Queue Size Determines the Width of Tunnels
16
Queue Size Determines the Width of Tunnels in the Formosan Subterranean Termite (Isoptera: Rhinotermitidae) Paul M. Bardunias & Nan-Yao Su Revised: 19 December 2008 / Accepted: 14 January 2010 / Published online: 3 February 2010 # Springer Science+Business Media, LLC 2010 Abstract We present a model of tunnel excavation by termites that requires no pheromone labeling of soil or work sites, but instead relies on tactile interactions and individuals who actively orient their movement. Potential termite excavators moved from the tunnel origin towards the distal end of the tunnel and formed a queue behind those termites at the digging face. Delayed termites excavated soil laterally from the tunnel wall at a position governed by their position in the queue of termites. By examining excavation under artificially induced conditions of longer and shorter queues of termites at the tunnel end, we showed that tunnel width increased with increased queue size and the rate of lateral excavation in a process we termed “digging pressure.” Keywords Excavation . tunnel . queue . coptotermes formosanus Introduction The construction of nests and galleries by groups of termites has been examined in a few termite species (Grasse 1959; Bruinsma 1979; Jones 1979). Grasse (1959) proposed the concept of stigmergy, an indirect mode of communication between termites whereby the work product of a builder acts to guide the response of subsequent workers, as a means to coordinate the labor of individual termites. In the context of termite construction this process relies on the application of various pheromones to mark sites or pathways and to identify the work product of construction to induce positive feedback loops, where deposition stimulates additional deposition. Based on empirical studies, self-organized nest building in J Insect Behav (2010) 23:189–204 DOI 10.1007/s10905-010-9206-z P. M. Bardunias (*) : N.-Y. Su Department of Entomology and Nematology, Ft. Lauderdale Research and Education Center, University of Florida, 3205 College Ave., Ft. Lauderdale, FL 33314, USA e-mail: [email protected]
Transcript of Queue Size Determines the Width of Tunnels

Queue Size Determines the Width of Tunnelsin the Formosan Subterranean Termite(Isoptera: Rhinotermitidae)
Paul M. Bardunias & Nan-Yao Su
Revised: 19 December 2008 /Accepted: 14 January 2010 /Published online: 3 February 2010# Springer Science+Business Media, LLC 2010
Abstract We present a model of tunnel excavation by termites that requires nopheromone labeling of soil or work sites, but instead relies on tactile interactions andindividuals who actively orient their movement. Potential termite excavators movedfrom the tunnel origin towards the distal end of the tunnel and formed a queuebehind those termites at the digging face. Delayed termites excavated soil laterallyfrom the tunnel wall at a position governed by their position in the queue of termites.By examining excavation under artificially induced conditions of longer and shorterqueues of termites at the tunnel end, we showed that tunnel width increased withincreased queue size and the rate of lateral excavation in a process we termed“digging pressure.”
Keywords Excavation . tunnel . queue . coptotermes formosanus
Introduction
The construction of nests and galleries by groups of termites has been examined in afew termite species (Grasse 1959; Bruinsma 1979; Jones 1979). Grasse (1959)proposed the concept of stigmergy, an indirect mode of communication betweentermites whereby the work product of a builder acts to guide the response ofsubsequent workers, as a means to coordinate the labor of individual termites. In thecontext of termite construction this process relies on the application of variouspheromones to mark sites or pathways and to identify the work product ofconstruction to induce positive feedback loops, where deposition stimulatesadditional deposition. Based on empirical studies, self-organized nest building in
J Insect Behav (2010) 23:189–204DOI 10.1007/s10905-010-9206-z
P. M. Bardunias (*) :N.-Y. SuDepartment of Entomology and Nematology, Ft. Lauderdale Research and Education Center,University of Florida, 3205 College Ave., Ft. Lauderdale, FL 33314, USAe-mail: [email protected]

termites has been extensively modeled (Courtois and Heymans 1991; Deneuborgand Franks 1995, 1997; Bonabeau et al. 1998a; O’Toole et al. 1999, 2003; Ladeleyand Bullock 2005). Excavation of tunnels (Sudd 1975; Buhl et al. 2005) and nests(Kramer 2005) has been modeled in ants based on a system of pheromone marking.In each case these models failed to produce elements of natural tunnels or requiredadditional information in the form of a simulated tactile sense or more pheromonetypes and added complexity to the marking system.
The overall morphology of termite tunnel complexes has been quantified(Reinhard et al. 1997; Pitts-Singer and Forschler 2000; Campora and Grace 2001;Su and Puche 2003; Su et al. 2004; Bardunias and Su 2005), but very little attentionhas been paid to the digging behavior of individuals. If we abandon pheromonelabeling as the guiding force in tunnel excavation, then an examination of thebehaviors of individuals and the manner in which their efforts are coordinated isneeded to establish an alternate means of focusing labor. Bonabeau et al. (1998a)touched upon the notion of an asymmetrical flux of individuals being directionally“convected” to provide a template for construction. Bardunias and Su (2009a),suggested that the location at which termites excavate or deposit is not governed bystigmergic communication from previous excavators, but instead results from aninteraction of the heading of termites moving through tunnels and the geometry ofthe tunnel wall. Bardunias and Su (2009b) showed that termite tunneling is activelyoriented away from a point of origin through dead reckoning and that the urge tomove away from the tunnel mouth led to the formation of queues of termites at thedistal end of the tunnel awaiting access to the digging face (the area of soilaccessible to the foremost excavator at the distal end of the tunnel). The bidirectionalflow of termites through tunnels, the movement of prospective excavators towardsthe tip and termites loaded with soil back toward the mouth, can serve to focus laborat the tunnel ends where excavators must halt or turn around (Bardunias and Su2009a). Increasing the number of termites at any point in the tunnel will change thetunnel environment, with an increase in worker-worker interactions that limitmovement options and access to some digging sites, while focusing labor on others.
A peculiar “halting” behavior has been seen in laborers of Nasutitermes costalis(Holmgren) when termites were unable to complete tasks because of trafficcongestion (Jones 1979). These termites stopped for a period of time and took noaction. Recent models have taken into account halting times, as well as factors suchas crowding and the periodicity of construction (O’Toole et al. 1999, 2003). Iftermites in queue at the distal end of a tunnel altered their behavior not by halting,but by switching tasks from attempting to move forward towards the end of thetunnel and the digging face to excavating from the tunnel wall at their currentlocation (lateral excavation), then we hypothesize that crowding and trafficcongestion may lead to tunnel widening.
In this study we observed the behavior of Formosan subterranean termites,Coptotermes formosanus Shiraki, in order to determine the location of excavationsites within tunnels as a whole and within the area of the tunnels where queueformation occurred. By creating two distinct sizes of excavator queue in tunnels, wealso examined how queue size affects the manner in which tunnels widen as theyelongate and the effect of increased queue size on the behavior of individualexcavators.
190 J Insect Behav (2010) 23:189–204

Methods
Experimental Set Up
Termites were collected from a C. formosanus colony as described by Su andScheffrahn (1986) in the summer of 2006. They were stored at 27±2°C in plasticboxes with thin, moist wood chips for no more than 30 d prior to experimentation.
The experimental arenas are similar to those used by Lee et al. (2008). Theyconsisted of three layers of clear acryl, top and bottom layers (13×13 cm×5 mm),and a 2 mm thick spacer in the middle to create a circular space (10 cm in diameter)in between (Fig. 1a). The enclosed space of the arena was filled with sand (150–500 μm sieves, Play Sand Bonsal, Miami, FL) moistened with deionized water ≈ 7%by sand weight (Fig. 1b). At the outer edge of the top layer, a 1.5 cm diameter holewas drilled through it and the middle spacer. A lidded cylinder (3 cm×2 cm) wasplaced over this hole to act as a termite release chamber (Fig. 1c). The chamber wasprovisioned with small slivers of pine wood and connected to the sand of the arenavia a 1 cm×4 mm pre-formed tunnel formed by a cut-out in the inner spacer(Fig. 1d). The arenas were placed horizontally on a raised platform and tunnelingactivity was recorded at room temperature (26°C) with a video camcorder (SonyGR-D22U, Tokyo, Japan) mounted below the arena.
Observations on Tunneling Behavior
In 5 separate trials, we introduced groups of 130 termites (117 workers and 13soldiers) into arenas and allowed them to excavate a single tunnel. In each trial, werecorded for 4 h from the first excavation event. We observed individuals withintunnels and at the tunnel tip in order to develop a general description of the
Fig. 1 Two-dimensional arenadesign. Arenas consist of twoclear 5 mm thick acryl plates and a2 mm thick central spacer betweenthem a, defining an openedcentral space filled with sand b.A cylindrical introductory cham-ber c connected to the sand-filledportion of the arena through a1 cm×4 mm channel d.
J Insect Behav (2010) 23:189–204 191191

behaviors involved in the excavation process. Additionally, we sampled 5 termitesfrom each of the 5 recordings of tunnels, 25 in total, and observed their behavior atthe tunnel tip in order to determine the number of times that they swept theirantennae across the tunnel wall at the digging face (the area of tunnel wall at the tipwhere a termite can take soil when at the very end of the tunnel) prior to excavation.
Location of Excavation Within Tunnels
From the 4 h recordings, one for each of the 5 groups of 130 termites (117 workersand 13 soldiers), we recorded whether excavation occurred either in the tunnel prioran excavator reaching the queue at distal end or within the length of tunnel occupiedby the queue. In order to provide a sufficient length of tunnel to clearly differentiatebetween these two areas within the tunnel, observations were made for 5 min fromthe point at which the tunnels had extended a distance of 1.5 cm beyond the initial1 cm length of pre-formed tunnel provided for the termites by the experimental setup (2.5 cm total length including the 1 cm preformed tunnel provided in the initialset up). Excavation events where sand was taken from the tunnel wall (excavationoccurring at the digging face at the tunnel tip was excluded) were recorded fortermites in queue at the distal end and in the rest of the tunnel prior to termitesreaching the queue.
Location of Excavation by Individuals in Queue
For a half hour after the tunnel had progressed 1 cm from the start of excavation(2 cm total) for each of five recordings excavated by groups of 130 termites (117workers and 13 soldiers), we recorded the distances (Fig. 2) of all digging eventsconducted by termites in queue behind an excavator occupying the digging face atthe tunnel tip that took place within 1 cm from the digging face, measured to thenearest 1 mm. The number of excavation events occurring in each of these ten
Fig. 2 A typical queue at the distal end of a tunnel. Each termite in queue is numbered successively (1–5),and arrows indicate the sites of excavation. Termite (1) is excavating sideways, but by definition, since itis the foremost excavator, this is part of the digging face at the distal end of the tunnel, indicated by theextent of the dotted line. Termites 2–4 are engaged in lateral excavation, excavation from the tunnel wallby those in queue. The white dashed lines indicate the tunnel width at successive distances from thedigging face: 0.05, 0.5, and 1.0 cm.
192 J Insect Behav (2010) 23:189–204

intervals of 1 mm width was compared by ANOVA and then grouped by a Dunn’spost-hoc test at α=0.05 (SAS Institute 1987). In addition, we compared the numberof excavation events at the digging face to the number of lateral excavation eventswith χ2 analysis at α=0.05 (SAS Institute 1987).
The average length of termite excavators was determined by measuring thedistance from the tip of the labrum to the end of the abdomen for 10 termites fromeach of the three colonies used in this study.
The Number of Termites and Queue Size
Two group sizes were employed to create two distinct levels of queue size in tunnels.We defined as “large queue” the queues that resulted from the recordings of 130termites (117 workers and 13 soldiers) described above. In order to create a “smallqueue” a second set of five trials with arenas that included 30 termites (27 workersand 3 soldiers) was conducted. Excavation was recorded for 4 h in each trialfollowing an initial tunnel distance of 1 cm beyond the 1 cm tunnel joining the arenaand introduction chamber (2 cm total). To verify that we created two levels of queuesize in the tunnels by altering the number of termites in the arenas, we recorded thelength of queues for both large and small treatments. Queue length was determinedby counting the number of termites in mutual contact (Fig. 2, 1–5) at the distal endof the tunnel at 5 min intervals over 60 min. This resulted in 12 counts for each offive replicates of the large queue treatment and five of the small queue. The meannumber of individuals at the distal end of the tunnel, including both the foremostexcavator and those in queue behind, for both treatments was compared by ANOVAand then grouped by a Dunn’s post-hoc test at α=0.05 (SAS Institute 1987).
Queue Size and Tunnel Profile Shape
The profile of burgeoning tunnels will show whether tunnel width is established byexcavation at the digging face, leading to roughly parallel tunnel walls, or ifexcavation from the tunnel wall behind the digging face by those in queue governswidth, imparting a wedge-shaped advance. In order to characterize the shape of theelongating tunnel near its distal end, we compared the width of the tunnels recordedfrom the large and small queue treatments above every 10 min for 60 min. Widthwas measured at distances suggested by preliminary observations of queue length:0.05 cm (the digging face), and 0.5 cm (the tunnel wall in the area generallyoccupied by the queue) and 1 cm (the tunnel wall generally at the end or behind thequeue) from the digging face to the nearest 0.25 mm (Fig. 2). Because ofheteroscedasticity and departures from normality in these data, we used the Kruskal-Wallace test (χ2 approximation) to detect significant difference among means andthen grouped by Dunn's post-hoc test at α=0.05 (SAS Institute 1987).
Queue Size and Tunnel Width
To ascertain if larger-queue tunnels were significantly wider than smaller-queuetunnels, the mean tunnel widths at 0.05 and 1.0 cm were compared between the 5small queue and 5 large queue tunnels using a t-test at α=0.05 (SAS Institute 1987).
J Insect Behav (2010) 23:189–204 193193

The 0.05 distance represents the width set by excavation at the digging face, whilethe 1.0 cm distance was chosen because it was the most distant from the tip andsubjected to possible lateral excavation for the longest period.
Queue Size and Tunnel Elongation
For each 10 min interval for both small and large queue sizes, we also measured thechange in tunnel length from origin to the distal end during the interval. Thiselongation distance over time provided a rate of tunnel extension. We compared therate of tunnel elongation between small and large queue size treatments with χ2
analysis at α=0.05 (SAS Institute 1987).
Queue Size and the Behavior of Individual Excavators
From the point where the tunnels progressed to a distance of 1.5 cm from the originin each of the 5 large queue and 5 small queue replicates, 15 individual excavatorswere followed from their initial contact with the queue to their leaving the distal endof the tunnel, and their actions recorded. The incidence of lateral excavation wascompared for large and small queue via χ2 analysis at α=0.05 (SAS Institute 1987).
We recorded the time spent delayed in the queue against the tunnel wall priorto lateral excavation, delayed in queue against the tunnel wall prior to excavationat the digging face, and in the queue bound on both flanks by other termites withno access to the walls. The time of delay in each case was recorded from thetime a termite initially came into physical contact with the members of the queueat the tunnel tip until it left the tunnel tip. The duration of a soil excavationevent, the time between the initial excavation at a digging site and leaving with aload of soil, both at the digging face and during lateral excavation from thetunnel wall, was recorded. These were compared between the two different queuesizes via a t-test at α=0.05.
For both small and large queue sizes, the actions of the 75 termites were tallied asthe percentage of individuals that alternately left without remaining in queue andexcavating, entered the queue with access to the tunnel wall or entered the queuebound between other termites. Subsequently, of those in queue, the proportion thatexcavated either laterally or at the digging face.
Results
Observations on Tunneling Behavior
Termites moved from the tunnel origin to the distal end, antennating the tunnel wallsas they passed (Fig. 3a), and may have excavated along the tunnel walls (Fig. 3b, a).Termites are slightly wider than they are high, so when they were packed tightly orpassed in a narrow tunnel, they tended to oriented themselves back to back tominimize the cross-sectional area of the pair (Fig. 3b). Upon arriving at the distalend of the tunnel (Fig. 3c, 1), if the digging face was unoccupied, they excavatedsoil (Fig. 3c, a). When the digging face was occupied, termites formed a queue
194 J Insect Behav (2010) 23:189–204

behind the foremost excavator (Fig. 3c, 2). While in queue, they pushed against thetermites in front of them and jostled those beside them in an attempt to moveforward. They often performed horizontal oscillatory movements (Stuart 1967),thought to have a communicative function, and in rare cases they gripped theabdomen or rear leg of termites ahead of them and pulled them out of line.Individuals in queue could also turn to the tunnel wall at their current position andexcavate laterally (Fig. 3c, b).
Upon arriving at the distal end of the tunnel, if the digging face was unoccupied,they antennated the soil an average of 2.48±1.0 (mean±SD, n=25) times beforeremoving soil. Each antennation event consisted of sweeping the antennae back andforth (Fig. 4, t-1, t-2) across the soil surface prior to choosing an excavation sitealong the tunnel surface at the digging face (Fig. 4, t-3).
After excavating a parcel of soil, termites walked backwards or turned around inthe tunnel and moved back towards the tunnel origin. They either unloaded beyondthe origin of the tunnel or pressed the parcels of soil against the tunnel wall,spreading the soil in a manner described by Bardunias and Su (2009a) (Fig. 3d, a).
Location of Excavation Within Tunnels
Individuals stopped and excavated laterally prior to reaching the queue at the tunneltip only rarely (6.7%, n=113 excavation events). The majority of excavation that
Fig. 3 a Termites antennate the walls as they travel down a tunnel. b A back to back posture is assumedas they pass in a narrow tunnel. A feature, a, in this case a bulge in the tunnel wall that blocks passage,may stimulate excavation. c Termites at the tunnel tip. Termite 1 excavates from the digging face, a, whiletermite 2 excavates laterally, b. d A termite drops a load of soil by pressing it against the tunnel wall, a.
J Insect Behav (2010) 23:189–204 195195

was not directed to the digging face at the distal end of the tunnel was done by thoseindividual in queue behind the foremost excavator.
Location of Excavation by Individuals in Queue
The distribution of excavation sites within 1 cm from the digging face for termites inqueue behind an excavator occupying the digging face at the tip is shown in Fig. 5a,while an example of termites in queue is presented in Fig. 5b for comparison. Froma minimum mean value at 1 mm, the occurrence of lateral excavation rose to asignificant peak at a distance of 3 mm, and then tapered to a second low at 7 mm. Asecond, smaller peak occurred at a distance of 9 mm from the tunnel tip. The averagenumber of excavation events was evenly divided between those that occurred at thetunnel tip, 540±15.378, and those that occurred along the tunnel wall within 1.0 cmfrom the tip, 507±7.368 (χ2=1.040, df=1, P=0.3078). There was no significantdifference between colonies in the length of the 10 termites taken from each, F (2,27)=2.47, p=0.103), so the measures were pooled to provide a mean of 4.42±0.18 mm (mean±SD).
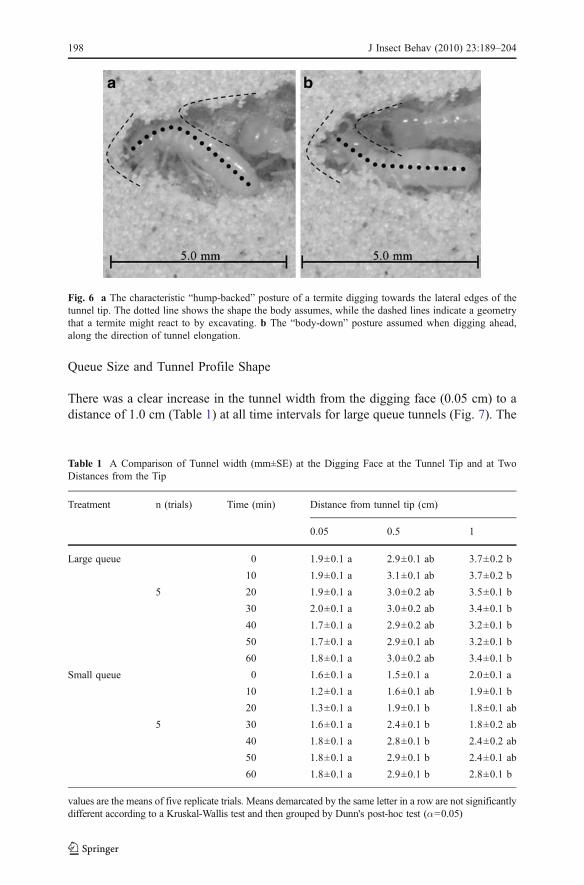
There was some variability in the postures that the foremost termite assumed, orwas pushed into while excavating: hump-backed, body-down, or unbent (Fig. 6).The location at the digging face from which the ≈ 4-5 mm long termites took soillimited the posture they could assume. When digging towards one side of thedigging face, they tended to assume a hump-backed stance as they lowered theirmouth parts to the soil, their thorax humped up and their abdomen down (Fig. 6a).The forward progress of the second termite in queue behind the foremost excavatorin this position was limited to about 3 mm from the digging face. When termites atthe digging face excavated directly ahead, along the direction of tunnel propagation,they were able to assume a posture that allowed the second termite in queue to movecloser to the digging face ( Fig. 6b), but no closer than the ≈ 1 mm width of thetermites head. If a termite completely blocked the tunnel with its body, access waslimited to a body-length from the digging face. There were zones at 0–2 mm and 6-
Fig. 4 a Antennation of the digging face at the tunnel tip in two successive snap shots (t-1 and t-2) andsubsequent excavation (t-3). The moniliform antennae deform extensively to match the relief of the wallprofiles, indicated by the dotted lines.
196 J Insect Behav (2010) 23:189–204
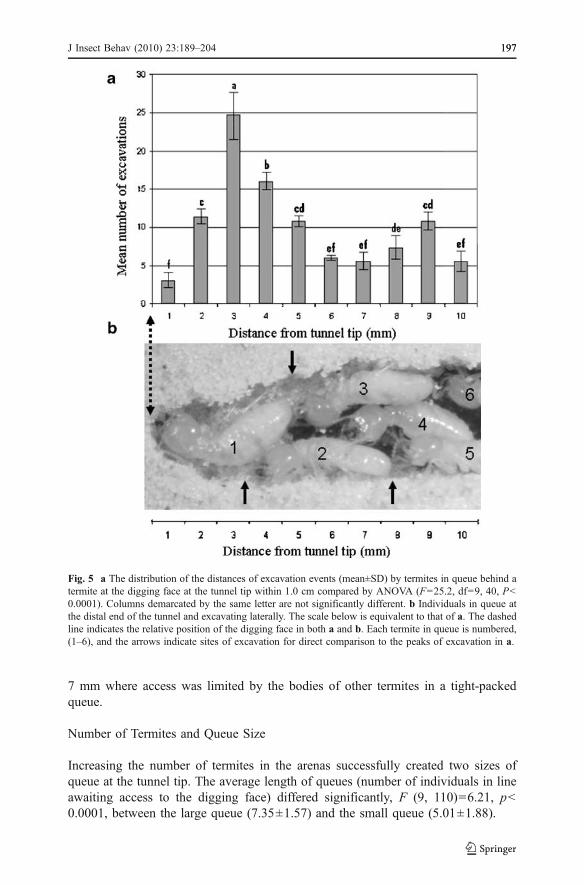
7 mm where access was limited by the bodies of other termites in a tight-packedqueue.
Number of Termites and Queue Size
Increasing the number of termites in the arenas successfully created two sizes ofqueue at the tunnel tip. The average length of queues (number of individuals in lineawaiting access to the digging face) differed significantly, F (9, 110)=6.21, p<0.0001, between the large queue (7.35±1.57) and the small queue (5.01±1.88).
Fig. 5 a The distribution of the distances of excavation events (mean±SD) by termites in queue behind atermite at the digging face at the tunnel tip within 1.0 cm compared by ANOVA (F=25.2, df=9, 40, P<0.0001). Columns demarcated by the same letter are not significantly different. b Individuals in queue atthe distal end of the tunnel and excavating laterally. The scale below is equivalent to that of a. The dashedline indicates the relative position of the digging face in both a and b. Each termite in queue is numbered,(1–6), and the arrows indicate sites of excavation for direct comparison to the peaks of excavation in a.
J Insect Behav (2010) 23:189–204 197197
Queue Size and Tunnel Profile Shape
There was a clear increase in the tunnel width from the digging face (0.05 cm) to adistance of 1.0 cm (Table 1) at all time intervals for large queue tunnels (Fig. 7). The
Fig. 6 a The characteristic “hump-backed” posture of a termite digging towards the lateral edges of thetunnel tip. The dotted line shows the shape the body assumes, while the dashed lines indicate a geometrythat a termite might react to by excavating. b The “body-down” posture assumed when digging ahead,along the direction of tunnel elongation.
Table 1 A Comparison of Tunnel width (mm±SE) at the Digging Face at the Tunnel Tip and at TwoDistances from the Tip
Treatment n (trials) Time (min) Distance from tunnel tip (cm)
0.05 0.5 1
Large queue 0 1.9±0.1 a 2.9±0.1 ab 3.7±0.2 b
10 1.9±0.1 a 3.1±0.1 ab 3.7±0.2 b
5 20 1.9±0.1 a 3.0±0.2 ab 3.5±0.1 b
30 2.0±0.1 a 3.0±0.2 ab 3.4±0.1 b
40 1.7±0.1 a 2.9±0.2 ab 3.2±0.1 b
50 1.7±0.1 a 2.9±0.1 ab 3.2±0.1 b
60 1.8±0.1 a 3.0±0.2 ab 3.4±0.1 b
Small queue 0 1.6±0.1 a 1.5±0.1 a 2.0±0.1 a
10 1.2±0.1 a 1.6±0.1 ab 1.9±0.1 b
20 1.3±0.1 a 1.9±0.1 b 1.8±0.1 ab
5 30 1.6±0.1 a 2.4±0.1 b 1.8±0.2 ab
40 1.8±0.1 a 2.8±0.1 b 2.4±0.2 ab
50 1.8±0.1 a 2.9±0.1 b 2.4±0.1 ab
60 1.8±0.1 a 2.9±0.1 b 2.8±0.1 b
values are the means of five replicate trials. Means demarcated by the same letter in a row are not significantlydifferent according to a Kruskal-Wallis test and then grouped by Dunn's post-hoc test (α=0.05)
198 J Insect Behav (2010) 23:189–204

data at 0.5 cm shows the trend towards widening, but the values were intermediateand not significantly different from both those at the digging face and at 1.0 cm.
The small queue tunnel data shows a more complex pattern that changed overtime (Table 1). At time 0, when the excavated tunnel was 1.0 cm in length, nosignificant difference in width at the various distances was recorded. As the tunnelselongated, there was a trend toward widening at the 0.5 cm distance, while at 1.0 cmthe tunnel was narrower. The apparent narrowing at 1.0 cm in the small queue datacan be explained by the elongation rate of ≈ 2 mm in each 10 s interval. The site onthe tunnel wall where the 1.0 cm value was recorded is roughly the same place the0.5 cm value was measured 2 time intervals earlier. The tunnel did not narrowbetween the two distances. What occurred is that the whole tunnel gradually becamewider, with the time delay explaining the difference in widths.
Queue Size and Tunnel Width
The mean widths of tunnels at 1.0 cm were 2.15±0.439 cm for small queue and 3.4±0.296 cm for large queue. These were significantly different (t=18.731, df=128, n=65,P<0.0001), and, more importantly, the average difference in width was enough toallow an additional termite to pack into the tunnel. The mean width at the digging face(0.05 cm), 1.58±0.280 cm for small queue and 1.8±0.201 cm for large queue, wasalso significantly different (t=5.208, df=128, n=65, P<0.0001), but the difference inthe means may be irrelevant to the mechanics of tunnel widening because thedifference is too small to allow an additional termite access to the digging face.
Queue Size and Tunnel Elongation
There was no significant difference (P=0.12, n=35) in the rate of tunnel elongation(mm/hr) between larger (15.3±1.4) and smaller queue (11.8±0.3) treatments.
Queue Size and the Behavior of Individual Excavators
The incidence of lateral excavation was significantly different between the large andsmall queue tunnels (χ2=4.446, df=1, P=0.0350). The mean time that excavatorsspent delayed in queue, with or without access to the walls, prior to excavation andthe duration of excavation for each queue size is shown in Table 2. There was nosignificant difference between large and small queue treatments for the times spentdelayed in queue prior to excavation or the duration of excavation. Figure 8 shows a
Fig. 7 The propagation of a tunnel at three time intervals, t-1, t-2, and t-3, traced from a large queuetunnel, showing the characteristic wedge-shaped advance of the tunnel due to lateral excavation behind thedigging face at the tunnel tip.
J Insect Behav (2010) 23:189–204 199199

flow chart for the percentage of termites in each possible state from their arrival atthe tunnel tip to excavation for termites in the large and small queue tunnels.
Discussion
Models of self-organized construction and excavation have previously relied onpheromones as a medium for information transfer between individuals to coordinatethe labor of agents with extremely simplified behavioral repertoires. Ourinvestigation of termite excavation breaks with previous theories of constructionand excavation. Rather than begin with the assumption that tunneling is governed bythe pheromone gradients and scent labels, we assumed that pheromones play nodirect role in the removal of soil, relegating the role of pheromones to trail markingand recruiting the labor force to tunnels. Here we present a scheme (Fig. 9) wherein
Table 2 A Comparison of the Delay in Queue Prior to Excavation and the Duration of Excavation EventsVia a t-test at α=0.05
Delay type n Time (sec±SD) t-value df P
Small queue n Large queue
Queue delay, lateral excavation 75 15.06±5.84 75 18.19±12.20 1.6345 62 0.1072a
Queue delay, digging face 75 18.76±7.62 75 22.29±12.97 1.3417 59 0.1848a
Queue delay bound 75 11.13±5.44 75 8.86±4.29 1.0967 21 0.2852a
Excavation time lateral 75 11.21±5.96 75 10.82±7.43 0.2226 63 0.8246a
Excavation time digging face 75 11.21±4.74 75 14.11±9.31 1.8363 83 0.0699a
aNS Not significantly different at α=0.05
In queue bound between other termites
Excavation at digging face
In queue with access to tunnel wall
Lateral excavation
18.6%
57.2%
42.8%
78.6%
40.7%
59.3%
Termites arriving at the tunnel tip
No excavation2.6%
(10.6%)
(62.5%)
(37.5%)
(88.0%)
(54.5%)
(44.5%)
(1.3%)
Fig. 8 The actions of individual termite excavators upon entering the queue at the tunnel tip, representedas the percentage of the total number of termites engaging in each action. Values in parentheses arederived from the small queue tunnels, the remaining values are from the large queue tunnels.
200 J Insect Behav (2010) 23:189–204
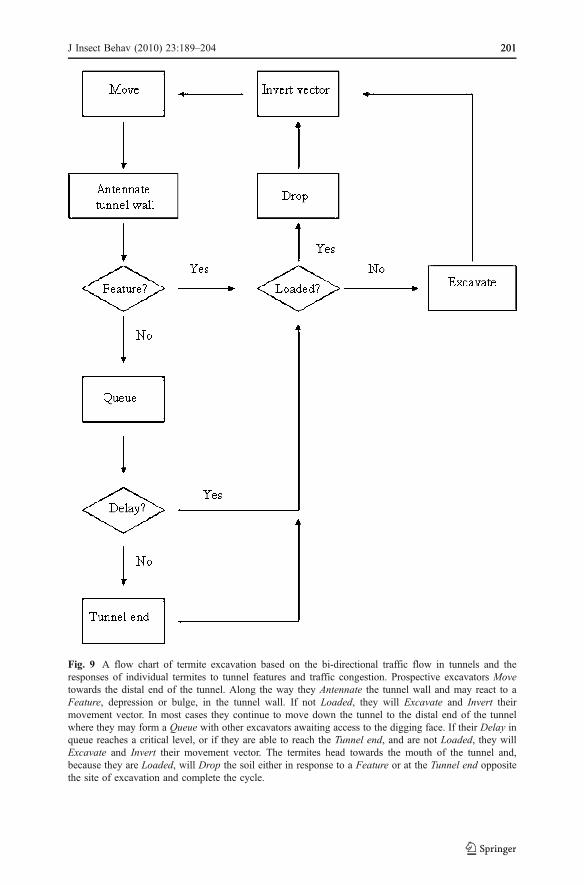
Fig. 9 A flow chart of termite excavation based on the bi-directional traffic flow in tunnels and theresponses of individual termites to tunnel features and traffic congestion. Prospective excavators Movetowards the distal end of the tunnel. Along the way they Antennate the tunnel wall and may react to aFeature, depression or bulge, in the tunnel wall. If not Loaded, they will Excavate and Invert theirmovement vector. In most cases they continue to move down the tunnel to the distal end of the tunnelwhere they may form a Queue with other excavators awaiting access to the digging face. If their Delay inqueue reaches a critical level, or if they are able to reach the Tunnel end, and are not Loaded, they willExcavate and Invert their movement vector. The termites head towards the mouth of the tunnel and,because they are Loaded, will Drop the soil either in response to a Feature or at the Tunnel end oppositethe site of excavation and complete the cycle.
J Insect Behav (2010) 23:189–204 201201

tactile interactions between individual termites and the tunnel walls are sufficient toorganize labor if we also endow individuals with the orientation ability suggested byBardunias and Su (2009b).
Termites repeatedly sweep the digging face with their antennae, pressing theentire length of each into the relief of the soil. This may indicate a tactile function,because chemoreceptors are present only on the distal segments of the antennae,while mechanoreceptors are present on all segments (Tarumingkeng et al. 1976). Themoniliform shape of termite antennae allows them to deform to match tunnel wallrelief, and the ability of termites to perceive the relief of tunnel walls could beanalogous to the advanced tactile system of star-nosed moles (Catania 1999).
Tunnel width was established not at the digging face, but by excavation along thewall further back in the tunnel. The distribution of excavation events was heavilybiased towards the digging face at the distal end of the tunnel and the zone of tunnelwall occupied by the queue, implicating the lateral excavation of those individuals inqueue as the determinant of tunnel width. The small size of the difference in tunnelwidths at the digging face at the tunnel end between large and small queue tunnelsand the similar rates of tunnel elongation suggest that most of the excavation of theadditional excavators in the larger queues was lateral excavation by those in queue.The time an individual spent delayed in queue prior to excavation did not differ withincreased queue size, nor did the duration of excavation. This uniformity of behavioramong termites in both treatments points to a simple additive effect (Bonabeau et al.1998b) resulting from the larger queue sizes as the source for the increase in tunnelwidth and not a group effect (Theraulaz and Bonabeau 1999). As queues becamelarger, the longer queue lengths put more termites in contact with the tunnel wall andmade the wait to reach the digging face longer, resulting in a higher proportion ofindividuals that excavated laterally.
We have termed this effect “digging pressure” because the manner in which thetunnel expands laterally in response to an increase in the number of excavators isanalogous to the effect of increasing pressure in a closed system. The time spent bythe foremost excavators at the digging face to choose a site and take on a load of soildefined the minimum delay of those awaiting access. The length of tunnel and thewalking speed of termites produced the round trip time for individual excavators.Longer trip times are expected to reduce pressure because termites will spend less oftheir total excavation and travel time in queue. Nowhere else will the diggingpressure be as high as in a stretch of tunnel occupied by a queue, such as the distalend of the tunnel or a zone of traffic congestion.
Digging pressure can produce tunnel features beyond initial width. The formationof queues due to the impeded flow of termites at any point in the tunnel, such asintersections or constrictions, will lead to widening. This will scale the tunnel widthto traffic flow and, coupled with the unloading of soil within the tunnel byexcavators (Bardunias and Su 2009a), will lead to homeostasis in tunnel width.
The choice of lateral excavation sites resulted from interactions betweenindividuals that limited the access of other excavators to specific areas of tunnelrather than from the detection of pheromonal marking. Because the second positionin queue was the most likely to be filled, and individuals at this position haveprobably been waiting the longest and are most likely to excavate laterally along theside wall, there was a peak of excavation between ≈ 2–5 mm from the digging face.
202 J Insect Behav (2010) 23:189–204

Beyond this, the bodies of termite excavators in queue block access to the tunnelwall for their ≈ 4-5 mm lengths, leading to another, smaller peak of excavation at ≈9 mm. Lateral excavation could have resulted from interplay between conflictingforces, as an urge to reach the tunnel tip may have waned with time spent in thequeue and been eclipsed by a rising urge to excavate. They may also have respondedto the tip-like geometry (Fig. 6, dashed line) formed by the tunnel wall on one sideand the body of a fellow excavator on the other as a cue for excavation (Lee et al.2008). Whatever the proximate cause of lateral excavation, it is clear from thedistribution of digging events that it is not caused by a simple response topheromone labeling of sites along the walls.
This termite tunneling system relies on the existence of discrete starting andending points to coordinate labor. As a tunnel widens with increased pressure, thewidth of the digging face at the distal end of the tunnel expands beyond a width thatallows one or two workers simultaneous access. When the distal end of the tunnelbroadens, the location of the digging face of the tunnel may become unfocussed andfragment into multiple discrete digging faces. The formation of multiple, discretedigging faces is the precursor to bifurcation and branching, and the emergence ofbranching and tunnel branch patterns from simple variations in queue size anddirected movement of excavators is the focus of forthcoming research.
Our findings cannot disprove the existence of pheromone markers in the self-organization of excavation in termites, and a pheromone could be invoked as analternate explanation for some features, such as the aggregation of termites at thedistal end of the tunnel, but we have shown that a number of key elements in theexcavation system are consistent with a tactile system. The difficulty in distinguish-ing between the possibilities that excavators are guided to excavation sites by scentand labeled work product or through their own ability to orient movement and tactileinteractions may justify a reexamination of previous work on the mechanics ofexcavation. It remains to be seen if a model with no pheromone labeling of worksites and individuals who actively orient is applicable to construction or toexcavation in other organisms or robotic systems.
Acknowledgements We thank R. Pepin and P. Ban for technical assistance and R. L. Yang for statisticanalysis (University of Florida) as well as G. Seedahmed for review of the manuscript (University ofFlorida). This research was supported by the Florida Agricultural Experiment Station and a grant fromUSDA-ARS under the grant agreement No. 58-6435-2-0023.
References
Bardunias PM, Su N-Y (2005) Comparison of Tunnel Geometry of Subterranean Termites (Isoptera:Rhinotermitidae) in “Two-dimensional” and “Three-dimensional” Arenas. Sociobiology 45:679–685
Bardunias PM, Su N-Y (2009a) Opposing headings of excavating and depositing termites facilitate branchformation in the Formosan subterranean termite. Animal Behav 78:755–759
Bardunias PM, Su N-Y (2009b) Dead reckoning in the tunnel propagation of the formosan subterraneantermite (Isoptera: Rhinotermitidae). Ann Entomol Soc Am 102:158–165
Bonabeau E, Theraulaz G, Deneubourg J-L, Franks NR, Rafelsberger O, Joly J-L, Blanco S (1998a) A modelfor the emergence of pillars, walls and royal chambers in termite nests. Phil Trans R Soc B 353:1561–1576
Bonabeau E, Theraulaz G, Deneubourg J-L (1998b) Latency time and absence of group effect. InsectesSoc 45:191–195
J Insect Behav (2010) 23:189–204 203203

Bruinsma OH (1979) An analysis of building behaviour of the termite Macrotermes subhyalinus(Rhambur). Ph. D. Thesis, Landbouwhogeschool, Wageningen, Netherlands
Buhl J, Deneubourg JL, Grimal A, Theraulaz G (2005) Self-organized digging activity in ant colonies.Behav Ecol Scociobiol 58:9–17
Campora CE, Grace JK (2001) Tunnel orientation and search pattern sequence of theFormosansubterranean termite (Isoptera: Rhinotermitidae). J Econ Ent 94:1193–1199
Catania KC (1999) A nose that looks like a hand and acts like an eye: the unusual mechanosensory systemof the star-nosed mole. J Comp Physiol A 185:376–372
Courtois PJ, Heymans F (1991) A simulation of the construction process of a termite nest. J Theor Biol153:469–475
Deneuborg J-L, Franks NR (1995) Collective control with out explicit coding: the case of communal nestexcavation. J Insect Behav 8:417–432
Deneuborg J-L, Franks NR (1997) Self-organizing nest construction in ants: the behaviour of individualworkers and the properties of the nest’s dynamics. Anim Behav 54:779–796
Grasse P-P (1959) La reconstruction du nid et les coordinations inter-individuelles chez Bellicositermesnataliensis et Cubitermes sp. La theore de la stigmergie: essai d’interpretation du comportment destermites constructeurs. Insectes Soc 6:41–81
Jones RJ (1979) Expansion of the nest of Nasutitermes coastalis. Insectes Soc 26:322–342Kramer RSS (2005) Three-dimensional ant nest excavation using stigmergic rules. The University of
Sussex, Brighton, M. S. thesisLadely D, Bullock S (2005) The role of logistic constraints in termite construction of chambers and
tunnels. J Theor Biol 234:551–564Lee S-H, Bardunias PM, Su N-Y, Yang R-L (2008) Behavioral response of termites to tunnel surface
irregularity. Behav Process 78:397–400O’Toole DV, Robinson PA, Myerscough MR (1999) Self organized criticality in termite architecture: a
role of crowding in ensuring ordered nest expansion. J Theor Biol 198:305–327O’Toole DV, Robinson PA, Myerscough MR (2003) Self organized criticality and emergent oscillations in
models of termite architecture with crowding. J Theor Biol 221:15–27Pitts-Singer TL, Forschler BT (2000) Influence of guidelines and passageways on tunnelling behavior of
Reticulitermes flavipes (Kollar) and R. virginicus (Banks) J. Insect Behav 13:273–290Reinhard J, Hertel H, Kaib M (1997) Systematic Search for food in the subterraneantermite Reticulitermes
santonensis De Feytaud (Isoptera, Rhinotermitidae). Insectes Soc 44:147–158SAS Institute (1987) SAS/STAT guide for personal computers, version 6 ed. SAS Institute, CaryStuart AM (1967) Alarm, defense and construction behaviour relationships in termites (Isoptera). Science
156:1123–1125Su N-Y, Scheffrahn RH (1986) A method to access, trap, and monitor field populations of the Formosan
subterranean termite (Isoptera: Rhinotermitidae) in the urban environment. Sociobiology 12:299–304Su N-Y, Puche H (2003) Tunneling activity of subterranean termites (Isoptera: Rhinotermitidae) in sand
with moisture gradients. J Econ Ent 96:88–93Su N-Y, Stith BM, Puche H, Bardunias PM (2004) Characterization of tunneling geometry of subterranean
termites (Isoptera: Rhinotermitidae) by computer simulation. Sociobiology 44:471–483Sudd JH (1975) A model of digging behaviour and tunnel production in ants. Insectes Soc 22:225–235Tarumingkeng RC, Coppel HC, Matsumura F (1976) Morphology and ultrastructure of the antennal
chemoreceptors and mechanoreceptors of worker Coptotermes formosanus Shiraki. Cell Tis Res173:173–178
Theraulaz G, Bonabeau E (1999) A brief history of stigmergy. Artificial Life 5:97–116
204 J Insect Behav (2010) 23:189–204