborated by thepaleoecological evidence from the region (Barnos-ky et al., 1987; Thompson et al., 1993).
Pollen assemblages from Carp Lake at theLGM contain abundant Artemisia and grass pol-len and do not have a strong modern-day analog.Whitlock and Bartlein (1997) interpreted theLGM assemblages at Carp Lake as representativeof a shrub steppe persisting in very cold and dryconditions. Elsewhere, similar pollen assemblageshave been interpreted as tundra (Mack et al.,1976) or temperate steppe (Thompson, 1992),and additional research is necessary. A furthercomplication is the fact that the pollen data lack
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10178

the taxonomic resolution necessary to identify dif-ferent grass taxa, or distinguish among shrub andherb Artemisia species. Also, pollen records rep-resent the vegetation of a large area, so it is di¤-cult to reconstruct local vegetation when forestand steppe may have been both present near thesite. Application of opal phytolith analysis (Piper-no, 1988) may provide a more detailed basis for insitu vegetation reconstructions in grassland eco-systems (Kiseleva, 1982; Blinnikov, 1994; Fred-lund and Tieszen, 1997)
A study of opal phytoliths from the KP-1 loesssection provided a new long vegetation recordfrom the center of Columbia Basin at ca. 400 melevation (Blinnikov et al., 2001). Phytolith rec-ords reconstruct a more localized vegetation sig-nal than pollen, and in the Paci¢c Northwest dif-ferent taxonomic groups of grasses, sedges,conifers, and some herbs and shrubs can be iden-ti¢ed by their phytoliths (Norgren, 1973). Accord-ing to the phytolith and other data from KP-1(Blinnikov, 1999; Blinnikov et al., 2001), thelate Pleistocene vegetation was alternatively grass-land or shrub steppe, with sagebrush and dry-adapted grasses (e.g., Poa, Stipa spp.) particularlycommon during the cold dry periods. At theLGM, the present-day Agropyron^Festuca grass-land site supported scattered Artemisia shrubswith Stipa, Poa, and Festuca as the main associ-ates. The closest modern analogs for such vegeta-tion are found in the subalpine zone of the Wal-lowa Mountains at elevations s 2600 m. Tracesof cicada burrows in loess (O'Geen and Busacca,2001), soil micromorphology (Tate, 1998), andcarbon and oxygen stable isotopes from pedogen-ic carbonates (Stevenson, 1997) from KP-1 cor-roborated the interpretation based on phytolithsthat a cold-adapted shrub steppe existed at thesite during the LGM.
In the present study, we extend the coverage ofpaleoecological sites in the Columbia Basin byexamining phytoliths from three additional loesssections located at higher elevations in southeast-ern Washington State (Fig. 1; Busacca, 1989;Baker et al., 1991). Together with the previouslyexamined KP-1 section (Blinnikov et al., 2001),the sites are located along elevational andpresent-day climate gradients. The vegetation
and climate history of the Columbia Basin atthese sites the last 100 000 years is comparedwith other long paleoenvironmental recordsfrom the region. This paper elucidates the re-sponse of the regional vegetation to the variedenvironmental conditions that a¡ected the Paci¢cNorthwest in the late Pleistocene.
2. Regional setting and study sites
2.1. Modern setting
The Columbia Basin today has a temperate dryclimate as a result of the rain shadow e¡ect fromthe Cascade Range located to the west. Most pre-cipitation occurs during winter. A strong precip-itation gradient extends from northwest to south-east across the Columbia Basin, from the driestsites at the base of the eastern slopes of the Cas-cades (6 200 mm mean annual precipitation) tothe Blue Mountains (s 1000 mm mean annualprecipitation) (Franklin and Dyrness, 1988;Fig. 2). Natural vegetation in the Columbia Basinincludes forests, grasslands, and sagebrush steppe.Three forest zones are found in the northern BlueMountains: Abies lasiocarpa^Picea engelmannii(s 1500 m elevation), Pseudotsuga menziesii^Abies grandis zone (900^1500 m elevation), andPinus ponderosa zone (600^1200 m elevation)(Franklin and Dyrness, 1988). Detailed descrip-tions of the speci¢c forest associations can befound in Daubenmire (1968). Three commonnon-forested vegetation types are sagebrushsteppe (Artemisia tridentata^Poa sandbergii andArtemisia tridentata^Agropyron spicatum associa-tions) on the driest sites at low elevations(6 400 m elevation), wheatgrass-dominated grass-land (Agropyron spicatum^Poa sandbergii andAgropyron spicatum^Festuca idahoensis associa-tions) on moderately dry sites between 400 and600 m elevation, and Idaho fescue-dominatedgrassland (Festuca idahoensis^Agropyron spica-tum, Festuca idahoensis^Symphoricarpos associa-tions) on mesic sites above 600 m elevation (Dau-benmire, 1970). Alpine grassland with Festucaviridula and Stipa occidentalis and sagebrush isfound in the nearby Wallowa Mountains above
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 79
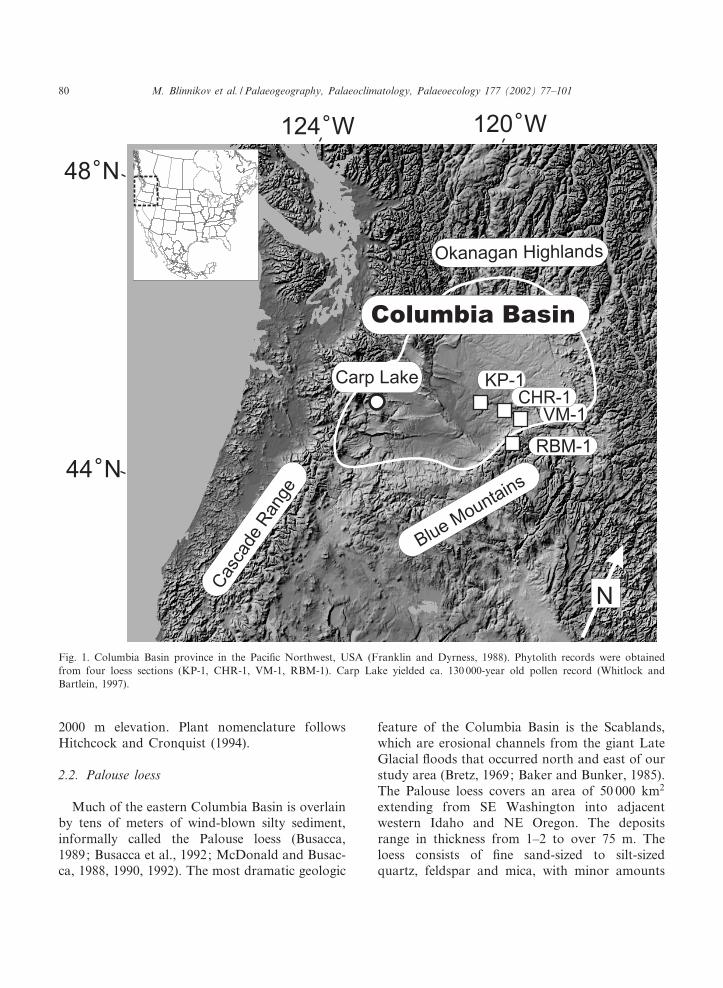
2000 m elevation. Plant nomenclature followsHitchcock and Cronquist (1994).
2.2. Palouse loess
Much of the eastern Columbia Basin is overlainby tens of meters of wind-blown silty sediment,informally called the Palouse loess (Busacca,1989; Busacca et al., 1992; McDonald and Busac-ca, 1988, 1990, 1992). The most dramatic geologic
feature of the Columbia Basin is the Scablands,which are erosional channels from the giant LateGlacial £oods that occurred north and east of ourstudy area (Bretz, 1969; Baker and Bunker, 1985).The Palouse loess covers an area of 50 000 km2
extending from SE Washington into adjacentwestern Idaho and NE Oregon. The depositsrange in thickness from 1^2 to over 75 m. Theloess consists of ¢ne sand-sized to silt-sizedquartz, feldspar and mica, with minor amounts
Fig. 1. Columbia Basin province in the Paci¢c Northwest, USA (Franklin and Dyrness, 1988). Phytolith records were obtainedfrom four loess sections (KP-1, CHR-1, VM-1, RBM-1). Carp Lake yielded ca. 130 000-year old pollen record (Whitlock andBartlein, 1997).
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10180

of heavy minerals and volcanic glass. The mainsource of the loess, apparently, was slack-waterdeposits of the Scabland Floods in the tributaryvalleys of the Columbia River (Busacca andMcDonald, 1994a). The total length of thePalouse loess record may exceed 2 million years(Kukla and Opdyke, 1980).
Several tephra layers and soil stratigraphicunits have been described from a few dozen loesssections spanning the last 70^100 ka. The mostimportant diagnostic tephra layers are MazamaAsh, which has been dated at 7.6 ka BP (Bacon,1983), Mount St. Helens tephra S, dated at 15.5 kaBP, and Mount St. Helens tephra C, which is ca.40 ka BP (Crandell et al., 1981; Berger and Bu-sacca, 1995). Overall, about 20 dates were ob-tained from loess sections in the area using ther-moluminescence dating (Berger and Busacca,1995; Richardson et al., 1997, 1999).
At the height of the last glaciation, the Cordil-leran Ice Sheet reached its southern limit in thenorthern Columbia Basin (Waitt and Thorson,1983). The maximum extent occurred between30 and 17 ka BP during marine isotope stage 2.The Fraser Glaciation was preceded by an inter-stadial event (30^50 ka BP), which was precededby another glacial advance in the early Wisconsin(s 50 ka BP). Studies of loess deposition patternsacross the Columbia Basin reveal two periods ofslow loess accumulation and apparent landscape
stability in the past 100 ka (Richardson et al.,1997; Blinnikov et al., 2001). These periods ledto the development of two widespread paleosolunits, the Washtucna Soil complex (ca. 17^37 kaBP) and the Devils Canyon Soil (ca. 67^83 ka BP)(McDonald and Busacca, 1992). Periods of fasterloess accumulation at the end of Devils Canyonand Washtucna Soil intervals coincided with thetime of catastrophic £ooding.
2.3. Study sites
Phytoliths were obtained from three loess sec-tions (CHR-1, VM-1, RBM-1) described by Bu-sacca and McDonald (1994b). These sections werelocated on £at surfaces away from streams tominimize the e¡ects of local topography on soilrecord preservation. In addition, the KP-1 sectiondata (Blinnikov et al., 2001) were considered tohelp interpret the other three sections and makeintra-regional comparisons (Fig. 1, Table 1).
3. Material and methods
Phytoliths were extracted from loess at Colora-do State University soil lab using standard wetoxidation and heavy £otation techniques (Kelly,1990; Stevenson, 1997). Approximately 20 g ofsoil was processed per sample. Organics were re-
Table 1Four loess sections from SE Washington State used in the study
Site Latitude Longitude Elevation MAT MAP Potential natural vegetation(PNV)
(N) (W) (m) (³C) (mm)
Kahlotus^Pasco-1 (KP-1) 46³31P10Q 118³37P30Q 410 12 220 Agropyron spicatum^Festucaidahoensis grassland nearArtemisia steppe
Cemetery Hill Road-1(CHR-1)
46³18P35Q 117³56P45Q 677 11 485 Festuca idahoensis^Agropyronspicatum grassland
Vernon Marll-1 (VM-1) 46³17P42Q 117³47P20Q 1095 8 790 Pseudotsuga menziesii^Abiesgrandis^Pinus ponderosa forest
Robinette Mountain-1(RBM-1)
46³08P20Q 117³56P30Q 1220 6 1000 Pseudotsuga menziesii^Abiesgrandis^Pinus ponderosa forestnear Abies lasiocarpa^Piceaengelmannii forest
MAT-mean annual temperature, MAP-mean annual precipitation. PNV is based on Ku«chler (1964) and Franklin and Dyrness(1988).
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 81

moved by heating 10 g of sample in 50 ml ofhydrogen peroxide (30%) in a heated bath. Diges-tion was allowed to proceed for 1.5 h. The re-maining residue was washed twice with distilledwater, boiled in hot hydrochloric acid (10%)for 15^20 min, washed twice, and dried. The siltfraction (5^100 Wm) was extracted by the
gravity sedimentation method (Pearsall, 1989).The opal phytoliths were extracted by £otationin a zinc bromide^hydrochloric acid mixturewith 2.3 g/cm3 speci¢c density.
Phytoliths were mounted in immersion oil forcounting. At least 300 phytoliths were tallied persample under a light microscope at 400U magni-
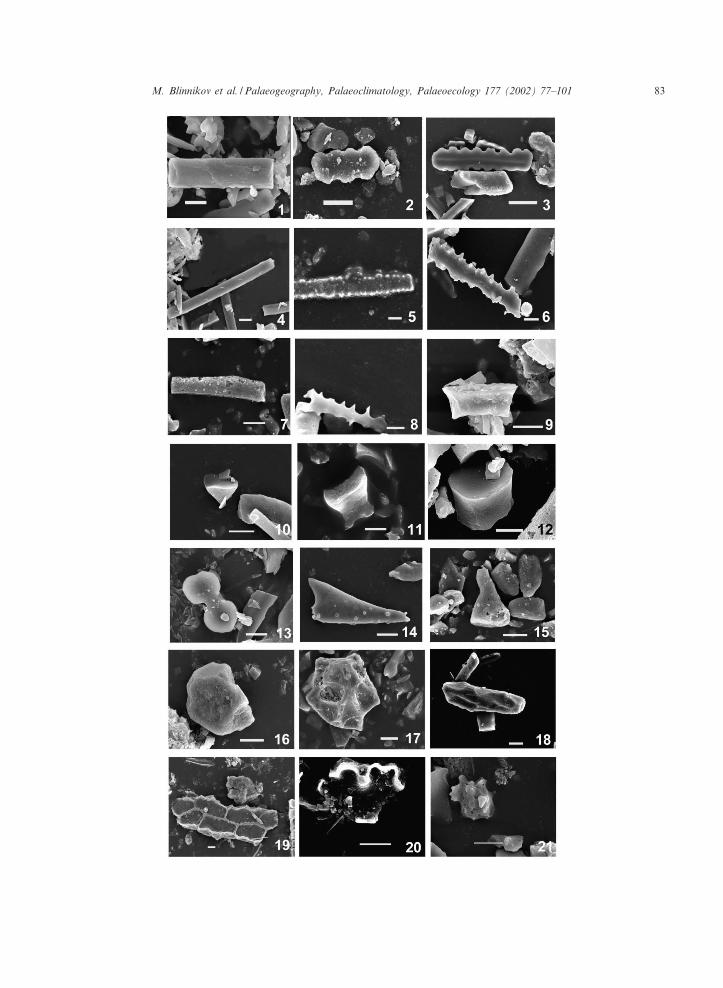
Fig. 3. Scanning electron micrograph images of the primary phytolith morphotypes recognized in this study. Bar is 10 Wm. Mor-photypes 1^15 are grass phytoliths, morphotypes 16^21 are non-grass phytoliths: (1) rectangular plate with straight edges,(2) short wavy plate with three to four undulations (e.g., Poa sandbergii, Koeleria cristata), (3) long wavy plate with s 5 undula-tions (e.g., Calamagrostis rubescens), (4) long cell with smooth parallel sides, (5) long cell indented, (6) long cell deeply indented,(7) long cell with angular sides, (8) dendritic long cell from seed epidermis, (9) elongated rondel (e.g., Agropyron spicatum),(10) rounded keeled rondel, (11) rounded horned rondel (e.g., Festuca idahoensis), (12) pyramidal rondel, (13) bilobate (e.g., Stipaspp.), (14) silici¢ed trichome, (15) silici¢ed hair base, (16) rounded blocky, (17) sculptured blocky, (18) elongated blocky, (19) epi-dermal polygonal, (20) epidermal anticlinal, (21) spiked (Pinus ponderosa-type).
Fig. 2. Location of loess stratigraphic sections and Carp Lake in relation to potential natural vegetation in the Columbia Basin.Potential natural vegetation type boundaries are drawn according to Ku«chler (1964) with names modi¢ed after Franklin and Dyr-ness (1988).
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10182
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 83

¢cation. The total phytolith count excluded otherbiological silica, such as diatoms and sponge spi-cules, occasionally found in the soil samples. Fi-nal phytolith diagrams were created in TILIA-GRAPH (Grimm, 1993). Phytolith zones wereestablished using sum of squares stratigraphicallyconstrained cluster analysis (CONISS) (Grimm,1993).
Identi¢cation of plants from phytoliths wasbased on a comparison with the reference phyto-lith collection obtained from 38 species of plantscommon to the region (Blinnikov, 1999). Grassphytoliths included plates, short wavy forms(three to four pronounced undulations, or manyslight undulations), long wavy forms (s 5 pro-nounced undulations), elongated rondels, roundedrondels with keel, rounded rondels with horns,pyramidal rondels, smooth long cells, indentedlong cells, deeply indented and angular long cells,phytoliths from glumes and seed epidermis (scuti-form and dendritic opal), Stipa-type bilobates, tri-chomes, and silici¢ed hairs. Non-grass phytolithsincluded blocky forms (including sculptured,smooth and elongated types) of Artemisia, Abies,and Picea, polygonal epidermal phytoliths, anti-clinal epidermal phytoliths, conical phytoliths ofCarex, spiked phytoliths of Pinus ponderosa, un-evenly thickened cell walls of Larix and Abies,`other conifer' category including silici¢ed trache-ids, and Asteraceae phytoliths including perfo-rated plates and segmented hairs (Fig. 3).
In addition, fossil phytolith assemblages weredirectly compared with 58 modern surface soil
analogs from eight vegetation types by usingsquared chord distance (SCD) as a dissimilaritymeasure (Overpeck et al., 1985). SCD suppressesnoise in the data and is robust with respect to thechoice of the number of morphotypes used in theanalysis, which makes it a preferred measure for¢nding analogs (Anderson et al., 1989). The ma-trix of SCD for 58 modern assemblages, both be-tween and within eight vegetation types, as de-¢ned in Blinnikov et al. (2001), was analyzed todetermine cuto¡ SCD values representing weak,moderate, and strong analogs. Histogramsof the within-type and between-type SCD val-ues are shown in Fig. 4. Based on the observedvalue distributions, we identi¢ed SCD valuesof less than 0.195 as `strong', 0.195^0.280 as`moderately strong', and 0.280^0.380 as `weak'analogs. SCD values s 0.380 were consideredno analog.
We assume that changes in phytolith percen-tages at di¡erent depths in the loess sections rep-resent vegetation composition at each site at thetime of deposition. We further assume that thereconstructed vegetation around each site wasrepresentative of the surrounding area. Consider-ing that little is known about the size of catch-ment area and phytolith taphonomy (Piperno,1988; Fredlund and Tieszen, 1994), extrapolatingphytolith data over surrounding area should beapproached with caution. Modern analog studies(Fredlund and Tieszen, 1994; Blinnikov, 1999)suggest that phytolith assemblages from modernsoils track vegetation composition at the stand
Fig. 4. Frequency distribution of SCD for (A) 1414 between-type and (B) 230 within-type pairs of samples.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10184
level (e.g., vegetation patches on the order of 1 hain size). We felt justi¢ed in making extrapolationsbecause the sites featured di¡erent associationstoday, which could be distinguished basedon the phytolith data from the top of each sec-tion.
4. Results
4.1. CHR-1 section
Twenty-four soil horizons were identi¢ed in the540-cm-deep pro¢le of the Cemetery Hill Road-1
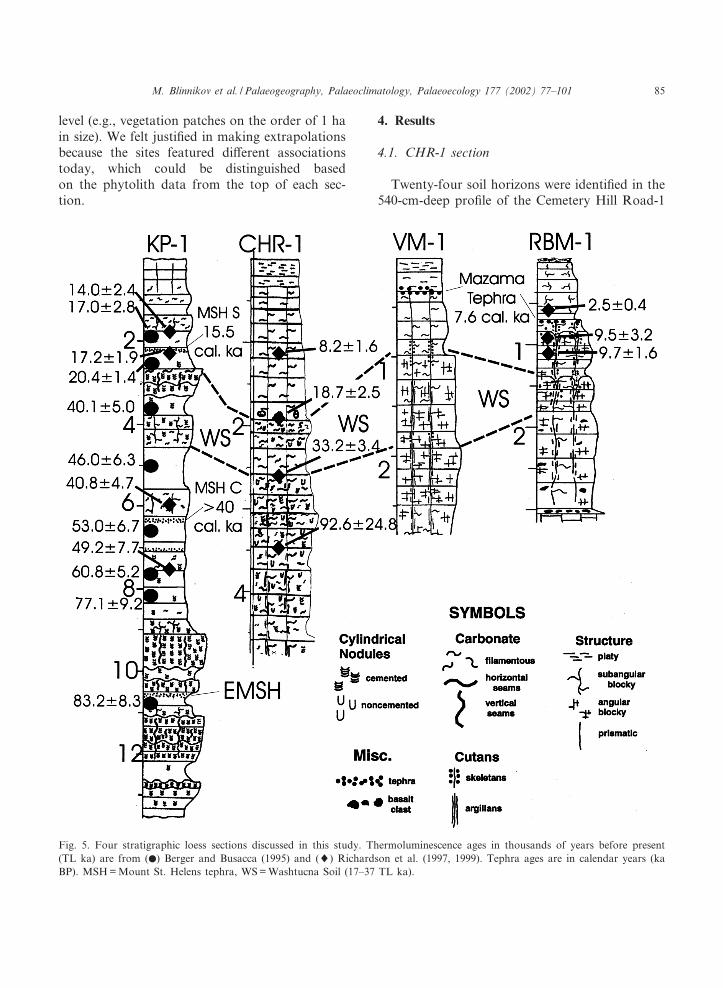
Fig. 5. Four stratigraphic loess sections discussed in this study. Thermoluminescence ages in thousands of years before present(TL ka) are from (b) Berger and Busacca (1995) and (8) Richardson et al. (1997, 1999). Tephra ages are in calendar years (kaBP). MSH = Mount St. Helens tephra, WS = Washtucna Soil (17^37 TL ka).
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 85

(CHR-1) section. Four dates were obtained fromthe upper 300 cm of the pro¢le (Fig. 5). TheWashtucna Soil complex, with its characteristicgray petrocalcic horizon, is found in the pro¢lebetween 196 and 258 cm depth. A thermolumines-cence date below the Washtucna Soil at 314 cmdepth was older than expected ^ 92.6 þ 24.8 ther-moluminescence (TL) ka BP (Richardson et al.,1999). Based on the correlation of paleosols indi¡erent sections, it seems that the actual datefor this horizon should be closer to 68 ka BP.The Washtucna Soil complex apparently devel-oped during the coldest part of the last glaciation,marine isotope stage 2 (33.2 þ 3.4 TL ka date at240 cm depth). Weakly altered loess overlies theWashtucna Soil complex above 192 cm depth.Two TL dates bracket the age of this loess unitbetween 18.7 þ 2.5 TL ka at 182 cm depth and8.2 þ 1.6 TL ka at 105 cm depth and suggestthat it is late Pleistocene to early Holocene inage. The modern soil extends down to 98 cmdepth. Nineteen phytolith samples were processedand analyzed from CHR-1 and three phytolithzones were identi¢ed by cluster analysis (Fig. 6).
Zone CHR-1a (323^542 cm depth) contains sixsamples with estimated age s 68 ka. Five lowerassemblages have uniformly high proportions of
grass phytoliths (around 90%), and low of non-grass phytoliths (6 10%), indicative of a grass-land. The most common were short wavy phyto-liths of Poa or Koeleria (35^40% of the total num-ber of all phytoliths) and di¡erent rondels,indicative of Festuca (25%). Elongated andhorned rondels, common in Agropyron and Festu-ca respectively, made up 5^10% of all phytolithsin this zone. Lack of bilobate phytoliths diagnos-tic of Stipa suggests that Agropyron and Festucawere the two probable contributors of the rondelsin this zone. Comparison with modern analogssupports this conclusion (Table 2). The closestanalog for the ¢ve lower samples came from Fes-tuca idahoensis-dominated grassland with Koeleriacristata, some Agropyron spicatum, and diverseherbs, from the Pataha River valley in the Uma-tilla National Forest, Washington. The elevation(1400 m) and climate data for this analog suggestthat conditions were colder and wetter at CHR-1during the Zone CHR-1a interval than today. Thetop assemblage from the zone at 354 cm depthrepresents a drier vegetation type. In this assem-blage, the proportion of short wavy and blockyphytoliths was higher suggesting an increased roleplayed by Poa sandbergii and Artemisia. The clos-est analog for the assemblage came from a
Fig. 6. Percentages of phytolith morphotypes in CHR-1 loess section.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10186

present-day Agropyron spicatum^Poa sandbergiigrassland (with Artemisia tridentata present with-in 10^15 m of the site) from central Oregon. Scar-city of deeply indented long cells and elongatedrondels of Agropyron in the fossil assemblagefrom CHR-1, however, suggests that A. spicatumplayed a smaller role at CHR-1 at that time rela-tive to the modern analog community. Thus,CHR-1 may have been a transitional site betweengrassland and sagebrush steppe at ca. 70 ka.
Zone CHR-1b (182^323 cm depth) contains sixsamples, three from the lower and three from theupper portions of the Washtucna Soil complex.Sample at 242 cm (chr13b) was dated at33.2 þ 3.4 TL ka. A sample immediately abovethe upper zone boundary was dated at 18.7 þ 2.5TL ka. The assemblages within Zone CHR-1brepresent communities during the time of marineisotope stages 2 and 3, including the LGM at21 ka BP. The phytolith assemblages are diverse,suggesting profound changes in vegetation com-position. Two lower assemblages contain a highproportion (50%) of non-grass phytoliths, primar-ily blocky forms of shrub Artemisia and someanticlinal phytoliths of herbs (e.g., Asteraceae).Grass phytoliths are represented by long cells(15%), rondels (10%) and short wavy phytoliths(5^20%), probably contributed by Poa and Festu-ca. The closest modern analogs for these two as-semblages are from two di¡erent sagebrush steppecommunities: warm and dry sagebrush steppe inBoardman Research Natural Area, Oregon andfrom cool and moist sagebrush steppe in an open-ing within Pinus ponderosa forest zone in UmatillaNational Forest, Washington (Table 2). The low-ermost assemblage had a higher proportion ofblocky phytoliths indicating sagebrush steppe atthe site, between 33 and 92 TL ka. If the actualdate for the bottom of the zone is 68 ka BP, thisperiod could represent early Wisconsin commun-ities corresponding to marine isotope stage 4. An-other period of abundant Artemisia phytoliths atCHR-1 occurs in the upper Washtucna Soil, atthe LGM (196^212 cm depth). Stipa-type bilo-bates (2^3%) were found in these samples, sug-gesting a dry period. At the same time, the lowerpresence of Agropyron indicates colder-than-present conditions. The modern analog for these
LGM samples came not from a sagebrush steppe,but from Festuca^Agropyron grassland near sage-brush steppe in Horse Ridge Research NaturalArea in central Oregon. The inferred climatewas cooler and drier than today.
Zone CHR-1c (0^182 cm depth) includes twolate Pleistocene assemblages from s 100 cm depthand ¢ve assemblages from the Holocene soil. Thelate Pleistocene assemblages displayed a more xe-ric character because of the greater presence ofArtemisia and Stipa phytoliths, and a higher pro-portion of wavy forms, probably of Poa sandber-gii. Sagebrush apparently disappeared at CHR-1by the early Holocene (8.2 þ 1.6 TL ka for a sam-ple at 110 cm depth). The Holocene assemblagescontained a high proportion of rondels (30%),which indicates the presence of Festuca, probablyFestuca idahoensis. The two late glacial sampleswere similar to Festuca^Koeleria grassland grow-ing today on the slopes of the Pataha River Valleyin the Blue Mountains at 1400 m elevation. Theseassemblages imply cooler and wetter climate thantoday. The upper three assemblages (early to midHolocene) were analogous to the modern mixedFestuca idahoensis-Agropyron spicatum grasslandin Adams County, Washington. The climate forthe mid Holocene implied by this modern analogwas drier than today (320 mm versus 485 mmMAP). Epidermal phytoliths from grass glumes(dendritic and scutiform opal), typical of weedyBromus tectorum and grain crops (Triticum, Hor-deum), were found in small quantities in the sec-ond sample from the top, but nowhere else in therecord. These seed phytoliths may record the ar-rival of the Euro-American settlers and the startof modern agriculture in the region.
4.2. VM-1 section
Fourteen soil horizons were described from a437-cm-deep pro¢le of the Vernon Marll-1 (VM-1) section on Cahill Mountain, of which 11 sam-ples were processed for phytoliths. Several paleo-sol horizons were described in the pro¢le, but nodirect dates are available (Fig. 7). Mazama Ash(7.6 ka BP) was found at 35^40 cm below thesurface. The Washtucna Soil complex appears asa prominent stratigraphic marker of the LGM
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 87
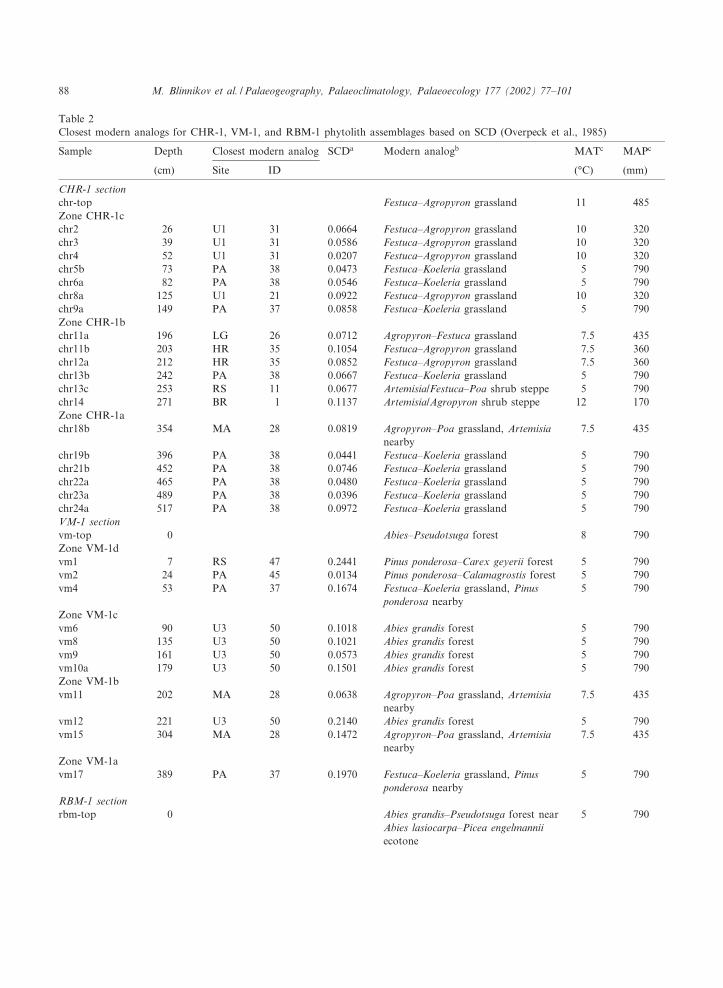
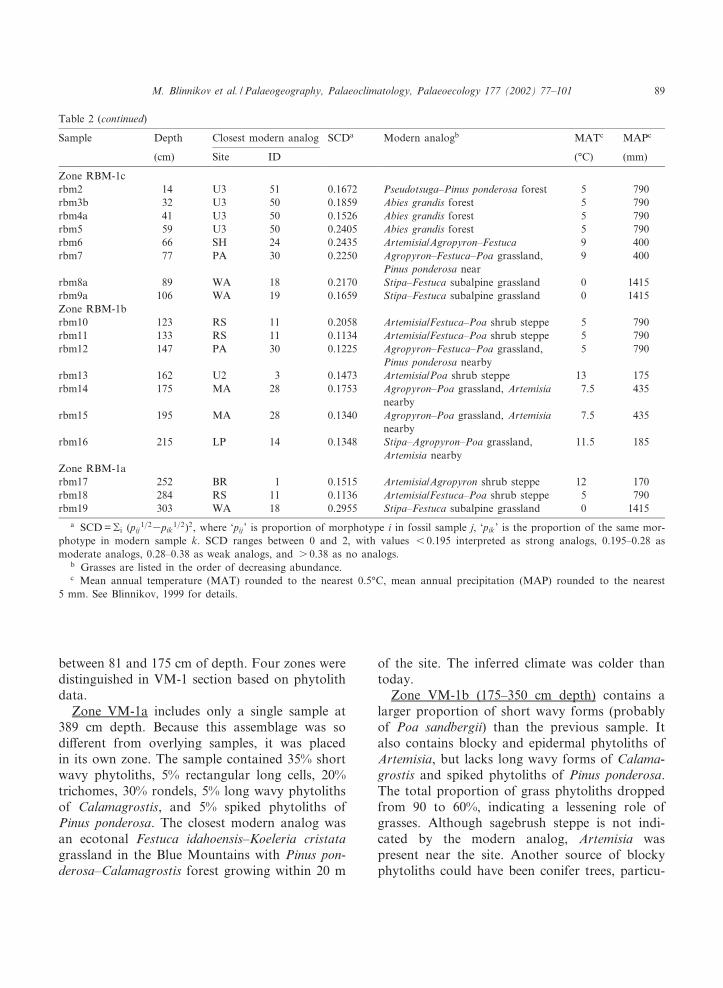
Table 2Closest modern analogs for CHR-1, VM-1, and RBM-1 phytolith assemblages based on SCD (Overpeck et al., 1985)
Sample Depth Closest modern analog SCDa Modern analogb MATc MAPc
(cm) Site ID (³C) (mm)
CHR-1 sectionchr-top Festuca^Agropyron grassland 11 485Zone CHR-1cchr2 26 U1 31 0.0664 Festuca^Agropyron grassland 10 320chr3 39 U1 31 0.0586 Festuca^Agropyron grassland 10 320chr4 52 U1 31 0.0207 Festuca^Agropyron grassland 10 320chr5b 73 PA 38 0.0473 Festuca^Koeleria grassland 5 790chr6a 82 PA 38 0.0546 Festuca^Koeleria grassland 5 790chr8a 125 U1 21 0.0922 Festuca^Agropyron grassland 10 320chr9a 149 PA 37 0.0858 Festuca^Koeleria grassland 5 790Zone CHR-1bchr11a 196 LG 26 0.0712 Agropyron^Festuca grassland 7.5 435chr11b 203 HR 35 0.1054 Festuca^Agropyron grassland 7.5 360chr12a 212 HR 35 0.0852 Festuca^Agropyron grassland 7.5 360chr13b 242 PA 38 0.0667 Festuca^Koeleria grassland 5 790chr13c 253 RS 11 0.0677 Artemisia/Festuca^Poa shrub steppe 5 790chr14 271 BR 1 0.1137 Artemisia/Agropyron shrub steppe 12 170Zone CHR-1achr18b 354 MA 28 0.0819 Agropyron^Poa grassland, Artemisia
nearby7.5 435
chr19b 396 PA 38 0.0441 Festuca^Koeleria grassland 5 790chr21b 452 PA 38 0.0746 Festuca^Koeleria grassland 5 790chr22a 465 PA 38 0.0480 Festuca^Koeleria grassland 5 790chr23a 489 PA 38 0.0396 Festuca^Koeleria grassland 5 790chr24a 517 PA 38 0.0972 Festuca^Koeleria grassland 5 790VM-1 sectionvm-top 0 Abies^Pseudotsuga forest 8 790Zone VM-1dvm1 7 RS 47 0.2441 Pinus ponderosa^Carex geyerii forest 5 790vm2 24 PA 45 0.0134 Pinus ponderosa^Calamagrostis forest 5 790vm4 53 PA 37 0.1674 Festuca^Koeleria grassland, Pinus
ponderosa nearby5 790
Zone VM-1cvm6 90 U3 50 0.1018 Abies grandis forest 5 790vm8 135 U3 50 0.1021 Abies grandis forest 5 790vm9 161 U3 50 0.0573 Abies grandis forest 5 790vm10a 179 U3 50 0.1501 Abies grandis forest 5 790Zone VM-1bvm11 202 MA 28 0.0638 Agropyron^Poa grassland, Artemisia
nearby7.5 435
vm12 221 U3 50 0.2140 Abies grandis forest 5 790vm15 304 MA 28 0.1472 Agropyron^Poa grassland, Artemisia
nearby7.5 435
Zone VM-1avm17 389 PA 37 0.1970 Festuca^Koeleria grassland, Pinus
ponderosa nearby5 790
RBM-1 sectionrbm-top 0 Abies grandis^Pseudotsuga forest near
Abies lasiocarpa^Picea engelmanniiecotone
5 790
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10188
between 81 and 175 cm of depth. Four zones weredistinguished in VM-1 section based on phytolithdata.
Zone VM-1a includes only a single sample at389 cm depth. Because this assemblage was sodi¡erent from overlying samples, it was placedin its own zone. The sample contained 35% shortwavy phytoliths, 5% rectangular long cells, 20%trichomes, 30% rondels, 5% long wavy phytolithsof Calamagrostis, and 5% spiked phytoliths ofPinus ponderosa. The closest modern analog wasan ecotonal Festuca idahoensis^Koeleria cristatagrassland in the Blue Mountains with Pinus pon-derosa^Calamagrostis forest growing within 20 m
of the site. The inferred climate was colder thantoday.
Zone VM-1b (175^350 cm depth) contains alarger proportion of short wavy forms (probablyof Poa sandbergii) than the previous sample. Italso contains blocky and epidermal phytoliths ofArtemisia, but lacks long wavy forms of Calama-grostis and spiked phytoliths of Pinus ponderosa.The total proportion of grass phytoliths droppedfrom 90 to 60%, indicating a lessening role ofgrasses. Although sagebrush steppe is not indi-cated by the modern analog, Artemisia waspresent near the site. Another source of blockyphytoliths could have been conifer trees, particu-
Table 2 (continued)
Sample Depth Closest modern analog SCDa Modern analogb MATc MAPc
(cm) Site ID (³C) (mm)
Zone RBM-1crbm2 14 U3 51 0.1672 Pseudotsuga^Pinus ponderosa forest 5 790rbm3b 32 U3 50 0.1859 Abies grandis forest 5 790rbm4a 41 U3 50 0.1526 Abies grandis forest 5 790rbm5 59 U3 50 0.2405 Abies grandis forest 5 790rbm6 66 SH 24 0.2435 Artemisia/Agropyron^Festuca 9 400rbm7 77 PA 30 0.2250 Agropyron^Festuca^Poa grassland,
Pinus ponderosa near9 400
rbm8a 89 WA 18 0.2170 Stipa^Festuca subalpine grassland 0 1415rbm9a 106 WA 19 0.1659 Stipa^Festuca subalpine grassland 0 1415Zone RBM-1brbm10 123 RS 11 0.2058 Artemisia/Festuca^Poa shrub steppe 5 790rbm11 133 RS 11 0.1134 Artemisia/Festuca^Poa shrub steppe 5 790rbm12 147 PA 30 0.1225 Agropyron^Festuca^Poa grassland,
Pinus ponderosa nearby5 790
rbm13 162 U2 3 0.1473 Artemisia/Poa shrub steppe 13 175rbm14 175 MA 28 0.1753 Agropyron^Poa grassland, Artemisia
nearby7.5 435
rbm15 195 MA 28 0.1340 Agropyron^Poa grassland, Artemisianearby
7.5 435
rbm16 215 LP 14 0.1348 Stipa^Agropyron^Poa grassland,Artemisia nearby
11.5 185
Zone RBM-1arbm17 252 BR 1 0.1515 Artemisia/Agropyron shrub steppe 12 170rbm18 284 RS 11 0.1136 Artemisia/Festuca^Poa shrub steppe 5 790rbm19 303 WA 18 0.2955 Stipa^Festuca subalpine grassland 0 1415
a SCD =gi (pij1=23pik
1=2)2, where `pij ' is proportion of morphotype i in fossil sample j, `pik' is the proportion of the same mor-photype in modern sample k. SCD ranges between 0 and 2, with values 6 0.195 interpreted as strong analogs, 0.195^0.28 asmoderate analogs, 0.28^0.38 as weak analogs, and s 0.38 as no analogs.
b Grasses are listed in the order of decreasing abundance.c Mean annual temperature (MAT) rounded to the nearest 0.5³C, mean annual precipitation (MAP) rounded to the nearest
5 mm. See Blinnikov, 1999 for details.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 89

larly Abies and Picea. The sample from 221 cmcontains phytoliths with unevenly thickened cellwalls and tracheid cells common in conifers(e.g., Abies, Larix). Some of the blocky forms inthis zone resemble small endodermal blocky phy-toliths of Picea. No diagnostic asterosclereids ofPseudotsuga were found in this sample. The clos-est modern analog for the assemblage was a forestcommunity with Abies grandis and Picea engel-mannii near Chase Mountain in the Umatilla Na-tional Forest. The inferred climate conditions forthe zone were colder and drier than at present.
Zone VM-1c (71^175 cm depth) represents thetime of the Washtucna Soil complex (includingthe LGM). High percentages of plates, smoothrectangular long cells, trichomes, and moderatevalues of blocky and `other conifer' phytolithsoccurred in this zone. The closest modern analogsuggests the presence of a conifer (Picea/Abies)forest at the site. Presence of Artemisia shrub can-not be ruled out given the occurrence of blockyforms. Regrettably, our modern database includesonly two samples from the present-day Abiesgrandis forest zone, and both are from closed-can-opy forests lacking Artemisia or bunchgrasses.
The closest modern analog for all four samplesin this zone was A. grandis forest with some Piceaengelmannii from the Chase Mountain in theUmatilla National Forest, Washington. TheLGM parkland at VM-1 site does not apparentlyhave a strong modern analog. The inferred cli-mate for this zone is colder and drier than today.
Zone VM-1d (0^71 cm depth) includes threeassemblages. A sample below Mazama Ash (i.e.,with the age s 7.6 ka BP) at 53 cm depth con-tained high percentages of grass phytoliths (90%),particularly rondels (30%), while the upper twosamples contained a lower amount of grass phy-toliths (80%), with long cells the most commongroup. This assemblage had as its closest analogFestuca^Koeleria grassland near Pinus ponderosaforest and could be late glacial in age. The uppertwo samples were most similar to Pinus ponde-rosa^Calamagrostis and Pinus ponderosa^Carexgeyerii forest stands near Rose Springs in theUmatilla National Forest. Both samples con-tained spiked phytoliths diagnostic of P. ponde-rosa and a high proportion of long wavy formsand trichomes of Calamagrostis rubescens. Con-trary to the expectations, no Carex phytoliths
Fig. 7. Percentages of phytolith morphotypes in VM-1 loess section.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10190

were found in the samples, although Carex geyeriiis common in nearby forests today. The inferredclimate is cooler and wetter than today.
4.3. RBM-1 section
Eighteen soil horizons were identi¢ed in the300-cm-deep pro¢le in the Robinette Mountain-1(RBM-1) section with a phytolith sample ob-tained from each (Fig. 8). RBM-1 is a high-eleva-tion site (1200 m) near the border of the Palousearea. Mazama Ash was found at 63^69 cm depthin the pro¢le. Three TL dates were available fromRBM: 2.5 þ 0.4 TL ka for the sample above Ma-zama Ash at 59 cm depth, 9.5 þ 3.2 TL ka for thesample at 93 cm depth, and 9.7 þ 1.6 TL ka forthe sample at 123 cm depth (Richardson et al.,1999). Three phytolith zones were distinguished.
Zone RBM-1a (240^305 cm depth) containsthree phytolith samples with high percentage ofnon-grass phytoliths (30^50%), including blockyforms of Artemisia, anticlinal phytoliths of herbsand deciduous shrubs, and a few spiked phyto-liths of Pinus ponderosa. Bilobate phytoliths ofStipa were present in moderate quantity (3^5%).
The percent of Artemisia forms increased towardsthe top of the zone, while that of grass phytolithsdecreased, and Pinus phytoliths disappeared.These changes suggest a transition from a mesicgrassland community with Stipa and scattered Pi-nus to a sagebrush community. Modern analogssupport such interpretation. The lowermost sam-ple had a weak analog (SCD = 0.2955), mesicStipa occidentalis^Festuca viridula subalpine grass-land from the Wallowa Mountains. The analogfor the next sample higher up in the pro¢le wasa drier Artemisia rigida/Festuca idahoensis^Poasandbergii shrub steppe, while the upper samplein the zone had a strong analog, Artemisia triden-tata/Agropyron spicatum shrub steppe fromBoardman Research Natural Area in Oregon, avery dry site today (Table 2). Based on these com-parisons, the conditions at RBM-1 were probablycool and moist at ¢rst, and then got progressivelydrier and perhaps warmer. The age of the zone isunknown, but it underlies the Washtucna Soilcomplex (17^37 TL ka) and is probably early Wis-consin in age (s 60 ka BP). The climate duringZone RBM-1a is di¤cult to interpret because ofthe wide range of climatic conditions implied by
Fig. 8. Percentages of phytolith morphotypes in RBM-1 loess section.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 91

modern analogs. Considering the topographic po-sition of the site, the three analogs likely indicatethat conditions were cool and moist.
Zone RBM-1b (118^240 cm depth) containsseven phytolith samples. All samples contain 5^40% non-grass phytoliths, mostly blocky and po-lygonal phytoliths of Artemisia and low percen-tages of conifer phytoliths (e.g., 6 2% Pinus pon-derosa phytoliths were found in four samples).The overall proportion of grass phytoliths washigher in this zone than in Zone RBM-1a, sug-gesting that the conditions were not as extremeand that the area supported well-developed grass-land, probably near the forest border. The smallbut consistent percentage of tree phytoliths sup-ports such a reconstruction. Among the grassphytoliths, the most abundant (20^40%) wereshort wavy phytoliths (Poa, Koeleria) and di¡er-ent rondels (Festuca).
The lower three samples indicate a dry grass-land/parkland ecotonal community in the pre-Washtucna Soil complex interval (s 37 TL ka),while the upper four samples (LGM age) weremost similar to sagebrush steppe community sam-ples from Zone RBM-1a. All samples in the zonehave strong modern analogs from present-dayStipa and Agropyron-dominated dry grasslandand Artemisia-dominated shrub steppe. Thegrass/non-grass ratio suggests a change from ashrub community near the bottom of the zoneto a Stipa-dominated grassland, to Agropyron-dominated grassland in the middle of the zone,and ¢nally back to Artemisia shrub communitynear the top of the zone. An apparent peak inrondels near the middle of the zone would gener-ally indicate an increase in Festuca. High percen-tages of rounded keeled rondels in this zone sug-gest the presence of an unknown grass species,particularly rich in this morphotype, but not in-cluded in our reference collection. The impliedclimate conditions were drier and probably colderthan today. The modern analogs do not indicatecolder conditions, but the lack of trees at theRBM-1 site suggests that it may have been abovethe tree line.
Zone RBM-1c (0^118 cm depth) contains eightphytolith samples from the Late Glacial and theHolocene. Mazama Ash (7.6 ka BP) separates
Late Glacial and early Holocene assemblagesfrom mid and late Holocene ones. Total percent-age of non-grass phytoliths changes from 40%near the bottom of the zone to 20% at the topof the zone. The blocky and epidermal polygonalphytoliths, indicative of Artemisia shrubs, gradu-ally decrease, while the proportions of di¡erentconifer phytoliths increase in the upper zone, sug-gesting gradual replacement of open shrub steppeand grassland by forest. Conical phytoliths ofCarex were found in three samples in the middleof the zone. Among the grass phytoliths, formscommon to open grasslands, e.g., short wavyforms (Poa, Koeleria) and bilobates (Stipa) atthe bottom of the zone, gave way at the top tolong wavy phytoliths, rectangular long cells, andtrichomes, probably of Calamagrostis rubescens (atypical forest species). Matching fossil samplesfrom Zone RBM-1c with modern analogs matchthe Late Glacial grassland with the present-daysubalpine Stipa^Festuca grassland from the Wal-lowa Mountains. This vegetation was replaced byparkland with Pinus ponderosa/Agropyron^Festucain the early Holocene. Eventually conifer forestwith Abies and Pseudotsuga as co-dominantsand P. ponderosa as an associate species was es-tablished at the site. The interpreted climate forZone RBM-1c was colder and wetter in the LateGlacial, and warmer and drier than present in theearly Holocene.
5. Discussion: environmental reconstructions
Phytolith data reveal the complex character ofvegetation and climate change in the ColumbiaBasin at di¡erent elevations during the last100 ka. Comparison of sites along a topographicgradient helps evaluate the spatial patterns of cli-mate and vegetation change, assuming that thephytolith records track zonal vegetation changes.The magnitude of change over the last 100 ka wasgreat. Some widespread taxa today had a morerestricted distribution in the past, for example,most trees and Agropyron during the late Pleisto-cene. On the other hand, Stipa, Poa, and Festucaexpanded their ranges during the same time. SomePleistocene assemblages have no modern analogs.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10192

Fig. 9 shows the inferred changes from the timeof the LGM to the present. Four periods areshown: the LGM ca. 21 ka BP near the top ofthe Washtucna Soil complex; the Late Glacial
interval ca. 14 ka BP immediately above Wash-tucna Soil and Mt. St. Helens tephra S (15.5 kaBP); the early Holocene ca. 10 ka BP immediatelybelow Mazama Ash (7.6 ka BP); and the present-
Fig. 9. Inferred vegetation in the Columbia Basin during the last 21 000 years based on the pollen record at Carp Lake (Whitlockand Bartlein, 1997) and phytoliths in four loess sections in the Columbia Basin (Blinnikov, 1999).
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 93

day samples. The Carp Lake record at 714 melevation on the western edge of the Basin wascorrelated with the phytolith records.
Fig. 10 provides an overview of vegetation andclimate change at four loess sites and Carp Lakeduring the last 100 ka. The chronological controlat CHR-1, VM-1, and RBM-1 sites below the
Washtucna Soil complex is poor, while the KP-1and Carp Lake have established chronologies thatcorrelate reasonably well (Blinnikov et al., 2001).Therefore, the early and mid Wisconsin vegeta-tion zone boundaries at CHR-1, VM-1, andRBM-1 were assigned only approximately basedon the stratigraphy and should be interpreted
Fig. 10. Inferred vegetation and climate history based on phytolith data from four loess sections in the Columbia Basin (Blinni-kov, 1999) and pollen data at Carp Lake (Whitlock and Bartlein, 1997). Estimated ages in TL ka BP for the four loess sectionsand in ka BP for the Carp Lake pollen. Marine isotope stages are from Martinson et al. (1987). Climate interpretations are rela-tive to the present-day conditions and based on modern analog matches. Washtucna Soil interval (17^37 TL ka) is shaded.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10194

with caution (Fig. 10). The ages for the KP-1section are uncalibrated TL ages calculated basedon an age^depth model (Blinnikov et al., 2001).The Carp Lake ages are calculated calendar yearsbased on a few radiocarbon dates and tephra(Whitlock and Bartlein, 1997).
5.1. Early Wisconsin (100^60 ka BP)
The phytolith records provided some interestinginsights into the character of the early and midWisconsin vegetation. Few paleoecological rec-ords from the Paci¢c Northwest describe vegeta-tion change during that time (Alley et al., 1986;Beiswenger, 1991; Heusser and Heusser, 1990;Worona and Whitlock, 1995), and none comefrom the Columbia Basin. In the early to middleWisconsin a glacial advance took place in the Pa-ci¢c Northwest (Baker et al., 1991; Clague et al.,1992), although the age and extent are not known.McDonald and Busacca (1988) provided evidenceof early Wisconsin post-glacial £oods in the Co-lumbia Basin, similar to the Late Glacial £oodsca. 14 ka (Baker and Bunker, 1985). The £oodsoccurred ca. 60^70 ka, during the glacial retreatof the early Wisconsin. Prior to 83 ka, conditionsat both KP-1 and Carp Lake were colder anddrier than today. A sagebrush steppe with Stipaand Poa existed at KP-1, and an open forest ofPinus and Picea existed at Carp Lake. The CarpLake record (714 m asl) for the early Wisconsinsuggests that a moderately cold and dry period atCarp Lake can be correlated with the marine iso-tope stage 4 (59^74 ka BP). A period of non-ana-log vegetation when a mixture of high- and low-elevation species was present at the lake occurredbetween 83 and 73 ka BP (marine isotope stage5a). This may have been a period of relativelywarm, but cooler-than-present, and wetter-than-present summers, caused by an unusual combina-tion of high summer insolation, modest global icevolumes, and cooler-than-present sea surface tem-peratures in the north Paci¢c (Whitlock and Bart-lein, 1997). An unusual assemblage was also de-tected ca. 77 TL ka at KP-1, when Pinusponderosa/Festuca^Carex parkland may have ex-isted at the site, implying cooler and wetter con-ditions than today.
The KP-1 section provides evidence of two dis-tinct intervals of sagebrush steppe in the earlyWisconsin, when conditions were colder and drieras evidenced by the stable isotopes at KP-1 (Ste-venson, 1997). The two intervals at KP-1 (ca.64 TL ka and prior to 83 TL ka) could be corre-lated with marine isotope stages 4 (59^74 ka BP)and 5b (85^94 ka BP) respectively. CHR-1 sitehad a distinct period of Artemisia shrub steppebelow the Washtucna Soil complex (s 37 TLka) in the zone with the probable age of ca. 68ka BP. At an earlier undated interval, cool moistconditions compared to the present existed atCHR-1, when the site supported Festuca^Koeleriagrassland, similar to that during the Late Glacialinterval. Four samples of the middle-slope VM-1section suggest overall cooler and wetter condi-tions, when grassland/Pinus ponderosa forest tran-sition existed at the site which today is occupiedby Abies^Pseudotsuga forest. Phytoliths at RBM-1 suggest that cooler and perhaps wetter condi-tions may have existed at the site in the earlyWisconsin, although chronological control is lack-ing at both sites. Overall, trees were less impor-tant than today in the eastern part of the Colum-bia Basin at all elevations, probably as a result oflower temperatures and available moisture.
5.2. Mid Wisconsin (60^40 ka BP)
The nature of climate and vegetation during themid Wisconsin interval in the Paci¢c Northwest isknown better than that of the early Wisconsin(Clague, 1978; Clague et al., 1992). Conditionsduring that time were cooler than today, but notas cold as at the LGM, with variable precipita-tion. West of the Columbia Basin, records fromcoastal British Columbia and Washington Statefrom 40 to 51 ka BP indicate a climate coolerby about 1³C, while the vegetation was similarto the present (Clague, 1978; Alley and Hicock,1986). Two paleosols described from the PurcellTrench in the south-central British Columbia in-dicate that the period around 42 ka was 3³C cold-er and somewhat wetter than today, as evidencedby the presence of spruce pollen (Alley et al.,1986). In the Oregon Coast Range, a pollen rec-ord from Little Lake shows cooler wetter climate
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 95

than today prior to 40 ka (Worona and Whitlock,1995). East of the Columbia Basin, a pollen rec-ord from Grays Lake (Beiswenger, 1991) in theeastern part of the Snake River Plain indicatescold and dry Artemisia steppe between 70 and30 ka.
Marine isotope records suggest that global con-ditions were variable during most of the mid Wis-consin, but generally temperatures were lowerthan today (Martinson et al., 1987, marine iso-tope stage 3, 28^59 ka BP). The KP-1 sectionprovides evidence of a moderately cool and dryperiod in the mid Wisconsin (42^59 TL ka), whenan Agropyron-dominated dry grassland, but notshrub steppe, existed at the site. It was duringthe time that Old Maid Coulee paleosol devel-oped. The CHR-1 site supported Artemisia-dom-inated shrub steppe with xeric grasses, also sug-gesting colder and drier conditions than atpresent. Drier conditions are inferred for theVM-1 site, where Agropyron-dominated grasslandexisted, with a few scattered conifer trees. Like-wise, site phytoliths from the RBM-1 section be-low the Washtucna Soil complex indicate drier-than-present conditions. The closest analog forthat sample, however, came from Agropyron^Stipa^Poa grassland, which would indicate condi-tions warmer than those observed at RBM-1 to-day. Carp Lake supported an open forest of Abiesand Pinus ponderosa during the mid Wisconsin(CL-4 between 31 and 43 ka BP and CL-5 be-tween 43 and 58 ka BP).
5.3. LGM (21 ka BP)
Based on our interpretation of phytolith andpollen data, the LGM vegetation was very di¡er-ent from that of today. Artemisia shrubs wereconsiderably more widespread, while trees wereabsent from all but middle-elevation sites (CarpLake at 714 m, VM-1 at 1095 m). The loss offorest suggests much drier and colder conditionsthan today. The pollen record at Carp Lake in-dicates a high presence of Artemisia, grasses, andscattered Picea (spruce) (Zone CL-3). Phytolithsat KP-1 (410 m) near the center of the ColumbiaBasin provided evidence for an extensive cover ofArtemisia shrubs, and Stipa, Poa, and Festuca
grasses. The presence of Artemisia shrubs at KP-1 is further con¢rmed by abundant cicada bur-rows in the Washtucna Soil complex (O'Geenand Busacca, 2001), the presence of ochric epipe-don, typical of aridisols in the Washtucna Soil(Tate, 1998), and the dry cold conditions inferredfrom the stable isotopes in KP-1 loess (Stevenson,1997). Further east, the CHR-1 site, at about thesame elevation as Carp Lake (Fig. 10), supporteda less xeric grassland with Poa, Festuca and someArtemisia, but no Stipa. The low Artemisia/grassratio suggests that CHR-1 received more moisturethan KP-1, located at the lower elevation. Condi-tions were colder and drier than today, as evi-denced by the lack of Agropyron and the presenceof Stipa and Poa. Another loess section, SAP-2,within the same vegetation zone today containsabundant cicada burrows in the Washtucna Soilcomplex, suggesting presence of `periglacialsteppe' with Artemisia (O'Geen and Busacca,2001). Although we did not ¢nd evidence forshrub steppe at CHR-1, the phytolith record sug-gests that some Artemisia was present in thegrassland.
At 1095 m elevation, the VM-1 site supportednon-analog conifer parkland with Artemisia,Abies, and Picea at the LGM. Apparently, treessurvived the LGM at this middle-slope site. Be-cause phytoliths register a more localized signalthan pollen, this con¢rms the earlier hypothesis(Mack et al., 1976; Barnosky et al., 1987) thatglacial refugia for conifer communities in the Co-lumbia Basin were located in the bordering high-lands, not in the lowlands to the south. Thus, ourdata con¢rm that at the LGM conifers were com-pressed between the expanded arid steppe in thelowlands and cold-adapted steppe higher in themountains (Barnosky, 1985a). Based on the phy-tolith evidence, the parkland zone apparently oc-cupied a narrow belt between 900 and 1150 m.
The RBM-1 site higher in the Blue Mountains(1220 m) supported an Artemisia/Stipa^Poa^Fes-tuca subalpine community, not unlike that foundin the Wallowa Mountains above 2200 m eleva-tion today. Presence of small amounts of coniferphytoliths in the LGM samples at RBM-1 sug-gests that the parkland with Abies and Piceawas not far away from the site, and scattered
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10196

conifers could have also grown locally. It is un-clear which species of grass contributed the un-usually high proportion of rounded keeled phyto-liths in the Washtucna Soil complex at this site.Cicada burrow data from another loess sectionnearby (RBM-2) did not indicate presence ofsagebrush near RBM-1 during the WashtucnaSoil complex interval (O'Geen and Busacca,2001), but it is unclear whether cicadas could sur-vive at this high elevation, regardless of the vege-tation composition. Thin sections of the formersoil surface at RBM-2 show soli£uction £owlobes, which is incompatible with existence of for-est.
Climate reconstructions at all ¢ve sites implythat the LGM was colder and drier than todayeverywhere in the Basin. This is consistent withpaleoclimate model simulations of the LGM forNorth America (COHMAP, 1988; Thompson etal., 1993; Bartlein et al., 1998). The simulatedPaci¢c Northwest climate was cold and dry as aresult of a strong temperature gradient south ofthe ice sheets, the southward displacement of thejet stream, and the development of a strong gla-cial anticyclone and easterly winds near the sur-face along the southern margin of the ice sheet.The phytolith data imply that the sites with sage-brush existed both at the high elevations and inthe lowlands. Sites at middle elevations (VM-1)supported some trees and were not as dry, andperhaps not as cold. The high-elevation siteswere colder both because of the orographic e¡ectand because of the greater exposure to the colddry easterly winds.
Phytolith data from the Columbia Basin haveimproved our understanding of the long-standingquestion about the nature of the full-glacial `tun-dra steppe' in the Columbia Basin (Barnosky,1985a). The reconstruction is supported by evi-dence from cicada burrows and stable isotopesat KP-1 and other sites across the Columbia Ba-sin. These data suggest that the lowlands sup-ported an extensive dry sagebrush shrub steppewith substantial presence of Stipa, Poa, and Fes-tuca grasses. This community was not a tundra ora grassland, but rather an arid shrubland withgrasses. Referring to this community as a `shrub
steppe' seems appropriate in view of our discoveryof abundant Stipa-type bilobates in the LGM-agedeposits. Clearly, the Stipa genus played a majorrole in the region during the late Pleistocene.Early Russian works on the origin of steppe de-¢ned Eurasian `steppe' as a biome based on thepresence of Stipa spp. (Alekhin, 1986).
5.4. Late Glacial interval (15^11 ka BP)
The magnitude of the climatic variations from15 to 11 ka was large (Thompson et al., 1993). Asthe ice sheets retreated from the full-glacial extent,the jet stream shifted north and brought moistureinto the region, the glacial anticyclone weakened,and easterly winds ceased (Bartlein et al., 1998).At the western edge of the Columbia Basin, CarpLake (Zone CL-3) at 714 m asl registers a briefperiod of abundant spruce (Picea) pollen ca.15^13 ka BP, followed by an abrupt increase ingrass and chenopod pollen ca. 13^10 ka BP (ZoneCL-2). The record suggests a shift from spruceparkland to temperate grassland. Phytoliths pro-vide additional information about the composi-tion of that grassland. Both KP-1 and CHR-1sites supported Festuca-dominated grassland dur-ing the Late Glacial, with the former site having amore xeric community (Festuca^Agropyron), com-pared to a more mesic community (Festuca^Koe-leria) at the latter site. Presence of spruce park-land near Carp Lake in the Late Glacial (15^13 kaBP) can be interpreted as an indication of coolerand wetter conditions than today. Similarly, Fes-tuca-dominated grassland at both KP-1 andCHR-1 implies cooler and wetter conditionsthan today. Cooler and wetter conditions werealso implied by phytoliths at VM-1 and RBM-1sites : the VM-1 site was Pinus ponderosa/Festucagrassland transition and RBM-1 supported Stipa^Festuca grassland, similar to the present-day sub-alpine community. Wetter conditions were in-ferred for 12 ka BP from all pollen records avail-able from the interior Paci¢c Northwest (Mack etal., 1978a,b,c; Thompson et al., 1993), as well asfrom the Puget Trough (Barnosky, 1981, 1985b;Barnosky et al., 1987) and western Oregon (Wor-ona and Whitlock, 1995).
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 97

5.5. Holocene (11^0 ka BP)
The early Holocene (11^7 ka BP) was an inter-val with a warm dry climate due to higher-than-present summer insolation and a strengtheningsubtropical high-pressure zone in the summer(Bartlein et al., 1998). All regional records fromthe Paci¢c Northwest indicate drier-than-presentconditions for the early (11 ka BP) and mid Ho-locene (6 7.5 ka BP) (Thompson et al., 1993).The Carp Lake (714 m asl) record indicates devel-opment of a grassland with chenopods in the vi-cinity of the lake between 13 and 9 ka BP, andPinus^Quercus woodland developed after 9 ka BP,implying drier and warmer conditions than today.Phytolith records from the Palouse support thisconclusion (Fig. 10). At KP-1, the Late GlacialFestuca-dominated grassland gave way to morexeric Agropyron-dominated grassland with Festu-ca idahoensis and Poa sandbergii in the early Ho-locene (ca. 9 TL ka). An increase in Agropyronindicates warm conditions, because today repre-sentatives of the genus (particularly the wide-spread Agropyron spicatum) are restricted to low-lands and middle elevations in the Paci¢cNorthwest (Hitchcock and Cronquist, 1994). Anincrease in sagebrush was observed at KP-1 in thephytolith sample from the mid Holocene (6 TLka), implying increasing dryness. Increases insagebrush abundance are also implied by in-creased abundance of cicada burrows in the midHolocene loess in SAP-2 and CLY-2 sections(O'Geen and Busacca, 2001). Both sections arelocated within the Agropyron^Festuca grasslandzone today, so the increase in sagebrush impliesdrier-than-present conditions. The increasing roleof Agropyron and the decreasing role of Koeleriaimply increasing aridity at CHR-1 from the LateGlacial to the early Holocene. CHR-1 supportedFestuca^Agropyron grassland during most of theHolocene, implying conditions similar to present.The two higher-elevation sites were forested dur-ing the Holocene. In the early Holocene, the Pinusponderosa forest zone was apparently located athigher elevations than today, with P. ponderosaplaying a major role in vegetation at the VM-1site. The site is above the P. ponderosa-dominatedforest today, although some pine is found near
the site. In the early Holocene, RBM-1 site sup-ported Agropyron-dominated grassland/P. ponde-rosa forest transition (or a parkland) implyingwarmer and drier conditions than today. Thepresent Abies grandis^Pseudotsuga forest was es-tablished at RBM-1 after 7.6 ka BP (the age ofMazama Ash). In the center of the Columbia Ba-sin, pollen records show development of drysteppe in the mid Holocene (Mehringer, 1985).Pinus ponderosa parkland was present in the Oka-nogan Highlands (Mack et al., 1978a,c).
6. Conclusions
Our study demonstrates the high potential ofphytolith analysis as a paleoecological tool.Changes in the proportions of di¡erent morpho-types were interpreted in terms of changes in veg-etation and climate during the last 100 000 years.Each of the three loess sections yielded abundantand distinct phytoliths at all depths. The lower-most site CHR-1 (677 m asl) is located within theFestuca idahoensis-dominated grassland today.Mesic grassland with Festuca was present duringan undated interval, likely in the early Wisconsin(s 75 ka). Between that period and prior to 37 kaBP, a shrub steppe with Artemisia existed at thesite. The site supported Festuca-dominated grass-land for the past 37 ka, including xeric Festuca^Poa grassland with some Artemisia during theLGM, a more mesic Festuca^Koeleria grasslandduring the late glacial interval (15^11 TL ka),and again more xeric Festuca^Agropyron grass-land in the Holocene (6 11 TL ka).
The intermediate site VM-1 (1095 m asl) sup-ports Abies grandis^Pseudotsuga forest today.Agropyron-dominated grassland existed at thesite in mid Wisconsin, and Pinus ponderosa forestwas at the site in the early Wisconsin. At theLGM, a non-analog community interpreted as aconifer parkland (with Abies, Picea, perhapsLarix) with some steppe elements (Artemisia,Stipa) existed at the site. The vegetation suggeststhat middle elevations within the Columbia Basinmay have supported trees during the LGM. Dur-ing the Late Glacial interval, conditions weremore moist and cool than at present, as suggested
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^10198

by the presence of subalpine Stipa^Festuca grass-land. The site supported a Pinus ponderosa forestduring the Holocene, which implies warmer, drierconditions.
The uppermost site RBM-1 (1200 m asl) sup-ports Abies grandis^Pseudotsuga forest with somePinus ponderosa today. A shrub steppe with Arte-misia shrubs and Stipa, Festuca, and Poa grasseswas present during the LGM. This communityappears similar to the one found at the sametime at low elevations (KP-1, CHR-1), based onthe phytolith record, while parkland persisted atVM-1. During the LGM, Artemisia shrubs andStipa were more abundant at the lower KP-1site than at CHR-1 or RBM-1. During the LateGlacial, Stipa^Festuca grassland was present atthe site, implying conditions close to thepresent-day subalpine. Agropyron-dominatedgrassland/Pinus ponderosa transition existed atthe site in the early Holocene. Phytolith datafrom the eastern part of the Columbia Basinand pollen record from Carp Lake (714 m asl)on the western margin of the Columbia Basinsuggest that sagebrush steppe existed across theColumbia Basin at low elevations at LGM. Phy-tolith and pollen data also suggest that conifertrees (Abies, Picea) were found at middle eleva-tions. Use of phytoliths allowed identi¢cation ofgrass genera that were present in the ColumbiaBasin at the LGM: Stipa, Poa, and Festuca.
Acknowledgements
The authors wish to express sincere thanks toEric McDonald, who contributed to the excava-tion and description of the loess sections, andBryan Stevenson and Eugene Kelly, who providedprocessed fossil phytolith samples and contributedto other aspects of this research project. We alsowish to thank Patrick Bartlein, Patricia McDowell,Gregory Retallack, Matt Wooller and two anon-ymous reviewers for their helpful comments on anearlier version of the manuscript. This paperpresents some of the results of Ph.D. dissertationresearch supported by Mazama and Sigma XiResearch Grants and Graduate Research Fellow-
ship of the University of Oregon to M.B., andNSF grants to C.W. and A.B.
References
Alekhin, V.V., 1986. Teoreticheskie problemy ¢totsenologii istepevedeniya (Theoretical problems of phytocoenology andsteppe science). Izdatel'stvo Moskovskogo Universiteta,Moscow (in Russian).
Alley, N.F., Hicock, S.R., 1986. The stratigraphy, palynology,and climatic signi¢cance of pre-middle Wisconsin Pleisto-cene sediments, southern Vancouver Island, British Colum-bia. Can. J. Earth Sci. 23, 369^383.
Alley, N.F., Valentine, K.W.G., Fulton, R.J., 1986. Paleocli-matic implications of middle Wisconsin pollen and a paleo-sol from the Purcell Trench, south-central British Columbia.Can. J. Earth Sci. 23, 1156^1168.
Anderson, P.M., Bartlein, P.J., Brubaker, L.B., Gajewski, K.,Ritchie, J.C., 1989. Modern analogues of late-Quaternarypollen spectra from the western interior of North America.J. Biogeogr. 16, 573^596.
Bacon, C.R., 1983. Eruptive history of Mt. Mazama and Cra-ter Lake caldera, Cascade Range, U.S.A. J. Volcanol. Geo-therm. Res. 18, 57^115.
Baker, V.R., Bunker, R.C., 1985. Cataclysmic Late Pleistocene£ooding from Glacial Lake Missoula: A review. Quat. Sci.Rev. 5, 1^41.
Baker, V.R., Bjornstad, B.N., Busacca, A.J., Fecht, K.R.,Kiver, E.P., Moody, U.L., Rigby, J.G., Stradling, D.F.,Tallman, A.M., 1991. Quaternary geology of the ColumbiaPlateau. In: Morrison, R. (Ed.), Quaternary Non-glacialGeology of the Conterminous United States. Geological So-ciety of America, Boulder, CO, pp. 215^250.
Barnosky, C.W., 1981. A record of late Quaternary vegetationfrom Davis Lake, southern Puget Lowland, Washington.Quat. Res. 16, 221^239.
Barnosky, C.W., 1985a. Late Quaternary vegetation in thesouthwestern Columbia Basin, Washington. Quat. Res. 23,109^122.
Barnosky, C.W., 1985b. Late Quaternary vegetation near Bat-tle Ground Lake, southern Puget Trough, Washington.Geol. Soc. Am. Bull. 96, 263^271.
Barnosky, C.W., Anderson, P.M., Bartlein, P.J., 1987. Thenorthwestern U.S. during deglaciation: Vegetational historyand paleoclimatic implications. In: Ruddiman, W.F.,Wright, Jr., H.E. (Eds.), North America and AdjacentOceans During the Last Deglaciation. Geological Societyof America, Boulder, CO, pp. 289^332.
Bartlein, P.J., Anderson, K.H., Anderson, P.M., Edwards,M.E., Mock, C.J., Thompson, R.S., Webb, R.S., Webb,T., III, Whitlock, C, 1998. Paleoclimate simulations forNorth America over the past 21,000 years: features of thesimulated climate and comparisons with paleoenvironmentaldata. Quat. Sci. Rev. 17, 549^585.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 99

Beiswenger, J.M., 1991. Late Quaternary vegetational historyof Grays Lake, Idaho. Ecol. Monogr. 61, 165^182.
Berger, G.W., Busacca, A.J., 1995. Thermoluminescence dat-ing of late Pleistocene loess and tephra from eastern Wash-ington and southern Oregon and implications for eruptivehistory of Mount St. Helens. J. Geophys. Res. 100 (B11),22361^22374.
Blinnikov, M., 1994. Phytolith analysis and the Holocene dy-namics of alpine vegetation. In: Onipchenko, V., Blinnikov,M. (Eds.), Experimental Investigation of Alpine Plant Com-munities in the Northwestern Caucasus. Vero«¡entlichungendes Geobotanischen Institutes der ETH, Stiftung Ru«bel,Zu«rich, Vol. 115, pp. 23^40.
Blinnikov, M.S., 1999. Late-Pleistocene History of the Colum-bia Basin Grassland Based on Phytolith Records in Loess.Ph.D. Dissertation, University of Oregon, Eugene, OR.
Blinnikov, M., Busacca, A., Whitlock, C., 2001. A new100,000-year record from the Columbia Basin, Washington,U.S.A. In: Meunier, J.D., Colin, F. (Eds.), Phytoliths ^Applications in Earth Science and Human History. A.A.Balkema, Rotterdam.
Bretz, J.H., 1969. The lake Missoula Floods and the Chan-neled Scablands. J. Geol. 77, 505^543.
Busacca, A.J., 1989. Long Quaternary record in Eastern Wash-ington, USA, interpreted from multiple buried paleosols inloess. Geoderma 45, 105^122.
Busacca, A.J., McDonald, E.V., 1994a. Regional sedimenta-tion of late Quaternary loess of the Columbia Plateau: sedi-ment source areas and loess distribution patterns. Bull.Washington Div. Geol. Earth Resources 80, 181^190.
Busacca, A.J., McDonald, E.V., The Geomorphology and Pa-leopedology of the Palouse and Channeled Scabland. Guide-book Prepared for the Soil Science Society of America An-nual Meeting, Seattle, WA, November 13^18, 1994.
Busacca, A.J., Nelstead, K.T., McDonald, E.V., Purser, M.D.,1992. Correlation of distal tephra layers of loess in theChanneled Scabland and Palouse of Washington State.Quat. Res. 37, 281^303.
Clague, J.J., 1978. Mid-Wisconsin climates of the Paci¢cNorthwest. Geol. Surv. Can. Pap. 78 (1B), 95^100.
Clague, J.J., Easterbrook, D.J., Hughes, O.L., Matthews, J.V.Jr., 1992. The Sangamonian and Early Wisconsinian stagesin western Canada and northwestern United States. In:Clark, P., Lea, P.D. (Eds.), The Last Interglacial-GlacialTransition in North America. Geological Society of AmericaSpecial Paper 270, Boulder, CO, pp. 253^268.
COHMAP Members, 1988. Climatic changes in the last 18,000years: Observations and model simulations. Science 241,1043^1052.
Coupland, R.T., 1992. Overview of the grassland of NorthAmerica. In: Natural Grasslands: Introduction and WesternHemisphere, Ecosystems of the World, Vol. 8A. Elsevier,New York, pp. 147^149.
Crandell, D.R., Mullineaux, D.R., Rubin, M., Soiker, E.,Kelly, M.L., 1981. Radiocarbon dates from volcanic depos-its at Mount St. Helens, Washington. US Geol. Surv. OpenFile Rep. 81, 844.
Daubenmire, R.F., 1968. Forest Vegetation of Eastern Wash-ington and Northern Idaho. Technical Bulletin of the Wash-ington Agricultural Experimental Station 60, Pullman, WA.
Daubenmire, R.F., 1970. Steppe Vegetation of Washington.Technical Bulletin of the Washington Agricultural Experi-mental Station 62, Pullman, WA.
Franklin, J.F., Dyrness, C.T., 1988. Natural Vegetation ofOregon and Washington. Oregon State University Press,Corvallis, OR.
Fredlund, G.G., Tieszen, L.L., 1994. Modern phytolith assem-blages from the North American Great Plains. J. Biogeogr.21, 321^335.
Fredlund, G.G., Tieszen, L.L., 1997. Phytolith and carbonevidence for Late Quaternary vegetation and climate changein the Southern Black Hills, South Dakota. Quat. Res. 47,206^217.
GRIP (Greenland Ice-Core Project) Members, 1993. Climateinstability during the last interglacial period recorded in theGRIP ice core. Nature 364, 203^207.
Grimm, E.C., 1993. TILIAGRAPH Version 1.8. Illinois StateMuseum Research and Collection Center, Spring¢eld, IL.
Heusser, C.J., 1985. Quaternary pollen records from the Paci¢cNorthwest coast: Aleutians to the Oregon-California bound-ary. In: Pollen Records of Late Quaternary North AmericanSediments. American Association of Stratigraphic Palynolo-gists Foundation, Boulder, CO, pp. 141^165.
Heusser, L.E., King, J.E., 1988. North America: with specialemphasis on the development of the Paci¢c coastal forestand prairie/forest boundary prior to the Last Glacial Max-imum. In: Huntley, B., Webb, T. III (Eds.), Vegetation His-tory. Kluwer, Dordrecht, pp. 193^211.
Heusser, C.J., Heusser, L.E., 1990. Long continental pollensequence from Washington State (USA): correlation ofupper levels with marine pollen-oxygen isotope stratigraphythrough substage 5e. Palaeogeogr. Palaeoclimatol. Palaeo-ecol. 79, 63^71.
Hitchcock, L., Cronquist, A., 1994. Flora of the Paci¢c North-west. University of Washington Press, Seattle, WA.
Kelly, E.F., 1990. Methods for Extracting Opal Phytolithsfrom Soil and Plant Material. Department of Agronomy,Colorado State University, Fort Collins, CO.
Kiseleva, N.K., 1982. Phytolith soil studies as a tool for theinvestigation of the history of East Mongolian steppes. Izv.Akad. Nauk SSSR, Ser. Geogr. 2, 95^106.
Ku«chler, A.W., 1964. Manual to Accompany the Map of Po-tential Natural Vegetation of the Conterminous UnitedStates. American Geographic Society Special Publication 36.
Kukla, G.J., Opdyke, N.D., 1980. Matuyama loess at Colum-bia Plateau, Washington. AMQUA Abstracts with Pro-grams 6, 122.
Leopold, E.B., Denton, M.F., 1987. Comparative age of grass-lands and steppe east and west of the northern RockyMountains. Ann. Missouri Bot. Gard. 74, 841^867.
Mack, R.N., Bryant, V.M., Jr., Fryxell, R., 1976. Pollen se-quence from the Columbia Basin, Washington: reappraisalof post-glacial vegetation. Am. Midland Nat. 95, 390^397.
Mack, R.N., Rutter, N.W., Bryant, V.M., Jr., Valastro, S.,
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101100

1978a. Late Quaternary pollen record for Big Meadow,Pend Oreille County, Washington. Ecology 59, 956^966.
Mack, R.N., Rutter, N.W., Valastro, S., 1978b. Late Quater-nary pollen record from the Sandpoil River Valley, Wash-ington. Can. J. Bot. 56, 1642^1650.
Mack, R.N., Rutter, N.W., Valastro, S., Bryant, V.M., Jr.,1978c. Late Quaternary vegetation history at Waits Lake,Colville River Valley, Washington. Bot. Gaz. 139, 499^506.
Martinson, D.G., Pisias, N.G., Hays, J.D., Imbrie, J., Moore,T.C., Jr., Shackleton, N.J., 1987. Age dating and the orbitaltheory of the ice ages: development of a long resolution 0 to300, 000-year chronostratigraphy. Quat. Res. 27, 1^29.
McDonald, E.V., Busacca, A.J., 1988. Record of pre-late Wis-consin giant £oods in the Channeled Scabland interpretedfrom loess deposits. Geology 16, 728^731.
McDonald, E.V., Busacca, A.J., 1990. Interaction between ag-grading geomorphic surfaces and the formation of a latePleistocene paleosol in the Palouse loess of eastern Wash-ington State. Geomorphology 3, 449^470.
McDonald, E.V., Busacca, A.J., 1992. Late Quaternary strat-igraphy of loess in the Channeled Scabland and Palouseregions of Washington State. Quat. Res. 38, 141^156.
Mehringer, P.J. Jr., 1985. Late Quaternary Pollen Recordsfrom the Interior Paci¢c Northwest and Northern GreatBasin of the United States. In: Pollen Records of Late Qua-ternary North American Sediments. American Associationof Stratigraphic Palynologists Foundation, Boulder, CO, pp.167^190.
Norgren, J.A., 1973. Distribution, Form and Signi¢cance ofPlant Opal in Oregon Soils. Ph.D. Dissertation, OregonState University, Corvallis, OR.
O'Geen, A.T., Busacca, A.J., 2001. Late Quaternary paleoen-vironments of the Columbia Plateau, PNW U.S., from fau-nal burrows in paleosols. Palaeogeography Palaeoclimatol-ogy Palaeoecology 169, 23^37.
Overpeck, J.T., Prentice, I.C., Webb, T., III, 1985. Quantita-tive interpretation of fossil pollen spectra: Dissimilarity co-e¤cients and the method of modern analogs. Quat. Res. 23,87^108.
Pearsall, D.M., 1989. Paleoethnobotany: A Handbook of Pro-cedures. Academic Press, San Diego, CA.
Piperno, D.R., 1988. Phytolith Analysis: An Archeologicaland Geological Perspective. Academic Press, San Diego,CA.
Richardson, C.A., McDonald, E.V., Busacca, A.J., 1997. Lu-
minescence dating of loess from the Northwest UnitedStates. Quat. Sci. Rev. 16, 403^415.
Richardson, C.A., McDonald, E.V., Busacca, A.J., 1999. Aluminescence chronology for loess deposition in WashingtonState and Oregon USA. Z. Geomorphol. 116, 77^95.
Stevenson, B.A., 1997. Stable Carbon and Oxygen Isotopes inSoils and Paleosols of the Palouse Loess, Eastern Washing-ton State: Modern Relationships and Applications for Pa-leoclimatic Reconstruction. Ph.D. Dissertation, ColoradoState University, Fort Collins, CO.
Tate, T.A., 1998. Micromorphology of Loessial Soils and Pa-leosols on Aggrading Landscapes of the Columbia Plateau.M.S. Thesis, Washington State University, Pullman, WA.
Thompson, R.S., 1992. Late Quaternary environments inRuby Valley, NV. Quat. Res. 37, 1^15.
Thompson, R.S., Whitlock, C., Bartlein, P.J., Harrison, S.P.,Spaulding, W.G., 1993. Climatic changes in the westernUnited States since 18,000 yr B.P. In: Wright, H.E. Jr.,Kutzbach, J.E., Webb, T. III, Ruddiman, W.F., Street-Per-rott, F.A., Bartlein, P.J. (Eds.), Global Climates Since theLast Glacial Maximum. University of Minnesota Press,Minneapolis, MN, pp. 468^513.
Waitt, R.B. Jr., Thorson, R.M., 1983. The Cordilleran icesheet in Washington, Idaho, and Montana. In: Wright,H.E. Jr. (Ed.), The Late Quaternary Environments of theUnited States. Volume I. Late Pleistocene. University ofMinnesota Press, Minneapolis, MN, pp. 53^70.
Webb, T. III, Ruddiman, W.F., Street-Perrott, F.A., Mark-graf, V., Kutzbach, J.E., Bartlein, P.J., Wright, H.E. Jr.,Prell, W.L., 1993. Climatic changes during the past 18,000years: regional syntheses, mechanisms and causes. In:Wright, H.E. Jr., Kutzbach, J.E., Webb, T. III, Ruddiman,W.F., Street-Perrott, F.A., Bartlein, P.J. (Eds.), Global Cli-mates Since the Last Glacial Maximum. University of Min-nesota Press, Minneapolis, MN, pp. 514^535.
Whitlock, C., 1992. Vegetational and climatic history of thePaci¢c Northwest during the last 20,000 years: Implicationsfor understanding present-day biodiversity. Northwest Envi-ron. J. 8, 5^28.
Whitlock, C., Bartlein, P.J., 1997. Vegetation and climatechange in northwest America during the past 125 kyr. Na-ture 388, 57^61.
Worona, M., Whitlock, C., 1995. Late Quaternary vegetationand climate history near Little Lake, central Coast Range,Oregon. Geol. Soc. Am. Bull. 107, 867^876.
PALAEO 2714 22-1-02
M. Blinnikov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 177 (2002) 77^101 101
Copyright © 2022 FDOKUMEN




















