
Rapid differentiation of species of Botryosphaeriaceae by PCR fingerprinting
10
Rapid differentiation of species of Botryosphaeriaceae by PCR fingerprinting Artur Alves a, * , Alan J.L. Phillips b , Isabel Henriques a , Anto ´nio Correia a a CESAM, Centro de Estudos do Ambiente e do Mar, Campus Universita ´ rio de Santiago, Departamento de Biologia, Universidade de Aveiro, 3810-193 Aveiro, Portugal b Centro de Recursos Microbiolo ´gicos, Faculdade de Cie ˆncias e Tecnologia, Universidade Nova de Lisboa, Quinta da Torre, 2829-516 Caparica, Portugal Received 2 June 2006; accepted 9 October 2006 Available online 20 December 2006 Abstract The fingerprinting methods referred to as MSP-PCR (microsatellite-primed polymerase chain reaction) and rep-PCR (repetitive-sequence- based polymerase chain reaction) were used to discriminate between species of Botryosphaeriaceae (Fungi, Ascomycota). Several primers were tested with both methods and each primer enabled clear differentiation of all the species tested. Cluster analysis of banding patterns of the isolates corresponded well with known species delineations based on morphology and phylogenetic analysis. The methods described in this paper provide simple and rapid procedures that can be used for routine differentiation of Botryosphaeriaceae isolates at the species level. The two methods are also useful for studying intraspecific variability. Ó 2006 Elsevier Masson SAS. All rights reserved. Keywords: Botryosphaeriaceae; Species differentiation; Fingerprinting; MSP-PCR; Rep-PCR 1. Introduction Species of Botryosphaeria sensu lato are well known path- ogens, saprophytes and endophytes in a wide range of woody angiosperm and gymnosperm hosts [4,9]. The classification and identification of Botryosphaeria species and related ana- morphic fungi have traditionally been based on morphology. While the morphology of the teleomorphic states differs little between species, a wide range of morphologies is seen in the anamorphs, and it is on this basis that species are distinguished [5,9,15]. Recently, Crous et al. [5] showed that the genus Botryosphaeria sensu lato is composed of distinct phyloge- netic lineages that can be defined on the basis of the anamorph genera. Thus, the genus name Botryosphaeria can only be applied to Botryosphaeria dothidea (type species) and closely related species. Species identification has relied heavily on morphological features of the conidia, including size, shape, color, septation, wall thickness and texture. However, some morphological characters exhibit extensive plasticity. Thus, size ranges of conidia of different species overlap, while age and state of maturity affect conidial pigmentation and septation. Morpho- logical characters can also be influenced by the substrate on which the fungus is growing. Moreover, a single species is able to colonize a wide range of hosts, and multiple species may be found on the same host [9,15]. Because species may differ in minor morphological features, identification can be a difficult task for those not familiar with these fungi. There- fore, there is a need for tools that can provide accurate and reproducible identifications of species of Botryosphaeriaceae. There has been increasing interest in the application of polymerase chain reaction (PCR) technology to identifica- tion of plant pathogenic fungi. Such methods offer the * Corresponding author. Tel.: þ351 234370970; fax: þ351 234426408. E-mail address: [email protected] (A. Alves). 0923-2508/$ - see front matter Ó 2006 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.resmic.2006.10.003 Research in Microbiology 158 (2007) 112e121 www.elsevier.com/locate/resmic
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Rapid differentiation of species of Botryosphaeriaceae by PCR fingerprinting

Research in Microbiology 158 (2007) 112e121www.elsevier.com/locate/resmic
Rapid differentiation of species of Botryosphaeriaceaeby PCR fingerprinting
Artur Alves a,*, Alan J.L. Phillips b, Isabel Henriques a, Antonio Correia a
a CESAM, Centro de Estudos do Ambiente e do Mar, Campus Universitario de Santiago, Departamento de Biologia,
Universidade de Aveiro, 3810-193 Aveiro, Portugalb Centro de Recursos Microbiologicos, Faculdade de Ciencias e Tecnologia, Universidade Nova de Lisboa,
Quinta da Torre, 2829-516 Caparica, Portugal
Received 2 June 2006; accepted 9 October 2006
Available online 20 December 2006
Abstract
The fingerprinting methods referred to as MSP-PCR (microsatellite-primed polymerase chain reaction) and rep-PCR (repetitive-sequence-based polymerase chain reaction) were used to discriminate between species of Botryosphaeriaceae (Fungi, Ascomycota). Several primerswere tested with both methods and each primer enabled clear differentiation of all the species tested. Cluster analysis of banding patterns ofthe isolates corresponded well with known species delineations based on morphology and phylogenetic analysis. The methods described inthis paper provide simple and rapid procedures that can be used for routine differentiation of Botryosphaeriaceae isolates at the species level.The two methods are also useful for studying intraspecific variability.� 2006 Elsevier Masson SAS. All rights reserved.
Keywords: Botryosphaeriaceae; Species differentiation; Fingerprinting; MSP-PCR; Rep-PCR
1. Introduction
Species of Botryosphaeria sensu lato are well known path-ogens, saprophytes and endophytes in a wide range of woodyangiosperm and gymnosperm hosts [4,9]. The classificationand identification of Botryosphaeria species and related ana-morphic fungi have traditionally been based on morphology.While the morphology of the teleomorphic states differs littlebetween species, a wide range of morphologies is seen in theanamorphs, and it is on this basis that species are distinguished[5,9,15]. Recently, Crous et al. [5] showed that the genusBotryosphaeria sensu lato is composed of distinct phyloge-netic lineages that can be defined on the basis of the anamorphgenera. Thus, the genus name Botryosphaeria can only be
* Corresponding author. Tel.: þ351 234370970; fax: þ351 234426408.
E-mail address: [email protected] (A. Alves).
0923-2508/$ - see front matter � 2006 Elsevier Masson SAS. All rights reserve
doi:10.1016/j.resmic.2006.10.003
applied to Botryosphaeria dothidea (type species) and closelyrelated species.
Species identification has relied heavily on morphologicalfeatures of the conidia, including size, shape, color, septation,wall thickness and texture. However, some morphologicalcharacters exhibit extensive plasticity. Thus, size ranges ofconidia of different species overlap, while age and state ofmaturity affect conidial pigmentation and septation. Morpho-logical characters can also be influenced by the substrate onwhich the fungus is growing. Moreover, a single species isable to colonize a wide range of hosts, and multiple speciesmay be found on the same host [9,15]. Because species maydiffer in minor morphological features, identification can bea difficult task for those not familiar with these fungi. There-fore, there is a need for tools that can provide accurate andreproducible identifications of species of Botryosphaeriaceae.
There has been increasing interest in the application ofpolymerase chain reaction (PCR) technology to identifica-tion of plant pathogenic fungi. Such methods offer the
d.

113A. Alves et al. / Research in Microbiology 158 (2007) 112e121
advantage of reducing or eliminating the need for lengthy cul-turing and difficult morphological identification procedures(e.g. [2,21]).
DNA-based techniques provide practical markers for mo-lecular typing of a range of species of fungi. These methodsare commonly used as tools in fungal taxonomy, allowingthe discrimination of isolates from intrageneric to strain levels[34]. Recently, Alves et al. [2] developed an ARDRA (ampli-fied ribosomal DNA restriction analysis) procedure for theidentification of ‘‘Botryosphaeria’’ species. The method relieson PCR amplification of a rDNA fragment followed by restric-tion analysis of the amplicons with frequent-cutting endonu-cleases. Using a combination of two restriction enzymes, itwas possible to identify 10 Botryosphaeria species.
PCR-based genomic fingerprinting is a good alternative tomethods that rely on specifically targeted primers. These tech-niques, which analyze the whole genome, have been shown tobe relatively robust and discriminatory [26,34]. Microsatellites(tandem repeats of 1e5 base pairs) and minisatellites (tandemrepeats of a basic motif 10e60 bp long) are ubiquitous compo-nents of eukaryotic genomes [41]. The microsatellite-primedpolymerase chain reaction (MSP-PCR) uses single primersto generate DNA fingerprints that are useful for discriminatingbetween fungal species and strains [24,25,37].
The repetitive-sequence-based polymerase chain reaction(rep-PCR) is a genomic fingerprinting technique that hasproven useful for molecular typing and identification ofbacterial species [39,40]. This technique is based on PCRamplification of DNA sequences located between specificinterspersed repeated sequences in prokaryotic genomes.Amplification generates an array of DNA fragments fromthe genomes of individual strains that can be separated inagarose gels, yielding highly specific DNA fingerprints.Several families of interspersed repetitive sequences are pres-ent in the genomes of bacterial species, e.g. the repetitiveextragenic palindromic (REP) sequence, the enterobacterialrepetitive intergenic consensus (ERIC) and the BOX element[26,29,39,40].
Although rep-PCR primers were developed for repetitiveelements in prokaryotic genomes, these primers have beenapplied with success to the fingerprinting of eukaryotic genomesincluding fungi [11]. Thus, rep-PCR primers have been used tocharacterize variability at inter- and/or intraspecific levels ofseveral fungal genera [6,7,13,16,22,38].
Despite the widespread application of these PCR tech-niques, few studies have been performed on the applicabilityof MSP-PCR [10,24,25,37] and rep-PCR [14,21,28] to identi-fication of fungal species.
The PCR-based method RAPD (random amplified poly-morphic DNA) has been used to characterize populations ofthe anamorphic species Sphaeropsis sapinea [32,35,36], whileMSP-PCR has been used to characterize populations ofB. dothidea [18e20]. The latter technique proved useful fordifferentiating between ‘‘Botryosphaeria’’ species [33,43].Zhou et al. [43] using MSP-PCR (or ISSR fingerprinting)were capable of discriminating between 10 ‘‘Botryosphaeria’’species and associated anamorphic fungi.
The purpose of the present work was to test the applicabil-ity of the MSP-PCR and rep-PCR genomic fingerprintingmethodologies as tools for differentiation of species in the Bo-tryosphaeriaceae. The potential of both methods for discrimi-nating between strains of the same species was also analyzed.
2. Materials and methods
2.1. Fungal strains and culture conditions
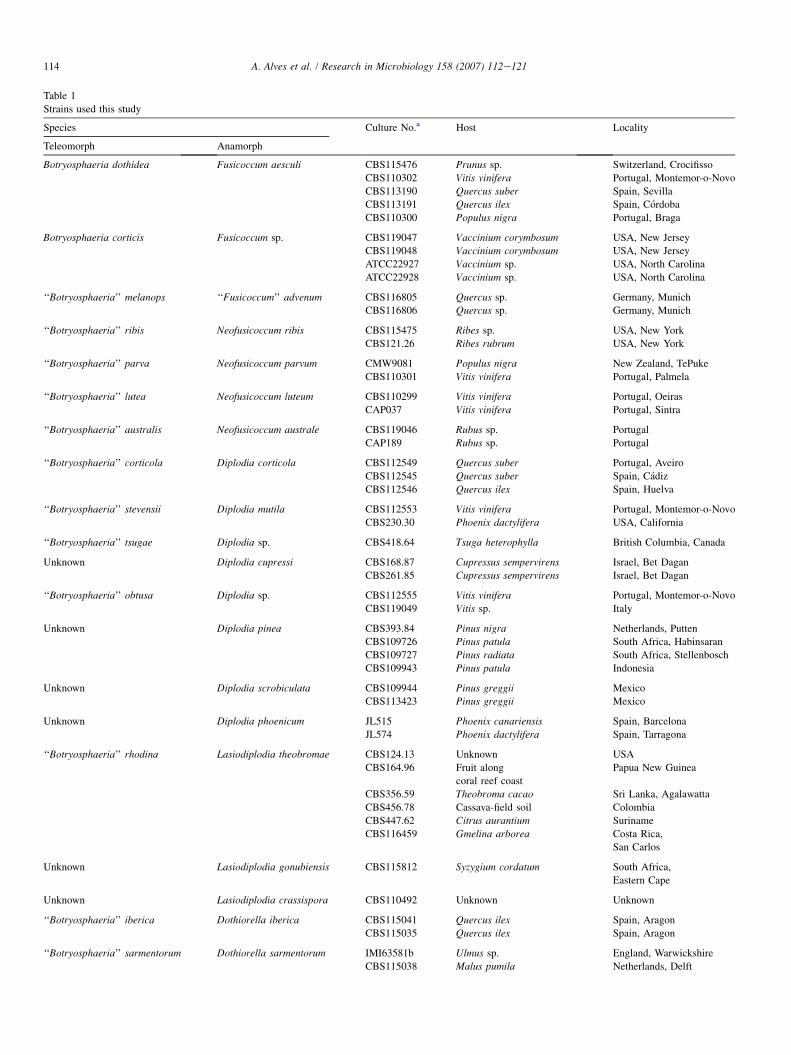
The fungal strains used in this study are listed in Table 1. Atotal of 53 strains representing 27 species were used. Cultureswere maintained on half-strength PDA (DIFCO) or oatmealagar [3].
2.2. DNA isolation
Fungal strains were grown in potato dextrose broth for 5days at approximately 25 �C. Genomic DNA was isolatedfrom fresh mycelium as described previously [1].
2.3. PCR amplification and gel electrophoresis
The primers used for each fingerprinting method are listedin Table 2. All primers were synthesized by MWG-BiotechAG (Ebersberg, Germany). All PCR reactions were carriedout with Taq polymerase, nucleotides and buffers from MBIFermentas (Vilnius, Lithuania), in a Bio-Rad iCycler ThermalCycler (Hercules, CA, USA). PCRs were performed in 25 mlreaction mixtures containing 1� PCR buffer (PCR bufferwithout MgCl2:PCR buffer with (NH4)2SO4, 1:1), 3 mMMgCl2, 200 mM each nucleotide, 20 pmol of each primer(for MSP-PCR primers and primer BOX) and 50 pmol ofeach primer (for REP and ERIC primers), 1 U of Taq polymer-ase, and 50e100 ng of purified template DNA. For each PCRreaction an initial denaturation step at 95 �C for 5 min wasdone; after that, 30 cycles of denaturation, annealing, andextension were performed according to Table 2. A final exten-sion step at 72 �C for 10 min (for MSP-PCR) and 65 �C for18 min (for rep-PCR) was included. Negative control reactionswithout any template DNA were carried out simultaneously.
PCR products were separated by electrophoresis in 2% aga-rose gels at 80 V for 3 h in Tris-acetate-EDTA buffer. A Gen-eRuler� DNA Ladder Mix (MBI Fermentas) was run at bothsides of each gel. The gels were stained with ethidium bromideand visualized on a UV transilluminator to assess PCRamplification.
2.4. Computer-assisted analysis of genomic fingerprints
Agarose gels were scanned on a Molecular Imager FX(BioRad) and recorded as TIFF images. The banding patternswere analyzed with GelCompar II software (Applied Maths).Gels were normalized using the same standard loaded atboth sides. The ‘‘rolling disk’’ background subtraction optionwas applied. DNA bands detected by the software were care-fully verified by visual examination to correct unsatisfactory
114 A. Alves et al. / Research in Microbiology 158 (2007) 112e121
Table 1
Strains used this study
Species Culture No.a Host Locality
Teleomorph Anamorph
Botryosphaeria dothidea Fusicoccum aesculi CBS115476 Prunus sp. Switzerland, Crocifisso
CBS110302 Vitis vinifera Portugal, Montemor-o-Novo
CBS113190 Quercus suber Spain, Sevilla
CBS113191 Quercus ilex Spain, Cordoba
CBS110300 Populus nigra Portugal, Braga
Botryosphaeria corticis Fusicoccum sp. CBS119047 Vaccinium corymbosum USA, New Jersey
CBS119048 Vaccinium corymbosum USA, New Jersey
ATCC22927 Vaccinium sp. USA, North Carolina
ATCC22928 Vaccinium sp. USA, North Carolina
‘‘Botryosphaeria’’ melanops ‘‘Fusicoccum’’ advenum CBS116805 Quercus sp. Germany, Munich
CBS116806 Quercus sp. Germany, Munich
‘‘Botryosphaeria’’ ribis Neofusicoccum ribis CBS115475 Ribes sp. USA, New York
CBS121.26 Ribes rubrum USA, New York
‘‘Botryosphaeria’’ parva Neofusicoccum parvum CMW9081 Populus nigra New Zealand, TePuke
CBS110301 Vitis vinifera Portugal, Palmela
‘‘Botryosphaeria’’ lutea Neofusicoccum luteum CBS110299 Vitis vinifera Portugal, Oeiras
CAP037 Vitis vinifera Portugal, Sintra
‘‘Botryosphaeria’’ australis Neofusicoccum australe CBS119046 Rubus sp. Portugal
CAP189 Rubus sp. Portugal
‘‘Botryosphaeria’’ corticola Diplodia corticola CBS112549 Quercus suber Portugal, Aveiro
CBS112545 Quercus suber Spain, Cadiz
CBS112546 Quercus ilex Spain, Huelva
‘‘Botryosphaeria’’ stevensii Diplodia mutila CBS112553 Vitis vinifera Portugal, Montemor-o-Novo
CBS230.30 Phoenix dactylifera USA, California
‘‘Botryosphaeria’’ tsugae Diplodia sp. CBS418.64 Tsuga heterophylla British Columbia, Canada
Unknown Diplodia cupressi CBS168.87 Cupressus sempervirens Israel, Bet Dagan
CBS261.85 Cupressus sempervirens Israel, Bet Dagan
‘‘Botryosphaeria’’ obtusa Diplodia sp. CBS112555 Vitis vinifera Portugal, Montemor-o-Novo
CBS119049 Vitis sp. Italy
Unknown Diplodia pinea CBS393.84 Pinus nigra Netherlands, Putten
CBS109726 Pinus patula South Africa, Habinsaran
CBS109727 Pinus radiata South Africa, Stellenbosch
CBS109943 Pinus patula Indonesia
Unknown Diplodia scrobiculata CBS109944 Pinus greggii Mexico
CBS113423 Pinus greggii Mexico
Unknown Diplodia phoenicum JL515 Phoenix canariensis Spain, Barcelona
JL574 Phoenix dactylifera Spain, Tarragona
‘‘Botryosphaeria’’ rhodina Lasiodiplodia theobromae CBS124.13 Unknown USA
CBS164.96 Fruit along
coral reef coast
Papua New Guinea
CBS356.59 Theobroma cacao Sri Lanka, Agalawatta
CBS456.78 Cassava-field soil Colombia
CBS447.62 Citrus aurantium Suriname
CBS116459 Gmelina arborea Costa Rica,
San Carlos
Unknown Lasiodiplodia gonubiensis CBS115812 Syzygium cordatum South Africa,
Eastern Cape
Unknown Lasiodiplodia crassispora CBS110492 Unknown Unknown
‘‘Botryosphaeria’’ iberica Dothiorella iberica CBS115041 Quercus ilex Spain, Aragon
CBS115035 Quercus ilex Spain, Aragon
‘‘Botryosphaeria’’ sarmentorum Dothiorella sarmentorum IMI63581b Ulmus sp. England, Warwickshire
CBS115038 Malus pumila Netherlands, Delft
115A. Alves et al. / Research in Microbiology 158 (2007) 112e121
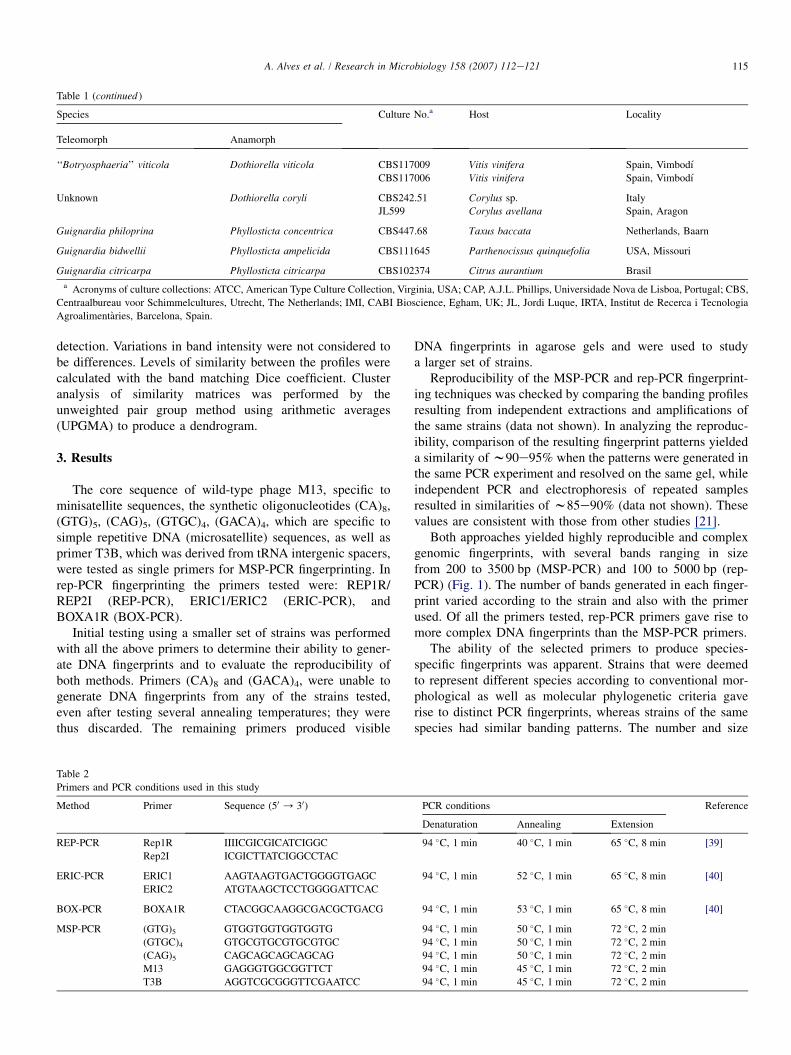
Table 1 (continued )
Species Culture No.a Host Locality
Teleomorph Anamorph
‘‘Botryosphaeria’’ viticola Dothiorella viticola CBS117009 Vitis vinifera Spain, Vimbodı
CBS117006 Vitis vinifera Spain, Vimbodı
Unknown Dothiorella coryli CBS242.51 Corylus sp. Italy
JL599 Corylus avellana Spain, Aragon
Guignardia philoprina Phyllosticta concentrica CBS447.68 Taxus baccata Netherlands, Baarn
Guignardia bidwellii Phyllosticta ampelicida CBS111645 Parthenocissus quinquefolia USA, Missouri
Guignardia citricarpa Phyllosticta citricarpa CBS102374 Citrus aurantium Brasil
a Acronyms of culture collections: ATCC, American Type Culture Collection, Virginia, USA; CAP, A.J.L. Phillips, Universidade Nova de Lisboa, Portugal; CBS,
Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands; IMI, CABI Bioscience, Egham, UK; JL, Jordi Luque, IRTA, Institut de Recerca i Tecnologia
Agroalimentaries, Barcelona, Spain.
detection. Variations in band intensity were not considered tobe differences. Levels of similarity between the profiles werecalculated with the band matching Dice coefficient. Clusteranalysis of similarity matrices was performed by theunweighted pair group method using arithmetic averages(UPGMA) to produce a dendrogram.
3. Results
The core sequence of wild-type phage M13, specific tominisatellite sequences, the synthetic oligonucleotides (CA)8,(GTG)5, (CAG)5, (GTGC)4, (GACA)4, which are specific tosimple repetitive DNA (microsatellite) sequences, as well asprimer T3B, which was derived from tRNA intergenic spacers,were tested as single primers for MSP-PCR fingerprinting. Inrep-PCR fingerprinting the primers tested were: REP1R/REP2I (REP-PCR), ERIC1/ERIC2 (ERIC-PCR), andBOXA1R (BOX-PCR).
Initial testing using a smaller set of strains was performedwith all the above primers to determine their ability to gener-ate DNA fingerprints and to evaluate the reproducibility ofboth methods. Primers (CA)8 and (GACA)4, were unable togenerate DNA fingerprints from any of the strains tested,even after testing several annealing temperatures; they werethus discarded. The remaining primers produced visible
DNA fingerprints in agarose gels and were used to studya larger set of strains.
Reproducibility of the MSP-PCR and rep-PCR fingerprint-ing techniques was checked by comparing the banding profilesresulting from independent extractions and amplifications ofthe same strains (data not shown). In analyzing the reproduc-ibility, comparison of the resulting fingerprint patterns yieldeda similarity of w90e95% when the patterns were generated inthe same PCR experiment and resolved on the same gel, whileindependent PCR and electrophoresis of repeated samplesresulted in similarities of w85e90% (data not shown). Thesevalues are consistent with those from other studies [21].
Both approaches yielded highly reproducible and complexgenomic fingerprints, with several bands ranging in sizefrom 200 to 3500 bp (MSP-PCR) and 100 to 5000 bp (rep-PCR) (Fig. 1). The number of bands generated in each finger-print varied according to the strain and also with the primerused. Of all the primers tested, rep-PCR primers gave rise tomore complex DNA fingerprints than the MSP-PCR primers.
The ability of the selected primers to produce species-specific fingerprints was apparent. Strains that were deemedto represent different species according to conventional mor-phological as well as molecular phylogenetic criteria gaverise to distinct PCR fingerprints, whereas strains of the samespecies had similar banding patterns. The number and size
Table 2
Primers and PCR conditions used in this study
Method Primer Sequence (50 / 30) PCR conditions Reference
Denaturation Annealing Extension
REP-PCR Rep1R IIIICGICGICATCIGGC 94 �C, 1 min 40 �C, 1 min 65 �C, 8 min [39]
Rep2I ICGICTTATCIGGCCTAC
ERIC-PCR ERIC1 AAGTAAGTGACTGGGGTGAGC 94 �C, 1 min 52 �C, 1 min 65 �C, 8 min [40]
ERIC2 ATGTAAGCTCCTGGGGATTCAC
BOX-PCR BOXA1R CTACGGCAAGGCGACGCTGACG 94 �C, 1 min 53 �C, 1 min 65 �C, 8 min [40]
MSP-PCR (GTG)5 GTGGTGGTGGTGGTG 94 �C, 1 min 50 �C, 1 min 72 �C, 2 min
(GTGC)4 GTGCGTGCGTGCGTGC 94 �C, 1 min 50 �C, 1 min 72 �C, 2 min
(CAG)5 CAGCAGCAGCAGCAG 94 �C, 1 min 50 �C, 1 min 72 �C, 2 min
M13 GAGGGTGGCGGTTCT 94 �C, 1 min 45 �C, 1 min 72 �C, 2 min
T3B AGGTCGCGGGTTCGAATCC 94 �C, 1 min 45 �C, 1 min 72 �C, 2 min

116 A. Alves et al. / Research in Microbiology 158 (2007) 112e121
Fig. 1. Examples of MSP-PCR and rep-PCR fingerprints obtained with primer (GTG)5 and the ERIC primer set. (M) GeneRuler DNA Ladder Mix; (1) L. gonu-
biensis CBS115812; (2) L. crassispora CBS110492; (3e6) L. theobromae CBS356.59, CBS456.78, CBS447.62 and CBS116459; (7) ‘B’. tsugae CBS418.64; (8,9)
D. pinea morphotype A CBS393.84 and CBS109727; (10,11) D. pinea morphotype C CBS109726 and CBS109943; (12,13) D. scrobiculata CBS109944 and
CBS113423.
of the multilocus PCR fingerprint bands were clearly species-specific. Based on the specific PCR fingerprints and the highinterspecies variation of these banding patterns, a clear distinc-tion between all species was possible. Although strains of thesame species always generated very similar fingerprints, bothmethodologies (MSP-PCR and rep-PCR) remained capable ofdetecting intraspecific variability.
Cluster analyses were performed on the genomic finger-prints generated by each of the primers tested. Samples ofthe dendrograms generated with the UPGMA method areshown in Figs. 2 and 3. For all primers analyzed the clusteringof the strains was species-specific. Strains belonging to thesame species typically clustered together at similarity valuesgreater than 80%. The only two exceptions found were thespecies Dothiorella viticola and Dothiorella coryli. The twostrains of each species did not cluster together in some ofthe analyses, and when they grouped together in the same clus-ter the similarity between the fingerprints was below 80%.Also, the isolates identified as Lasiodiplodia theobromaeformed three separate and well defined clusters, each with sim-ilarity values well above 80%.
MSP-PCR and rep-PCR fingerprints and cluster analysisrevealed no relationships at the genus level. Thus, speciesbelonging to the same genus were not grouped into clearlydefined and separate clusters.
The fingerprints obtained with all the MSP-PCR primerswere subjected to combined cluster analysis, and the samewas performed for BOX, ERIC, and REP fingerprints. Inboth analyses, the dendrograms produced were similar to
those produced by cluster analysis of the non-combinedfingerprints.
4. Discussion
The identification of species in the Botryosphaeriaceae canbe a difficult task if based solely on morphological and cul-tural characters. In this respect, the application of molecularmethods, particularly PCR-based techniques, may provide animportant contribution.
MSP-PCR and rep-PCR fingerprinting have proven usefulin the molecular typing of fungal strains [6,7,13,16,22,38].Recently, some studies have addressed its application in theidentification of species of yeasts [14,24,25,37] and filamen-tous fungi [10,21,28], showing that these methodologieshave great potential as diagnostic tools. In a previous work,Zhou et al. [43] reported that MSP-PCR, which they referredto as ISSR fingerprinting, enabled the differentiation of 10Botryosphaeria species. In the present study, we tested theapplicability of these fingerprinting methods to the discrimina-tion of 27 species.
Of all the primers tested, only the microsatellite repeats(CA)8 and (GACA)4 were unable to generate visible DNAfingerprints. It is possible that these repeats are not presentin the genomes of these fungi. Another possibility is thatthe primer annealing sites are at such a distance that amplifica-tion by Taq polymerase is impossible. Zhou et al. [43] reportedthat no tetra-nucleotide repeat primers worked well forBotryosphaeria species. We tested two tetra-nucleotide repeat

117A. Alves et al. / Research in Microbiology 158 (2007) 112e121
Fig. 2. UPGMA cluster analysis based on the Dice coefficient of MSP-PCR fingerprints obtained with primer (GTG)5. Similarity is indicated as a percentage.
primers, namely (GACA)4 and (GTGC)4. The resultsobtained with (GACA)4 are in agreement with those reportedby Zhou et al. [43], but the primer (GTGC)4 generatedfingerprints with all species studied.
Primers REP, ERIC, and BOX used for rep-PCR finger-printing all proved useful for generating highly reproducibleand discriminatory fingerprints with all species tested. Theseprimers are derived from repetitive sequences present in

118 A. Alves et al. / Research in Microbiology 158 (2007) 112e121
Fig. 3. UPGMA cluster analysis based on the Dice coefficient of rep-PCR fingerprints obtained with primers ERIC1/ERIC2 (ERIC-PCR). Similarity is indicated as
a percentage.

119A. Alves et al. / Research in Microbiology 158 (2007) 112e121
prokaryotic genomes [39,40]. It is not known whether these re-petitive sequences are also present in eukaryotic genomes, butit was shown that these primers are able to generate finger-prints in a wide diversity of eukaryotes [11]. Some studieshave shown that these primers amplify regions that are notnecessarily related to the prokaryotic REP, ERIC and BOXregions [12,23].
The MSP-PCR and rep-PCR fingerprinting methods pre-sented here led to clear differentiation of the isolates at the spe-cies level. Fingerprinting profiles generated by all the primerstested readily discriminated between each of the 27 species.Cluster analysis further supported this observation and clusterscorresponding to each species are readily identified in the den-drograms. As can be seen in the dendrograms, clusters formedat similarity values �80% correspond to known species. Bothprocedures appear to be excellent tools for the differentiationof even cryptic species which can be easily distinguished bythe fingerprints generated with any of the MSP-PCR andrep-PCR primers tested. A clear separation of the speciesaccording to the genus was not possible. MSP-PCR and rep-PCR fingerprinting methods analyze regions of the genomethat are extremely variable and are thus better suited for taxo-nomic studies at or below the species level [29,41].
In recent years, sequence analyses of the ITS region alone[1,9,15,42] or in combination with introns of protein-encodinggenes such as b-tubulin and the translation elongation factor1-alpha (EF1-a) [17,27,30,31] have been used to study rela-tionships among species and to distinguish closely related‘‘Botryosphaeria’’ species. Some cryptic species like Neofusi-coccum ribis and Neofusicoccum parvum or Neofusicoccumluteum and Neofusicoccum australis, are very similar in mor-phological terms, and reliable identifications are difficult toaccomplish without the use of molecular methods [30,31]. Infact, nucleotide sequences of the ITS region alone cannotclearly discriminate between the species and it is necessaryto combine this region with partial sequence of the EF1-a gene [30,31]. These four species are easily separated by theirMSP-PCR or rep-PCR fingerprints and form separate clustersreadily identified by cluster analyses.
Dothiorella sarmentorum, Dothiorella iberica, and D. viti-cola are recently described species that can be readily distin-guished based on teleomorph features. However, this form isextremely rare, and the anamorphic forms are more abundantin nature. The anamorphs can be separated by minor differ-ences in the size and shape of the conidia, thus complicatingthe identification [17,27]. In the same way as other ‘‘Botryos-phaeria’’ species, these can be differentiated by sequenceanalysis of the ITS and EF1-a regions [17,27]. The DNA fin-gerprints generated by MSP-PCR and rep-PCR allow a cleardifferentiation of these species.
Two isolates of D. viticola were analyzed in this study. Inthe majority of the analyses, these isolates did not appear inthe same cluster. When describing D. viticola, Luque et al.[17] reported that isolate CBS117006 exhibited some differ-ences in culture morphology from the other isolates studied.Furthermore, this isolate differed in ITS and EF1-a sequences(one substitution and one deletion in ITS and nine substitutions
in EF1-a). These differences in nucleotide sequences andcolony morphology were regarded as intraspecific variation.The results present here also show that strain CBS117006 ismarkedly different from the other D. viticola strain tested(CBS117009). Due to the differences in ITS and EF1-asequences as well as MSP-PCR and rep-PCR fingerprints, itis likely that strain CBS 117006 identified as D. viticola mayin fact represent a separate taxon. In order to confirm thishypothesis a larger set of similar isolates would need to bestudied.
The same possibility may be true for the two strains iden-tified as D. coryli (CBS242.51 and JL599) which have gener-ated divergent MSP-PCR and rep-PCR fingerprints. Thesestrains differ by one nucleotide substitution in the ITS region,as well as five nucleotide substitutions and one deletion in theEF1-a region (data not shown).
The isolates of L. theobromae analyzed in this studyrevealed an extremely high genetic diversity forming threeseparate groups that can be easily identified in cluster analy-ses. These isolates, although initially identified as membersof the species L. theobromae, are now deemed to representthree separate taxa based on morphological and phylogeneticevidences (Alves et al., unpublished).
Diplodia scrobiculata is clearly separated from Diplodiapinea on the basis of the DNA fingerprints produced by thetwo methods. Also, strains belonging to different D. pinea mor-photypes (A and C), although very closely related, can be sep-arated by their MSP-PCR and rep-PCR fingerprints. It is notclear whether these morphotypes represent distinct taxa [8],but due to the large number of bands shared in the DNA finger-prints it is likely that the differences between the morphotypesreflect only intraspecific variability. This, however, can only beverified through comparison of a larger number of isolates.
The PCR fingerprinting methods tested, although producingfingerprints that are very similar within strains of the samespecies, are also capable of detecting intraspecific variation.As can be seen in the fingerprints, it is possible to identifypolymorphic bands between strains of the same species.MSP-PCR and rep-PCR fingerprinting can be used to studygenetic variability and population structure of species in theBotryosphaeriaceae, and both are a good alternative toRAPD analysis. The major advantage of MSP-PCR and rep-PCR when compared to RAPD is the capacity to generatehighly reproducible results. Higher reproducibility is relatedto the fact that longer primers are used and consequentlyPCR reactions are carried out at higher annealing temperatures[34,41].
Previous studies with other fungi have shown that MSP-PCR and rep-PCR fingerprinting are well suited for detectingintraspecific variability (e.g. [6,13,16]). The high discrimina-tory potential of these methods is particularly useful whendealing with closely related or cryptic species. Both methodsclearly differentiated between cryptic species in the Botryos-phaeriaceae. Nevertheless, in order to fully understand inter-and intraspecific variation within this group of fungi, furtherstudies with a larger number of isolates and species shouldbe performed.

120 A. Alves et al. / Research in Microbiology 158 (2007) 112e121
Reproducibility is extremely important in any PCR-basedfingerprinting method. In this work, particular attention wasgiven to this aspect during optimization of the PCR reactions.In order to obtain reproducible fingerprint patterns, it wasessential to maintain standardized conditions, such as theconcentrations of reagents (especially magnesium and primers)and cycling conditions. The addition of 5% DMSO to the reac-tion proved useful to help the amplification of some difficulttemplates and also increased reproducibility. Slight variationsin the PCR reaction conditions were directly related to varia-tions in the fingerprints. These minor changes, however, donot affect the discriminatory power of the procedure.
We have observed some variability in fingerprint patternspredominantly in the intensity of weaker bands. However,when the experimental conditions (PCR and electrophoresis)were strictly maintained, multiple tests of an individual straingenerated identical fingerprints.
The results of this study demonstrate the usefulness ofMSP-PCR and rep-PCR genomic fingerprinting as comple-mentary or alternate strategies for differentiation of speciesin the Botryosphaeriaceae. Recently an ARDRA procedurewas described that could be used to identify ‘‘Botryosphaeria’’species [2]. Like ARDRA, MSP-PCR and rep-PCR finger-printing are simple and fast alternatives to sequencing whichcan be extremely useful when dealing with large sets ofisolates. These tools will be very useful for screening largecollections of known species and sorting them into thosedefined taxa based on reference cultures. It will also be usefulto characterize undefined collections, in order to group isolatesinto potential species groups. The advantages of the threemethods are simplicity, universal availability of PCR primers,reproducibility and amenability to computer database analysis.It is possible to build databases of fingerprints that can be usedfor routine identification of isolates by simple comparison offingerprints with those of reference cultures.
The major advantage of MSP-PCR and rep-PCR over AR-DRA is the fact that there is no need for a step involving thedigestion of PCR amplicons with restriction endonucleases. Inorder to discriminate between 10 ‘‘Botryosphaeria’’ spp. it wasnecessary to use combinations of two restriction enzymes inthe ARDRA procedure [2]. Using only a single primer or setof primers in MSP-PCR and rep-PCR, it is possible to achievehigher levels of resolution. This makes the procedure muchfaster and easier, and greatly reduces the cost.
Pounder et al. [28] tested the performance of rep-PCR usingthe DiversiLab system for identification of dermatophytes.Those authors reported that the turnaround time for both iden-tification and typing results is <24 h once a pure culture isavailable, versus several days to weeks for fungal identifica-tion, and 2 days when using sequencing for identification.Similar timescales are applicable to the differentiation ofbotryosphaeriaceous fungi.
Acknowledgments
This work was financed by the Fundac~ao para a Ciencia ea Tecnologia (FCT) under project POCTI/AGR/44348/2002
(Portugal). A. Alves and I. Henriques were supported by grantnumbers SFRH/BD/10389/2002 and SFRH/BD/5275/2001from FCT. M.E. Sanchez and A. Trapero (University of Cor-doba, Spain), J. Luque (IRTA, Spain) and M.J. Wingfield(FABI, University of Pretoria, South Africa) donated someof the strains included in this work. The authors are thankfulto Dr. Paula Teixeira (Escola Superior de Biotecnologia,Universidade Catolica Portuguesa) for allowing the use ofthe GelCompar II software.
References
[1] A. Alves, A. Correia, J. Luque, A.J.L. Phillips, Botryosphaeria corticola
sp. nov. on Quercus species, with notes and description of Botryosphae-
ria stevensii and its anamorph Diplodia mutila, Mycologia 96 (2004)
598e613.
[2] A. Alves, A.J.L. Phillips, I. Henriques, A. Correia, Evaluation of ampli-
fied ribosomal DNA restriction analysis (ARDRA) as a method for the
identification of Botryosphaeria species, FEMS Microbiol. Lett. 245
(2005) 221e229.
[3] Anonymous, Plant Pathologists Pocketbook, Kew, England, 1968.
[4] M.E. Barr, Prodromus to Class Loculoascomycetes Published by the
author, Amherst, Massachusetts, 1987.
[5] P.W. Crous, B. Slippers, M.J. Wingfield, J. Reeder, W.F.O. Marasas,
A.J.L. Phillips, A. Alves, T. Burgess, P. Barber, J.Z. Groenewald, Phy-
logenetic lineages in the Botryosphaeriaceae, Stud. Mycol. 55 (2006)
235e253.
[6] P.C. Czembor, E. Arseniuk, Study of genetic variability among monop-
ycnidial and monopycnidiospore isolates derived from single pycnidia
of Stagonospora spp. and Septoria tritici with the use of RAPD-PCR,
MP-PCR and rep-PCR techniques, J. Phytopathol. 147 (1999) 539e546.
[7] M.C.C. de Arruda, R.N.G. Miller, M.A.S.V. Ferreira, M.S.S. Felipe,
Comparison of Crinipellis perniciosa isolates from Brazil by ERIC repet-
itive element sequence-based PCR genomic fingerprinting, Plant Pathol.
52 (2003) 236e244.
[8] J. de Wet, T. Burgess, B. Slippers, O. Preisig, B.D. Wingfield,
M.J. Wingfield, Multiple gene genealogies and microsatellite markers re-
flect relationships between morphotypes of Sphaeropsis sapinea and dis-
tinguish a new species of Diplodia, Mycol. Res. 107 (2003) 557e566.
[9] S. Denman, P.W. Crous, J.E. Taylor, J.-C. Kang, I. Pascoe,
M.J. Wingfield, An overview of the taxonomic history of Botryosphaeria,
and a re-evaluation of its anamorphs based on morphology and ITS
rDNA phylogeny, Stud. Mycol. 45 (2000) 129e140.
[10] E. Faggi, G. Pini, E. Campisi, C. Bertellini, E. Difonzo, F. Mancianti,
Application of PCR to distinguish common species of dermatophytes,
J. Clin. Microbiol. 39 (2001) 3382e3385.
[11] M. Gillings, M. Holley, Amplification of anonymous DNA fragments
using pairs of long primers generates reproducible DNA fingerprints
that are sensitive to genetic variation, Electrophoresis 18 (1997) 1512e1518.
[12] M. Gillings, M. Holley, Repetitive element PCR fingerprinting (rep-PCR)
using enterobacterial repetitive intergenic consensus (ERIC) primers is
not necessarily directed at ERIC elements, Lett. Appl. Microbiol. 25
(1997) 17e21.
[13] P. Godoy, J. Cano, J. Gene, J. Guarro, A.L. Hofling-Lima, A.L. Colombo,
Genotyping of 44 isolates of Fusarium solani, the main agent of fungal
keratitis in Brazil, J. Clin. Microbiol. 42 (2004) 4494e4497.
[14] N. Hierro, �A. Gonzalez, A. Mas, J.M. Guillamon, New PCR-based
methods for yeast identification, J. Appl. Microbiol. 97 (2004) 792e801.
[15] K.A. Jacobs, S.A. Rehner, Comparison of cultural and morphological
characters and ITS sequences in anamorphs of Botryosphaeria and
related taxa, Mycologia 90 (1998) 601e610.
[16] M. Jedryczka, T. Rouxel, M.H. Balesdent, Rep-PCR based genomic fin-
gerprinting of isolates of Leptosphaeria maculans from Poland, Eur. J.
Plant Pathol. 105 (1999) 813e823.

121A. Alves et al. / Research in Microbiology 158 (2007) 112e121
[17] J. Luque, S. Martos, A.J.L. Phillips, Botryosphaeria viticola sp. nov. on
grapevines: a new species with a Dothiorella anamorph, Mycologia 97
(2005) 1111e1121.
[18] Z. Ma, T.J. Michailides, Characterization of Botryosphaeria dothidea
isolates collected from pistachio and other plant hosts in California,
Phytopathology 92 (2002) 519e526.
[19] Z. Ma, E.W.A. Boehm, Y. Luo, T.J. Michailides, Population structure of
Botryosphaeria dothidea from pistachio and other hosts in California,
Phytopathology 91 (2001) 665e672.
[20] Z. Ma, Y. Luo, T.J. Michailides, Spatiotemporal changes in the popula-
tion structure of Botryosphaeria dothidea from California pistachio
orchards, Phytopathology 94 (2004) 326e332.
[21] J.G. McDonald, E. Wong, G.P. White, Differentiation of Tilletia species
by rep-PCR genomic fingerprinting, Plant Dis. 84 (2000) 1121e1125.
[22] A. Mehta, Y.R. Mehta, Y.B. Rosato, ERIC and REP-PCR banding pat-
terns and sequence analysis of the internal transcribed spacer of rDNA
of Stemphylium solani isolates from cotton, Curr. Microbiol. 44 (2002)
323e328.
[23] A. Mehta, Y.R. Mehta, Y.B. Rosato, ERIC- and REP-PCR amplify non-
repetitive fragments from the genome of Drechslera avenae and
Stemphylium solani, FEMS Microbiol. Lett. 211 (2002) 51e55.
[24] W. Meyer, G.N. Latouche, H.M. Daniel, M. Thanos, T.G. Mitchell,
D. Yarrow, G. Schonian, T.C. Sorrell, Identification of pathogenic yeasts
of the imperfect genus Candida by polymerase chain reaction fingerprint-
ing, Electrophoresis 18 (1997) 1548e1559.
[25] W. Meyer, T.G. Mitchell, E.Z. Freedman, R. Vilgalys, Hybridization
probes for conventional DNA fingerprinting used as single primers in
the polymerase chain reaction to distinguish strains of Cryptococcus neo-
formans, J. Clin. Microbiol. 31 (1993) 2274e2280.
[26] D.M. Olive, P. Bean, Principles and applications of methods for DNA-
based typing of microbial organisms, J. Clin. Microbiol. 37 (1999)
1661e1669.
[27] A.J.L. Phillips, A. Alves, A. Correia, J. Luque, Two new species of
Botryosphaeria with brown, one-septate ascospores and Dothiorellaanamorphs, Mycologia 97 (2005) 513e529.
[28] J.I. Pounder, S. Williams, D. Hansen, M. Healy, K. Reece, G.L. Woods,
Repetitive-sequence-PCR-based DNA fingerprinting using the Diversi-
Lab system for identification of commonly encountered dermatophytes,
J. Clin. Microbiol. 43 (2005) 2141e2147.
[29] J.L.W. Rademaker, F.J. de Bruijn, DNA markers: protocols, applications
and overviews, in: G. Caetano-Anolles, P.M. Gresshoff (Eds.), John
Wiley and Sons, New York, 1997, pp. 151e171.
[30] B. Slippers, P.W. Crous, S. Denman, T.A. Coutinho, B.D. Wingfield,
M.J. Wingfield, Combined multiple gene genealogies and phenotypic
characters differentiate several species previously identified as Botryos-phaeria dothidea, Mycologia 96 (2004) 83e101.
[31] B. Slippers, G. Fourie, P.W. Crous, T.A. Coutinho, B.D. Wingfield,
M.J. Wingfield, Multiple gene sequences delimit Botryosphaeria
australis sp. nov. from B. lutea, Mycologia 96 (2004) 1030e1041.
[32] D.R. Smith, G.R. Stanosz, Confirmation of two distinct populations of
Sphaeropsis sapinea in the North Central United States using RAPDs,
Phytopathology 85 (1995) 699e704.
[33] D.R. Smith, T.J. Michailides, G.R. Stanosz, Differentiation of a Fusicoc-
cum sp. causing panicle and shoot blight on California pistachio trees
from Botryosphaeria dothidea, Plant Dis. 85 (2001) 1235e1240.
[34] D.R. Soll, The ins and outs of DNA fingerprinting the infectious fungi,
Clin. Microbiol. Rev. 13 (2000) 332e370.
[35] G.R. Stanosz, D.R. Smith, M.A. Guthmiller, Characterization of Sphaer-
opsis sapinea from the West Central United States by means of random
amplified polymorphic DNA markers analysis, Plant Dis. 80 (1996)
1175e1178.
[36] G.R. Stanosz, W.J. Swart, D.R. Smith, RAPD marker and isozyme char-
acterization of Sphaeropsis sapinea from diverse coniferous hosts and
locations, Mycol. Res. 103 (1999) 1193e1202.
[37] M. Thanos, G. Schonian, W. Meyer, C. Schweynoch, Y. Graser,
T.G. Mitchell, W. Presber, H.J. Tietz, Rapid identification of Candida
species by DNA fingerprinting with PCR, J. Clin. Microbiol. 34 (1996)
615e621.
[38] A. van Belkum, W.G.V. Quint, B.E. de Pauw, W.J.G. Melchers, J.F. Meis,
Typing of Aspergillus species and Aspergillus fumigatus isolates by
interrepeat polymerase chain reaction, J. Clin. Microbiol. 31 (1993)
2502e2505.
[39] J. Versalovic, T. Koeuth, J.R. Lupski, Distribution of repetitive DNA
sequences in eubacteria and application to fingerprinting of bacterial
genomes, Nucleic Acids Res. 19 (1991) 6823e6831.
[40] J. Versalovic, M. Schneider, F.J. de Bruijn, J.R. Lupski, Genomic finger-
printing of bacteria using repetitive sequence-based polymerase chain
reaction, Methods Mol. Cell. Biol. 5 (1994) 25e40.
[41] J.M. Vogel, P.A. Scolnik, in: G. Caetano-Anolles, P.M. Gresshoff (Eds.),
DNA Markers: Protocols, Applications and Overviews, John Wiley and
Sons, New York, 1997, pp. 133e150.
[42] S. Zhou, G.R. Stanosz, Relationships among Botryosphaeria species and
associated anamorphic fungi inferred from the analyses of ITS and 5.8S
rDNA sequences, Mycologia 93 (2001) 516e527.
[43] S. Zhou, D.R. Smith, G.R. Stanosz, Differentiation of Botryosphaeria
species and related anamorphic fungi using Inter Simple or Short
Sequence Repeat (ISSR) fingerprinting, Mycol. Res. 105 (2001)
919e926.




















