
Purification and primary structure of a mannose/glucose-binding lectin from Parkia biglobosa Jacq....
9
Purification and primary structure of a mannose/glucose-binding lectin from Parkia biglobosa Jacq. seeds with antinociceptive and anti-inflammatory properties Helton C. Silva a , Alfa U. Bari a , Bruno Anderson M. Rocha a , Kyria S. Nascimento a , Edson L. Ponte b , Alana F. Pires b , Plínio Delatorre c , Edson H. Teixeira d , Henri Debray e , Ana Maria S. Assreuy b , Celso S. Nagano f and Benildo S. Cavada a * Parkia biglobosa (subfamily Mimosoideae), a typical tree from African savannas, possess a seed lectin that was purified by combination of ammonium sulfate precipitation and affinity chromatography on a Sephadex G-100 column. The P. biglobosa lectin (PBL) strongly agglutinated rabbit erythrocytes, an effect that was inhibited by D-mannose and D-glucose-derived sugars, especially α-methyl-D-mannopyranoside and N-acetyl-D-glucosamine. The hemagglutinating activity of PBL was maintained after incubation at a wide range of temperature and pH and also was independent of divalent cations. By sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis, PBL exhibited an electrophoretic profile consisting of a single band with apparent molecular mass of 45 kDa. An analysis using electrospray ionization–mass spectrometry indicated that purified lectin possesses a molecular average mass of 47 562 ± 4 Da, and the analysis by gel filtration showed that PBL is a dimer in solution. The complete amino acid sequence of PBL, as determined using tandem mass spectrometry, consists of 443 amino acid residues. PBL is composed of a single non-glycosylated polypeptide chain of three tandemly arranged jacalin-related domains. Sequence heterogeneity was found in six positions, indicating that the PBL preparations contain highly homologous isolectins. PBL showed important antinociceptive activity associated to the inhibition of inflammatory process. Copyright © 2013 John Wiley & Sons, Ltd. Supporting information may be found in the online version of this paper. Keywords: lectin; Parkia biglobosa; subfamily Mimosoideae INTRODUCTION The interaction of carbohydrates with proteins is a crucial step in many biological processes including cell–cell recognition, adhesion, metastasis, bacterial and viral infection, and inflamma- tion (Gabius and Gabius, 1997; Taylor and Drickamer, 2003). Lectins are defined as structurally heterogeneous proteins possessing at least one non-catalytic domain by which they reversibly bind to specific monosaccharides or oligosaccharides (Van Damme et al., 1998). These proteins are ubiquitous in nature and are found in all types of living organisms. In fact, lectins have been used for decades as model systems for the study of protein–sugar interactions because of their wide range of specificities (Loris et al., 1998; Sharon and Lis, 2004). Despite of the lectins wide distribution, the best-studied are those obtained from plants, especially from the family Leguminosae (van Damme et al., 1998), subfamily Papilinoidae. However, few lectins from the Mimosoideae species have been reported (Suvachittanonts and Peutpaiboon, 1992; Utarabhand and Akkayanont, 1995; Cavada et al., 1997, 2000, 2006; Guzman-Partida et al., 2004; Kaur et al., 2005; Santi-Gadelha et al., 2008). In the Mimosoideae subfamily, the glucose/ mannose-specific lectin isolated from Parkia platycephala seeds * Correspondence to: B. S. Cavada, Department of Biochemistry and Molecular Biology, Federal University of Ceará, Mister Hull s/n Bloco 907, Lab. 1075, PO Box 6043, 60440-970, Fortaleza, CE, Brazil. E-mail: [email protected] a H. C. Silva, A. U. Bari, B. A. M. Rocha, K. S. Nascimento, B. S. Cavada BioMol-Group, Department of Biochemistry and Molecular Biology, Federal University of Ceará, PO Box 6043, 60440-970, Fortaleza, CE, Brazil b E. L. Ponte, A. F. Pires, A. M. S. Assreuy BioMol-Group, Institute of Biomedical Sciences, State University of Ceará, Fortaleza, CE, Brazil c P. Delatorre BioMol-Group, Department of Molecular Biology, Federal University of Paraíba, João Pessoa, PB, Brazil d E. H. Teixeira BioMol-Group, Department of Pathology and Legal Medicine, Faculty of Medicine, Federal University of Ceará, Fortaleza, CE, Brazil e H. Debray University of Science and Technology of Lille, Lille, France f C. S. Nagano BioMol-Group, Department of Fishing Engineering, Federal University of Ceará, Fortaleza, CE, Brazil Research Article Received: 3 May 2013, Revised: 7 June 2013, Accepted: 7 June 2013, Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/jmr.2289 J. Mol. Recognit. 2013; 26: 470–478 Copyright © 2013 John Wiley & Sons, Ltd. 470
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Purification and primary structure of a mannose/glucose-binding lectin from Parkia biglobosa Jacq....

Purification and primary structure of amannose/glucose-binding lectin from Parkiabiglobosa Jacq. seeds with antinociceptiveand anti-inflammatory propertiesHelton C. Silvaa, Alfa U. Baria, Bruno Anderson M. Rochaa,Kyria S. Nascimentoa, Edson L. Ponteb, Alana F. Piresb, Plínio Delatorrec,Edson H. Teixeirad, Henri Debraye, Ana Maria S. Assreuyb,Celso S. Naganof and Benildo S. Cavadaa*
Parkia biglobosa (subfamily Mimosoideae), a typical tree from African savannas, possess a seed lectin that was purifiedby combination of ammonium sulfate precipitation and affinity chromatography on a Sephadex G-100 column. TheP.biglobosa lectin (PBL) strongly agglutinated rabbit erythrocytes, an effect that was inhibited by D-mannose andD-glucose-derived sugars, especially α-methyl-D-mannopyranoside and N-acetyl-D-glucosamine. The hemagglutinatingactivity of PBL was maintained after incubation at a wide range of temperature and pH and also was independent ofdivalent cations. By sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis, PBL exhibited an electrophoreticprofile consisting of a single bandwith apparentmolecularmass of 45kDa. An analysis using electrospray ionization–massspectrometry indicated that purified lectin possesses a molecular average mass of 47562±4Da, and the analysis by gelfiltration showed that PBL is a dimer in solution. The complete amino acid sequence of PBL, as determined using tandemmass spectrometry, consists of 443 amino acid residues. PBL is composed of a single non-glycosylated polypeptide chain ofthree tandemly arranged jacalin-related domains. Sequence heterogeneity was found in six positions, indicating that thePBL preparations contain highly homologous isolectins. PBL showed important antinociceptive activity associated to theinhibition of inflammatory process. Copyright © 2013 John Wiley & Sons, Ltd.Supporting information may be found in the online version of this paper.
Keywords: lectin; Parkia biglobosa; subfamily Mimosoideae
INTRODUCTION
The interaction of carbohydrates with proteins is a crucial step inmany biological processes including cell–cell recognition,adhesion, metastasis, bacterial and viral infection, and inflamma-tion (Gabius and Gabius, 1997; Taylor and Drickamer, 2003).
Lectins are defined as structurally heterogeneous proteinspossessing at least one non-catalytic domain by which theyreversibly bind to specific monosaccharides or oligosaccharides(Van Damme et al., 1998). These proteins are ubiquitous innature and are found in all types of living organisms. In fact,lectins have been used for decades as model systems for thestudy of protein–sugar interactions because of their wide rangeof specificities (Loris et al., 1998; Sharon and Lis, 2004).
Despite of the lectins wide distribution, the best-studied arethose obtained from plants, especially from the familyLeguminosae (van Damme et al., 1998), subfamily Papilinoidae.However, few lectins from the Mimosoideae species have beenreported (Suvachittanonts and Peutpaiboon, 1992; Utarabhandand Akkayanont, 1995; Cavada et al., 1997, 2000, 2006;Guzman-Partida et al., 2004; Kaur et al., 2005; Santi-Gadelhaet al., 2008). In the Mimosoideae subfamily, the glucose/mannose-specific lectin isolated from Parkia platycephala seeds
* Correspondence to: B. S. Cavada, Department of Biochemistry and MolecularBiology, Federal University of Ceará, Mister Hull s/n Bloco 907, Lab. 1075, POBox 6043, 60440-970, Fortaleza, CE, Brazil.E-mail: [email protected]
a H. C. Silva, A. U. Bari, B. A. M. Rocha, K. S. Nascimento, B. S. CavadaBioMol-Group, Department of Biochemistry and Molecular Biology, FederalUniversity of Ceará, PO Box 6043, 60440-970, Fortaleza, CE, Brazil
b E. L. Ponte, A. F. Pires, A. M. S. AssreuyBioMol-Group, Institute of Biomedical Sciences, State University of Ceará,Fortaleza, CE, Brazil
c P. DelatorreBioMol-Group, Department of Molecular Biology, Federal University of Paraíba,João Pessoa, PB, Brazil
d E. H. TeixeiraBioMol-Group, Department of Pathology and Legal Medicine, Faculty ofMedicine, Federal University of Ceará, Fortaleza, CE, Brazil
e H. DebrayUniversity of Science and Technology of Lille, Lille, France
f C. S. NaganoBioMol-Group, Department of Fishing Engineering, Federal University of Ceará,Fortaleza, CE, Brazil
Research Article
Received: 3 May 2013, Revised: 7 June 2013, Accepted: 7 June 2013, Published online in Wiley Online Library
(wileyonlinelibrary.com) DOI: 10.1002/jmr.2289
J. Mol. Recognit. 2013; 26: 470–478 Copyright © 2013 John Wiley & Sons, Ltd.
470

(PPL-1) was the first characterized with regard to the primary(Mann et al., 2001) and three-dimensional structure (Gallegodel Sol et al., 2005). PPL-1 contains a single, non-glycosylatedpolypeptide chain composed of three tandemly arrangedjacalin-related domains. The resolution of PPL-1 crystal structureshowed a novel circular arrangement of β-prism domains,highlighting the adaptability of a β-prism fold as building blocksin the evolution of plant lectins (Gallego del Sol et al., 2005).Parkia is a pantropical genus of trees comprising around 30
species found in the neotropics from Honduras to southeasternBrazil, West Africa, the northern part of Malaysia, and southernThailand (Hopkins, 1984), being the most primitive groupwithin Leguminosae. Parkia biglobosa is an African tree,reaching about 20m in height, widespread in the Sudanianand Guinean savannas, highly valued for its brown seeds thatare ground into a pungent nutritious spice or condiment(Teklehaimanot, 2004).Among the lectins belonging to Parkia genera, similar biolog-
ical properties have been reported, such as mitogenic effect onhuman lymphocytes (Suvachittanonts and Jaranchavanapet,2000), recognition of meningothelial tumor (Beltrão et al., 2003)and antiviral activity (Favacho et al., 2007). However, no studyof antinociceptive and anti-inflammatory activities involvinglectins of this taxa has been related.The present work aimed to purify and reveal the amino acid
sequence and the antinociceptive and anti-inflammatory effectsof a new mannose/glucose-binding jacalin-related lectin from P.biglobosa Jacq. seeds. This is the first report of a jacalin-relatedlectin able to negatively modulate the cellular events ofinflammation and inflammatory pain.
MATERIALS AND METHODS
Lectin purification
Mature seeds of P. biglobosa (collected in Bolama, GuineaBissau) were ground into a fine powder using a coffee mill.The powder was incubated in 1:10 (w/v) 100mM sodiumacetate buffer, pH 4.0, containing 150mM NaCl at room tem-perature under continuous stirring for 3 h before centrifugationat 9000 g for 20min. The crude extract was precipitated withammonium sulfate (0–60% saturation) and centrifuged at10 000 g. The pellet was resuspended, dialyzed in 50mM Tris–HCl buffer, pH 7.6, containing 150mM NaCl, and recentrifuged.The resultant supernatant (fraction 0–60%) was applied to across-linked dextran column (12 × 2 cm Sephadex G-100Superfine, Sigma-Aldrich, St. Louis, MO, USA), previously equili-brated with the same solution. After removing unbound material,the lectin was eluted with 200mM D-glucose in equilibrium solution.Fractions of 2.5ml were collected at 1ml/min. The eluted sampleswere pooled, dialyzed (25 kDa cutoff membrane) against distilledwater, and freeze-dried. The purity of the isolated protein wasassessed by sodium dodecyl sulfate–polyacrylamide gel electro-phoresis (SDS–PAGE). The pure lectin was named P.biglobosalectin (PBL).
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
The purification process of the PBL was monitored by SDS–PAGEcarried out as described by Laemmli (1970) on slabs of 12% poly-acrylamide in the absence or presence of β-mercaptoethanol(2%) using a Mini-Protean II apparatus (Bio-Rad, Milan, Italy).
Electrophoresis was conducted at a constant current of 25mAfor 90min. The protein bands were visualized by stainingwith Coomassie Brilliant Blue R-250. The molecular markerswere phosphorylase b (97kDa), albumin (66 kDa), ovalbumin(45 kDa), carbonic anhydrase (30 kDa), trypsin inhibitor (20.1 kDa)and α-lactalbumin (14.4 kDa).
Protein and carbohydrate dosage
Protein content was determined using bovine serum albuminas standard (Bradford, 1976). Absorbance at 280 nm was alsoused to estimate protein concentration in the chromato-graphic fractions.
Neutral sugar content of PBL was estimated using the phenolsulfuric acid method with lactose as standard (Dubois et al.,1956). The presence of covalently bound carbohydrate wasconfirmed, staining the protein bands from electrophoresis gelwith periodic acid-Schiff reagent (Zacharius et al., 1969).
Hemagglutination assay
Hemagglutination was carried out in microtitration plates usingnative or enzyme-treated rabbit erythrocytes (Ainouz et al., 1992).The hemagglutination activity was given as a titer (the reciprocalof the highest twofold dilution) or expressed as the minimumprotein concentration (μg/ml), which produced macroscopicallyvisible agglutination and denoted as hemagglutinating unit (HU).Specific activity was defined as the HU per milligram of protein.Lectin carbohydrate-binding specificity was assessed by theminimalinhibitory concentration of sugar, giving full inhibition of agglutina-tion (Ramos et al., 1996). Twofold serial dilutions (150mMNaCl) wereprepared from the following saccharides: D-glucose, D-galactose,D-mannose, D-arabinose, D-fructose, N-acetyl-D-glucosamine, α-methyl-D-galactopyranoside, α-methyl-D-mannopyranoside, lactose,and lactulose (100mM initial concentration). The hemagglutinationinhibition titer was recorded as the inhibitor highest dilution, whichinhibited agglutination produced by 4 HU after a 60-min incubation.
Physicochemical assays
Assay of PBL (0.5mg/ml) at different pH and temperature, in thepresence of divalent cations, was carried out to establish itsstability. To measure the influence of pH, PBL was dialyzed for24 h against buffers (at 100mM) containing 150mM NaCl :sodium citrate (pH 4.0 and 6.0), sodium acetate (pH 5.0), sodiumphosphate (pH 7.0), Tris–HCl (pH 8.0) and glycine–NaOH (pH 9.0and 10.0). The influence of divalent cations was measured afterdialysis of PBL against 100mM ethylenediaminetetraacetic acid(EDTA) containing 150mM NaCl for 24 h at 4 °C, followed by anew dialysis against 150mM NaCl to remove EDTA excess fromthe sample. The recovered activity was determined afteraddition of CaCl2 and MnCl2 (10mM). The influence of tempera-ture was determined by incubation of PBL for 60min (10–100 °C).Samples were centrifuged at 9000 g for 10min before thehemagglutination assays.
Molecular mass determination
The average molecular mass of PBL was determined byelectrospray ionization–mass spectrometry (ESI–MS) using ahybrid quadrupole/ion mobility separator/orthogonal accelera-tion time-of-flight mass spectrometer (SYNAPT HDMS System,Waters Corp., Milford, MA, USA). PBL (10μmol/μl in 50%
PURIFICATION OF A LECTIN FROM Parkia biglobosa Jacq. SEEDS
J. Mol. Recognit. 2013; 26: 470–478 Copyright © 2013 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmr
471

acetonitrile with 0.1% formic acid) was infused into the system at0.2μl/min. The capillary and cone voltages were set at 3.5 kV and40 V, respectively. The source temperature was maintained at100 °C, and nitrogen was used as a drying gas (flow rate of150 l/h). The instrument was calibrated with average m/z(charges +12 to +26) of horse heart myoglobin. Data acquisitionwas performed atm/z range of 1000–6000 using Mass Lynx 4.0 soft-ware (Waters), and the multiply charge spectra were deconvolutedusing maximum entropy techniques (Ferrige et al., 1992). PBL nativemolecular mass was estimated by gel filtration in a TSK G3000SWxLcolumn (7.8mm ID×30.0 cm L) (Tosoh Bioscience, Stuttgart,Germany) coupled to high-performance liquid chromatography(LC) system (ÄKTA purifier, Amersham Biosciences) at 0.5ml/min.The column was equilibrated with 50mM Tris–HCl buffer, pH7.6,containing 500mM NaCl and calibrated with standard proteins: β-amylase (200kDa), conalbumin (75kDa), ovalbumin (43 kDa) andcarbonic anhydrase (29 kDa).
Proteolytic cleavages
The protein digestion was performed as previously described byShevchenko and co-workers (2006). For this, PBL was submittedto SDS–PAGE, and the bands were excised and bleached with50mM ammonium bicarbonate in 50% acetonitrile. Bands weredehydrated in 100% acetonitrile and dried in a Speedvac(LabConco, Kansas City, MO, USA). For proteolytic cleavage,performed at 37 °C overnight, gels were rehydrated with thefollowing enzyme solutions: 50mM ammonium bicarbonatecontaining trypsin (Promega, Madison, WI, USA), chymotrypsin(Sigma-Aldrich), endoproteinase Asp-N (Roche, Penzberg, Germany),or endoproteinase Glu-C (Roche) (1:100 w/w; enzyme : substrate ratio);100mM Tris–HCl (pH7.6) containing 50mM NaCl; 50mM CaCl2containing thermolysin (Sigma-Aldrich) (1:50 w/w; enzyme : sub-strate ratio); and 10mM HCl containing pepsin (Sigma-Aldrich).The obtained peptides were extracted in a solution of 50% acetoni-trile with 5% formic acid and concentrated in Speedvac.
Tandem mass spectrometry analysis
The peptides obtained in the different proteolytic cleavageswere sep-arated on a BEH300 C18 column (100μm×100mm) using ananoAcquity™ system (Waters) and eluted with acetonitrile gradient(10–85%), containing 0.1% formic acid at 600μl/min. The LC systemwas connected to a nanoelectrospray mass spectrometer source(SYNAPTHDMS System). Themass spectrometer was operated in pos-itive mode, using a source temperature of 80 °C and capillary voltageof 3.5kV. The instrument was calibrated with fragments of the doubleprotonated ion [Glu1]-fibrinopeptide B (Sigma-Aldrich) (m/z785.84).The LC–MS/MS experiment was performed according to thedata-dependent acquisition function, selecting the MS/MS doublyor triply charged precursor ions, which were fragmented bycollision-induced dissociation (CID) using argon as collision gas andramp collision energy that varied according to the charge state ofthe selected precursor ion. The data were processed and analyzedusing Proteinlynx (Waters), and the search parameter was thefragmentation pattern of the peptides. The CID spectra wereinterpreted manually using the Peptide Sequencing tool from MassLynx 4.0 software (Waters).
Primary structure analysis
Primary sequence was submitted to Basic Local Alignment SearchTool (Altschul et al., 1997), and the proteins with suitable e-value
were selected to sequence alignments at ESPript 2.2 (Gouet et al.,2003). The identity score was determined using ClustalW, and thetheoretical pI was determined using ProtParam (Gasteiger et al.,2005). The theoretical average molecular mass of the amino acidsequence was calculated using the PeptideMass tool (http://web.expasy.org/peptide_mass/) (Gasteiger et al., 2005), and the presenceof N-glycosylation sites was evaluated using the NetNGlyc tool(http://www.cbs.dtu.dk/services/NetNGlyc/) (Gupta et al., 2004).
Biological assays
Animals
Mice (25–30g) were raised and housed (n=6 per cage) in roomswith a controlled 12/12h light/dark cycle at 25 °C with free accessto food and water. Experimental protocols were approved by theInstitutional Animal Care and Use Committee of the State Universityof Ceará (CEUA no. 10130208-8/40), Fortaleza, CE, Brazil, inaccordance with the Guide for the Care and Use of LaboratoryAnimals of the US Department of Health and Human Services (NIHpublication no. 85-23, revised 1985).
Writhing test
Mice received 0.6% acetic acid (v/v; 0.1ml/10g body weight) byintraperitoneal (i.p.) route, and the number of abdominal writhes(typical contractions of the abdominal musculature followed byhind-limb stretches) was counted from 10 to 30min after injection(Koster et al., 1959). PBL (0.01–1mg/kg) or saline (50μl/10g bodyweight) was injected intravenously (i.v.), and indomethacin (5mg/kg) was injected i.p. 30min before administration of acetic acid.Results were expressed as the number of writhes.
Peritonitis model
Carrageenan (1mg), N-formyl-methionyl-leucyl-phenylalanine(fMLP; 50ng) or NaCl 0.9% (sterile saline; 100μl/10g body mass)was injected in the animal peritoneal cavities 4h before euthanasia.Peritoneal cells were harvested with 3ml saline (5 IU heparin) fortotal and differential leukocyte counts (Cytopro® CytocentrifugeModel 7620, Wescor Biomedical Systems, Logan, UT, USA) (Souzaand Ferreira, 1985). Results were expressed as the number of leuko-cytes per microliter of peritoneal fluid. Animals received saline orPBL (0.001–1mg/kg) i.v. 30min before administration of stimulus.Results were expressed as the number leukocytes per milliliter.
Statistical analysis
Data were presented as mean± SEM (n= 6) and tested using theone-way analysis of variance and the Bonferroni test. Statisticaldifferences were set for p values less than 0.05.
RESULTS AND DISCUSSION
Crude protein extracts of P. biglobosa seeds possess relativelyhigh levels of hemagglutinating activity against rabbit bloodcells, either native or treated with proteolytic enzymes. Thepresence of lectin in P. biglobosa seeds was first demon-strated by the research group of Professor H. Debray in theLaboratoire de Chimie Biologique et Unité Mixte deRecherche (Université des Sciences et Technologies de Lille,France) and purified by Sepharose–mannose affinity chroma-tography (data not published).
H. C. SILVA ET AL.
wileyonlinelibrary.com/journal/jmr Copyright © 2013 John Wiley & Sons, Ltd. J. Mol. Recognit. 2013; 26: 470–478
472
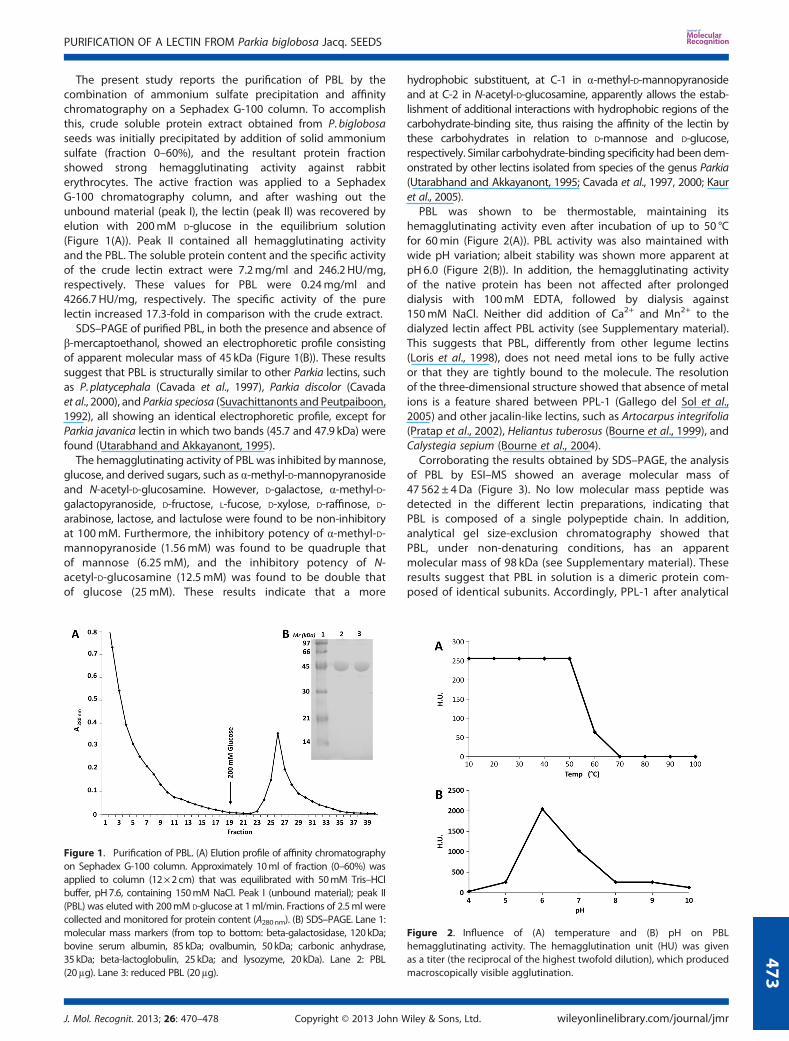
The present study reports the purification of PBL by thecombination of ammonium sulfate precipitation and affinitychromatography on a Sephadex G-100 column. To accomplishthis, crude soluble protein extract obtained from P. biglobosaseeds was initially precipitated by addition of solid ammoniumsulfate (fraction 0–60%), and the resultant protein fractionshowed strong hemagglutinating activity against rabbiterythrocytes. The active fraction was applied to a SephadexG-100 chromatography column, and after washing out theunbound material (peak I), the lectin (peak II) was recovered byelution with 200mM D-glucose in the equilibrium solution(Figure 1(A)). Peak II contained all hemagglutinating activityand the PBL. The soluble protein content and the specific activityof the crude lectin extract were 7.2mg/ml and 246.2 HU/mg,respectively. These values for PBL were 0.24mg/ml and4266.7 HU/mg, respectively. The specific activity of the purelectin increased 17.3-fold in comparison with the crude extract.SDS–PAGE of purified PBL, in both the presence and absence of
β-mercaptoethanol, showed an electrophoretic profile consistingof apparent molecular mass of 45 kDa (Figure 1(B)). These resultssuggest that PBL is structurally similar to other Parkia lectins, suchas P.platycephala (Cavada et al., 1997), Parkia discolor (Cavadaet al., 2000), and Parkia speciosa (Suvachittanonts and Peutpaiboon,1992), all showing an identical electrophoretic profile, except forParkia javanica lectin in which two bands (45.7 and 47.9 kDa) werefound (Utarabhand and Akkayanont, 1995).The hemagglutinating activity of PBL was inhibited bymannose,
glucose, and derived sugars, such as α-methyl-D-mannopyranosideand N-acetyl-D-glucosamine. However, D-galactose, α-methyl-D-galactopyranoside, D-fructose, L-fucose, D-xylose, D-raffinose, D-arabinose, lactose, and lactulose were found to be non-inhibitoryat 100mM. Furthermore, the inhibitory potency of α-methyl-D-mannopyranoside (1.56mM) was found to be quadruple thatof mannose (6.25mM), and the inhibitory potency of N-acetyl-D-glucosamine (12.5mM) was found to be double thatof glucose (25mM). These results indicate that a more
hydrophobic substituent, at C-1 in α-methyl-D-mannopyranosideand at C-2 in N-acetyl-D-glucosamine, apparently allows the estab-lishment of additional interactions with hydrophobic regions of thecarbohydrate-binding site, thus raising the affinity of the lectin bythese carbohydrates in relation to D-mannose and D-glucose,respectively. Similar carbohydrate-binding specificity had been dem-onstrated by other lectins isolated from species of the genus Parkia(Utarabhand and Akkayanont, 1995; Cavada et al., 1997, 2000; Kauret al., 2005).
PBL was shown to be thermostable, maintaining itshemagglutinating activity even after incubation of up to 50 °Cfor 60min (Figure 2(A)). PBL activity was also maintained withwide pH variation; albeit stability was shown more apparent atpH 6.0 (Figure 2(B)). In addition, the hemagglutinating activityof the native protein has been not affected after prolongeddialysis with 100mM EDTA, followed by dialysis against150mM NaCl. Neither did addition of Ca2+ and Mn2+ to thedialyzed lectin affect PBL activity (see Supplementary material).This suggests that PBL, differently from other legume lectins(Loris et al., 1998), does not need metal ions to be fully activeor that they are tightly bound to the molecule. The resolutionof the three-dimensional structure showed that absence of metalions is a feature shared between PPL-1 (Gallego del Sol et al.,2005) and other jacalin-like lectins, such as Artocarpus integrifolia(Pratap et al., 2002), Heliantus tuberosus (Bourne et al., 1999), andCalystegia sepium (Bourne et al., 2004).
Corroborating the results obtained by SDS–PAGE, the analysisof PBL by ESI–MS showed an average molecular mass of47 562± 4Da (Figure 3). No low molecular mass peptide wasdetected in the different lectin preparations, indicating thatPBL is composed of a single polypeptide chain. In addition,analytical gel size-exclusion chromatography showed thatPBL, under non-denaturing conditions, has an apparentmolecular mass of 98 kDa (see Supplementary material). Theseresults suggest that PBL in solution is a dimeric protein com-posed of identical subunits. Accordingly, PPL-1 after analytical
Figure 1. Purification of PBL. (A) Elution profile of affinity chromatographyon Sephadex G-100 column. Approximately 10ml of fraction (0–60%) wasapplied to column (12×2cm) that was equilibrated with 50mM Tris–HClbuffer, pH7.6, containing 150mM NaCl. Peak I (unbound material); peak II(PBL) was eluted with 200mM D-glucose at 1ml/min. Fractions of 2.5ml werecollected and monitored for protein content (A280nm). (B) SDS–PAGE. Lane 1:molecular mass markers (from top to bottom: beta-galactosidase, 120kDa;bovine serum albumin, 85 kDa; ovalbumin, 50 kDa; carbonic anhydrase,35 kDa; beta-lactoglobulin, 25 kDa; and lysozyme, 20kDa). Lane 2: PBL(20μg). Lane 3: reduced PBL (20μg).
Figure 2. Influence of (A) temperature and (B) pH on PBLhemagglutinating activity. The hemagglutination unit (HU) was givenas a titer (the reciprocal of the highest twofold dilution), which producedmacroscopically visible agglutination.
PURIFICATION OF A LECTIN FROM Parkia biglobosa Jacq. SEEDS
J. Mol. Recognit. 2013; 26: 470–478 Copyright © 2013 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmr
473

ultracentrifugation equilibrium sedimentation behaves as adimer (Mann et al., 2001).
The primary structure of PBL was established by sets ofoverlaps peptides obtained by proteolytic digestions. The aminoacid sequences of the peptides were determined by CID–MS/MS,in which 64 peptides were sequenced, resulting in a total of 443amino acid residues (Figure 4). Sequence heterogeneity wasfound in the positions D/S83, D/A163, G/R211, D/S346, A/G347,and D/S376, indicating that PBL preparations contain highlyhomologous isolectins. The CID spectra of two isoforms ofpeptide T18, in which sequence-specific y-ions were used forthe structure determination, showed the replacement of Ala byGly in position 347 (Figure 5).
Comparisons with other sequences from the National Centerfor Biotechnology Information data bank revealed that PBL
displays sequence similarity with mannose-binding jacalin-related lectins, such as those from A. integrifolia (TrEMBLaccession code: Q7M1T4), H. tuberosus (TrEMBL accession code:Q8S3V3), Castanea crenata (SwissProt accession code: P82859),and, mainly, with P. platycephala (SwissProt accession code:P83304), with which it shares 94% of sequence similarity. Theselectins are widespread among various taxonomically distantgroups, including alga (Mori et al., 2005) and plants, such asPteridophyta (Tateno et al., 2003), Spermatophyta (Nakamuraet al., 2002), and Angiosperm (Bourne et al., 1999; Zhang et al.,2000; Mann et al., 2001). This suggests that mannose-bindingjacalin-related lectins are a primitive protein group and wereconserved along the evolutionary process. Moreover, they showsequence similarity with proteins inducible by the planthormones jasmonic acid and salicylic acid (Abebe et al., 2005;Ma et al., 2010; Xiang et al., 2011) and with saltstress-inducedgene products (Zhang et al., 2000) of a number of differentplants, suggesting a common ancestry for jacalin-related lectinsand inducible defense proteins.Amino acid sequence alignment between PBL and PPL-1 using
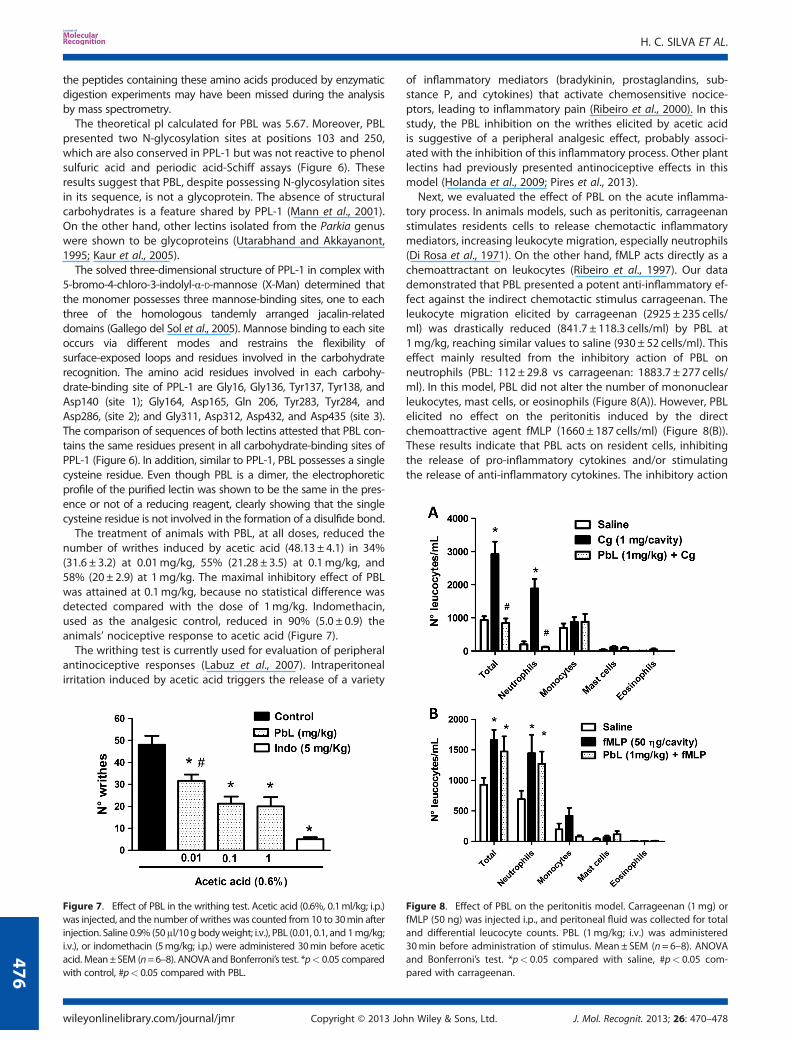
ESPript 2.1 (Gouet et al., 2003) revealed that, similar to PPL-1(Mannet al., 2001), PBL is a single polypeptide chain composed of threetandemly arranged jacalin-related domains (Figure 6). ConsideringAsp83, Asp163, Gly211, Asp346, Ala347, and Asp376 in PBL, thecalculated averaged mass of the 443 amino acid sequence was47236Da, which is 326Da lower than that determined by massspectrometry. PPL-1 presents three amino acid residues at its N-terminus (Ser-Leu-Lys), which were not found in PBL (Figure 6).However, considering that PBL possesses the same three amino acidresidues in N-terminus, the calculated mass increased to 47565Da,which is notably close to the mass observed using ESI–MS(47562±4). This suggests that PBLmay contain these and possessesthe same sequence that PPL-1 has at the N-terminus; nevertheless,
Figure 3. Electrospray ionization–mass spectrometry analysis of PBL.Deconvoluted mass spectra showing the isotopic average mass of PBLobtained using a hybrid mass spectrometer (SYNAPT HDMS System).
Figure 4. PBL amino acid sequence assembled from sequences of overlapping degradation products generated by proteolytic cleavage. Trypsin (T),chymotrypsin (C), endoproteinase Asp-N (D), endoproteinase Glu-C (G), thermolysin (Th), and pepsin (P). The letter X in PBL represents residues of leucine orisoleucine, which cannot be distinguished by mass.
H. C. SILVA ET AL.
wileyonlinelibrary.com/journal/jmr Copyright © 2013 John Wiley & Sons, Ltd. J. Mol. Recognit. 2013; 26: 470–478
474
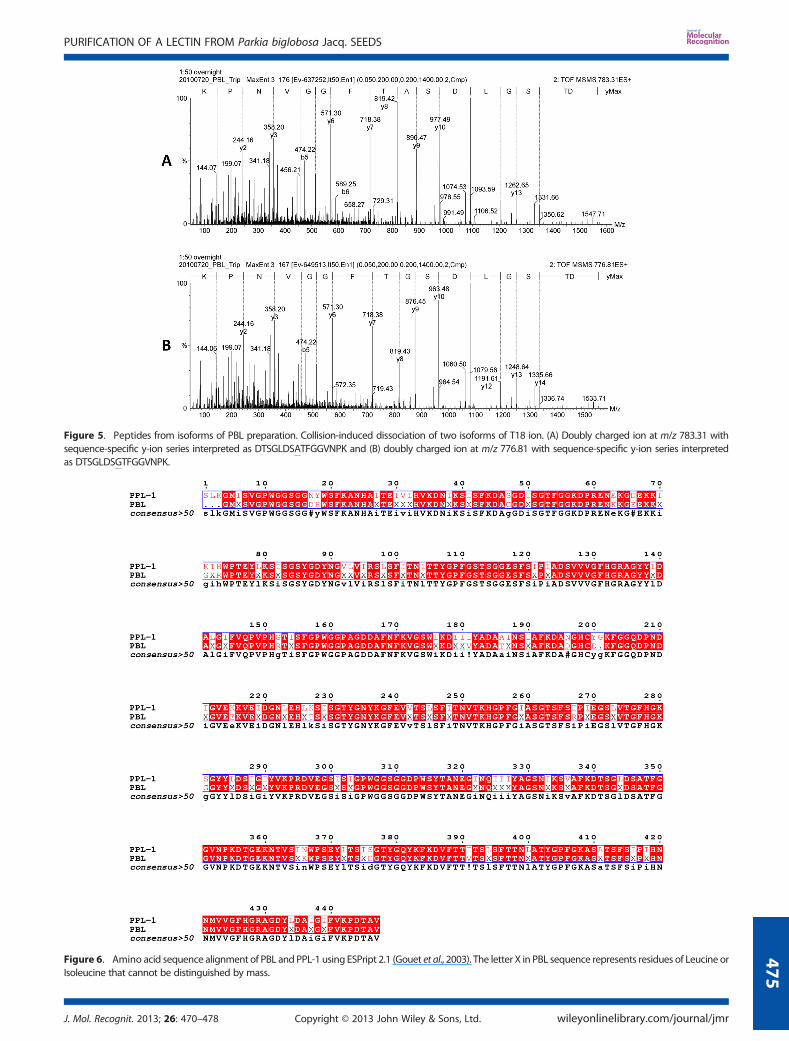
Figure 5. Peptides from isoforms of PBL preparation. Collision-induced dissociation of two isoforms of T18 ion. (A) Doubly charged ion at m/z 783.31 withsequence-specific y-ion series interpreted as DTSGLDSATFGGVNPK and (B) doubly charged ion at m/z 776.81 with sequence-specific y-ion series interpretedas DTSGLDSGTFGGVNPK.
Figure 6. Amino acid sequence alignment of PBL and PPL-1 using ESPript 2.1 (Gouet et al., 2003). The letter X in PBL sequence represents residues of Leucine orIsoleucine that cannot be distinguished by mass.
PURIFICATION OF A LECTIN FROM Parkia biglobosa Jacq. SEEDS
J. Mol. Recognit. 2013; 26: 470–478 Copyright © 2013 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmr
475
the peptides containing these amino acids produced by enzymaticdigestion experiments may have been missed during the analysisby mass spectrometry.
The theoretical pI calculated for PBL was 5.67. Moreover, PBLpresented two N-glycosylation sites at positions 103 and 250,which are also conserved in PPL-1 but was not reactive to phenolsulfuric acid and periodic acid-Schiff assays (Figure 6). Theseresults suggest that PBL, despite possessing N-glycosylation sitesin its sequence, is not a glycoprotein. The absence of structuralcarbohydrates is a feature shared by PPL-1 (Mann et al., 2001).On the other hand, other lectins isolated from the Parkia genuswere shown to be glycoproteins (Utarabhand and Akkayanont,1995; Kaur et al., 2005).
The solved three-dimensional structure of PPL-1 in complex with5-bromo-4-chloro-3-indolyl-α-D-mannose (X-Man) determined thatthe monomer possesses three mannose-binding sites, one to eachthree of the homologous tandemly arranged jacalin-relateddomains (Gallego del Sol et al., 2005). Mannose binding to each siteoccurs via different modes and restrains the flexibility ofsurface-exposed loops and residues involved in the carbohydraterecognition. The amino acid residues involved in each carbohy-drate-binding site of PPL-1 are Gly16, Gly136, Tyr137, Tyr138, andAsp140 (site 1); Gly164, Asp165, Gln 206, Tyr283, Tyr284, andAsp286, (site 2); and Gly311, Asp312, Asp432, and Asp435 (site 3).The comparison of sequences of both lectins attested that PBL con-tains the same residues present in all carbohydrate-binding sites ofPPL-1 (Figure 6). In addition, similar to PPL-1, PBL possesses a singlecysteine residue. Even though PBL is a dimer, the electrophoreticprofile of the purified lectin was shown to be the same in the pres-ence or not of a reducing reagent, clearly showing that the singlecysteine residue is not involved in the formation of a disulfide bond.
The treatment of animals with PBL, at all doses, reduced thenumber of writhes induced by acetic acid (48.13 ± 4.1) in 34%(31.6 ± 3.2) at 0.01mg/kg, 55% (21.28 ± 3.5) at 0.1mg/kg, and58% (20± 2.9) at 1mg/kg. The maximal inhibitory effect of PBLwas attained at 0.1mg/kg, because no statistical difference wasdetected compared with the dose of 1mg/kg. Indomethacin,used as the analgesic control, reduced in 90% (5.0 ± 0.9) theanimals’ nociceptive response to acetic acid (Figure 7).
The writhing test is currently used for evaluation of peripheralantinociceptive responses (Labuz et al., 2007). Intraperitonealirritation induced by acetic acid triggers the release of a variety
of inflammatory mediators (bradykinin, prostaglandins, sub-stance P, and cytokines) that activate chemosensitive nocice-ptors, leading to inflammatory pain (Ribeiro et al., 2000). In thisstudy, the PBL inhibition on the writhes elicited by acetic acidis suggestive of a peripheral analgesic effect, probably associ-ated with the inhibition of this inflammatory process. Other plantlectins had previously presented antinociceptive effects in thismodel (Holanda et al., 2009; Pires et al., 2013).Next, we evaluated the effect of PBL on the acute inflamma-
tory process. In animals models, such as peritonitis, carrageenanstimulates residents cells to release chemotactic inflammatorymediators, increasing leukocyte migration, especially neutrophils(Di Rosa et al., 1971). On the other hand, fMLP acts directly as achemoattractant on leukocytes (Ribeiro et al., 1997). Our datademonstrated that PBL presented a potent anti-inflammatory ef-fect against the indirect chemotactic stimulus carrageenan. Theleukocyte migration elicited by carrageenan (2925 ± 235 cells/ml) was drastically reduced (841.7 ± 118.3 cells/ml) by PBL at1mg/kg, reaching similar values to saline (930 ± 52 cells/ml). Thiseffect mainly resulted from the inhibitory action of PBL onneutrophils (PBL: 112 ± 29.8 vs carrageenan: 1883.7 ± 277 cells/ml). In this model, PBL did not alter the number of mononuclearleukocytes, mast cells, or eosinophils (Figure 8(A)). However, PBLelicited no effect on the peritonitis induced by the directchemoattractive agent fMLP (1660 ± 187 cells/ml) (Figure 8(B)).These results indicate that PBL acts on resident cells, inhibitingthe release of pro-inflammatory cytokines and/or stimulatingthe release of anti-inflammatory cytokines. The inhibitory action
Figure 7. Effect of PBL in the writhing test. Acetic acid (0.6%, 0.1ml/kg; i.p.)was injected, and the number of writhes was counted from 10 to 30min afterinjection. Saline 0.9% (50μl/10g bodyweight; i.v.), PBL (0.01, 0.1, and 1mg/kg;i.v.), or indomethacin (5mg/kg; i.p.) were administered 30min before aceticacid. Mean±SEM (n=6–8). ANOVA and Bonferroni’s test. *p< 0.05 comparedwith control, #p< 0.05 compared with PBL.
Figure 8. Effect of PBL on the peritonitis model. Carrageenan (1mg) orfMLP (50 ng) was injected i.p., and peritoneal fluid was collected for totaland differential leucocyte counts. PBL (1mg/kg; i.v.) was administered30min before administration of stimulus. Mean± SEM (n=6–8). ANOVAand Bonferroni’s test. *p< 0.05 compared with saline, #p< 0.05 com-pared with carrageenan.
H. C. SILVA ET AL.
wileyonlinelibrary.com/journal/jmr Copyright © 2013 John Wiley & Sons, Ltd. J. Mol. Recognit. 2013; 26: 470–478
476

of glucose–mannose-binding lectins from the Diocleinae subtribehad already been described (Assreuy et al., 1997; Alencar et al.,1999), but this is the first report of a jacalin-related lectin able tonegatively modulate the cellular events of inflammation andinflammatory pain.
CONCLUSION
The results reported here clearly demonstrate the purification andcharacterization of a new mannose/glucose-specific lectin fromP. biglobosa seeds (PBL), which, similar to the glucose/mannose-specific lectin isolated from P.platycephala seeds, is a single,non-glycosylated polypeptide chain composed of three tandemlyarranged jacalin-related domains. PBL exhibits important
anti-inflammatory activity, associated to leukocyte mobilization,and antinociceptive activity, associated to inflammatory pain.These biological activities highlight the importance of sugar–protein interactions in rodent leukocytes.
Acknowledgements
This studywas partly financed by Coordenação de Aperfeiçoamentode Pessoal de Nível Superior (CAPES), Conselho Nacional deDesenvolvimento Científico e Tecnológico (CNPq), and FundaçãoCearense de Apoio ao Desenvolvimento Científico e Tecnológico(FUNCAP). M. David Martin helped with the English editing of themanuscript. B. A.M. R., K. S. N., C. S. N., P. D., E. H. T., A.M. S. A., and B. S.C. are senior investigators of CNPq (Brazil).
REFERENCES
Abebe T, Skadsen RW, Kaeppler HF. 2005. A proximal upstream sequencecontrols tissue-specific expression of Lem2, a salicylate induciblebarley lectin-like gene. Planta 221: 170–183.
Ainouz IL, Sampaio AH, Benevides NMB, Freitas ALP, Costa FHF, Carvalho MR,Joventino FP. 1992. Agglutination of enzyme treated erythrocytes byBrazilian marine algae extracts. Botânica Marinha. 35: 447–479.
Alencar NM, Teixeira EH, Assreuy AM, Cavada BS, Flores CA, Ribeiro RA.1999. Leguminous lectins as tools for studying the role of sugarresidues in leukocyte recruitment. Mediators Inflamm. 8(2): 107–113.
Altschul SF, Maddem TL, Scharffer AA, Zhang L, Zhang Z, Miller W, LipmanDL.1997. Gapped BLAST and PSI-BLAST: a new generation of proteindatabase search programs. Nucleic Acids Res. 25: 3389–3402.
Assreuy AM, Shibuya MD, Martins GJ, De Souza ML, Cavada BS, MoreiraRA, Oliveira JT, Ribeiro RA, Flores CA.1997. Anti-inflammatory effectof glucose-mannose binding lectins isolated from Brazilian beans.Mediators Inflamm. 6(3): 201–10.
Beltrão EIC, Medeiros PL, Rodrigues OG, Figueredo-Silva J, Valenc¸a MM,Coelho LCBB, Carvalho LB Jr. 2003. Parkia pendula lectin as histochemis-try marker for meningothelial tumor. Eur. J. Histochem. 47(2):139–142.
Bourne Y, Zamboni V, Barre A, Peumans WJ, van Damme EJM, Rouge P.1999. Helianthus tuberosus lectin reveals a widespread scaffold formannose-binding lectins. Structure 7: 1473–1482.
Bourne Y, Roig-Zamboni V, Barre A, Peumans WJ, Houlès-Astoul C, VanDamme EJM 2004. The crystal structure of the Calystegia sepiumagglutinin reveals a novel quaternary arrangement of lectin subunitswith a b-prism fold. J. Biol. Chem. 279: 527–533.
Bradford MMA. 1976. A rapid and sensitive method for the quantification ofmicrogram quantities of protein utilizing the principles of protein-dyebinding. Anal. Biochem. 72: 248–254.
Cavada BS, Santos CF, Grangeiro TB, Moreira Silva LIMS, Campos MJO, SousaFAM, Calvete JJ. 1997. Isolation and partial characterization of a lectinfrom Parkia platycephala Benth seeds. Physiol. Mol. Biol. Plants. 3:109–115.
Cavada BS, Madeira SVF, Calvete JJ, Sousa LAG, Bomfim LR, DantasAR, Lopes MC, Grangeiro TB, Freitas BT, Pinto VPT, Leite KB,Ramos MV. 2000. Purification, chemical, and immunochemicalproperties of a new lectin from Mimosoideae (Parkia discolor).Prep. Biochem. Biotech. 30: 271–280.
Cavada BS, Moreno FBB, Rocha BAM, Azevedo WF Jr, Castellón RE,Goersch GV, Nagano CS, Souza EP, Nascimento KS, Radis-BaptistaG, Delatorre P, Leroy Y, Toyama MH, Pinto VPT, Sampaio AH,Barettino D, Debray H, Calvete JJ, Sanz L. 2006. cDNA cloning and1.75 A crystal structure determination of PPL2, an endochitinaseand N-acetylglucosamine-binding hemagglutinin from Parkiaplatycephala seeds. FEBS J. 273: 3962–3974.
Di Rosa M, Giroud JP, Willoughby DA. 1971. Studies on the mediators of theacute inflammatory response induced in rats in different sites bycarrageenan and turpentine. J. Pathol. 104: 15–29.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Shith F. 1956. Colorimetricmethod for determination of sugar and related substances. Anal.Biochem. 28: 350–356.
Favacho ARM, Cintra EA, Coelho LCBB, Linhares MIS. 2007. In vitro activityevaluation of Parkia pendula seed lectin against human cytomegalovirusand herpes virus 6. Biologicals 35: 189–194.
Ferrige AG, Seddon MJ, Green BN, Jarvis SA, Skilling J. 1992.Disentangling electrospray spectra with maximum entropy. RapidCommun. Mass Spectrom. 6: 707–711.
Gabius HJ, Gabius S. 1997. Glycoscience. Status and perspectives.Weinheim, Germany: Chapman & Hall.
Gallego del Sol F, Nagano CS, Cavada BS, Calvete JJ. 2005. The first crystalstructure of a Mimosoideae lectin reveals a novel quaternaryarrangement of a widespread domain. J. Mol. Biol. 353: 574–583.
Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins MR, Appel RD,Bairoch A. 2005. Protein identification and analysis tools on theExPASy server. In The Proteomics Protocols Handbook, Walker JM(ed). Humana Press: Totowa, NJ, USA; 571–607.
Gouet P, Robert X, Courcelle E. 2003. ESPript/ENDscript: extracting and ren-dering sequence and 3D information from atomic structure of proteins.Nucleic Acids Res. 31: 3320–3323.
Gupta R, Jung E, Brunak S. 2004. Prediction of N-glycosylation sites in hu-man proteins. In preparation.
Guzman-Partida AM, Robles-Burgueno MR, Ortega-Nieblas M,Vazquez-Moreno I. 2004. Purification and characterization of complexcarbohydrate specific isolectins from wild legume seeds: Acaciaconstricta is (vinorama) highly homologous to Phaseolus vulgaris lectins.Biochimie 86: 335–42.
Holanda FR, Coelho-de-Sousa AN, Assreuy AMS, Leal-Cardoso JH,Pires AF, Nascimento KS, Teixeira CS, Cavada BS, Santos CF.2009. Antinociceptive activity of lectins from Diocleinae seedson acetic acid-induced writhing test in mice. Protein Pept. Lett.16: 1088–1092.
Hopkins HC. 1984. Floral biology and pollination ecology of the neotrop-ical species of Parkia. J. Ecol. 72: 01–23.
Kaur N, Singh J, Kamboj SS, Agrewala JN, KaurM. 2005. Two novel lectins fromParkia biglandulosa and Parkia roxburghii: isolation, physicochemicalcharacterization, mitogenicity and anti-proliferative activity. Protein Pept.Lett. 12(6): 585–95.
Koster R, Anderson M, Debeer EJ. 1959. Acetic-acid for analgesic screen-ing. Fed. Proc. 18: 412.
Labuz D, Mousa SA, Schafer M, Stein C, Machelska H. 2007. Relative con-tribution of peripheral versus central opioid receptors toantinociception. Brain Res. 1160: 30–38.
Laemmli UK. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227: 680–685.
Loris R, Hamelryck T, Bouckaert J, Wyns L. 1998. Legume lectin structure.Biochim. Biophys. Acta 1383: 9–36.
Ma QH, Tian B, Li YL. 2010. Overexpression of a wheat jasmonate-regulatedlectin increases pathogen resistance. Biochimie 92: 187–193.
Mann K, Farias CMSA, Gallego del Sol F, Santos CF, Grangeiro TB, NaganoCS, Cavada BS, Calvete JJ. 2001. The amino-acid sequence of the glu-cose/mannose-specific lectin isolated from Parkia platycephala seedsreveal three tandemly arranged jacalin-related domains. Eur. J.Biochem. 218: 4414–4422.
Mori T, O’Keefe BR, Sowder RC, Bringans S, Gardella R, Berg S, CochranP, Turpin JA, Buckheit RW Jr, McMahon JB, Boyd MR. 2005. Isola-tion and characterization of griffithsin, a novel HIV-inactivatingprotein, from the red alga Griffithsia sp. J. Biol. Chem. 280:9345–9353.
PURIFICATION OF A LECTIN FROM Parkia biglobosa Jacq. SEEDS
J. Mol. Recognit. 2013; 26: 470–478 Copyright © 2013 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmr
477

Nakamura S, Ikegami A, Matsumura Y, Nakanishi T, Nomura K. 2002. Molec-ular cloning and expression of the mannose/glucose specific lectinfrom Castanea crenata cotyledons. J. Biochem. 131(2): 241–246.
Pires AF, Assreuy AMS, Lopes EA, Celedônio NR, Soares CE, Rodrigues NV,Sousa PL, Benevides RG, Nagano CS, Cavada BS, Leal-Cardoso JH,Coelho-de-Souza AN, Santos CF. 2013. Opioid-like antinociceptive effectsof oral administration of a lectin purified from the seeds of Canavaliabrasiliensis. Fundam. Clin. Pharmacol. 27: 201–209.
Pratap JV, Jeyaprakash AA, Rani PG, Sekar K, Surolia A, VijayanM. 2002. Crystalstructures of artocarpin, a Moraceae lectin with mannose specificity, andits complex with methyl-alpha-D-mannose: implications to the genera-tion of carbohydrate specificity. J. Mol. Biol. 317(2): 237–47.
Ramos MV, Moreira RA, Cavada BS, Oliveira JTA, Rouge P. 1996. Interac-tion of lectins from the sub tribe Diocleinae with specific ligands.R. Bras. Fisiol. Veg. 8: 193–199.
Ribeiro RA, Souza-Filho MV, Souza MH, Oliveira SH, Costa CH, Cunha FQ,Ferreira HS. 1997. Role of resident mast cells and macrophages inthe neutrophil migration induced by LTB4, fMLP and C5a des arg.Int. Arch. Allergy Immunol. 112(1): 27–35.
Ribeiro RA, Vale ML, Thomazzi SM, Paschoalato AB, Poole S, Ferreira SH,Cunha FQ. 2000. Involvement of resident macrophages and mastcells in the writing nociceptive response induced by zimosan andacetic acid in mice. Eur. J. Pharmacol. 387: 111–118
Santi-Gadelha T, Rocha BAM, Oliveira CC, Aragão KS, Marinho ES, GadelhaCAA, Toyama MH, Pinto VPT, Nagano CS, Delatorre P, Martins JL, GalvaniFR, Sampaio AH, Debray H, Cavada BS. 2008. Purification of a PHA-likechitin-binding protein from Acacia farnesiana seeds: a time-dependentoligomerization protein. Appl. Biochem. Biotechnol. 150: 97–111.
Sharon N, Lis H. 2004. History of lectins: from hemagglutinins to biologi-cal recognition molecules. Glycobiology 14: 53–62.
Shevchenko A, Tomas H, Havlis J, Olsen JV, Mann M. 2006. In gel diges-tion for mass spectrometric characterization of proteins andproteomes. Nat. Protoc. 1: 2856–2860.
Souza GEP, Ferreira SH. 1985. Blockade by antimacrophage serum of themigration of PMN neutrophils into the inflamed peritoneal cavity.Agents Actions 17: 97–103.
Suvachittanonts W, Jaranchavanapet P. 2000. Mitogenic effect of Parkiaspeciosa seed lectin on human lymphocytes. Planta Med. 66: 699–704.
Suvachittanonts W, Peutpaiboon A. 1992. Lectin from Parkia speciosaseeds. Phytochemistry 31: 4065–4070.
Tateno H, Winter HC, Petryniak J, Goldstein IJ. 2003. Purification,characterization, molecular cloning, and expression of novelmembers of jacalin-related lectins from rhizomes of the true fernPhlebodium aureum (L) J. Smith (polypodiaceae). J. Biol. Chem. 278(13):10891–10899.
Taylor ME, Drickamer K. 2003. Introduction to glycobiology, OxfordUniversity Press: Oxford, UK.
Teklehaimanot Z 2004. Exploiting the potential of indigenous agrofor-estry trees: Parkia biglobosa and Vitellaria paradoxa in sub-SaharanAfrica. Agroforestry Systems 61: 207–220.
Utarabhand P, Akkayanont P. 1995. Purification of a lectin from Parkiajavanica beans. Phytochemistry 38: 281–285.
Van Damme EJM, Peumans WJ, Barre A, Rougé P. 1998. Plant lectins: acomposite of several distinct families of structurally and evolutionaryrelated proteins with diverse biological roles. Cr. Rev. Plant Sci. 17:575–692.
Xiang Y, Song M, Wei Z, Tong J, Zhang L, Xiao L, Ma Z, Wang Y. 2011. Ajacalin-related lectin-like gene in wheat is a component of the plantdefence system. J. Exp. Bot. 62(15): 5471–83.
Zacharius RM, Zell TE, Morrison JH, Woodlock JJ. 1969. Glycoproteinstaining following electrophoresis on acrylamide gels. Anal. Biochem.30(1): 148–152.
Zhang W, Peumans WJ, Barre A, Astoul CH, Rovira P, Rougé P, Proost P,Truffa-Bachi P, Jalali AA, Van Damme EJ. 2000. Isolation and charac-terization of a jacalin-related mannose-binding lectin from salt-stressed rice (Oryza sativa) plants. Planta 210(6): 970–8.
H. C. SILVA ET AL.
wileyonlinelibrary.com/journal/jmr Copyright © 2013 John Wiley & Sons, Ltd. J. Mol. Recognit. 2013; 26: 470–478
478




















