
RecA Proteins from Deinococcus geothermalis and Deinococcus murrayi - Cloning, Purification and...
13
RESEARCH ARTICLE Open Access RecA Proteins from Deinococcus geothermalis and Deinococcus murrayi - Cloning, Purification and Biochemical Characterisation Marta Wanarska, Beata Krawczyk, Piotr Hildebrandt and Józef Kur * Abstract Background: Escherichia coli RecA plays a crucial role in recombinational processes, the induction of SOS responses and mutagenic lesion bypasses. It has also been demonstrated that RecA protein is indispensable when it comes to the reassembly of shattered chromosomes in g-irradiated Deinococcus radiodurans, one of the most radiation-resistant organisms known. Moreover, some functional differences between E. coli and D. radiodurans RecA proteins have also been shown. Results: In this study, recA genes from Deinococcus geothermalis and Deinococcus murrayi, bacteria that are slightly thermophilic and extremely g-radiation resistant, were isolated, cloned and expressed in E. coli. After production and purification, the biochemical properties of DgeRecA and DmuRecA proteins were determined. Both proteins continued to exist in the solutions as heterogenous populations of oligomeric forms. The DNA binding by DgeRecA and DmuRecA proteins is stimulated by Mg 2+ ions. Furthermore, both proteins bind more readily to ssDNA when ssDNA and dsDNA are in the same reaction mixture. Both proteins are slightly thermostable and were completely inactivated in 10 s at 80°C. Both proteins hydrolyze ATP and dATP in the presence of ssDNA or complementary ssDNA and dsDNA, but not in the absence of DNA or in the presence of dsDNA only, and dATP was hydrolyzed more rapidly than ATP. They were also able to promote DNA strand exchange reactions by a pathway common for other RecA proteins. However, we did not obtain DNA strand exchange products when reactions were performed on an inverse pathway, characteristic for RecA of D. radiodurans. Conclusions: The characterization of DgeRecA and DmuRecA proteins made in this study indicates that the unique properties of D. radiodurans RecA are probably not common among RecA proteins from Deinococcus sp. Background Deinococcus geothermalis DSM 11302 and Deinococcus murrayi DSM 11303 are gram positive, nonmotile, sphe- rical bacteria living in aerobic conditions. Cells that divide as tetrads are very common in both species. D. geothermalis and D. murrayi form orange-pigmented colonies, are slightly thermophilic with an optimum growth temperature of between 45-50°C, but differ in optimum pH for growth. D. geothermalis DSM 11302 is slightly acidophilic and grows optimally at pH 6.5, while D. murrayi DSM 11303 is slightly alcaliphilic with an optimum pH for growth of 8.0, although both species were isolated from the hot springs which had alkaline pH values ranging from 8.6 to 8.9. D. geothermalis and D. murrayi were isolated from hot springs at São Pedro do Sul and Alcafache in central Portugal, respectively. The isolation of the acidophilic D. geothermalis strain from an alkaline site suggests that it can colonize the microenvironments of alkaline hot springs, such as bio- films, where the pH is lowered by other microorganisms [1]. D. geothermalis is also able to grow on metallic sur- faces of printing paper machines and it is known as an efficient primary biofilm, formerly functioning as an adhesion platform for secondary biofilm bacteria [2-4]. D. geothermalis and D. murrayi display an increased gamma radiation resistance, as would normally be found in the genus Deinococcus [1]. The ability of these species to withstand high doses of ionizing radiation might result from an efficient RecA-dependent DSB repair * Correspondence: [email protected] Department of Microbiology, Chemical Faculty, Gdańsk University of Technology, Narutowicza 11/12, 80-233 Gdańsk, Poland Wanarska et al. BMC Molecular Biology 2011, 12:17 http://www.biomedcentral.com/1471-2199/12/17 © 2011 Wanarska et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of RecA Proteins from Deinococcus geothermalis and Deinococcus murrayi - Cloning, Purification and...

RESEARCH ARTICLE Open Access
RecA Proteins from Deinococcus geothermalis andDeinococcus murrayi - Cloning, Purification andBiochemical CharacterisationMarta Wanarska, Beata Krawczyk, Piotr Hildebrandt and Józef Kur*
Abstract
Background: Escherichia coli RecA plays a crucial role in recombinational processes, the induction of SOSresponses and mutagenic lesion bypasses. It has also been demonstrated that RecA protein is indispensable whenit comes to the reassembly of shattered chromosomes in g-irradiated Deinococcus radiodurans, one of the mostradiation-resistant organisms known. Moreover, some functional differences between E. coli and D. radioduransRecA proteins have also been shown.
Results: In this study, recA genes from Deinococcus geothermalis and Deinococcus murrayi, bacteria that are slightlythermophilic and extremely g-radiation resistant, were isolated, cloned and expressed in E. coli. After productionand purification, the biochemical properties of DgeRecA and DmuRecA proteins were determined. Both proteinscontinued to exist in the solutions as heterogenous populations of oligomeric forms. The DNA binding byDgeRecA and DmuRecA proteins is stimulated by Mg2+ ions. Furthermore, both proteins bind more readily tossDNA when ssDNA and dsDNA are in the same reaction mixture. Both proteins are slightly thermostable and werecompletely inactivated in 10 s at 80°C. Both proteins hydrolyze ATP and dATP in the presence of ssDNA orcomplementary ssDNA and dsDNA, but not in the absence of DNA or in the presence of dsDNA only, and dATPwas hydrolyzed more rapidly than ATP. They were also able to promote DNA strand exchange reactions by apathway common for other RecA proteins. However, we did not obtain DNA strand exchange products whenreactions were performed on an inverse pathway, characteristic for RecA of D. radiodurans.
Conclusions: The characterization of DgeRecA and DmuRecA proteins made in this study indicates that the uniqueproperties of D. radiodurans RecA are probably not common among RecA proteins from Deinococcus sp.
BackgroundDeinococcus geothermalis DSM 11302 and Deinococcusmurrayi DSM 11303 are gram positive, nonmotile, sphe-rical bacteria living in aerobic conditions. Cells thatdivide as tetrads are very common in both species. D.geothermalis and D. murrayi form orange-pigmentedcolonies, are slightly thermophilic with an optimumgrowth temperature of between 45-50°C, but differ inoptimum pH for growth. D. geothermalis DSM 11302 isslightly acidophilic and grows optimally at pH 6.5, whileD. murrayi DSM 11303 is slightly alcaliphilic with anoptimum pH for growth of 8.0, although both specieswere isolated from the hot springs which had alkaline
pH values ranging from 8.6 to 8.9. D. geothermalis andD. murrayi were isolated from hot springs at São Pedrodo Sul and Alcafache in central Portugal, respectively.The isolation of the acidophilic D. geothermalis strainfrom an alkaline site suggests that it can colonize themicroenvironments of alkaline hot springs, such as bio-films, where the pH is lowered by other microorganisms[1]. D. geothermalis is also able to grow on metallic sur-faces of printing paper machines and it is known as anefficient primary biofilm, formerly functioning as anadhesion platform for secondary biofilm bacteria [2-4].D. geothermalis and D. murrayi display an increasedgamma radiation resistance, as would normally be foundin the genus Deinococcus [1]. The ability of these speciesto withstand high doses of ionizing radiation mightresult from an efficient RecA-dependent DSB repair
* Correspondence: [email protected] of Microbiology, Chemical Faculty, Gdańsk University ofTechnology, Narutowicza 11/12, 80-233 Gdańsk, Poland
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
© 2011 Wanarska et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the CreativeCommons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, andreproduction in any medium, provided the original work is properly cited.

system, similar to that recently described in Deinococcusradiodurans [5-7].RecA protein is a crucial DNA dependent ATPase
involved in DNA repair and homologous recombination.RecA proteins are found in most microorganisms withinthe Bacteria domain, but some insects’ and clams’ endo-cellular bacterial symbionts such as Buchnera aphidicolaAPS, B. aphidicola Sg, Blochmannia floridanus, B. penn-sylvanicus, Wigglesworthia glossinidia, Vesiomyosociusokutanii and Ruthia magnifica lack the recA gene[8-13]. Its analogues such as RadA [14] or Rad51 [15,16]are common in Archaea and Eucarya domains organ-isms. The product of the uvsX gene of the bacteriophageT4 also displays many RecA-like properties [17].RecA protein of E. coli, the best characterized RecA, is
a multifunctional protein involved in homologousrecombination [18], recombinational DNA repair [19]and SOS response to DNA damage and arrest of DNAreplication [20]. RecA filaments on ssDNA acts as acoprotease which facilitates the autoproteolysis of LexAprotein. This results in the derepression of genes in theSOS regulon [21,22]. RecA coprotease activity also facili-tates the autocatalytic cleavage of the UmuD protein tothe activated UmuD’, a component of DNA polymeraseV (UmuD’2C) [23-27]. Moreover RecA nucleoproteinfilament transfers RecA-ATP complex to polymerase Vto form an active mutasome UmuD’2C-RecA-ATPwhich catalyzes translesion DNA synthesis [28].In vitro, in the presence of Mg2+ ions and ATP, dATP
or nonhydrolyzable ATP analogue ATP-g-S, RecAassembles around single-stranded DNA into a catalycallyactive helical filaments [29-32]. No ATP or dATPhydrolysis is needed for nucleoprotein filament forma-tion, although these nucleotides are hydrolyzed by RecAin the presence of ssDNA [33], during the disassemblyof filaments [34]. The active RecA filament is able tosearch out a homology between bound, single-strandedDNA and double-stranded molecules, and catalyzes thehomologous pairing of DNA stands. These reactionsalso do not require cofactor hydrolysis and can occur inthe presence of ATP-g-S [35,36]. The RecA protein of E.coli promotes both three-strand exchange reactionbetween homologous ssDNA and dsDNA molecules,and four-strand exchange between a duplex DNA with asingle-stranded tail and a full dsDNA, where the strandexchange reaction is initiated in the single-strandedregion [37,38]. Although homologous pairing and DNAstrand exchange can occur in the three-strand exchangereaction without ATP hydrolysis [36], ATP hydrolysisrenders RecA protein-mediated DNA strand exchangeunidirectional (5’ to 3’ with respect to the single-stranded DNA). In the presence of ATP-g-S DNAstrand exchange is bidirectional and limited in extent[39]. Moreover, ATP hydrolysis allows a heterologous
sequence bypass in one of the DNA substrates [40,41],and is indispensable in the four-strand exchange reac-tion [41,42]. In contrast to E. coli RecA, the RecA pro-tein of D. radiodurans is able to promote the DNAstrand exchange through an inverse pathway, where thedouble-stranded DNA is bound first and the homolo-gous single-stranded DNA second [43].The aim of the present study was to clone, sequence
and overexpress D. geothermalis DSM 11302 and D.murrayi DSM 11303 recA genes in E. coli. A biochem-ical characterization of recombinant DgeRecA andDmuRecA proteins was performed.
ResultsCloning, expression and purification of D. geothermalisand D. murrayi RecA proteinsThe primers for amplification of D. geothermalis DSM11302 recA gene were designed on the basis of theknown recA gene sequence of D. geothermalis DSM11300 [GenBank: CP000359]. The obtained PCR pro-duct was cloned into a PCR-Blunt vector andsequenced. The nucleotide sequence of D. geothermalisDSM 11302 recA gene is available from the GenBankdatabase under accession number EF447285.The predicted DgeRecA monomer protein contains
358 amino acid residues. A homology search performedusing a version 3 FASTA programme at the EBI (Eur-opean Bioinformatics Institute) revealed that the aminoacid sequence of D. geothermalis DSM 11302 RecAshares 100% identity with D. geothermalis DSM 11300RecA protein [GenBank: ABF46432], 87.6% identity and95.9% similarity with D. radiodurans R1 RecA protein[GenBank: AAF11887], 71.1% identity and 91.6%similarity with Thermus aquaticus YT-1 RecA protein[GenBank: AAA19796], 71.0% identity and 89.8% similar-ity with T. thermophilus HB8 or T. thermophilus HB27RecA proteins [GenBank: BAD71641 and AAS81808,respectively], 70.6% identity and 89.8% similarity withMeiothermus ruber DSM 1279 RecA protein [GenBank:ADD27511], 67.6% identity and 87.2% similarity withM. silvanus DSM 9946 RecA protein [GenBank:ADH62770], and 60.9% identity and 84.1% similarity withE. coli RecA protein [GenBank: CAA23618].The D. murrayi DSM 11303 recA gene sequence was
obtained using a two step procedure. In first step, theinternal fragment of the recA gene was amplified usingdegenerated primers designed on the basis of an align-ment of recA gene sequences from bacteria belonging tothe Deinococcus-Thermus group. In the second step,flanking regions were amplified by inverse PCR. Theobtained partial sequences of the D. murrayi recA genewere then aligned and the primers for amplification ofthe gene were designed. Afterwards, the PCR productwas cloned using a CloneJET™ PCR Cloning Kit and
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 2 of 13

sequenced. The nucleotide sequence of D. murrayi DSM11303 recA gene was deposited in the GenBank databaseunder accession number HM004587.The DmuRecA protein contains 359 amino acid resi-
dues. The deduced amino acid sequence of D. murrayiDSM 11303 RecA shows 94.4% identity and 97.8% simi-larity with D. geothermalis DSM 11300 RecA protein,86.9% identity and 96.9% similarity with D. radioduransR1 RecA protein, 71.4% identity and 90.7% similaritywith Thermus aquaticus YT-1 RecA protein, 70.7%identity and 89.2% similarity with T. thermophilus HB8RecA protein, 70.3% identity and 89.5% similarity withMeiothermus ruber DSM 1279 RecA protein, 69.6%identity and 87.8% similarity with T. thermophilus HB27RecA protein, 67.0% identity and 85.7% similarity withM. silvanus DSM 9946 RecA protein, and 62.3% identityand 86.0% similarity with E. coli RecA protein. The mul-tiple sequence alignment of RecA proteins from bacteriaof the genus Deinococcus, Thermus and Meiothermus,and E. coli RecA protein is shown in Figure 1.In the next step, recombinant plasmids (pET-30Ek/
LIC-DgeRecA and pET-30Ek/LIC-DmuRecA for bio-synthesis of RecA proteins from D. geothermalis and D.murrayi in E. coli T7 expression system) were con-structed. After recA gene expression in the E. coli BLR(DE3) (recA-) cells, DgeRecA protein was purified byheat treatment of lysate at 60°C for 20 min and ionexchange chromatography. In the case of purification ofDmuRecA protein heat treatment was omitted due to itslow thermostability. The purity of DgeRecA and DmuR-ecA proteins at every step of production and purifica-tion was checked by SDS-PAGE after CoomassieBrilliant Blue R staining (Figure 2).The applied overexpression and purification systems
produced about 63 mg of DgeRecA and 50 mg ofDmuRecA proteins from 1 L of E. coli culture.Analysis of the purified proteins by SDS-PAGE
revealed major bands with a molecular mass of about 40kDa for both DgeRecA and DmuRecA proteins (Figure2, lanes 6 and 11), which agreed with the amino acidsequence calculation; 38.157 kDa and 38.170 kDa,respectively. Summary of the purification process isshown in Table 1.
Oligomeric states of DgeRecA and DmuRecA proteinsOligomeric states of DgeRecA and DmuRecA proteins ata concentration range between 3 and 58 μM in 25 mMpotassium phosphate buffer pH 7.5 containing 1 M KClwere analyzed by gel filtration. The oligomeric states ofexamined proteins depend highly on their level of con-centration. At high concentrations in solutions (58 and29 μM) both RecA proteins exist as a heterogenouspopulation of oligomeric forms ranging in size fromdimers (elution volume near 14 ml) to long protein
filaments and highly aggregated structures (elution nearthe void volume of 8.25 ml). Furthermore in such condi-tions, a greater percentage of DgeRecA and DmuRecAproteins exist as big oligomers, although in the case ofDmuRecA significant amounts of dimers and trimers(elution volume near 14 and 13 ml, respectively) werealso present. At 6 and 3 μM concentrations both pro-teins eluted at volumes corresponding mainly to thesmall oligomers and probably monomers in the case ofDmuRecA protein (Figure 3).
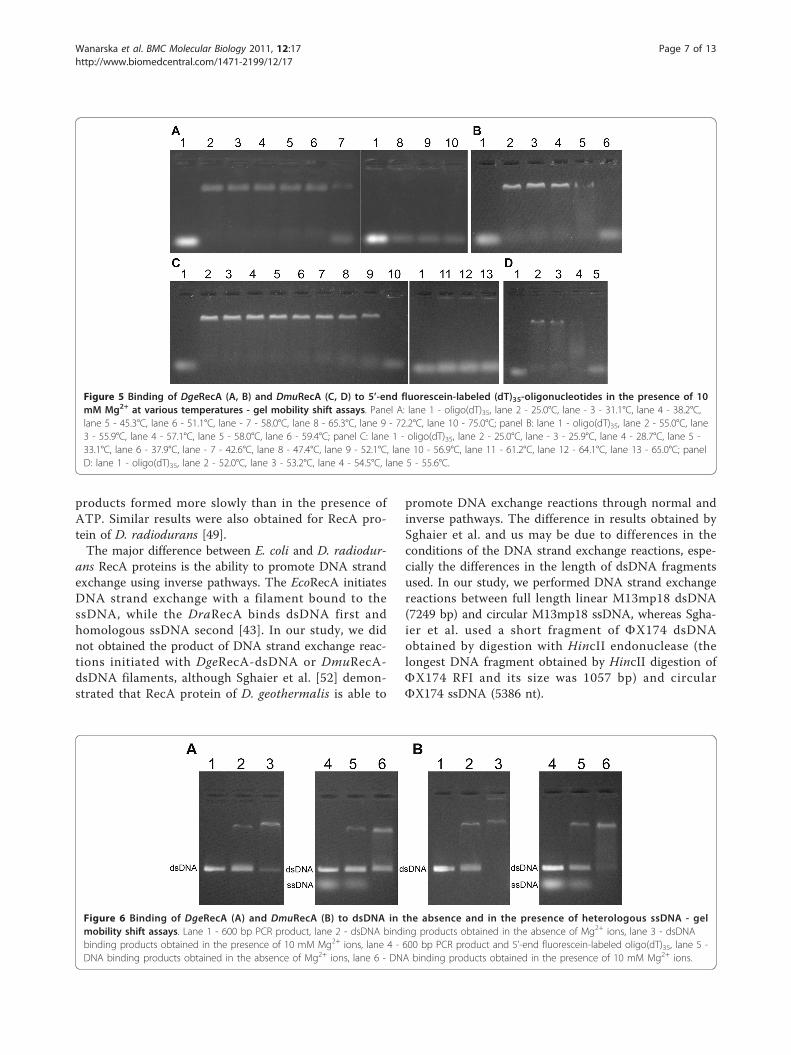
ssDNA-binding propertiesTo determine the ability of DgeRecA and DmuRecA pro-teins to bind ssDNA, we carried out agarose gel mobilityassays with 5’-end fluorescein-labelled (dT)35-oligonu-cleotides. The assays were carried out with increasingconcentration of Mg2+ ions and at various temperaturesbetween 25 and 75°C. We observed, that ssDNA bindingby RecA proteins was stimulated by Mg2+ ions and wasthe most efficient at 8 and 10 mM Mg2+ (Figure 4).DgeRecA was able to bind oligo(dT)35 at temperaturesbetween 25 and 58°C while DmuRecA exhibited activitybetween 25 and 54.5°C, although at the highest tempera-tures, 58°C for DgeRecA and 54.5°C for DmuRecA, thessDNA binding was very slight (Figure 5).
dsDNA-binding propertiesWe examined also the ability of RecA proteins from D.geothermalis and D. murrayi to bind dsDNA in theabsence of magnesium ions and in the presence of 10mM Mg2+. Both proteins were able to bind 600 bp PCRproducts and the dsDNA binding was stimulated byMg2+ ions as well as in the case of ssDNA binding (Fig-ure 6A and 6B, lanes 2 and 3). Afterwards, to determinewhat kind of DNA is more readily bound by DgeRecAand DmuRecA proteins we carried out the reactionswhere 5’-end fluorescein-labelled oligo(dT)35 and 600 bpPCR product were in the same test tube. When ssDNAand dsDNA were included in the reaction mixture,ssDNA was bound preferentially by both RecA proteins,both in the absence and presence of magnesium ions(Figure 6A and 6B, lanes 5 and 6). We observed alsothat DmuRecA bound dsDNA more efficient than DgeR-ecA in all conditions tested.
Thermostability of DgeRecA and DmuRecA proteinsThermostability of RecA proteins was characterized asthe longest time period needed for complete loss of visi-ble ssDNA binding activity investigated by gel-mobilityshift assay after incubation at given temperature. DgeR-ecA and DmuRecA were incubated at temperaturesbetween 50 and 80°C for 10 s - 180 minutes, in theabsence or in the presence of 10 mM Mg2+. No signifi-cant loss of protein activity was observed after
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 3 of 13

incubation at 50°C for 180 min. DgeRecA was also stablefor 180 min at 57°C, while DmuRecA lost activity within15 min at this temperature. Both proteins incubated athigher temperatures displayed lower thermostability andat 80°C were completely inactivated in 10 s. The RecAprotein from D. geothermalis was more thermostablethan RecA from D. murrayi (Table 2). Moreover magne-sium ions have no effect on the thermal stability neitherDgeRecA nor DmuRecA.
ATP and dATP hydrolysisATP and dATP hydrolysis catalyzed by RecA proteinsfrom D. geothermalis and D. murrayi was monitoredwith a coupled spectrophotometric assay. We studiedthe DNA and temperature dependence of DgeRecA andDmuRecA activity. We also measured the ATPase
activity of both proteins at various concentrations ofATP and dATP to determine their kinetic parameters.We found no detectable ATPase and dATPase activ-
ities of DgeRecA and DmuRecA in the absence of DNAand in the presence of dsDNA only. Both proteins how-ever were able to hydrolyze ATP and dATP in the pre-sence of ssDNA and in the DNA strand exchangeconditions, although the rates of ssDNA-dependentATP or dATP hydrolysis were higher. Furthermore,both proteins hydrolyzed dATP more rapidly than ATPeither in the presence of ssDNA or ssDNA and dsDNAin the same reaction mixture (Tables 3 and 4). Therates of ssDNA-dependent ATP and dATP hydrolysisincreased with temperature for both proteins andpeaked at 42°C and 45°C for DmuRecA and DgeRecA,respectively (Table 5). Kinetic parameters for ATPase
Figure 1 Multiple sequence alignment of RecA proteins from bacteria belonging to the Deinococcus-Thermus group and E. coli RecAprotein. DgeRecA - Deinococcus geothermalis DSM 11302 RecA, DmuRecA - D. murrayi DSM 11303 RecA, DraRecA - D. radiodurans R1 RecA,TthHB27Rec - Thermus thermophilus HB27 RecA, TaqRecA - T. aquaticus YT-1 RecA,TthHB8RecA - T. thermophilus HB8 RecA, MsiRecA - Meiothermussilvanus DSM 9946 RecA, MruRecA - M. ruber DSM 1279 RecA, EcoRecA - E. coli RecA. Three levels of conserved residues are indicated by blue(100%), green (80%) and grey (60%) backgrounds. The alignment was performed using Clustalx 2.0.11 program.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 4 of 13

and dATPase activity of DgeRecA and DmuRecA pro-teins are displayed in Tables 6 and 7.
DNA strand exchange reactionsThe ability of DgeRecA and DmuRecA proteins to pro-mote DNA strand exchange reaction was investigated inthe presence of ATP or dATP, ATP-regenerating systemand DgeSSB protein (obtained according to a procedureby Filipkowski et al. [44]), at 42°C for DmuRecA and 45°C for DgeRecA. At first, filaments composed of ssDNAand RecA protein were formed. Afterwards, linear com-plementary dsDNA was added to start DNA strand
exchange. After incubation and at various time periods,reactions were halted by protein degradation. As shownin Figure 7, DgeRecA and DmuRecA promoted homolo-gous DNA strand exchange either in the presence ofATP or dATP as evidenced by forming nicked circulardsDNA products, although the products appeared earlierwhen reactions were performed with ATP. MoreoverDNA strand exchange occurred faster when DmuRecAwas used. We also performed DNA strand exchangereactions via an inverse pathway, where RecA proteinswere preincubated with the linear dsDNA and circularssDNA was then added to initiate reactions, however wedo not observe nicked circular dsDNA products evenafter 90 min of incubation.
DiscussionIn this article, the cloning, purification and initial char-acterization of RecA proteins from slightly thermophilicand extremely radioresistant bacteria D. geothermalisand D. murrayi are described. The DgeRecA and DmuR-ecA exhibit many properties common to this class ofproteins. In the absence of DNA both proteins self-assemble into a variety of oligomers, their size stronglydepending on the concentration of protein in the solu-tion, a characteristic of RecA proteins. Although oligo-meric populations of T. aquaticus and T. thermophilusRecA proteins contain a large percentage of hexamers[45-48], it was impossible in our study to determine thepreferred oligomeric state of DgeRecA and DmuRecA.RecA proteins of D. geothermalis and D. murrayi bind
both ssDNA and dsDNA like other RecA proteins.
Figure 2 SDS-PAGE analysis of the fractions obtained by expression and purification of DgeRecA and DmuRecA proteins. Lane 1 - LMWSDS Marker (Amersham Biosciences AB): 97, 66, 45, 30, 20.1 and 14.4 kDa, lane 2 - cell extract of the E. coli BLR(DE3) + pET-30Ek/LIC-DgeRecA,lane 3 - DgeRecA after heat treatment, lane 4 - DgeRecA after chromatography on Fractogel EMD DEAE column, lane 5 - DgeRecA afterchromatography on ResourceQ column, lane 6 - DgeRecA after chromatography on MonoQ column, lane 7 - Unstained Protein MolecularWeight Marker (Fermentas): 116, 66.2, 45, 35, 25, 18.4 and 14.4 kDa, lane 8 - cell extract of E. coli BLR(DE3) + pET-30Ek/LIC-DmuRecA, lane 9 -DmuRecA after chromatography on Fractogel EMD DEAE column, lane 10 - DmuRecA after chromatography on ResourceQ column, lane 11 -DmuRecA after chromatography on MonoQ column.
Table 1 Summary of the purification of DgeRecA andDmuRecA proteins obtained from 1 L of E. coli BLR(DE3)culture
Protein Purificationstep
Total protein(mg)
Recovery of protein(%)
DgeRecA Cell extract 760 100
Heat treatment 341 45
Fractogel EMD 121 16
DEAE column
ResourceQcolumn
75 10
MonoQ column 63 8
DmuRecA Cell extract 510 100
Fractogel EMD 356 70
DEAE column
ResourceQcolumn
93 18
MonoQ column 50 10
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 5 of 13

However, it was demonstrated that RecA protein of D.radiodurans binds preferentially to double-strandedDNA even when ssDNA is present in the reaction mix-ture [49,50]. In contrast to DraRecA, DgeRecA andDmuRecA bind more readily to single-stranded DNAwhen both ssDNA and dsDNA are in the same reactionmixture.RecA proteins of D. geothermalis and D. murrayi are
DNA-dependent ATPases. In the absence of DNA, ATPor dATP, hydrolysis was not detected. A similar result wasdemonstrated for D. radiodurans RecA [50]. In contrast,ATP hydrolysis in the absence of exogenous DNA wasdetected for E. coli RecA protein, although the rate wassignificantly reduced [33]. Unlike DraRecA protein[49,50], the DgeRecA and DmuRecA proteins were notable to perform ATP or dATP hydrolysis in the presenceof dsDNA at pH 7.5. The same results were shown for T.thermophilus and E. coli RecA proteins [33,51], althoughEcoRecA protein hydrolyses ATP in the presence ofdsDNA at lower pH (optimum near pH 6) [33]. The ATPand dATP hydrolysis catalyzed by RecA proteins of D.geothermalis and D. murrayi was stimulated by single-stranded DNA, and dATP was hydrolyzed faster thanATP. These results are consistent with the results obtainedfor RecA proteins of E. coli and D. radiodurans [33,49].Both DgeRecA and DmuRecA as well as EcoRecA and
DraRecA were able to promote DNA strand exchangeusing ATP or dATP as a cofactor [41,49]. However, inthe presence of dATP nicked circular heteroduplex
Figure 3 Effects of DgeRecA (A) and DmuRecA (B) proteins concentrations on the elution profiles of gel filtration. In panel A elutionvolume of 9.73 ml corresponding to the 14-meric protein; elution volume of 10.15 ml corresponding to the 12-meric protein; elution volumes of10.60 and 12.52 ml corresponding to 10-meric and tetrameric proteins, respectively; elution volumes of 10.79 and 13.58 ml corresponding to 9-meric and trimeric proteins, respectively; elution volumes of 11.10 and 14.24 ml corresponding to octameric and dimeric proteins, respectively. Inpanel B elution volumes of 8.44, 8.48, 8.38 and 8.26 ml corresponding to protein agregates; elution volumes of 9.94, 13.22 and 14.29 mlcorresponding to 13-meric, trimeric and dimeric proteins, respectively; elution volumes of 10.56, 13.30 and 14.35 ml corresponding to 10-meric,trimeric and dimeric proteins, respectively; elution volume of 11.12 ml corresponding to the octameric protein; elution volume of 13.24 mlcorresponding to the trimeric protein; elution volume of 14.46 ml corresponding to the dimeric protein.
Figure 4 Binding of DgeRecA (A) and DmuRecA (B) to 5’-endfluorescein-labeled (dT)35-oligonucleotides in the presence ofvarious Mg2+ ions concentrations at 37°C - gel mobility shiftassays. Lane 1 - oligo(dT)35, lane 2 - 0 mM Mg2+, lane 3 - 2 mMMg2+, lane 4 - 4 mM Mg2+, lane 5 - 6 mM Mg2+, lane 6 - 8 mMMg2+, lane 7 - 10 mM Mg2+.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 6 of 13
products formed more slowly than in the presence ofATP. Similar results were also obtained for RecA pro-tein of D. radiodurans [49].The major difference between E. coli and D. radiodur-
ans RecA proteins is the ability to promote DNA strandexchange using inverse pathways. The EcoRecA initiatesDNA strand exchange with a filament bound to thessDNA, while the DraRecA binds dsDNA first andhomologous ssDNA second [43]. In our study, we didnot obtained the product of DNA strand exchange reac-tions initiated with DgeRecA-dsDNA or DmuRecA-dsDNA filaments, although Sghaier et al. [52] demon-strated that RecA protein of D. geothermalis is able to
promote DNA exchange reactions through normal andinverse pathways. The difference in results obtained bySghaier et al. and us may be due to differences in theconditions of the DNA strand exchange reactions, espe-cially the differences in the length of dsDNA fragmentsused. In our study, we performed DNA strand exchangereactions between full length linear M13mp18 dsDNA(7249 bp) and circular M13mp18 ssDNA, whereas Sgha-ier et al. used a short fragment of FX174 dsDNAobtained by digestion with HincII endonuclease (thelongest DNA fragment obtained by HincII digestion ofFX174 RFI and its size was 1057 bp) and circularFX174 ssDNA (5386 nt).
Figure 5 Binding of DgeRecA (A, B) and DmuRecA (C, D) to 5’-end fluorescein-labeled (dT)35-oligonucleotides in the presence of 10mM Mg2+ at various temperatures - gel mobility shift assays. Panel A: lane 1 - oligo(dT)35, lane 2 - 25.0°C, lane - 3 - 31.1°C, lane 4 - 38.2°C,lane 5 - 45.3°C, lane 6 - 51.1°C, lane - 7 - 58.0°C, lane 8 - 65.3°C, lane 9 - 72.2°C, lane 10 - 75.0°C; panel B: lane 1 - oligo(dT)35, lane 2 - 55.0°C, lane3 - 55.9°C, lane 4 - 57.1°C, lane 5 - 58.0°C, lane 6 - 59.4°C; panel C: lane 1 - oligo(dT)35, lane 2 - 25.0°C, lane - 3 - 25.9°C, lane 4 - 28.7°C, lane 5 -33.1°C, lane 6 - 37.9°C, lane - 7 - 42.6°C, lane 8 - 47.4°C, lane 9 - 52.1°C, lane 10 - 56.9°C, lane 11 - 61.2°C, lane 12 - 64.1°C, lane 13 - 65.0°C; panelD: lane 1 - oligo(dT)35, lane 2 - 52.0°C, lane 3 - 53.2°C, lane 4 - 54.5°C, lane 5 - 55.6°C.
Figure 6 Binding of DgeRecA (A) and DmuRecA (B) to dsDNA in the absence and in the presence of heterologous ssDNA - gelmobility shift assays. Lane 1 - 600 bp PCR product, lane 2 - dsDNA binding products obtained in the absence of Mg2+ ions, lane 3 - dsDNAbinding products obtained in the presence of 10 mM Mg2+ ions, lane 4 - 600 bp PCR product and 5’-end fluorescein-labeled oligo(dT)35, lane 5 -DNA binding products obtained in the absence of Mg2+ ions, lane 6 - DNA binding products obtained in the presence of 10 mM Mg2+ ions.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 7 of 13

RecA proteins of D. geothermalis and D. murrayi areactive at elevated temperatures similarly to RecA of T.thermophilus. The optimum temperature for TthRecAATPase activity is 65°C [51], and for DgeRecA andDmuRecA proteins is 45 and 42°C, respectively. How-ever, they are able to bind ssDNA at temperaturesreaching 54.5°C for DmuRecA and 58°C for DgeRecA.Moreover, the thermostability assay revealed that theDgeRecA and DmuRecA are less thermostable thanTthRecA which remains still active after a 10-s incuba-tion at 85°C (data not shown), whereas DgeRecA andDmuRecA are completely inactivated in 10 s at 80°C.
ConclusionsThe properties of RecA proteins of slightly thermophilicand extremely radioresistant bacteria D. geothermalis andD. murrayi obtained in this study demonstrate that theyare close functional homologues of the other RecA pro-teins. Although it has previously been shown that theRecA protein of the extremely radioresistant bacteriumD. radiodurans exhibits some unusual properties, in ourstudy we did not observe such properties in the case ofDgeRecA and DmuRecA. This suggests the uniqueness ofDraRecA even among RecA proteins from microorgan-isms of the Deinococcus genus. On the other hand, it hasbeen also demonstrated that the dsDNA-binding prefer-ences displayed by DraRecA are not necessary for dou-ble-stranded breaks repair in g-irradiated D. radiodurans.The rapid reconstruction of an intact genome occurringthrough an extended synthesis-dependent strand
annealing process (ESDSA) followed by a DNA recombi-nation requires a 5’-3’ single-stranded DNA exonucleaseRecJ, a RecQ helicase and a RecF, RecO and RecR pro-teins to act together to promote loading of RecA ontosingle-stranded DNA [53].
MethodsBacterial strains and growth conditionsD. geothermalis DSM 11302 and D. murrayi DSM 11303were purchased from Deutsche Sammlung von Mikroor-ganismen und Zellkulturen (Braunschweig, Germany).They were grown in 50 ml modified Luria-Bertani med-ium pH 7.2 containing 1 g peptone K, 1 g yeast extractand 7.4 g sea salt in 1000 ml distilled water. D. geother-malis was grown for 30 h at 50°C and D. murrayi wasgrown for 30 h at 47°C. Afterwards genomic DNA wasisolated using Genomic Mini AX Bacteria (A&A Bio-technology, Poland).
Cloning of the D. geothermalis recA geneThe recA gene was amplified by PCR using genomicDNA from D. geothermalis DSM 11302 as the template.The primers used were: DGRAFNde 5’ CGACATAT-GAGCAAGGAACAACCCCAAGGA 3’, containingrecognition site for NdeI endonuclease (underlined) andDGRARHnd 5’ ACAAAGCTTACTCTGC-CAAGGCGGGC 3’, containing recognition site for Hin-dIII endonuclease (underlined). The start and stopcodons are bolded. The reaction mixture consisted of0.13 μg of D. geothermalis DNA, 0.2 μM of each primer,200 μM of each dNTP, 2 mM MgSO4 and 1 U ofDelta3 DNA polymerase (DNA-Gdańsk, Poland) in 1 ×PCR buffer (20 mM Tris-HCl pH 8.8, 10 mM KCl, 10mM (NH4)2SO4, 0.1% Triton X-100). PCR reaction wasperformed using the following conditions: 95°C - 2 min,(95°C - 1 min, 61°C - 1 min, 72°C - 1 min; 30 cycles),72°C - 5 min. The PCR product was cloned into thepCR-Blunt vector (Invitrogen, Carlsbad, California,USA) and sequenced. The obtained pCR-Blunt-NdeI-DgeRecA-HindIII plasmid was then digested with NdeIand HindIII endonucleases, and the DNA fragment con-taining the recA gene was cloned into pET-30 Ek/LIC
Table 2 Thermostability of DgeRecA and DmuRecAproteins
Temperature (°C) Maximum incubation time (min)
DgeRecA DmuRecA
50 180 180
57 180 15
62 20 8
71 1 0.67
75 0.50 0.33
80 0a 0a
a Complete lost of activity after shorter than 10 s incubation time
Table 3 Effect of DNA on the rate of ATP and dATPhydrolysis catalyzed by DgeRecA.
Substrate Rate of hydrolysis (μM/min)
absence ofDNA
dsDNA ssDNA ssDNA anddsDNAa
ATP not detected notdetected
26.8 ±0.40
20.3 ± 0.36
dATP not detected notdetected
31.6 ±0.63
23.7 ± 0.43
Reactions conditions: 37°C, 2.27 mM ATP or dATP.a DNA strand exchange conditions
Table 4 Effect of DNA on the rate of ATP and dATPhydrolysis catalyzed by DmuRecA
Substrate Rate of hydrolysis (μM/min)
absence ofDNA
dsDNA ssDNA ssDNA anddsDNAa
ATP not detected notdetected
34.4 ±0.58
21.2 ± 0.44
dATP not detected notdetected
47.0 ±0.71
24.6 ± 0.49
Reactions conditions: 37°C, 2.27 mM ATP or dATP.a DNA strand exchange conditions
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 8 of 13

expression vector (Novagen, Beeston, Nottingham, Eng-land) digested with the same restriction enzymes. Theresulting recombinant plasmid pET-30Ek/LIC-DgeRecAwas used for the production of D. geothermalis RecAprotein in E. coli.
Isolation and cloning of the D. murrayi recA geneIn order to obtain an internal part of the recA genefrom D. murrayi DSM 11303, sequences encoding RecAproteins of Deinococcus geothermalis DSM 1300, D.geothemalis DSM 11302, Deinococcus radiodurans R1,Thermus aquaticus YT-1, Thermus thermophilus HB8and T. thermophilus HB27 obtained from the GenBankdatabase were aligned using the ClustalX program, ver-sion 1.8. Based on the alignment, degenerated primersInvRecA1 5’ GAGTCSGGSGGCAAGACCAC 3’ andInvRecA2 5’ TCCTTSCCCTGGCCSAKGC 3’ weredesigned and synthesized. The PCR reaction was per-formed in the mixture containing: 0.2 μM of each pri-mer, 0.2 μg of D. murrayi DSM 11303 genomic DNA,200 μM of each dNTP, 3 mM MgCl2 and 1 U of DNApolymerase Hypernova (DNA-Gdańsk, Poland) in 1 ×buffer Hypernova (10 mM Tris-HCl pH 8.8, 50 mMKCl, 0.15% Triton X-100). The reaction mixture wasincubated for 2 min at 95°C, followed by 30 cycles at95°C for 1 min, 59°C for 1 min, 72°C for 1 min, and afinal incubation for 5 min at 72°C. The obtained PCRproduct was then purified from an agarose gel bandusing Gel-Out kit (A&A Biotechnology, Poland), clonedinto pJET1.2/blunt vector CloneJET™ PCR Cloning Kit(Fermentas, Vilnius, Lithuania) and sequenced. After-wards, the inverse PCR was performed for obtaining offlanking regions. The D. murrayi genomic DNA wasfirst digested with DraI restriction endonuclease andthen restriction fragments were ligated upon itself to
form circles. In the second step, the PCR amplificationwas performed using ligation products as a templateand two primers: recAInvFor 5’ CCGCAAGATTGGG-CAGCCCGTCAAGA 3’ and recAInvRev 5’ACCCGACCGCACGAGCAGCTCCATG 3’, designedon the basis of previously obtained partial sequence ofD. murrayi recA gene. The reaction mixture containedalso 200 μM of each dNTP, 3 mM MgCl2 and 1 U ofDNA polymerase Hypernova (DNA-Gdańsk, Poland) in1 × buffer Hypernova. DNA amplification was per-formed using the following conditions: 95°C - 3 min,(95°C - 1.5 min, 66°C - 1 min, 72°C - 5 min) 30 cyclesand 72°C - 15 min after the final cycle. The PCR pro-duct was purified from an agarose gel band, cloned intopJET1.2/blunt vector and sequenced. Afterwards, frag-ments of D. murrayi genomic DNA sequence werealignment and the full sequence of recA gene wasobtained.The D. murrayi DSM 11303 recA gene was then
amplified using the forward primer FDMRecANdeI 5’ATTACATATGAGCAAGGACAACCCCAAGGACTTC3’, and the reverse primer RDMRecAXhoI 5’TATTCTCGAGTTACTCCGCGACAGCGGGCAC 3’,containing NdeI and XhoI recognition sites, respectively(underlined). The start and stop codons are bolded. ThePCR reaction mixture contained: 0.2 μM of each primer,0.2 μg of D. murrayi genomic DNA, 200 μM of eachdNTP, 3 mM MgCl2 and 1 U of DNA polymeraseHypernova (DNA-Gdańsk, Poland) in 1 × Hypernovabuffer. The reaction mixture was incubated for 3 min at96°C, followed by 5 cycles at 95°C for 1 min, 58°C for 1min, 72°C for 1 min and 25 cycles at 95°C for 1 min,63°C for 1 min, 72°C for 1 min, and a final incubationfor 5 min at 72°C. Afterwards, PCR product was purifiedfrom an agarose gel band, cloned into pJET1.2/blunt
Table 5 Effect of temperature on the rate of ssDNA-dependent ATP and dATP hydrolysis catalyzed by DgeRecA andDmuRecA
Protein Substrate Rate of hydrolysis (μM/min)
32°C 37°C 42°C 45°C 50°C
DgeRecA ATP 19.8 ± 0.26 26.8 ± 0.40 31.2 ± 0.56 34.1 ± 0.78 26.6 ± 0.43
dATP 10.3 ± 0.18 31.6 ± 0.63 53.7 ± 1.18 65.7 ± 1.71 56.5 ± 1.30
DmuRecA ATP 15.6 ± 0.23 34.4 ± 0.58 55.2 ± 1.16 47.6 ± 1.20 34.5 ± 0.62
dATP 24.2 ± 0.34 47.0 ± 0.71 70.3 ± 1.76 59.9 ± 1.38 43.9 ± 0.88
Reaction conditions: 2.27 mM ATP or dATP.
Table 6 Kinetic parameters for ATP and dATP hydrolysis catalyzed by DgeRecA
Substrate DNA T (°C) KM (mM) kcat (s-1) kcat/KM (s-1 M-1) na
ATP ssDNA 37 0.56 ± 0.018 0.15 ± 0.004 2.7 ± 0.12 × 102 2.9
ssDNA and dsDNA 37 0.13 ± 0.006 0.13 ± 0.006 1.0 ± 0.04 × 103 1.4
dATP ssDNA 37 0.62 ± 0.023 0.21 ± 0.007 3.3 ± 0.10 × 102 1.2
ssDNA and dsDNA 37 0.36 ± 0.016 0.15 ± 0.007 4.2 ± 0.16 × 102 1.0a Hill coefficient.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 9 of 13

vector and sequenced. The obtained pJET-NdeI-DmuR-ecA-XhoI plasmid was then digested with NdeI andXhoI endonucleases, and the DNA fragment containingthe recA gene was cloned into pET-30 Ek/LIC expres-sion vector (Novagen) digested with the same restrictionenzymes. The resulting recombinant plasmid pET-30Ek/LIC-DmuRecA was used for the production of D. mur-rayi RecA protein in E. coli.
Production and purification of recombinant DgeRecA andDmuRecA proteinsOverproduction of DgeRecA and DmuRecA proteins wasperformed in the E. coli BLR(DE3) cells (Novagen) car-rying the pET-30Ek/LIC-DgeRecA or pET-30Ek/LICD-muRecA plasmids. Cells were grown at 37°C in 750 mlLB medium (1% peptone K, 0.5% yeast extract, 1%
NaCl) containing 20 μg/ml of kanamycin. At OD600 0.5IPTG was added to the final concentration of 1 mMand cultivation was continued for 4 hours. Subsequently,cultures were centrifuged (4612 × g, 10 min, 4°C) andpellets were resuspended in 75 ml of buffer A1 (20 mMpotassium phosphate buffer pH 6.0, 50 mM KCl, 1 mMEDTA, 1 mM DTT) in the case of DgeRecA proteinproduction or buffer A2 (20 mM potassium phosphatebuffer pH 6.5, 50 mM KCl, 1 mM EDTA, 1 mM DTT )for the DmuRecA. Samples were sonicated seven timesfor 30 s at 0°C and centrifuged (4612 × g, 10 min, 4°C).The supernatant containing DgeRecA protein was heat-treated at 60°C for 20 min, cooled on ice and centri-fuged again (18000 × g, 30 min, 4°C).DgeRecA and DmuRecA proteins were then purified
using Fractogel EMD DEAE column (Merck, Darmstadt,
Table 7 Kinetic parameters for ATP and dATP hydrolysis catalyzed by DmuRecA
Substrate DNA T (°C) KM (mM) kcat (s-1) kcat/KM (s-1 M-1) na
ATP ssDNA 37 0.31 ± 0.009 0.13 ± 0.004 4.3 ± 0.18 × 102 1.1
ssDNA and dsDNA 37 0.28 ± 0.009 0.067 ± 0.0021 2.4 ± 0.08 × 102 1.5
dATP ssDNA 37 0.28 ± 0.007 0.14 ± 0.004 5.0 ± 0.25 × 102 1.0
ssDNA and dsDNA 37 0.22 ± 0.007 0.073 ± 0.0023 3.4 ± 0.05 × 102 1.3a Hill coefficient
Figure 7 DNA strand exchange promoted by DgeRecA (A, C) and DmuRecA (B, D) proteins. Panels A and B: lane 1 - M13mp18 ssDNA,lane 2 - M13mp18 dsDNA digested with SmaI, lane 3 - reaction products formed after 15 min, lane 4 - reaction products formed after 30 min,lane 5 - reaction products formed after 45 min, lane 6 - reaction products formed after 60 min, lane 7 - reaction products formed after 75 min,lane 8 - reaction products formed after 90 min. P - nicked circular dsDNA. Panels C and D: plots of the quantified product formation for thereactions in panels A and B.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 10 of 13

Germany) equilibrated with buffer A1 or A2, respec-tively. RecA proteins were eluted with linear gradient of0.05-1.5 M KCl in the appropriate buffer. Fractions con-taining D. geothermalis or D. murrayi RecA werepooled, dialyzed against buffer A1 or A2 and loadedonto ResourceQ column (Amersham Biosciences AB,Uppsala, Sweden). Elution was performed by linear gra-dient of 0.05-0.425 M KCl in buffer A1 or A2. Fractionscontaining RecA proteins were dialyzed against bufferA1 or A2 once again and purified using MonoQ column(Amersham Biosciences AB). DgeRecA and DmuRecAwere eluted with linear gradient of 0.05-0.35 M KCl inappropriate buffer. Collected fractions containing puri-fied RecA proteins were then checked for nucleases con-taminations by incubating them with M13mp18 RFIDNA (24 μM of nucleotides), linear dsDNA M13mp18(24 μM of nucleotides) and circular ssDNA M13mp18(12 μM of nucleotides) in the 20 mM potassium phos-phate buffer pH 7.5 containing 10 mM MgCl2. The con-centration of RecA proteins was 10 μM. Reactionmixtures were incubated at 37°C for 2 h and thenEDTA, SDS and Proteinase K were added to the finalconcentrations of 10 mM, 1% and 2 mg/ml, respectively.Samples were incubated at room temperature for 30min and separated by electrophoresis in 1% agarose gelswith ethidium bromide. DNAs were also incubated withProteinase K only, as a control sample. We did notobserve any exo- or endonuclease activities neither inRecA proteins preparations nor in Proteinase K. After-wards the RecA proteins were dialyzed against 100 mM(NH4)HCO3 and lyophilized for long-time storage.The purity of proteins was investigated by SDS-PAGE.
The concentration of RecA proteins at every step of pro-duction and purification was measured by Bradford method[54]. Protein concentration in the samples for biochemicaltests was determined by A280 nm using an extinction coeffi-cient of 11,920 M-1 cm-1, which was calculated from theamino acid sequences of DgeRecA and DmuRecA proteins.
Estimation of the native molecular massThe molecular mass of DgeRecA and DmuRecA proteinswere estimated using analytical gel filtration chromatogra-phy. Samples containing various concentrations of purifiedproteins were loaded onto Superdex 200 HR 10/30 col-umn (Amersham Biosciences AB) equilibrated with 25mM potassium phosphate buffer pH 7.5 containing 1.0 MKCl and eluted with the same buffer at a flow rate of 0.5ml/min. The elution profiles were monitored by recordingthe absorbance at 214 nm. The molecular weights ofDgeRecA and DmuRecA oligomers were determined bycomparison with those of standard proteins: thyroglobulin(669 kDa), apoferritin (443 kDa), b-amylase (200 kDa),alcohol dehydrogenase (150 kDa), bovine serum albumin(66 kDa) and carbon anhydrase (29 kDa).
ssDNA binding activityThe ability of DgeRecA and DmuRecA proteins to bindssDNA was investigated by electrophoretic mobility shiftof fluorescein 5’-end-labelled oligo(dT)35 nucleotides.25 μl reaction mixtures containing 5.6 μM (nucleotides)oligo(dT)35, 110 μM ATP-g-S, 300 μM ATP, from 0 to10 mM MgCl2 and 9 μM RecA protein in 20 mM potas-sium phosphate buffer pH 7.5 were incubated at 25-75°C for 30 min in the dark. Afterwards samples wereseparated by electrophoresis in 0.8% agarose gels.Results were visualized using UV transilluminator.
dsDNA binding activityThe ability of DgeRecA and DmuRecA proteins to binddsDNA was investigated by electrophoretic mobilityshift of 600 bp PCR product (l DNA amplified using 5’GGACAAGGTCGCTGAAGCCTTCG 3’ and 5’TCCTGACGGGCGGTATATTTCTCC 3’ primers). 25μl reaction mixtures containing 30 μM (nucleotides)dsDNA or 30 μM dsDNA and 14 μM ssDNA (fluores-cein 5’-end-labelled oligo(dT)35), 180 μM ATP-g-S, 500μM ATP, 0 or 10 mM MgCl2 and 12 μM RecA proteinin 20 mM potassium phosphate buffer pH 7.5 wereincubated at 37°C for 30 min. Afterwards samples wereseparated by electrophoresis in 0.8% agarose gels con-taining ethidium bromide. Results were visualized usingUV transilluminator.
Thermostability of the RecA proteinsThe thermostability of DgeRecA and DmuRecA wasdetermined by ssDNA binding activity after incubationof proteins at various temperatures for a set of time per-iods. The samples containing 11.8 μM RecA protein and0 or 10 mM MgCl2 in 20 mM potassium phosphatebuffer pH 7.5 were incubated at 50-80°C for 10 s to180 min, cooled on ice and centrifuged (13500 × g, 10min). Aliquots (19 μl) were then taken and fluorescein5’-end-labelled oligo(dT)35 nucleotides, ATP, ATP-g-Sand MgCl2 were added to the concentrations of 5.6 μM,300 μM, 110 μM and 10 mM in the final volume of25 μl, respectively. Subsequently reaction mixtures wereincubated at 37°C for 30 min in the dark and separatedby electrophoresis in 0.8% agarose gels. Results werevisualized using UV transilluminator.
ATPase assayHydrolysis of ATP (or dATP) by DgeRecA and DmuR-ecA was monitored by an enzyme coupled method.Regeneration of ATP from ADP and phosphoenolpyru-vate catalyzed by pyruvate kinase was coupled with theconversion of NADH to NAD+ catalyzed by lactatedehydrogenase, which was monitored by a decrease inabsorbance at 355 nm. The absorbance was measuredusing PerkinElmer multilabel plate reader Victor3 V
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 11 of 13

(Centre of Excellence ChemBioFarm, Gdańsk Universityof Technology, Gdańsk, Poland).The reaction mixtures consisted of: 3.03 μM RecA pro-
tein, 11.56 mM MgCl2, 1.25 U of lactate dehydrogenasefrom rabbit muscle, 0.75 U of pyruvate kinase from rabbitmuscle, 3.3 mM NADH, 2.27 mM PEP, 0.38-2.27 mMATP (or dATP) and 20 mM Tris-HCl buffer pH 7.5 to theend volume of 75 μl. Reactions were carried out in theabsence of DNA and in the presence of 12.16 μM (nucleo-tides) ssDNA of SygB1a oligonucleotide 5’CGTTCGAAACGATGAGGAAGACTTATTGGCTGATGGCGCTTTTCGCGGTGCTCATGTTCCCTGAAAAGTTCCTTTGGGGT 3’ (80 nt) and/or 1,312 μM (nucleo-tides) of XbaI-linearized dsDNA of pBADTOPObAQaplasmid (5727 bp) containing complementary sequence tothe SygB1a oligonucleotide. ATP (or dATP) hydrolysisreactions were performed at 32, 37, 42, 45 and 50°C.
DNA strand exchange reactionsRecA-dependent DNA strand exchange reactions werecarried out between circular ssDNA of M13mp18 andlinear dsDNA of M13mp18 obtained by SmaI digestion.All reactions were carried out in solutions containing 20mM Tris-HCl buffer pH 7.5, 5 μM RecA protein, 9 μM(nucleotides) ssDNA, 18 μM (nucleotides) dsDNA, 0.3μM DgeSSB protein [44], 1.2 mM DTT, 10 mM MgCl2,3 mM ATP and an ATP-regenerating system (0.5 U pyr-uvate kinase from rabbit muscle, 3 mM phosphoenol-pyruvate) in 25 μl. A preincubation of ssDNA withDgeRecA at 45°C or with DmuRecA at 42°C for 5 minwas followed by addition of ATP or dATP and SSB.After an additional 5 min incubation, linear dsDNA wasadded to start the strand exchange reactions. The reac-tions were carried out at 45°C when DgeRecA was usedor at 42°C in the case of DmuRecA for 15, 30, 45, 60,75 and 90 min and stopped by addition of EDTA, SDSand Proteinase K to the final concentrations of 10 mM,1% and 2 mg/ml, respectively. Samples were incubatedat room temperature for 30-90 min and then separatedby electrophoresis in 0.8% agarose gels. Results werevisualized and photographed with UV light using Versa-Doc™ Imaging System (Bio-Rad Laboratories, Hercules,California, USA) after staining of gels with ethidiumbromide. The DNA bands were quantified with QuantityOne software, version 4.3.1 (Bio-Rad Laboratories). Theband corresponding to nicked circular dsDNA productwas quantified as the fraction of the total DNA in agiven gel lane, excluding the band corresponding to thessDNA. In the case of inverse DNA strand exchangereactions, RecA proteins were preincubated with the lin-ear dsDNA and ATP or dATP for 15-60 min. ThessDNA was then added to start the reactions, and theSSB was added 5 min later.
AbbreviationsDSB: DNA double-strand break; DgeRecA: Deinococcus geothermalis RecAprotein; DmuRecA: Deinococcus murrayi RecA protein; DraRecA: Deinococcusradiodurans RecA protein; TthRecA: Thermus thermophilus RecA protein;EcoRecA: Escherichia coli RecA protein; DgeSSB: Deinococcus geothermalissingle-stranded DNA-binding protein; ATP-γ-S: Adenosine 5’-[γ-thio]triphosphate; IPTG: Isopropyl β-D-thiogalactopyranoside.
AcknowledgementsThis work was supported by the Polish Ministry of Science and HigherEducation Grant N N302 067934 to J.K. We thank Dr. P. Filipkowski for pBAD-DgeSSB plasmid used in this study.
Authors’ contributionsMW participated in the design of the study, partially characterized of RecAproteins and drafted the manuscript; PH obtained and partially characterizedof RecA proteins; BK partially characterized of RecA proteins; JK correctedand edited the final version of manuscript. All authors read and approvedthe final manuscript.
Received: 24 January 2011 Accepted: 22 April 2011Published: 22 April 2011
References1. Ferreira AC, Nobre MF, Rainey FA, Silva MT, Wait R, Burghardt J, Chung AP,
da Costa MS: Deinococcus geothermalis sp. nov. and Deinococcus murrayisp. nov., two extremely radiation-resistant and slightly thermophilicspecies from hot springs. Int J Syst Bacteriol 1997, 47:939-947.
2. Väisänen OM, Weber A, Bennasar A, Rainey FA, Busse HJ, Salkinoja-Salonen MS: Microbial communities of printing paper machines. J ApplMicrobiol 1998, 84:1069-1084.
3. Kolari M, Nuutinen J, Salkinoja-Salonen MS: Mechanisms of biofilmformation in paper machine by Bacillus species: the role of Deinococcusgeothermalis. J Ind Microbiol Biotechnol 2001, 27:343-351.
4. Peltola M, Kanto Öqvist C, Ekman J, Kosonen M, Jokela S, Kolari M,Korhonen P, Salkinoja-Salonen M: Quantitative contributions of bacteriaand of Deinococcus geothermalis to deposits and slimes in paperindustry. J Ind Microbiol Biotechnol 2008, 35:1651-1657.
5. Zahradka K, Slade D, Bailone A, Sommer S, Averbeck D, Petranovic M,Lindner AB, Radman M: Reassembly of shattered chromosomes inDeinococcus radiodurans. Nature 2006, 443:569-573.
6. Slade D, Lindner AB, Paul G, Radman M: Recombination and replication inDNA repair of heavily irradiated Deinococcus radiodurans. Cell 2009,136:1044-1055.
7. Repar J, Cvjetan S, Slade D, Radman M, Zahradka D, Zahradka K: RecAprotein assures fidelity of DNA repair and genome stability inDeinococcus radiodurans. DNA Repair 2010, 9:1151-1161.
8. Shigenobu S, Watanabe H, Masahira Hattori M, Sakaki Y, Ishikawa H:Genome sequence of the endocellular bacterial symbiont of aphidsBuchnera sp. APS. Nature 2000, 407:81-86.
9. Gil R, Silva FJ, Zientz E, Delmotte F, González-Candelas F, Latorre A,Rausell C, Kamerbeek J, Gadau J, Hölldobler B, van Ham RCHJ, Gross R,Moya A: The genome sequence of Blochmannia floridanus: comparativeanalysis of reduced genomes. Proc Natl Acad Sci USA 2003, 100:9388-9393.
10. Akman L, Yamashita A, Watanabe H, Oshima K, Shiba T, Hattori M, Aksoy S:Genome sequence of the endocellular obligate symbiont of tsetse flies,Wigglesworthia glossinidia. Nat Genet 2002, 32:402-407.
11. Kuwahara H, Yoshida T, Takaki Y, Shimamura S, Shinro Nishi S, MaikoHarada M, Kazuyo Matsuyama K, Takishita K, Kawato M, Uematsu K,Fujiwara Y, Sato T, Kato C, Kitagawa M, Kato I, Maruyama T: Reducedgenome of the thioautotrophic intracellular symbiont in a deep-seaclam, Calyptogena okutanii. Curr Biol 2007, 17:881-886.
12. Newton ILG, Woyke T, Auchtung TA, Dilly GF, Dutton RJ, Fisher MC,Fontanez KM, Lau E, Stewart FJ, Richardson PM, Barry KW, Saunders E,Detter JC, Wu D, Eisen JA, Cavanaugh CM: The Calyptogena magnificachemoautotrophic symbiont genome. Science 2007, 315:998-1000.
13. Kuwahara H, Takaki Y, Yoshida T, Shimamura S, Takishita K, Reimer JD,Kato C, Maruyama T: Reductive genome evolution in chemoautotrophicintracellular symbionts of deep-sea Calyptogena clams. Extremophiles2008, 12:365-374.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 12 of 13

14. Seitz EM, Brockman JP, Sandler SJ, Clark AJ, Kowalczykowski SC: RadAprotein is an archaeal RecA protein homolog that catalyzes DNA strandexchange. Genes Dev 1998, 12:1248-1253.
15. Ogawa T, Yu X, Shinohara A, Egelman EH: Similarity of the yeast RAD51filament to the bacterial RecA filament. Science 1993, 259:1896-1899.
16. Benson FE, Stasiak A, West SC: Purification and characterization of thehuman Rad51 protein, an analogue of E. coli RecA. Embo J 1994,13:5764-5771.
17. Fujisawa H, Yonesaki T, Minagawa T: Sequence of the T4 recombinationgene, uvsX, and its comparison with that of the recA gene of Escherichiacoli. Nucleic Acids Res 1985, 13:7473-7481.
18. Kowalczykowski SC, Dixon DA, Eggleston AK, Lauder SD, Rehrauer WM:Biochemistry of homologous recombination in Escherichia coli. MicrobiolRev 1994, 58:401-465.
19. Kuzminov A: Recombinational repair of DNA damage in Escherichia coliand bacteriophage λ. Microbiol Mol Biol Rev 1999, 63:751-813.
20. Lusetti SL, Cox MM: The bacterial RecA protein and the recombinationalDNA repair of stalled replication forks. Annu Rev Biochem 2002, 71:71-100.
21. Little JW: Mechanism of specific LexA cleavage - autodigestion and therole of RecA coprotease. Biochimie 1991, 73:411-421.
22. Shinagawa H: SOS response as an adaptative response to DNA damagein prokaryotes. EXS 1996, 77:221-235.
23. Burckhardt SE, Woodgate R, Scheuremann RH, Echols H: UmuDmutagenesis protein of Escherichia coli: overproduction, purification, andcleavage by RecA. Proc Natl Acad Sci USA 1988, 85:1811-1815.
24. Nohmi T, Battista JR, Dodson LA, Walker GC: RecA-mediated cleavageactivates UmuD for mutagenesis: mechanistic relationship betweentranscriptional derepression and posttranslational activation. Proc NatlAcad Sci USA 1988, 85:1816-1820.
25. Shinagawa H, Iwasaki T, Kato T, Nakata A: RecA-protein dependentcleavage of UmuD protein and SOS mutagenesis. Proc Natl Acad Sci USA1988, 85:1806-1810.
26. Bruck I, Woodgate R, McEntee K, Goodman MF: Purification of a solubleUmuD’C complex from Escherichia coli: cooperative binding of UmuD’Cto single-stranded DNA. J Biol Chem 1996, 271:10767-10774.
27. Tang M, Shen X, Frank EG, O’Donnell M, Woodgate R, Goodman MF:UmuD’2C is an error-prone DNA polymerase, Escherichia coli pol V. ProcNatl Acad Sci USA 1999, 96:8919-8924.
28. Jiang Q, Karata K, Woodgate R, Cox MM, Goodman MF: The active form ofDNA polymerase V is UmuD’2C-RecA-ATP. Nature 2009, 460:359-363.
29. Egelman EH, Stasiak A, Structure of helical RecA-DNA complexes:Complexes formed in the presence of ATP-gamma-S or ATP. J Mol Biol1986, 191:677-697.
30. Di Capua E, Schnarr M, Ruigrok RWH, Lindner P, Timmins PA: Complexes ofRecA protein in solution. A study by small angle neutron scattering.J Mol Biol 1990, 214:557-570.
31. Nordén B, Elvingson C, Kubista M, Sjöberg B, Ryberg H, Ryberg M,Mortensen K, Takahashi M: Structure of RecA-DNA complexes studied bycombination of linear dichroism and small-angle neutron scatteringmeasurements on flow-oriented samples. J Mol Biol 1992, 226:1175-1191.
32. Yu X, Egelman EH: Structural data suggest that the active and inactiveforms of the RecA filament are not simply interconvertible. J Mol Biol1992, 227:334-346.
33. Weinstock GM, McEntee K, Lehman IR: Hydrolysis of nucleosidetriphosphates catalyzed by the recA protein of Escherichia coli.Characterization of ATP hydrolysis. J Biol Chem 1981, 256:8829-8834.
34. Arenson TA, Tsodikov OV, Cox MM: Quantitative analysis of the kinetics ofend-dependent disassembly of RecA filaments from ssDNA. J Mol Biol1999, 288:391-401.
35. Honigberg SM, Gonda DK, Flory J, Radding CM: The pairing activity ofstable nucleoprotein filaments made from recA protein, single-strandedDNA, and adenosine 5’-(gamma-thio)triphosphate. J Biol Chem 1985,260:11845-11851.
36. Menetski JP, Bear DG, Kowalczykowski SC: Stable DNA heteroduplexformation catalyzed by the Escherichia coli RecA protein in the absenceof ATP hydrolysis. Proc Natl Acad Sci USA 1990, 87:21-25.
37. Cox MM, Lehman IR: RecA protein-promoted DNA strand exchange.Stable complexes of recA protein and single-stranded DNA formed inthe presence of ATP and single-stranded DNA binding protein. J BiolChem 1982, 257:8523-8532.
38. Chow SA, Chiu SK, Wong BC: RecA protein-promoted homologous pairingand strand exchange between intact and partially single-strandedduplex DNA. J Mol Biol 1992, 223:79-93.
39. Jain SK, Cox MM, Inman RB: On the role of ATP hydrolysis in RecAprotein-mediated DNA strand exchange. III. Unidirectional branchmigration and extensive hybrid DNA formation. J Biol Chem 1994,269:20653-20661.
40. Kim JI, Cox MM, Inman RB: On the role of ATP hydrolysis in RecA protein-mediated DNA strand exchange. I. Bypassing a short heterologous insertin one DNA substrate. J Biol Chem 1992, 267:16438-16443.
41. Shan Q, Cox MM, Inman RB: DNA strand exchange promoted by RecAK72R. Two reaction phases with different Mg2+ requirements. J BiolChem 1996, 271:5712-5724.
42. Kim JI, Cox MM, Inman RB: On the role of ATP hydrolysis in RecA protein-mediated DNA strand exchange. II. Four-strand exchanges. J Biol Chem1992, 67:16444-16449.
43. Kim JI, Cox MM: The RecA protein of Deinococcus radiodurans andEscherichia coli promote DNA strand exchange via inverse pathways.Proc Natl Acad Sci USA 2002, 99:7917-7921.
44. Filipkowski P, Duraj-Thatte A, Kur J: Novel thermostable single-strandedDNA-binding protein (SSB) from Deinococcus geothermalis. Arch Microbiol2006, 186:129-137.
45. Brenner SL, Zlotnick A, Griffith JD: RecA protein self-assembly. Multiplediscrete aggregation states. J Mol Biol 1988, 204:959-972.
46. Brenner SL, Zlotnick A, Stafford WF III: RecA protein self-assembly. II.Analytical equilibrium ultracentrifugation studies of the entropy-drivenself-association of RecA. J Mol Biol 1990, 216:949-964.
47. Yu X, Angov E, Camerini-Otero RD, Egelman EH: Structural polymorphismof the RecA protein from the thermophilic bacterium Thermus aquaticus.Biophys J 1995, 69:2728-2738.
48. Masui R, Mikawa T, Kato R, Kuramitsu S: Characterization of the oligomericstates of RecA protein: monomeric RecA protein can form anucleoprotein filament. Biochemistry 1998, 37:14788-14797.
49. Kim JI, Sharma AK, Abbott SN, Wood EA, Dwyer DW, Jambura A,Minton KW, Inman RB, Daly MJ, Cox MM: RecA protein from the extremelyradioresistant bacterium Deinococcus radiodurans: expression,purification, and characterization. J Bacteriol 2002, 184:1649-1660.
50. Kim JI: Analysis of double stranded DNA-dependent activities ofDeinococcus radiodurans RecA protein. J Microbiol 2006, 44:508-514.
51. Kato R, Kuramitsu S: Characterization of thermostable RecA protein andanalysis of its interaction with single-stranded DNA. Eur J Biochem 1999,259:592-601.
52. Sghaier H, Satoh K, Ohba H, Narumi I: Assessing the role of RecA proteinin the radioresistant bacterium Deinococcus geothermalis. Afr J Bichem Res2010, 4:111-118.
53. Bentchikou E, Servant P, Coste G, Sommer S: A major role of the RecFORpathway in DNA double-strand-break repair through ESDSA inDeinococcus radiodurans. PLoS Genetics 2010, 6:e1000774.
54. Bradford MM: A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 1976, 72:248-254.
doi:10.1186/1471-2199-12-17Cite this article as: Wanarska et al.: RecA Proteins from Deinococcusgeothermalis and Deinococcus murrayi - Cloning, Purification andBiochemical Characterisation. BMC Molecular Biology 2011 12:17.
Wanarska et al. BMC Molecular Biology 2011, 12:17http://www.biomedcentral.com/1471-2199/12/17
Page 13 of 13




















